Embed Size (px)
Citation preview

- Is there limiting similarity in the phenology of fleshy fruits? - 617
Journal of Vegetation Science 16: 617-624, 2005© IAVS; Opulus Press Uppsala.
AbstractQuestion: Is there evidence for limiting similarity in thetiming of fruit production by a bird-dispersed plant commu-nity? Is the rate of fruit removal in each plant species in-versely related to fruit availability in other species? Cansimple measurements of fruit phenologies (i.e. temporalchanges in fruit availability) obscure important fruit attributesthat influence their removal by birds?Location: Vancouver Island, British Columbia, Canada.Methods: Periods of fruit availability were measured in tenwoody angiosperm species for two years. In the second year,the fate of individual fruits was quantified to disentangledates of fruit maturation, removal and mortality from meas-urements of availability.Results: Null model analyses of fruit availability distribu-tions showed no evidence for limiting similarity. However,fruit removal rates of most plant species were correlated withtheir relative abundance in the community, indicating fruitswere removed more rapidly when other fruits were lessabundant. Species with similar periods of fruit availabilityoften had different dates of fruit maturation, rates of fruitremoval and fruit persistence times, indicating fruit avail-ability measurements can obscure important bird-fruit inter-actions.Conclusions: Competition for dispersers appears to occur.However, it has not resulted in limiting similarity in fruitavailability distributions. A likely explanation for this dis-crepancy is that fruit availability distributions often con-found several important fruit attributes that can independ-ently influence fruit removal by birds.
Keywords: Competition; Frugivory; Null model; Seed dis-persal; Vancouver Island.
Nomenclature: Hitchcock & Cronquist (1994).
Is there limiting similarity in the phenology of fleshy fruits?
Burns, K.C.
Department of Organismic Biology, Ecology & Evolution, University of California, Los Angeles, USA; Bamfield MarineSciences Centre, Bamfield, British Columbia, Canada; Present address: School of Biological Sciences, Victoria Univer-
sity of Wellington, P.O. Box 600, Wellington, New Zealand; Fax +64 44636873; E-mail [email protected]
Introduction
One of the longest standing questions in ecology iswhether interspecific competition limits the similar-ity of co-existing species (Hutchinson 1959). Thisquestion has experienced a resurgence of interest inrecent years (Losos 2000). Rekindled interest in lim-iting similarity has been fuelled by careful considera-tion of the methods used to document its existence(see Dayan & Simberloff 2005). To help avoid con-troversy, which has plagued the subject in the past,detailed methodological criteria have now been es-tablished to help ensure the credibility of empiricaltests (Schluter 2000a, b). For example, the phenotypictrait under investigation must directly correspondwith resource use, phenotypic patterns must differfrom randomized expectations and evidence for com-petition must be established.
Scores of studies have tested for limiting similar-ity in fruit phenologies (i.e. ‘community-wide char-acter displacement’; Poulin et al. 1999; Wilson 1999;Thies & Kalko 2004). Differences in fruit productiontimes among plant species that share seed disperserscould reduce competition for seed dispersers (Snow1965). However, the rigorous methods outlined abovehave not been applied to the phenology of fleshyfruits, and evidence for the phenomenon is mixed(Wheelwright 1985; Gotelli & Graves 1996).
Here, I test for limiting similarity in the phenol-ogy of a bird-dispersed plant community in westernCanada. Seasonal patterns in fruit availability of tenspecies of trees and shrubs were assessed with anewly derived null model. Seasonal changes in fruitremoval by birds were evaluated to test whether plantsmight compete for seed dispersers. Lastly, fruit matu-ration dates, fruit removal rates and fruit persistencetimes were measured to evaluate whether interspecificdifferences in fruit availability distributions accu-rately reflect interspecific differences in dispersal bybirds.

618 Burns, K.C.
Methods
All data were collected on the west coast of Van-couver Island, British Columbia, Canada. The areaexperiences a temperate climate, with warm tempera-tures and moderate precipitation occurring from Maythrough September, and almost continuous rainfall andcooler temperatures from October through April. Theprevailing vegetation type is coastal conifer forest,which is dominated by Thuja plicata, Tsuga hetero-phylla and Picea sitchensis. Klinka et al. (1989) andCody (1999) described the site’s vegetation, climate,geology and natural history.
Ten species of woody angiosperms are commonbelow the canopy of conifer forest:Gaultheria shallon Lonicera involucrataPyrus fusca Rhamnus purshianaRibes bracteosum Rubus parviflorusRubus spectabilis Sambucus racemosaVaccinium ovatum Vaccinium parvifolium.
All produce fleshy fruits that are commonly con-sumed by four species of fruit-eating birds. Detailed
descriptions of the local seed dispersal mutualism,including bird-fruit abundances patterns (Burns 2004a),the effects of seed dispersal on plant recruitment (Burns2004b, 2005a) and how fruit colour influences fruitconsumption by birds (Burns & Dalen 2002; Burns2005b, c) can be found elsewhere.
Phenological observations to test for limiting simi-larity in fruit phenologies were made over two fruitingseasons. In 1998, the phenology of each species wasquantified with periodic observations of a single branchon ten plants of each species (except S. racemosa, N =5, and P. fusca, N = 9). A total of 20 censuses wereconducted throughout the fruiting season at approxi-mately 10-day intervals (10.8 ± 0.5, range: 9–16).During each census, the total number of ripe fruits onmarked branches was noted.
As in most studies (e.g. Smith-Ramírez et al. 1998;Burns & Dalen 2002; Thies & Kalko 2004), fruitphenologies were defined as temporal changes in theavailability of fruits to frugivores. Each species’ phe-nology can be broken into two components, averagefruiting times and fruiting intervals. Average fruiting
Fig. 1. Fruit phenologies of ten bird-dispersed fruit species in a temperate rainforest in western Canada in 1998. Bars represent theproportion of plants with ripe fruits available during each census.
- Is there limiting similarity in the phenology of fleshy fruits? - 619
times were calculated from the frequency distributionof marked plants bearing ripe fruits during each census(see Fig. 1). Fruiting intervals were defined as onestandard deviation around average dates.
Slightly different methods were used to quantifyfruit phenologies in 1999, to disentangle dates of fruitmaturation, removal and mortality from traditionalmeasurements of fruit availability. The fruiting stalksof six fruits on 12 individuals of each species weremarked with small pieces of green tape (1 cm2) andfollowed throughout the fruiting season within one dayof the previous year’s censuses. Observations in 1998indicated that: (1) characteristic markings were leftbehind on fruiting stalks after removal by birds, (2)fruits rarely fell from parent plants without showingevidence of desiccation, stem damage or pathogenicattack prior to fruit fall and (3) following fruit damage,fruits were avoided by birds and usually fell fromparent plants while still attached to stalks. Therefore,dates of fruit maturation, removal and mortality werequantified through inspection of fruiting stalks and theprior status of fruits, which was then cross-checked bysearching the ground below marked plants.
Marchinko et al.’s (2004) null model was employedto test for non-random patterns in fruit availabilitydistributions. The null model is based on the index d/w,which is commonly used to characterize phenotypicoverlap in theoretical studies (e.g. MacArthur 1972).In this study, d/w was defined as the difference be-tween average fruiting times of two adjacent species inthe fruiting sequence divided by the sum of their fruit-ing intervals.
To test for non-random fruiting patterns, plantspecies were first arranged throughout the fruitingseason according to their average dates of availability.The index d/w was then calculated for all adjacentspecies pairs in the sequence of fruit availability (i.e. L.involucrata and R. spectabilis, R. spectabilis and S.racemosa, … V. ovatum and P. fusca, see Fig. 1) . Thevariance of these values was then calculated to obtain asingle metric characterizing community-level differ-entiation in fruit phenologies. To determine whetherthe observed variance in d/w is lower than would beexpected by chance, the observed variance was com-pared to expected variance values generated in MonteCarlo simulations.
Ashton et al. (1988) demonstrated that randomizingphenologies within the observed range in average fruit-ing times inflates expected values of phenologicaloverlap and gives an excess of Type I errors. Toremove this ‘edge effect’, the first and last species inthe seasonal sequence of fruit production were heldconstant during all iterations (following Simberloff &Boecklen 1981), so the length of the observed and
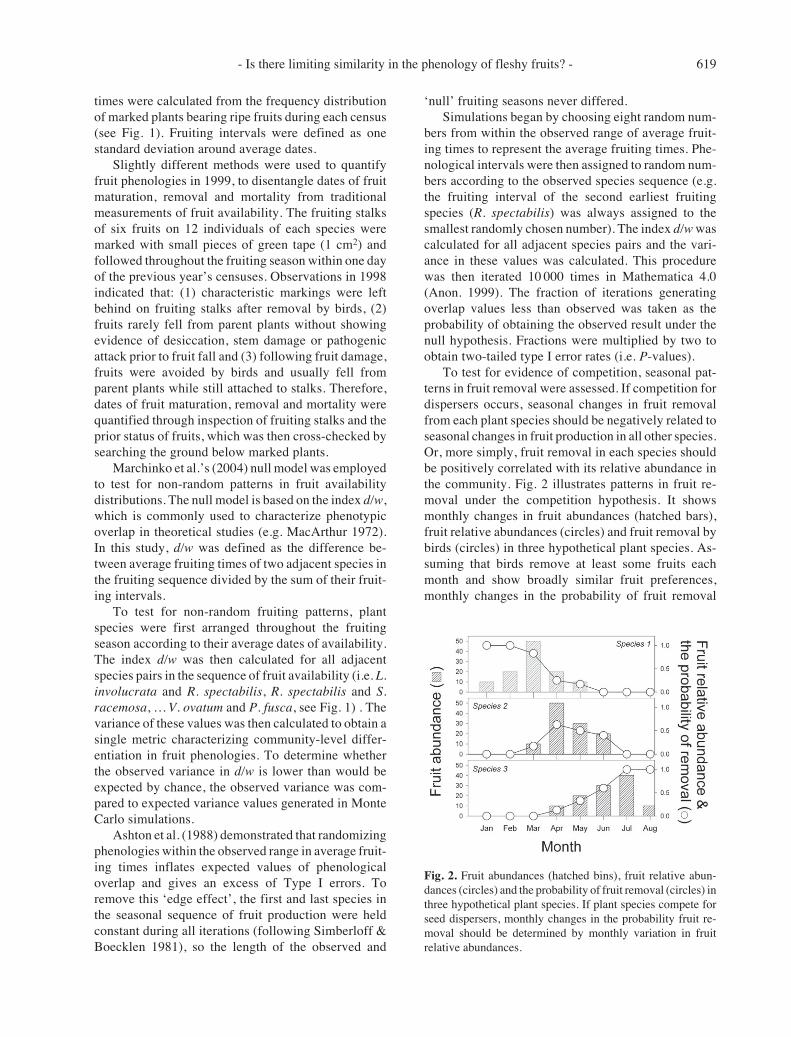
Fig. 2. Fruit abundances (hatched bins), fruit relative abun-dances (circles) and the probability of fruit removal (circles) inthree hypothetical plant species. If plant species compete forseed dispersers, monthly changes in the probability fruit re-moval should be determined by monthly variation in fruitrelative abundances.
‘null’ fruiting seasons never differed.Simulations began by choosing eight random num-
bers from within the observed range of average fruit-ing times to represent the average fruiting times. Phe-nological intervals were then assigned to random num-bers according to the observed species sequence (e.g.the fruiting interval of the second earliest fruitingspecies (R. spectabilis) was always assigned to thesmallest randomly chosen number). The index d/w wascalculated for all adjacent species pairs and the vari-ance in these values was calculated. This procedurewas then iterated 10 000 times in Mathematica 4.0(Anon. 1999). The fraction of iterations generatingoverlap values less than observed was taken as theprobability of obtaining the observed result under thenull hypothesis. Fractions were multiplied by two toobtain two-tailed type I error rates (i.e. P-values).
To test for evidence of competition, seasonal pat-terns in fruit removal were assessed. If competition fordispersers occurs, seasonal changes in fruit removalfrom each plant species should be negatively related toseasonal changes in fruit production in all other species.Or, more simply, fruit removal in each species shouldbe positively correlated with its relative abundance inthe community. Fig. 2 illustrates patterns in fruit re-moval under the competition hypothesis. It showsmonthly changes in fruit abundances (hatched bars),fruit relative abundances (circles) and fruit removal bybirds (circles) in three hypothetical plant species. As-suming that birds remove at least some fruits eachmonth and show broadly similar fruit preferences,monthly changes in the probability of fruit removal

620 Burns, K.C.
should equal fruit relative abundances if competitionoccurs. Conversely if competition does not occur, fruitremoval should be correlated with fruit abundances.
To test these predictions, seasonal changes in fruitremoval were subject to analysis of covariance (ancova).The number of fruits removed from each species dur-ing each census interval was used as the dependentvariable. The relative abundance of each species dur-ing each census interval (the number of fruits availablein each species divided by the total number of all fruitsavailable) was used one covariate. The abundance ofeach species during each census interval was used as asecond covariate. However, fruit abundances were cor-related with fruit relative abundances (r = 0.405, p <0.001). Covariation between fruit abundances and fruitrelative abundances complicates establishing whichcovariate has the strongest influence on fruit removal.Therefore, to disentangle the effects of fruit abun-dances and fruit relative abundances on fruit removal,the analysis was conducted using Type-I sums-of-squares. Fruit abundances were entered into the modelfirst, and after removing variation in fruit removalassociated with fruit abundances, the relationship be-tween fruit removal and fruit relative abundances wasassessed. Potential differences between adjusted means(i.e. intercepts) in the relationships between fruit re-moval and fruit relative abundances for each specieswere assessed next. Potential differences in the slopesof relationships for each species were assessed last. Alldata were log (+ 1) transformed to conform to assump-tions. This analysis was conducted using the GLMprocedure in SPSS (Anon. 2002).
To evaluate whether interspecific differences infruit availability distributions accurately reflectinterspecific differences in dispersal by birds, dates offruit maturation and fruit removal were graphicallycompared to fruit availability distributions. Graphicalcomparisons were used to identify species pairs withstrongly overlapping fruit availability distributions.Once identified, average dates of fruit maturation,removal and availability were compared betweenspecies with analysis of variance (Anova). Samplesizes were based on the number of plants marked forobservation (N = 12). Therefore, values for individu-ally marked fruits on each plant were averaged prior toanalysis. Data were log transformed to conform toassumptions when necessary.
Results
Null model analyses showed no support for limit-ing similarity in fruit phenologies in either year (Figs.1 and 3). Approximately one in four randomizations(2480 / 10000) were more evenly displaced than theobserved phenological pattern in the first year (P =0.4960). Results from the second year were similar(2206 / 10000, P = 0.4412).
Although phenological patterns showed little evi-dence for limiting similarity, patterns in fruit removalsuggest that plants compete for seed dispersers (Fig.4). Results of Ancova showed that the number of fruitsremoved from each species during each census inter-val was positively associated with their abundances(F1,61 = 7.141, P = 0.010, Fig. 4). However, afterremoving the effect of fruit abundances on fruit re-moval, a significant amount of variation in fruit re-moval could be attributed to fruit relative abundances(F1,61 = 9.964, P = 0.003). The adjusted means (i.e.intercepts) did not differ among species (F9,72 = 1.174,P = 0.325), and the slope of relationships also did notdiffer (F10,72 = 1.709, P = 0.095). Therefore, relation-ships between fruit removal and fruit relative abun-dances were generally similar among plant species.Birds also appeared to be in limited supply, as approxi-mately one-third of all ripe fruits produced were neverdispersed (30.8% ± 5.6%; range: 8% - 58%).
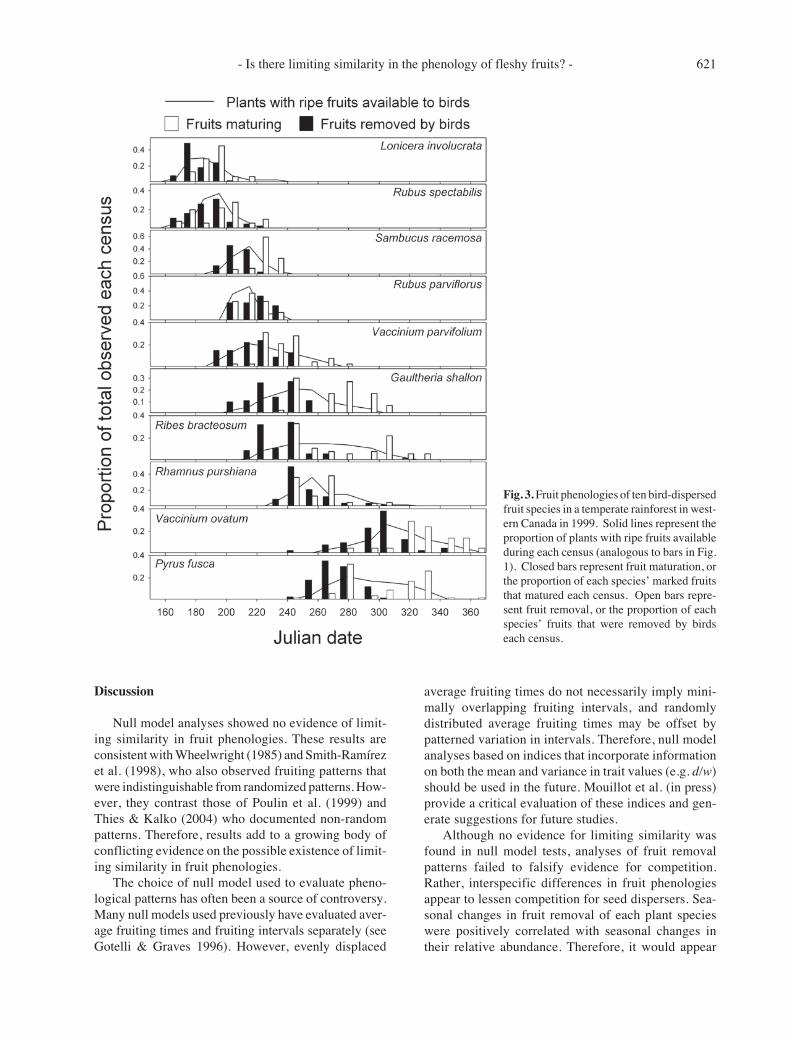
Dates of fruit maturation and removal sometimesvaried independently of fruit availability distributions.Graphical comparisons of fruit availability distribu-tions (Figs. 1 and 3) revealed that one species pair(Sambucus racemosa and Rubus parviflorus) was par-ticularly similar. ‘Anova’ showed that both species hadsimilar dates of fruit availability (F1,22 = 0.90, P =0.354). However, they had different dates of fruit matu-ration (F1,22 = 12.01, P = 0.002) and removal (F1,22 =5.22, P = 0.033). Interestingly, differences in averagedates of maturation and removal were inverted be-tween species; S. racemosa matured prior to R.parviflorus, yet R. parviflorus was removed more rap-idly than S. racemosa. However, S. racemosa ulti-mately dispersed a greater fraction of marked fruits(78%) than did R. parviflorus (42%).
- Is there limiting similarity in the phenology of fleshy fruits? - 621
Discussion
Null model analyses showed no evidence of limit-ing similarity in fruit phenologies. These results areconsistent with Wheelwright (1985) and Smith-Ramírezet al. (1998), who also observed fruiting patterns thatwere indistinguishable from randomized patterns. How-ever, they contrast those of Poulin et al. (1999) andThies & Kalko (2004) who documented non-randompatterns. Therefore, results add to a growing body ofconflicting evidence on the possible existence of limit-ing similarity in fruit phenologies.
The choice of null model used to evaluate pheno-logical patterns has often been a source of controversy.Many null models used previously have evaluated aver-age fruiting times and fruiting intervals separately (seeGotelli & Graves 1996). However, evenly displaced
average fruiting times do not necessarily imply mini-mally overlapping fruiting intervals, and randomlydistributed average fruiting times may be offset bypatterned variation in intervals. Therefore, null modelanalyses based on indices that incorporate informationon both the mean and variance in trait values (e.g. d/w)should be used in the future. Mouillot et al. (in press)provide a critical evaluation of these indices and gen-erate suggestions for future studies.
Although no evidence for limiting similarity wasfound in null model tests, analyses of fruit removalpatterns failed to falsify evidence for competition.Rather, interspecific differences in fruit phenologiesappear to lessen competition for seed dispersers. Sea-sonal changes in fruit removal of each plant specieswere positively correlated with seasonal changes intheir relative abundance. Therefore, it would appear
Fig. 3. Fruit phenologies of ten bird-dispersedfruit species in a temperate rainforest in west-ern Canada in 1999. Solid lines represent theproportion of plants with ripe fruits availableduring each census (analogous to bars in Fig.1). Closed bars represent fruit maturation, orthe proportion of each species’ marked fruitsthat matured each census. Open bars repre-sent fruit removal, or the proportion of eachspecies’ fruits that were removed by birdseach census.

622 Burns, K.C.
that most fruit species were more likely to be removedduring periods of low phenological overlap with otherspecies. This result also suggests that different fru-givorous bird species show broadly similar fruit pref-erences, and that all birds consume different types offruits in accordance with their availability in the com-munity. Fruit-eating birds also appeared to be in lim-ited supply, as nearly one in three fruits that reachedmaturity were never dispersed.
Evidence for competition for dispersers has beenfrequently documented (e.g. Manasse & Howe 1983;Alcántara et al. 1997; Carlo et al. 2003; Saracco et al.2005). In particular, the results reported here are con-sistent with those of Wheelwright (1985), who quanti-fied the fruit phenologies of 23 lauraceous tree speciesin Costa Rica. He also found evidence for interspecificcompetition for dispersers, yet no support for limitingsimilarity in phenological patterns.
Nevertheless, testing for interspecific competitionby analysing patterns in fruit removal provides onlyindirect evidence for competition. Experimental ma-nipulations that control for other factors, such as thespatial distributions of plants (see Mário & Lewinsohn2004), would provide more direct evidence for inter-specific competition.
Evidence for competition in the absence of limitingsimilarity in fruit phenologies is somewhat curious. Ifthe process thought to generate staggered phenologicalpatterns occurs, why would the pattern not result fromit? Evolutionary constraints are commonly observed instudies of seed dispersal mutualisms (Herrera 2002a).
Therefore, phylogenetic effects, allometric relation-ships, environmental factors, or geographic variationin the composition of competitors may have constrainedthe evolutionary development of staggered phenologi-cal patterns (Thompson 1994; Smith-Ramírez et al.1998; Herrera 2002b). However, another possibility isthat interspecific differences in fruit availability distri-butions do not accurately reflect interspecific differ-ences in the use of seed dispersers.
An important criterion for demonstrating evidencefor limiting similarity (i.e. community-wide characterdisplacement) is that the phenotypic trait under inves-tigation must accurately reflect resource use (see Dayan& Simberloff 2005). Patterns in fruit maturation, re-moval and availability suggest that this may not be thecase. Rubus parviflorus and Sambucus racemosa hadvery similar fruit availability distributions in both years(Figs. 1 and 3). Therefore, these two species providesome of the strongest evidence against limiting simi-larity in null model analyses. However, similarity intheir fruit availability distributions masked underlyingdifferences in fruit maturation dates, removal rates andpersistence times, which appear to influence fruit re-moval by birds. Sambucus racemosa matured fruitprior to R. parviflorus. However, birds prefer R.parviflorus fruits (Burns 2005c), and most R. parvi-florus fruits were removed before S. racemosa fruits.Nevertheless, S. racemosa fruits persisted on parentplants until later in the season, when they were eventu-ally removed. As a result, both species appear to gainaccess to seed dispersers through differences in fruit
Fig. 4. Relationships between fruit removal andfruit relative abundances in ten bird-dispersedplant species in western Canada. Data pointsrepresent the number of fruits removed from eachplant species during each census interval. Sepa-rate best fit lines are shown for each plant species,which are labelled by their genus-species initials.Positive relationships indicate that fruits wereremoved more rapidly when they co-occurredwith fewer numbers of fruits produced by otherplant species.

- Is there limiting similarity in the phenology of fleshy fruits? - 623
maturation dates, removal rates and persistence times,even though their fruit availability distributions werevirtually identical. Consequently, null model analysesof fruit availability distributions do not accurately testfor limiting similarity. Future tests for limiting similar-ity in fleshy fruits would probably benefit from criti-cally evaluating the phenological measurement used innull model analyses.
The study of ecology has often been criticized forfailing to resolve controversial issues. Once-populartheoretical predictions are often abandoned in favourof more recent ideas, once their utility is questioned(Rosenzweig 1995). This often leaves a wake of unre-solved issues in ecology’s path. Limiting similarity infruit phenologies is a good example of this situation. Itwas a popular topic in the late 1970s and early 1980s,but has since then lost its popularity. More recent tests(e.g. Smith-Ramírez et al. 1998; Poulin et al. 1999;Thies & Kalko 2004) illustrate that we still have muchto learn by testing the hypothesis and that we are notyet able to determine how frequently limiting similar-ity in fruit phenologies occurs. Results from this studysuggest that more detailed consideration of the meth-ods used to test for limiting similarity may generatenew insight into the phenomenon.
Acknowledgements. Funding was provided by the FulbrightFoundation. I would also like to thank T. Case, M. Cody, J.Dalen, H. Hespinheide, S. Law, R. Vance and S. Zaklan fortheir assistance and advice. Two anonymous reviewers pro-vided helpful comments to the manuscript. I am especiallygrateful to W.J. Stubbs and J.B. Wilson who identifiedseveral inconsistencies in the rational of a previous draft.
References
Anon. 1999. Mathematica 4.0. Wolfram Research Inc., Chi-cago, IL, US.
Anon. 2002. Release 11.5.1 for Windows. SPSS Inc., Chi-cago, IL, US.
Alcántara, J.M., Rey, P.J., Valera, F., Sánchez-Lafuente,A.M. & Gutiérrez, J.E. 1997. Habitat alteration and plantintra-specific competition for seed dispersers. An exam-ple with Olea europaea var. sylvestris. Oikos 79: 291-300.
Ashton, P.S., Givnish, T.J. & Appanah, S. 1988. Staggeredflowering in the Dipterocarpaceae: New insights intofloral induction and the evolution of mast fruiting in theaseasonal tropics. Am. Nat. 132: 44-66.
Burns, K.C. 2004a. Scale and macroecological patterns inseed dispersal mutualisms. Global Ecol. Biogeogr. 13:289-293.
Burns, K.C. 2004b. Relationships between the demographyand distribution of two bird-dispersed plants in an island
archipelago. J. Biogeogr. 31: 1935-1943.Burns, K.C. 2005a. A multi-scale test for dispersal filters in
an island plant community. Ecography 28: 552-560.Burns, K.C. 2005b. Effects of bi-colored displays on avian
fruit color preferences in a color polymorphic plant. J.Torrey Bot. Soc. 132: 505-509.
Burns, K.C. 2005c. Does mimicry occur in fleshy-fruits?Evol. Ecol. Res. 7: 1067-1076.
Burns, K.C. & Dalen, J.L. 2002. Foliage color contrasts andadaptive fruit color variation in a bird dispersed plantcommunity. Oikos 96: 463-469.
Carlo, T.A., Collazo, J.A. & Groom, M.J. 2003. Avian fruitpreferences across a Puerto Rican forested landscape:pattern consistency and implications for seed removal.Oecologia 134: 119-131.
Cody, M.L. 1999. Assembly rules at different scales in plantand animal communities. In: Weiher, E. & Keddy, P.(eds.) Ecological assembly rules, pp. 165-205. Cam-bridge University Press, Cambridge, UK.
Dayan, T. & Simberloff, D. 2005 Ecological and commu-nity-wide character displacement: the next generation.Ecol. Lett. 8: 875-894.
Gotelli, N.J. & Graves, G.R. 1996. Null Models in Ecology.Smithsonian Press, Washington D.C., US.
Herrera, C.M. 2002a. Seed dispersal by vertebrates. In:Herrera, C.M. & Pellmyr, O. (eds.) Plant-animal inter-actions: an evolutionary approach, pp. 185-208. Black-well Science, Oxford, UK.
Herrera, C.M. 2002b. Correlated evolution of fruit and leafsize in bird-dispersed plants: species-level variance infruit traits explained a little further? Oikos 97: 426-432.
Hitchcock, L.C. & Cronquist, A. 1994. Flora of the PacificNorthwest. University of Washington Press, Seattle,WA, US.
Hutchinson, G.E. 1959. Homage to Santa Rosalia, or why arethere so many kinds of animals? Am. Nat. 93: 145-159.
Klinka, K., Krajina, V.J., Ceska, A. & Scagal, A.M. 1989.Indicator plants of coastal British Columbia. Universityof British Columbia Press, Vancouver, CA.
Losos, J.B. 2000. Ecological character displacement and thestudy of adaptation. Proc. Natl. Acad. Sci. 97: 5693-5695.
MacArthur, R.H. 1972. Geographical ecology. PrincetonPress, Princeton, NJ, US.
Manasse, R.S. & Howe, H.F. 1983. Competition for disper-sal agents among tropical trees: influences of neigh-bours. Oecologia 59: 185-190.
Marchinko, K.B., Nishizaki, M.T. & Burns, K.C. 2004.Community-wide character displacement in barnacles: anew perspective for past observations. Ecol. Lett. 7: 114-120.
Mário, A.-N. & Lewinsohn, T.M. 2004. Small-scale auto-correlation and the interpretation of relationships be-tween phenological parameters. J. Veg. Sci. 15: 561-568.
Mouillot, D., Stubbs, W., Faure, M., Dumay, O., Tomasini,J.A., Wilson, J.B. & Chi, T.D. In press. Niche overlapestimates based on quantitative functional traits: a newfamily of non-parametric indices. Oecologia.
Poulin, B., Wright, S.J., Lefebvre, G. & Calderón, O. 1999.

624 Burns, K.C.
Interspecific synchrony and asynchrony in the fruitingphenologies of congeneric bird-dispersed plants inPanama. J. Trop. Ecol. 15: 231-227.
Rosenzweig, M.L. 1995. Species diversity in space and time.Cambridge University Press, Cambridge, UK.
Saracco, J.F., Collazo, J.A., Groom, M.J. & Carlo, T.A.2005. Crop size and fruit neighbourhood effects on birdvisitation to fruiting Schefflera morototoni trees in PuertoRico. Biotropica 37: 81-87.
Schluter, D. 2000a. Introduction to the symposium: speciesinteractions and adaptive radiation. Am. Nat. 156: S1-S4.
Schluter, D. 2000b. Ecological character displacement inadaptive radiation. Am. Nat. 156: S4-S16.
Simberloff, D. & Boecklen, W. 1981. Santa Rosalia recon-sidered: size ratios and competition. Evolution 35: 1206-1228.
Smith-Ramírez, C., Armesto, J.J. & Figueroa. J. 1998. Flow-ering, fruiting and seed germination in Chilean rainforest
myrtaceae: ecological and phylogenetic constraints. PlantEcol. 136: 119-131.
Snow, D.W. 1965. A possible selective factor in the evolu-tion of fruiting seasons in tropical forest. Oikos 15: 274-281.
Thies, W. & Kalko, E.K. 2004. Phenology of neotropicalpepper plants (Piperaceae) and their association withtheir main seed dispersers, two short-tailed bats, Carolliaperspicillata and C. castanea (Phyllostomidae). Oikos104: 362-376.
Thompson, J.H. 1994. The coevolutionary process. Univer-sity of Chicago Press, Chicago, IL, US.
Wheelwright, N.T. 1985. Competition for dispersers, and thetiming of flowering and fruiting in a guild of tropicaltrees. Oikos 44: 465-477.
Wilson, J.B. 1999. Assembly rules in plant communities. In:Weiher, E. & Keddy, P. (eds.) Ecological assembly rules,pp. 130-164. Cambridge University Press, Cambridge,UK.
Received 17 September 2004;Accepted 16 September 2005.
Co-ordinating Editor: J.B. Wilson.