Embed Size (px)
Citation preview

Alterations in DICER and miRNA levels with cigarette smoke
1
A microRNA processing defect in smokers' macrophages is linked to SUMOylation of the
endonuclease DICER
Thomas J. Gross, Linda S. Powers, Ryan L. Boudreau, Brandi Brink, Anna Reisetter,
Khushboo Goel, Alicia K. Gerke, Ihab H. Hassan and Martha M. Monick
From the Department of Medicine, Carver College of Medicine,
University of Iowa, Iowa City, Iowa, 52242
* Running title: Alterations in DICER and miRNA levels with cigarette smoke
To whom correspondence should be addressed: Thomas J. Gross, Division of Pulmonary, Critical
Care, and Occupational Medicine, University of Iowa, Iowa City, IA 52242, Phone: (319) 356-6268,
Email: [email protected]
Keywords: COPD, DICER, inflammation, miRNA, macrophages
Background: Smoking causes a global down
regulation in alveolar macrophage miRNA
expression.
Results: Cigarette smoke exposure modifies the
RNA endonuclease DICER, resulting in a
microRNA-processing defect.
Conclusion: Cigarette smoke alters alveolar
macrophage microRNA expression, in part, by
SUMOylation of DICER.
Significance: This is the first description of an
environmental exposure causing changes in
microRNA expression via post-translational
modification of Dicer.
ABSTRACT
Despite the fact that alveolar macrophages
play an important role in smoking-related disease,
little is known about what regulates their
pathophysiologic phenotype. Evaluating smoker
macrophages, we found significant down
regulation of multiple microRNAs (miRNAs).
This project investigates the hypothesis that
cigarette smoke alters mature miRNA expression
in lung macrophages by inhibiting processing of
primary miRNA transcripts. Studies on smoker
alveolar macrophages showed a defect in miRNA
maturation. Studies on the miRNA biogenesis
machinery led us to focus on the cytosolic RNA
endonuclease, DICER. DICER cleaves the stem-
loop structure from pre-miRNAs, allowing them to
dissociate into their mature 20-22 nucleotide
single stranded form. DICER activity assays
confirmed impaired DICER activity following
cigarette smoke exposure. Further protein studies
demonstrated a decreased expression of the native
217 kD form of DICER and an accumulation of
high molecular weight forms with cigarette smoke
exposure. This molecular mass shift was shown to
contain SUMO moieties and could be blocked by
silencing RNA directed at the primary
SUMOylating ligase, Ubc9. In determining the
cigarette smoke components responsible for
changes in DICER, we found that N-acetyl-
cysteine (NAC), an anti-oxidant and anti-
aldehyde, protected DICER protein and activity
from cigarette smoke extract (CSE). This massive
down regulation of miRNAs (driven in part by
alterations in DICER) may be an important
regulator of the disease promoting macrophage
phenotype found in smoker's lungs.
INTRODUCTION
COPD is characterized by inflammation in
both small airways and alveoli eventually leading
to irreversible airflow limitation and parenchymal
emphysema. Lung pathology in patients with
COPD has been largely attributed to neutrophil
elastases and macrophage proteinases (1,2). While
multiple lung cell types contribute to the
pathogenesis of COPD, evidence strongly supports
a key role for alveolar macrophages (3-5). Inhaled
tobacco smoke triggers disease-relevant changes
in alveolar macrophage gene expression (i.e.
MMP12) (6,7). While the regulation of disease-
relevant genes is complex and multifactorial,
increasing experimental evidence suggests that
macrophage gene expression is strongly
influenced by miRNAs (8,9). Our laboratory has
http://www.jbc.org/cgi/doi/10.1074/jbc.M114.565473The latest version is at JBC Papers in Press. Published on March 25, 2014 as Manuscript M114.565473
Copyright 2014 by The American Society for Biochemistry and Molecular Biology, Inc.
by guest on October 12, 2019
http://ww
w.jbc.org/
Dow
nloaded from

Alterations in DICER and miRNA levels with cigarette smoke
2
demonstrated that cigarette smoking is associated
with an impressive decrease in alveolar
macrophage miRNA expression (10). Detailing
the mechanism(s) behind this significant decrease
in miRNA levels could shed insight into the
pathophysiology of smoking-related lung diseases
and improve understanding of the adverse health
effects of cigarettes in general.
The role of miRNA levels in regulating
cellular genomic tone is an area of active
investigation (11-13). Primary miRNAs are
transcribed in the nucleus and processed into
precursor miRNA (pre-miRNA) by a nuclear
endoribonuclease, Drosha, in complex with a
double-stranded RNA binding protein, Pasha
(known as DGCR8 in vertebrates). The pre-
miRNA is an approximately 70 base pair double-
stranded RNA stem-loop structure that contains
the imbedded 22-nucleotide sequence
complementary to the mRNA target of interest.
Pre-miRNAs are exported from the nucleus where
the cytoplasmic endoribonuclease, DICER, excises
the stem-loop yielding a 20 to 25 base-pair RNA
duplex (14,15). In the cytosol, DICER, in complex
with other protein components (transactivating
response RNA-binding protein TRBP, protein
kinase RNA activator PACT, and Argonaut
protein Ago2) binds the duplex, slices out the anti-
complementary “passenger” RNA strand, and
targets it for destruction. The mature miRNA and
processing proteins form an RNA-induced
silencing complex (RISC), which represses
translation or enhances degradation of the targeted
complementary mRNA. Thus, up or down
regulation of an miRNA can result in specific
regulation of gene expression. The miRNA
biogenesis pathway has several potential points for
regulatory control. In this manuscript, we present
evidence that cigarette smoke exposure leads to a
post-translational modification of DICER,
resulting in decreased ribonuclease activity and
contributing to a generalized decrease in miRNA
expression in smokers' alveolar macrophages.
EXPERIMENTAL PROCEDURES
Reagents and antibodies: Antibodies used in
this study were obtained from a variety of sources:
anti-HA, Roche Biochemical Laboratories,
#11867423001; anti-DICER, Abcam, #ab14601;
anti-PACT, Abcam, #ab75749; anti-TRBP,
Abcam, #ab42018; anti actin, Abcam, #ab8226;
anti-UBC9, Abcam, ab21193; anti-SUMO 2/3,
Abcam, #ab3742; anti-ubiquitin, Cell Signaling,
#3936. Horseradish peroxidase conjugated anti-
rabbit (sc-2004), anti-mouse (sc-2005) and anti-
goat (sc-2020) antibodies were all obtained from
Santa Cruz Biotechnology. Culture media used in
experiments included serum free RPMI tissue
culture media with Glutamax (Invitrogen, 61870-
036) and DMEM (Invitrogen, 11095) with 10%
FCS (Hyclone Fetal Bovine Serum, SH30071).
Ethics statement: The University of Iowa
Institutional Review Board approved all
procedures and protocols described in this
communication. Written informed consent was
obtained and all clinical investigation has been
conducted according to the principles expressed in
the Declaration of Helsinki.
Alveolar macrophage donors: Subjects were
recruited from the community through the Iowa
Institute for Clinical and Translational Science
(ICTS) Clinical Core. Inclusion criteria for
smoking subjects required at least a 10 pack-year
history of ongoing cigarette smoking, while the
nonsmoker control subjects were self-reported
never smokers. Subjects were excluded if they had
any significant co-morbid conditions such as
pregnancy or other acute or chronic disease such
as pre-existing asthma, interstitial lung disease or
cardiovascular disease. Subjects were also
excluded if a baseline spirometry revealed the
forced expiratory volume in the first second was
less than 60% of the predicted value based on
National Health and Nutrition Examination Survey
III data set. None of the subjects were using
inhaled corticosteroids.
Bronchoalveolar lavage: After informed
consent was obtained, subjects underwent standard
flexible bronchoscopy (16). Local anesthesia with
lidocaine instillation into the upper airway was
followed by bronchoalveolar lavage (BAL)
whereby 20 ml of normal saline was instilled into
a tertiary bronchus up to five times in three
different lung segments. The first collection out of
five was discarded to avoid possible contamination
with upper airway secretions or lidocaine. The
remaining lavage was filtered through sterile
gauze and centrifuged at 200xg for 5 minutes to
pellet cellular material. The resulting pellet was
suspended in phosphate buffered saline (PBS) and
centrifuged at 200xg for 5 minutes. A sample of
the cells were labeled with Wright stain and
by guest on October 12, 2019
http://ww
w.jbc.org/
Dow
nloaded from

Alterations in DICER and miRNA levels with cigarette smoke
3
microscopically examined to determine the
proportion of the cells that were macrophages (17-
20). Aliquots of 5x106
cells were stored at -80°C
until RNA isolation procedure was performed.
Cell yields from bronchoscopy of nonsmokers
averaged 25±3 x 106 cells and for smokers
averaged, 67±4 x 106. In all three cohorts, the
procedure generated a relatively pure population
of alveolar macrophages with fewer than 5%
neutrophils or lymphocytes in the bronchoalveolar
lavage fluid.
CSE production and exposure: Cigarette
smoke extract was prepared as previously
described (21,22). CSE solutions were prepared
using a modification of the method of Blue and
Janoff (23). 10 mls of sterile serum free RPMI
tissue culture media with Glutamax was drawn
into a 50-ml plastic syringe. Then, 40 ml of
cigarette smoke was then drawn into the syringe
and mixed with the medium by vigorously shaking
them together. The smoke was expelled and the
process repeated until one cigarette was used up.
The generated CSE solution was filtered (0.22 μm)
to remove large particles (24,25). The resulting
solution was designated a 100% CSE solution and
was used immediately after generation. For the
CSE dialysis experiment, we dialyzed 5% CSE
overnight using Thermo Scientific’s 2000 and
10,000 MWCO Slide-A-Lyzer Dialysis Cassette
GP cassettes. RNA isolation: RNA was isolated from
alveolar macrophages using the mirVana miRNA
Isolation kit (Applied Biosystems (ABI)). The
quantity and quality of the RNA samples was
assessed using an Experion Automated
Electrophoresis Station (Bio-Rad). The RNA
quality indicator was above 8 for all samples
where values of greater than 8 indicate primarily
intact RNA on a scale of 1-10. After preparation,
RNA samples were stored at -80° C until use.
miRNA expression analysis: RNA from
alveolar macrophages of nonsmokers and active
smokers was reverse transcribed with MultiScribe
Reverse Transcriptase (ABI) using Megaplex
Primers version 2.0 (ABI). Changes in miRNA
expression were then determined using human
TaqMan Low Density Arrays version 2.0 (ABI).
Ct values calculated using SDS version 2.4 (ABI)
were exported to the Partek Genomics Suite to
calculate smoker-to-nonsmoker expression ratios.
Robust multiarray averaging (RMA)-normalized
data were subjected to an ANOVA model with
linear contrasts to calculate p-values. Individual
miRNA-specific assays were done using ABI kits
specific for the mature miRNA or for the primary
transcript (before processing).
mRNA expression analysis: Genome-wide
macrophage mRNA expression was obtained
using the GeneChip Human Exon 1.0 ST Arrays
(Affymetrix). Generation of labeled cDNA,
hybridizations, and scanning of the microarray
were performed under contract by the University
of Iowa DNA facility. The resulting data were
analyzed using the Partek Genomics Suite version
6.5 (Partek). The data were assessed for quality
and subjected to RMA normalization. The
normalized data were then analyzed using an
ANOVA model with linear contrasts to calculate
p-values and smoker-to-nonsmoker expression
ratios. The false discovery rate (FDR) step-up
method (26) was applied to correct for multiple
testing. The expression data has been deposited in
NCBI Geo repository (GSE34517). To generate
the data shown in figure3E, we mined RNA array
data performed on alveolar macrophages from 43
nonsmokers and 42 smokers. The log 2 values
obtained by the initial Partek analysis for the gene
DICER are shown in an individual value plot.
Possible significance between the two groups was
assessed by Student’s t test.
Whole cell protein isolation: Whole cell
protein was obtained by lysing the cells on ice for
20 minutes in 200 l of lysis buffer (0.05 M Tris
pH 7.4, 0.15 M NaCl, 1% NP-40, with added
protease and phosphatase inhibitors: 1 protease
Minitab (Roche Biochemical)/10 ml and 100 ul
100X phosphatase inhibitor cocktail (Calbiochem)
/10 ml. The lysates were sonicated for 20 seconds,
kept at 4o for 30 minutes, spun at 15,000 g for 10
minutes and the supernatant saved. Protein
determinations were made using the Bradford
Protein assay from Bio-Rad. Cell lysates were
stored at –70o until use.
Western analysis: Whole cell protein was
obtained as previously described (27). Following
protein purification, Western analysis was
performed following protocols as described by
Reisetter et al (16). The following antibodies were
used: anti-HA, Roche Biochemical Laboratories,
#11867423001; anti-DICER, Abcam, #ab14601;
anti-PACT, Abcam, #ab75749; anti-TRBP,
Abcam, #ab42018; anti actin, Abcam, #ab8226;
by guest on October 12, 2019
http://ww
w.jbc.org/
Dow
nloaded from

Alterations in DICER and miRNA levels with cigarette smoke
4
anti-UBC9, Abcam, ab21193; anti SUMO 2/3,
Abcam, #ab3742; and anti-ubiquitin, Cell
Signaling, #3936.
Immunoprecipitation (IP): IP of DICER was
performed on alveolar macrophages or THP-1
cells. Immunoprecipitation of cellular protein was
performed using the protein G Dynabead
immunoprecipitation kit (Invitrogen) as per the
manufacturer's instructions. Immunoprecipitation
was used in the protein modification assays and in
the DICER activity assay. DICER was pulled
down using an antibody to DICER (Abcam,
#ab14601).
In vitro DICER activity assay: An in vitro
DICER activity assay was used to test the effect of
smoking on DICER activity (28). Briefly, whole
cell protein extracts were adjusted to 1 mg/ml and
25 ul of the lysate incubated with 125 pmoles of a
27mer RNA duplex DICER substrate (DsiRNAs,
IDT) and incubated at 37oC for 8 hours. To
evaluate the conversion of 27mer dsRNA substrate
to 21mer dsRNA product, 2.5 ul of extract post
incubation was resolved on a 3.5-20% non-
denaturing PAGE gel and visualized with SYBR
green fluorescent dye. DICER activity was
calculated based on the percentage of 27mer
converted to a 21mer RNA strand. The fluorescent
image from a BioRad VersaDoc Imaging System
was converted to a tiff file and analyzed for pixels
per band using Image J, a public domain Java
image-processing program inspired by NIH Image.
In vivo DICER activity assay: To measure
Dicer activity within cells, vectors were made
expressing pre-miRNAs for two of the highly
smoking-altered miRNAs (miR-146b and miR-
708). Hela cells were transfected with the pre-
miRNA constructs and expression allowed to
proceed overnight. Cells were then exposed to
CSE for a short time (1 and 3 hours, chosen to
show Dicer altered but not depleted) and RNA
harvested. Generation of mature miRs was
determined by both qRT-PCR and Northern
analysis.
Pre-miRNA cloning: Primers for the H1
promoter that included sequence for the pre-miR
of interest were obtained from Integrated DNA
Technologies (IDT, Coralville, Iowa). These
included: H1 promoter forward primer, 5'-
ccatggaattcgaacgctgacg-3' and reverse primers for
either miR-146b 5'-aaaactcgagaaaaaacagaactgagt
ccacagggcattgctagagctcacagcctatggaattcagttctcagg
gaaagagtggtctcagacagaac-3' or miR-708, 3'-aaaact
cgagaaaaaactagaagctcacagtctagttgtgttcatgtgcaagtca
tttacccccagctagattgtaagctccttgggaaagagtggtctcagac
agaac-3'. PCR was performed using the H1
plasmid as a substrate (as described in (29)). The
resulting product (H1 promoter followed by the
specific pre-miR) was cloned into a TOPO cloning
vector (pCR 4-TOPO TA cloning vector) using the
manufacturer's protocol. Kanamycin/ampicillin
resistant clones were selected, amplified and DNA
isolated. Selected clones were grown up and
sequenced to verify the insert.
Northern analysis: Isolated RNA was run on
a 12% polyacrylamide/19:1 bis/7M urea/0.5x TBE
gel in 0.5xTBE. The gel was blotted on to
Hybond N+ membrane and probed with a P32
end-
labeled oligo designed to bind both the mature
miR and pre-miRNA sequence (miR-146b:
5'GAGCCTATGGAATTCAGTTCTCA and 3'GC
AGAACTGAGTCCACTGGGCA; miR-708:5'GA
GCCTATGGAATTCAGTTCTCA and 3’GCCAG
AACTGAGTCCACAGGGCA). For both
miR146b and miR-708, the 5' arm of the duplex
produces the major mature product. The 3' probes
were used as a control. Blots were hybridized
overnight, washed and images obtained with x-ray
film.
Ubc9 depletion with siRNA: Control siRNA
(Santa Cruz #44230) or target-specific 19-25 nt
siRNA designed to knock down Ubc9 was
transfected into Hela cells at a final concentration
of 50 nM using Lipofectamine 2000 (Invitrogen).
Transfection mixtures were assembled as
previously described (30). Cells were incubated
with 200 μl of transfection mixture and 2 ml of
antibiotic-free medium for 16-18 h. Transfection
efficiency was assessed with using a fluorescent
control siRNA (Santa Cruz #36869). Knock down
of Ubc9 was confirmed with Western Blotting. Glutathione assay: A glutathione assay was
performed on whole cell lysates using GSH-Glo
Glutathione Assay from Promega (Cat# V6911)
according to manufacturer's instructions. Statistical analysis: Unpaired Student t-test
and one-way ANOVA were used to determine
significance between experimental groups. Data
are presented as means ± SEM. The program used
for data analysis was GraphPad Prism 5.00 (San
Diego, CA).
by guest on October 12, 2019
http://ww
w.jbc.org/
Dow
nloaded from

Alterations in DICER and miRNA levels with cigarette smoke
5
RESULTS
Alveolar macrophages from chronic active
cigarette smokers demonstrate decreased mature
miRNA transcript expression when compared to
cells from nonsmokers. Our lab has previously
shown that smoker’s alveolar macrophages are
characterized by a decrease in expression of a
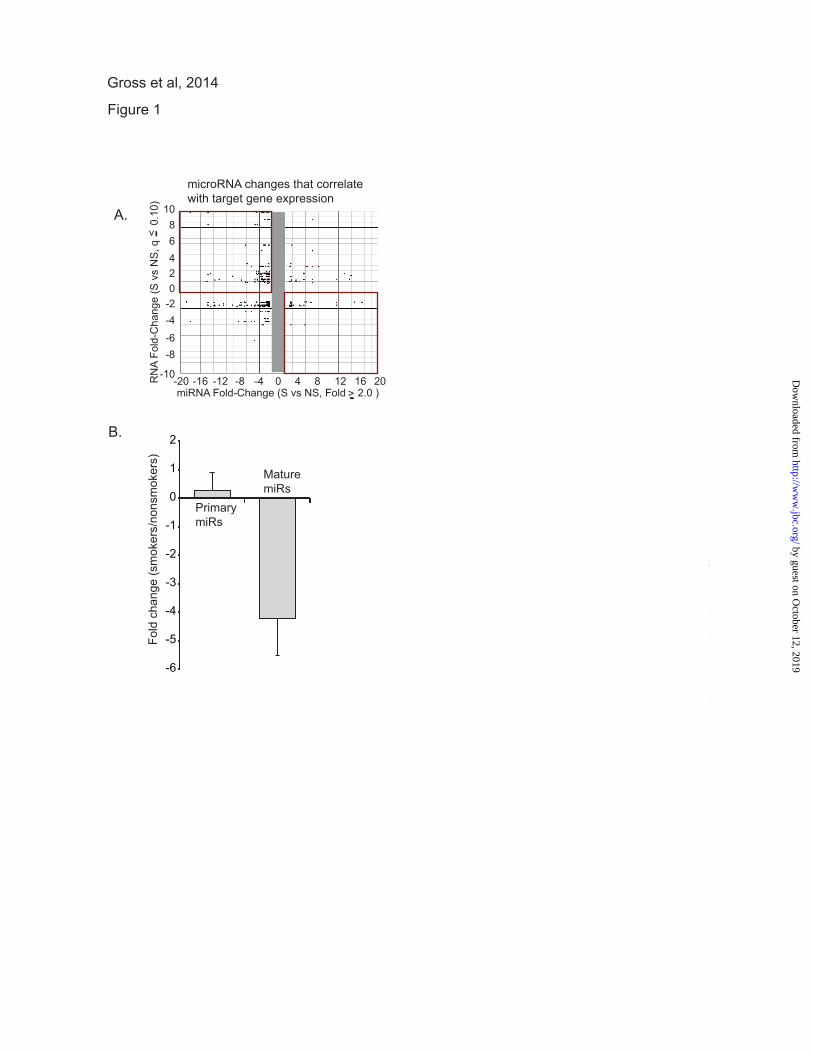
wide variety of miRNAs (10). In Figure 1A, we
show an analysis of the changes in miRNA that
are linked to mRNA changes between nonsmoker
and smoker alveolar macrophages These values
were extracted from the larger data set reported in
reference (10). The data for this figure was
generated from lists of altered miRNAs (> 2 fold
change) and their potential targets (determined by
Target Scan and TarBase) and significantly altered
mRNAs (q≤ 0.10) (Gene Chip Human Exon Array
1.0 ST arrays). The q value (false discovery rate)
for the mRNAs was calculated using the Partek
GS implementation of the step-up method. Gene
IDs common to both lists (miRNA targets and
mRNAs) were plotted. Each dot represents a
gene/miRNA pair as predicted by Target Scan
and/or TarBase. Of particular note are the gene/miRNA pairs
identified in the upper left and lower right
quadrants. The upper left quadrant (more mRNA
and less linked miRNA) and lower right (less
mRNA and more linked miRNA) quadrants
demonstrate multiple reciprocal relationships
between miRNA changes and mRNA changes
with smoking. Some of the genes included in the
upper left quadrant of this figure include: MMP12,
a matrix metalloproteinase linked to emphysema;
SPRY2, a possible regulator of the DICER
complex; SPARC, a regulator of extracellular
matrix; and gamma glutamylcysteine synthetase,
the first rate limiting enzyme in glutathione
synthesis. The data were significant both in the
consistency of the changes caused by smoking and
by the presence of smoking-linked changes in
genes involved in disease, i.e. MMP12, other
matrix-related genes, inflammatory genes and
angiogenesis factors (4,5,31-35). Thus, the
decrease in miRNA expression appears to have
consequences in mRNA levels and, likely, protein
expression. In order to dissect the mechanism underlying
this significant decrease in expression of miRNAs,
we selected eight miRNAs that were significantly
down regulated in our published data set (miR-
708, miR-200a, miR-210, miR-187, miR-149,
miR-429, miR-146b-3p, miR-200c) (10). Using
alveolar macrophages from four nonsmoking
control subjects and four active smokers, we
performed qRT-PCR for both mature miRNA
levels and primary transcripts. Primary miRNA
transcripts are transcribed from the genome and
include flanking regions and one or more stem
loop structures; a difference in primary miRNA
levels would imply a change in nuclear miRNA
transcription rate or alteration in primary miRNA
stability. Our assay for the mature transcript
measured only the 21/22 nucleotide miRNA; a
change in levels of only the mature miRNAs
would imply alterations in processing or stability.
Figure 1B demonstrates no decrease in the amount
of primary miRNA transcripts in smoker alveolar
macrophages. If anything, there may have been a
subtle accumulation as has been previously
reported with inhibition of DICER (36). In
contrast, consistent with our prior observations,
there was a significant comparative decrease in the
level of the mature miRNA transcripts.
Cigarette smoke exposure reduces DICER
complex activity. Because cigarette smoke-induced
alterations in miRNA levels occurred somewhere
between nuclear transcription and loading onto the
RISC-complex, we initiated studies to explore
miRNA processing activity. As these studies
required quantities of cellular protein difficult to
consistently isolate from the number of
macrophages retrieved from nonsmokers, we used
the human monocytic leukemia cell line, THP-1,
as an in vitro model. In this bioactivity assay,
FITC-labeled RNA oligomers (27mer) are
incubated with aliquots of whole cell lysates.
DICER-like activity is detected by demonstrating
cleavage of the oligomers to 21mer strands with a
band shift on gel electrophoresis. Figure 2A is a
methods development assay, showing a dose
response in whole cell lysates ability to cleave the
27mer to a 21mer. There is clear cleavage of the
27mer that varies in proportion with the total cell
protein. In Figure 2B, THP-1 cells were cultured
for 6 hours with or without 5% CSE. Whole cell
lysates were collected and an activity assay run
using 10 ug of protein. Shown in Figure 2B is a
representative gel and quantification from three
identical experiments. To more directly attribute
the measured endonuclease activity to DICER,
lysates from untreated and CSE-exposed cells
by guest on October 12, 2019
http://ww
w.jbc.org/
Dow
nloaded from

Alterations in DICER and miRNA levels with cigarette smoke
6
were subject to immunoprecipitation using a
DICER-specific antibody. As a control, parallel
samples went through the immunoprecipitation
protocol with a nonspecific IgG. 10% of the
recovered sample was incubated with a set amount
of the FITC-labeled 27mer. As shown in Figure
2C, we observed a significant decrease in DICER
activity in the IP sample recovered from smoke-
exposed cells (shown on the right is densitometry
from three separate experiments). Cigarette-smoke
exposure reduces DICER-like RNA endonuclease
activity.
To confirm the effect of cigarette smoke
exposure on mi-RNA processing in an in vivo
model, Hela cells were transfected with a construct
expressing either pre-miR-146b or pre-miR-708.
24 hours after transfection, cells were exposed to
CSE for varying times. RNA was isolated and
pre-miRNA processing assessed by both Northern
analysis and qRT-PCR. Figure 3 demonstrates
that CSE exposure leads to a decrease in
processing of pre-miRNAs into their respective
mature transcripts (Figure 3A, miR-146b and
Figure 3B, miR-708). Experiments using the 3'
Northern probes showed extremely low levels of
mature mi-RNAs, consistent with what is known
about the respective miRNAs (data not shown).
Both the Northern blot and qRT-PCR
demonstrated decreased processing of the pre-
miRNAs after CSE exposure. This in vivo assay of
pre-miRNA processing demonstrates that
cigarette-smoke exposure reduces stem-loop
cleavage activity and specific mature miRNA
levels. While we cannot exclude a concurrent
effect at the initial primary miRNA processing
step in the nucleus (e.g. DROSHA), it seems clear
that smoke exposure disrupts extra-nuclear,
terminal miRNA processing.
Cigarette-smoke decreases DICER protein
levels in exposed cells. Given the dramatic
decreases in both the levels of mature miRNA and
DICER activity after smoke exposure, we further
investigated the proteins involved in the terminal
steps of miRNA processing. Newly isolated
human alveolar macrophages retrieved from
nonsmokers were incubated for six hours in cell
culture media supplemented with freshly prepared
CSE at final concentrations ranging from 0 to 5%.
Whole cell lysates were subject to standard
Western analysis using antibodies to human
DICER, PACT, TRBP, and Ago2, with
constitutively expressed actin serving as a
control for equal protein loading. CSE-exposed
alveolar macrophages from nonsmokers
demonstrated a dose dependent change in the
molecular mass of DICER immunoreactive protein
even at very low concentrations of CSE exposure
(Figure 4A). Figure 4B demonstrates that as early
as one hour after in vitro exposure to 2% CSE,
there was a progressive loss of the 217 kD band
and appearance of multiple higher molecular
weight bands. For other proteins involved in the
cytosolic processing of pre-miRNAs, we found no
change in levels of TRBP, Ago2, or actin
demonstrating the DICER changes were not a
nonspecific cytotoxic effect from the smoke
extract. PACT was the only other complex protein
to show a change with CSE exposure. PACT also
demonstrated a relative decrease at the higher
concentrations of CSE, though this was less
pronounced than the effect on DICER (Figure 4C).
Interestingly, PACT also manifests a shift to a
higher molecular mass form after smoke exposure
similar to that seen in DICER (Figure 4C).
Because the effect of CSE on Dicer was more
pronounced that the changes in PACT, subsequent
experiments focused on DICER. Along with the
effect of CSE on DICER protein in alveolar
macrophages, similar decreases in native sized
DICER (271 kD) and the appearance of higher
molecular weight immunoreactive DICER forms
were also found in cell lysates from THP-1 and
Hela cells (Figure 4D). As the Western Blots in all
of Figure 4 were run under standard reducing
conditions, the shift to a higher molecular mass
DICER was not due to protein-protein
aggregation, but more likely a smoke exposure-
induced post-translational modification of the
DICER protein.
To assess the effect of chronic in vivo
cigarette smoke exposure on DICER protein,
whole cell lysates of alveolar macrophages were
obtained from active smokers and nonsmokers and
analyzed for DICER expression. Western Blot
analysis shown in Figures 5A & 5B confirms that
freshly isolated alveolar macrophages from active
smokers contain significantly less DICER protein
than nonsmoker’s cells. The higher molecular
weight DICER bands were also found in cells
from chronic smokers, although at seemingly
lower levels than after in vitro exposure (Figures 3
vs 4B). Expression array analysis of freshly
by guest on October 12, 2019
http://ww
w.jbc.org/
Dow
nloaded from

Alterations in DICER and miRNA levels with cigarette smoke
7
isolated alveolar macrophages showed no
difference in DICER mRNA levels between
smokers and nonsmokers confirming that smoke-
induced reductions in DICER gene transcription
were not a major contributor to our observations
(Figure 5C) Thus, cigarette smoke exposure
results in a broad decrease in mature miRNA
transcripts. This is associated with a post-
translational modification resulting in functional,
quantitative and qualitative changes in the DICER
protein.
Cigarette-smoke exposure induces post-
translational SUMOylation of DICER. The
dramatic change in apparent molecular mass on
reducing gel analysis suggested a post-
translational modification. Previously work from
out lab identified a block in the autophagy
pathway of smoke-exposed alveolar macrophages
with accumulation of high-molecular mass protein
complexes targeted for degradation. These
complexes contained both ubiquitin and SUMO
2/3 polymers. This directed us to explore whether
this high molecular mass DICER also contained
ubiquitin or SUMO 2/3 adducts.
Freshly isolated alveolar macrophages were
held in culture with or without media
supplemented with CSE. Equal amounts of protein
from whole cell lysates were subjected to
immunoprecipitation with anti-DICER antibody or
a control IgG. The proteins retrieved were
separated using reducing gel electrophoresis and
Western blots probed with anti-DICER, anti-
SUMO, or anti-ubiquitin antibodies. As seen in
Figure 6A, CSE resulted in a notable reduction in
immunoprecipitable DICER with appearance of
faint higher molecular mass bands; no similar
proteins were retrieved using a nonspecific control
antibody. When anti-DICER immunoprecipitates
were probed with antibody recognizing SUMO
2/3, there was considerable SUMO
immunoreactivity clearly detected within the high
molecular mass bands (Figure 6A). Interestingly,
although there was a reduction in total DICER
protein after CSE exposure, the SUMO 2/3
antibody suggested much of this native DICER
(217 kD) was also SUMOylated. Antibody
recognizing ubiquitin moieties also detected
immunoreactivity in the high molecular mass
DICER forms, albeit at comparatively lower levels
than seen with anti-SUMO 2/3.
DICER contains several candidate lysine
SUMOylation sites (see Supplement 1). We next
asked if blocking a key enzyme in the
SUMOylation cascade would alter the appearance
of HMW DICER forms after cigarette-smoke
exposure. The SUMO conjugating (E2) enzyme
Ubc9 catalyzes the formation of isopeptide bonds
between the C-terminus of SUMO and the amino
group of lysine in the target protein. Hela cells
were treated with varying concentration of
silencing RNA (siRNA) specific to Ubc9 or a
control siRNA and then exposed to CSE. Whole
cell lysates were then extracted and analyzed by
Western blotting. As demonstrated in Figure 6C,
the siRNA specific for Ubc9 reduced levels of
immunoreactive Ubc9 in control and CSE-exposed
cells. CSE exposure reduced levels of
immunoreactive DICER and again resulted in the
appearance of higher molecular mass forms in the
cells not exposed to smoke. Interestingly, though
the Ubc9 siRNA attenuated the CSE-induced
change in molecular mass of DICER; it did not
appear to block the reduction in immunoreactive
native DICER (217 kD). The reason for this
unexpected result is part of ongoing studies in the
lab. We also found that smoke exposure resulted
in minor increases in Ubc9 mass detected by
Western blotting (Figure 6B) that might account
for some SUMOylation change. This finding
certainly does not exclude a concurrent smoke-
induced alteration in Ubc9 activity (54, 55) and/or
effects on function of SUMOlase enzymes.
As a composite, these data show that cigarette
smoke exposure results in post-translation
modification of DICER protein that is due, at least
in part, to changes in SUMOylation.
Cigarette smoke extract induced DICER
modification is blocked by NAC and reproduced
by acrolein. Cigarette smoke contains hundreds of
known toxic chemicals that exist in both vapor and
particulate phases. As our cigarette smoke extract
was generated using an aqueous-based medium,
we knew the target compound was water-soluble.
Figure 7A demonstrates that overnight incubation
does not impact the ability of CSE to induce a shift
to higher molecular mass forms of DICER in
THP-1 cells. The DICER modifying activity is lost
when CSE is dialyzed overnight using membranes
with molecular mass cutoff > 2000 kD. Thus, the
active ingredient(s) in CSE are low molecular
weight, water soluble, and stable over many hours.
by guest on October 12, 2019
http://ww
w.jbc.org/
Dow
nloaded from

Alterations in DICER and miRNA levels with cigarette smoke
8
As many of the toxic compounds generated
by the combustion of tobacco are potent oxidants,
we examined whether DICER could be protected
from CSE effects using the potent cellular
antioxidant, NAC. Cells were incubated with or
without 2 % CSE as per prior experiments and the
media concurrently supplemented with NAC at a
final concentration of 2 mM for 6 hours. Whole
cell lysates were subjected to Western blotting and
probed with anti-DICER antibody. As seen in
Figure 7B, despite the brief incubation period,
NAC supplementation increased intracellular
glutathione levels in both control and CSE
exposed cells. Importantly, NAC treatment
prevented the CSE-mediated decrease in DICER
protein levels and blocked the shift to higher
molecular mass forms.
We next tested whether the DICER protein
protection seen with NAC led to increased DICER
activity. Using the in vivo activity assay shown in
Figure 3, we transfected a pre-miRNA-146b
expressing vector into Hela cells. Cells were
exposed to CSE with and without NAC (added to
the cell cultures 15 minutes before the CSE).
Figure 7C shows that NAC reverses the loss of
pree-miRNA-146b cleavage seen with CSE
(shown is a representative Northern and
densitometry from three separate experiments).
As increasing cellular glutathione levels leads
to protection from many forms of oxidant stress,
we tested two classic candidate molecular species
commonly associated with cigarette smoke,
hydrogen peroxide (H2O2) and acrolein (a pure
aldehyde compound). As shown in Figure 7D,
supplementing the media of THP-1 cells with
concentrations of H2O2 approaching the limits of
cellular viability did not reproduce the effect of
CSE on immunoreactive DICER levels. In
contrast, acrolein at concentrations within the
range seen in cigarette smoke vapor and in the
epithelial lining fluid of active smokers (37-39)
faithfully reproduced both the decrease in DICER
protein levels and the appearance of higher
molecular mass forms seen after CSE exposure.
DISCUSSION
The major conclusions of the current study
are that cigarette smoke exposure alters miRNA
expression in human alveolar macrophages via
disruption of the complex maturation process that
occurs between miRNA transcription and
generation of the mature transcript. In addition we
demonstrate that the RNA endonuclease DICER,
an essential component of the miRNA biogenesis
pathway, is altered by cigarette smoke exposure.
These conclusions are based on the following
experimental evidence. We show that a group of
miRNAs with significantly decreased expression
in smoker’s cells is not decreased at the
transcriptional level. This suggests a defect in the
processing of the miRNA transcript. We
demonstrate that the lysates of smoke-exposed
cells contain less dsRNA endonuclease activity
with both an in vitro and in vivo assay.
Transfecting in expression plasmids for pre-
miRNAs, we demonstrate that smoke exposure
decreases the conversion of over-expressed pre-
miR146b and pre-miR708 to mature miRNA.
In smoker's alveolar macrophages, we found
alterations in DICER, the RNA endonuclease
responsible for the final processing of the miRNA
stem-loop structure. Exposure to cigarette smoke
led to decreased activity and amounts of the
endonuclease DICER. We demonstrated that
cigarette smoke-exposed cells expressed an
immunoreactive DICER of increased molecular
mass (in reducing conditions). This change in
mass appears to be linked to the addition of
SUMO chains to DICER. Finally, we
demonstrated that the smoking-induced
modifications to intracellular DICER were
blocked by incubation with N-acetylcysteine, were
reproduced by exposure to acrolein, and were less
evident with exposure to H2O2; these data
robustly support a role for aldehydes in cigarette
smoke-induced DICER protein modification and
consequent decrease in miRNA expression.
Collectively, this evidence strongly supports the
concept that alveolar macrophage phenotype in
smoker's lungs is determined, in part, by gene
expression changes driven by a DICER-linked
processing defect in the miRNA system (Figure
7E).
There is increasing evidence that cigarette-
smoking impacts pulmonary pathophysiology
through alterations in miRNA expression in the
lung. We have previously shown that miRNAs are
decreased in alveolar macrophages from smokers
(10). In that paper, we found that smoking down
regulated 1/3 of the tested miRNAs. In addition,
we found a number of correlations between down
regulated miRNAs and up regulated mRNAs,
by guest on October 12, 2019
http://ww
w.jbc.org/
Dow
nloaded from

Alterations in DICER and miRNA levels with cigarette smoke
9
including the emphysema relevant gene, MMP12.
Other studies have supported our discovery of
reduced miRNAs in smoker lung cells. Schembri,
et al, demonstrated differences in miRNA
expression in epithelial cells isolated from the
airways of smokers as compared with cells from
never smokers (40). Similarly, the induced sputum
from active smokers contained a different pattern
of miRNA than the sputum from nonsmokers (41).
In both of these studies, a general decrease in
miRNA expression was noted in the smoking
subjects. No clear differences could be identified
when smokers with and without clinical COPD
were compared, though both groups differed from
nonsmokers (41). Our findings in the alveolar
macrophages retrieved from active smokers show
that miRNA down regulation in response to
cigarette smoke is not restricted to the airway
epithelium. This is in line with the findings of
whole lung miRNA profiles from smoke-exposed
rats (42). Furthermore, our findings that both
THP-1 and Hela cells manifest similar DICER
modifications offer the possibility that cigarette
smoke-induced defects in miRNA processing may
be a more universal cellular response.
These changes in miRNA levels are likely not
simply epiphenomena as all four labs have found
reciprocal changes in mRNA coding genes
relevant to smoking-induced lung disease,
including inflammatory cytokine receptors,
nuclear transcription factors, and
metalloproteinases. In this study we extend our
initial work to demonstrate that the decrease in
miRNAs targeting some of these genes of interest
is not due to a change in miRNA transcription or
intranuclear stability, as there was little difference
in the levels of primary miRNA transcripts
between smokers and nonsmokers despite a
dramatic decrease in mature miRNA strands.
Thus, the smoking-induced decrease in miRNA
levels results from changes in miRNA maturation.
The importance of miRNAs in modulating
health and disease is well accepted. However, how
miRNA levels are modulated remains
incompletely defined. The biogenesis of miRNAs
is complex, but involves a process leading from
nuclear transcription through a final cytosolic
maturational processing. A key step in this
maturation is the cleavage of the stem loop
structure by the cytoplasmic RNA endonuclease,
DICER. Our demonstration that DICER activity is
reduced in smoke-exposed cells led us to focus on
this enzyme. The regulation of DICER appears to
be complex with evidence for autophagy-mediated
protein degradation as well as post-transcriptional
repression of DICER mRNA by the miRNA let-7a
(43). Our lab has previously established that
cigarette smoke induces an autophagic block in
alveolar macrophages and we found no change in
DICER mRNA in smoke exposed cells (9,20).
This led us to postulate that cigarette smoke
must induce some novel change in DICER protein
itself. Immunoblotting of cell lysates demonstrated
the appearance of DICER species in Western gels
(in reducing conditions) of higher molecular mass
suggesting a post-translational protein
modification. This was followed at later times by a
decrease in total DICER mass. By over-expressing
pre-miRNAs, we were able to show a defect in
DICER/complex activity and impaired maturation
at time points when higher molecular weight
DICER species were apparent without notable
change in total DICER mass.
Further analysis of DICER protein provided
evidence for low-level ubiquitination that might
lead to decreased activity or proteasome targeting
of DICER. However, a more significant finding
was an increase in SUMO 2/3 staining of
immunoprecipitated DICER (217 KD). The high
molecular weight species generated by cigarette
smoke exposure were blocked with inhibition of
the dedicated SUMO ligase, Ubc9. SUMO protein
adducts are seen in response to a variety of cellular
stresses including oxidant injury, heat shock, and
DNA strand damage (44-46). DICER has ten
lysine moieties that fulfill described SUMO
consensus sequences (Ψ-K-x-D/E where Ψ is a
hydrophobic residue) (47,48) (Supplement #1).
While Ubc9 may modify many proteins as part of
their respective regulatory processes, this
represents the first description of a change in
DICER function due to post-translational
SUMOylation. Further studies are needed to
dissect whether this may be a potential target for
manipulation of miRNA expression.
Cigarette smoke is a complex potpourri of gas
phase chemicals and particulate biotoxins. These
include potent oxidants and reactive aldehydes
(49,50). We found that smoke-induced
quantitative and qualitative changes in cellular
DICER were abrogated by the concurrent
administration of the potent anti-oxidant and
by guest on October 12, 2019
http://ww
w.jbc.org/
Dow
nloaded from

Alterations in DICER and miRNA levels with cigarette smoke
10
aldehyde-neutralizing agent, NAC. In addition, the
smoke effects on DICER were faithfully
reproduced with acrolein exposure, but not
hydrogen peroxide. Thus, acrolein delivered via
cigarette smoke inhalation might reduce cellular
DICER activity leading to alterations in miRNA
levels and, ultimately, gene expression. Such a
model has been described in cultured endothelial
cells where acrolein exposure resulted in decreases
in miRNA and increased expression of their
respective mRNA targets (39). As acrolein is a
byproduct of the incomplete combustion of many
natural and man-made materials, aldehyde-
induced changes in miRNA expression due to
alterations in DICER activity may prove to be a
universal mechanism of inhalational pollutant
toxicity (37).
The accumulation of alveolar macrophages
in the distal air space is a defining characteristic of
several smoking-associated lung diseases. The
alveolar macrophage is poised as an ideal cell to
reflect the toxic effects of inhaled smoke and
situated to respond in ways that may initiate and/or
perpetuate disease. Cigarette smoke has been
shown to have profound effects on gene
expression profiles of many cells including
alveolar macrophages (5,51-53). The mechanisms
behind this response are complex and likely due to
both genetic and epigenetic responses. This study
describes a novel impact of cigarette-smoke
exposure on the miRNA-processing pathway in
alveolar macrophages. Smoking-induced
alterations in miRNA maturation could
significantly regulate changes in the expression of
disease-relevant genes and represents a new target
for potential therapeutic intervention.
BIBLIOGRPAHY
1. Martin, T. R., Raghu, G., Maunder, R. J., and Springmeyer, S. C. (1985) The effects of chronic
bronchitis and chronic air-flow obstruction on lung cell populations recovered by bronchoalveolar
lavage. Am Rev Respir Dis 132, 254-260
2. Janoff, A., and Scherer, J. (1968) Mediators of inflammation in leukocyte lysosomes. IX.
Elastinolytic activity in granules of human polymorphonuclear leukocytes. J Exp Med 128, 1137-
1155
3. Shapiro, S. D., and Ingenito, E. P. (2005) The Pathogenesis of Chronic Obstructive Pulmonary
Disease: Advances in the Past 100 Years. Am. J. Respir. Cell Mol. Biol. 32, 367-372
4. Tetley, T. D. (2002) Macrophages and the pathogenesis of COPD. Chest 121, 156S-159S
5. Shaykhiev, R., Krause, A., Salit, J., Strulovici-Barel, Y., Harvey, B. G., O'Connor, T. P., and
Crystal, R. G. (2009) Smoking-dependent reprogramming of alveolar macrophage polarization:
implication for pathogenesis of chronic obstructive pulmonary disease. J Immunol 183, 2867-
2883
6. Hautamaki, R. D., Kobayashi, D. K., Senior, R. M., and Shapiro, S. D. (1997) Requirement for
macrophage elastase for cigarette smoke-induced emphysema in mice. Science 277, 2002-2004
7. Shapiro, S. D. (1999) The macrophage in chronic obstructive pulmonary disease. Am J Respir
Crit Care Med 160, S29-32
8. Chaudhuri, A. A., So, A. Y., Sinha, N., Gibson, W. S., Taganov, K. D., O'Connell, R. M., and
Baltimore, D. (2011) MiRNA-125b potentiates macrophage activation. Journal of immunology
187, 5062-5068
by guest on October 12, 2019
http://ww
w.jbc.org/
Dow
nloaded from

Alterations in DICER and miRNA levels with cigarette smoke
11
9. Graff, J. W., Dickson, A. M., Clay, G., McCaffrey, A. P., and Wilson, M. E. (2012) Identifying
functional miRNAs in macrophages with polarized phenotypes. J Biol Chem 287, 21816-21825
10. Graff, J. W., Powers, L. S., Dickson, A. M., Kim, J., Reisetter, A. C., Hassan, I. H., Kremens, K.,
Gross, T. J., Wilson, M. E., and Monick, M. M. (2012) Cigarette smoking decreases global
miRNA expression in human alveolar macrophages. PLoS ONE 7, e44066
11. Baek, D., Villen, J., Shin, C., Camargo, F. D., Gygi, S. P., and Bartel, D. P. (2008) The impact of
miRNAs on protein output. Nature 455, 64-71
12. Farh, K. K., Grimson, A., Jan, C., Lewis, B. P., Johnston, W. K., Lim, L. P., Burge, C. B., and
Bartel, D. P. (2005) The widespread impact of mammalian MiRNAs on mRNA repression and
evolution. Science 310, 1817-1821
13. Selbach, M., Schwanhausser, B., Thierfelder, N., Fang, Z., Khanin, R., and Rajewsky, N. (2008)
Widespread changes in protein synthesis induced by miRNAs. Nature 455, 58-63
14. Macrae, I. J., Zhou, K., Li, F., Repic, A., Brooks, A. N., Cande, W. Z., Adams, P. D., and
Doudna, J. A. (2006) Structural basis for double-stranded RNA processing by Dicer. Science 311,
195-198
15. Park, J. E., Heo, I., Tian, Y., Simanshu, D. K., Chang, H., Jee, D., Patel, D. J., and Kim, V. N.
(2011) Dicer recognizes the 5' end of RNA for efficient and accurate processing. Nature 475,
201-205
16. Reisetter, A. C., Stebounova, L. V., Baltrusaitis, J., Powers, L., Gupta, A., Grassian, V. H., and
Monick, M. M. (2011) Induction of inflammasome-dependent pyroptosis by carbon black
nanoparticles. J Biol Chem 286, 21844-21852
17. Monick, M. M., Carter, A. B., Gudmundsson, G., Geist, L. J., and Hunninghake, G. W. (1998)
Changes in PKC isoforms in human alveolar macrophages compared with blood monocytes. Am J
Physiol 275, L389-397
18. Monick, M. M., Powers, L. S., Barrett, C. W., Hinde, S., Ashare, A., Groskreutz, D. J., Nyunoya,
T., Coleman, M., Spitz, D. R., and Hunninghake, G. W. (2008) Constitutive ERK MAPK activity
regulates macrophage ATP production and mitochondrial integrity. J Immunol 180, 7485-7496
19. Monick, M. M., Powers, L. S., Gross, T. J., Flaherty, D. M., Barrett, C. W., and Hunninghake, G.
W. (2006) Active ERK Contributes to Protein Translation by Preventing JNK-Dependent
Inhibition of Protein Phosphatase 1. J Immunol 177, 1636-1645
20. Monick, M. M., Powers, L. S., Walters, K., Lovan, N., Zhang, M., Gerke, A., Hansdottir, S., and
Hunninghake, G. W. (2010) Identification of an autophagy defect in smokers' alveolar
macrophages. J Immunol 185, 5425-5435
21. Nyunoya, T., Monick, M. M., Klingelhutz, A., Yarovinsky, T. O., Cagley, J. R., and
Hunninghake, G. W. (2006) Cigarette smoke induces cellular senescence. Am J Respir Cell Mol
Biol 35, 681-688
22. Nyunoya, T., Monick, M. M., Klingelhutz, A. L., Glaser, H., Cagley, J. R., Brown, C. O.,
Matsumoto, E., Aykin-Burns, N., Spitz, D. R., Oshima, J., and Hunninghake, G. W. (2009)
Cigarette smoke induces cellular senescence via Werner's syndrome protein down-regulation. Am
J Respir Crit Care Med 179, 279-287
23. Blue, M. L., and Janoff, A. (1978) Possible mechanisms of emphysema in cigarette smokers.
Release of elastase from human polymorphonuclear leukocytes by cigarette smoke condensate in
vitro. Am Rev Respir Dis 117, 317-325
24. Panayiotidis, M. I., Stabler, S. P., Allen, R. H., Ahmad, A., and White, C. W. (2004) Cigarette
smoke extract increases S-adenosylmethionine and cystathionine in human lung epithelial-like
(A549) cells. Chem Biol Interact 147, 87-97
25. Carnevali, S., Petruzzelli, S., Longoni, B., Vanacore, R., Barale, R., Cipollini, M., Scatena, F.,
Paggiaro, P., Celi, A., and Giuntini, C. (2003) Cigarette smoke extract induces oxidative stress
and apoptosis in human lung fibroblasts. Am J Physiol Lung Cell Mol Physiol 284, L955-963
by guest on October 12, 2019
http://ww
w.jbc.org/
Dow
nloaded from

Alterations in DICER and miRNA levels with cigarette smoke
12
26. Benjamini Y, Krieger A, and Yekutieli D. (2001) Two staged linear step up FDR controlling
procedure. . Tel Aviv University, and Department of Statistics, Wharton School, University of
Pennsylvania
27. Philibert, R. A., Sears, R. A., Powers, L. S., Nash, E., Bair, T., Gerke, A. K., Hassan, I., Thomas,
C. P., Gross, T. J., and Monick, M. M. (2012) Coordinated DNA methylation and gene
expression changes in smoker alveolar macrophages: specific effects on VEGF receptor 1
expression. J Leukoc Biol 92, 621-631
28. Kim, D. H., Behlke, M. A., Rose, S. D., Chang, M. S., Choi, S., and Rossi, J. J. (2005) Synthetic
dsRNA Dicer substrates enhance RNAi potency and efficacy. Nat Biotechnol 23, 222-226
29. Boudreau, R. L., Martins, I., and Davidson, B. L. (2009) Artificial miRNAs as siRNA shuttles:
improved safety as compared to shRNAs in vitro and in vivo. Mol Ther 17, 169-175
30. Manzel, L. J., Chin, C. L., Behlke, M. A., and Look, D. C. (2009) Regulation of bacteria-induced
intercellular adhesion molecule-1 by CCAAT/enhancer binding proteins. American journal of
respiratory cell and molecular biology 40, 200-210
31. Houghton, A. M., Quintero, P. A., Perkins, D. L., Kobayashi, D. K., Kelley, D. G., Marconcini,
L. A., Mecham, R. P., Senior, R. M., and Shapiro, S. D. (2006) Elastin fragments drive disease
progression in a murine model of emphysema. J Clin Invest 116, 753-759
32. Belaaouaj, A., Shipley, J. M., Kobayashi, D. K., Zimonjic, D. B., Popescu, N., Silverman, G. A.,
and Shapiro, S. D. (1995) Human macrophage metalloelastase. Genomic organization,
chromosomal location, gene linkage, and tissue-specific expression. J Biol Chem 270, 14568-
14575
33. Robbins, C. S., Bauer, C. M., Vujicic, N., Gaschler, G. J., Lichty, B. D., Brown, E. G., and
Stampfli, M. R. (2006) Cigarette smoke impacts immune inflammatory responses to influenza in
mice. Am J Respir Crit Care Med 174, 1342-1351
34. Birrell, M. A., Wong, S., Catley, M. C., and Belvisi, M. G. (2008) Impact of tobacco-smoke on
key signaling pathways in the innate immune response in lung macrophages. J Cell Physiol 214,
27-37
35. Yang, S. R., Chida, A. S., Bauter, M. R., Shafiq, N., Seweryniak, K., Maggirwar, S. B., Kilty, I.,
and Rahman, I. (2006) Cigarette smoke induces proinflammatory cytokine release by activation
of NF-kappaB and posttranslational modifications of histone deacetylase in macrophages. Am J
Physiol Lung Cell Mol Physiol 291, L46-57
36. Koscianska, E., Starega-Roslan, J., and Krzyzosiak, W. J. (2011) The role of Dicer protein
partners in the processing of miRNA precursors. PLoS One 6, e28548
37. Stevens, J. F., and Maier, C. S. (2008) Acrolein: sources, metabolism, and biomolecular
interactions relevant to human health and disease. Molecular nutrition & food research 52, 7-25
38. Eiserich, J. P., van der Vliet, A., Handelman, G. J., Halliwell, B., and Cross, C. E. (1995) Dietary
antioxidants and cigarette smoke-induced biomolecular damage: a complex interaction. The
American journal of clinical nutrition 62, 1490S-1500S
39. Talbot, P., DiCarlantonio, G., Knoll, M., and Gomez, C. (1998) Identification of cigarette smoke
components that alter functioning of hamster (Mesocricetus auratus) oviducts in vitro. Biology of
reproduction 58, 1047-1053
40. Schembri, F., Sridhar, S., Perdomo, C., Gustafson, A. M., Zhang, X., Ergun, A., Lu, J., Liu, G.,
Zhang, X., Bowers, J., Vaziri, C., Ott, K., Sensinger, K., Collins, J. J., Brody, J. S., Getts, R.,
Lenburg, M. E., and Spira, A. (2009) MiRNAs as modulators of smoking-induced gene
expression changes in human airway epithelium. Proc Natl Acad Sci U S A 106, 2319-2324
41. Pottelberge, G. R., Mestdagh, P., Bracke, K. R., Thas, O., Durme, Y. M., Joos, G. F.,
Vandesompele, J., and Brusselle, G. G. (2011) MiRNA expression in induced sputum of smokers
and patients with chronic obstructive pulmonary disease. Am J Respir Crit Care Med 183, 898-
906
by guest on October 12, 2019
http://ww
w.jbc.org/
Dow
nloaded from

Alterations in DICER and miRNA levels with cigarette smoke
13
42. Izzotti, A., Calin, G. A., Arrigo, P., Steele, V. E., Croce, C. M., and De Flora, S. (2009)
Downregulation of miRNA expression in the lungs of rats exposed to cigarette smoke. Faseb J
23, 806-812
43. Gibbings, D., Mostowy, S., Jay, F., Schwab, Y., Cossart, P., and Voinnet, O. (2012) Selective
autophagy degrades DICER and AGO2 and regulates miRNA activity. Nat Cell Biol 14, 1314-
1321
44. Bossis, G., and Melchior, F. (2006) SUMO: regulating the regulator. Cell division 1, 13
45. Bossis, G., and Melchior, F. (2006) Regulation of SUMOylation by reversible oxidation of
SUMO conjugating enzymes. Molecular cell 21, 349-357
46. Tempe, D., Piechaczyk, M., and Bossis, G. (2008) SUMO under stress. Biochem Soc Trans 36,
874-878
47. Teng, S., Luo, H., and Wang, L. (2012) Predicting protein sumoylation sites from sequence
features. Amino acids 43, 447-455
48. Sampson, D. A., Wang, M., and Matunis, M. J. (2001) The small ubiquitin-like modifier-1
(SUMO-1) consensus sequence mediates Ubc9 binding and is essential for SUMO-1
modification. J Biol Chem 276, 21664-21669
49. Pryor, W. A., and Stone, K. (1993) Oxidants in cigarette smoke. Radicals, hydrogen peroxide,
peroxynitrate, and peroxynitrite. Ann N Y Acad Sci 686, 12-27; discussion 27-18
50. Facchinetti, F., Amadei, F., Geppetti, P., Tarantini, F., Di Serio, C., Dragotto, A., Gigli, P. M.,
Catinella, S., Civelli, M., and Patacchini, R. (2007) Alpha,beta-unsaturated aldehydes in cigarette
smoke release inflammatory mediators from human macrophages. Am J Respir Cell Mol Biol 37,
617-623
51. Spira, A., Beane, J., Shah, V., Liu, G., Schembri, F., Yang, X., Palma, J., and Brody, J. S. (2004)
Effects of cigarette smoke on the human airway epithelial cell transcriptome. Proc Natl Acad Sci
U S A 101, 10143-10148
52. Spira, A., Schembri, F., Beane, J., Shah, V., Liu, G., and Brody, J. S. (2004) Impact of cigarette
smoke on the normal airway transcriptome. Chest 125, 115S
53. Zeskind, J. E., Lenburg, M. E., and Spira, A. (2008) Translating the COPD transcriptome:
insights into pathogenesis and tools for clinical management. Proc Am Thorac Soc 5, 834-841
Acknowledgements. We thank Jiselle Vertinez and Richard Starr for technical assistance.
FOOTNOTES
*This project was supported by NIH R01 HL079901, NIH RO1 HL096625 and R21HL109589 to M. M.
This research was funded in part by the National Institute for Environmental Health Sciences through the
University of Iowa Environmental Health Sciences Research Center, NIEHS/NIH P30 ES005605 and
Grant Number UL1RR024979 from the National Center for Research Resources (NCRR), a part of the
National Institutes of Health (NIH).
1. To whom correspondence should be addressed: Thomas J. Gross, Division of Pulmonary, Critical Care,
and Occupational Medicine, University of Iowa, Iowa City, IA 52242, Phone: (319) 356-6268, Email:
[email protected] 2. Department of Medicine
3. The abbreviations used are: COPD, Chronic Obstructive Pulmonary Disease; CSE, cigarette smoke
extract; SUMO, Small Ubiquitin-like Modifier; siRNA, small interfering RNA; NAC, N-acetyl cysteine
FIGURE LEGENDS
Figure 1. In vivo cigarette smoke exposure alters miRNA profiles in alveolar macrophages. A.
Differentially expressed miRNAs in smoker's alveolar macrophages are down regulated and link to up
regulated mRNAs. Alveolar macrophages were isolated from 4 active smokers (> 10 pack years) and 4
by guest on October 12, 2019
http://ww
w.jbc.org/
Dow
nloaded from

Alterations in DICER and miRNA levels with cigarette smoke
14
never smokers. An analysis of mRNA expression was performed using Human Exon Array 1.0 ST arrays
from Affymetrix. MiRNA expression was analyzed using TaqMan Low Density Arrays version 2.0
(ABI). Altered miRNAs (> 2 fold change) were analyzed for potential mRNA targets using TargetScan
and TarBase. The miRNA target information was compared to significantly altered mRNAs (q≤ 0.10).
The q value (false discovery rate) for the mRNAs was calculated using the Partek GS implementation of
the step-up method. Gene IDs common to both lists (miRNA targets and mRNAs) were plotted. Each
dot represents a gene/miRNA pair as predicted by Target Scan and/or TarBase. The upper right quadrant
is of particular interest as it identifies down regulated miRNAs that link to up regulated mRNAs. B.
MiRNAs that are down regulated in smoker's alveolar macrophages have intact transcriptional profiles. A
panel of down regulated miRNAs (miR-708, miR-200a, miR-210, miR-187, miR-149, miR-429, miR-
146b-3p, miR-200c) was tested in RNA isolated from smoker and never smoker alveolar macrophages.
Each miRNA was tested using primers specific for the mature miRNA and primers specific for the
primary miRNA transcript (the stem loop structure with accompanying 5' and 3' ends). The data is a
composite of results from 4 smokers and 4 never smokers with significance determined by Student's t test.
Figure 2. Cigarette smoke exposure decreases dsRNA endonuclease activity. To test the effect of
cigarette smoke on DICER activity, we used an in vitro assay of endonuclease cleavage of a fluorescent
27mer target. A. To validate the assay, varying amounts of whole cell lysate protein (THP-1 cells) were
incubated with the FITC-27mer. Shown is a gel demonstrating a dose dependent cleavage of the 27mer
target dsRNA. B. Whole cell lysates from Control THP-1 cells and CSE exposed THP-1 cells (5% CSE
for 6 hours) were incubated with the FITC-27mer. Shown is a fluorescent image of the DNA gel and
densitometry from three separate experiments. Significance was determined using Student's t test. C.
DICER was immunoprecipitated from 500 ug THP-1 protein (control and CSE exposed, 5% for 6 hours).
20% of the resulting protein coated beads (pulled down with an anti-DICER antibody or a nonspecific
IgG) were mixed with the FITC-27mer to assay for DICER-specific ribonuclease activity. Shown is the
fluorescent image of a DNA gel showing both the original 27mer and the cleaved 21mer product.
Densitometry is from three separate experiments.
Figure 3. Cigarette smoke exposure decreases pre-miRNA processing activity. Two highly CSE
regulated miRNAs were chosen for an in vivo pre-miRNA processing. A+B. Transfection with either
pre-miR-146b (panel A) or pre-miR-708 (panel B) and processing of the respective miRNAs was
evaluated by Northern blot and qRT-PCR. Hela cells were transfected with a plasmid expressing pre-
miRNAs under control of the constitutive H1 promoter. 24 hours after transfection, cells were exposed to
1 or 3 hours of CSE and RNA isolated. Northern analysis was performed to demonstrate the relative
degree of pre-miRNA processing. qRT-PCR to compare pre-miRNA to mature miRNA levels was also
performed. All assays were done in triplicate and statistical analysis for both the densitometry data and
qRT-PCR performed (Student's ttest).
Figure 4. DICER protein is modified by cigarette smoke exposure. A. Human alveolar macrophages
freshly isolated from nonsmokers were exposed to varying concentrations of CSE for 6 hours (0.5% to
5.0%). Whole cell lysates were obtained and Western analysis performed for DICER, PACT, TRBP, and
AGO2. Equal loading of the gel was determined by staining for actin. B. Normal human alveolar
macrophages were exposed to CSE (2%) for varying times (1 to 6 hours) Whole cell lysates were
obtained and Western analysis performed for DICER. C. Normal human alveolar macrophages were
exposed to CSE at varying concentrations and whole cell lysates were run on a 7.5 % PAGE SDS gel to
optimize transfer of high molecular weight proteins. PACT protein levels were determined by Western
analysis. D. THP-1 cells were exposed to varying concentrations of CSE for 6 hours; Hela cells were
exposed to CSE for 3 and 6-hour time points. Whole cell lysates were obtained and Western analysis
performed for DICER.
by guest on October 12, 2019
http://ww
w.jbc.org/
Dow
nloaded from

Alterations in DICER and miRNA levels with cigarette smoke
15
Figure 5. Smoking alters alveolar macrophage DICER protein and not DICER mRNA. A. Human
alveolar macrophages were obtained from active smokers and never smokers. Whole cell lysates were
obtained immediately (baseline sample) and Western analysis was performed for DICER. Shown on the
right is densitometry. Significance was determined using Student's t test. B. Human alveolar macrophages
were obtained from active smokers and never smokers. Fresh whole cell lysates were immediately
analyzed by Western blotting and probed for DICER. The gel was optimized to maximize visualization of
the high molecular weight bands (5% gel with a long transfer). C. Total RNA was obtained from freshly
isolated human alveolar macrophages (43 nonsmoker subjects and 42 smoker subjects). The mRNA levels
for DICER were analyzed using an Affymetrix array (Gene Chip Human Exon Array 1.0 ST arrays).
Shown are the log2 values for DICER, as determined by Partek analysis of the raw data. There is no
difference in DICER mRNA levels between smokers and nonsmokers.
Figure 6. Cigarette smoke exposure increases conjugation of SUMO chains to the protein DICER.
A. Alveolar macrophages from nonsmokers were exposed to 2% CSE in vitro for 6 hours. Whole cell
lysates were isolated and 500 ug of protein was used for immunoprecipitation using an anti-DICER
antibody. The resulting samples were divided into thirds and Western analysis performed for DICER,
SUMO 2/3 and ubiquitin. As a control, 500 ug of whole cell lysate protein was also subjected to
immunoprecipitation with a nonspecific IgG. B. Human alveolar macrophages were exposed to varying
concentrations of CSE. Western analysis was performed for Ubc9. C. Hela cells were incubated with
Ubc9-specific or a control siRNA. The cells were then exposed to 5% CSE for 6 hours and whole cell
protein obtained. Western analysis was performed for DICER, Ubc9 (verifying knockdown) and actin.
Figure 7. DICER modification by cigarette smoke is inhibited by N-acetylcysteine. A. To explore
the chemical properties of the DICER-active component of cigarette smoke, CSE was dialyzed overnight
in tubing with cutoffs of 2 kD or 10 kD. THP-1 cells were then exposed to freshly prepared CSE, CSE
that sat in the refrigerator over night (control for dialysis) or the dialyzed CSE for 6 hours. Western
analysis for DICER was performed on whole cell lysates. B. THP-1 cells were exposed to CSE alone
(2%) or CSE in the presence of NAC (2 mM) for 6 hours. Duplicate assays were set up. One set of cells
was utilized for glutathione assays and the other set of cells were lysed and whole cell protein obtained
for Western analysis for DICER. C. Hela cells were transfected with a plasmid that expresses pre-
miRNA-146b under control of the constitutive H1 promoter. 24 hours following transfection cells were
exposed to CSE (10%) with and without added NAC (2 mM) for three hours. RNA was isolated and
Northern analysis for miRNA-146b performed. Shown is a representative blot and densitometry from
three separate experiments. D. THP-1 cells were exposed to varying concentrations of H2O2 or acrolein
for 6 hours. Whole cell lysates were obtained and Western analysis for DICER performed. E. This
diagram represents an overview of the conclusions supported by the study data.
by guest on October 12, 2019
http://ww
w.jbc.org/
Dow
nloaded from
Gross et al, 2014
-6
-5
-4
-3
-2
-1
0
1
2
Fold
cha
nge
(sm
oker
s/no
nsm
oker
s)
PrimarymiRs
MaturemiRs
Figure 1
A.
B.
1086420
-2-4-6-8
-10-20 -16 -12 -8 -4 0 4 8 12 16 20
microRNA changes that correlate with target gene expression
RN
A Fo
ld-C
hang
e (S
vs
NS
, q
0.1
0)<
miRNA Fold-Change (S vs NS, Fold 2.0 )<
by guest on October 12, 2019
http://ww
w.jbc.org/
Dow
nloaded from

Cell lysate (ug)Assay Time (h)
258
108
58
08
Gross et al, 2014
Figure 2
A.
B.
27mer
21mer
Olig
o (D
siR
NA
)
Con
trol
5% C
SE
Con
trol
5% C
SE
IP aDicer IP IgG
C.
Con
trol
CS
E
27mer
21mer
27mer21mer
Non
e
Dic
er A
b
Dic
er A
b
Control CSE
Con
trol I
gG
Con
trol I
gG
Den
sito
met
ry o
f 21m
er(A
rbitr
ary
untis
)
Den
sito
met
ry o
f 21m
er(A
rbitr
ary
num
bers
)
IP antibody
p< 0.01
0
2000
4000
6000
8000
Oligo Con CSE Con CSE0
5001000150020002500300035004000 p<0.01
by guest on October 12, 2019
http://ww
w.jbc.org/
Dow
nloaded from

Con
trol
No
trans
fect
ion
CS
E 1
h
CS
E 3
h
Con
trol
No
trans
fect
ion
CS
E 1
h
CS
E 3
h
30nt20nt
50nt60nt
20nt
70nt
Pre miR-146b
miR-146b
Pre miR-708
miR-708
Gross et al, 2014
Figure 3.
5’UGAGAACUGAAUUCCAUAGGCUG
3’GG-UCUUGACU-CAGGUGUCCCGU
UGAGCUCUAGCAA
Pre miR-146b (58nt)mature
passenger
Pre miR-708 (68nt)mature
passenger
5’AAGGAGCUUACAAUCUAGCUGGGG
3’GAUCUUCGAGUGUCAGAUCAAC--A
GUAAAUGACUU
GCACAUGAAC
A
Control CSE 1h CSE 3h
Nor
ther
n de
nsito
met
ry, R
atio
(m
atur
e m
iR-1
46b/
pre
miR
-146
b)N
orth
ern
dens
itom
etry
, Rat
io
(mat
ure
miR
-708
/pre
miR
-708
)
miR
-708
RN
A (m
iR-7
08/R
NU
48)
miR
-146
b R
NA
(miR
-146
b/R
NU
48)
A.
B.
Control CSE 1h CSE 3h Control CSE 1h CSE 3h
Control CSE 1h CSE 3h
0
2
4
6
8
10
12
14
****
** p<0.01** p<0.01
**
**
p<0.01*
*
p<0.05 *
*
p<0.05
0
0.1
0.2
0.3
0.4
0.5
0.6
0.000.050.100.150.200.250.300.350.400.45
0
5
10
15
20
25
30
by guest on October 12, 2019
http://ww
w.jbc.org/
Dow
nloaded from

Gross et al, 2014
Figure 4.
220 kD
mol wtmarker
DICER, 217 kD
Con
trol
CS
E 1
h
CS
E 3
h
CS
E 6
h
hmw DICER
β Actin, 42 kD
A. B.
C. D.
Human alveolar macrophages
Human alveolar macrophages
Con
trol
0.5
% C
SE
2.0
% C
SE
1.0
% C
SE
5.0
% C
SE
PACT, 34 kD
DICER, 217 kD
TRBP, 39 kD
AGO2, 97 kD
hmw DICER
β actin, 42 kD
β actin, 42 kD
Con
trol
2.5%
CS
E
5.0%
CS
E
7.5%
CS
E
10%
CS
E
DICER, 217 kDhmw DICER
THP1 Cells (human acute monocytic leukemia cell line)
Con
trol
CS
E 0
.5%
CS
E 1
.0%
CS
E 2
.0%
CS
E 5
.0%
Hela cells (human cell line derived from cervical cancer)
DICER, 217 kDhmw DICER
β actin, 42 kD
Con
trol
0.5
% C
SE
2.0
% C
SE
1.0
% C
SE
5.0
% C
SE
PACT, 34 kD
hmw PACT
Human alveolar macrophagesβ actin, 42 kD
Con
trol
CS
E 1
0% 3
h
by guest on October 12, 2019
http://ww
w.jbc.org/
Dow
nloaded from

Gross et al, 2014
Figure 5.
Non
smok
er 1
Non
smok
er 2
Non
smok
er 1
Non
smok
er 2
Non
smok
er 3
Non
smok
er 4
Sm
oker
1
Sm
oker
2
Sm
oker
1
Sm
oker
2
Sm
oker
3
Sm
oker
4
Pk yrs
Pk yrs
0 0 0 0 25 14 12 32
DICER, 217 kD
β Actin, 42 kD
220
mw
0
500
1000
1500
2000
2500
3000
3500
4000
Nonsmokers Smokers
Den
sito
met
ry (a
rbitr
ary
units
)
p<0.05
DICER, 217 kD
hmw DICER
β Actin, 42 kD
A.
B. C.
Human alveolar macrophages
0 0 10 20
Human alveolar macrophages Nonsmokers (n=43) Smokers (n=42
Dic
er m
RN
A (lo
g2 v
alue
s)9.0
8.5
8.0
7.5
7.0
6.5
by guest on October 12, 2019
http://ww
w.jbc.org/
Dow
nloaded from

Gross et al, 2014
Figure 6.
B.
C.
A.
Con CSE Con CSE Con CSE Con CSE
anti-Dicer
Ubc9, 18kD
anti-Sumo 2/3 anti-Ub
Pull down
DICER, 217 kD
DICER Ab DICER Ab DICER AbControl IgG
hmw DICER
Western Ab
DICER, 217 kD
Ubc9, 18 kD
siRNA amts
Ubc9 siR
NA
con siR
NA
CSE + Ubc9 siR
NA
CSE + con siR
NA
hmw DICER
β actin, 42 kD
β actin, 42 kD
Con
trol
0.5
% C
SE
2.0
% C
SE
1.0
% C
SE
Human alveolar macrophages
Human alveolar macrophages
by guest on October 12, 2019
http://ww
w.jbc.org/
Dow
nloaded from

Con
trol
5% fr
esh
CS
E
5% 2
K d
ialy
zed
CS
E
5% 1
0K d
ialy
zed
CS
E
5% C
SE
(4o
over
nigh
t)DICER 217 kD
hmw DICER
β actin, 42 kD
β actin, 42 kD
β actin, 42 kD
DICER, 217 kD
Con
trol
NA
C
CS
E
CS
E +
NA
C
hmw DICER
β actin, 42 kD
Gross et al, 2014
Figure 7
A.
B.
C.
D. E.
Con
trol
5% C
SE
50 u
M a
crol
ein
100
uM a
crol
ein
150
uM a
crol
ein
250
uM a
crol
ein
Con
trol
5% C
SE
100
uM H
2O2
250
uM H
2O2
500
uM H
2O2
1 m
M H
2O2
DICER, 217 kDhmw DICER
DICER, 217 kDhmw DICER
0
400
800
1200
1600
2000
ControlNACCSE
NAC + CSE
Glu
tath
ione
(uM
)
Cigarette smoke
Aldehydes/oxidants
Dicer modification
Large scale decreasein mature microRNAs
Increase in selected gene expression
MicroRNA processing defect
Con
trol
No
trans
fect
ion
CS
E 3
h
NA
C+C
SE
NA
C
Pre miR-146b
miR-146b
Control
Den
sito
met
ryR
atio
(miR
-146
b/P
re m
iR-1
46b)
NAC CSE Both0.00
0.05
0.10
0.15
0.20
0.25
*
p<0.05
by guest on October 12, 2019
http://ww
w.jbc.org/
Dow
nloaded from

Khushboo Goel, Alicia K. Gerke, Ihab H. Hassan and Martha M. MonickThomas J. Gross, Linda S. Powers, Ryan L. Boudreau, Brandi Brink, Anna Reisetter,
the endonuclease DICERA microRNA processing defect in smokers' macrophages is linked to SUMOylation of
published online March 25, 2014J. Biol. Chem.
10.1074/jbc.M114.565473Access the most updated version of this article at doi:
Alerts:
When a correction for this article is posted•
When this article is cited•
to choose from all of JBC's e-mail alertsClick here
Supplemental material:
http://www.jbc.org/content/suppl/2014/03/25/M114.565473.DC1
by guest on October 12, 2019
http://ww
w.jbc.org/
Dow
nloaded from