Embed Size (px)
Citation preview

Proceedings of the Fourth International Workshop on Lobster Biology and Management, 1993
THE INFLUENCE OF TEMPERATURE, SALINITY, AND
POSTLARVAI, TRANSPORT ON THE DISTRIBUTION OF JUVENILE SPINY LOBSTERS, PANULIRUS ARGUS (LATREILLE, 1804), IN
FLORIDA BAY
BY
JENNIFER M. FIELD 1) and MARK J. BUTLER, IV2)
Department of Biological Sciences, Old Dominion University, Norfolk, VA 23529-0266, U.S.A.
ABSTRACT
Florida Bay is the major nursery for the Caribbean spiny lobster, Panulirus argus, population in south Florida. This region is characterized by a series of shallow hardbottom or seagrass-covered basins separated by carbonate mudbanks that serve as barriers to water circulation and presuma- bly to transport of planktonic larvae. Temperatures fluctuate dramatically in the bay and salinities there range from 35 ppt-50 ppt. In this study, we investigated the physiological tolerance of P. argus postlarvae to various combinations of temperature and salinity representative of the condi- tions found in Florida Bay and also determined the extent of postlarval recruitment into the interior of the bay.
We measured postlarval settlement, juvenile abundances, and postsettlement habitat availabil- ity monthly (March 1992-July 1992) along 5 transects extending north from the main Florida Keys into the interior of Florida Bay. Concurrently, P. argus postlarvae were reared in the laboratory, in a completely crossed design, at four temperatures (18°C, 22°C, 29°C and 33°C) and four salinities (25, 35, 45 and 50 ppt). Survival, time:-to-metamorphosis, and growth to the first juvenile stage were measured.
Few postlarvae settled at sites beyond the emergent banks ringing Florida Bay, and lobsters were only found in one basin (Twin Keys Basin) where habitat and environmental conditions were favorable. Laboratory results indicate that at high (33°C) and low temperatures (18°C) survival at salinities other than 35 ppt is greatly reduced. Our results indicate that postlarvae are not regularly transported into the interior of Florida Bay and that the lack of suitable nursery habitat and high salinity further limit recruitment, especially at extreme temperatures. Given the present conditions in Florida Bay, recruitment of P. argus is restricted to the southernmost reaches of Florida Bay nearest the Florida Keys.
RÉSUMÉ
La baie de Floride est la principale nursery pour la population caraïbe de la langouste Panulirus argus en Floride du Sud. Cette région est caractérisée par une série de bassins peu profonds à fond dur ou couvert d'algues, séparés par des bancs de vase carbonatée qui servent de barrière à la circulation de l'eau et, suppose-t-on, au transport des larves planctoniques. Les températures
1) Currcnt address: Oak Ridge National Laboratories, Environmental Sciences Division, P.O. Box 2008, Oak Ridge, TN 37831-6037, U.S.A. 2) Corresponding author.

27
fluctuent de façon spectaculaire dans la baie et les salinités vont de 35 ppt à 50 ppt. Au cours de cette étude, nous avons testé la tolérance physiologique des postlarves de P. argus à différentes combinaisons de température et salinité, représentatives des conditions observées dans la baie de Floride, et aussi déterminé l'importance du recrutement postlarvaire à l'intérieur de la baie.
Nous avons mesuré chaque mois (mars 1992-juillet 1992) la sédentarisation postlarvaire, les abondances de juvéniles et la disponibilité des habitats de post-sédentarisation le long de 5 transects s'étendant du nord des principales Florida Keys jusqu'à l'intérieur de la baie de Floride. Simultanément, des postlarves de P. argus étaient élevées au laboratoire, sous les combinaisons complètes de 4 températures (18°C, 22°C, 29°C et 33°C) et de 4 salinités (25, 35, 45 et 50 ppt). La survie, le temps jusqu'à la métamorphose et la croissance jusqu'au premier stade juvénile ont été mesurés.
Peu de postlarves se sont sédentarisées à des endroits situés au-delà des bancs émergés entourant la Baie de Floride, et les langoustes ont été trouvées dans un seul bassin (Twin Keys Basin) où l'habitat et les conditions d'environnement étaient favorables. Les résultats en laboratoire indi- quent que, à haute (33°C) et basse (18°C) températures, la survie à des salinités autres que 35 ppt était fortement réduite. Nos résultats indiquent que les postlarves ne sont pas régulièrement transportées vers l'intérieur de la baie de Floride et que le manque d'un habitat de nursery approprié ainsi qu'une salinité élevée limitent davantage le recrutement, en particulier aux températures extrêmes. Etant donné les conditions actuelles dans la baie de Floride, le recrute- ment de P. argus est restreint aux approches les plus méridionales de la baie de Floride, au plus près des Florida keys.
INTRODUCTION
The Caribbean spiny lobster, Panulirus argus (Latreille, 1804), has a complex life cycle, which in south Florida is intricately tied to the Florida Bay nursery. Postlarvae arrive along the coast of south Florida each month during the new
moon and swim (Calinski & Lyons, 1983) or are transported at night on rising tides through the channels between the Florida Keys and into Florida Bay
(Sweat, 1968; Witham et al., 1968; Little, 1977; Little & Milano, 1980; Marx,
1986). In the bay, postlarvae settle primarily into clumps of the red macroalgae Laurencia spp. (Marx & Herrnkind, 1985; Marx, 1986; Herrnkind & Butler, 1986; Butler & Herrnkind, 1991) where they metamorphose into the benthic
juvenile stage. Juvenile lobsters are solitary and remain within the algae until
reaching 20 mm carapace length (CL) when they become gregarious and reside
under sponges, coral heads, solution holes, seagrass undercuts and other struc-
tures (hereafter referred to as "post-algal" stage juveniles; Herrnkind et al., in
press). After 2-3 years, the lobsters migrate, as adults, to the reef (Davis 1978; Davis & Dodrill, 1980, 1989; Kanciruk, 1980).
Processes limiting recruitment of P. argu.s in Florida Bay may operate at both
local and regional scales. Locally, the availability of postlarvae arriving at an
area establish a minimum level of recruitment (Herrnkind & Butler, in press), while the availability of appropriate macroalgae settlement substrate and post- settlement shelter (e.g., sponges, octocorals, etc.) may limit post-settlement survival of juveniles (Marx & Herrnkind, 1985; Herrnkind & Butler, 1986; Smith & Herrnkind, 1992). On a large scale, the physical structure and condi-
tion of Florida Bay may impose regional limits to recruitment. Florida Bay is
characterized by a series of shallow (<3 m) seagrass-covered or hardbottom

28
(carbonate bedrock with patches of sponges, octocorals and small coral heads) basins separated by seagrass-covered carbonate mudbanks <1 1 m in depth
(Zieman et al., 1989). These mudbanks restrict water circulation in the bay and
Holmquist et al. ( 1989a) have suggested that transport of planktonic larvae into
these basins may be inhibited. Most basins are hypersaline (35-50 ppt) due to
reduced freshwater flow from the Everglades and high rates of evaporation
(Enos & Perkins, 1979; Fourqurean et al., 1992; Walters et al., 1992), and
seasonal fluctuations in temperature can be dramatic (Holmquist et al., 1989b). Therefore, physiological tolerance to temperature and salinity may influence
survival of postlarvae gaining access to these basins. Recently, hypersaline conditions within Florida Bay have sparked a cascade of disturbances, includ-
ing the loss of seagrass meadows (Robblee et al., 1992) and blooms of cya- nobacteria that led to the mass destruction of sponges in the bay (Butler et al.,
unpubl. ms.). Thus, the declining availability of sponges, which serve as shelter
for post-algal stage juvenile lobsters, may also create a recruitment bottleneck
on a regional scale (Wahle & Steneck, 1991; Butler & Herrnkind, in press). There appears to be a northern boundary within Florida Bay beyond which
P. argus is not found (Davis & Dodrill, 1989). Is this boundary a result of
physical barriers to transport, limited settlement or post-settlement habitat, or
constraints set by the physiological tolerance of postlarvae to the temperature and salinity conditions present in the region? Only onc study of postlarval
transport and recruitment has been conducted in Florida Bay (Butler & Herrn-
kind, 1992; Herrnkind & Butler, in press). That study did not, however, include
the interior regions of the bay beyond the fringing mudbanks where the spiny lobster distributional boundary lies. Moreover, there is no detailed information
on the distribution and structure of the hardbottom habitats in these areas, and
thus their suitability as lobster nurseries. Similarly, the physiological tolerances
of P. argus postlarvae and juveniles to the temperatures and salinities typical of
Florida Bay are not known. In this study, we evaluated three mechanisms that
might determine the northern limit to the distribution of P. argus in Florida Bay;
(i) postlarval transport, (ii) settlement and postsettlement habitat suitability, and (iii) physiological tolerance of postlarvae to spccific temperature and
salinity regimes.
MATERIALS AND METHODS
Laboratory methods
We examined the effects of temperature and salinity on Panulirus argus post- larvae in a laboratory study conducted from February 1992 to August 1992 at
the Keys Marine Laboratory, Long Key, Florida. Postlarvae were collected at
night on rising new moon tides using stationary plankton nets (1 1 m2 diameter,

29
750 pm mesh) deployed from the Tom's Harbor Channel Bridge, Florida, and
were retained overnight in the laboratory in water of the same temperature and
salinity from which they were collected. The following morning, postlarvae were placed into 400 ml experimental beakers ( postlarva/beaker). Postlarvae
were randomly assigned to treatments and were gradually exposed over 8-12 hrs to the treatment conditions by slowly changing the salinity and temperature within each experimental beaker. Postlarvae were exposed, in a completely crossed design, to four temperatures (18° and 22°C representative of winter
temperatures; and 29° and 33°C representative of summer temperatures) and
four salinities (25, 35, 45, 50 ppt). Postlarvae were exposed to the 18° and 22°C treatments during winter and early spring, and the 29" and 33"C treatments
during summer. These treatments span a range of environmental conditions
present within Florida Bay (Fourquerean et al., 1992). The laboratory experiments were conducted in water tables, each with a
separate sea water system that recirculated sea water between a 40 1 reservoir
(equipped with a carbon and subgravel filter) and up to 20 experimental beakers at a rate of 0.3 1/hr. A 2 cm2 piece of air conditioning filter (which mimics macroalgal structure) was placed in each beaker as a shelter. The
number of replicates per treatment varied from 13 to 52, depending on the
number of postlarvae captured from the plankton. Experimental temperatures were maintained using heaters or by chilling the laboratory room. Salinities
were maintained by adding sea salt (Instant Ocean Brand) or freshwater to
natural seawater. Temperature and salinity were monitored twice daily; tem-
peratures were maintained to within ± 2°C, and salinities were maintained to
within ± 1 ppt. Dissolved oxygen, pH, nitrate nitrogen and ammonia were
monitored weekly. We observed postlarvae twice daily and recorded survival,
time-to-metamorphosis (TTM) to the first benthic juvenile stage, and juvenile
length following metamorphosis (mm carapace length; CL). Survival of postlarvae was analyzed using a three-way log-linear categorical
analysis to test for an interaction between temperature, salinity, and postlarval survival. If the three-way interaction was not significant, we then used two-way
contigency tables analyses to examine the relationship between various tem-
perature or salinity treatments and postlarval survival. The effect of tempera- ture and salinity on postlarval time-to-metamorphosis was analyzed using a
two-factor model I ANOVA on In transformed data. Similarly, we used a two-
factor model I ANOVA on square root transformed data to test the effect of
temperature and salinity on the carapace length of first stagc juveniles at
metamorphosis. The Ryan-Einot-Gabriel-Welsch Multiple F test was employed to determine the significance of the levels of temperature and salinity for both
analyses. The alpha level for significance was set at 0.025 to insure an experi- mentwise error rate of 0.05 for the two separate ANOVAs.


31
at a particular site for a few months may not accurately reflect the annual
potential for natural settlement in the area. Therefore, we censused the post-
algal stage juvenile lobster population near the benthic collector sites as an
independent estimate of recent recruitment. Lobsters < 35 mm CL are not
nomadic (Kanciruk, 1980; Herrnkind & Butler, 1986) and arc likely to have
settled in the vicinity near their capture within the last few months (Yoshimura & Yamakawa, 1988). Post-algal stage juveniles are also diurnal crevice dwellers
and can be visually censused, whereas algal-dwelling juveniles cannot. Surveys of post-algal stage juveniles are, thus, useful measures of recent recruitment
because the number of juveniles ? 35 mm CL at a site represents an integration of settlement and post-settlement survival over the previous 3-5 months (For- cucci et al., in press). Surveys were conducted by two divers who searched and
collected lobsters from natural shelters (e.g., sponges, octocorals, solution
holes, etc.) in an area of hardbottom for 30 minutes (1 diver hour/site), yielding a catch-per-unit-effort index of post-algal juvenile abundance. All lobsters
found were measured (nearest 0.1 mm) and released. Diver surveys were not
conducted in Johnson Key Basin (site 21), Ninemile Bank (site 22), Panhandle
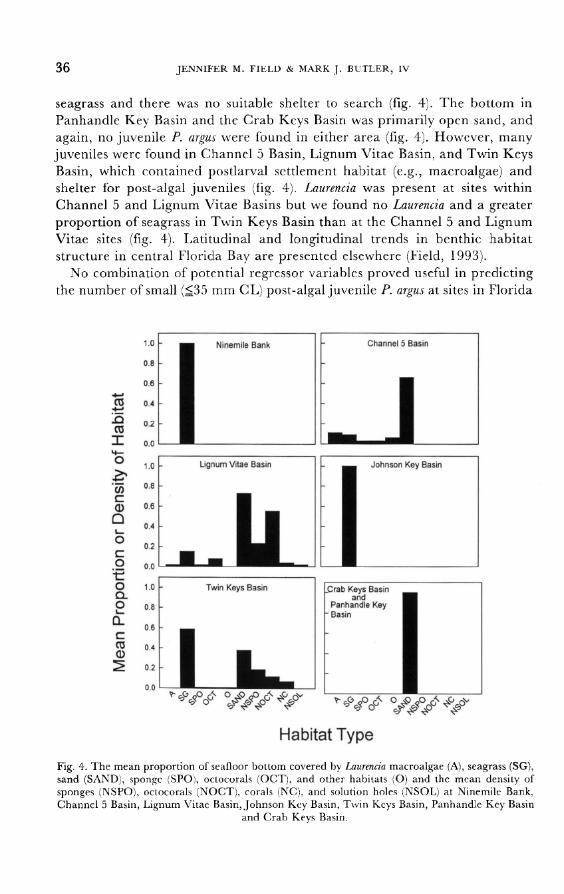
Key Basin (site 23) or the Crab Keys Basin (site 24) due to the lack of suitable hardbottom habitat or juvenile shelter in those areas.
At the same sites where juvenile lobster surveys were conducted, we also
quantified habitat structure. Four haphazardly-positioned 25 m line-transects
were established at each site and the proportion of bottom covered by macro-
algae (primarly Laurencia spp.), seagrass (Thalassia testudinum Koenig), sponge, octocoral, and other potential post-algal stage lobster shelters (e.g., coral heads
and solution holes) recorded. At a subset of sites (10 total), four 25 m X 2 m belt-
transects were established over the original line transects to estimate the density of sponges, octocorals, coral heads, solution holes and other potential lobster shelters. The shortest possible distance (km) to each site from the inlets between
the main islands of the Florida Keys (i.e., the source of postlarvae for the bay), and the number of mudbanks that postlarvae would pass over enroute to a
specific site were also determined, as was the temperature and salinity at each
site.
We used a multiple regression analysis (best-fit approach) to determine
whether the number of settling postlarvae, recently-recruited juveniles 35
mm CL) or larger resident juveniles (>35 mm CL) could be predicted using the
following factors as potential regressor variables: distance to channel, number of banks crossed, salinity, number of postlarvae/benthic collector/month, the
proportion of bottom covered by Laurencia, seagrass, sponge, octocoral or other
habitats, and the density of sponges, octocorals, corals, and solution holes.
Because only a subset of the sites included shelter densities estimated from belt-
transects, as opposed to the line-transect data (proportion of bottom cover) collected at all sites, separate sets of multiple regression analyses were per- formed on these two data sets. An important assumption of multiple regression

32
analysis is that the regressor variables not be correlated. We used Spearmans rank correlation analysis to test for significant correlations among the non-
normally distributed regressor variables collected from the line-transect sam-
pling method, and Pearsons correlation analysis to test for correlations among the normally distributed regressor variables collected using the belt-transect
sampling method. Several regressor variables collected from the line-transect
sampling method could not be transformed to meet the assumption of nor-
mality for multiple regression, therefore, these data were ranked prior to
analysis (Iman & Conover, 1979; Conover & Iman, 1981). Separate analyses were run on the line- or belt-transect habitat data to predict: (i) number of
juvenile lobsters 35 mm CL/site, (ii) number of juvenile lobsters >35 mm CL/
site, and (iii) number of settling postlarvae/site. The dependent variables, the
habitat sampling method employed, and the uncorrelated regressor variables
used for each analysis are listed in table I.
TABLE I
Summary of the multiple regression analyses listing the predicted variables, the
types of habitat data (i.e., data obtained from line- or belt-transects), and the
uncorrelated regressor variables used in each analysis
Predicted variable Habitat data used Uncorrelated regressor variables
Number of juvenile Line-transect Number of mudbanks Panulirus argus <_35 mm CI. Proportion of algae (square root transformed) Proportion of spongcs
Proportion of octocorals
Belt-transect Density of sponges Density of octocorals (In transformed) Density of corals
Number of juvenile Line-transect Proportion of algae Panulirus argus >35 mm CL Proportion of octocorals (square root transformed) Proportion of other shelters
Proportion of sand
Belt-transect Salinity Density of octocorals (In transformed)
. Density of corals Density of solution holcs
Number of postlarvae Line-transect Number of mudbanks settling (In transformed) '
Salinity Proportion of sand
Belt-transect Salinity Density of spongcs Density of octocorals (In transformed) Density of solution holes

33
RESULTS
Laboratory study
The three-way interaction between temperature, salinity, and survival of P.
argus postlarvae to the first benthic juvenile stage approached significance (x2 =
15.57; df = 9; p - 0.08). However, the two-way interactions between the
various levels of temperature or salinity and postlarval survival were all highly
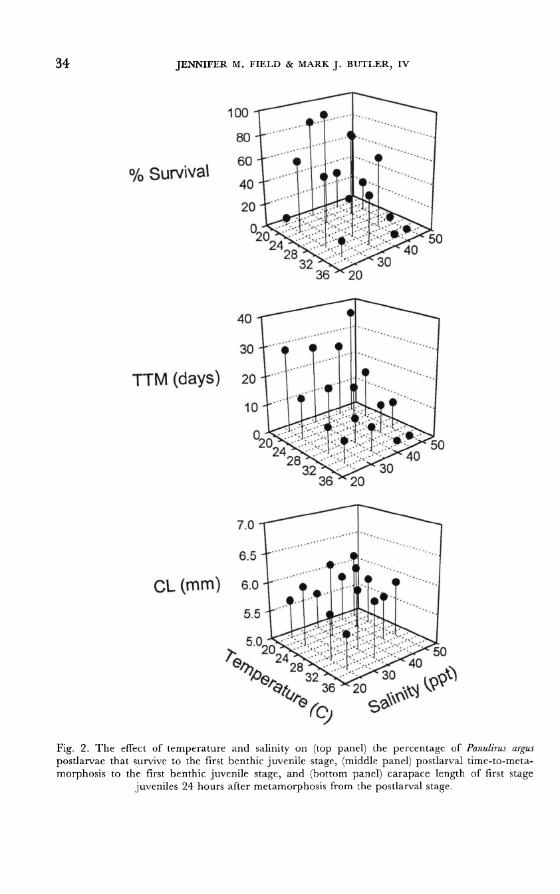
significant in all cases). Survival of postlarvae was highest at 22°C
(55%) and 35 ppt (76%; fig. 2). At temperatures above and below 22°C and
salinities other than 35 ppt, survival of postlarvae declined. When postlarvae were reared at the highest temperature and salinities (i.e., 33°C and 45 and 50
ppt), none survived and < 10% survived extreme salinities (i.e., 25 and 50 ppt) when reared at 18°C. Temperature and salinity interact to influence TTM of
postlarvae (F = 3.15; df = 7; p = 0.004). Postlarval TTM increased with
decreasing temperatures and was greater for postlarvae reared at salinities >45
ppt (fig. 2). However, TTM of postlarvae reared in 35 and 45 ppt was not
significantly different. The interaction of temperature and salinity also signifi-
cantly influenced the size of first stage juveniles at metamorphosis (F = 3.67; df = 7; p = 0.001). Size at metamorphosis was greatest for postlarvae raised at
salinities of 35 ppt and temperatures of 22°C, but the maximum difference
among treatment means (0.31 mm CL) was within measurement error and is
probably not of ecological significance.
Field study
Postlarvae settled in benthic collectors in areas south of the first set of
mudbanks, including: Channel 5 Basin (sites 1-7), several sites within the
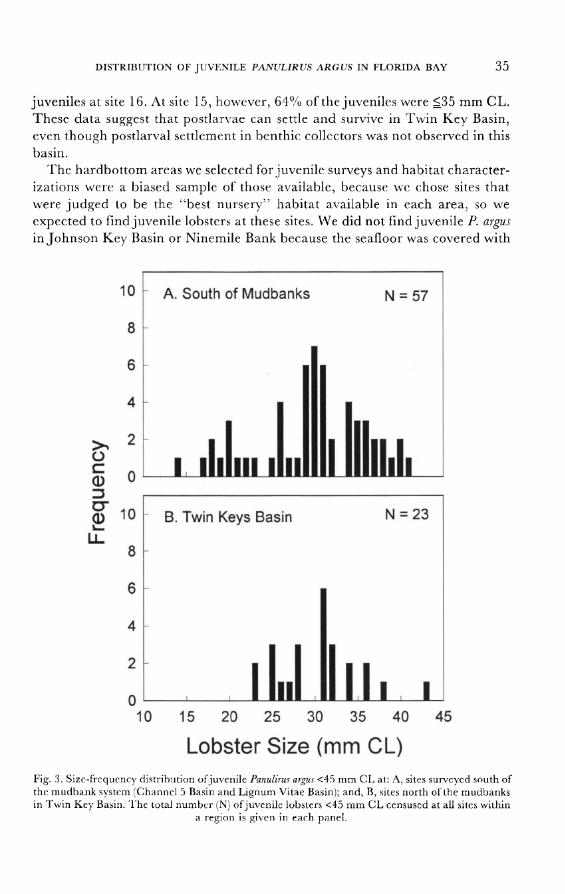
Lignum Vitae Basin (sites 10, 17 and 18), and at Ninemile Bank (site 22). Most of these sites (sites 2, 4-10, 13, 14 and 18) also contained juvenile P. argus <35
mm CL (size range 14-<45 mm CL; fig. 3a) indicating that postlarval settle-
ment and recruitment to the postalgal juvenile stage occurred in regions south of the banks. No postlarvae were found within benthic collectors in Johnson
Key Basin (site 21), the Twin Keys Basin (sites 11, 12 and 19), in Panhandle
Key Basin (site 23) or the Crab Keys Basin (site 24) indicating that P. argus
postlarvae are not typically transported across mudbanks and into the interior
basins of Florida Bay. Similarly, no juvenile lobsters <_35 mm CL were found in
Johnson Key Basin, Panhandle Key Basin or the Crab Keys Basin. In Twin Key Basin, however, juveniles were collected from hardbottom habitat in the cen-
tral and western sections of the basin (fig. 3b). At the Twin Key Basin site
nearest Twin Key Bank (site 11), 22 juvenile lobsters (size range: 28.7->60 mm
CL) were collected; 41 % of which were <_35 mm CL. At sites 15 and 16, 5 km
and 6.7 km northeast of the bank (respectively), the abundance of juvenile lobsters decreased; 14 juveniles (size range 28->60 mm CL) at site 15, and 2

37
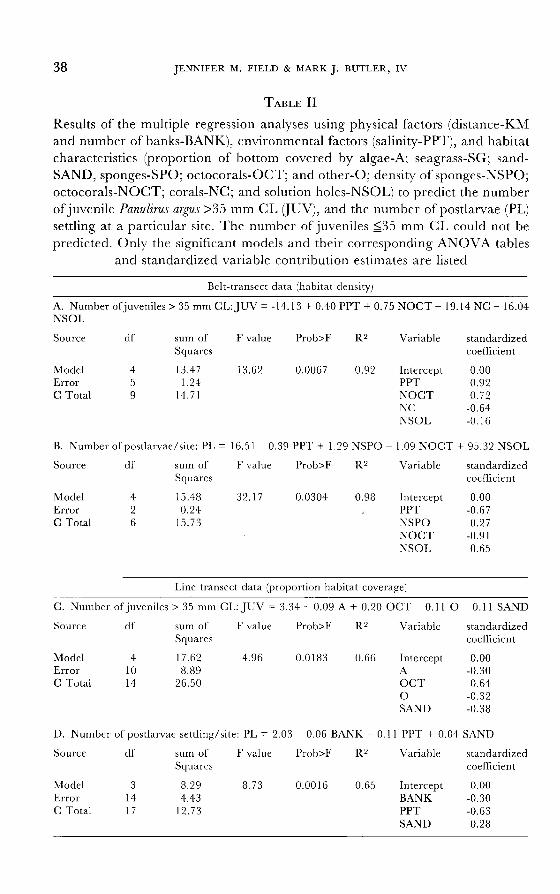
Bay. However, the number of large (>35 mm CL) juveniles/site and the
number of postlarvae settling/site could be predicted using regressor variables
collected from both line-transect and belt-transect sampling methods (table II). The habitat density data from the belt transects yielded the best models, with
multiple R2 values >0.90, as compared to R2 values of approximately 0.65 for
models incorporating proportional bottom coverage data. However, the den-
sity-based models included fewer sites and, therefore, contain fewer degrees of
freedom than the models based on bottom coverage. Examination of the standardized coefficients for the density-based habitat
data indicates that salinity, and the density of octocorals and hard corals are,
respectively, the most influential variables in the model predicting the number
of juveniles >35 mm CL (R`' = 0.92; table II). The number of postlarvae settling at a site was best predicted by the density of octocorals, salinity, and the density of solution holes at a site, respectively, (R2 = 0.98; table II). For the models
incorporating bottom coverage of various habitats (i.e., line-transect data), the
number of juvenile P. argus >35 mm CL was most influenced by the proportion of bottom covered by octocorals (R2 = 0.66; table II). The number ofpostlarvae settling at a site was negatively related to salinity and the minimum number of
banks over which postlarvae pass enroute to a site (R2 = 0.65; table II).
DISCUSSION
Implications of postlarval physiological tolerances for recruitment in
Florida Bay
At typical south Florida winter water temperatures and salinities (22°C and
35 ppt), TTM for P. argus postlarvae is 15 days; in the summer TTM decreases
to 9 days at 29°C and 35 ppt. Examination of water temperature records for
Florida Bay for the past 5 years indicate that winter water temperatures
occasionally dip below 20°C (i.e., 13-18°C) and summer temperatures can
exceed 32°C. The consequences of such a thermal regime for recruitment of
postlarvae into Florida Bay may be manifested in effects on both TTM and
survival. Time-to-metamorphosis for P. argus postlarvae raised at 18°C and 35
ppt in our study was approximately 27 days, as compared to 8 days at 33°C. Because postlarvae recruit throughout the year in south Florida, those entering Florida Bay during cold winter months could potentially be transported further
into the bay due to their prolonged postlarval period. However, physical conditions in the bay may nullify this potential. North winds are common in the
winter and may set up south or southeast flowing surface currents that might limit postlarval movement into the bay. During the summer, postlarval recruit-
ment is probably restricted to areas close to the southern chain of keys due to
the rapid TTM at warm temperatures. The laboratory results reported here also suggest that recruitment of postlar-
vae in Florida Bay is likely to vary geographically and seasonally due to the
38
TABLE II
Results of the multiple regression analyses using physical factors (distance-KM and number of banks-BANK), environmental factors (salinity-PPT), and habitat characteristics (proportion of bottom covered by algae-A; seagrass-SG; sand-
SAND, sponges-SPO; octocorals-OCT; and other-O; density of sponges-NSPO; octocorals-NOCT; corals-NC; and solution holes-NSOL) to predict the number
of juvenile Panulirus argus >35 mm CL (JUV), and the number of postlarvae (PL)
settling at a particular site. The number of juveniles <_35 mm CL could not be
predicted. Only the significant models and their corresponding ANOVA tables
and standardized variable contribution estimates are listed

40
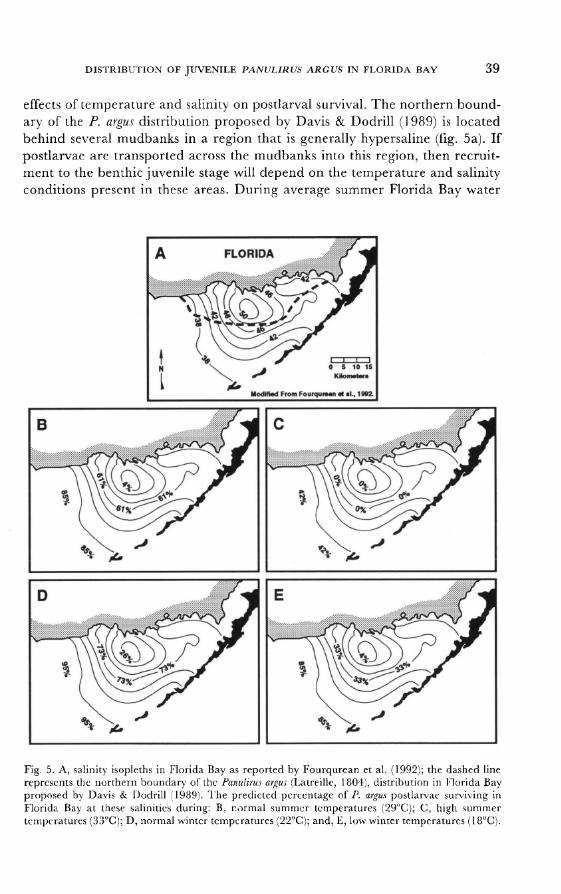
temperatures (i.e., 29°C), up to 61 % of recruiting postlarvae could tolerate a
salinity of 45 ppt, whereas only 4% could withstand a salinity of 50 ppt (fig. 5b). Under these conditions, we would expect low postlarval survival and poor recruitment to the early benthic juvenile stage in the interior regions (i.e., at the
northern P. argus boundary) of the bay. No postlarvae could tolerate extreme
summer temperatures of 33°C and salinities >45 ppt; therefore, recruitment to
the bays interior would be unlikely (fig. 5c). Cooler winter temperatures would
also reduce postlarval survival. Approximately 73% of the recruiting postlarvae could withstand winter Florida Bay water temperatures of 22°C at a salinity of
45 ppt, and 26% could withstand a salinity of 50 ppt (fig. 5d). However,
periodic cold snaps, combined with high salinities in the interior of the bay would be lethal to P. argus postlarvae; postlarval survival is only 4% at 18°C and
50 ppt (fig. 5e). Thus, the typical salinity and temperature regimes within Florida Bay proba-
bly limit recruitment of P. argus in the interior portions of the bay. In fact, the
distributional boundary for P. argus in Florida Bay first reported by Davis &
Dodrill (1989) is well described by the expected postlarval survival probabilities associated with salinity-temperature tolerance isopleths (fig. 5). Salinities
between 45-50 ppt and extreme seasonal temperatures (<20°C or >30°C) may be the best descriptor of this geographic boundary.
Plans are underway to restore much of the natural freshwater flow and
hydroperiod within the Everglades. Florida Bay would be the ultimate recipient of this freshwater, so these measures are likely to lower salinities within the bay. A moderate reduction in salinities to near 35 ppt would extend the potential
nursery region northward, assuming adequate postlarval transport and habitat
availability. However, a large decline in salinity (<35 ppt) in the bay would be
as detrimental to P. argus recruitment as high salinities. Postlarval survival at 25
ppt is about 60% at 22-29°C, but at extreme temperatures (18° and 33°C)
postlarval survival decreases to 6% and 13%, respectively. A laboratory study
by Witham et al. (1968) also indicated that low salinities found in the St. Lucie
(Florida ) estuary limit P. argus recruitment. Thus, the geographic limits of P.
argus recruitment in Florida Bay are not only determined by the present
hypersaline conditions, but would also be affected by the hyposalinc conditions
resulting from restoration of the Everglades freshwater flow.
Implications of geography and habitat structure for postlarval recruitment
in Florida Bay
Geographic features in the nursery, such as the number and distribution of
mudbanks, not only restrict water circulation within Florida Bay, but may also
inhibit the transport of postlarvae and, thus, the recruitment of juvenile P.
argus. No postlarvae were found in benthic collectors beyond the first series of
mudbanks suggesting that the banks may indeed be barriers to postlarval

41
transport. Furthermore, field surveys show that fewer than 12% of the juvenile P. argus collected at sites near the northern boundary of the P. argus distribution
in Florida Bay were small individuals ?35 mm CL as compared to 37%
elsewhere (fig. 3). Although large individuals arc nomadic and can immigrate into the bay's interior, juveniles <_35 mm CL are not thought to move far from
where they initially settle as postlarvae (Herrnkind et al., in press). Thus, the
paucity of small lobsters in northern Florida Bay reinforces the benthic collec-
tor data and collectively suggests that postlarval settlement is limited in the
interior regions of Florida Bay. Twin Keys Basin is the only northern basin we surveyed that contained
juvenile P. argus and it is unique among the basins that lie to the north of the
mudbank barrier in terms of physical structure and habitat diversity. The
mudbank to the south of Twin Keys Basin is dissected by several wide, rela-
tively deep channels, so water circulation may not be as restricted in this basin
as it is in others. Perhaps most important is the presence of large hardbottom
areas in Twin Keys Basin that contain abundant postlarval settlement habitat
(e.g., Laurencia) and postsettlement shelter (e.g., sponges, octocorals, etc.). This
type of habitat does not occur in most of the other basins, which contain either
seagrass (Johnson Key Basin) or silted, open hardbottom (e.g., Panhandle Key Basin and the Crab Keys Basin; fig. 4). Apparently few postlarvae reach Twin
Keys Basin, but those that do are more likely to find suitable settlement and
post-settlement habitat there than in adjacent basins behind mudbanks.
Nursery habitat structure plays an important role in determining the distri-
bution of P. argus elsewhere in Florida Bay (Herrnkind & Butler, in press). The
availability of suitable macroalgal settlement habitat influences postlarval sur-
vival to the first benthic juvenile stage (Herrnkind & Butlcr, 1986; Herrnkind et
al., 1988; Smith & Herrnkind, 1992), and the presence of sponges, coral heads, octocorals and other structures is critical to the survival of post-algal stage
juveniles (Smith & Herrnkind, 1992). Thus, by investigating habitat charac-
teristics at a particular site, and combining these data with geographic and
water quality information, we may be able to predict postlarval settlement and
the number of juvenile P. argus found at a site.
The multiple regression model predicting the magnitude of postlarval settle-
ment in an area was significant (R2 = 0.65; table II) and best predicted by the minimum number of banks that postlarvae pass over enroute to a site, salinity, and the proportion of bottom covered by sand. Banks are barriers to physical
transport, high salinities decrease postlarval survival, and both of these predic- tor variables were negatively related to postlarval settlement. The proportion of
bottom covered by sand explained the least variance in the model but it was
positively associated with settlement, which is difficult to explain. Postlarvae
that settle on open sand arc quickly consumed (Herrnkind & Butler, 1986; Smith & Herrnkind, 1992), but hardbottom nursery areas often contain large
expanses of open sand bottom. The model incorporating post-algal shelter

42
densities, rather than percent cover to predict the settlement of postlarvae,
yielded even better results (R2 = 0.98; table II). Again, salinity was important in
predicting postlarval settlement, and we know that abnormal salinites nega-
tively influence lobster survival, but the negative relationship between postlar- val settlement and the most influential variable in the model (the density of
octocorals at a site) is not easily interpreted. The density of solution holes in an
area was also positively associated with the magnitude of postlarval settlement, and was as influential in the model as salinity. Solution holes are indicative of
hardbottom habitat, which may again explain this association.
Both models predicting the number of large juvenile P. argus (i.e., those >35
mm CL) found at a site were also significant. The model based on habitat
coverage (Rz = 0.66; table II) indicated that the number of large juveniles at a
site was positively and strongly associated with octocoral cover. Octocorals are
characteristic of hardbottom areas, their abundance is significantly correlated
with sponge abundance (r = 0.66; p<0.001 ), and juvenile lobsters are often
found under these two shelter types. The association between large juveniles and algae, sand, and the "other" habitat categories (e.g., small coral heads and
solution holes) was negative, but was much weaker than that with octocorals. The size class designation ">35 mm CL" includes a broad range of sizes some of which use these types of shelters (e.g., sub-adults that are 50-75 mm CL often
reside under small coral heads or in solution holes). Among-site differences in
the distribution of individuals of different sizes within this broad size category is
likely to produce very different habitat-specific associations. The model incor-
porating shelter densities (table II) again yielded better predictions (R2 = 0.92) for the number of large juveniles (>35 mm CL) found at a site than the model
incorporating percent cover data. Octocorals were again positively associated
with the number of juveniles whereas coral and solution hole densities were
negatively associated with juvenile abundance. Salinity also had a strong influ-
ence on the number of juveniles >35 mm CL found in region. No one has
exposed juvenile P. argus to the temperatures and salinity conditions present in
Florida Bay, so we cannot be certain that hypersaline conditions are as detri-
mental to juvenile P. argus as they are to postlarvae. No combination of
regressor variables proved useful in predicting the number of small juvenile lobsters mm CL) at a site. Small juveniles <35 mm CL are the most
difficult size class to accurately census, and their abundance at a site may be a
poor reflection of present habitat (e.g., algal abundance) and environmental
conditions (e.g., salinity).
Conclusions
The number of P. argus postlarvae entering the Florida Bay nursery each
month is highly variable (Little, 1977; Forcucci et al., in press) and their
transport into the bay's interior basins appears limited, presumably due to the

43
extensive mudbank system that restricts water circulation in the bay. The
survival of postlarvae that arrive in these basins depends on the salinity and
temperature regimes there and the availability of settlement and post-settle- ment habitat. Salinities in central Florida Bay typically exceed 45 ppt and when
water temperatures are extreme (<20°C or >30°C), postlarval survival, hence
recruitment to later juvenile stages, will be poor. Given the high salinities in the
central bay, postlarval survival is unlikely to exceed 25% at normal
temperatures. The presence of macroalgal settlement habitat for postlarvae and shelter for
postalgal juveniles may also limit recruitment in portions of Florida Bay. With
the exception of Twin Keys Basin, the interior basins studied here lacked
suitable spiny lobster nursery habitat. Thus, the intricate mudbank system,
high salinities, occasionally extreme temperatures, and distribution of nursery habitat in Florida Bay all conspire to limit recruitment of P. argus in the interior
portions of the bay. If so, P. argus recruitment in south Florida may be limited to
a much smaller region, adjacent to the Florida Keys, than was previously
thought. Projected changes in the delivery of freshwater from the Everglades to
Florida Bay are likely to alter the present distribution of nursery areas within
the bay and could potentially enlarge the region suitable for recruitment, given sufficient postlarval abundance and nursery habitat.
ACKNOWLEDGMENTS
We thank W. Herrnkind, J. Hunt, M. Childress, M. Kuhlmann, C. Bowling, T. Matthews, R. Bertlesen, B. Sharp, C. Acosta, G. Hyde, S. Gornak, and J.
Kipp for their assistance and suggestions throughout the study. Many others
also assisted us in collecting postlarvae and we owe them our thanks. For the
use of the Keys Marine Laboratory, we thank the Florida Institute of Ocean-
ography, the Florida Department of Environmental Protection - Florida
Marine Research Institute, and the staff of the Keys Marine Laboratory. The
Everglades National Park permitted us access to many sites and provided historic temperature and salinity data for portions of Florida Bay. This reserach
was supported by a Lerner-Gray Marine Research (American Museum of
Natural History) Grant to J. Field, and by a Florida Sea Grant award (R/LR-
B-30) to M. Butler and W. Herrnkind.
LITERATURE CITED
BUTLER, M. J., IV & W. F. HERRNKIND, 1991. Effect of microhabitat cues on the metamorphosis of pueruli of the spiny lobster, Panulirus argus. Journ. Crust. Biol., 11:23-28.
- - & --, 1992. Spiny lobster recruitment in South Florida: quantitative experimcnts and management implications. Proc. Gulf Carib. Fish. Inst., 41: 508-515.
- - & - - , in press. Spiny lobster recruitment: field tests of ecological bottlenecks and the potential for population enhancement. Ecol. appl.
CALINSKI, M. D. & W. G. LYONS, 1983. Swimming bchavior of the puerulus of the spiny lobster, Panulirus argus (Latreille) (Crustacca: Palinuridae). Journ. Crust. Biol., 3: 329-335.

44
CONOVER, W. J. & R. L. IMAN, 1981. Rank transformations as a bridge between parametric and nonparametric statistics. Amer. Stat., 35: 124-129.
DAVIS, G. E., 1978. Management recommendations for juvenile spiny lobsters, Panulirus argus, in Biscayne National Monument, Florida. U.S. Natl. Park Ser. S. Florida Res. Center Rep., M-530, 32 pp.
DAVIS, G. E. & J. W. DODRILL, 1980. Marine parks and sanctuaries for spiny lobster fisheries management. Proc. Gulf. Caribb. Fish. Inst., 32: 194-207.
- - & --,1989. Recreational fishery and population dynamics of spiny lobsters, Panulirus argus, in Florida Bay, Everglades National Park, 1977-1980. Bull. mar. Sci., 44: 78-88.
ENOS, P. & R. D. PERKINS, 1979. Evolution of Florida Bay from island statigraphy. Bull. geol. Soc. America, 90: 59-83.
FIELD, J. M., 1993. The influence of temperature, salinity, and postlarval transport of the distribution of juvenile spiny lobster, Panulirus argus, in Florida Bay. (MS Thesis, Old Dominion University, Norfolk, VA, U.S.A.).
FORCUCCI, D., M.J. BUTLER, IV & J. H. HUNT, in press. Population dynamics of juvenile Caribbean spiny lobster, Panulirus argus, in Florida Bay, FL (USA). Bull. mar. Sci.
FOURQUREAN, J. W., J. C. ZIEMAN & G. V. N. POWELL, 1992. Phosphorus limitation of primary production in Florida Bay: Evidence for C:N:P ratios of dominant seagrass Thalassia les- tudinum. Limnol. Oceanogr., 37: 162-171.
HERRNKIND, W. F. & M. J. BUTLER, IV, 1986. Factors regulating postlarval settlement and juvenile microhabitat use by spiny lobsters Panulirus argus. Mar. Ecol. Progr. Ser., 34: 23-30.
- - & - --, 1994. Settlement of spiny lobster, Panulirus argus (Latreille, 1804), in Florida: pattern without predictability. Crustaccana, 67 (1): 46-64 (this volume).
HERRNKIND, W. F. & M. J. BUTLER, IV & R. A. TANKERSLEY, 1988. The effects of siltation on recruitment of spiny lobsters, Panulirus argus. Fish. Bull., 86: 331-338.
HERRNKIND, W. F., P. JERNAKOFF & M. J. BUTLER, IV, in press. Puerulus and post-puerulus ecology. In: B. PHILLIPS, S. COBB &J. KITTAKA (eds.), Spiny lobster management: current situation and perspectives. (Blackwell Press, London).
HOLMQUIST, J. G., G. V. N. POWELL & S. M. SOGARD, 1989a. Decapod and stomatopod commu- nities of seagrass-covered mud banks in Florida Bay: inter- and intra-bank heterogeneity with special reference to isolated subenvironments. Bull. mar. Sci., 44:251-262.
- - & --, 1989b. Sediment, water level and water temperature characteristics of Florida Bay's grass covered mud-banks. Bull. mar. Sci., 44: 348-364.
IMAN, R. L. & W.J. CONOVER, 1979. The use of rank transformation in regression. Technometrics, 21: 499-509.
KANCIRUK, P., 1980. Ecology of juvenile and adult Palinuridae (spiny lobsters). In : J. S. COBB & B. F. PHILLIPS (eds.). The biology and management of lobsters, 2: 59-92. (Academic Press, New York).
LITTLE, E.J.,Jr., 1977. Observations on recruitment of postlarval spiny lobsters, Panulirus argus, to the south Florida coast. Florida mar. Res. Publ., 29, 35 pp.
LITTLE, E. J., Jr. & G. R. MILANO, 1980. Techniques to monitor postlarval recruitment of postlarval spiny lobsters, Panulirus argus, to the Florida Keys. Florida mar. Res. Publ., 37, 16 pp.
MARX, J. M., 1986. Settlement of spiny lobster, Panulirus argus, pucruli in South Florida: an evaluation from two perspectives. Canadian Journ. Fish. aquat. Sci., 43: 2221-2227.
MARX, J. M. & W. F. HERRNKIND, 1985. Macroalgae (Rhodophyta: Laurencia spp.) as habitat for juvenile spiny lobster, Panulirus argus. Bull. mar. Sci., 36: 423-431.
ROBBLEE, M.,J. R. BARBER, P. B. CARLSON, M.J. DURAKO, J. W. FOURQUREAN, L. K. MUEHLSTEIN, D. PORTER, L. A. YAKRBO, R. T. ZIEMAN & J. C. ZIEMAN, 1991. Mass mortality of the tropical seagrass Thalassia testudinum in Florida Bay (USA). Mar. Ecol. Progr. Ser., 71: 297-299.
SMITH, K. N. & W. F. HERRNKIND, 1992. Predation on early juvenile spiny lobsters, Panulirus argus (Lat.): influence of size and shelter. Journ. exp. mar. Biol. Ecol., 64: 1-16.
SWEAT, D. E., 1968. Growth and tagging studies on Panulirus argus (Latreille) in the Florida Keys. Florida Bd Conserv. mar. Res. Lab. tech. Ser., 57: 30 pp.
WAHLE, R. A. & R. S. STENECK, 1991. Recruitment habitats and nursery grounds of the American lobster Homarus americanus: a demographic bottleneck? Mar. Ecol. Progr. Ser., 69: 231-243.
WALTERS, C. L. GUNDERSON & C. S. HOLLING, 1992. Experimental policies for water management in the Everglades. Ecol. appl., 2: 189-202.

45
WITHAM, R., R. M. INGLE & E. A. JOYCE, Jr., 1968. Physiological and ecological studies of Panulirus argus from the St. Lucie estuary. Florida Bd Conserv. mar. Res. Lab. tech. Ser., 53, 31 pp.
YOSHIMURA, T. & H. YAMAKAWA, 1988. Ecological investigations of settled postlarval and juvenile stages of the Japanese spiny lobster Panulirus japonicus at Kominato, Japan. Journ. Crust. Biol., 8: 524-531.
ZIEMAN, J. C., J. W. FOURQUREAN & R. L. IVERSON, 1989. The distribution and abundance of seagrass in Florida Bay. Bull. mar. Sci., 44: 292-311.





























