Embed Size (px)
Citation preview

Jejunal Perfusion of Simple and Conjugated Folates
in Tropical Sprue
JosE J. CORCINO, ANNM. REISENAUER, and CHARLEsH. HALSTED
From the Departments of Medicine, University of Puerto Rico School ofMedicine, San Juan, Puerto Rico 00936 and University of California,Davis, California 95616
A B S T R A C T Absorption of labeled simple 3',5',9'-Hpteroylmonoglutamate, (1[H]PG-1) and conjugatedpteroyl-4u ["C] glutamyl-Y-hexaglutamate, ( [I4C] PG-7),fo-lates was assessed in six patients with tropical sprue,before and after 6 mo of treatment, utilizing jejunalperfusion and urinary recovery techniques. Degradationproducts of [14C]PG-7 which were produced during per-fusion were identified by DEAE-cellulose column chro-matography. Jejunal mucosal activities of folate con-jugase, lactase, sucrase, and maltase were measured inevery patient. Malabsorption of both [8H] PG-1 and[14C]PG-7 was found in every untreated patient, withsignificant improvement after therapy. The urinary ex-cretion of 'H and 14C paralleled the luminal disappear-ance of both isotopes. The chromatographic patterns ofintraluminal degradation products of [14C]PG-7 obtainedduring perfusion did not differ from those previouslyfound in normal subjects and were similar in studiesperformed before and after treatment. The activity offolate conjugase was increased in the mucosa of the un-treated patients when compared to the post-treatmentlevels while the activities of mucosal lactase, sucrase,and maltase were originally low and increased signifi-
This work was presented in part at the National Meetingof the American Federation for Clinical Research, 1975,(Clin. Res. 23: 250A.). This work was reported at theAnnual Meeting of the American Society for Clinical Nu-trition, 1975, (Am. J. Clin. Nutr. 28: 418.). Part of thiswork was published in abstract form in 1975, (Gastroenter-ology. 68: 908.). This work was reported at the Xth Inter-national Congress of Nutrition in Kyoto, Japan in 1975,p. 41 (Abstr.)
Dr. Corcino is the recipient of an Academic Career De-velopment Award no. 5 K 07-AM 70696-02. Dr. Halstedis the recipient of Award 465 from the Nutrition Founda-tion, New York.
Received for publication 27 September 1975 and in revisedform 15 March 1976.
cantly after therapy. These observations suggest thatfolate conjugase originates at a different mucosal locusthan the brush border disaccharidases, and are consistentwith previous evidence that folate conjugase is an intra-cellular enzyme. The present studies have demonstratedunequivocal malabsorption of both simple and conjugatedfolates in tropical sprue. In tropical sprue, folate malab-sorption is the reflection of impaired folate transportand not of impaired hydrolysis.
INTRODUCTION
Folate deficiency is almost always present in PuertoRican patients with tropical sprue (1) and is probablya reflection of the generalized intestinal malabsorptionwhich characterizes this syndrome (2). Although morethan 85% of the dietary folates are present as conju-gated pteroylpolyglutamates (3), scant attention hasbeen paid to the assessment of the absorption of thesefolates in tropical sprue. The intestinal mucosa is a po-tent source of the -v-carboxypeptidase known as folateconjugase which is required for the cleavage of they-peptide linkages present in conjugated folates (4, 5).In vivo studies in the dog and the human have shownthat cleavage of pteroylpolyglutamates to pteroylmono-glutamate (simple folate) occurs during or soon afterintestinal absorption (4-6). Thus, the malabsorption ofconjugated folates may result from decreased mucosalhydrolysis, decreased intestinal transport of pteroyl-monoglutamate, or a combination of these factors. Therecent development of more refined techniques to deter-mine folate conjugase activity in intestinal mucosa (7)and to measure intestinal luminal disappearance, as wellas luminal degradation products of pure synthetic con-jugated folate (8), prompted us to study the simultane-ous absorption of separately labeled simple and conju-gated folates in six patients with tropical sprue.
The Journal of Clinical Investigation Volume 58 August 1976 298-305298
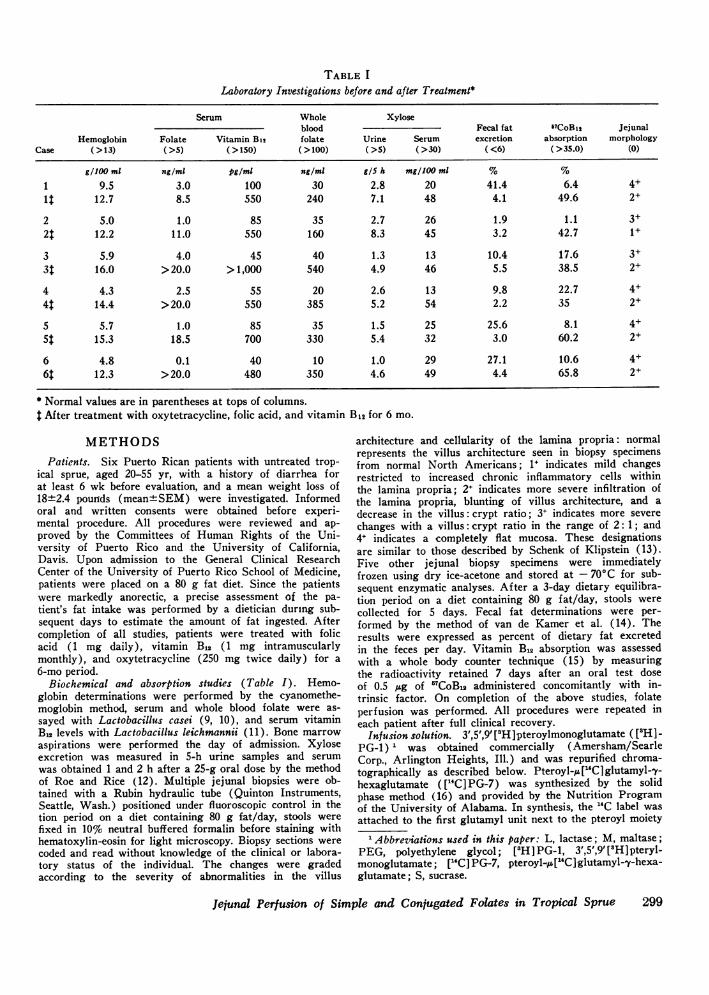
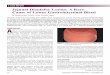
TABLE ILaboratory Investigations before and after Treatment*
Serum Whole Xyloseblood Fecal fat 67CoBi2 Jejunal
Hemoglobin Folate Vitamin B12 folate Urine Serum excretion absorption morphologyCase (>13) (>5) (>150) (>100) (>5) (>30) (<6) (>35.0) (0)
g/100 ml ng/ml pg/ml ng/ml g/S h mg/100 ml % %
1 9.5 3.0 100 30 2.8 20 41.4 6.4 4+it 12.7 8.5 550 240 7.1 48 4.1 49.6 2+
2 5.0 1.0 85 35 2.7 26 1.9 1.1 3+24 12.2 11.0 550 160 8.3 45 3.2 42.7 1+
3 5.9 4.0 45 40 1.3 13 10.4 17.6 3+3t 16.0 > 20.0 > 1,000 540 4.9 46 5.5 38.5 2+
4 4.3 2.5 55 20 2.6 13 9.8 22.7 4+4$ 14.4 >20.0 550 385 5.2 54 2.2 35 2+
5 5.7 1.0 85 35 1.5 25 25.6 8.1 4+5t 15.3 18.5 700 330 5.4 32 3.0 60.2 2+
6 4.8 0.1 40 10 1.0 29 27.1 10.6 4+6t 12.3 >20.0 480 350 4.6 49 4.4 65.8 2+
* Normal values are in parentheses at tops of columns.After treatment with oxytetracycline, folic acid, and vitamin B12 for 6 mo.
METHODSPatients. Six Puerto Rican patients with untreated trop-
ical sprue, aged 20-55 yr, with a history of diarrhea forat least 6 wk before evaluation, and a mean weight loss of18±2.4 pounids (mean+SEM) were investigated. Informedoral and written consents were obtained before experi-mental procedure. All procedures were reviewed and ap-proved by the Committees of Human Rights of the Uni-versity of Puerto Rico and the University of California,Davis. Upon admission to the General Clinical ResearchCenter of the University of Puerto Rico School of Medicine,patients were placed on a 80 g fat diet. Since the patientswere markedly anorectic, a precise assessment of the pa-tient's fat intake was performed by a dietician during sub-sequent days to estimate the amount of fat ingested. Aftercompletion of all studies, patients were treated with folicacid (1 mg daily), vitamin B12 (1 mg intramuscularlymonthly), and oxytetracycline (250 mg twice daily) for a6-mo period.
Biochemical and absorption studies (Table I). Hemo-globin determinations were performed by the cyanomethe-moglobin method, serum and whole blood folate were as-sayed with Lactobacillus casei (9, 10), and serum vitaminB12 levels with Lactobacillus leichmannii (11). Bone marrowaspirations were performed the day of admission. Xyloseexcretion was measured in 5-h urine samples and serumwas obtained 1 and 2 h after a 25-g oral dose by the methodof Roe and Rice (12). Multiple jejunal biopsies were ob-tained with a Rubin hydraulic tube (Quinton Instruments,Seattle, Wash.) positioned under fluoroscopic control in thetion period on a diet containing 80 g fat/day, stools werefixed in 10% neutral buffered formalin before staining withhematoxylin-eosin for light microscopy. Biopsy sections werecoded and read without knowledge of the clinical or labora-tory status of the individual. The changes were gradedaccording to the severity of abnormalities in the villus
architecture and cellularity of the lamina propria: normalrepresents the villus architecture seen in biopsy specimensfrom normal North Americans; 1+ indicates mild changesrestricted to increased chronic inflammatory cells withinthe lamina propria; 2+ indicates more severe infiltration ofthe lamina propria, blunting of villus architecture, and adecrease in the villus: crypt ratio; 3+ indicates more severechanges with a villus : crypt ratio in the range of 2: 1 ; and4+ indicates a completely flat mucosa. These designationsare similar to those described by Schenk of Klipstein (13).Five other jejunal biopsy specimens were immediatelyfrozen using dry ice-acetone and stored at -70°C for sub-sequent enzymatic analyses. After a 3-day dietary equilibra-tion period on a diet containing 80 g fat/day, stools werecollected for 5 days. Fecal fat determinations were per-formed by the method of van de Kamer et al. (14). Theresults were expressed as percent of dietary fat excretedin the feces per day. Vitamin B12 absorption was assessedwith a whole body counter technique (15) by measuringthe radioactivity retained 7 days after an oral test doseof 0.5 ,ug of '7CoBna administered concomitantly with in-trinsic factor. On completion of the above studies, folateperfusion was performed. All procedures were repeated ineach patient after full clinical recovery.
Infusion solution. 3',5',9[3H]pteroylmonoglutamate ([5H] -
PG-1) 1 was obtained commercially (Amersham/SearleCorp., Arlington Heights, Ill.) and was repurified chroma-tographically as described below. Pteroyl-A[`4C1glutamyl-'y-hexaglutamate (["C] PG-7) was synthesized by the solidphase method (16) and provided by the Nutrition Programof the University of Alabama. In synthesis, the "C label wasattached to the first glutamyl unit next to the pteroyl moiety
'Abbreviations used in this paper: L, lactase; M, maltase;PEG, polyethylene glycol; [3H] PG-1, 3',5',9'['H]pteryl-monoglutamate; ["C] PG-7, pteroyl-p[`C]glutamyl--y-hexa-glutamate; S, sucrase.
Jejunal Perfusion of Simple and Conjugated Folates in Tropical Sprue 299

and was thus retained as part of the folate molecule afterhydrolysis. Before use, the folates were purified by columnchromatography and quantitated spectrophotometrically asdescribed. The sp act of each folate was approximately 1.8x 106 cpm/,umol. The perfusion solution was prepared as 1liter of isotonic sodium chloride containing 0.75 ,&mol eachof [3H] PG-1 and ["C] PG-7 and 10 g of polyethylene glycol4,000 (PEG). The pH was adjusted to 7.0 before use.
Procedure. The small bowel was intubated after anovernight fast with three fused polyvinyl tubes which wereweighed by a mercury bag and positioned under fluoroscopiccontrol so that the infusion port lay at least 10 cm beyondthe ligament of Treitz. The aspiration ports were located 15and 30 cm downstream from the infusion port. The testsolution was infused into the jejunum with a peristaltic in-fusion pump (Model 1023, Harvard Apparatus Co., Millis,Mass.) at a constant rate of 9.2 ml/min. A 40-min equilibra-tion period was allowed after which samples of intestinalcontents were collected by siphonage, over ice, at the twodistal openings at a rate of 1 ml/min for 60 min. Afterremoval of a 7-ml aliquot from each aspirate, the contentswere brought to pH 2.8 by the addition of 10% trichloro-acetic acid and stored at -70°C until subjected to columnchromatography.
Each subject received an intramuscular injection of 15mg of folic acid (Lederle Laboratories, Pearl River, N. Y.)immediately and 24 h after perfusion to flush the labeledfolates from the tissues. Urine was collected for 48 h fromthe start of the infusion, its volume measured, and a l-mlsample counted in the manner described below. The percentof urinary recovery of each labeled folate was calculated,taking into account that after intestinal aspiration eachpatient retained -800 ml of the infused solution containing0.6 umol of each labeled folate. Creatinine clearances wereperformed simultaneously.
Assays. Aliquots of each solution and each intestinalaspirate were used to determined the concentration of PEG,the radioactivity of each label, and concentrations of sodiumand chloride. A modified turbidometric method was used todetermine the concentrations of PEG (17). After decoloriza-tion with 37% H202, 1 ml of each sample was preparedfor counting by adding 10 ml of scintillation fluid (Scintisol,Isolab, Inc., Akron, Ohio). Radioactivity was assessed in aBeckman LS-230 liquid scintillation counter (Beckman In-struments Inc., Fullerton, Calif.) set for double isotopecounting so that there was no drift of 'H to the "C countingchannel and a correctable spillover of 0.330 of "C countsto the 'H channel. The counting efficiencies were 65% for"C and 25%o for 3H. Disappearance of each label from the30-cm perfused segment was calculated as previously de-scribed (8). Transintestinal movement of water, sodium, andchloride was calculated for the distal 15-cm segment usingconventional formulas (18). The frozen and acidified re-maining aliquot was thawed, neutralized, and then subjectedto column chromatography on DEAE cellulose chloridewith a linear sodium gradient to separate degradation prod-ucts of ['C] PG-7, as previously described (8).
In vzitro studies. 25 ml of intestinal juice, obtained bysiphonage from the jejunum of an untreated patient andfrozen at - 70°C was thawed and diluted three times insaline to approximate intraluminal concentrations duringperfusion. One-half was adjusted to pH 4.5 and the otherhalf to pH 6.5; ["C] PG-7 was added to each portion inthe same concentrations, 0.75 IAM, as employed in the in vivoperfusion studies. After incubation at 37°C for 15 min, thereaction was stopped by acidification with 10%o trichloro-
acetic acid to pH 2.8 and the mixture frozen at - 70°C. Thereactant mixtures were subsequently subjected to columnchromatography.
Column chromatography. Analytical ion exchange chro-matography using DEAEcellulose chloride was employed toidentify degradation products of ['C] PG-7 in aspiratesobtained from the proximal and distal ports during theperfusion procedures and from the in vitro incubation mix-ture. The chromatographic procedures were identical tothose previously described (8), except that the final ana-lytical column measured 17 X 0.9 cm and 6-ml fractionswere collected. This modification was necessary because ofthe use of a smaller dose of administered folate than thatpreviously employed and permitted adequate separation andidentification of radioactive peaks. Synthetic, spectrallypure, nonradioactive markers for pteroylmono- (PG-1),pteroyldi- (PG-2), and pteroyltriglutamate (PG-3) wereadded to each sample before column application.
Enzymatic assays. Folate conjugase in the mucosal biop-sies was determined by the charcoal precipitation method ofKrumdieck and Baugh (7) utilizing pteroylglutamyl-,yglu-tamyl["4C]glutamic acid (synthesized and provided by Dr.Carlos L. Krumdieck) as a substrate. Based on preliminaryexperiments showing saturation kinetics at a substrate con-centration between 10 and 30 nmol/1.5 ml reaction tube, aconcentration of 20 nmol per tube, a pH of 4.5, and incu-bation time of 15 min was employed in each assay. Activitiesof lactase, sucrase, and maltase were determined by themethod of Dahlqvist (19). Mucosal protein was analyzedby the method of Lowry et al. (20). Results were expressedas nmol/mg protein per 15 min for folate conjugase andas U/g protein per h for lactase (L), sucrase (S), andmaltase (M).
In carrying out these studies, the clinical and laboratoryevaluation of the patients as well as the assays for PEGand electrolytes were performed at the University of PuertoRico, whereas after shipment of the frozen materials on dryice, the isotope counting, column chromatography and en-zymatic assays were performed at the University of Cali-fornia, Davis. All data were analyzed using the paired ttest. Thus, each patient served as his own control, beforeand after treatment.
RESULTS
Biochemical and absorption studies (Table I)
Before treatment, all six patients were anemic withlow levels of serum and red cell folate and serum B12.The values for absorption of D-xylose and vitamin Biwere markedly low in all six patients; steatorrhea wassignificant in five out of the six. The jejunal mucosalabnormality was graded as 4+ in four patients and 3+ intwo. After 6 mo of treatment, each patient had regainedhis normal weight and bowel habits. The post-treatmenthemoglobin levels were greater than 12 g/100 ml andthe levels of serum and red cell folate and serum B-awere within or greater than the normal range. Valuesof serum folate greater than 20 ng/ml and of serum Bsgreater than 1,000 pg/ml in the treated patients reflectthe recent administration of either vitamin. The in-testinal absorption of D-xylose, dietary fat, and vitaminBu were normal after treatment. The jejunal morphol-
300 J. J. Corcino, A. Reisenauer, and C. H. Halsted
70-
*0-
<
40-
ro
FIGURE 1 Luminal disappearance of labeled [3H]pteroyl-monoglutamate and ["C]pteroylheptaglutamate from theperfused jejunum before and after 6-mo treatment, ex-pressed as percent of infused compound per 30-cm jejunalsegment. The closed circles (@) represent original valuesand the open circles (0) the values obtained after treat-ment. Bar and dotted lines represent mean±+SEM.
ogy improved by two grades in all but one patient. Al-though none of the post-treatment biopsies were normalby North American standards, the histology was con-sistent with the previously reported jejunal mucosalabnormalities present in asymptomatic Puerto Ricans(21).
30
,0
20-
15-cm aspirate
FIGURE 2 Recovery of each label in the urine collected for48 h, expressed as percent of administered radioactivity afteradministration by jejunal perfusion of [3H]pteroylmono-glutamate and ["C]pteroylheptaglumate. The closed circles(-)represent original values and the open (0) the valuesobtained after treatment. Bar and dotted lines representmean+SEM.
30-cm aspirate
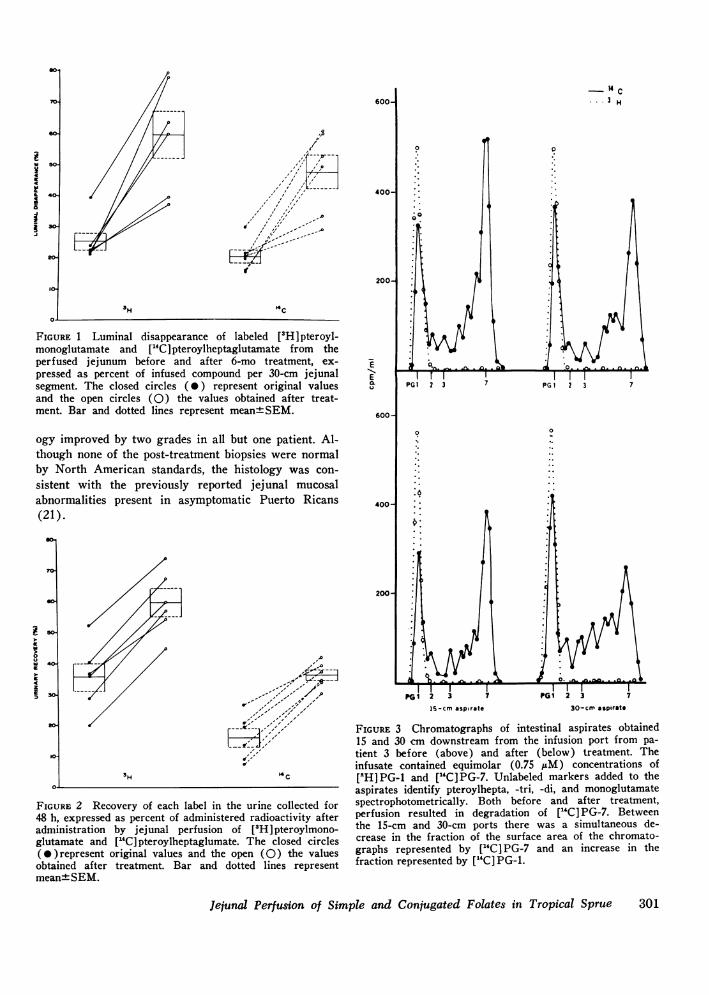
FIGURE 3 Chromatographs of intestinal aspirates obtained15 and 30 cm downstream from the infusion port from pa-tient 3 before (above) and after (below) treatment. Theinfusate contained equimolar (0.75 ttM) concentrations of["H]PG-1 and [(C]PG-7. Unlabeled markers added to theaspirates identify pteroylhepta, -tri, -di, and monoglutamatespectrophotometrically. Both before and after treatment,perfusion resulted in degradation of ["C] PG-7. Betweenthe 15-cm and 30-cm ports there was a simultaneous de-crease in the fraction of the surface area of the chromato-graphs represented by ["C] PG-7 and an increase in thefraction represented by [14C] PG-1.
Jejunal Perfusion of Simple and Conjugated Folates in Tropical Sprue
E
'4c
1c
301
600-
E- 600E
400 pH 4.5 pH 6.5
200
PG 1 2 3 7 PG1 3 7
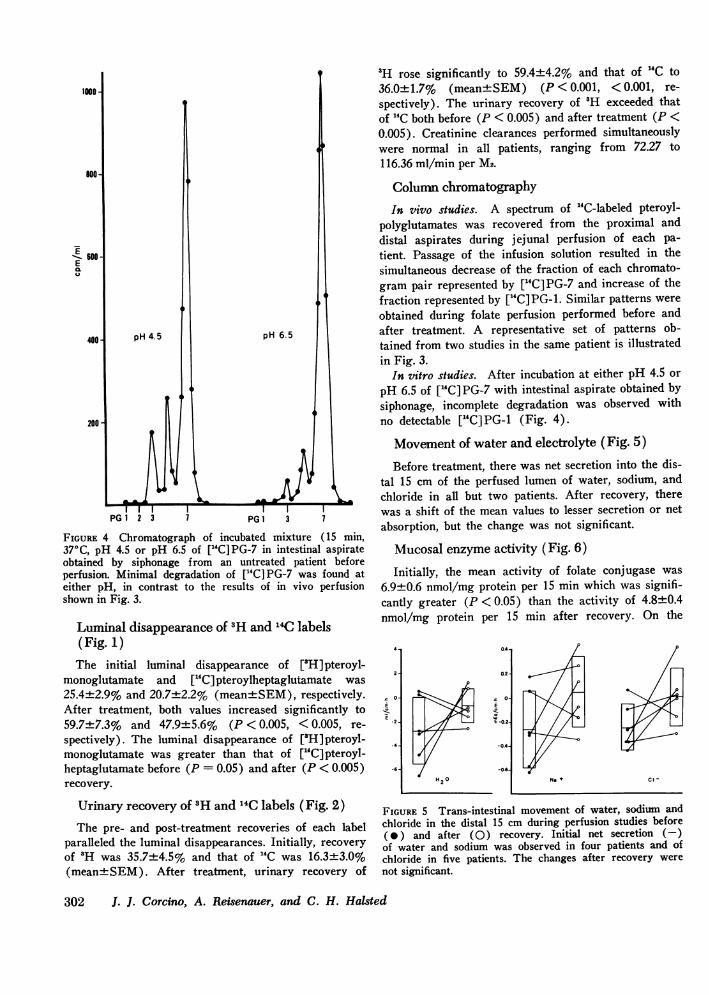
FIGURE 4 Chromatograph of incubated mixture (15 min,37°C, pH 4.5 or pH 6.5 of ["Cl PG-7 in intestinal aspirateobtained by siphonage from an untreated patient beforeperfusion. Minimal degradation of ["C] PG-7 was found ateither pH, in contrast to the results of in vivo perfusionshown in Fig. 3.
Luminal disappearance of 3H and 14C labels(Fig. 1)
The initial luminal disappearance of [aH]pteroyl-monoglutamate and ["C] pteroylheptaglutamate was25.4±2.9% and 20.7±2.2% (mean±SEM), respectively.After treatment, both values increased significantly to59.7±7.3% and 47.9±5.6% (P <0.005, < 0.005, re-spectively). The luminal disappearance of ['H]pteroyl-monoglutamate was greater than that of ["C]pteroyl-heptaglutamate before (P = 0.05) and after (P < 0.005)recovery.
Urinary recovery of 3H and 14C labels (Fig. 2)
The pre- and post-treatment recoveries of each labelparalleled the luminal disappearances. Initially, recoveryof 3H was 35.7+4.5% and that of 14C was 16.3±3.0%(mean+SEM). After treatment, urinary recovery of
3H rose significantly to 59.4±4.2% and that of 14C to36.0±1.7% (mean±SEM) (P < 0.001, < 0.001, re-spectively). The urinary recovery of 3H exceeded thatof 14C both before (P < 0.005) and after treatment (P <0.005). Creatinine clearances performed simultaneouslywere normal in all patients, ranging from 72.27 to116.36 m/min per M2.
Column chromatographyIn vivo studies. A spectrum of 14C-labeled pteroyl-
polyglutamates was recovered from the proximal anddistal aspirates during jejunal perfusion of each pa-tient. Passage of the infusion solution resulted in thesimultaneous decrease of the fraction of each chromato-gram pair represented by [14C] PG-7 and increase of thefraction represented by [14C]PG-i. Similar patterns wereobtained during folate perfusion performed before andafter treatment. A representative set of patterns ob-tained from two studies in the same patient is illustratedin Fig. 3.
In vitro studies. After incubation at either pH 4.5 or
pH 6.5 of [GC]PG-7 with intestinal aspirate obtained bysiphonage, incomplete degradation was observed withno detectable [14C]PG-i (Fig. 4).
Movemnent of water and electrolyte (Fig. 5)Before treatment, there was net secretion into the dis-
tal 15 cm of the perfused lumen of water, sodium, andchloride in all but two patients. After recovery, therewas a shift of the mean values to lesser secretion or netabsorption, but the change was not significant.
Mucosal enzyme activity (Fig. 6)Initially, the mean activity of folate conjugase was
6.9±0.6 nmol/mg protein per 15 min which was signifi-cantly greater (P < 0.05) than the activity of 4.8±0.4nmol/mg protein per 15 min after recovery. On the
04-
0.2-
C O-E
E -0.21
-0.4
-0.6-
0H2
Na41 if4
FIGURE 5 Trans-intestinal movement of water, sodium andchloride in the distal 15 cm during perfusion studies before(*) and after (0) recovery. Initial net secretion (-)of water and sodium was observed in four patients and ofchloride in five patients. The changes after recovery werenot significant.
302 J. J. Corcino, A. Reisenauer, and C. H. Halsted
4 1
2
'. 0-
i-Y-
E.2
-4-
-6-
No +
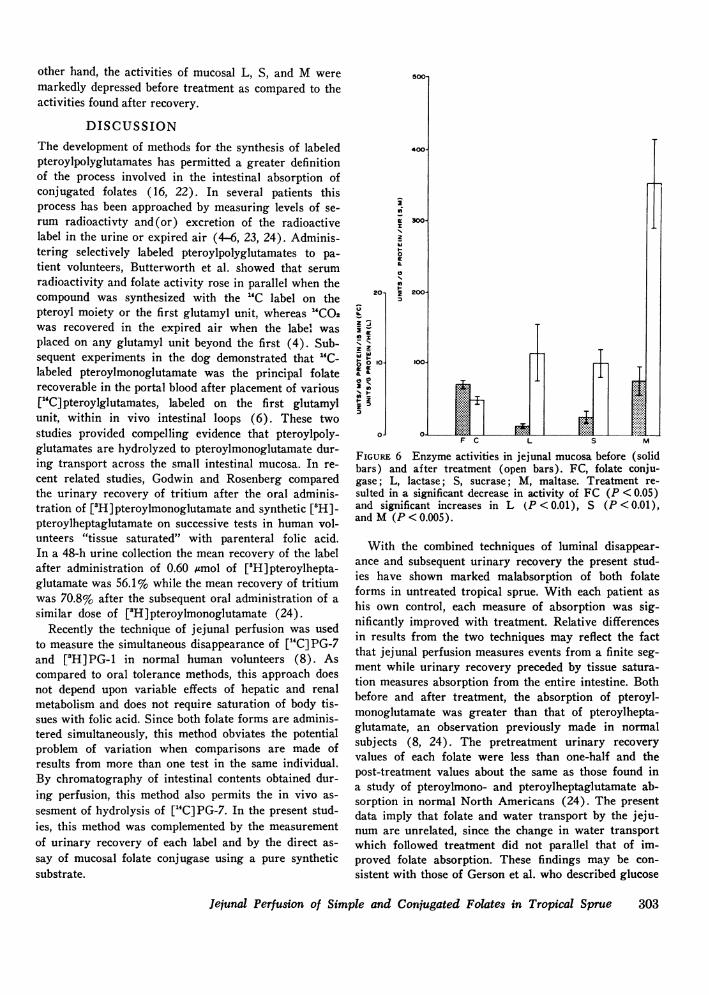
other hand, the activities of mucosal L, S, and M weremarkedly depressed before treatment as compared to theactivities found after recovery.
DISCUSSIONThe development of methods for the synthesis of labeledpteroylpolyglutamates has permitted a greater definitionof the process involved in the intestinal absorption ofconjugated folates (16, 22). In several patients thisprocess has been approached by measuring levels of se-rum radioactivty and(or) excretion of the radioactivelabel in the urine or expired air (4-6, 23, 24). Adminis-tering selectively labeled pteroylpolyglutamates to pa-tient volunteers, Butterworth et al. showed that serumradioactivity and folate activity rose in parallel when thecompound was synthesized with the "C label on thepteroyl moiety or the first glutamyl unit, whereas "CO2was recovered in the expired air when the label wasplaced on any glutamyl unit beyond the first (4). Sub-sequent experiments in the dog demonstrated that 14C-labeled pteroylmonoglutamate was the principal folaterecoverable in the portal blood after placement of various["C] pteroylglutamates, labeled on the first glutamylunit, within in vivo intestinal loops (6). These twostudies provided compelling evidence that pteroylpoly-glutamates are hydrolyzed to pteroylmonoglutamate dur-ing transport across the small intestinal mucosa. In re-cent related studies, Godwin and Rosenberg comparedthe urinary recovery of tritium after the oral adminis-tration of [3H]pteroylmonoglutamate and synthetic [8H]-pteroylheptaglutamate on successive tests in human vol-unteers "tissue saturated" with parenteral folic acid.In a 48-h urine collection the mean recovery of the labelafter administration of 0.60 umol of [3H]pteroylhepta-glutamate was 56.1% while the mean recovery of tritiumwas 70.8% after the subsequent oral administration of asimilar dose of [3H]pteroylmonoglutamate (24).
Recently the technique of jejunal perfusion was usedto measure the simultaneous disappearance of [14C] PG-7and [3H]PG-1 in normal human volunteers (8). Ascompared to oral tolerance methods, this approach doesnot depend upon variable effects of hepatic and renalmetabolism and does not require saturation of body tis-sues with folic acid. Since both folate forms are adminis-tered simultaneously, this method obviates the potentialproblem of variation when comparisons are made ofresults from more than one test in the same individual.By chromatography of intestinal contents obtained dur-ing perfusion, this method also permits the in vivo as-sesment of hydrolysis of [14C] PG-7. In the present stud-ies, this method was complemented by the measurementof urinary recovery of each label and by the direct as-say of mucosal folate conjugase using a pure syntheticsubstrate.
ZZ
z
10
01
i
z
0
2
FIGURE 6 Enzyme activities in jejunal mucosa before (solidbars) and after treatment (open bars). FC, folate conju-gase; L, lactase; S, sucrase; M, maltase. Treatment re-sulted in a significant decrease in activity of FC (P < 0.05)and significant increases in L (P < 0.01), S (P < 0.01),andM (P<0.005).
With the combined techniques of luminal disappear-ance and subsequent urinary recovery the present stud-ies have shown marked malabsorption of both folateforms in untreated tropical sprue. With each patient ashis own control, each measure of absorption was sig-nificantly improved with treatment. Relative differencesin results from the two techniques may reflect the factthat jejunal perfusion measures events from a finite seg-ment while urinary recovery preceded by tissue satura-tion measures absorption from the entire intestine. Bothbefore and after treatment, the absorption of pteroyl-monoglutamate was greater than that of pteroylhepta-glutamate, an observation previously made in normalsubjects (8, 24). The pretreatment urinary recoveryvalues of each folate were less than one-half and thepost-treatment values about the same as those found ina study of pteroylmono- and pteroylheptaglutamate ab-sorption in normal North Americans (24). The presentdata imply that folate and water transport by the jeju-num are unrelated, since the change in water transportwhich followed treatment did not parallel that of im-proved folate absorption. These findings may be con-sistent with those of Gerson et al. who described glucose
Jejunal Perfusion of Simple and Conjugated Folates in Tropical Sprue 303

enhancement of water and folic acid uptake from theperfused jejunum, but no apparent relationship of wa-ter and folic acid uptake from a saline solution (25).The results confirm the findings of Hoffbrand et al.(26) who found malabsorption of equimolar amountsof unlabeled pteroylmonoglutamate and pteroylheptaglu-tamate with successive oral tolerance tests and measure-ment of serum levels in eight patients with tropicalsprue. The present data are more conclusive since themethod used in the latter study required prior treatmentof the patients with a tissue-saturating dose of folic acid.
The data obtained by chromatographic analysis of theintestinal aspirate and by enzymatic assays of the mu-cosal biopsies imply that folate malabsorption in tropi-cal sprue is not caused by deficient hydrolysis of pteroyl-polyglutamate but reflects decreased intestinal transportof its principal degradation product. In vivo hydrolysisof ["C] PG-7 was shown by the identification of achromatographic spectrum of degradation products inthe intestinal aspirates obtained during perfusion whichwas similar to those previously described in normalsubjects (8), and which was similar in studies per-formed both before and after treatment (Fig. 3). Asin the previous study of normals (8), the present datasuggest that hydrolysis of ["C]PG-7 is a consequence ofits contact with the intestinal mucosa. Insignificant deg-radation of ["C] PG-7 was found in vitro at two differ-ent pH levels and under conditions which were designedto mimic the folate and intestinal juice concentrationsobtained during in vivo perfusion (Fig. 4). Direct en-zymatic assays of jejunal mucosa obtained by biopsyshowed that folate conjugase is actually increased inuntreated tropical sprue compared to values obtainedafter treatment, the reverse of the pattern of pre- andpost-treatment activities of jejunal disaccharidases (Fig.6). Folate conjugase activities in the post-treatment bi-opsies were similar to a series of North American con-trols (27). Conceivably, increased folate conjugase inthe mucosa of untreated tropical sprue could reflect thegreater cellular infiltration of the lamina propria. Thecell type responsible for mucosal folate conjugase hasnot been precisely defined. However, by measurement offolate conjugase activity in peripheral lymphocytes,Jigerstad et al. have shown indirectly that the cells ofthe lamina propria are an unlikely enzyme source (28).Halsted et al. have recently shown deconjugation of["C]PG-7 by isolated mucosal epithelial cells (29).
Present evidence from three different in vitro studiessuggests that mammalian intestinal mucosal folate con-jugase is intracellular and not a surface brush borderenzyme (29-31). The present studies indirectly and par-tially support this concept by showing that the mucosalactivity of folate conjugase was not affected by tropicalsprue in parallel to known surface disaccharidase en-
zyme activity. If folate conjugase is indeed an intra-cellular enzyme, the finding of intraluminal hydrolyticproducts of ["C]PG-7 during its jejunal perfusion mustrepresent a process of back diffusion after the parentmolecule has been transported intact into the mucosalcell, as suggested by recent in vitro studies (29). Onthe other hand, the possibility that products of hydrolysisof ["C]PG-7 appear in the lumen as a result of reactionwith a surface-active folate conjugase before mucosaltransport has not been totally excluded by the availabledata.
ACKNOWLEDGMENTSWATe are grateful to Doctors C. E. Butterworth, Jr., C. L.Krumdieck, and C. M. Baugh, Nutrition Program, Univer-sity of Alabama, who supplied the labeled folates used inthese studies. We also wish to thank Ms. M. Maldonadoand tht nursing, technical, and secretarial staff at theGeneral Clinical Research Center of the University ofPuerto Rico for their assistance.
This work was supported by grants from the ResearchCorporation, New York; grant RR-63-13 from the GeneralClinical Research Centers Branch of the Division of Re-search Resources, National Institutes of Health; and grant1 RO 1-AM 18330-01 from the National Institutes of Health.Folate synthesis was supported by grant AM 08644 fromthe National Institutes of Health and grant 1 C-3 fromthe American Cancer Society.
REFERENCES1. Klipstein, F. A. 1968. Tropical Sprue. Gastroenterology.
54: 275-293.2. Corcino, J. J. 1975. Recent advances in tropical sprue.
In Intestinal Absorption and Malabsorption. T. Z.Csaky, editor. Raven Press, New York. 285-299.
3. Butterworth, C. E., Jr., R. Santini, Jr., and W. B. From-meyer, Jr. 1963. The pteroylglutamate components ofAmerican diets as determined by chromatographic frac-tionation. J. Clin. Invest. 42: 1929-1939.
4. Butterworth, C. E., Jr., C. M. Baugh, and C. L. Krum-dieck. 1969. A study of folate absorption and metabolismin man utilizing carbon-14-labeled polyglutamates syn-thesized by the solid phase method. J. Clin. Invest. 48:1131-1142.
5. Rosenberg, I. H., R. R. Streiff, H. A. Godwin, andW. B. Castle. 1969. Absorption of polyglutamatic folate:participation of deconjugating enzymes of the intestinalmucosa. N. Engl. J. Med. 280: 985-988.
6. Baugh, C. M., C. L. Krumdieck, H. J. Baker, and C. E.Butterworth, Jr. 1971. Studies on the absorption andmetabolism of folic acid. I. Folate absorption in the dogafter exposure of isolated intestinal segments to syn-thetic pteroylpolyglutamates of various chain lengths.J. Clin. Invest. 50: 2009-2021.
7. Krumdieck, C. L., and C. M. Baugh. 1970. Radioactiveassay of folic acid polyglutamate conjugase(s). Anal.Biochem. 35: 123-129.
8. Halsted, C. H., C. M. Baugh, and C. E. Butterworth,Jr. 1975. Jejunal perfusion of simple and conjugatedfolates in man. Gastroenterology. 68: 261-269.
9. Herbert, V., H. Baker, 0. Frank, I. Pasher, H. Sobotka,and L. R. Wasserman. 1960. The measurement of folicacid activity in serum: a diagnostic aid in the differentia-tion of the megaloblastic anemias. Blood. 15: 228-235.
304 J. J. Corcino, A. Reisenauer, and C. H. Halsted

10. Grossowicz, N., F. Mandelbaum-Shavit, R. Davidoff,and J. Aronovich. 1962. Microbiologic determination offolic acid derivates in blood. Blood. 20: 609-616.
11. Spray, G. H. 1955. An improved method for the rapidestimation of vitamin Bn in serum. Clin. Sci. (Oxf.).14: 661-667.
12. Roe, J. H., and E. W. Rice. 1948. Photometric methodfor determination of free pentoses in animal tissues. J.Biol. Chem. 173: 507-512.
13. Schenk, E. A., and F. A. Klipstein. 1972. Appendix toSession II: A protocol for the evaluation of small bowelbiopsies. Am. J. Clin. Nutr. 25: 1108-1117.
14. van de Kamer, J. H., H. ten Bokkel Huinink, and H. A.Weyers. 1949. Rapid method for the determination offat in feces. J. Biol. Chem. 177: 347-355.
15. Corcino, J. J., R. C. Dietrich, and A. E. Lanaro. 1972.Assessment of vitamin B2 absorption in tropical sprueutilizing a whole body counter. Bol. Asoc. Med. P. R.64: 275. (Abstr.)
16. Krumdieck, C. L., and C. M. Baugh. 1969. The solid-phase synthesis of polyglutamates of folic acid. Biochem-istry. 8: 1568-1572.
17. Malawer, S. J., and D. W. Powell. 1967. An improvedturbidometric analysis of polyethylene glycol utilizing anemulsifier. Gastroenterology. 53: 250-256.
18. Cooper, H., R. Levitan, J. S. Fordtran, and F. J. In-gelfinger. 1966. A method for studying absorption ofwater and solute from the human small intestine. Gas-troenterology. 50: 1-7.
19. Dahlqvist, A. 1968. Assay of intestinal disaccharidases.Anal. Biochem. 22: 99-107.
20. Lowry, 0. H., N. J. Rosenbrough, A. L. Farr, and R.J. Randall. 1951. Protein measurements with Folinphenol reagent. J. Biol. Chem. 193: 265-275.
21. Klipstein, F. A., I. Beauchamp, J. J. Corcino, M. Mal-donado, J. T. Tomasini, N. Maldonado, C. Rubio, andE. A. Schenk. 1972. Nutritional status and intestinalfunction among rural populations of the West Indies.
II. Barrio Nuevo, Puerto Rico. Gastroentterology. 63:758-767.
22. Godwin, H. A., I. H. Rosenberg, C. R. Ferenz, P. M.Jacobs, and J. Meienhofer. 1972. The synthesis of bio-logically active pteroyloligo--y-L-glutamates (folic acidconjugates). Evaluation of [3H]pteroylheptaglutamatefor metabolic studies. J. Biol. Chem. 247: 2266-2271.
23. Godwin, H. A., and I. H. Rosenberg. 1970. Absorptionof synthetic "cold" and tritium-labeled pteroylheptaglu-tamic acid. J. Clin. Invest. 49: 35a. (Abstr.)
24. Godwin, H. A., and I. H. Rosenberg. 1975. Comparativestudies of the intestinal absorption of [3H]pteroylmono-glutamate and [3H]pteroylheptaglutamate in man. Gas-troenterology. 69: 364-373.
25. Gerson, C. D., N. Cohen, G. W. Hepner, N. Brown,V. Herbert, and H. D. Janowitz. 1971. Folic acid ab-sorption in man: enhancing effect of glucose. Gastro-enterology. 61: 224-227.
26. Hoffbrand, A. V., T. F. Necheles, N. Maldornado, E.Horta, and R. Santini. 1969. Malabsorption of folatepolyglutamates in tropical sprue. Br. Med. J. 2: 543-547.
27. Halsted, C. H., A. M. Reisenauer, and J. J. Corcino.1975. Effect of tropical sprue on hydrolysis and absorp-tion of conjugated folates. Gastroenterology. 68: 5a.
28. Jagerstad, M., K. Lindstrand, A. Norden, A-K. Westes-son, and T. Lindberg. 1974. The folate conjugase activityof the intestinal mucosa in celiac disease. Scand. J. Gas-troenterol. 9: 255-259.
29. Halsted, C. H., A. M. Reisenauer, C. Back, and G.Gotterer. 1976. In vitro uptake and metabolism of pter-oylpolyglutamate by rat small intestine. J. Nutr. 106:485492.
30. Hoffbrand, A. V., and T. J. Peters. 1969. The sub-cellular localization of pteroyl polyglutamate hydrolaseand folate in guinea pig intestinal mucosa. Biochim.Biophys. Acta. 192: 479-485.
31. Rosenberg, I. H., and H. A. Godwin. 1971. The diges-tion and absorption of dietary folate. Gastroenterology.60: 445-463.
Jejunal Perfusion of Simple and Conjugated Folates in Tropical Sprue 305