Embed Size (px)
Citation preview

Investigations of the Pathway of Incorporation and Functionof Lamin A in the Nuclear LaminaJOANNA A. DYER,1 BIRGITTE E. LANE,2 AND CHRISTOPHER J. HUTCHISON1*1Department of Biological Sciences, The University of Dundee, Dundee DD1 4HN, Scotland2Department of Anatomy and Physiology, The University of Dundee Dundee, DD1 4HN, Scotland
KEY WORDS lamins; lamina; nuclear envelope; Xenopus egg extracts
ABSTRACT Cell-free extracts of Xenopus eggs were used to investigate how differing laminsub-types are incorporated into a lamina and how lamina composition influences DNA replication.Purified recombinant human lamin A (HlaminA) was inoculated into egg extracts, which support invitro nuclear assembly. The route of incorporation of Hlamin A into the lamina was compared toXenopus lamin B3 (XlaminB3), the major endogenous lamin sub-type in egg extracts. While XlaminB3 was incorporated into the lamina directly, HlaminA first accumulated at nucleoplasmic foci,before entering the lamina. When HlaminA was inoculated into extracts depleted of XlaminB3 itentered nuclei efficiently and was incorporated into nucleoplasmic foci. However, in the absence ofXlaminB3, HlaminA remained in the foci and did not enter the nuclear envelope. When HlaminAentered the nuclear envelope, it did not influence DNA replication. Nuclei containing HlaminAinitiated DNA replication on queue and in addition the spatial distribution of replication centres inthese nuclei was identical to controls. Taken together, these results suggest that the incorporation ofA-type lamins into the nuclear envelope is dependent upon the presence of B-type lamins. However,the presence of A-type lamins alone is not sufficient to influence the spatial and temporal order ofDNA replication. Microsc. Res. Tech. 45:1–12, 1999. r 1999 Wiley-Liss, Inc.
INTRODUCTIONThe nuclear lamina consists of a network of interme-
diate type filaments that line the nucleoplasmic face ofthe inner nuclear membrane and that interconnectsnuclear pores (reviewed by Hutchison et al., 1994).Lamina filaments are composed of type V intermediatefilaments, termed lamins (reviewed by Quinlan et al.,1995). Lamins fall into two subfamilies termed A-typeand B-type. Lamins A and C comprise the A-typesubfamily and are synthesised from alternatively splicedtranscripts of the same gene (Fisher et al., 1986). Incontrast, at least three B-type genes are found invertebrates and are classified as lamins B1, B2, and B3(Stick, 1988). Lamins have been assigned a number offunctions, including the targeting of nuclear membranevesicles to chromatin after mitosis (Benevente andKrohne, 1986; Burke and Gerace, 1986), the correctassembly of nuclear pores (Goldberg et al., 1995), and,indirectly, in the organisation of replication domains(Jenkins et al., 1995; Zhang et al., 1996).
A-type and B-type lamins differ is several respectsincluding their behaviour at mitosis (Gerace and Blo-bel, 1980), post-translational processing (Beck et al.,1990; Farnsworth et al., 1989), and expression. B-typelamins are ubiquitous components of all cells althoughdifferent cells do express different B-type lamins (Ben-evente et al., 1985; Hoger et al., 1988; Stick andHausen, 1985). In contrast, the expression of A-typelamins is restricted to differentiated tissues (Lehner etal., 1987; Rober et al., 1989) and is first observed inembryos at the time of organogenesis. Consistent withthis observation, pluripotent cells can be induced toexpress A-type lamins in culture by treatments that
induce differentiation (Lebel et al., 1987; Rober et al.,1990) while loss of A-type lamin expression is associ-ated with dedifferentiation in some tumour cells (Bro-ers et al., 1993; Paulin-Levasseur et al., 1989). Thecorrelation between A-type lamin expression and celldifferentiation has led some workers to speculate thatA-type lamins may facilitate differential gene expres-sion by anchoring chromatin at the nuclear envelope orby sequestering inhibitors (reviewed by Nigg, 1989).Consistent with this hypothesis, lamin A displays ahigher affinity for chromatin binding than do B-typelamins (Hoger et al., 1991). In addition, lamins A and Cbind the negative growth regulator p110Rb (Ozaki et al.,1994). Moreover, ectopic expression of lamin A in myo-blasts appears to promote differentiation (Lourim andLim, 1990).
Despite this evidence, the precise function of A-typelamins is unclear and in particular there is currently noinformation regarding the function of A-type lamins inrelation to their structure. The nuclear lamina can beorganised differently in different cells. In embyroniccells that express only B-type lamins, the lamina is arelatively flat 2-dimensional lattice (Aebi et al., 1986;Goldberg and Allen, 1992). In contrast, in some cellsthat express multiple lamin types (particularly syno-vial cells) the lamina is a thick 3-dimensional structurethat extends some 200 nm into the nucleoplasm and is
Contract grant sponsor: Wellcome Trust; Contract grant sponsor: CancerResearch Campaign.
*Correspondence to: Christopher J. Hutchison, Department of BiologicalSciences, The University of Dundee, Dundee DD1 4HN, Scotland.
Received 1 October 1998; accepted in revised form 19 November 1998
MICROSCOPY RESEARCH AND TECHNIQUE 45:1–12 (1999)
r 1999 WILEY-LISS, INC.

in close contact with peripheral heterochromatin(Hoeger et al., 1990). Several questions arise from thesesimple observations. In particular, the nature of laminafilaments in situations in which more than one lamintype is expressed is unknown. While lamins do formheterotypic associations in vitro (Ellis et al., 1997), it isnot known whether different lamin sub-types co-existwithin the same filaments. In addition, the distributionof chromatin in cells that express A-type lamins isdistinctly different when compared to cells that expressonly B-type lamins and this is reflected by changes inpatterns of DNA replication (Kill et al., 1991).
To address these questions, we have investigated theassembly of lamin A into the nuclear lamina, whenadded to cell-free extracts of Xenopus eggs that supportnuclear assembly in vitro (Blow and Laskey, 1986;Hutchison et al., 1987). The extracts are derived frommeiotic cells that contain only B-type lamins (Lourimand Krohne, 1993). Therefore, the extracts provide anexperimental situation in which the behaviour andinfluence of A-type lamins can be investigated whenadded as purified exogenous proteins. Recombinantlamin A, when added to the extracts, did form hetero-typic associations with the endogenous lamin B3. How-ever, the pathway of lamin A assembly into the nuclearlamina differed from lamin B3 in that lamin A firstaccumulated at nucleoplasmic foci, prior to its incorpo-ration into the lamina. When the extracts were de-pleted of endogenous lamin B3, lamin A was stillincorporated into nucleoplasmic foci, but failed to accu-mulate at the nuclear envelope, suggesting that laminA is not assembled into lamina filaments in the absenceof B-type lamins. The timing of DNA replication and thedistribution of replication domains were both unaf-fected by the presence of lamin A in the lamina,indicating that lamin A alone is not sufficient to influ-ence the distribution of replication domains.
MATERIALS AND METHODSAntibodies
The following antibodies were used in this investiga-tion: human auto-immune serum 1008 that detectslamins A and C (see below); mAb L6 8A7, which detectsXenopus lamins B1, B2, and B3 (Stick and Hausen, 1985)and human lamins A and C (Dyer et al., 1997); mAb L65D5, which detects Xenopus lamin B3 (Stick, 1988);mAb JoL2, which detects human lamins A and C (Dyeret al., 1997); LN43, which detects lamin B2 (Kill andHutchison, 1995); and PC7, which detects PCNA from arange of species (Waseem and Lane, 1990).
Serum 1008 was used in immunofluorescence assays,since previous studies had revealed that mAbs againstA-type lamins could be unreliable markers of lamindistribution due to epitope masking (Dyer et al., 1997).Preliminary investigations using auto-immune serum1008 in replication assays demonstrated that it con-tained anti-DNA antibodies in addition to anti-laminA/C antibodies. For this reason, serum 1008 was preab-sorbed against double- and single-stranded DNA priorto use. Each resin was prepared as follows: 1g of DNAcellulose (containing 4 mg DNA, Sigma, St. Louis, MO)was added to 10 ml of 4 M KCl. The resin was mixed andallowed to settle, the supernatant decanted off, and theprocess repeated. The resin was washed by suspensionin 150 ml of extraction buffer (E.B.: 100 mM KCl, 5 mM
MgCl2, 2 mM b-mercaptoethanol, 20 mM Hepes-KOH,pH 7.5), allowed to settle and the supernatant decantedoff. This was repeated and 0.02% sodium azide wasadded to the resin. The preparation was stored at 4°C.To pre-absorb 1008, 1 ml of the prepared resin wasadded to 9 ml of phosphate buffered saline (PBS)containing 10 µl of serum 1008 and mixed for 1 hour atroom temperature by end-over-end rotation. The resinwas pelleted by centrifugation at 1,000 rpm in a SorvalEconospin bench top centrifuge (Du Pont, Wilmington,DE) for 10 min. The supernatant, containing the anti-body, was collected supplemented with 0.02% sodiumazide and 1% newborn calf serum and stored as aworking dilution of the antiserum.
Preparation of Cell Free ExtractsFrom Xenopus Eggs
Extracts were prepared from unfertilised Xenopuseggs by centrifugation according to the method de-scribed by Hutchison (1994). For every 1 ml of extractprepared, approximately 3,000 eggs were used. FemaleXenopus laevis frogs were stimulated to lay, 16 hoursbefore the eggs were required, by injection of 600 i.u.human chorionic gonadotrophic hormone (Chorulon,Intervet Laboratories, Cambridge, UK) into the dorsallymph sac. Eggs were collected in 110 mM NaCl toprevent activation before dejellying in 5 mM DTT, 110mM NaCl, 20 mM Tris-HCl (pH8.5), containing 4%(w/v) Phenol Red (Sigma). Once eggs were dejellied,they were washed three times in 110 mM NaCl. Duringthe last wash, the eggs were inspected for any abnor-malities or evidence of necrosis. Necrotic eggs werediscarded. The dejellied eggs were further washedthree times in ice-cold extraction buffer (EB, 100 mMKCl, 5 mM MgCl2, 2 mM b-mercaptoethanol, 20 mMHepes-KOH, pH 7.5). The eggs were transferred tochilled 3-ml centrifugation tubes and allowed to settlebefore excess buffer was removed. The eggs were thensubjected to centrifugation at 10,000g for 10 minutes at4°C. During centrifugation, the eggs rupture and theextract separates into three fractions: a yellow lipidcap; a cytosolic layer, which varied in colour and texturefrom extract to extract; a large insoluble pellet consist-ing of the yolk platelets, pigment, and chromosomesfrom the germinal vesicles. All of the cytosolic fractionbetween the lipid cap and pellet was removed andmixed with cytochalasin B (final concentration of 50µm/ml) to disrupt microfilaments. This fraction wasclarified by centrifugation to remove excess lipid andpigment. The resulting straw-coloured extract wasmixed with aprotinin (final concentration of 10 µg/ml)and 5% (v/v) glycerol before snap freezing and storagein liquid nitrogen. The activity of all extracts wasassessed using standard procedures (Hutchison, 1994).
Preparation of Sperm TemplatesSperm templates were prepared by a modification of
the method of Hutchison et al. (1987). Approximately90 million sperm per testes were obtained. Male frogswere killed by cervical dislocation and pithed. The bodycavity was opened up by a mid-ventral incision. Theorgans of the intestinal tract were moved to one side toreveal the testis, which are off-white in colour and 5–8mm in length. The testis were removed and rinsed inSUNaSp (250 mM sucrose, 75 mM NaCl, 0.5 mM
2 J.A. DYER ET AL.

spermidine, 0.15 mM spermine) and homogenised in 3ml of SUNaSp in a Wheaton-Dounce glass homogeniserusing a loose-fitting pestle. The homogeniser was incu-bated on ice while particulate matter settled. Thesperm suspension was pipetted off, and dimethyl sul-phoxide (DMSO) and newborn calf serum (NCS) wereboth added to a concentration of 10% (v/v). The concen-tration of sperm was calculated using a haemocytom-eter before sperm were aliquoted at 2 x 105 per vial andstored at -80°C.
Prior to use in replication assays, the sperm werethawed rapidly and diluted in 3 ml of SUNaSp. Thesperm were pelleted by centrifugation at 2,000 rpm in aSorval Econospin bench top centrifuge (DuPont). Thepellet was resuspended in 200 µl of SUNaSP and thenumber of sperm estimated using a haemocytometer.Demembranation of the sperm was carried out byincubating the sperm with 4 µl of 10 mg/ml lysolecithinfor 90 seconds while agitating the tube gently. To stopthe reaction, 3 ml of SUNaSp containing 3 mg/ml BSAwere added and the sperm pelleted by centrifugation.The pellet was resuspended in SUNaSp to give a finalconcentration of 2 x 105 sperm per µl.
Preparation of Recombinant Lamin APurified recombinant lamin A (Moir et al., 1991) was
generously provided by Dr. Murray Stewart (MRC,Cambridge, UK) as a 6 mg ml-1 suspension in 6 M urea,50 mM Tris, pH 8.0, and 5 mM DTT. Prior to use, thelamin A was dialysed in 20-µl aliquots using the Pierce(Rockford, IL) microdialysis system 100. The lamin Awas dialysed at 4°C for 4 hours against Buffer A (3 MUrea, 50 mM Tris, pH 8.0, 2 mM DTT) and overnight at4°C against buffer B (50 mM Tris, pH 8.0, 500 mMNaCl, and 1 mM DTT) or modified extraction buffer(500 mM KCl, 20 mM Hepes, pH 7.4, MgCl2). Typically60% of the lamin A was precipitated during dialysis. Todetermine the state and concentration of the solublelamin A after dialysis, samples of lamin A removed beforeand after dialysis were compared by SDS PAGE and silverstaining or with a Bradford assay. Dialysed recombi-nant lamin A was added to egg extracts at a finalconcentration of 80 µg ml-1 for nuclear assembly assays.
Replication AssaysDemembranated sperm heads were incubated in
extract at a density of 2 x 105/100 µl extract in thepresence of (unless otherwise stated) an energy regener-ating system: 2 mM ATP, 20 mM creatinine phosphate,and 0.5 mg/ml creatinine phosphokinase. Replicationwas monitored by the incorporation of either Biotin-11-dUTP added as a continuous label, or Biotin-16-dUTPadded as a pulse label, 30 minutes prior to fixation. Theworking concentration of both thymidine analogueswas 4 µM. The final reaction mix was divided into 10-µlaliquots and incubated at 21°C. At regular time inter-vals throughout the incubation, aliquots were removedand fixed by the addition of 200 µl of 1 mM ethyleneglycol bis(succinidyl succinate) dihydrochloride (EGS)in one-third strength extraction buffer. The sampleswere incubated at 37°C for 30 minutes. Pronuclei wererecovered onto glass coverslips by centrifugationthrough 30% glycerol in one-third strength extractionbuffer, at 1,600 rpm using a Sorvall Econospin centri-fuge (Du Pont). After removing the glycerol, coverslips
were left for 30 minutes at room temperature to air dry.This allowed the pronuclei to adhere to the coverslipsbefore the primary antibody was added. The primaryantibody was incubated on the coverslips overnight at4°C. The coverslips were rinsed in PBS before additionof the appropriate FITC or TRITC conjugated second-ary antibody (DakoPatts, Carpinteria, CA; diluted 1:25and 1:40, respectively, with PBS/1% (v/v) NCS) and/orTexas Red-streptavidin (Amersham, Arlington Heights,IL; diluted 1:50 with PBS/1% (v/v) NCS) to visualiseBiotin-dUTP incorporation. Incubations with second-ary antibodies were either 4°C for 4 hours or at roomtemperature for 1 hour. Finally the coverslips werewashed in PBS and each coverslip was mounted in 10 µlof Mowiol (Aldrich, Gillingham, UK) containing DAPIat a concentration of 2 µg/ml (Boeringer-Mannheimer,Indianapolis, IN). The slides were viewed using a Zeiss(Welwyn Garden City, UK) photomicroscope.
Xenopus Pronuclear IsolationPronuclei formed in Xenopus extracts were isolated
as follows: replication assays were established as abovebut the reaction mix was divided into 40-µl aliquots.Nuclear reformation was monitored by fluorescencemicroscopy as follows: 2 µl of sample was withdrawnand diluted in 5 µl of wet mount (15 mM Pipes, pH 7.2,80 mM KCl, 5 mM EDTA, 15 mM NaCl, 10 µg/ml DAPI,3.3 % formaldehyde) and viewed using a Zeiss photomi-croscope. When over 80% of the sperm had assembledinto pronuclei (approximately 3 hours), all the aliquotswere pooled and diluted in 3 ml of ice-cold nuclearisolation buffer (NIB, 60 mM KCl, 15 mM NaCl, 15 mMTris-HCl, pH 7.4, 10 µg/ml leupeptin, and 10 µg/mlaprotinin). The dilution was layered over 1 ml of a 60%percoll cushion (v/v in NIB). Pronuclei were recoveredat the percoll interface using a pasteur pipette anddiluted in 500 µl of NIB. The pronuclei were thenlayered over a sucrose cushion (25% w/v in NIB) andpelleted at 4,000g in a bench top Eppendorf centrifugefor 10 minutes. If the pellet appeared dirty, then thelast step was repeated. The pellet was finally resus-pended in 1 x SDS sample buffer to give 1 x 105
pronuclei/10 µl volume.
Immuno-Depletion of Lamin B3 From ExtractsLamin B3 was immuno-depleted from Xenopus egg
extracts using the mAb L6 5D5 linked to magneticDynabeads (Dynal, Wirrel Merseyside, UK). Mock deple-tions using the mAbs LN43 and PC 7 were alsoperformed (Waseem and Lane, 1990). To immuno-deplete lamin B3 from 250 µl of extract, 400 µl ofParamagnetic Dynabeads were washed twice in PBS/1%NCS and three times in PBS. Between each wash, thebeads were recovered using a Dynal magnet particelseparater. The beads were resuspended in 800 µl of L65D5 tissue culture supernatant and left to incubateovernight at 4°C on a rotary wheel. A parallel incuba-tion was set up with either 800 µl of the mAb PC 7tissue culture supernatant. After incubation in theprimary antibody, the beads were collected using themagnet. The beads were washed 3 times in PBS andtwice in SUNaSp. The beads were resuspended inSUNaSp and each suspension divided into 2. Onealiquot was left on the rotary wheel and the secondaliquot was pelleted on the magnet and the buffer
3LAMIN A ASSEMBLY AND FUNCTION
removed. The pelleted beads were resuspended in 250µl of extract and left on a rotary wheel at roomtemperature for 30 minutes (incubation 1). After thistime the Dynabeads were removed from the extract(pellet 1) and the second aliquot of beads pelleted andresuspended in the partially depleted extract. Thesebeads were incubated for a further 30 minutes (incuba-tion 2). Again the beads were pelleted on the magnet(pellet 2). All the beads were washed three times in PBSand resuspened in 200 µl of 2 x SDS SB. Five microli-ters of the depleted extract was diluted at 1:50 in SDSSB for Western blot analysis, which was necessary todemonstrate that the depletion had been successful.The remaining extract was used immediately or frozenin liquid nitrogen for future use.
Immuno-Isolation of Inoculated Human Lamin AHuman lamin A was inoculated into Xenopus egg
extract and immuno-isolated from the extract usingantibodies bound to Dynabeads. The antibodies JoL 2(anti-lamin A), L6 5D5 (anti-lamin B3), and PC 7(anti-PCNA) were bound to Dynabeads using the proto-col outlined above. Each antibody (960 µl) was incu-bated with 480 µl of Dynabead suspension to form anantibody/bead complex. Approximately 21 µg of dial-ysed human lamin A (in 54 µl of dialysis buffer) wasadded to 450 µl of Xenopus egg extract in the presenceof an energy regenerating system. The extract wasdivided into 50 µl aliquots and incubated at 21°C (series1). As a control, an equal volume of dialysis buffer(54 µl) was added to 450 µl of extract containing anenergy regenerating system, divided into 50 µl aliquotsand incubated at 21°C (series 2). One hour after addi-tion of lamin A aliquots were removed from each seriesand antibody-Dynabeads complexes added and incu-bated overnight at 4°C on rotary wheel. The Dynabeadswere collected using a magnet and washed 3 x in PBS.The beads were resuspended in 40 µl of 1 x SDS SB andanalysed by SDS-PAGE and Western blotting.
Bio-Rad Protein Assay (Bradford)To estimate the concentration of soluble lamin A after
dialysis a Bio-Rad (Hemel Hempstead, UK) proteinassay (microassay procedure) was performed accordingto the manufacturer’s instructions using a BSA stan-dard curve.
SDS PAGE and Immunoblotting.SDS-PAGE and immunoblotting were carried out as
described previously (Dyer et al., 1997). Resolved pro-teins were detected by staining with Coomassie bril-liant blue or by ammoniacal silver staining as recom-mended by Harlow and Lane (1988). This methodpermits detection of proteins down to a concentration of1–10 ng per band. Western blotting was also performed
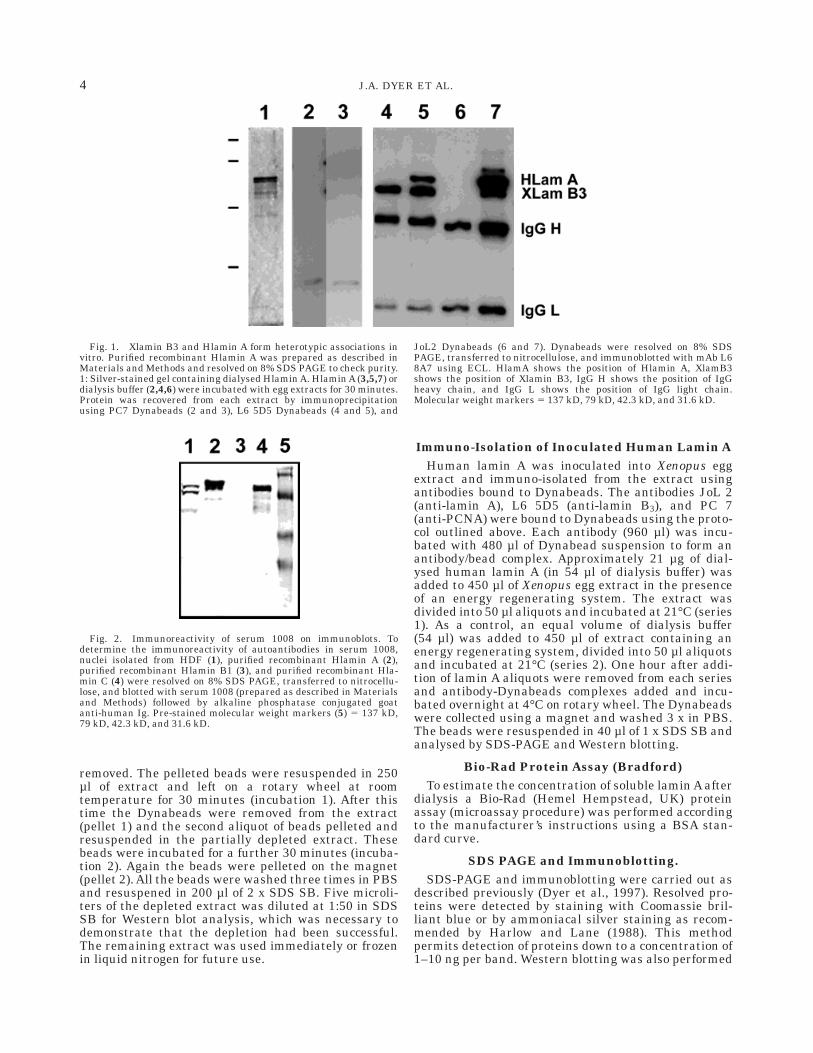
Fig. 1. Xlamin B3 and Hlamin A form heterotypic associations invitro. Purified recombinant Hlamin A was prepared as described inMaterials and Methods and resolved on 8% SDS PAGE to check purity.1: Silver-stained gel containing dialysed Hlamin A. Hlamin A (3,5,7) ordialysis buffer (2,4,6) were incubated with egg extracts for 30 minutes.Protein was recovered from each extract by immunoprecipitationusing PC7 Dynabeads (2 and 3), L6 5D5 Dynabeads (4 and 5), and
JoL2 Dynabeads (6 and 7). Dynabeads were resolved on 8% SDSPAGE, transferred to nitrocellulose, and immunoblotted with mAb L68A7 using ECL. HlamA shows the position of Hlamin A, XlamB3shows the position of Xlamin B3, IgG H shows the position of IgGheavy chain, and IgG L shows the position of IgG light chain.Molecular weight markers 5 137 kD, 79 kD, 42.3 kD, and 31.6 kD.
Fig. 2. Immunoreactivity of serum 1008 on immunoblots. Todetermine the immunoreactivity of autoantibodies in serum 1008,nuclei isolated from HDF (1), purified recombinant Hlamin A (2),purified recombinant Hlamin B1 (3), and purified recombinant Hla-min C (4) were resolved on 8% SDS PAGE, transferred to nitrocellu-lose, and blotted with serum 1008 (prepared as described in Materialsand Methods) followed by alkaline phosphatase conjugated goatanti-human Ig. Pre-stained molecular weight markers (5) 5 137 kD,79 kD, 42.3 kD, and 31.6 kD.
4 J.A. DYER ET AL.

as described by Dyer et al. (1997). Antigens were visual-ised using both colourimetric and ECL detection systems.
RESULTSRecombinant lamin A forms heterotypic associations
with lamin B3 in egg extractsTo investigate associations between human lamin A
(HlaminA) and Xenopus lamin B3 (XlaminB3), purifiedrecombinant Hlamin A (Fig. 1.1) was added to Xenopusegg extracts at a final concentration of 21 µg ml-1. As acontrol, extracts were supplemented with dialysis buffer.One hour after addition of Hlamin A or buffer to the
extracts, aliquots of each extract were immunoprecipi-tated with mAb L6 5D5 (which reacts with lamin B3),mAb JoL 2 (which reacts with Hlamins A and C), or as acontrol mAb PC7 (which reacts with PCNA). Immuno-precipitates were resolved on SDS PAGE and immuno-blotted. Immunodetection of lamins was performedwith L6 8A7 that recognises both B-type Xlamins and Atype Hlamins. As expected, no lamins were detected inPCNA immunoprecipitates (PC7 antibody, Fig. 1.2,3).Lamin B3 was the only lamin detected in lamin B3immunoprecipitates (L6 5D5 antibody) from extractssupplemented with buffer (Fig. 1.4). In contrast, Xla-
Fig. 3. Immunoreactivity of serum 1008 determined by indirectimmunofluorescence. HDF (a,b) were fixed with methanol:acetoneand stained with serum 1008 followed by FITC conjugated goatanti-human Ig (b). The distribution of DNA was revealed with DAPI(a). Alternatively, sperm pronuclei assembled over 60 minutes in vitro
(c– f) were fixed with EGS and stained with serum 1008 followed byFITC conjugated goat anti-human Ig (f) or L6 5D5 followed by FITCconjugated goat anti-mouse Ig (d). The distribution of DNA wasrevealed with DAPI (c and e). Scale bars 5 10 µm.
5LAMIN A ASSEMBLY AND FUNCTION
min B3 and Hlamin A were both detected in lamin B3immunoprecipitates (L6 5D5 antibody) from extractssupplemented with Hlamin A (Fig. 1.5). No lamins weredetected in Hlamin A immunoprecipitates (JoL 2 anti-body) from extracts supplemented with buffer (Fig. 1.6).In contrast, both Xlamin B3 and Hlamin A were de-tected in Hlamin A immunoprecipitates (JoL2 anti-body) from extracts supplemented with Hlamin A (Fig.1.7). Since JoL2 does not cross-react with B-type laminsand L6 5D5 does not cross-react with A-type lamins, thepresence of Hlamin A in L6 5D5 immunoprecipitates(Fig. 1.5) and Xlamin B3 in JoL 2 immunoprecipitates(Fig. 1.7) implies that Hlamin A forms heterotypicassociations with Xlamin B3 when added to egg ex-tracts.
Pathways of Hlamin A and Xlamin B3Incorporation Into the Nuclear Envelope Differ
To determine whether Hlamin A is incorporated intothe nuclear envelope of in vitro assembled nuclei,indirect immunofluorescence and confocal microscopywere performed. For these investigations, we used ahuman auto antibody against lamins A and C termed
1008. The specificity of 1008 was first determined byimmunoblotting. Recombinant Hlamins A, C, and B1together with nuclei isolated from human dermal fibro-blasts (HDF) were resolved on SDS-PAGE and immuno-blotted with 1008. 1008 reacted strongly with HlaminsA and C (Fig. 2, lanes 2 and 4, respectively) but did notreact with Hlamin B1 (Fig. 2, lane 2). In addition, 1008reacted strongly with two bands in isolated HDF nucleimigrating at the predicted Mr of lamins A and C (Fig. 2,lane 1). 1008 also reacted to a lesser extent with a bandmigrating at ,48 kDa. The appearance of this bandwas variable and may have been due to partial degrada-tion of the lamins. Finally, 1008 did not react with anybands when isolated Xenopus sperm pronuclei wereemployed for immunoblotting (data not shown).
Next, we tested the reactivity of 1008 in indirectimmunofluorescence assays. 1008 produced strongnuclear envelope fluorescence when used to stain HDF(Fig. 3 a and b). In contrast, 1008 did not stain spermpronuclei (Fig. 3e and f), whereas in parallel prepara-tions (prepared after 60 minutes incubation in eggextracts) the nuclear envelope of ,96% of nuclei werestained strongly with L6 5D5 (Fig. 3c and d). Taken
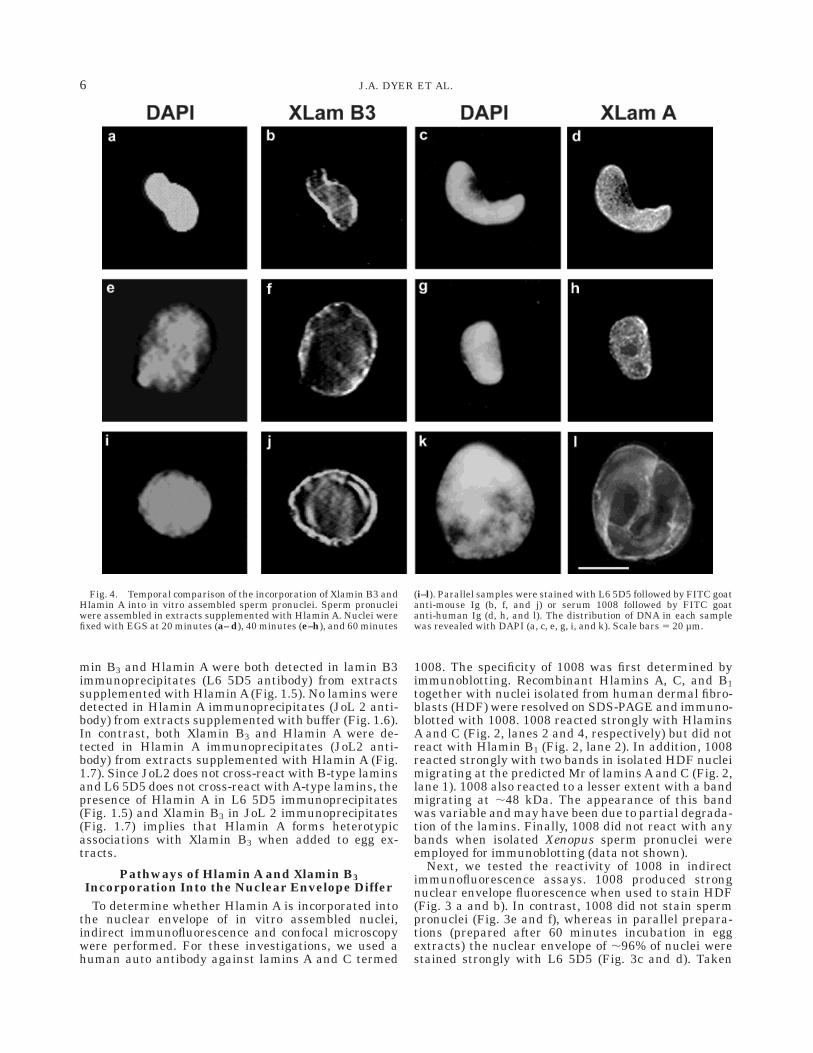
Fig. 4. Temporal comparison of the incorporation of Xlamin B3 andHlamin A into in vitro assembled sperm pronuclei. Sperm pronucleiwere assembled in extracts supplemented with Hlamin A. Nuclei werefixed with EGS at 20 minutes (a– d), 40 minutes (e–h), and 60 minutes
(i–l). Parallel samples were stained with L6 5D5 followed by FITC goatanti-mouse Ig (b, f, and j) or serum 1008 followed by FITC goatanti-human Ig (d, h, and l). The distribution of DNA in each samplewas revealed with DAPI (a, c, e, g, i, and k). Scale bars 5 20 µm.
6 J.A. DYER ET AL.

together these results show that serum 1008 reactsspecifically with lamins A and C and does not cross-react with components of Xenopus sperm pronuclei.Therefore, we used serum 1008 and L6 5D5 to comparethe pathways of Hlamin A and Xlamin B3 incorporationinto the nuclear envelope of in vitro assembled spermpronuclei. Xenopus egg extracts were supplementedwith Hlamin A as described above. Sperm pronucleiwere then assembled in the extracts and prepared at20-minute intervals for indirect immunofluorescence.Lamin B3 was first detected in in vitro assembled nucleiafter 20 minutes when ,35% of nuclei displayed strongnuclear rim staining with L6 5D5 (Figs. 4a,b, 5).Typically, nuclei at this time point were sausage shaped.After 40 minutes incubation, 85% of nuclei displayedstrong nuclear rim staining with L6 5D5 (Figs. 4e,f, 5).Typically, nuclei were fully rounded at this time point.By 60 minutes, nuclei had adopted the typical large sizeof sperm pronuclei and displayed continuous rim stain-ing with L6 5D5 (Fig. 4i,j, 5). The incorporation ofHlamin A into sperm pronuclei differed both temporallyand spatially. After 20 minutes incubation, only 15% ofsperm pronuclei had accumulated Hlamin A (as de-tected by 1008) (Fig 5). Moreover, these nuclei dis-played a predominantly punctate nucleoplasmic stain-
ing pattern with 1008 (Fig. 4c and d). After 40 minutesincubation, the fraction of nuclei stained with 1008 hadincreased to 65% (Fig. 5) and with the adoption of amore rounded morphology, increased nuclear rim stain-ing was observed (Fig. 4g and h). After 60 minutesincubation, nuclei were rounded and enlarged and mostof these displayed nuclear rim staining that was indis-tinguishable from staining with L6 5D5 (compare Fig.4k and l with Fig. 3c and d). To confirm that Hlamin Awas distributed primarily within the nucleoplasm after20– 40 minutes incubation, samples stained with 1008were examined with a laser scanning confocal micro-scope. Z-sections were collected at 1-µm steps throughsperm pronuclei and mid-sections were projected. Twoexamples of mid-sections are displayed in Figure 6 andcompared to a typical mid-section stained with L6 5D5.L6 5D5 gives rise predominantly to a nuclear rim stain(Fig. 6a). In contrast, 1008 stains granular structuresthat are distributed throughout the nucleoplasm (Fig.6b). Taken together, these data suggest that the inco-poration of Xlamin B3 and Hlamin A into the nuclearenvelope occurs through different pathways. Moreover,Xlamin B3 is incorporated into the nuclear envelopebefore Hlamin A.
Fig. 5. Xlamin B3 is incorporated into the lamina before Xlamin A.Sperm pronuclei were assembled into extracts supplemented withHlamin A. The appearance of Xlamin B3 and Hlamin A was investi-gated in samples of nuclei at 20-minute intervals by indirect immuno-
fluorescence as described in Figure 4. At each time point, 200 nucleiwere scored on each of duplicate slides and the percentage showingnuclear incorporation of each lamin was expressed. Squares 5 XlaminB3, circles 5 Hlamin A.
7LAMIN A ASSEMBLY AND FUNCTION
To determine whether the incorporation of Hlamin Ainto the nuclear envelope is dependent upon the pres-ence of Xlamin B3, immunodepletion experiments wereperformed. Xlamin B3 was depleted from egg extractsusing L6 5D5 coupled to Dynabeads. As a control mock,depleted extracts were prepared using PC7 coupled toDynabeads. The depleted extracts and mock depletedextracts were supplemented with Hlamin A and itsincorporation into the nuclear envelope was investi-gated. Nuclei assembled in mock-depleted extractsincorporated lamin B3 into the nuclear envelope effi-ciently (Fig. 7a and b). In contrast, even after prolongedincubation, lamin B3 was not observed in the nuclearenvelope of nuclei assembled in L6 5D5 depleted ex-tracts (Fig. 7c and d). Hlamin A did appear at thenuclear envelope of nuclei assembled in mock-depletedextracts after 60 minutes incubation (Fig. 7e and f). Incontrast, Hlamin A remained within nucleoplasmic fociin nuclei assembled in L6 5D5 depleted extracts (Fig. 7gand h). These results suggest that the incorporation ofHlamin A into the nuclear envelope of in vitro as-sembled nuclei is dependent upon the presence ofXlamin B3.
The presence of Hlamin A in the nuclear envelopedoes not alter the timing of DNA replication or thedistribution of replication centres.
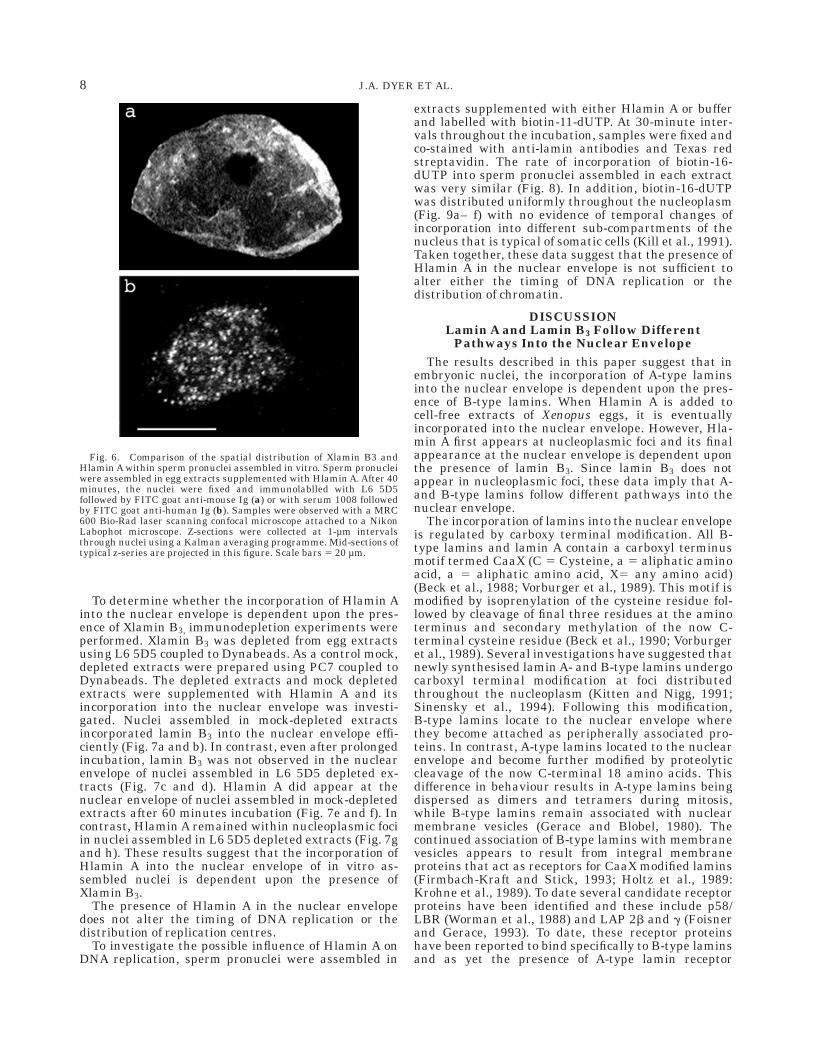
To investigate the possible influence of Hlamin A onDNA replication, sperm pronuclei were assembled in
extracts supplemented with either Hlamin A or bufferand labelled with biotin-11-dUTP. At 30-minute inter-vals throughout the incubation, samples were fixed andco-stained with anti-lamin antibodies and Texas redstreptavidin. The rate of incorporation of biotin-16-dUTP into sperm pronuclei assembled in each extractwas very similar (Fig. 8). In addition, biotin-16-dUTPwas distributed uniformly throughout the nucleoplasm(Fig. 9a– f) with no evidence of temporal changes ofincorporation into different sub-compartments of thenucleus that is typical of somatic cells (Kill et al., 1991).Taken together, these data suggest that the presence ofHlamin A in the nuclear envelope is not sufficient toalter either the timing of DNA replication or thedistribution of chromatin.
DISCUSSIONLamin A and Lamin B3 Follow Different
Pathways Into the Nuclear EnvelopeThe results described in this paper suggest that in
embryonic nuclei, the incorporation of A-type laminsinto the nuclear envelope is dependent upon the pres-ence of B-type lamins. When Hlamin A is added tocell-free extracts of Xenopus eggs, it is eventuallyincorporated into the nuclear envelope. However, Hla-min A first appears at nucleoplasmic foci and its finalappearance at the nuclear envelope is dependent uponthe presence of lamin B3. Since lamin B3 does notappear in nucleoplasmic foci, these data imply that A-and B-type lamins follow different pathways into thenuclear envelope.
The incorporation of lamins into the nuclear envelopeis regulated by carboxy terminal modification. All B-type lamins and lamin A contain a carboxyl terminusmotif termed CaaX (C 5 Cysteine, a 5 aliphatic aminoacid, a 5 aliphatic amino acid, X5 any amino acid)(Beck et al., 1988; Vorburger et al., 1989). This motif ismodified by isoprenylation of the cysteine residue fol-lowed by cleavage of final three residues at the aminoterminus and secondary methylation of the now C-terminal cysteine residue (Beck et al., 1990; Vorburgeret al., 1989). Several investigations have suggested thatnewly synthesised lamin A- and B-type lamins undergocarboxyl terminal modification at foci distributedthroughout the nucleoplasm (Kitten and Nigg, 1991;Sinensky et al., 1994). Following this modification,B-type lamins locate to the nuclear envelope wherethey become attached as peripherally associated pro-teins. In contrast, A-type lamins located to the nuclearenvelope and become further modified by proteolyticcleavage of the now C-terminal 18 amino acids. Thisdifference in behaviour results in A-type lamins beingdispersed as dimers and tetramers during mitosis,while B-type lamins remain associated with nuclearmembrane vesicles (Gerace and Blobel, 1980). Thecontinued association of B-type lamins with membranevesicles appears to result from integral membraneproteins that act as receptors for CaaX modified lamins(Firmbach-Kraft and Stick, 1993; Holtz et al., 1989:Krohne et al., 1989). To date several candidate receptorproteins have been identified and these include p58/LBR (Worman et al., 1988) and LAP 2b and g (Foisnerand Gerace, 1993). To date, these receptor proteinshave been reported to bind specifically to B-type laminsand as yet the presence of A-type lamin receptor
Fig. 6. Comparison of the spatial distribution of Xlamin B3 andHlamin A within sperm pronuclei assembled in vitro. Sperm pronucleiwere assembled in egg extracts supplemented with Hlamin A. After 40minutes, the nuclei were fixed and immunolablled with L6 5D5followed by FITC goat anti-mouse Ig (a) or with serum 1008 followedby FITC goat anti-human Ig (b). Samples were observed with a MRC600 Bio-Rad laser scanning confocal microscope attached to a NikonLabophot microscope. Z-sections were collected at 1-µm intervalsthrough nuclei using a Kalman averaging programme. Mid-sections oftypical z-series are projected in this figure. Scale bars 5 20 µm.
8 J.A. DYER ET AL.

proteins at the nuclear envelope have not been re-ported. Moreover, several reports have suggested thatthe pathway of A-type lamin incorporation into thenuclear envelope differs from that of B-type lamins(Bridger et al., 1993; Goldman et al., 1992). Here wepresent evidence that the incorporation of A-type lam-ins into the nuclear envelope is dependent upon theprior assembly of B-type lamins into a lamina. Thisdata implies that the association of A-type lamins with
the nuclear envelope is indirect and results from theincorporation of A-type lamins into pre-existing orforming filaments. The data, therefore, suggest thatlamina filaments consist of heterotypic assemblies ofA- and B-type lamin dimers and that the nucleation ofthese filaments depends upon B-type lamins. We specu-late that this hierarchy exists because B-type laminsare first concentrated at the nuclear envelope by virtueof their association with integral membrane proteins
Fig. 7. The incorporation of Hlamin A into the nuclear envelope isdependent upon the presence of Xlamin B3. Xenopus egg extracts weredepleted of Xlamin B3 using L6 5D5 linked to paramagnetic immun-obeads (c,d,g,h) or mock depleted with PC7 antibodies, also linked toparagmanetic immunobeads (a,b,e,f). Sperm pronuclei were as-sembled in the extracts for periods of up to 90 minutes. The nuclei
were fixed and stained with L6 5D5 to detect lamin B3 (a– d) or withserum 1008 to detect Hlamin A (e– h). The distribution of DNA in eachsample was detected with DAPI (a, c, e, and g). The intensely stainedround particles (diameter 5 4 µm) observed in some micrographs arethe immunobeads. Scale bars 5 10 µm.
9LAMIN A ASSEMBLY AND FUNCTION

that act as specific receptors as well as nucleatingpoints for filament assembly (Yang et al., 1997) whereasA-type lamins have no specific receptors but rapidlyassociate with B-type lamin filaments once they start toassemble.
Lamin A Is Not Sufficient to InfluenceChromatin Organisation
A-type lamin expression has been linked to differen-tiation both in vivo and in vitro (Benevente et al., 1985;Guilly et al., 1990; Lourim and Lim, 1990; Rober et al.,1989; Wolin et al., 1987). It has been proposed that onefunction of A-type lamins is to influence gene expres-sion by altering chromatin organisation (Nigg, 1989).Indeed, changes in the distribution of chromatin dooccur during embryogenesis at the same time as changesin lamin expression (Leibovici et al., 1992; Stick andHausen, 1985). Temporal changes in the distribution ofreplication domains is considered a reliable marker ofdiffering organisations of chromatin within the nucleus(Ferreira et al., 1997; Kill et al., 1991; Nakamura et al.,1986). In early embryos that contain no heterochroma-tin, only a single pattern of DNA replication is observed(e.g., Hutchison, 1995). Later during embryogenesis,different patterns of DNA replication are observed andthe timing at which these new patterns of DNA replica-
tion emerge corresponds to times at which recognisabledifferences in the organisation of chromatin occurs(Leibovici et al., 1992). If altered chromatin organisa-tions are a function of lamin expression, one mightexpect that ectopic expression of A-type lamins inembryonic systems would generate distinct chromatindomains that are recognised by different patterns ofDNA replication. The accumulation of Hlamin A in thenuclear lamina did not influence either the temporalappearance or spatial distribution of replication do-mains in in vitro assembled nuclei. Therefore, lamin Aalone is not sufficient to reorganise chromatin. It ispossible that chromatin organisation is a function oflamin C rather than lamin A. Alternatively, alteredchromatin organisation may result from interactionsbetween lamin A and chromatin associated proteins.Chromatin-associated proteins have been reported tobind to lamin A, including the negative growth regula-tor p110RB (Mancini et al., 1994; Ozaki et al., 1994).Perhaps the appearance of different chromatin do-mains during embryogenesis is a function of alteredlamin expression coupled with the expression of chroma-tin binding proteins. The assay system that we haveestablished would allow us to test this hypothesis byexogenously adding combinations of lamins and chroma-tin-associated proteins.
Fig. 8. The presence of Hlamin A in the nucleus does not affect thetiming of DNA replication. Sperm pronuclei assembled in the presence(squares) or absence (circles) of Hlamin A were pulse labelled withbiotin-16-dUTP. Thirty-minute pulses were applied over a period of120 minutes. Following each pulse label, nuclei were fixed and stained
with Texas Red streptavidin. Two hundred nuclei were scored at eachtime point and for each sample on each duplicate slides and thenumber of biotin positive nuclei (replicating) were expressed as apercentage. The results shown in this example were typical of fivereplicate experiments.
10 J.A. DYER ET AL.

ACKNOWLEDGMENTSThe authors thank Professor Mike Kerr (Ninewells
Teaching Hospital, Dundee) for the supply of humanautoimmune sera and Dr. Murray Stewart (MRC Labo-ratory of Molecular Biology, Cambridge) for the supplyof purified recombinant Hlamins A, C, and B1. Thiswork was supported by the award of a Wellcome TrustPrize Studentship to J.A.D. and grants from the CancerResearch Campaign to E.B.L. and C.J.H.
REFERENCESAebi U, Cohn J, Buhle L Gerace L. 1986. The nuclear lamina is a
meshwork of intermediate type filaments. Nature 323:560– 564.Beck L, Hosick T Sinensky M. 1988. Incorporation of a product of
mevalonic acid metabolism into protein of chinese hamster ovarycell nuclei. J Cell Biol 107:1307– 1316.
Beck L, Hosick T, Sinensky M. 1990. Isoprenylation is required for theprocessing of the lamin A precursor. J Cell Biol 110:1489–1499.
Benevente R Krohne G. 1986. Involvement of nuclear lamins inpostmitotic reorganisation of chromatin as demonstrated by micro-injection of lamin antibodies. J Cell Biol103:1847–1854.
Benevente R, Krohne G Franke WW. 1985. Cell type specific expres-sion of nuclear lamina proteins during development of Xenopuslaevis. Cell 41:177–190.
Blow JJ, Laskey RA. 1986. Initiation of DNA replication in nuclei andpurified DNA by a cell-free extract of Xenopus eggs. Cell 47:577–587.
Bridger JM, Kill IR, O’Farrell M Hutchison CJ. 1993. Internal lamistructures within G1 nuclei of human dermal fibroblasts. J Cell Sci104:297– 306.
Broers JL.V Raymond Y, Klein-Rot M, Kuijpers H, Wagenaar S,Ramaekers FCS. 1993. Nuclear A-type lamins are differentiallyexpressed in human lung cancer sub-types. Am J Pathol 143:211–219.
Burke B, Gerace L. 1986. A cell-free system to study reassembly of thenuclear envelope at the end of mitosis. Cell 44:639– 652.
Dyer J.A, Kill IR, Pugh G, Quinlan RA, Lane EB, Hutchison CJ. 1997.Cell cycle changes in A-type lamin associations detected in humandermal fibroblasts using monoclonal antibodies. Chrom Res 5:383–394.
Ellis DJ, Jenkins HE, Whitfield WGF, Hutchison CJ. 1997. GTS-laminfusion proteins act as dominant negative mutants in Xenopus egg
extract and reveal the function of the lamina in DNA replication. JCell Sci 110:2507– 2518.
Farnsworth C, Wolda S, Gelb M, Glomset J. 1989. Human lamin Bcontains farnesylated cystein residue. J Biol Chem 264:20422–20429.
Ferreira J, Paolella G, Ramos C, Lamond AI. 1997. Spatial organiza-tion of large-scale chromatin domains in the nucleus: a magnifiedview of single chromosome territories. J Cell Biol 139:1597– 1566.
Firmbach-Kraft I, Stick R. 1993. The role of CaaX-dependant modifica-tions in membrane association of Xenopus nuclear lamin B3 duringmeiosis and the fate of B3 in transfected mitotic cells. J Cell Biol123:1661– 1670.
Fisher D, Chaudhary N, Blobel G. 1986. cDNA sequencing of nuclearlamin A and C reveals primary and secondary structure homology tointermediat filament proteins. Proc Natl Acad Sci USA 83:6450–6454..
Foisner R, Gerace L. 1993. Integral membrane proteins of the nuclearenvelope interact with lamins and chromosomes, and binding ismodulated by mitotic phosphorylation. Cell 73:1267– 1279.
Gerace L, Blobel G. 1980. The nuclear envelope lamina is reversiblypolymerised during mitosis. Cell 19:277– 287.
Goldberg M, Allen T. 1993. The nuclear pore complex: three dimen-sional surface structure revealed by field emission in-lens scanningelectron microscopy, with underlying structure uncovered by prote-olysis. J Cell Sci 106:261– 274.
Goldberg M, Jenkins HE, Allen TD, Whitfield WGF, Hutchison CJ.1995. Xenopus lamin B3 has a direct role in the assembly of areplication competent nucleus: evidence from cell-free egg extracts.J Cell Sci 108:3451– 3461.
Goldman AE, Moir RD, Montag-Lowry M, Stewart M, Goldman RD.1992. Pathway of incorporation of microinjected lamin A into thenuclear envelope. J Cell Biol 119:725– 735.
Guilly MN, Bensussan A, Bourge JF, Bornens M, Courvalin JC. 1987.A human T lymphoblastic cell line lacks lamins A and C. EMBO J6:3795– 3799.
Harlow L, Lane D. 1988. Antibodies: a laboratory manual. Cold SpringHarbor, NY: Cold Spring Harbor Laboratory Press.
Hoger TH, Krohne G, Franke WW. 1988. Amino acid sequence andmolecular characterisation of murin lamin B as deduced from cDNAclones. Eur J Cell Biol 47:283– 290.
Hoger TH, Zatloukal K, Waizenegger I, Krohne G. 1990. Characterisa-tion of a second highly conserved B-type lamin present in cellspreviously thought to contain only a single B-type lamin. Chromo-soma 99:379– 390.
Fig. 9. The presence of Hlamin A in the nuclear envelope does notaffect the spatial distribution of replication centres. Sperm pronucleiassembled in the presence (a–c) or absence (d–f) or Hlamin A werepulse labelled with biotin-16-dUTP. Thrity minutes after each pulsewas applied, nuclei were fixed, and stained with serum 1008 to detect
Hlamin A (c,f), Texas Red streptavidin to detect replication centres(b,e) and DAPI to detect the distribution of DNA (a,d). The micro-graphs show the typical examples of nuclei fixed at 60 minutes andviewed with a Zeiss Axioskop equipped for epifluorescence. Scalebars 5 15 µm.
11LAMIN A ASSEMBLY AND FUNCTION

Hoger TH, Krohne G, Kleinschmidt JA. 1991. Interaction of Xenopuslamins A and LII with chromatin in vitro mediated by a sequenceelement in the carboxyterminal domain. Exp Cell Res 197:280–289.
Holtz D, Tanaka RA, Hartwig J, McKeon F. 1989. The CaaX motif oflamin A functions in conjunction with the nuclear localisation signalto target assembly to the nuclear envelope. Cell 59:969–977.
Hutchison CJ. 1994. The use of cell free extracts of Xenopus eggs forstudying DNA replication in vitro. In: Fantes P, Brooks R, editors.The cell cycle: a practical approach. IRL Press at Oxford UniversityPress, p 177–196.
Hutchison CJ. 1995. Local and global changes in the distribution ofreplication centres in rapidly expanding nuclei. Chromosome Res3:16– 26.
Hutchison CJ, Cox R, Drepaul R-S, Gomperts M, Ford CC. 1987.Periodic DNA synthesis in cell-free extracts of Xenopus eggs. EMBOJ 6:2003– 2010.
Hutchison CJ, Bridger JM, Cox LS, Kill IR. 1994. Weaving a patternfrom disparate threads: lamin function in nuclear assembly andDNA replication. J Cell Sci 107:3259– 3269.
Jenkins H.E, Holleman T, Lyon C, Lane EB, Stick R, Hutchison CJ.1995. Nuclei which lack a lamina accumulate karyophilic proteinsand assemble a nuclear matrix. J Cell Sci 106:275– 285.
Kill IR, Bridger JM, Campbell KH.S, Moldonado-Codina G, HutchisonC. 1991. The timing of the formation and usage of replicationclusters in S-phase nuclei of human diploid fibroblasts. J Cell Sci100:869– 876.
Kill IR, Hutchinson CJ. 1995. S-phase phosphorylation of humanlamin B2. FEBS Lett. 377:26–30.
Kitten G, Nigg E. 1991. The CaaX motif is required for isoprenylation,carboxyl methylation and nuclear membrane association of laminB2. J Cell Biol 113:13–23.
Krohne G, Waizenegger I, Hoger TH. 1989. The conserved carboxyl-terminal cysteine of nuclear lamins is essential for lamin associationwith the nuclear envelop. J Cell Biol 109:2003– 2011.
Lebel S, Lampron C, Royal A, Raymond Y. 1987. Lamins A and Cappear during retinoic acid-induced differentiation of mouse emby-ronal carcinoma cells. J Cell Biol 105:1099– 1104..
Lehner CF, Stick R, Eppenberger HM. Nigg EA. 1987. Differentialexpression of nuclear lamin proteins during chicken development. JCell Biol 105:577– 587.
Leibovici M, Monod G, Geraudie J, Bravo R, Mechali M. 1992. Nucleardistribution of PCNA during embryonic development in Xenopuslaevis: a reinvestigation of early cell cycles. J Cell Sci 102:63–69.
Lourim D, Lim JJ-C. 1992. Expression of wild-type and nuclearlocalization deficient human lamin A in chick myogenic cells. J CellSci 103:863–874.
Lourim D, Krohne G. 1993. Membrane-associated lamins in Xenopusegg extracts: identification of two vesicle populations. J Cell Sci123:501– 512.
Moir RD, Donaldson AD, Stewart M. 1991. Expression in Eschericia
coli of human lamins A and C, influence of head and tail domains onassembly properties and paracrystal formation. J Cell Sci 99:363–372.
Mancini M, Shan B, Nickerson J, Penman S, Lee W-H. 1994. Theretinoblastoma gene product is a cell cycle dependent nuclearmatrix associated protein. Proc Natl Acad Sci USA 91:418– 422.
Nakamura H, Morita T, Sato C. 1986. Structural organisation ofreplicon domains during DNA synthetic phase in the mammaliannucleus. Exp Cell Res 165:291– 297.
Nigg EA. 1989. The nuclear envelope. Curr Opin Cell Biol 1:435– 440.Ozaki T, Saijo M, Murakimi K, Enomoto H, Taya Y, Sakiyama S. 1994.
Complex formation between lamin A and the retinoblastoma geneproduct: identification of the domain on lamin A required for itsinteraction. Oncogene 9:2649– 2653.
Paulin-Levasseur M, Scherbarth A, Giese G, Roser K, Bohn W, TraubP. 1989. Expression of vimentin and nuclear lamins during the invitro differentiation of human promyelocytic leukemia cells HL–60.Eur J Cell Biol 50:453– 461.
Quinlan R, Hutchison CJ, Lane EB. 1995. Intermediate filamentproteins: protein profiles. 2:801–952..
Rober R-A, Weber K, Osborn M. 1989. Differential timing of nuclearlamin A/C expression in the various organs of the mouse embyro andthe young animal: a developmental study. Development 105:365–378.
Rober R-A, Sauter H, Weber K, Osborn M. 1990. Cells of the cellularimmune and hemopoietic system of the mouse lack lamins A/C:distinction versus other somatic cells. J Cell Sci 95:587– 598.
Sinesky M, Fantle K, Trujillo M, McLain T, Kupfer A, Dalton M. 1994.The processing pathway of prelamin A. J Cell Sci 107:61– 67.
Stick R. 1988. cDNA cloning of the developmentally regulated laminLiii of Xenopus laevis. EMBO J 7:3189– 3197.
Stick R, Hausen P. 1985. Changes in the nuclear lamina compositionduring early development of Xenopus laevis. Cell 41:191– 200.
Vorburger K, Kitten GT, Nigg EA. 1989. Modification of nuclear laminproteins by a mevalonic acid derivative occurs in reticulocyte lysatesand requires the cysteine resiude of the C-terminal CxxM motif.EMBO J 8:4007– 4013.
Waseem N, Lane DP. 1990. Monoclonal antibody analysis of theproliferating cell nuclear antigen (PCNA). J Cell Sci 96:121– 129.
Wolin S, Krohne G, Kirschner M. 1987. A new lamin in Xenopussomatic tissue displays strong homology to human lamin A. EMBO J6:3809– 3818.
Worman HJ, Yuan J, Blobel G, Georgatos SD. 1988. A lamin B receptorin the nuclear envelope. Proc Natl Acad Sci USA 85:8531– 8534.
Yang L, Guan T, Gerace L. 1997. Lamin-binding fragment of LAP2inhibits increase in nuclear volume during the cell cycle andprogression into S-phase. J Cell Biol 139:1077– 1087.
Zhang C, Jenkins HE, Goldberg M, Allen TD, Hutchison CJ. 1996.Nuclear lamina and nuclear matrix organisation in sperm pronucleiassembled in Xenopus egg extract. J Cell Sci 109: 2275– 2286.
12 J.A. DYER ET AL.





























