Embed Size (px)
Citation preview

SUMMARY
Phytoplasmas for which 16S rDNA sequences areavailable have been classified into 20 major phylogenet-ic groups or subclades. Further phytoplasmas havebeen assigned to these groups, according to other mole-cular data such as RFLP analysis of PCR-amplified ri-bosomal DNA, nucleic acid hybridization, and serologi-cal comparison. A total of 75 phytoplasmas were distin-guishable among the molecularly characterized phy-topathogenic mollicutes.
RIASSUNTO
STATO ATTUALE DELLA CLASSIFICAZIONE MOLECOLA-RE DEI FITOPLASMI. I fitoplasmi, dei quali la sequenzanucleotidica del 16S rDNA era nota, sono stati classifi-cati in 20 gruppi filogenetici principali. Sulla base di da-ti ottenuti mediante analisi di RFLP del DNA riboso-male amplificato mediante PCR, ibridazione di acidinucleici e sierologia, altri fitoplasmi sono stati assegnatiai gruppi suddetti. Un totale di 75 distinti taxa è statoindividuato tra tutti i mollicuti fitopatogeni caratterizza-ti con metodiche molecolari.
Key words: phylogeny, taxonomy, phytoplasmal dis-eases.
INTRODUCTION
Phytoplasmas are nonhelical mollicutes associatedwith diseases of several hundred plant species (McCoyet al., 1989). These disorders are characterized byflower malformation, other growth aberrations, yellow-ing and/or decline and are collectively referred to asyellows diseases. They were thought to be caused byviruses until a group in Japan (Doi et al., 1967) reco-gnized in diseased plants wall-less, pleomorphic bodies
Corresponding author: E. SeemüllerFax: +49. 6221. 861222E-Mail: [email protected]
which resembled in morphology and ultrastructure my-coplasmas known to be pathogens of humans and ani-mals. Due to this similarity the structures detected inplants were called mycoplasma-like organisms (MLOs),but the failure of all attempts to culture phytoplasmasunder axenic conditions made their characterizationdifficult and prevented their definitive classification.
A new era in phytoplasma research began whenDNA-based methods were introduced about a decadeago, following the development of procedures to ex-tract and enrich phytoplasmal DNA from infectedplants or insects (Kirkpatrick et al., 1987; Lee andDavis, 1988; Sears et al., 1989; Kollar et al., 1990). Phy-toplasmal DNA could then be cloned and sequenced,and methods applied for amplification of this DNA us-ing PCR. A wealth of molecular data on phytoplasmadiversity and on the relationships of the phytoplasmaswas generated. In several comprehensive studies onphylogeny and taxonomy of the phytoplasmas, manyphytoplasmas from several phylogenetic groups havebeen examined, using either sequence or RFLP analysisof ribosomal DNA. However, there is no recent publi-cation in which the available data are combined to pre-sent a comprehensive classification scheme. Thus, theaim of this review is to compile the results of bothDNA-based and serological studies and present a pic-ture of the current status of phytoplasma differentiationand classification. This will be done by grouping thephytoplasmas phylogenetically and assigning other mol-ecularly characterized phytoplasmas, for which full-length 16S rDNA sequences were not available, to thegroups delineated.
PHYTOPLASMA PHYLOGENY
The first molecular indication of the mycoplasmal na-ture of the phytoplasmas was obtained by estimating theG+C content of highly enriched phytoplasmal DNA.The very low values obtained (23.0 to 26.2 mol %) sug-gested a close relationship to the culturable mollicutes(Kollar and Seemüller, 1989). More evidence came from
Journal of Plant Pathology (1998), 80 (1), 3-26 Edizioni ETS Pisa, 1998 3
INVITED REVIEW
CURRENT STATUS OF MOLECULAR CLASSIFICATION OF THE PHYTOPLASMASE. Seemüller 1, C. Marcone 1,2, U. Lauer 1, A. Ragozzino 2 and M. Göschl 3
1 Biologische Bundesanstalt, Institut für Pflanzenschutz im Obstbau, D-69221 Dossenheim, Germany2 Istituto di Patologia Vegetale, Università di Napoli ‘Federico II’, Via Università 100, I-80055 Portici, Napoli, Italy
3 LION Bioscience AG, Im Neuenheimer Feld 517, D-69120 Heidelberg, Germany

estimation of genome size, found by Neimark and Kirk-patrick (1993) to lie between 600 and 1150 kb, which iswithin the range for culturable mollicutes.
Evidence on the phylogeny of phytoplasmas can beobtained using comparative sequence analysis of theconserved 16S rRNA gene, which was proposed byWoese (1987) as a universal phylogenetic marker forclassifying major groups of prokaryotes. This approachhas been used to determine relationships of culturablemollicutes with each other and with walled bacteria(Weisburg et al., 1989).
Lim and Sears (1989) and Kuske and Kirkpatrick(1992) were the first to clone and completely sequence16S rRNA genes of phytoplasmas. Sequence compar-isons indicated that the two aster yellows (AY) phyto-plasmas which were examined are more closely relatedto culturable mollicutes than to other bacteria. Whenthe 16S rRNA gene became readily accessible by PCRamplification, Namba et al. (1993), Gundersen et al.(1994), and Seemüller et al. (1994) sequenced full-length or nearly full-length 16S rRNA genes of 37 rep-resentative strains. Comparison showed that these weremore closely related to each other than to any otherprokaryote, and indicated that the phytoplasmas repre-sent a monophyletic clade of organisms more closely re-lated to other mollicutes than to walled bacteria. Theclosest relatives of the phytoplasmas are some achole-plasmas such as A. palmae and A. modicum (Davis etal., 1997a) of the anaeroplasma clade as defined byWeisburg et al. (1989). These relationships have beenlargely confirmed by sequence analysis of two con-served ribosomal protein genes (Lim and Sears, 1992;Gundersen et al., 1994).
As a consequence of these studies, the InternationalCommittee of Systematic Bacteriology (ICSB) Subcom-mittee on the Taxonomy of Mollicutes (1993, 1997) hasagreed to replace the trivial name MLO by the termphytoplasma. This name will also, under the provisionaltaxonomic status Candidatus (Murray and Schleifer,1994), become the genus name of the plant pathogenicmycoplasmas.
METHODS TO DIFFERENTIATE AND CLASSIFYPHYTOPLASMAS
SYMPTOMATOLOGY AND HOST RANGE. Differentiationand classification of the culturable mollicutes is basedon both phenotypic and genotypic characteristics (ICSBSubcommittee on the Taxonomy of Mollicutes, 1995).
Since phytoplasmas have not been grown in axenic cul-ture, genotypic data are difficult to obtain and most ofthe phenotypic traits are still not applicable for theirclassification. Thus phytoplasmas were for a long timedescribed and differentiated according to the symptomsthey induce, the host plant affected, and sometimes alsothe geographic area where they occur (e.g. Europeanaster yellows). A particular role in symptomatologic clas-sification was played by the experimental host Catharan-thus roseus (periwinkle), to which many phytoplasmashave been experimentally transmitted and in which theyinduce specific symptoms. A classification of phytoplas-mas maintained in periwinkle was attempted by Marwitz(1990), and his grouping shows some similarities to thecurrent genetic classification. However, in several casesphytoplasmas were grouped together, which are geneti-cally distinctly different (e.g. the apple proliferation andX-disease agents), whereas others considered to be dif-ferent are genetically closely related (e.g. the agents ofAY and paulownia witches’-broom).
The specificity of insect vectors of phytoplasmas hasalso been used as a means of differentiation. For exam-ple, the two symptomatically similar phytoplasmoses af-fecting grapevine in France, flavescence dorée and boisnoir, were correctly thought to be caused by two differ-ent agents because the vector of flavescence dorée(Scaphoideus titanus) did not transmit bois noir (Caud-well et al., 1971). However, vector transmission is acomplex process, being a three-way interaction betweenpathogen, vector and host (Chiykowski and Sinha,1990), and is thus a difficult tool for use in taxonomy.
SEROLOGY. Polyclonal antisera and monoclonal anti-bodies have been produced against various phytoplas-mas (for reviews see Kirkpatrick, 1991; Lee and Davis,1992). Immunogens were usually extracted and en-riched from infected plants, less often from infested in-sect vectors. As antisera prepared with plant-derivedimmunogens often showed strong cross reactivity withplant antigens, and as insect-derived immunogens wererarely available, the production of specific, high-titerantisera proved to be difficult. Thus most of the anti-bodies available were prepared against isolates main-tained in high-titer hosts such as periwinkle or herba-ceous plants, and were suited to detect and identifyphytoplasmas in samples from hosts with a sufficientlyhigh titer. However, due to the comparatively highspecificity of antibodies, serology has provided onlylimited information about the interrelatedness of phyto-plasmas and none about relationships with culturablemollicutes and walled bacteria. Thus, the future role ofserological methods will mainly be the differentiation of
4 Phytoplasma classification Journal of Plant Pathology (1998), 80 (1), 3-26

closely related strains that were difficult to distinguishat the level of rDNA or other conserved sequences.
DOT AND SOUTHERN BLOT ANALYSIS. Randomlycloned fragments of chromosomal phytoplasmal DNAhave been widely used in dot blot hybridization assays(Kirkpatrick et al., 1987; Lee and Davis, 1988; Bonnetet al., 1990; Lee et al., 1991). The probes usuallyshowed a considerably broader detection range thanantibodies. Depending on the sequence selected, theprobes varied in specificity and allowed definition ofwider or narrower relationships. In particular by usingprobes from different phytoplasmas and with differentspecificity, it was possible to identify groups of geneti-cally different phytoplasmas and to distinguish phyto-plasmas within the groups.
Southern blot hybridization and analysis of the re-sulting RFLP patterns provided more information ongenetic relationships than dot blot assays, which arerarely used now. Strains that were indistinguishable bydot hybridization could be differentiated by RFLPanalysis (Bonnet et al., 1990). This method also revealedconsiderable genetic variation between AY strains(Kuske et al., 1991; Lee et al., 1992a; Schneider andSeemüller, 1994b) and between apple proliferation iso-lates (Kison et al., 1994), and could further distinguishfruit tree phytoplasmas within the apple proliferationgroup (Kison et al., 1997). Thus, using appropriate re-striction enzymes and probes, Southern blot analysis isstill an important tool in phytoplasma differentiation.Especially for the distinction of closely related strains, itmay allow finer differentiation than RFLP analysis ofPCR-amplified conserved sequences. Beside the limitedhybridization range of most probes, the major disad-vantage of Southern blotting is the insufficient sensitivi-ty of the probes in comparative work with phytoplas-mas from low-titer hosts.
RFLP ANALYSIS OF PCR-AMPLIFIED CONSERVEDDNA. In contrast to antibodies and hybridizationprobes, the 16S rRNA gene is universal in prokaryotesand possesses both conserved and variable regions thatmake it useful for taxonomic studies. After 16S rDNAbecame readily accessible by PCR amplification, RFLPanalysis of PCR-amplified rDNA was employed to com-prehensively classify the phytoplasmas on a molecularbasis for the first time. This work showed that similarphytoplasmas, e. g. from the same host, exhibit similarprofiles whereas isolates from different hosts or differentgeographic areas differ in their RFLP patterns. Usingfrequently cutting restriction endonucleases, such asAluI, RsaI and others, approximately 12 major groups
and several subgroups were distinguishable (Lee et al.,1993c; Schneider et al., 1993). At present, RFLP analysisof PCR-amplified rDNA is the method of choice for rou-tine differentiation and classification of phytoplasmas.As the information increases with the number of restric-tion sites, a large fragment is often preferred that com-prises the entire 16S rRNA gene and the 16S-23S rDNAspacer region (Schneider et al., 1995b). Differentiationcan also be improved by increasing the number of re-striction enzymes. Lee et al. (1993c) used 15 enzymes forthe classification of many diverse phytoplasmas.
However, from the data available, it appears thatclassification based on RFLP analysis of the 16S rRNAgene alone does not reflect the full range of phenotypicdiversity. This gene does not always seem sufficientlyvariable to allow distinction of phytoplasmas that differin plant host or vector specificity. More variable are nu-cleic acid sequences of ribosomal protein genes, whichhave enabled identification of more RFLP groups than16S rDNA data. The combined RFLP analysis of bothsequences resulted in the most detailed differentiationof the AY and X-disease group phytoplasmas so far(Gundersen et al., 1996). Another less conserved se-quence, recently examined, is the gene encoding theelongation factor Tu (tuf gene). Due to its variability, amore detailed subdivision of AY group phytoplasmaswas obtained than by 16S rDNA analysis (Schneider etal., 1997a). RFLP analysis of randomly cloned chromo-somal DNA fragments has also been employed, mainlyto differentiate closely related phytoplasmas (Jarauschet al., 1994; Daire et al., 1997).
SEQUENCE ANALYSIS OF CONSERVED DNA. Sequenceanalysis of the 16S rRNA gene is the standard methodfor the phylogenetic classification of prokaryotes. Majorefforts were undertaken by Namba et al. (1993), Gun-dersen et al. (1994), and Seemüller et al. (1994) in se-quencing full-length or nearly full-length 16S rRNAgenes of representative strains from major phytoplasmagroups established by RFLP analysis of 16S rDNA. Inthese and other studies, in which more than 40 phyto-plasmas were included, approximately 12 groups couldbe identified as distinct subclades of the phytoplasmaclade. These subclades are considered to represent theequivalent of distinct species (ICSB Subcommittee onthe Taxonomy of Mollicutes, 1997). The classificationbased on sequence analysis largely corresponds to thegrouping obtained by RFLP analysis of PCR-amplified16S rDNA. The few differences observed indicate thatthe sequence of a large molecule, such as the 16S rRNAgene, reflects phylogenetic distance more accuratelythan restriction patterns which depend on significantly
Journal of Plant Pathology (1998), 80 (1), 3-26 Seemüller et al. 5

fewer genetic characters. The two 16S rRNA geneswhich occur in phytoplasmas (Schneider andSeemüller, 1994a), seem to differ little from each otherand do not markedly affect the results of sequenceanalysis. The only pair of heterogeneous 16S rRNAgenes, which has been sequenced, differed at four nu-cleotide positions (Liefting et al., 1996).
Analysis of another conserved sequence, the spacerregion between the 16S and 23S rRNA genes, results in aclassification scheme similar to that derived from 16SrDNA data. Although the spacer region is less commonlyused in phylogenetic studies, it is an attractive moleculebecause it is considerably shorter than the 16S rRNAgene (220-250 bp versus ca. 1530 bp) and thus easier tosequence. In addition, the spacer region is, apart fromthe highly conserved tRNAIle gene that is apparently pre-sent in all phytoplasmas, less conserved than the 16SrDNA. For this reason, more detailed differentiation ispossible. More than 60 phytoplasma strains have beendifferentiated and classified by analysing the sequence oftheir 16S-23S spacer region (Kirkpatrick et al., 1994;Schneider et al., 1995b). Nucleotide sequence analysis ofthe conserved ribosomal protein L22 gene has also beenused. This fragment supported the designation of sub-clades and their phylogenetic positions within the phyto-plasma clade as obtained by 16S rDNA sequence analy-sis (Gundersen et al., 1994).
DIFFERENTIATION AND CLASSIFICATIONOF PHYTOPLASMAS
PHYLOGENETIC GROUPING. The taxonomy of the un-culturable phytoplasmas is a particular problem be-cause many criteria important in the taxonomy of cul-turable bacteria are only attainable with difficulty ornot at all. This applies mainly to suitable phenotypic in-formation. Thus, the criteria of modern bacterial taxon-omy as a polyphasic approach to classification, in whichgenotypic, phylogenetic, and phenotypic information isintegrated (Stackebrandt and Goebel, 1994), cannot beemployed in phytoplasmology. As similar problems ex-ist with other nonculturable prokaryotes, a formal clas-sification of such bacteria, which are extensively charac-terized by molecular techniques, has been proposed,using a Candidatus prefix (Murray and Schleifer, 1994).The Working Team on Phytoplasmas of the Interna-tional Research Programme of Comparative Mycoplas-mology (IRPCM) of the International Organization forMycoplasmology (IOM) has, at its meetings in 1994and 1996 at Bordeaux (France) and Orlando (Florida),proposed to adopt this new classification based on 16SrRNA gene sequences and describe each subclade (ma-
jor group) defined to the tentative species level as Can-didatus Phytoplasma species. The ICSB Subcommitteeon the Taxonomy of Mollicutes (1993, 1997) has agreedto this proposal and adopted the policy of basing phy-toplasma taxonomy on phylogeny. So far, two phyto-plasmas have been described, Candidatus Phytoplasmaaurantifolia (Zreik et al., 1995) and Candidatus P. aus-traliense (Davis et al., 1997a) (see also Table 2).
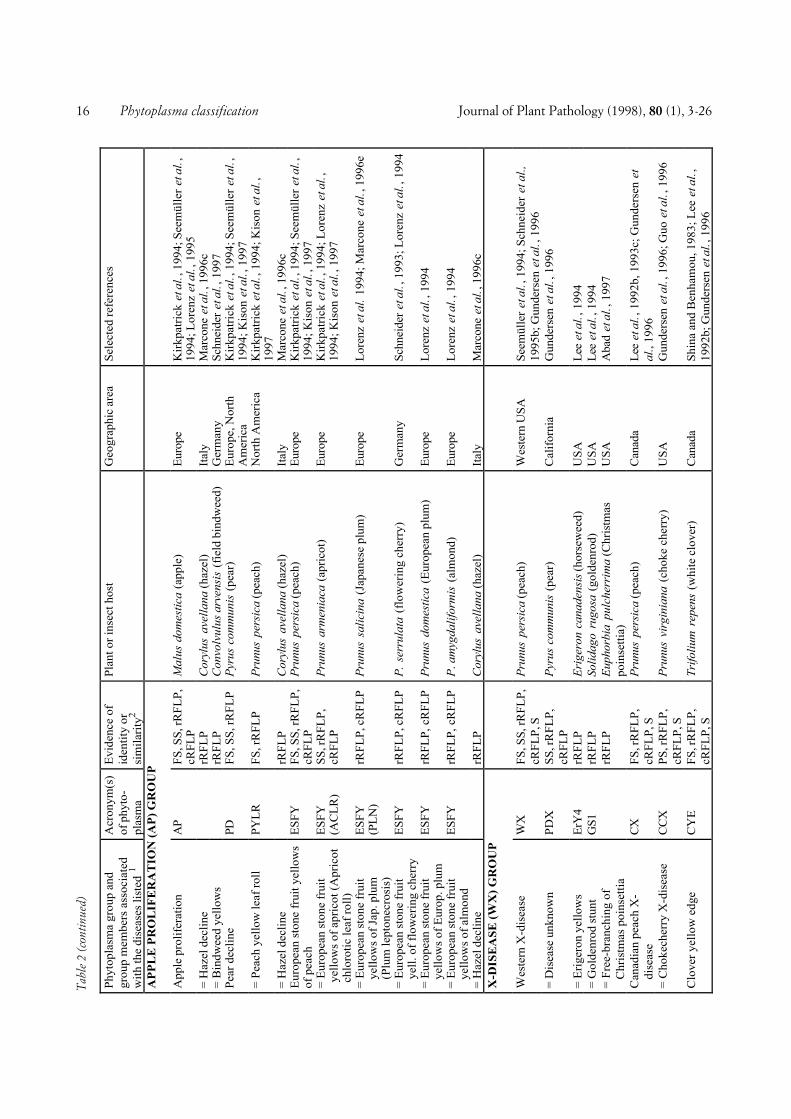
In our attempt at classification, 48 full-length ornearly full-length phytoplasma 16S rDNA sequencesavailable from the EMBL databank were examined. Inaddition, the 16S rRNA gene sequence of nine otherphytoplasmas was determined as described bySeemüller et al., (1994) and included in the analysis.These phytoplasmas were strains ALY, BGWL, EVY,IAWB, IBS, PDI, PEP, PYLR, and RuS (see Fig. 1 forabbreviations). Analysis of the 57 phytoplasmas re-vealed a greater diversity than known from previouswork, in which fewer phytoplasmas were examined. Byincluding all strains and by defining some groups in anarrower sense than previously, twenty distinct sub-clades consisting of groups of related phytoplasmas orsingle phytoplasmas were recognized (Fig. 1). The sin-gle-phytoplasma subclades are in the following also re-ferred to as groups. The subclades delineated includethe Australian grapevine yellows (AUSGY), Italianbindweed stolbur (IBS), stolbur (STOL), aster yellows(AY), buckthorn witches’-broom (BWB), spartiumwitches’-broom (SpaWB), apple proliferation (AP), X-disease (WX), Italian alfalfa witches’-broom (IAWB),faba bean phyllody (FBP), pigeon pea witches’-broom(PPWB), cirsium phyllody (CirP), sugarcane white leaf(SCWL), Bermuda grass white leaf (BGWL), Tanzan-ian lethal decline (TLD), lethal yellowing (LY), loofahwitches’-broom (LfWB), ash yellows (AshY), cloverproliferation (CP), and elm yellows (EY) groups.Groups AUSGY, IBS, BWB, SpaWB, IAWB, CirP,BGWL, and TLD were newly defined on the basis ofbranching patterns and by differences in the 16S rDNAsequence homology of at least 2.3% (AUSGY group)and 2.4% (IAWB group), or greater. The IRPCMWorking Team on Phytoplasmas had earlier agreed to athreshold of 2.5% to separate the stolbur phytoplasmagroup from the AY agents (Meetings 1994 and 1996 atBordeaux and Orlando, respectively).
Due to the finer subclade differentiation, the groupsas delineated here are less heterogeneous than in previ-ous schemes. Of the larger subclades, the AY, appleproliferation, and elm yellows groups are relatively ho-mogenous, differing in their levels of 16S rDNA se-quence homology by not more than 1.7%, 1.6%, and1.2%, respectively. Other subclades such as the sugar-
6 Phytoplasma classification Journal of Plant Pathology (1998), 80 (1), 3-26

cane white leaf, faba bean phyllody, and X-diseasegroups are still relatively heterogeneous. This applies inparticular for the sugarcane white leaf group, in whichthe evolutionary distance between the members variesbetween 2.3 and 2.7%. Thus, each of the group mem-bers may eventually be considered to be sufficiently dif-ferent from the others to be distinguished at the specieslevel. Within the X-disease and faba bean phyllodygroups the greatest differences in the 16S rDNA se-quence homology levels are 2.2 and 2.5%.
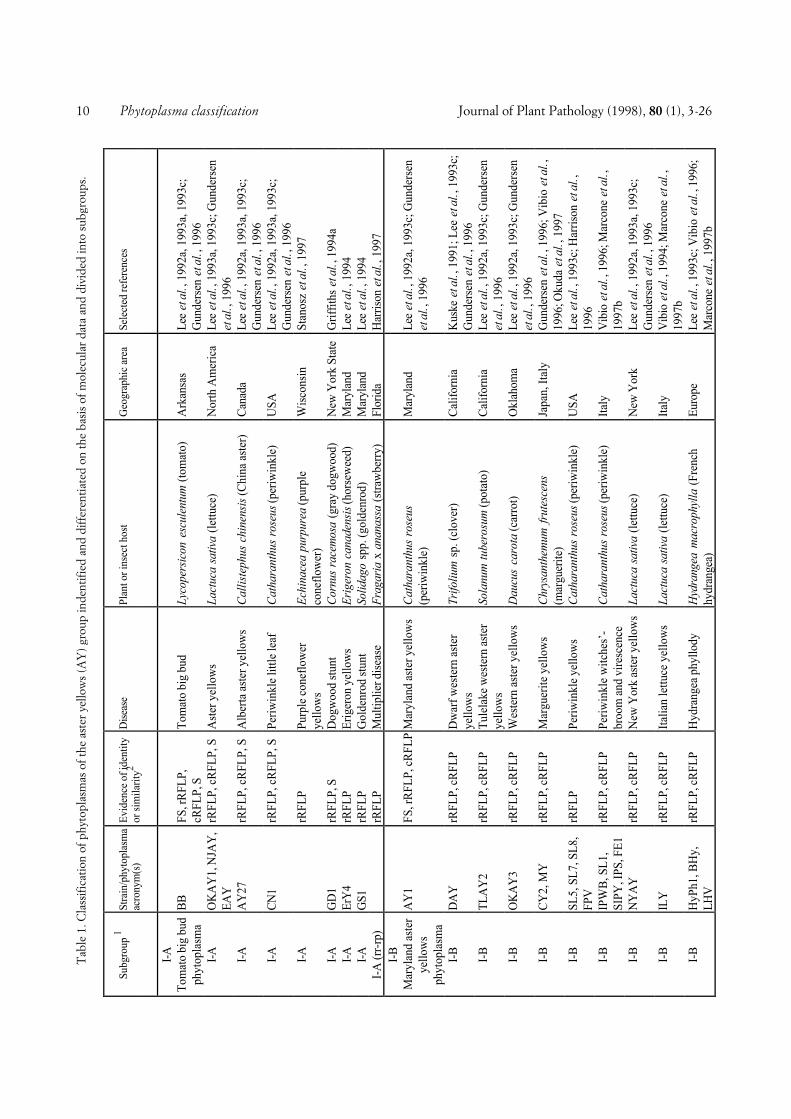
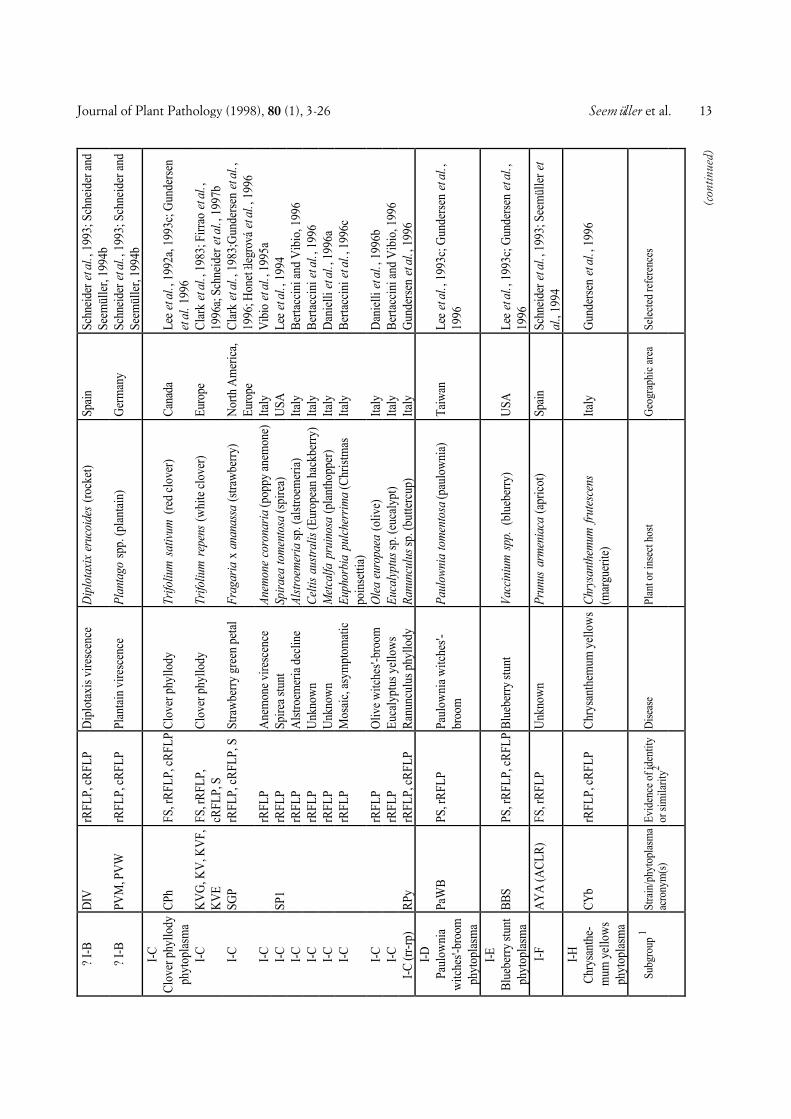
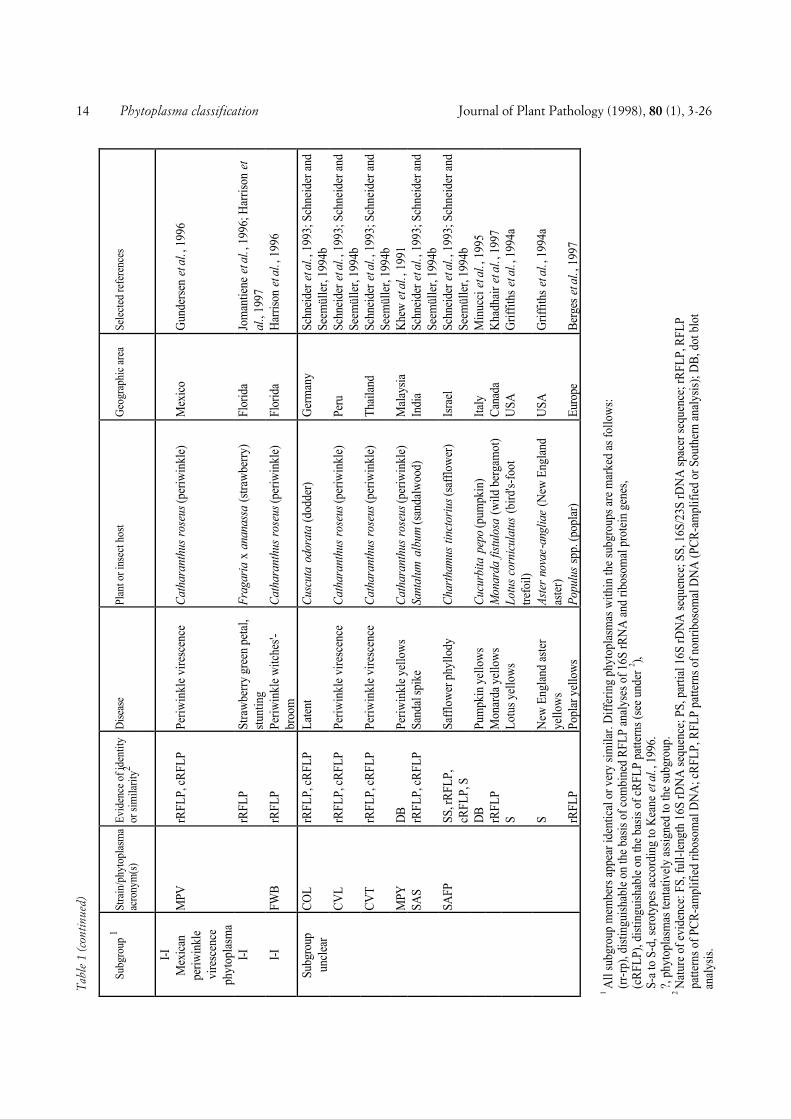
COMPREHENSIVE CLASSIFICATION AND DIFFERENTIA-TION. A total of 246 phytoplasmas (Tables 1 and 2) ex-amined using DNA-based and/or serological methodswere divided into major phylogenetic groups or sub-clades as described above. Table 1 shows 103 phyto-plasmas placed within the largest subclade, the AYgroup, grouped according to a scheme proposed by Leeet al. (1993c) and Gundersen et al. (1994, 1996), basedon 16S rDNA RFLP or putative restriction site analysis.In this scheme, 8 subgroups (I-A through I-I) are distin-guished. The number of phytoplasmas placed in varioussubgroups varies from only one (I-D, I-E, I-F, I-H) toseveral or many, I-B being by far the largest subgroup.All phytoplasmas of a subgroup are considered to bevery similar or identical, because they did not differ inthe comparisons made. However, within subgroup I-A,I-B, and I-C, a total of nine additional phytoplasmascan be distinguished from the other subgroup membersby combined RFLP analyses of 16S rRNA and riboso-mal protein gene sequences (Gundersen et al., 1996),Southern blot analysis, and/or serological comparison.Some of the AY phytoplasmas included in Table 1 werenot sufficiently characterized to be assigned to one ofthe established subgroups.
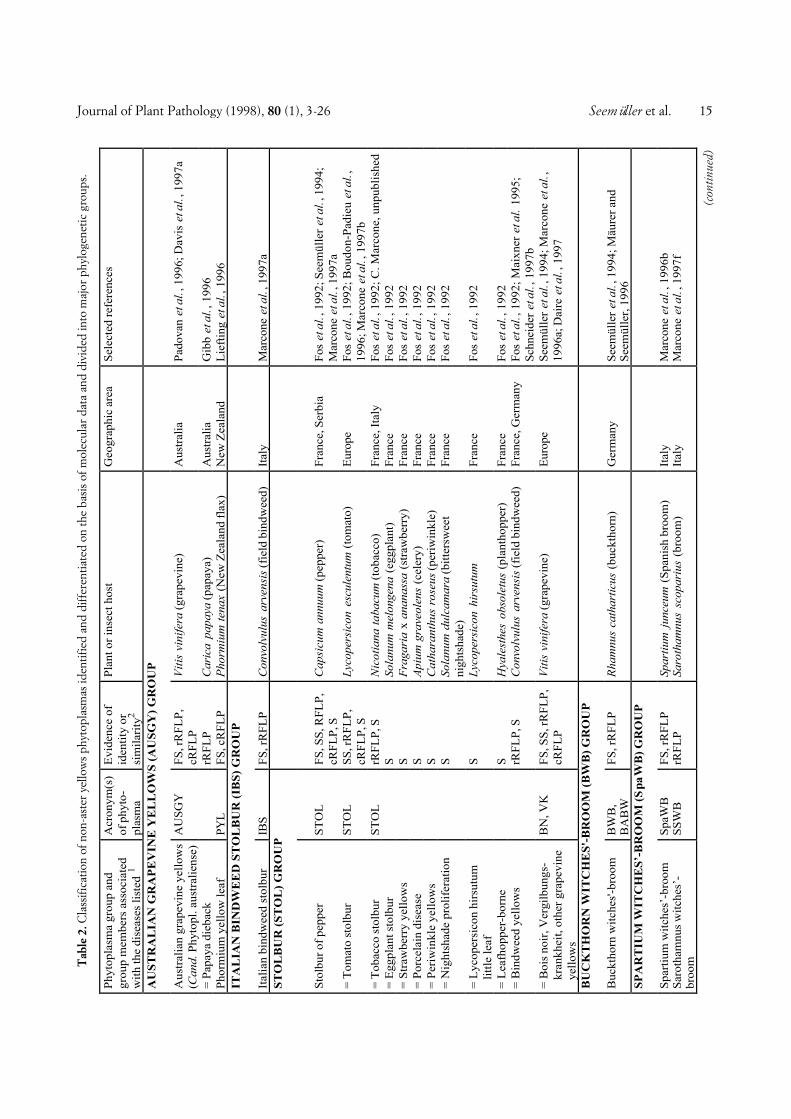
The 143 non-AY phytoplasmas are classified in a sim-ilar way, except that the major groups and the phyto-plasmas within the groups are named according to thediseases the phytoplasmas are associated with. Some-times a geographic attribute is included into the name tofurther specify the pathogen (e.g. Tanzanian coconutlethal decline phytoplasma). The non-AY phytoplasmasare divided into 19 major groups (Table 2). In mostgroups there were one or more phytoplasmas that werenot distinguishable from the taxon preceeding them inthe list so that they were treated as identical or very simi-lar. Among the non-AY phytoplasmas, 58 taxonomicunits were distinguishable, unevenly distributed amongthe groups. There are several groups consisting of onlyone taxon whereas in other groups, up to 11 taxa (X-dis-ease group) were differentiated. In Table 2, ten phyto-plasmas were included which were only partially charac-terized, so that only group assignment was possible.
GEOGRAPHIC DISTRIBUTION AND PLANTASSOCIATION
Phytoplasmas occur worldwide, but there are sig-nificant differences in the distribution of the variousgroups and subgroups. For example, AY subgroup I-A phytoplasmas have only been reported in NorthAmerica whereas members of subgroup I-B are wide-spread in North America, Europe and Japan. Phyto-plasmas of the stolbur and ash yellows groups haveonly been identified in Europe and North America, re-spectively, whereas phytoplasmas of the faba beanphyllody group are only known from Asia and Aus-tralia. Apple proliferation group phytoplasmas haveonly been reported in Europe with the exception ofthe peach yellow leaf roll agent, which occurs in west-ern USA. However, the current information about dis-tribution is likely to be just a temporary picture sub-ject to change with further research. There is still verylittle known about the occurrence of phytoplasmas invast areas including the former Soviet Union, Africa,and South America.
Tables 1 and 2 suggest that phytoplasmas may differconsiderably in host specificity. In the AY group, mem-bers of subgroup I-B appear to have a wide host rangesince this group occurs in many plant species. In con-trast, paulownia witches’-broom (subgroup I-D), blue-berry stunt (subgroup I-E), and maize bushy stunt phy-toplasmas (rr-rp variant of subgroup I-B) have eachbeen identified only in one plant species. The fruit treephytoplasmas of the apple proliferation group alsoseem to preferentially infect one host. The apple prolif-eration (Seemüller et al., 1994), pear decline, and Euro-pean stone fruit yellows phytoplasmas, from whichmore than one 16S rRNA gene has been sequenced,show nearly 100% homology whereas between the vari-ous pathogens clear differences exist. The ash yellowsphytoplasma also seems to have a narrow host range.Similarly, phytoplasmas of the lethal yellowing andTanzanian lethal decline groups appear to infect onlypalms. On the other hand, phytoplasmas of the fababean phyllody group, in particular the sunn hempwitches’-broom phytoplasma, occur in many plantspecies. The well-defined stolbur phytoplasma has alsobeen identified in many plant species.
However, host specificity is still not well understoodand thus differentiation of phytoplasmas at the hostspecificity level difficult. Also, it seems not always bepossible to distinguish phytoplasmas differing is hostspecificity by RFLP analysis of PCR-amplified con-served sequences. It is thus conceivable that the numer-
Journal of Plant Pathology (1998), 80 (1), 3-26 Seemüller et al. 7

ous isolates of AY subgroup I-B and the sunn hempwitches’-broom phytoplasma are not as homogeneousas they appear at present. Additional work using othermolecular markers or other methods might revealgreater genetic diversity. It may then become possibleto correlate genotype and host specificity more closely.However, as most or all phytoplasmas grow in periwin-kle and induce specific symptoms in this host, theremay be no strict host specificity in phytoplasmas. Hostrange may also depend on the interaction with insectvectors.
CONCLUSIONS AND OUTLOOK
This review reflects the tremendous progress made inthe last decade in differentiating and classifying the phy-toplasmas and elucidating their genetic diversity. It waspossible to divide the phytoplasmas examined into 20major groups, in which a total of 75 phytoplasmas couldbe distinguished. The major groups are distinctly differ-ent and are planned, according to the consensusreached, to be delineated to the species level. Such a for-mal description as Candidatus species will undoubtedlyimprove the current situation, complicated by confusinghistorical and preliminary classification schemes. How-ever, the numbers of distinct taxa within the groups isconstantly increasing. Therefore, despite the narrowerdefinition of some major groups proposed in this paper,some of the subclades, in particular the AY, faba beanphyllody, X-disease, sugarcane white leaf, and appleproliferation groups, are genetically and/or pathological-ly still too diverse, and they cannot each be described asa single taxon, in a satisfactory working sense.
Therefore, the current concept for phytoplasma clas-sification should be expanded and adapted to the exist-ing diversity. One possibility would be to define eachgroup as narrowly as possible. We also propose to de-scribe only those major groups as Candidatus speciesthat consist of only one taxon, or of phytoplasmas thatare very closely related. For the other groups, consistingof several distinct phytoplasmas, all clearly different tax-onomic units within the groups should be described sep-arately, preferably as species. Examples of such addition-al putative taxa within subclades are the rice yellowdwarf phytoplasma and strain BVK of the sugarcanewhite leaf group, the European stone fruit yellows andpear decline phytoplasmas of the apple proliferationgroup, and the tsuwabuki witches’-broom phytoplasmaof the X-disease group. Other taxa which are moreclosely related to a phytoplasma described as a tentativespecies may be distinguished as a subspecies althoughdistinction at the species level or the use of infrasubspe-
cific ranks such as pathovar, serovar and strain is pre-ferred (ICSB Subcommittee on the Taxonomy of Molli-cutes, 1995). However, at the moment it is unclear howmany of the molecularly distinguishable phytoplasmaswill eventually be considered to be sufficiently differentto be described at one of the levels discussed. One mark-er, such as a single conserved restriction site, may not al-ways be sufficient to define a useful taxonomic unit.
According to the criteria recommended by the ICSB,bacteria belonging to the same species should exhibit atleast 70% homology of their total DNA (Wayne et al.,1987). However, DNA homologies between the noncul-turable phytoplasmas are difficult to determine, and sothe sequence of the conserved 16S rRNA gene was se-lected as the basis for phytoplasma classification. Un-fortunately, there is no clear correlation between se-quence homology of the 16S rDNA gene and that ofthe entire genome, and there is no threshold level de-fined for designating species on the basis of 16S rDNAsequence similarity. Comparisons show that bacteriasharing 16S rDNA similarity of about 99% may vary inthe homology of their total DNA between 23.5 and93%. However, most of the species that have been ex-amined differ in their 16S rDNA sequence from relatedspecies of the same genus in at least 1.5% of the se-quence positions, and usually more (Fox et al., 1992).Thus, threshold levels of less than 2.5% for definingspecies may be appropriate to classify the pathologicallydiverse but phylogenetically coherent phytoplasmas,which differ in their 16S rDNA sequence by not morethan 14.0%. This evolutionary distance corresponds tothat within the phylogenetic groups of the culturablemollicutes as defined by Weisburg et al. (1989), whichconsist of much fewer taxonomic units. Thus, differ-ences in the 16S rDNA sequence may not be sufficientto be the sole basis for phytoplasma classification, inwhich many distinct taxonomic units are phylogeneti-cally closely related. For their differentiation, additionalcriteria may have to be included such as less conservedgenes, pathological aspects, and geographical distribu-tion. However, only such phylogenetically closely relat-ed taxa should be delineated which are genetically andpathologically adequately characterized. Any taxonomicwork should be done with caution, keeping the conser-vative nature of any classification system in mind andtaking practical considerations into account.
ACKNOWLEDGEMENTS
We thank Bernd Schneider for critical reading of themanuscript and valuable suggestions, and Robert Milnefor improving and correcting the text.
8 Phytoplasma classification Journal of Plant Pathology (1998), 80 (1), 3-26

Journal of Plant Pathology (1998), 80 (1), 3-26 Seemüller et al. 9
Fig. 1. Phylogenetic dendrogram of the phytoplasmas examined, generated by using the Phylogeny Inference Package (PHYLIP), Version 3.57c (J.Felsenstein, Department of Genetics, University of Washington, Seattle, WA 98195). The bar represents a phylogenetic distance of 10%. Phyloge-netic groups defined are shown in the right hand column. Group abbreviations correspond to the acronyms of the representative strains of thegroups (see also text). Phytoplasma strain abbreviations and sequence accession numbers (from bottom): PYL, phormium yellow leaf (U43569); P.austral., Candidatus Phytoplasma australiense (AUSGY) (L76865); IBS, Italian bindweed stolbur (Y16391); VK, Vergilbungskrankheit (grapevineyellows) (X76428); STOL, stolbur of pepper (X76427); KVG, clover phyllody/Germany (X83870); CPh, clover phyllody/Canada (L33762); BB,tomato big bud/Arkansas (L33760); AYA (formerly ACLR), aster yellows from apricot (X68338); OAY, Oenothera aster yellows (M30790); RP,rape phyllody (U89378); OY, onion yellows (D12569); AY1, Maryland aster yellows (L33767); AAY, American aster yellows (X68373); SAY, west-ern severe aster yellows (M86340); BWB, buckthorn witches’-broom (X76431); SpaWB (formerly SPAR), spartium witches’-broom (X92869); PD,pear decline/Germany (X76425); PYLR, peach yellow leaf roll (Y16394); PDI, pear decline/Italy (Y16392); AT, apple proliferation (X68375); ES-FY, European stone fruit yellows/Germany (X68374); ESFYC, European stone fruit yellows/Czech Republic (Y11933); CX, Canadian X-disease(L33733); WX, western X-disease (L04682); TWB, tsuwabuki witches’-broom (D12580); VAC, vaccinium witches’-broom (X76430); ICPh, Italianclover phyllody (X77482); CYE, clover yellow edge (L33766); IAWB, Italian alfalfa witches’-broom (Y16390); PEP, picris echioides phyllody(Y16393); FBP, faba bean phyllody (X83432); P.aurant., Candidatus P. aurantifolia (WBDL), (U15442); TBB, tomato big bud/Australia (Y08173);SUNHP, sunn hemp witches’-broom (X76433); PnWB, peanut witches’-broom (L33765); SPWB, sweet potato witches’-broom (L33770); EVY,echium vulgare yellows (Y16389); CPPWB, Caribbean pigeon pea witches’-broom (U18763); PPWB, pigeon pea witches’-broom (L33735); CirP,cirsium phyllody (X83438); BVK, from leafhopper Psammotettix cephalotes (X76429); RYD, rice yellow dwarf (D12581); SCWL, sugarcane whiteleaf (X76432); BGWL, Bermuda grass white leaf (Y16388); TLD, Tanzanian lethal decline of coconut (X80117); LY, coconut lethal yellowing(U18747); YLD, Yucatan lethal decline of coconut (U18753); LfWB, loofah witches’-broom (L33764); AshY, ash yellows (X68339); BLL, brinjallittle leaf (X83431); CP, clover proliferation (L33761); EY, elm yellows/USA (L33763); ULW, elm yellows/France (X68376); ALY, alder yellows(Y16387); RuS, rubus stunt (Y16395); FD, flavescence dorée (X76560). Clostridium innocuum was used as an outgroup and three Acholeplasmaspecies were included for comparison. See Tables 1 and 2 for further information about the phytoplasmas examined.
10 Phytoplasma classification Journal of Plant Pathology (1998), 80 (1), 3-26
Tab
le 1
. Cla
ssifi
catio
n of
phy
topl
asm
as o
f the
ast
er y
ello
ws
(AY
) gro
up in
dent
ified
and
diff
eren
tiate
d on
the
basi
s of
mol
ecul
ar d
ata
and
divi
ded
into
sub
grou
ps.
Subg
roup
1St
rain
/phy
topl
asm
aac
rony
m(s
)E
vide
nce
of id
entit
yor
sim
ilari
ty2
Dis
ease
Plan
t or i
nsec
t hos
tG
eogr
aphi
c ar
eaSe
lect
ed re
fere
nces
I-A
Tom
ato
big
bud
phyt
opla
sma
BB
FS, r
RFL
P,cR
FLP,
ST
omat
o bi
g bu
dLy
cope
rsic
on e
scul
entu
m (t
omat
o)A
rkan
sas
Lee
et a
l., 1
992a
, 199
3a, 1
993c
;G
unde
rsen
et a
l., 1
996
I-A
OK
AY
1, N
JAY
,EA
YrR
FLP,
cR
FLP,
SA
ster
yel
low
sLa
ctuc
a sa
tiva
(let
tuce
)N
orth
Am
eric
aLe
eet
al.
, 199
3a, 1
993c
; Gun
ders
enet
al.
, 199
6I-
AA
Y27
rRFL
P, c
RFL
P, S
Alb
erta
ast
er y
ello
ws
Cal
liste
phus
chi
nens
is (C
hina
ast
er)
Can
ada
Lee
et a
l., 1
992a
, 199
3a, 1
993c
;G
unde
rsen
et a
l., 1
996
I-A
CN
1rR
FLP,
cR
FLP,
SPe
riw
inkl
e lit
tle le
afC
atha
rant
hus
rose
us (p
eriw
inkl
e)U
SALe
eet
al.
, 199
2a, 1
993a
, 199
3c;
Gun
ders
enet
al.
, 199
6I-
ArR
FLP
Purp
le c
onef
low
erye
llow
sE
chin
acea
pur
pure
a (p
urpl
eco
nefl
ower
)W
isco
nsin
Stan
osz
et a
l., 1
997
I-A
GD
1rR
FLP,
SD
ogw
ood
stun
tC
ornu
s ra
cem
osa
(gra
y do
gwoo
d)N
ew Y
ork
Stat
eG
riff
iths
et a
l., 1
994a
I-A
ErY
4rR
FLP
Eri
gero
n ye
llow
sE
rige
ron
cana
dens
is (h
orse
wee
d)M
aryl
and
Lee
et a
l., 1
994
I-A
GS1
rRFL
PG
olde
nrod
stu
ntSo
lidag
o sp
p. (g
olde
nrod
)M
aryl
and
Lee
et a
l., 1
994
I-A
(rr-
rp)
rRFL
PM
ultip
lier d
isea
seF
raga
ria
xan
anas
sa (s
traw
berr
y)Fl
orid
aH
arris
onet
al.
, 199
7I-
BM
aryl
and
aste
rye
llow
sph
ytop
lasm
a
AY
1FS
, rR
FLP,
cR
FLP
Mar
ylan
d as
ter y
ello
ws
Cat
hara
nthu
s ro
seus
(per
iwin
kle)
Mar
ylan
dLe
eet
al.
, 199
2a, 1
993c
; Gun
ders
enet
al.
, 199
6
I-B
DA
YrR
FLP,
cR
FLP
Dw
arf w
este
rn a
ster
yello
ws
Trifo
lium
sp.
(clo
ver)
Cal
ifor
nia
Kus
keet
al.
, 199
1; L
eeet
al.
, 199
3c;
Gun
ders
enet
al.
, 199
6I-
BT
LA
Y2
rRFL
P, c
RFL
PT
ulel
ake
wes
tern
ast
erye
llow
sSo
lanu
m tu
bero
sum
(pot
ato)
Cal
ifor
nia
Lee
et a
l., 1
992a
, 199
3c; G
unde
rsen
et a
l., 1
996
I-B
OK
AY
3rR
FLP,
cR
FLP
Wes
tern
ast
er y
ello
ws
Dau
cus
caro
ta (c
arro
t)O
klah
oma
Lee
et a
l., 1
992a
, 199
3c; G
unde
rsen
et a
l., 1
996
I-B
CY
2, M
YrR
FLP,
cR
FLP
Mar
guer
ite y
ello
ws
Chr
ysan
them
um fr
utes
cens
(mar
guer
ite)
Japa
n, It
aly
Gun
ders
enet
al.
, 199
6; V
ibio
et a
l.,
1996
; Oku
daet
al.
, 199
7I-
BSL
5, S
L7,
SL
8,FP
VrR
FLP
Peri
win
kle
yello
ws
Cat
hara
nthu
s ro
seus
(per
iwin
kle)
USA
Lee
et a
l., 1
993c
; Har
riso
net
al.
,19
96I-
BIP
WB
, SL
1,SI
PY, I
PS, F
E1
rRFL
P, c
RFL
PPe
riw
inkl
e w
itche
sÕ-
broo
m a
nd v
ires
cenc
eC
atha
rant
hus
rose
us (p
eriw
inkl
e)It
aly
Vib
ioet
al.
, 199
6; M
arco
neet
al.
,19
97b
I-B
NY
AY
rRFL
P, c
RFL
PN
ew Y
ork
aste
r yel
low
sLa
ctuc
a sa
tiva
(let
tuce
)N
ew Y
ork
Lee
et a
l., 1
992a
, 199
3a, 1
993c
;G
unde
rsen
et a
l., 1
996
I-B
ILY
rRFL
P, c
RFL
PIt
alia
n le
ttuce
yel
low
sLa
ctuc
a sa
tiva
(let
tuce
)It
aly
Vib
ioet
al.
, 199
4; M
arco
neet
al.
,19
97b
I-B
HyP
h1, B
Hy,
LHV
rRFL
P, c
RFL
PH
ydra
ngea
phy
llody
Hyd
rang
ea m
acro
phyl
la (F
renc
hhy
dran
gea)
Euro
peLe
eet
al.
, 199
3c; V
ibio
et a
l., 1
996;
Mar
cone
et a
l., 1
997b

Journal of Plant Pathology (1998), 80 (1), 3-26 Seemüller et al. 11
(con
tinu
ed)
I-B
AA
YFS
, rR
FLP,
Am
eric
an a
ster
yel
low
sFl
orid
aSc
hnei
dere
t al.,
199
3; S
eem
�lle
ret
al.,
1994
; Mar
cone
et a
l., 1
997a
I-B
OY
PS, S
S, rR
FLP
Oni
on y
ello
ws a
ndvi
resc
ence
Alliu
m c
epa
(oni
on)
Japa
n, It
aly
Nam
baet
al.,
199
3; S
em�l
lere
t al.,
1994
; Vib
ioet
al.,
199
5bI-
BG
IYrR
FLP,
cR
FLP
Gla
diol
us y
ello
ws
Gla
diol
us h
ybrid
(gla
diol
us)
Euro
peB
erta
ccin
i and
Vib
io, 1
996;
Vib
ioet
al.,
1996
I-B
ICY
rRFL
PIta
lian
cabb
age
yello
ws
Bras
sica
ole
race
a va
r.ca
pita
ta(c
abba
ge)
Italy
Mar
cone
et a
l., 1
997b
Ber
tacc
ini a
nd V
ibio
, 199
6I-
BIB
PrR
FLP
Bro
ccol
i phy
llody
Bras
sica
ole
race
a va
r.ita
lica
(spr
outin
g br
occo
li)Ita
lyM
arco
neet
al.,
199
7b
I-B
SIK
PrR
FLP
Kal
e ph
yllo
dyBr
assi
ca o
lera
cea
var.
palm
ifolia
(kal
e)Ita
lyM
arco
neet
al.,
199
7b
I-B
TPV
rRFL
PTu
rnip
vire
scen
ceBr
assi
ca r
apa
var.
rapi
fera
(tur
nip)
Italy
Mar
cone
et a
l., 1
997b
I-B
rRFL
PB
ig b
udLy
cope
rsic
on e
scul
entu
m (t
omat
o)Ita
ly, J
apan
Mar
cone
et a
l., 1
997b
; Oku
daet
al.,
1997
I-B
SIC
VrR
FLP
Cal
endu
la v
iresc
ence
Cal
endu
la o
ffici
nalis
(pot
mar
igol
d)Ita
lyM
arco
neet
al.,
199
7aI-
BPV
rRFL
PPa
pave
r vire
scen
cePa
pave
r rh
oeas
(cor
n po
ppy)
Italy
Mar
cone
et a
l., 1
997c
I-B
WR
YrR
FLP
Wild
radi
sh y
ello
ws
Raph
anus
rap
hani
stru
m (w
ild ra
dish
)Ita
lyM
arco
neet
al.,
199
7cI-
BPO
YrR
FLP
Portu
laca
yel
low
sPo
rtul
aca
oler
acea
(pur
slan
e)Ita
lyM
arco
neet
al.,
199
7c; S
chne
ider
etal
., 19
97b
I-B
rRFL
PC
ycla
men
vire
scen
ceC
ycla
men
per
sicu
m (c
ycla
men
)G
erm
any
Schn
eide
ret a
l., 1
997b
I-B
ErY
rRFL
PEr
iger
on y
ello
ws
Erig
eron
can
aden
sis (
hors
ewee
d)U
SALe
eet
al.,
199
4I-
BrR
FLP
Als
troem
eria
dec
line
Alst
roem
eria
sp. (
alst
roem
eria
)Ita
lyB
erta
ccin
i and
Vib
io, 1
996
I-B
rRFL
PR
anun
culu
s phy
llody
Ranu
ncul
us a
siat
icus
(Per
sian
butte
rcup
)Ita
lyV
ibio
et a
l., 1
997
I-B
rRFL
PM
allo
w y
ello
ws
Mal
va s
p. (m
allo
w)
Italy
Vib
ioet
al.,
199
7I-
BrR
FLP,
cR
FLP
Rag
wee
d ye
llow
sAm
bros
ia a
rtem
isiif
olia
(com
mon
ragw
eed)
Flor
ida
Har
rison
et a
l., 1
996
I-B
rRFL
P, c
RFL
PD
ogfe
nnel
yel
low
sEu
pato
rium
cap
illifo
lium
(dog
fenn
el)
Flor
ida
Har
rison
et a
l., 1
996
I-B
rRFL
PPo
plar
witc
hes'-
broo
mPo
pulu
s ni
gra
'Ital
ica'
(Lom
bard
ypo
plar
)Fr
ance
Ber
ges
et a
l., 1
997
I-B
JHW
rRFL
PM
itsub
a w
itche
s'-br
oom
Cry
ptot
aeni
a ja
poni
ca (J
apan
ese
hone
wor
t)Ja
pan
Oku
daet
al.,
199
7
I-B
MD
FSM
ulbe
rry
dwar
fM
orus
bom
byci
s (m
ulbe
rry)
Japa
nN
amba
et a
l., 1
993
I-B
GC
WrR
FLP
Chr
ysan
them
umw
itche
s'-br
oom
Chr
ysan
them
um c
oron
ariu
m(c
row
n da
isy)
Japa
nO
kuda
et a
l., 1
997
I-B
EDrR
FLP
Eggp
lant
dw
arf
Sola
num
mel
onge
na (e
ggpl
ant)
Japa
nO
kuda
et a
l., 1
997
I-B
rRFL
PU
nkno
wn
Prun
us a
rmen
iaca
(apr
icot
)Ita
lyLe
eet
al.,
199
5a

12 Phytoplasma classification Journal of Plant Pathology (1998), 80 (1), 3-26
Tab
le 1
(con
tinu
ed)
Subg
roup
1St
rain
/phy
topl
asm
aac
rony
m(s
)Ev
iden
ce o
f ide
ntity
or s
imila
rity2
Dis
ease
Plan
t or i
nsec
t hos
tG
eogr
aphi
c ar
eaSe
lect
ed re
fere
nces
I-B
rRFL
PU
nkno
wn
Prun
us p
ersi
ca v
ar.n
ecta
rina
(nec
tarin
e)Ita
lyLe
eet
al.,
199
5a
I-B
rRFL
PU
nkno
wn
Prun
us s
alic
ina
(Jap
anes
e pl
um)
Italy
Lee
et a
l., 1
995a
I-B
rRFL
PU
nkno
wn
Trifo
lium
pra
tens
e (r
ed c
love
r)C
anad
aLe
eet
al.,
199
4I-
BrR
FLP
Unk
now
nSo
lanu
m tu
bero
sum
(pot
ato)
Can
ada
Lee
et a
l., 1
994
I-B
rRFL
PU
nkno
wn
Cel
tis a
ustr
alis
(Eur
opea
n ha
ckbe
rry)
Italy
Ber
tacc
inie
t al.,
199
6bI-
BrR
FLP
Unk
now
nU
lmus
sp.
(elm
)Ita
lyLe
eet
al.,
199
5bI-
BrR
FLP
Gra
pevi
ne y
ello
ws
Vitis
vin
ifera
(gra
pevi
ne)
Italy
Alm
aet
al.,
199
6I-
BrR
FLP
Unk
now
nM
etca
lfa p
ruin
osa
(pla
ntho
pper
)Ita
lyD
anie
lliet
al.,
199
6aI-
BrR
FLP
Mos
aic,
Fre
e-br
anch
ing,
asym
ptom
atic
Euph
orbi
a pu
lche
rrim
a (C
hris
tmas
poin
setti
a)Ita
lyB
erta
ccin
iet a
l., 1
996a
, 199
6c
I-B
rRFL
PO
live
witc
hes'-
broo
mO
lea
euro
paea
(oliv
e)Ita
lyD
anie
lliet
al.,
199
6bI-
BrR
FLP
Euca
lypt
us y
ello
ws
Euca
lypt
us sp
p. (e
ucal
ypt)
Italy
Ber
tacc
ini a
nd V
ibio
, 199
6I-
BrR
FLP
Myr
tle y
ello
ws
Myr
tus
com
mun
is (m
yrtle
)Ita
lyB
erta
ccin
i and
Vib
io, 1
996
I-B
(rr-
rp)
IOW
BrR
FLP,
cR
FLP
Ipom
oea
obsc
ura
witc
hes'-
broo
mIp
omoe
a ob
scur
aTa
iwan
Lee
et a
l., 1
993c
; Gun
ders
enet
al.,
1996
I-B
(rr-
rp)
MB
SrR
FLP,
cR
FLP
Mai
ze b
ushy
stu
ntZe
a m
ays
(cor
n)M
exic
o, O
hio
Gun
ders
enet
al.,
199
6I-
B (c
RFL
P)O
AY
FS, S
S, rR
FLP,
cRFL
PO
enot
hera
vire
scen
ceO
enot
hera
hoo
keri
(eve
ning
prim
rose
)M
ichi
gan
Lim
and
Sea
rs, 1
989;
Kus
keet
al.
1991
; Gun
ders
enet
al.,
199
6I-
B (c
RFL
P, S
-a)
CL,
RV
rRFL
P, c
RFL
P, S
Chl
oran
tieBr
assi
ca n
apus
(rap
e)Fr
ance
Kus
keet
al.
1991
; Kea
neet
al.,
199
6;V
ibio
et a
l., 1
996;
I-B
(S-a
)SA
YFS
, SS,
rRFL
P,cR
FLP,
SSe
vere
wes
tern
ast
erye
llow
sAp
ium
gra
veol
ens
(cel
ery)
Cal
iforn
iaK
uske
and
Kirk
patri
ck, 1
992;
Lee
etal
., 19
93c;
Kea
neet
al.,
199
6I-
B (S
-a)
CC
rRFL
P, c
RFL
P, S
Cac
tus v
iresc
ence
Opu
ntia
sp.
(cac
tus)
UK
Kea
neet
al.,
199
6; V
ibio
et a
l., 1
996
I-B
(S-b
)PY
rRFL
P, c
RFL
P, S
Prim
ula
yello
ws
Prim
ula
sp. (
prim
rose
)G
erm
any
Kea
neet
al.,
199
6; V
ibio
et a
l., 1
996
I-B
(S-c
)K
DrR
FLP,
cR
FLP,
SK
ools
ard
Bras
sica
sp. (
cabb
age)
UK
Kea
neet
al.,
199
6; V
ibio
et a
l., 1
996
I-B
(cR
FLP,
S-
d)EA
Y, A
V21
92rR
FLP,
cR
FLP,
SS, S
Euro
pean
ast
er y
ello
ws
Cal
liste
phus
chi
nens
is (C
hina
ast
er)
Ger
man
yK
uske
et a
l., 1
991;
Kea
neet
al.,
1996
; Vib
ioet
al.,
199
6?
I-B
rRFL
PC
arda
ria p
hyllo
dyC
arda
ria
drab
a (h
oary
cre
ss)
Ger
man
ySc
hnei
der
et a
l., 1
997b
? I-
BrR
FLP
Bun
ias p
hyllo
dyBu
nias
ori
enta
lis (h
ill m
usta
rd)
Ger
man
ySc
hnei
der
et a
l., 1
997b
? I-
BrR
FLP
Stel
laria
yel
low
sSt
ella
ria
med
ia (c
omm
on c
hick
wee
d)G
erm
any
Schn
eide
ret
al.,
199
7b?
I-B
rRFL
PC
arro
t pro
lifer
atio
nD
aucu
s ca
rota
(car
rot)
Ger
man
ySc
hnei
der
et a
l., 1
997b
? I-
BA
KV
rRFL
P, c
RFL
PC
olum
bine
vire
scen
ceAq
uile
gia
alpi
na (c
olum
bine
)G
erm
any
Schn
eide
ret
al.,
199
3; S
chne
ider
and
Seem
�lle
r, 19
94b
? I-
BD
EVrR
FLP,
cR
FLP
Lark
spur
vire
scen
ceD
elph
iniu
m h
ybrid
(lar
kspu
r)G
erm
any
Schn
eide
ret
al.,
199
3; S
chne
ider
and
Seem
�lle
r, 19
94b
Journal of Plant Pathology (1998), 80 (1), 3-26 Seemüller et al. 13
(con
tinu
ed)
? I-
BD
IVrR
FLP,
cR
FLP
Dip
lota
xis v
iresc
ence
Dip
lota
xix
eruc
oide
s (r
ocke
t)Sp
ain
Schn
eide
ret a
l., 1
993;
Sch
neid
er a
ndSe
em�l
ler,
1994
b?
I-B
PVM
, PV
WrR
FLP,
cR
FLP
Plan
tain
vire
scen
cePl
anta
go sp
p. (p
lant
ain)
Ger
man
ySc
hnei
dere
t al.,
199
3; S
chne
ider
and
Seem
�lle
r, 19
94b
I-C
Clo
ver p
hyllo
dyph
ytop
lasm
aC
PhFS
, rR
FLP,
cR
FLP
Clo
ver p
hyllo
dyTr
ifoliu
m s
ativ
um (
red
clov
er)
Can
ada
Lee
et a
l., 1
992a
, 199
3c; G
unde
rsen
et a
l. 19
96I-
CK
VG
, KV
, KV
F,K
VE
FS, r
RFL
P,cR
FLP,
SC
love
r phy
llody
Trifo
lium
rep
ens
(whi
te c
love
r)Eu
rope
Cla
rket
al.,
198
3; F
irrao
et a
l.,19
96a;
Sch
neid
eret
al.,
199
7bI-
CSG
PrR
FLP,
cR
FLP,
SSt
raw
berr
y gr
een
peta
lFr
agar
ia x
anan
assa
(stra
wbe
rry)
Nor
th A
mer
ica,
Euro
peC
lark
et a
l., 1
983;
Gun
ders
enet
al.,
1996
; Hon
etłl
egro
v�et
al.,
199
6I-
CrR
FLP
Ane
mon
e vi
resc
ence
Anem
one
coro
nari
a (p
oppy
ane
mon
e)Ita
lyV
ibio
et a
l., 1
995a
I-C
SP1
rRFL
PSp
irea
stun
tSp
irae
a to
men
tosa
(spi
rea)
USA
Lee
et a
l., 1
994
I-C
rRFL
PA
lstro
emer
ia d
eclin
eAl
stro
emer
ia sp
. (al
stro
emer
ia)
Italy
Ber
tacc
ini a
nd V
ibio
, 199
6I-
CrR
FLP
Unk
now
nC
eltis
aus
tral
is (E
urop
ean
hack
berr
y)Ita
lyB
erta
ccin
iet a
l., 1
996
I-C
rRFL
PU
nkno
wn
Met
calfa
pru
inos
a (p
lant
hopp
er)
Italy
Dan
ielli
et a
l., 1
996a
I-C
rRFL
PM
osai
c, a
sym
ptom
atic
Euph
orbi
a pu
lche
rrim
a (C
hris
tmas
poin
setti
a)Ita
lyB
erta
ccin
iet a
l., 1
996c
I-C
rRFL
PO
live
witc
hes'-
broo
mO
lea
euro
paea
(oliv
e)Ita
lyD
anie
lliet
al.,
199
6bI-
CrR
FLP
Euca
lypt
us y
ello
ws
Euca
lypt
us sp
. (eu
caly
pt)
Italy
Ber
tacc
ini a
nd V
ibio
, 199
6I-
C (r
r-rp
)R
PyrR
FLP,
cR
FLP
Ran
uncu
lus p
hyllo
dyRa
nunc
ulus
sp. (
butte
rcup
)Ita
lyG
unde
rsen
et a
l., 1
996
I-DPa
ulow
nia
witc
hes'-
broo
mph
ytop
lasm
a
PaW
BPS
, rR
FLP
Paul
owni
a w
itche
s'-br
oom
Paul
owni
a to
men
tosa
(pau
low
nia)
Taiw
anLe
eet
al.,
199
3c; G
unde
rsen
et a
l.,19
96
I-EB
lueb
erry
stun
tph
ytop
lasm
aB
BS
PS, r
RFL
P, c
RFL
PB
lueb
erry
stun
tVa
ccin
ium
spp
. (b
lueb
erry
)U
SALe
eet
al.,
199
3c; G
unde
rsen
et a
l.,19
96I-
FA
YA
(AC
LR)
FS, r
RFL
PU
nkno
wn
Prun
us a
rmen
iaca
(apr
icot
)Sp
ain
Schn
eide
ret a
l., 1
993;
See
m�l
lere
tal
., 19
94I-H
Chr
ysan
the-
mum
yel
low
sph
ytop
lasm
a
CY
brR
FLP,
cR
FLP
Chr
ysan
them
um y
ello
ws
Chr
ysan
them
um fr
utes
cens
(mar
guer
ite)
Italy
Gun
ders
enet
al.,
199
6
Subg
roup
1St
rain
/phy
topl
asm
aac
rony
m(s
)Ev
iden
ce o
f ide
ntity
or si
mila
rity2
Dis
ease
Plan
t or i
nsec
t hos
tG
eogr
aphi
c ar
eaSe
lect
ed re
fere
nces
14 Phytoplasma classification Journal of Plant Pathology (1998), 80 (1), 3-26
Tab
le 1
(con
tinu
ed)
Subg
roup
1St
rain
/phy
topl
asm
aac
rony
m(s
)Ev
iden
ce o
f ide
ntity
or s
imila
rity2
Dis
ease
Plan
t or i
nsec
t hos
tG
eogr
aphi
c ar
eaSe
lect
ed re
fere
nces
I-IM
exic
anpe
riwin
kle
vire
scen
ceph
ytop
lasm
a
MPV
rRFL
P, c
RFL
PPe
riwin
kle
vire
scen
ceC
atha
rant
hus
rose
us (p
eriw
inkl
e)M
exic
oG
unde
rsen
et a
l., 1
996
I-IrR
FLP
Stra
wbe
rry
gree
n pe
tal,
stun
ting
Frag
aria
xan
anas
sa (s
traw
berr
y)Fl
orid
aJo
man
tiene
et a
l., 1
996;
Har
rison
etal
., 19
97I-I
FWB
rRFL
PPe
riwin
kle
witc
hes'-
broo
mC
atha
rant
hus
rose
us (p
eriw
inkl
e)Fl
orid
aH
arris
onet
al.,
199
6
Subg
roup
uncl
ear
CO
LrR
FLP,
cR
FLP
Late
ntC
uscu
ta o
dora
ta (d
odde
r)G
erm
any
Schn
eide
ret
al.,
199
3; S
chne
ider
and
Seem
�lle
r, 19
94b
CV
LrR
FLP,
cR
FLP
Periw
inkl
e vi
resc
ence
Cat
hara
nthu
s ro
seus
(per
iwin
kle)
Peru
Schn
eide
ret
al.,
199
3; S
chne
ider
and
Seem
�lle
r, 19
94b
CV
TrR
FLP,
cR
FLP
Periw
inkl
e vi
resc
ence
Cat
hara
nthu
s ro
seus
(per
iwin
kle)
Thai
land
Schn
eide
ret
al.,
199
3; S
chne
ider
and
Seem
�lle
r, 19
94b
MPY
DB
Periw
inkl
e ye
llow
sC
atha
rant
hus
rose
us (p
eriw
inkl
e)M
alay
sia
Khe
wet
al.,
199
1SA
SrR
FLP,
cR
FLP
Sand
al sp
ike
Sant
alum
alb
um (s
anda
lwoo
d)In
dia
Schn
eide
ret
al.,
199
3; S
chne
ider
and
Seem
�lle
r, 19
94b
SAFP
SS, r
RFL
P,cR
FLP,
SSa
fflo
wer
phy
llody
Cha
rtha
mus
tinc
tori
us (s
afflo
wer
)Is
rael
Schn
eide
ret
al.,
199
3; S
chne
ider
and
Seem
�lle
r, 19
94b
DB
Pum
pkin
yel
low
sC
ucur
bita
pep
o (p
umpk
in)
Italy
Min
ucci
et a
l., 1
995
rRFL
PM
onar
da y
ello
ws
Mon
arda
fist
ulos
a (w
ild b
erga
mot
)C
anad
aK
hadh
air
et a
l., 1
997
SLo
tus y
ello
ws
Lotu
s co
rnic
ulat
us (b
ird's-
foot
trefo
il)U
SAG
riffit
hset
al.,
199
4a
SN
ew E
ngla
nd a
ster
yello
ws
Aste
r no
vae-
angl
iae
(New
Eng
land
aste
r)U
SAG
riffit
hset
al.,
199
4a
rRFL
PPo
plar
yel
low
sPo
pulu
s sp
p. (p
opla
r)Eu
rope
Ber
ges
et a
l., 1
997
1A
ll su
bgro
up m
embe
rs a
ppea
r ide
ntic
al o
r ver
y si
mila
r. D
iffer
ing
phyt
opla
smas
with
in th
e su
bgro
ups
are
mar
ked
as fo
llow
s: (
rr-r
p), d
istin
guis
habl
e on
the
basi
s of
com
bine
d R
FLP
anal
yses
of 1
6S rR
NA
and
ribo
som
al p
rote
in g
enes
, (
cRFL
P), d
istin
guis
habl
e on
the
basi
s of
cR
FLP
patte
rns
(see
und
er2 ),
S-a
to S
-d, s
erot
ypes
acc
ordi
ng to
Kea
neet
al.,
199
6. ?
, phy
topl
asm
as te
ntat
ivel
y as
sign
ed to
the
subg
roup
.2N
atur
e of
evi
denc
e: F
S, fu
ll-le
ngth
16S
rDN
A s
eque
nce;
PS,
par
tial 1
6S rD
NA
seq
uenc
e; S
S, 1
6S/2
3S rD
NA
spa
cer s
eque
nce;
rRFL
P, R
FLP
pat
tern
s of
PC
R-a
mpl
ified
ribo
som
al D
NA
; cR
FLP,
RFL
P pa
ttern
s of
non
ribos
omal
DN
A (P
CR
-am
plifi
ed o
r Sou
ther
n an
alys
is);
DB
, dot
blo
t a
naly
sis.
Journal of Plant Pathology (1998), 80 (1), 3-26 Seemüller et al. 15
Tab
le 2
.Cla
ssifi
catio
n of
non
-ast
er y
ello
ws
phyt
opla
smas
iden
tifie
d an
d di
ffer
entia
ted
on th
e ba
sis
of m
olec
ular
dat
a an
d di
vide
d in
to m
ajor
phy
loge
netic
gro
ups.
(con
tinu
ed)
Phy
topl
asm
a gr
oup
and
grou
p m
embe
rs a
ssoc
iate
dw
ith
the
dise
ases
list
ed 1
Acr
onym
(s)
of p
hyto
-pl
asm
a
Evi
denc
e of
iden
tity
or
sim
ilar
ity2
Pla
nt o
r in
sect
hos
tG
eogr
aphi
c ar
eaS
elec
ted
refe
renc
es
AU
ST
RA
LIA
N G
RA
PE
VIN
E Y
EL
LO
WS
(A
US
GY
) G
RO
UP
Aus
tral
ian
grap
evin
e ye
llow
s(C
and.
Phy
topl
. aus
tral
iens
e)A
US
GY
FS
, rR
FL
P,
cRF
LP
Vit
is v
inif
era
(gra
pevi
ne)
Aus
tral
iaP
adov
anet
al.
, 199
6; D
avis
et a
l., 1
997a
= P
apay
a di
ebac
krR
FL
PC
aric
a p
apay
a (p
apay
a)A
ustr
alia
Gib
bet
al.
, 199
6P
horm
ium
yel
low
leaf
PYL
FS
, cR
FL
PP
horm
ium
ten
ax (
New
Zea
land
fla
x)N
ew Z
eala
ndL
ieft
ing
et a
l., 1
996
ITA
LIA
N B
IND
WE
ED
ST
OL
BU
R (
IBS
) G
RO
UP
Ital
ian
bind
wee
d st
olbu
rIB
SF
S, r
RF
LP
Con
volv
ulus
arv
ensi
s (f
ield
bin
dwee
d)It
aly
Mar
cone
et a
l., 1
997a
ST
OL
BU
R (
ST
OL
) G
RO
UP
Sto
lbur
of
pepp
erS
TO
LF
S, S
S, R
FL
P,
cRF
LP
, SC
apsi
cum
ann
uum
(pe
pper
)F
ranc
e, S
erbi
aF
oset
al.
, 199
2; S
eem
�lle
ret
al.
, 199
4;M
arco
neet
al.
, 199
7a=
Tom
ato
stol
bur
ST
OL
SS
, rR
FL
P,
cRF
LP
, SL
ycop
ersi
con
esc
ulen
tum
(tom
ato)
Eur
ope
Fos
et a
l., 1
992;
Bou
don-
Pad
ieu
et a
l.,
1996
; Mar
cone
et a
l., 1
997b
= T
obac
co s
tolb
urS
TO
LrR
FL
P, S
Nic
otia
na t
abac
um (
toba
cco)
Fra
nce,
Ita
lyF
oset
al.
, 199
2; C
. Mar
cone
, unp
ubli
shed
= E
ggpl
ant s
tolb
urS
Sola
num
mel
onge
na (
eggp
lant
)F
ranc
eF
oset
al.
, 199
2=
Str
awbe
rry
yell
ows
SF
raga
ria
xan
anas
sa (
stra
wbe
rry)
Fra
nce
Fos
et a
l., 1
992
= P
orce
lain
dis
ease
SA
pium
gra
veol
ens
(cel
ery)
Fra
nce
Fos
et a
l., 1
992
= P
eriw
inkl
e ye
llow
sS
Cat
hara
nthu
s ro
seus
(pe
riw
inkl
e)F
ranc
eF
oset
al.
, 199
2=
Nig
htsh
ade
prol
ifer
atio
nS
Sola
num
dul
cam
ara
(bit
ters
wee
tni
ghts
hade
)F
ranc
eF
oset
al.
, 199
2
= L
ycop
ersi
con
hirs
utum
li
ttle
leaf
SL
ycop
ersi
con
hir
sutu
mF
ranc
eF
oset
al.
, 199
2
= L
eafh
oppe
r-bo
rne
SH
yale
sthe
s ob
sole
tus
(pla
ntho
pper
)F
ranc
eF
oset
al.
, 199
2=
Bin
dwee
d ye
llow
srR
FL
P, S
Con
volv
ulus
arv
ensi
s (f
ield
bin
dwee
d)F
ranc
e, G
erm
any
Fos
et a
l., 1
992;
Mai
xner
et a
l. 1
995;
Sch
neid
eret
al.
, 199
7b=
Boi
s no
ir, V
ergi
lbun
gs-
kr
ankh
eit,
othe
r gr
apev
ine
ye
llow
s
BN
, VK
FS
, SS
, rR
FL
P,
cRF
LP
Vit
is v
inif
era
(gra
pevi
ne)
Eur
ope
See
m�l
ler
et a
l., 1
994;
Mar
cone
et a
l.,
1996
a; D
aire
et a
l., 1
997
BU
CK
TH
OR
N W
ITC
HE
S'-
BR
OO
M (
BW
B)
GR
OU
P
Buc
ktho
rn w
itch
es'-b
room
BW
B,
BA
BW
FS
, rR
FL
PR
ham
nus
cath
arti
cus
(buc
ktho
rn)
Ger
man
yS
eem
�lle
ret
al.
, 199
4; M
�ure
r an
dS
eem
�lle
r, 1
996
SP
AR
TIU
M W
ITC
HE
SÕ-
BR
OO
M (
Spa
WB
) G
RO
UP
Spa
rtiu
m w
itch
esÕ-
broo
mS
paW
BF
S, r
RF
LP
Spar
tium
jun
ceum
(S
pani
sh b
room
)It
aly
Mar
cone
et a
l., 1
996b
Sar
otha
mnu
s w
itch
esÕ-
broo
mS
SW
BrR
FL
PSa
roth
amnu
s sc
opar
ius
(bro
om)
Ital
yM
arco
neet
al.
, 199
7f
16 Phytoplasma classification Journal of Plant Pathology (1998), 80 (1), 3-26
Tab
le 2
(con
tinu
ed)
Phy
topl
asm
a gr
oup
and
grou
p m
embe
rs a
ssoc
iate
dw
ith
the
dise
ases
list
ed 1
Acr
onym
(s)
of p
hyto
-pl
asm
a
Evi
denc
e of
iden
tity
or
sim
ilar
ity2
Pla
nt o
r in
sect
hos
tG
eogr
aphi
c ar
eaS
elec
ted
refe
renc
es
AP
PL
E P
RO
LIF
ER
AT
ION
(A
P)
GR
OU
P
App
le p
roli
fera
tion
AP
FS
, SS
, rR
FL
P,
cRF
LP
Mal
us d
omes
tica
(app
le)
Eur
ope
Kir
kpat
rick
et a
l., 1
994;
See
m�l
ler
et a
l.,
1994
; Lor
enz
et a
l., 1
995
= H
azel
dec
line
rRF
LP
Cor
ylus
ave
llan
a (h
azel
)It
aly
Mar
cone
et a
l., 1
996c
= B
indw
eed
yell
ows
rRF
LP
Con
volv
ulus
arv
ensi
s (f
ield
bin
dwee
d)G
erm
any
Sch
neid
eret
al.
, 199
7P
ear
decl
ine
S, S
S, r
RF
LP
Pyr
us c
omm
unis
(pe
ar)
Eur
ope,
Nor
thA
mer
ica
Kir
kpat
rick
et a
l., 1
994;
See
m�l
ler
et a
l.,
1994
; Kis
onet
al.
, 199
7=
Pea
ch y
ello
w le
af r
oll
PY
LR
FS
, rR
FL
PP
runu
s pe
rsic
a (p
each
)N
orth
Am
eric
aK
irkp
atri
cket
al.
, 199
4; K
ison
et a
l.,
1997
= H
azel
dec
line
rRF
LP
Cor
ylus
ave
llan
a (h
azel
)It
aly
Mar
cone
et a
l., 1
996c
Eur
opea
n st
one
frui
t yel
low
sof
pea
chE
SF
YF
S, S
S, r
RF
LP
,cR
FL
PP
runu
s pe
rsic
a (p
each
)E
urop
eK
irkp
atri
cket
al.
, 199
4; S
eem
�lle
ret
al.
,19
94; K
ison
et a
l., 1
997
= E
urop
ean
ston
e fr
uit
yel
low
s of
apr
icot
(A
pric
ot c
hlor
otic
leaf
rol
l)
ES
FY
(AC
LR
)S
S, r
RF
LP
,cR
FL
PP
runu
s ar
men
iaca
(apr
icot
)E
urop
eK
irkp
atri
cket
al.
, 199
4; L
oren
zet
al.
,19
94; K
ison
et a
l., 1
997
= E
urop
ean
ston
e fr
uit
ye
llow
s of
Jap
. plu
m
(Plu
m le
pton
ecro
sis)
ES
FY
(PL
N)
rRF
LP
, cR
FL
PP
runu
s sa
licin
a (
Japa
nese
plu
m)
Eur
ope
Lor
enz
et a
l. 1
994;
Mar
cone
et a
l., 1
996e
= E
urop
ean
ston
e fr
uit
ye
ll. o
f fl
ower
ing
cher
ryE
SF
YrR
FL
P, c
RF
LP
P.
serr
ulat
a (
flow
erin
g ch
erry
)G
erm
any
Sch
neid
eret
al.
, 199
3; L
oren
zet
al.
, 199
4
= E
urop
ean
ston
e fr
uit
ye
llow
s of
Eur
op. p
lum
ES
FY
rRF
LP
,cR
FL
PP
runu
s do
mes
tica
(E
urop
ean
plum
)E
urop
eL
oren
zet
al.
, 199
4
= E
urop
ean
ston
e fr
uit
ye
llow
s of
alm
ond
ES
FY
rRF
LP
,cR
FL
PP
.am
ygda
lifo
rmis
(al
mon
d)E
urop
eL
oren
zet
al.
, 199
4
= H
azel
dec
line
rRF
LP
Cor
ylus
ave
llan
a (h
azel
)It
aly
Mar
cone
et a
l., 1
996c
X-D
ISE
AS
E (
WX
) G
RO
UP
Wes
tern
X-d
isea
seW
XF
S, S
S, r
RF
LP
,cR
FL
P, S
Pru
nus
pers
ica
(pea
ch)
Wes
tern
US
AS
eem
�lle
ret
al.
, 199
4; S
chne
ider
et a
l.,19
95b;
Gun
ders
enet
al.
, 199
6=
Dis
ease
unk
now
nP
DX
SS
, rR
FL
P,
cRF
LP
Pyr
us c
omm
unis
(pe
ar)
Cal
ifor
nia
Gun
ders
enet
al.
, 199
6
= E
rige
ron
yell
ows
ErY
4rR
FL
PE
rige
ron
cana
dens
is (h
orse
wee
d)U
SA
Lee
et a
l., 1
994
= G
olde
nrod
stu
ntG
S1rR
FL
PSo
lida
go r
ugos
a (
gold
enro
d)U
SA
Lee
et a
l., 1
994
= F
ree-
bran
chin
g of
C
hris
tmas
poi
nset
tia
rRF
LP
Eup
horb
ia p
ulch
erri
ma
(Chr
istm
aspo
inse
ttia
)U
SA
Aba
det
al.
, 199
7
Can
adia
n pe
ach
X-
di
seas
eC
XF
S, r
RF
LP
,cR
FL
P, S
Pru
nus
pers
ica
(pea
ch)
Can
ada
Lee
et a
l., 1
992b
, 199
3c; G
unde
rsen
etal
., 19
96=
Cho
kech
erry
X-d
isea
seC
CX
PS
, rR
FL
P,
cRF
LP
, SP
runu
s vi
rgin
iana
(ch
oke
cher
ry)
US
AG
unde
rsen
et a
l., 1
996;
Guo
et a
l., 1
996
Clo
ver
yell
ow e
dge
CY
EF
S, r
RF
LP
,cR
FL
P, S
Tri
foliu
m r
epen
s (w
hite
clo
ver)
Can
ada
Shi
na a
nd B
enha
mou
, 198
3; L
eeet
al.
,19
92b;
Gun
ders
enet
al.
, 199
6

Journal of Plant Pathology (1998), 80 (1), 3-26 Seemüller et al. 17
(con
tinu
ed)
= G
enti
an w
itch
es'-b
room
GW
rRF
LP
Gen
tian
a sp
. (ge
ntia
n)Ja
pan
Oku
daet
al.
, 199
7=
Gra
pevi
ne y
ello
ws
FD
VA
,F
DU
rRF
LP
, cR
FL
P,
SV
itis
vin
ifer
a (g
rape
vine
)U
SA
, Ita
lyP
rinc
eet
al.
, 199
3; C
hen
et a
l., 1
993;
Mar
cone
et a
l., 1
996a
= O
xeye
phy
llod
yO
xPrR
FL
P, c
RF
LP
Chr
ysan
them
um l
euca
nthe
mum
(oxe
ye d
aisy
)It
aly
Fir
rao
et a
l., 1
996a
= T
arax
acum
yel
low
s=
TaY
rRF
LP
, cR
FL
PT
arax
acum
off
icin
ale
(da
ndel
ion)
Ital
yF
irra
oet
al.
, 199
6a=
Cre
pis
yell
ows
= C
reY
rRF
LP
, cR
FL
PC
repi
s bi
enni
s (r
ough
haw
k's
bear
d)It
aly
Fir
rao
et a
l., 1
996a
= F
ree-
bran
chin
g of
C
hris
tmas
poi
nset
tia
rRF
LP
Eup
horb
ia p
ulch
erri
ma
(Chr
istm
aspo
inse
ttia
)U
SA
Aba
det
al.
, 199
7
Mil
kwee
d ye
llow
sM
WY
rRF
LP
, cR
FL
PA
scle
pias
syr
iaca
(m
ilkw
eed)
US
AG
riff
iths
et a
l., 1
994c
; G
unde
rsen
et a
l.,
1996
Ital
ian
clov
er p
hyll
ody
ICP
hF
S, r
RF
LP
,cR
FL
P, S
Tri
foli
um s
p. (
clov
er)
Ital
yF
irra
oet
al.
, 199
6a, 1
996b
Pec
an b
unch
PB
rRF
LP
, cR
FL
PC
arya
ill
inoe
nsis
(pe
can)
US
AG
unde
rsen
et a
l., 1
996
Gol
denr
od y
ello
ws
GR
1rR
FL
P, c
RF
LP
Soli
dago
rug
osa
(go
lden
rod)
US
AG
riff
iths
et a
l., 1
994c
; G
unde
rsen
et a
l.,
1996
Wal
nut w
itch
es'-b
room
WW
BS
S, r
RF
LP
,cR
FL
P, S
Jugl
ans
nigr
a (
blac
k w
alnu
t)U
SA
Kir
kpat
rick
et a
l., 1
994;
Gun
ders
enet
al.
,19
96; F
irra
oet
al.
, 199
6b S
pire
a st
unt
SP
1rR
FL
P, c
RF
LP
Spir
aea
sp. (
spir
ea)
US
AG
riff
iths
et a
l., 1
994c
; G
unde
rsen
et a
l.,
1996
Fre
e-br
anch
ing
ofC
hris
tmas
poi
nset
tia
rRF
LP
Eup
horb
ia p
ulch
erri
ma
(Chr
istm
aspo
inse
ttia
)U
SA
Lee
et a
l., 1
997a
Vac
cini
um w
itch
es'-b
room
VA
CF
S, S
S, r
RF
LP
,S
Vac
cini
um m
yrti
llus
(bl
uebe
rry)
Ger
man
yS
chne
ider
et a
l., 1
993;
Kir
kpat
rick
et a
l.,
1994
; See
m�l
ler
et a
l., 1
994
Tsu
wab
uki w
itch
es'-b
room
TW
BF
S, r
RF
LP
Far
fugi
um j
apon
icum
(ts
uwab
uki)
Japa
nN
amba
et a
l., 1
993;
Kir
kpat
rick
et a
l.,
1994
; Oku
daet
al.
, 199
7?
Gre
en V
alle
y X
-dis
ease
GV
XrR
FL
P, c
RF
LP
Pru
nus
aviu
m (
cher
ry)
Cal
ifor
nia
Ahr
ens
et a
l., 1
993
? P
each
yel
low
sPY
cRF
LP
Pru
nus
pers
ica
(pea
ch)
US
AK
irkp
atri
ck, 1
991
? P
each
ros
ette
PR
SS
, cR
FL
P, S
Pru
nus
pers
ica
(pea
ch)
US
AK
irkp
atri
ck, 1
991;
Fir
rao
et a
l., 1
996b
? B
rasi
lian
tom
ato
big
bud
BT
BB
rRF
LP
, SL
ycop
ersi
con
esc
ulen
tum
(to
mat
o)B
rasi
lB
oudo
n-P
adie
uet
al.
, 199
6?
Dod
onea
yel
low
sD
oYS
S, D
BD
odon
aea
vis
cosa
Haw
aii
Bor
thet
al.
, 199
5?
Hor
sew
eed
phyl
lody
rRF
LP
Eri
gero
n ca
nade
nsis
(ho
rsew
eed)
US
AS
chne
ider
et a
l., 1
997b
ITA
LIA
N A
LF
AL
FA
WIT
CH
ES
'-B
RO
OM
(IA
WB
) G
RO
UP
Ital
ian
alfa
lfa
wit
ches
'-bro
omIA
WB
FS
, rR
FL
PM
edic
ago
sat
iva
(al
falf
a)It
aly
Mar
cone
et a
l., 1
997b
Pic
ris
echi
oide
s ph
yllo
dyP
EP
FS
, rR
FL
PP
icri
s ec
hioi
des
(bri
stly
oxt
ongu
e)It
aly
Mar
cone
et a
l., 1
997a
FA
BA
BE
AN
PH
YL
LO
DY
(F
BP
) G
RO
UP
Fab
a be
an p
hyll
ody
FB
PF
S, S
S, r
RF
LP
Vic
ia f
aba
(fa
ba b
ean)
Sud
anK
irkp
atri
cket
al.
, 199
4; S
chne
ider
et a
l.,
1995
a=
Cro
tala
ria
phyl
lody
CrP
(C
JP)
rRF
LP
, SC
rota
lari
a j
unce
a (
sunn
hem
p)T
hail
and
Sae
edet
al.
, 199
4; S
chne
ider
et a
l., 1
995a
= C
rota
lari
a sa
ltia
na
phyl
lody
.C
SP
rRF
LP
, SC
rota
lari
a s
alti
ana
Sud
anS
aeed
et a
l., 1
994;
Sch
neid
eret
al.
, 199
5a
= S
oybe
an p
hyll
ody
Soy
PrR
FL
P, S
Gly
cine
max
(so
ybea
n)T
hail
and
Sae
edet
al.
, 199
4; S
chne
ider
et a
l., 1
995a
Cot
ton
phyl
lody
CoP
rRF
LP
Gos
sypi
um h
irsu
tum
(co
tton
)B
urki
na F
aso
Sch
neid
eret
al.,
199
7b; C
. Mar
cone
,un
publ
ishe
d re
sult

18 Phytoplasma classification Journal of Plant Pathology (1998), 80 (1), 3-26
Tab
le 2
(con
tinu
ed)
Phyt
opla
sma
grou
p an
dgr
oup
mem
bers
ass
ocia
ted
with
the
dise
ases
list
ed 1
Acr
onym
(s)
of p
hyto
-pl
asm
a
Evi
denc
e of
iden
tity
orsi
mila
rity
2
Plan
t or
inse
ct h
ost
Geo
grap
hic
area
Sele
cted
refe
renc
es
Sunn
hem
p w
itche
s'-br
oom
SUN
HP
FS, r
RFL
P, S
Cro
tala
ria
junc
ea (s
unn
hem
p)T
haila
ndSa
eed
et a
l., 1
994;
Sch
neid
eret
al.
, 199
5a=
Cro
tala
ria
spp.
witc
hes'-
br
oom
rRFL
PC
rota
lari
a sp
p.A
ustr
alia
Dav
iset
al.
, 199
7b
= Se
sam
e ph
yllo
dySe
PrR
FLP,
SSe
sam
um i
ndic
um (
sesa
me)
Tha
iland
Saee
det
al.
, 199
4; S
chne
ider
et a
l.,
1995
a; D
avis
et a
l., 1
997b
= C
leom
e ph
yllo
dyC
LP
(C
mP)
rRFL
P, S
Cle
ome
visc
osa
Tha
iland
Schn
eide
ret
al.
, 199
5a=
Aus
tral
ian
pear
dec
line
rRFL
P, D
BP
yrus
com
mun
is (p
ear)
Aus
tral
iaSc
hnei
der
and
Gib
b, 1
997
= A
ustr
alia
n to
mat
o bi
g bu
dT
BB
FS, r
RFL
P, S
Lyco
pers
icon
esc
ulen
tum
(po
tato
)A
ustr
alia
Schn
eide
ret
al.
, 199
5a; M
inuc
ciet
al.
,19
96; D
avis
et a
l., 1
997b
= Ju
te p
hyllo
dyJu
PrR
FLP,
SC
orch
orus
sp.
(ju
te)
Min
ucci
et a
l., 1
996
= Pa
paya
yel
low
cri
nkle
PYC
rRFL
PC
aric
a p
apay
a (p
apay
a)A
ustr
alia
Dav
iset
al.
, 199
7b=
Papa
ya m
osai
crR
FLP
Car
ica
pap
aya
(pap
aya)
Aus
tral
iaD
avis
et a
l., 1
997b
= E
ggpl
ant l
ittle
leaf
EL
LrR
FLP,
cR
FLP
Sola
num
mel
onge
na (e
ggpl
ant)
Aus
tral
iaD
avis
et a
l., 1
997b
= A
ngel
s' tr
umpe
t litt
le le
afrR
FLP
Bru
gman
sia
can
dida
(an
gels
' tru
mpe
t)A
ustr
alia
Dav
iset
al.
, 199
7b=
Wild
goo
sebe
rry
little
leaf
rRFL
PP
hysa
lis m
inim
a (w
ild g
oose
berr
y)A
ustr
alia
Dav
iset
al.
, 199
7b=
Zuc
chin
i litt
le le
afrR
FLP
Cuc
urbi
ta p
epo
(zuc
chin
i)A
ustr
alia
Dav
iset
al.
, 199
7b=
Pean
ut w
itche
s'-br
oom
rRFL
PA
rach
is h
ypog
ea (p
eanu
t)A
ustr
alia
Dav
iset
al.
, 199
7b=
Aus
tral
ian
clov
er p
hyllo
dyrR
FLP
Trifo
lium
rep
ens
(whi
te c
love
r)A
ustr
alia
Dav
iset
al.
, 199
7b=
Vig
na w
itche
s'-br
oom
rRFL
PV
igna
spp
. (co
wpe
a an
d ot
hers
)A
ustr
alia
Dav
iset
al.
, 199
7b=
Ger
bera
phy
llody
rRFL
PG
erbe
ra s
p. (
gerb
era)
Aus
tral
iaD
avis
et a
l., 1
997b
= N
iga
little
leaf
rRFL
PG
uizo
tia a
byss
inic
a (n
iga)
Aus
tral
iaD
avis
et a
l., 1
997b
= L
ettu
ce p
hyllo
dyrR
FLP
Lact
uca
sativ
a (l
ettu
ce)
Aus
tral
iaD
avis
et a
l., 1
997b
= E
upho
rbia
littl
e le
afrR
FLP
Eup
horb
ia m
illii
Aus
tral
iaD
avis
et a
l., 1
997b
= R
ough
cha
inw
eed
w
itche
s'-br
oom
rRFL
PA
lysi
carp
us r
ugos
us (
roug
hch
ainw
eed)
Aus
tral
iaD
avis
et a
l., 1
997b
= M
acro
ptili
um li
ttle
leaf
an
d w
itche
s'-br
oom
rRFL
PM
acro
ptili
um sp
p.A
ustr
alia
Dav
iset
al.
, 199
7b
= V
elve
t bea
n w
itche
s'-
broo
mrR
FLP
Muc
una
pru
rien
s (v
elve
t bea
n)A
ustr
alia
Dav
iset
al.
, 199
7b
= Fl
anne
l wee
d lit
tle le
afrR
FLP
Sida
cor
difo
lia (f
lann
el w
eed)
Aus
tral
iaD
avis
et a
l., 1
997b
= Pe
renn
ial p
hlox
phy
llody
rRFL
PP
hlox
sp.
(pe
renn
ial p
hlox
)A
ustr
alia
Dav
iset
al.
, 199
7b=
Mar
sh g
rape
frui
t die
back
rRFL
PC
itrus
par
adis
i (m
arsh
gra
pefr
uit)
Aus
tral
iaD
avis
et a
l., 1
997b
= B
ellv
ine
little
leaf
rRFL
PIp
omoe
a p
lebe
ia (
Bel
lvin
e)A
ustr
alia
Dav
iset
al.
, 199
7b=
Swee
t pot
ato
little
leaf
SPL
SIp
omoe
a b
atat
as (s
wee
t pot
ato)
Aus
tral
iaM
inuc
ciet
al.
, 199
6Sw
eet p
otat
o lit
tle le
afSP
LL
rRFL
P, c
RFL
PIp
omoe
a b
atat
as (s
wee
t pot
ato)
Aus
tral
iaD
avis
et a
l., 1
997b
Lim
e w
itche
s'-br
oom
(Can
d. P
hyt.
aura
ntif
olia
)W
BD
LFS
, cR
FLP
Citr
us a
uran
tifol
ia (l
ime)
Om
anZ
reik
et a
l., 1
995
Swee
t pot
ato
witc
hes
broo
mSP
WB
FS, r
RFL
PIp
omoe
a b
atat
as (s
wee
t pot
ato)
Tai
wan
Lee
et a
l., 1
993c
; Gun
ders
enet
al.
, 199
4Pe
anut
witc
hes'-
broo
mPn
WB
FS, c
RFL
PA
rach
is h
ypog
aea
(pea
nut)
Tai
wan
Che
n an
d L
in, 1
997
= R
ed b
ird
cact
us w
itche
s'-
broo
mR
BC
WB
rRFL
PP
edila
nthu
s tit
hym
aloi
des
(red
bir
dca
ctus
)T
aiw
anL
eeet
al.
, 199
3c
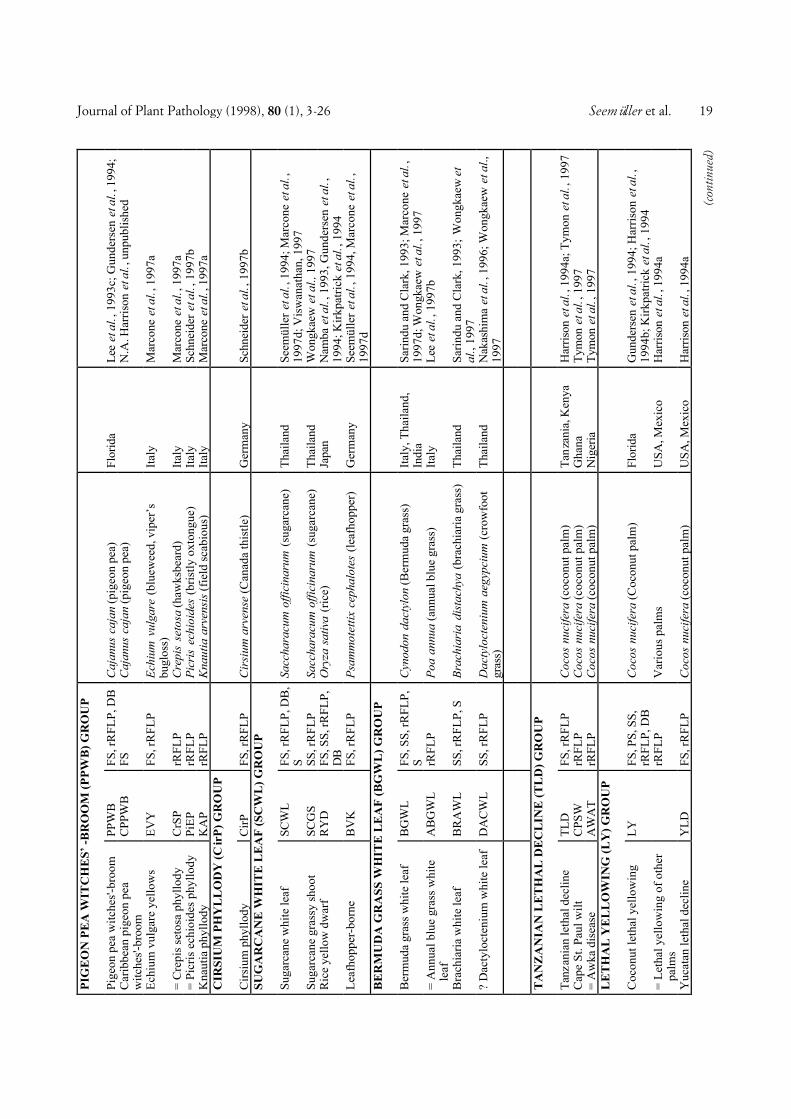
Journal of Plant Pathology (1998), 80 (1), 3-26 Seemüller et al. 19
(con
tinu
ed)
PIG
EO
N P
EA
WIT
CH
ES
Õ -B
RO
OM
(P
PW
B)
GR
OU
P
Pig
eon
pea
wit
ches
'-bro
omP
PW
BF
S, r
RF
LP
, DB
Caj
anus
caj
an (
pige
on p
ea)
Flor
ida
Lee
et a
l., 1
993c
; Gun
ders
enet
al.
, 199
4;C
arib
bean
pig
eon
pea
witc
hes'
-bro
omC
PPW
BF
SC
ajan
us c
ajan
(pi
geon
pea
)N
.A. H
arri
son
et a
l., u
npub
lish
ed
Ech
ium
vul
gare
yel
low
sE
VY
FS
, rR
FL
PE
chiu
m v
ulga
re (
blue
wee
d, v
iper
Õsbu
glos
s)It
aly
Mar
cone
et a
l., 1
997a
= C
repi
s se
tosa
phy
llod
yC
rSP
rRF
LP
Cre
pis
seto
sa (h
awks
bear
d)It
aly
Mar
cone
et a
l., 1
997a
= P
icri
s ec
hioi
des
phyl
lody
PiE
PrR
FL
PP
icri
s ec
hioi
des
(bri
stly
oxt
ongu
e)It
aly
Schn
eide
ret
al.
, 199
7bK
naut
ia p
hyll
ody
KA
PrR
FL
PK
naut
ia a
rven
sis
(fie
ld s
cabi
ous)
Ital
yM
arco
neet
al.
, 199
7aC
IRS
IUM
PH
YL
LO
DY
(C
irP
) G
RO
UP
Cir
sium
phy
llod
yC
irP
FS
, rR
FL
PC
irsi
um a
rven
se (
Can
ada
this
tle)
Ger
man
ySc
hnei
der
et a
l., 1
997b
SU
GA
RC
AN
E W
HIT
E L
EA
F (
SC
WL
) G
RO
UP
Sug
arca
ne w
hite
leaf
SCW
LF
S, r
RF
LP
, DB
,S
Sacc
hara
cum
offi
cina
rum
(su
garc
ane)
Tha
iland
See
m�l
ler
et a
l., 1
994;
Mar
cone
et a
l.,
1997
d; V
isw
anat
han,
199
7S
ugar
cane
gra
ssy
shoo
tS
CG
SS
S, r
RF
LP
Sacc
hara
cum
offi
cina
rum
(su
garc
ane)
Tha
iland
Won
gkae
wet
al.,
199
7R
ice
yell
ow d
war
fR
YD
FS
, SS
, rR
FL
P,
DB
Ory
za s
ativ
a (
rice
)Ja
pan
Nam
baet
al.
, 199
3, G
unde
rsen
et a
l.,
1994
; Kir
kpat
rick
et a
l., 1
994
Lea
fhop
per-
born
eB
VK
FS
, rR
FL
PP
sam
mot
etti
x ce
phal
otes
(lea
fhop
per)
Ger
man
yS
eem
�lle
ret
al.
, 199
4, M
arco
neet
al.
,19
97d
BE
RM
UD
A G
RA
SS
WH
ITE
LE
AF
(B
GW
L)
GR
OU
P
Ber
mud
a gr
ass
whi
te le
afB
GW
LF
S, S
S, r
RF
LP
,S
Cyn
odon
dac
tylo
n (B
erm
uda
gras
s)It
aly,
Tha
ilan
d,In
dia
Sar
indu
and
Cla
rk, 1
993;
Mar
cone
et a
l.,
1997
d; W
ongk
aew
et a
l., 1
997
= A
nnua
l blu
e gr
ass
whi
te
leaf
AB
GW
LrR
FL
PP
oa a
nnua
(an
nual
blu
e gr
ass)
Ital
yL
eeet
al.
, 199
7b
Bra
chia
ria
whi
te le
afB
RA
WL
SS
, rR
FL
P, S
Bra
chia
ria
dis
tach
ya(b
rach
iari
a gr
ass)
Tha
iland
Sar
indu
and
Cla
rk, 1
993;
Won
gkae
w e
tal
., 19
97?
Dac
tylo
cten
ium
whi
te le
afD
AC
WL
SS
, rR
FL
PD
acty
loct
eniu
m a
egyp
cium
(cro
wfo
otgr
ass)
Tha
iland
Nak
ashi
ma
et a
l., 1
996;
Won
gkae
wet
al.,
1997
TA
NZ
AN
IAN
LE
TH
AL
DE
CL
INE
(T
LD
) G
RO
UP
Tan
zani
an le
thal
dec
line
TL
DF
S, r
RF
LP
Coc
os n
ucif
era
(coc
onut
pal
m)
Tan
zani
a, K
enya
Har
riso
net
al.
, 199
4a; T
ymon
et a
l., 1
997
Cap
e S
t. P
aul w
ilt
CP
SW
rRF
LP
Coc
os n
ucif
era
(coc
onut
pal
m)
Gha
naT
ymon
et a
l., 1
997
= A
wka
dis
ease
AW
AT
rRF
LP
Coc
os n
ucif
era
(coc
onut
pal
m)
Nig
eria
Tym
onet
al.
, 199
7L
ET
HA
L Y
EL
LO
WIN
G (
LY
) G
RO
UP
Coc
onut
leth
al y
ello
win
gL
YF
S, P
S, S
S,
rRF
LP
, DB
Coc
os n
ucif
era
(Coc
onut
pal
m)
Flor
ida
Gun
ders
enet
al.
, 199
4; H
arri
son
et a
l.,
1994
b; K
irkp
atri
cket
al.
, 199
4=
Let
hal y
ello
win
g of
oth
er
palm
srR
FL
PV
ario
us p
alm
sU
SA
, Mex
ico
Har
riso
net
al.
, 199
4a
Yuc
atan
leth
al d
ecli
neY
LD
FS
, rR
FL
PC
ocos
nuc
ifer
a (c
ocon
ut p
alm
)U
SA
, Mex
ico
Har
riso
net
al.
, 199
4a
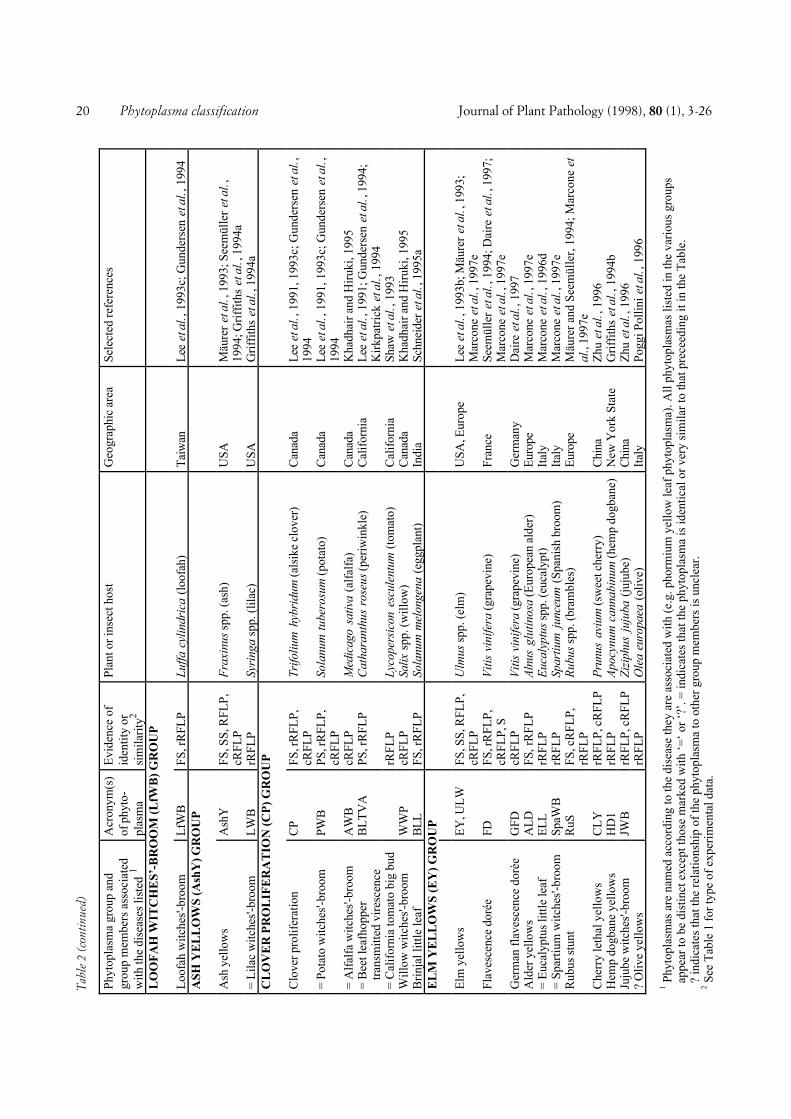
20 Phytoplasma classification Journal of Plant Pathology (1998), 80 (1), 3-26
Tab
le 2
(con
tinu
ed)
Phyt
opla
sma
grou
p an
dgr
oup
mem
bers
ass
ocia
ted
with
the
dise
ases
list
ed 1
Acr
onym
(s)
of p
hyto
-pl
asm
a
Evi
denc
e of
iden
tity
orsi
mila
rity
2
Plan
t or
inse
ct h
ost
Geo
grap
hic
area
Sele
cted
refe
renc
es
LO
OF
AH
WIT
CH
ESÕ
-BR
OO
M (L
fWB
) GR
OU
P
Loo
fah
witc
hes'-
broo
mL
fWB
FS, r
RFL
PLu
ffa c
ylin
dric
a (l
oofa
h)T
aiw
anL
eeet
al.
, 199
3c; G
unde
rsen
et a
l., 1
994
ASH
YE
LL
OW
S (A
shY
) GR
OU
P
Ash
yel
low
sA
shY
FS, S
S, R
FLP,
cRFL
PF
raxi
nus
spp.
(ash
)U
SAM
�ure
ret
al.
, 199
3; S
eem
�lle
ret
al.
,19
94; G
riff
iths
et a
l., 1
994a
= L
ilac
witc
hes'-
broo
mL
WB
rRFL
PSy
ring
a sp
p. (
lilac
)U
SAG
riff
iths
et a
l., 1
994a
CL
OV
ER
PR
OL
IFE
RA
TIO
N (C
P) G
RO
UP
Clo
ver p
rolif
erat
ion
CP
FS, r
RFL
P,cR
FLP
Trifo
lium
hyb
ridu
m (a
lsik
e cl
over
)C
anad
aL
eeet
al.
, 199
1, 1
993c
; Gun
ders
enet
al.
,19
94=
Pota
to w
itche
s'-br
oom
PWB
PS, r
RFL
P,cR
FLP
Sola
num
tub
eros
um (
pota
to)
Can
ada
Lee
et a
l., 1
991,
199
3c; G
unde
rsen
et a
l.,
1994
= A
lfal
fa w
itche
s'-br
oom
AW
BcR
FLP
Med
icag
o s
ativ
a (a
lfal
fa)
Can
ada
Kha
dhai
r and
Hir
uki,
1995
= B
eet l
eafh
oppe
r
tran
smitt
ed v
ires
cenc
eB
LT
VA
PS, r
RFL
PC
atha
rant
hus
rose
us (p
eriw
inkl
e)C
alif
orni
aL
eeet
al.
, 199
1; G
unde
rsen
et a
l., 1
994;
Kir
kpat
rick
et a
l., 1
994
= C
alif
orni
a to
mat
o bi
g bu
drR
FLP
Lyco
pers
icon
esc
ulen
tum
(tom
ato)
Cal
ifor
nia
Shaw
et a
l., 1
993
Will
ow w
itche
s'-br
oom
WW
PcR
FLP
Salix
spp
. (w
illow
)C
anad
aK
hadh
air a
nd H
iruk
i, 19
95B
rinj
al li
ttle
leaf
BL
LFS
, rR
FLP
Sola
num
mel
onge
na (e
ggpl
ant)
Indi
aSc
hnei
der
et a
l., 1
995a
EL
M Y
EL
LO
WS
(EY
) GR
OU
P
Elm
yel
low
sE
Y, U
LW
FS, S
S, R
FLP,
cRFL
PU
lmus
spp
. (el
m)
USA
, Eur
ope
Lee
et a
l., 1
993b
; M�u
rer
et a
l., 1
993;
Mar
cone
et a
l., 1
997e
Flav
esce
nce
dor�
eFD
FS, r
RFL
P,cR
FLP,
SV
itis
vini
fera
(gra
pevi
ne)
Fran
ceSe
em�l
ler
et a
l., 1
994;
Dai
reet
al.
, 199
7;M
arco
neet
al.
, 199
7eG
erm
an f
lave
scen
ce d
or�e
GFD
cRFL
PV
itis
vini
fera
(gra
pevi
ne)
Ger
man
yD
aire
et a
l., 1
997
Ald
er y
ello
ws
AL
DFS
,rR
FLP
Aln
us g
lutin
osa
(Eur
opea
n al
der)
Eur
ope
Mar
cone
et a
l., 1
997e
= E
ucal
yptu
s lit
tle le
afE
LL
rRFL
PE
ucal
yptu
s sp
p. (e
ucal
ypt)
Ital
yM
arco
neet
al.
, 199
6d=
Spar
tium
witc
hes'-
broo
mSp
aWB
rRFL
PSp
artiu
m ju
nceu
m (S
pani
sh b
room
)It
aly
Mar
cone
et a
l., 1
997e
Rub
us s
tunt
RuS
FS, c
RFL
P,rR
FLP
Rub
us s
pp. (
bram
bles
)E
urop
eM
�ure
r an
d Se
em�l
ler,
199
4; M
arco
neet
al.,
1997
eC
herr
y le
thal
yel
low
sC
LY
rRFL
P, c
RFL
PP
runu
s av
ium
(sw
eet c
herr
y)C
hina
Zhu
et a
l., 1
996
Hem
p do
gban
e ye
llow
sH
D1
rRFL
PA
pocy
num
can
nabi
num
(hem
p do
gban
e)N
ew Y
ork
Stat
eG
riff
iths
et a
l., 1
994b
Juju
be w
itche
s'-br
oom
JWB
rRFL
P, c
RFL
PZi
ziph
us ju
juba
(ju
jube
)C
hina
Zhu
et a
l., 1
996
? O
live
yello
ws
rRFL
PO
lea
euro
paea
(ol
ive)
Ital
yPo
ggi P
ollin
iet
al.
, 199
6
1 Phy
topl
asm
as a
re n
amed
acc
ordi
ng to
the
dise
ase
they
are
ass
ocia
ted
with
(e.
g. p
horm
ium
yel
low
leaf
phy
topl
asm
a). A
ll ph
ytop
lasm
as li
sted
in th
e va
riou
s gr
oups
app
ear
to b
e di
stin
ct e
xcep
t tho
se m
arke
d w
ith Ô
=Ô o
r Ô?
Õ. =
indi
cate
s th
at th
e ph
ytop
lasm
a is
iden
tical
or
very
sim
ilar
to th
at p
rece
edin
g it
in th
e T
able
. ?
indi
cate
s th
at th
e re
latio
nshi
p of
the
phyt
opla
sma
to o
ther
gro
up m
embe
rs is
unc
lear
.2 S
ee T
able
1 f
or ty
pe o
f ex
peri
men
tal d
ata.

REFERENCES
Abad J.A., Randall C., Moyer J.W., 1997. Genomic diversityand molecular characterization of poinsettia phytoplasmas.Phytopathology 87 : S1.
Ahrens U., Lorenz K.-H., Seemüller E., 1993. Genetic diversi-ty among mycoplasmalike organisms associated with stonefruit diseases. Molecular Plant-Microbe Interactions 6: 686-691.
Alma A., Davis R.E., Vibio M., Danielli A., Bosco D., ArzoneA., Bertaccini A., 1996. Mixed infection of grapevines innorthern Italy by phytoplasmas including 16S rRNA RFLPsubgroup 16SrI-B strains previously unreported in thishost. Plant Disease 80: 418-421.
Berges R., Cousin M.T., Roux J., Mäurer R., Seemüller E.,1997. Detection of phytoplasma infections in decliningPopulus nigra ‘Italica’ trees and molecular differentiation ofthe aster yellows phytoplasmas identified in various Populusspecies. European Journal of Forest Pathology 27: 33-43.
Bertaccini A., Bellardi M.G., Vibio M., 1996a. «Free-branch-ing» della poinsettia e presenza di fitoplasmi. Atti Conveg-no Annuale SIPaV, Udine 1996, C10-C11.
Bertaccini A., Mittempergher L., Vibio M., 1996b. Identifica-tion of phytoplasmas associated with a decline of Euro-pean hackberry (Celtis australis). Annals of Applied Biology128: 245-253.
Bertaccini A., Vibio M., 1996. Phytoplasma identification inItaly: Results of 16S rDNA analysis. The Fifth InternationalWorkshop on Phytoplasmas, Bejing 1996, 13-14.
Bertaccini A., Vibio M., Bellardi M.G., 1996c. Virus diseasesof ornamental shrubs: X. Euphorbia pulcherrima Willd. in-fected by viruses and phytoplasmas. PhytopathologiaMediterranea 35: 129-132.
Bonnet F., Saillard C., Kollar A., Seemüller E., Bové J.M.,1990. Detection and differentiation of the mycoplasmalikeorganism associated with apple proliferation disease usingcloned DNA probes. Molecular Plant-Microbe Interactions3: 438-443.
Borth W.B., Hu J.S., Kirkpatrick B.C., Gardner D.E., Ger-man T.L., 1995. Occurrence of phytoplasmas in Hawaii.Plant Disease 79: 1094-1097.
Boudon-Padieu E., Cousin M.T., Daire X., Roux J., 1996.Molecular and serological characterization of phytoplasmaisolates inducing tomato stolbur symptoms in tomato.IOM Letters 4: 167-168.
Caudwell A., Larrue J., Kuszala C., Bachelier J.-C., 1971. Plu-ralité des jaunisses de la vigne. Annales de Phytopathologie3: 95-105.
Chen K.H., Guo J.R., Wu X.Y., Loi N., Carraro L., GuoY.H., Chen Y.D., Osler R., Pearson R., Chen T.A., 1993.Comparison of monoclonal antibodies, DNA probes, andPCR for detection of the grapevine yellows disease agent.Phytopathology 83: 915-922.
Chen M.-F., Lin C.-P., 1997. DNA probes and PCR primersfor the detection of a phytoplasma associated with peanutwitches’ broom. European Journal of Plant Pathology 103:137-145.
Chiykowski L.N., Sinha R.C., 1990. Differentiation of MLOdiseases by means of symptomatology and vector transmis-sion. Zentralblatt für Bakteriologie, Suppl. 20: 280-287.
Clark M.F., Barbara D.J., Davies D.L., 1983. Production andcharacteristics of antisera to Spiroplasma citri and cloverphyllody-associated antigens derived from plants. Annalsof Applied Biology 103: 251-259.
Daire X., Clair D., Reinert W., Boudon-Padieu E., 1997. De-tection and differentiation of grapevine yellows phytoplas-mas belonging to the elm yellows group and to the stolbursubgroup by PCR amplification of non-ribosomal DNA.European Journal of Plant Pathology 103: 507-514.
Danielli A., Bertaccini A., Vibio M., Mori N., Murari E.,Posenato P., Girolami V., 1996a. Detection and molecularcharacterization of phytoplasmas in the planthopper Met-calfa pruinosa (Say) (Homoptera: Flatidae). PhytopathologiaMediterranea 35: 62-65.
Danielli A., Bertaccini A., Vibio M., Rapetti S., 1996b. Identi-ficazione di fitoplasmi associati allo scopazzo dell’ olivo.Atti Convegno Annuale SIPaV, Udine 1996, C28-C29.
Davis R.E., Dally E.L., Gundersen D.E., Lee I.-M., Habili N.,1997a. «Candidatus phytoplasma australiense,» a new phy-toplasma taxon associated with Australian grapevine yel-lows. International Journal of Systematic Bacteriology 47:262-269.
Davis R.I., Schneider B., Gibb K.S., 1997b. Detection anddifferentiation of phytoplasmas in Australia. AustralianJournal of Agricultural Research 48: 535-544.
Doi Y., Teranaka M., Yora K., Asuyama H., 1967. Mycoplas-ma- or PLT group-like microorganisms found in thephloem elements of plants infected with mulberry dwarf,potato witches’ broom, aster yellows, or paulownia witch-es’ broom. Annals of the Phytopathological Society of Japan33: 259-266.
Firrao G., Carraro L., Gobbi E., Locci R., 1996a. Molecularcharacterization of a phytoplasma causing phyllody inclover and other herbaceous hosts in Northern Italy. Euro-pean Journal of Plant Pathology 102: 817-822.
Journal of Plant Pathology (1998), 80 (1), 3-26 Seemüller et al. 21

Firrao G., Scott S., Smart C., Carraro L., Chang C.-J.,Seemüller E., Kirkpatrick B.C., 1996b. Serological andmolecular genetic characterization of five members of theX-disease phytoplasma clade. IOM Letters 4: 278-279.
Fos A., Danet J.L., Zreik L., Garnier M., Bové J.M., 1992.Use of a monoclonal antibody to detect the stolbur my-coplasmalike organism in plants and insects and to identifya vector in France. Plant Disease 76: 1092-1096.
Fox G.E., Wisotzkey J.D., Jurtshuk P.J., 1992. How close isclose: 16S rRNA sequence identity may not be sufficient toguarantee species identity. International Journal of System-atic Bacteriology 42: 166-170.
Gibb K.S., Persley D.M., Schneider B., Thomas J.E., 1996.Phytoplasmas associated with papaya diseases in Australia.Plant Disease 80: 174-178.
Griffiths H.M., Gundersen D.E., Sinclair W.A., Lee I.-M.,Davis R.E., 1994c. Mycoplasmalike organisms from milk-weed, goldenrod, and spirea represent two new 16S rRNAsubgroups and three new strain subclusters related topeach X-disease MLOs. Canadian Journal of Plant Patholo-gy 16: 255-260.
Griffiths H.M., Sinclair W.A., Davis R.E., Lee I.-M., DallyE.L., Guo Y.-H., Chen T.A., Hibben C.R., 1994a. Charac-terization of mycoplasmalike organisms from Fraxinus, Sy-ringa, and associated plants from geographically diversesites. Phytopathology 84: 119-126.
Griffiths H.M., Sinclair W.A., Gundersen D.E., Lee I.-M.,Davis R.E., 1994b. Characterization of mycoplasmalike or-ganisms detected in plants in the vicinity of natural out-breaks of ash yellows and elm yellows in North America.IOM Letters 3: 259-260.
Gundersen D.E., Lee I.-M., Rehner S.A., Davis R.E., Kings-bury D.T., 1994. Phylogeny of mycoplasmalike organisms(phytoplasmas): a basis for their classification. Journal ofBacteriology 176: 5244-5254.
Gundersen D.E., Lee I.-M., Schaff D.A., Harrison N.A.,Chang C.J., Davis R.E., Kingsbury D.T., 1996. Genomicdiversity and differentiation among phytoplasma strains in16S rRNA groups I (aster yellows and related phytoplas-mas) and III (X disease and related phytoplasmas). Inter-national Journal of Systematic Bacteriology 46: 64-75.
Guo Y.H., Walla J.A., Cheng Z.M., Lee I.-M., 1996. X-dis-ease confirmation and distribution in chokecherry inNorth Dakota. Plant Disease 80: 95-102.
Harrison N.A., Legard D.E., Dibonito R., Richardson P.A.,1997. Detection and differentiation of phytoplasmas asso-ciated with diseases of strawberry in Florida. Plant Disease81: 230.
Harrison N.A., Richardson P.A., Jones P., Tymon A.M.,Eden-Green S.J., Mpunami A.A., 1994a. Comparative in-vestigation of MLOs associated with Caribbean andAfrican coconut lethal decline diseases by DNA hybridiza-tion and PCR assays. Plant Disease 78: 507-511.
Harrison N.A., Richardson P.A., Kramer J.B., Tsai J.H.,1994b. Detection of the mycoplasma-like organism associ-ated with lethal yellowing disease of palms in Florida bypolymerase chain reaction. Plant Pathology 43: 998-1008.
Harrison N.A., Tsai J.H., Richardson P.A., 1996. Etiologicaland epidemiological investigations of periwinkle witches’broom disease in Florida. IOM Letters 4: 197-198.
Honetłlegrová J.F., Vibio M., Bertaccini A., 1996. Electronmicroscopy and molecular identification of phytoplasmasassociated with strawberry green petals in the Czech Re-public. European Journal of Plant Pathology 102: 831-835.
International Committee on Systematic Bacteriology, Sub-committee on the Taxonomy of Mollicutes., 1993. Minutesof the Interim Meetings, 1 and 2 August, 1992, Ames,Iowa. International Journal of Systematic Bacteriology 43:394-397.
International Committee on Systematic Bacteriology, Sub-committee on the Taxonomy of Mollicutes., 1995. Revisedminimum standards for description of new species of theclass Mollicutes (division Tenericutes). International Jour-nal of Systematic Bacteriology 45: 605-612.
International Committee on Systematic Bacteriology, Sub-committee on the Taxonomy of Mollicutes., 1997. Minutesof the interim meetings, 12 and 18 July 1996, Orlando,Florida, USA. International Journal of Systematic Bacteriol-ogy 47: 911-914.
Jarausch W., Saillard C., Dosba F., Bové J.-M., 1994. Differ-entiation of mycoplasmalike organisms (MLOs) in Euro-pean fruit trees by PCR using specific primers derivedfrom the sequence of a chromosomal fragment of the ap-ple proliferation MLO. Applied and Environmental Micro-biology 60: 2916-2923.
Jomantiene R., Davis R.E., Maas J., Dally E.L., 1996. Phyto-plasmas associated with diseases of strawberries in Florida.Phytopathology 86: S123.
Keane G., Edwards A., Clark M.F., 1996. Differentiation ofgroup 16Sr-IB aster yellows phytoplasmas with monoclon-al antibodies. BCPC Symposium Proceedings: Diagnostic inCrop Production, No. 65: 263-268.
Khadhair A.H., Hiruki C., 1995. The molecular genetic relat-edness of willow witches’-broom phytoplasma to theclover proliferation group. Proceedings of the Japan Acade-my Series B - Physical and Biological Sciences 71: 145-147.
22 Phytoplasma classification Journal of Plant Pathology (1998), 80 (1), 3-26

Khadhair A.H., Hwang S.F., Chang K.F., Howard R.J., 1997.Molecular identification of aster yellows phytoplasmas inpurple conflower and monarda based on PCR amplifica-tion and RFLP analysis of 16S rDNA sequences. Zeitschriftfür Pflanzenkrankheiten und Pflanzenschutz 104: 403-410.
Khew K.L., Davis R.E., Ong C.A., Lee I.-M., Su H.J., TsaiM.C., 1991. Detection of a Malaysian mycoplasmalike or-ganism (MLO) and its differentiation from other Asian,European, and North American MLOs by use of clonedchromosomal and extrachromosomal MLO DNA probes.Journal of Plant Protection in the Tropics 8: 167-180.
Kirkpatrick B.C. 1991. Mycoplasma-like organisms-plant andinvertebrate pathogens. In: Balows A., Trüper H.G.,Dworkin M., Harder W., Schleifer K.H. (eds.). The Pro-caryotes, Vol. 3., pp. 4050-4067. Springer, New York.
Kirkpatrick B., Smart C., Gardner S., Gao J.-L., Ahrens U.,Mäurer R., Schneider B., Lorenz K.-H., Seemüller E., Har-rison N., Namba S., Daire X., 1994. Phylogenetic relation-ship of plant pathogenic MLOs established by 16/23S rD-NA spacer sequences. IOM Letters 3: 228-229.
Kirkpatrick B.C., Stenger D.C., Morris T.J., Purcell A.H.,1987. Cloning and detection of DNA from a nonculturableplant pathogenic mycoplasma-like organism. Science 238:197-200.
Kison H., Kirkpatrick B.C., Seemüller E., 1997. Genetic com-parison of the peach yellow leaf roll agent with Europeanfruit tree phytoplasmas of the apple proliferation group.Plant Pathology 46: 538-544.
Kison H., Schneider B., Seemüller E., 1994. Restriction frag-ment length polymorphism within the apple proliferationmycoplasmalike organism. Journal of Phytopathology 141:395-401.
Kollar A., Seemüller E., 1989. Base composition of the DNAof mycoplasmalike organisms associated with various plantdiseases. Journal of Phytopathology 127: 177-186.
Kollar A., Seemüller E., Bonnet F., Saillard C., Bové J.M.,1990. Isolation of the DNA of various plant pathogenicmycoplasmalike organisms from infected plants. Phy-topathology 80: 233-237.
Kuske C.R., Kirkpatrick B.C., 1992. Phylogenetic relation-ships between the western aster yellows mycoplasmalikeorganism and other prokaryotes established by 16S rRNAgene sequence. International Journal of Systematic Bacteri-ology 42: 226-233.
Kuske C.R., Kirkpatrick B.C., Seemüller E., 1991. Differentia-tion of virescence MLOs using western aster yellows my-coplasma-like organism chromosomal DNA probes andrestriction fragment length polymorphism analysis. Journalof General Microbiology 137: 153-159.
Lee I.-M., Bertaccini A., Vibio M., Gundersen D.E., 1995a.Detection of multiple phytoplasmas in perennial fruit treeswith decline symptoms in Italy. Phytopathology 85: 728-735.
Lee I.-M., Bertaccini A., Vibio M., Gundersen D.E., DavisR.E., Mittempergher L., Conti M., Gennari F., 1995b. De-tection and characterization of phytoplasmas associatedwith disease in Ulmus and Rubus in northern and centralItaly. Phytopathologia Mediterranea 34: 174-183.
Lee I.-M., Davis R.E., 1988. Detection and investigation ofgenetic relatedness among aster yellows and other my-coplasmalike organisms by using cloned DNA and RNAprobes. Molecular Plant-Microbe Interactions 1: 303-310.
Lee I.-M., Davis R.E. 1992. Mycoplasmas which infect plantsand insects. In: Maniloff J., McElhaney R.N., Finch L.R.,Baseman J.B. (eds.). Mycoplasmas - molecular biology andpathogenesis, pp. 379-390. American Society for Microbi-ology, Washington, D.C.
Lee I.-M., Davis R.E., Chen T.-A., Chiykowski L.N., FletcherJ., Hiruki C., Schaff D.A., 1992a. A genotype-based systemfor identification and classification of mycoplasmalike or-ganisms (MLOs) in the aster yellow MLO strain cluster.Phytopathology 82: 977-986.
Lee I.-M., Davis R.E., Hiruki C., 1991. Genetic interrelated-ness among clover proliferation mycoplasmalike organisms(MLOs) and other MLOs investigated by nucleic acid hy-bridization and restriction fragment length polymorphismanalyses. Applied and Environmental Microbiology 57:3565-3569.
Lee I.-M., Davis R.E., Hsu H.-T., 1993a. Differentiation ofstrains in the aster yellows mycoplasmalike organism straincluster by serological assay with monoclonal antibodies.Plant Disease 77: 815-817.
Lee I.-M., Davis R.E., Sinclair W.A., DeWitt N.D., Conti M.,1993b. Genetic relatedness of mycoplasmalike organismsdetected in Ulmus spp. in the United States and Italy bymeans of DNA probes and polymerase chain reactions.Phytopathology 83: 829-833.
Lee I.-M., Gundersen D.E., Davis R.E., Chiykowski L.N.,1992b. Identification and analysis of a genomic strain clus-ter of mycoplasmalike organisms associated with Canadianpeach (eastern) X disease, western X-disease, and cloveryellow edge. Journal of Bacteriology 174: 6694-6698.
Lee I.-M., Gundersen D.E., Hammond R.W., Davis R.E.,1994. Use of mycoplasmalike organism (MLO) group-spe-cific oligonucleotide primers for nested-PCR assays to de-tect mixed-MLO infections in a single host plant. Phy-topathology 84: 559-566.
Journal of Plant Pathology (1998), 80 (1), 3-26 Seemüller et al. 23

Lee I.-M., Hammond R.W., Davis R.E., Gundersen D.E.,1993c. Universal amplification and analysis of pathogen16S rDNA for classification and identification of my-coplasmalike organisms. Phytopathology 83: 834-842.
Lee I.-M., Klopmeyer M., Bartoszyk I.M., Gundersen-RindalD.E., Chou T.S., Thomson K.L., Eisenreich R., 1997a.Phytoplasma induced free-branching in commercial poin-settia cultivars. Nature Biotechnology 15: 178-182.
Lee I.-M., Pastore M., Vibio M., Danielli A., Attathorn S.,Davis R.E., Bertaccini A., 1997b. Detection and character-ization of a phytoplasma associated with annual blue grass(Poa annua) white leaf disease in southern Italy. EuropeanJournal of Plant Pathology 103: 251-254.
Liefting L.W., Andersen M.T., Beever R.E., Gardener R.C.,Forster R.L.S., 1996. Sequence heterogeneity in the two16S rRNA genes of phormium little leaf phytoplasma. Ap-plied and Environmental Microbiology 62: 3133-3139.
Lim P.-O., Sears B.B., 1989. 16S rRNA sequence indicatesthat plant-pathogenic mycoplasmalike organisms are evo-lutionarly distinct from animal mollicutes. Journal of Bacte-riology 171: 5901-5906.
Lim P.-O., Sears B.B., 1992. Evolutionary relationships of aplant-pathogenic mycoplasmalike organism and Achole-plasma laidlawii deduced from two ribosomal protein genesequences. Journal of Bacteriology 174: 2606-2611.
Lorenz K.-H., Dosba F., Poggi Pollini C., Llacer G.,Seemüller E., 1994. Phytoplasma diseases of Prunusspecies in Europe are caused by genetically similar organ-isms. Zeitschrift für Pflanzenkrankheiten und Pflanzen-schutz 101: 567-575.
Lorenz K.-H., Schneider B., Ahrens U., Seemüller E., 1995.Detection of the apple proliferation and pear decline phy-toplasmas by PCR amplification of ribosomal and nonri-bosomal DNA. Phytopathology 85: 771-776.
Mäurer R., Seemüller E., 1994. Nature and genetic related-ness of the mycoplasma-like organism causing rubus stuntin Europe. Plant Pathology 44: 244-249.
Mäurer R., Seemüller E., 1996. Witches’ broom of Rhamnuscatharticus: a new phytoplasma disease. Journal of Phy-topathology 144: 221-223.
Mäurer R., Seemüller E., Sinclair W.A., 1993. Genetic relat-edness of mycoplasmalike organisms affecting elm, alder,and ash in Europe and North America. Phytopathology 83:971-976.
Maixner M., Ahrens U., Seemüller E., 1995. Detection of theGerman grapevine yellows (Vergilbungskrankheit) MLOin grapevine, alternative hosts and a vector by a specificPCR procedure. European Journal of Plant Pathology 101:241-250.
Marcone C., Ragozzino A., Credi R., Seemüller E., 1996a. De-tection and characterization of phytoplasmas infectinggrapevine in southern Italy and their genetic relatedness toother grapevine yellows phytoplasmas. PhytopathologiaMediterranea 35: 207-213.
Marcone C., Ragozzino A., Schneider B., Lauer U., SmartC.D., Seemüller E., 1996b. Genetic characterization andclassification of two phytoplasmas associated with spar-tium witches’-broom disease. Plant Disease 80: 365-371.
Marcone C., Ragozzino A., Seemüller E., 1996c. Associationof phytoplasmas with the decline of European hazel insouthern Italy. Plant Pathology 45: 857-863.
Marcone C., Ragozzino A., Seemüller E., 1996d. Detection ofan elm yellows-related phytoplasma in eucalyptus trees af-fected by little-leaf disease in Italy. Plant Disease 80: 669-673.
Marcone C., Ragozzino A., Seemüller E., 1996e. Europeanstone fruit yellows phytoplasma as the cause of peach veinenlargement and other yellows and decline diseases ofstone fruits in southern Italy. Journal of Phytopathology144: 559-564.
Marcone C., Ragozzino A., Seemüller E., 1997a. Detectionand identification of phytoplasmas in yellows-diseasedweeds in Italy. Plant Pathology 46: 530-537.
Marcone C., Ragozzino A., Seemüller E., 1997b. Detectionand identification of phytoplasmas infecting vegetable, or-namental, and forage crops in southern Italy. Journal ofPlant Pathology 79: 211-217.
Marcone C., Ragozzino A., Seemüller E., 1997c. Detectionand identification of phytoplasmas infecting wild plantspecies in Southern Italy. Proceedings 10th Congress of theMediterranean Phytopathological Union, Montpellier 1997,251-255.
Marcone C., Ragozzino A., Seemüller E., 1997d. Detection ofBermuda grass white leaf diseases in Italy and characteriza-tion of the associated phytoplasma by RFLP analysis. PlantDisease 81: 862-866.
Marcone C., Ragozzino A., Seemüller E., 1997e. Identificationand characterization of the phytoplasma associated withelm yellows in southern Italy and its relatedness to otherphytoplasmas of the elm yellows group. European Journalof Forest Pathology 27: 45-54.
Marcone C., Ragozzino A., Seemüller E., 1997f. Witches’broom of Sarothamnus scoparius: a new disease associatedwith a phytoplasma related to the spartium witches’-broom agent. Journal of Phytopathology 145: 159-161.
Marwitz R., 1990. Diversity of yellows disease agents in plantinfections. Zentralblatt für Bakteriologie, Suppl. 20: 431-434.
24 Phytoplasma classification Journal of Plant Pathology (1998), 80 (1), 3-26

McCoy R.E., Caudwell A., Chang, C.J., Chen T.A., Chiykows-ki L.N., Cousin M.T., Dale J.L., de Leeuw G.T.N., GolinoD.A., Hackett K.J., Kirkpatrick B.C., Marwitz R., PetzoldH., Sinha R.C., Sugiura M., Whitcomb R.F., Yang I.L.,Zhu B.M., Seemüller E. 1989. Plant diseases associatedwith mycoplasma-like organisms. In: Whitcomb R.F., Tul-ly J.G. (eds.). The Mycoplasmas, Vol. V., pp. 545-640.Academic Press, San Diego.
Minucci C., Rajan J., Clark M.F., 1996. Differentiation of phy-toplasmas associated with sweet potato little leaf and otherphytoplasmas in the faba bean phyllody cluster. BritishCrop Protection Council Symposium Proceedings, Diagnos-tics in Crop Production, Warwick 1996, No. 65, 61-66.
Minucci C., Ramasso E., Dellavalle G., Lisa V., Masenga V.,Boccardo G., 1995. Caratterizzazione di un organismosimile a micoplasmi in zucchino in Liguria. Informatore Fi-topatologico 45: 61-64.
Murray R.G.E., Schleifer K.H., 1994. Taxonomic notes: aproposal for recording the properties of putative taxa ofprocaryotes. International Journal of Systematic Bacteriolo-gy 44: 174-176.
Nakashima, K., Hayashi, T., Chaleeprom, W., Wongkaew, P.,Sirithorn, P., 1996. Complex phytoplasma flora in north-east Thailand as revealed by 16S rDNA analysis. Annals ofthe Phytopathological Society of Japan 62: 57-60.
Namba S., Oyaizu S., Kato S., Iwanami S., Tsuchizaki T.,1993. Phylogenetic diversity of phytopathogenic mycoplas-malike organisms. International Journal of Systematic Bac-teriology 43: 461-467.
Neimark H.C., Kirkpatrick B.C., 1993. Isolation and charac-terization of full-length chromosomes from non-culturableplant-pathogenic mycoplasma-like organisms. MolecularMicrobiology 7: 21-28.
Okuda S., Prince J.P., Davis R.E., Dally E.L., Lee I.-M., Mo-gen B., Kato S., 1997. Two groups of phytoplasmas fromJapan distinguished on the basis of amplification and re-striction analysis of 16S rDNA. Plant Disease 81: 301-305.
Padovan A.C., Gibb K.S., Daire X., Boudon-Padieu E., 1996.A comparison of the phytoplasma associated with Aus-tralian grapevine yellows to other phytoplasmas ingrapevine. Vitis 35: 189-194.
Poggi Pollini C., Bissani R., Giunchedi L., Vindimian E.,1996. First report of phytoplasma infection in olive trees(Olea europea L.). Journal of Phytopathology 144: 109-111.
Prince J.P., Davis R.E., Wolf T.K., Lee I.-M., Mogen B.D.,Dally E.L., Bertaccini A., Credi R., Barba M., 1993. Mole-cular detection of diverse mycoplasmalike organisms(MLOs) associated with grapevine yellows and their classi-fication with aster yellows, X-disease, and elm yellowsMLOs. Phytopathology 83: 1130-1137.
Saeed E.M., Sarindu N., Davies D.L., Clark M.F., Roux J.,Cousin M.T., 1994. Use of monoclonal antibodies to identi-fy mycoplasmalike organisms (MLOs) from the Sudan andfrom Thailand. Journal of Phytopathology 142: 345-349.
Sarindu N., Clark M.F., 1993. Antibody production and iden-tity of MLOs associated with sugar-cane whiteleaf diseaseand bermuda-grass whiteleaf disease from Thailand. PlantPathology 42: 396-402.
Schneider B., Ahrens U., Kirkpatrick B.C., Seemüller E.,1993. Classification of plant-pathogenic mycoplasma-likeorganisms using restriction-site analysis of PCR-amplified16S rDNA. Journal of General Microbiology 139: 519-527.
Schneider B., Cousin M.T., Klinkong S., Seemüller E., 1995a.Taxonomic relatedness and phylogenetic positions of phy-toplasmas associated with disease of faba bean, sunnhemp,sesame, soybean, and eggplant. Zeitschrift fürPflanzenkrankheiten und Pflanzenschutz 102: 225-232.
Schneider B., Gibb K.S., 1997. Detection of phytoplasmas indeclining pears in southern Australia. Plant Disease 81:254-258.
Schneider B., Gibb K.S., Seemüller E., 1997a. Sequence andRFLP analysis of the elongation factor Tu gene used in dif-ferentiation and classification of phytoplasmas. Microbiolo-gy 143: 3381-3389.
Schneider B., Marcone C., Kampmann M., Ragozzino A., Led-erer W., Cousin M.T., Seemüller E., 1997b. Characteriza-tion and classification of phytoplasmas from wild and culti-vated plants by RFLP and sequence analysis of ribosomalDNA. European Journal of Plant Pathology 103: 675-686.
Schneider B., Seemüller E., 1994a. Presence of two sets of ri-bosomal genes in phytopathogenic mollicutes. Applied andEnvironmental Microbiology 60: 3409-3412.
Schneider B., Seemüller E., 1994b. Studies on the taxonomicrelationships of mycoplasmalike organisms by Southernblot analysis. Journal of Phytopathology 141: 173-185.
Schneider B., Seemüller E., Smart C.D., Kirkpatrick B.C.1995b. Phylogenetic classification of plant pathogenic my-coplasma-like organisms or phytoplasmas. In: Razin S.,Tully J.G. (eds.). Molecular and diagnostic procedures inmycoplasmology, Vol. I., pp. 369-380. Academic Press,San Diego, CA.
Sears B.B., Lim P.-O., Holland N., Kirkpatrick B.C., Klom-parens K.L., 1989. Isolation and characterization of DNAfrom a mycoplasmalike organism. Molecular Plant-MicrobeInteractions 2: 175-180.
Seemüller E., Schneider B., Mäurer R., Ahrens U., Daire X.,Kison H., Lorenz K.-H., Firrao G., Avinent L., Sears B.B.,
Journal of Plant Pathology (1998), 80 (1), 3-26 Seemüller et al. 25

Stackebrandt E., 1994. Phylogenetic classification of phy-topathogenic mollicutes by sequence analysis of 16S ribo-somal DNA. International Journal of Systematic Bacteriolo-gy 44: 440-446.
Shaw M.E., Kirkpatrick B.C., Golino D.A., 1993. The beetleafhopper-transmitted virescence agent causes tomato bigbud disease in California. Plant Disease 77: 290-295.
Sinha R.C., Benhamou N., 1983. Detection of mycoplasmalikeantigens from aster yellows-diseased plants by two serolog-ical procedures. Phytopathology 73: 1199-1202.
Stackebrandt E., Goebel B.M., 1994. Taxonomic note: a placefor DNA-DNA reassociation and the 16S rRNA sequenceanalysis in the present species definition in bacteriology.International Journal of Systematic Bacteriology 44: 846-849.
Stanosz G.R., Heimann M.F., Lee I.-M., 1997. Purple cone-flower is a host of the aster yellows phytoplasma. PlantDisease 81: 424.
Tymon A.M., Jones P., Harrison N.A., 1997. Detection anddifferentiation of African coconut phytoplasmas: RFLPanalysis of PCR-amplified 16S rDNA and DNA hybridisa-tion. Annals of Applied Biology 131: 91-102.
Vibio M., Bertaccini A., Hostachy B., Rapetti S., 1997. Differ-ent phytoplasmas asssociated with identical symptoms:The case of ranunculus phyllody. Proceedings 10th Con-gress of the Mediterranean Phytopathological Union, Mont-pellier 1997, 331-335.
Vibio M., Bertaccini A., Lee I.-M., Davis R.E., Clark M.F.,1996. Differentiation and classification of aster yellows andrelated European phytoplasmas. Phytopathologia Mediter-ranea 35: 33-42.
Vibio M., Bertaccini A., Lee I.-M., Davis R.E., Minucci C.,Milne R.G., 1994. Etiology of lettuce yellows in Italy: ge-netic characterization of associated mycoplasmalike organ-isms. Phytopathologia Mediterranea 33: 179-186.
Vibio M., Bertaccini A., Schilir E., Rapetti S., 1995a. Carat-terizzazione molecolare di fitoplasmi associati a virescenzain anemone. Informatore Fitopatologico 45: 60-62.
Vibio M., Camele I., Bertaccini A., Rana G.L., D’ Aloisio V.,Benni A., 1995b. Metodi diagnostici molecolari per l’ indi-viduazione di infezioni da fitoplasmi in cipolla. L’Informa-tore Agrario 35: 75-77.
Viswanathan R., 1997. Detection of phytoplasmas associatedwith grassy shoot disease of sugarcane by ELISA tech-niques. Zeitschrift für Pflanzenkrankheiten und Pflanzen-schutz 104: 9-16.
Wayne L.G., Brenner D.J., Colwell R.R., Grimont P.A.D.,Kandler O., Krichevsky M.I., Moore L.H., Moore W.E.C.,Murray R.G.E., Stackebrandt E., Starr M.P., Truper H.G.,1987. Report of the ad hoc committee on reconciliation ofapproaches to bacterial systematics. International Journalof Systematic Bacteriology 37: 463-464.
Weisburg W.G., Tully J.G., Rose D.L., Petzel J.P., Oyaizu H.,Yang D., Mandelco L., Sechrest J., Lawrence T.G., VanEtten J., Maniloff J., Woese C.R., 1989. A phylogeneticanalysis of the mycoplasmas: basis for their classification.Journal of Bacteriology 171: 6455-6467.
Woese C.R., 1987. Bacterial evolution. Microbiological Review51: 221-271.
Wongkaew P., Hanboonsong Y., Sirithorn P., Choosai C.,Boonkrong S., Tinnangwattana T., Kitchareonpanya R.,Damak S., 1997. Differentiation of phytoplasmas associat-ed with sugarcane and gramineous weed white leaf diseaseand sugarcane grassy shoot by RFLP and sequencing. The-oretical and Applied Genetics 95: 660-663.
Zhu S.F., Lee I.-M., Gundersen D.E., Zhang C.L., Hadidi A.,1996. Phytoplasmas associated with cherry lethal yellowsand jujube witches’-broom in China represent a new Can-didatus subspecies level taxon. IOM Letters 4: 218.
Zreik L., Carle P., Bové J.M., Garnier M., 1995. Characteriza-tion of the mycoplasmalike organism associated withwitches’-broom disease of lime and proposition of a Candi-tatus taxon for the organism, «Canditatus Phytoplasma au-rantifolia». International Journal of Systematic Bacteriology45: 449-453.
26 Phytoplasma classification Journal of Plant Pathology (1998), 80 (1), 3-26





























