Embed Size (px)
Citation preview

Introduction: MHC class II-restricted antigen presentationJim Miller* and Andrea J. Sant†
THE CONCEPT THAT class II-restricted antigens must beprocessed by viable antigen presenting cells prior topresentation to CD4-positive T cells was first proposedby Ziegler and Unanue in 1981.1 Antigen processingwas defined as a degradative process based on theinhibitory capacity of lysosomotropic reagents2 and,later, protease inhibitors.3 This concept was con-firmed by the demonstration that in-vitro proteolysisof antigen could substitute for the intracellularprocessing events.4 That T cells recognize peptideepitopes was consistent with previous work that haddemonstrated that CD4-positive T cells were depend-ent on the primary amino acid sequence of shortsegments of foreign proteins. Direct binding studies5,6
demonstrated that class II molecules could formstable complexes with synthetic, antigenic peptides.When these in-vitro class II–peptide complexes wereinserted into artificial, planer membranes, they servedas effective ligands for T-cell stimulation.7 Collectively,these data led to the concept that foreign antigen wasinternalized and degraded into peptides that formedstable complexes with class II molecules. These classII–peptide complexes were expressed on the cellsurface of antigen presenting cells where they servedas recognition molecules for CD4-positive T cells.
During the time that the general framework of classII-restricted antigen presentation was being defined,antigen presentation to class I restricted cells wasthought to be mediated by simultaneous recognitionof intact protein antigens and class I moleculesexpressed at the cell surface. This apparent bias forclass I to associate with endogenously synthesizedplasma membrane proteins and for class II to asso-ciate with soluble, exogenous antigens was supportedby work from Braciale and colleagues.8 Class I antigenrecognition was re-evaluated in 1986 following AlainTownsend’s work demonstrating that class I-restrictedantigenic epitopes could also be defined by syntheticpeptides.9 Thus, MHC-restricted recognition of pep-tide antigens was a common theme for both class Iand class II presentation.
Understanding the molecular events that wereinvolved in the generation of class I-peptide com-plexes was initiated in 1989 by the discovery of mutantcell lines that were defective in this process.10 Thegenes associated with this defect were subsequentlyidentified as peptide transporters (TAP). TAP areintegral membrane proteins in the endoplasmicreticulum (ER) that actively transport peptides fromthe cytosol into the lumen of the ER, where they areavailable for class I binding (for review see ref 11).The TAP proteins have recently been shown tointeract directly with class I,12,13 suggesting that theymay be involved in both transport of peptide acrossthe ER membrane and loading the peptides onto classI molecules. The pool of cytosolic peptides is in partgenerated from a large, multi-subunit proteolyticcomplex, called the proteosome. Two components ofthe proteosome (LMP) are actually encoded in theMHC.14 Interestingly, the TAP genes are also linked toMHC, indicating that in addition to the structuralclass I genes, the MHC contains genes that encodesome of the cellular machinery involved in theformation of MHC-peptide complexes. Both thegenetic and biochemical analyses led to the conceptthat class I-peptide association is mediated by aregulated series of protein–protein interactions thatgenerates peptides in the cytosol, transports them intothe ER, and directly loads them into class I duringclass I folding and assembly with â2-microglobulin.
Many studies on class II function have focused onthe unique ability of class II molecules to assemblewith peptides that are generated in the endosomal/lysosomal pathway. Early studies from Cresswell15 andlater from Neefjes et al16 indicated that class IIfollowed an unconventional route from the Golgi tothe cell surface, being diverted first into endosomalcompartments. These endosomal compartments havesince been identified as an unusual late endosomalcompartment, called the MIIC (ref 17, see review byHarding, this issue). This compartment is thought tobe a major site in which class II encounters and bindsto peptides derived from endocytosed proteins.Whether this and/or other compartments are thesites for antigen processing is not resolved (see reviewby Jensen, this issue). Class II-restricted antigen
From the Committee on Immunology and Departments ofMolecular Genetics and Cell Biology, and †Pathology, University ofChicago, Chicago, IL 60637, USA
©1995 Academic Press Ltd
seminars in IMMUNOLOGY, Vol 7, 1995: pp 343–346
343

presentation is facilitated through the action of atleast two accessory proteins. Invariant chain (Ii) is anon-polymorphic protein that associates with class IIearly in biosynthesis in the ER and is proteolyticallycleaved from class II in endosomal compartments (forreview see ref 18). Ii plays a role in class II assembly,folding, egress from the ER, and targeting class II intoendosomes. Ii can also regulate class II–-peptideloading, both by protecting the class II peptidebinding site in the ER and by enhancing peptidegeneration or loading in endosomes (see reviews byGermain and by Ceman and Sant, this issue). HLA-DM is an MHC-linked, class II-like heterodimer that iscolocalized with class II in MIIC compartments and insome cases appears to be critical for the generation ofclass II–peptide complexes.19-23 Interestingly, neitherIi or HLA-DM are essential components for the class IIantigen presentation pathway, suggesting that unlikeclass I, class II–peptide loading may not be a single,localized and orchestrated event. Rather, class II maybe capable of associating with a wide array of peptidesgenerated in a variety of intracellular compartmentsand only some of these events may be mediated by Iior HLA-DM.
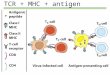
One common feature to class I and class IImolecules is that peptide can be thought of as anintegral component of the complex. Both class I andclass II molecules are biochemically more stable in thepresence of peptide and it is clear from the crystalstructure of these molecules that peptide is tightlyfolded into the final complex (see review by Schafer etal, this issue). Interestingly, the peptide binding sitesdiffer in that class I binds peptide of defined lengthwith predictable contact residues for the N- andC-termini of the peptide, whereas the peptide bindingsite in class II is open at both ends allowing forassociation with peptides of variable length. Thisdichotomy was clearly illustrated when endogenouslyloaded peptides were eluted from class I24,25 and classII26-28 molecules; class I contained discrete peptides ofmostly nine amino acids and class II contained nestedsets of peptides varying in length at both the N- andC-termini. The source of peptides found associatedwith class II were derived from both exogenous andendogenous cellular proteins (see review byRudensky, this issue). This raises the question as tohow, in the face of a sea of host proteins and derivedpeptides, antigen presenting cells manage to producesufficient peptide–MHC complexes from foreign anti-gens to activate T cells. One answer is that T cells canbe activated by very small numbers of specific pep-tide–MHC complexes, possibly in the order of only a
few hundred.29,30 Interestingly, these small numbersof complexes may serially engage many TcR throughrapid association and dissociation.31
This issue of Seminars in Immunology contains reviewsthat address many of the key issues involved in MHCclass II-restricted antigen presentation, from process-ing of internalized protein antigens, through theintracellular transport of class II–invariant chaincomplexes, and to the functional expression of class IIpeptide complexes at the cell surface.
Although it is simple to follow the catabolism ofinternalized proteins, delineation of the precise orderof unfolding, proteolysis and class II association hasnot been possible. This is compounded by theextremely small number of class II–peptide complexesrequired to activate T cells. These issues are addressedin the first review by Peter Jensen, with a particularemphasis on the mechanism and importance ofacidification and disulfide bond reduction in proteinunfolding and antigen processing. One issue that isbrought to light is whether class II molecules associatewith unfolded proteins prior to or after antigenproteolysis. There is precedent for both pathways andthe former may account for some of the strikingphenomena of epitope dominance and, followingtrimming by exopeptidases, the generation of nestedsets of peptides.
One of the more exciting results in the last few yearsis the identification of a specific compartment whereclass II–peptide association may take place. Althoughthis MIIC is clearly an important site for class II–peptide binding, it is not clear whether this is the onlyintracellular compartment whereby class II can accesspeptide. Likewise, it is not certain whether this andother peptide binding compartments are identical inall cell types. The identification of class II–peptidebinding compartments and the generation of thesecompartments in different cell types is reviewed byCliff Harding.
In addition to targeting of class II into MIICcompartments, several other steps in class II transportare regulated. For example, many of the class IIpolymorphic residues are in regions that interactbetween class II αâ chains. Differences in theseregions can determine the efficiency of dimer forma-tion, a step that is essential for exit from the ER andtransport to the Golgi. In addition, access to the classII peptide binding site must be regulated to inhibitformation of complexes with various partially foldedand misfolded proteins that class II might encounterduring the biosynthetic pathway. Ron Germain dis-cusses how invariant chain can function as a surrogate
Introduction
344

peptide in the ER, facilitating class II assembly andprotecting the class II peptide binding site and howconventional peptides can modulate class II con-formation in endosomes by stabilizing the structure ofclass II dimers. Thus, assembly of class II and foldingof the class II peptide binding site may regulate atleast two essential intracellular transport steps, egressfrom the ER to the Golgi and release from endosomesto the cell surface.
In addition to the ability of Ii to function as asurrogate and replaceable peptide in class II, Ii canhave direct effects on the efficiency of antigenpresentation. However, this ability is not universal andmany antigenic epitopes are presented equally well inthe presence and absence of Ii. Likewise, HLA-DMcan modulate antigen presentation, but in an allele-dependent fashion. Efficient antigen presentation inthe absence of Ii or HLA-DM raises questions aboutboth the regulation of intracellular transport of classII and the mechanism of class II–peptide loading.Stephanie Ceman and Andrea Sant discuss the natureof Ii-dependent and Ii-independent antigens, thevarious pathways whereby Ii can facilitate antigenprocessing, and the role of HLA-DM.
Crystallization of class II proteins revealed a pro-pensity for the molecule to form a dimer of dimers, orsuperdimer. Although superdimer formation was acommon feature to several crystal forms of class II andappealing for some models of T cell activation, it wasnot until recently that the formation of class IIsuperdimers has been detected biochemically. PeterSchafer, Sue Pierce and Ted Jardetzky review both thestructural and biochemical data on this superdimerand discuss the implications such a superdimer has onthe cognate interaction between antigen-presentingcells and T cells.
The vast majority of class II molecules that areexpressed at the cell surface are associated withpeptide and under normal conditions, most of thesepeptides are derived from endogenous antigens. Thearray of peptides that can be displayed at the cellsurface is determined by the structure of the class IIpeptide binding site and so different MHC haplotypeswill present different sets of peptides. This phenome-non may account for the finding that susceptibility tomany autoimmune diseases can be MHC-associated.Alexander Rudensky reviews the generation of endo-genous peptide–class II complexes and how theymight be generated in a tissue specific manner. Healso discusses how the array of class II–peptidecomplexes can shape the repertoire of T cells through
positive and negative selection events during T celldevelopment in the thymus.
In the past few years we have learned much aboutthe cell biology of class II-restricted antigen presenta-tion. We have identified many of the players andclarified many of the events in this process. However,there are many areas that remain unresolved. Onecommon theme in immune regulation is that thereare often parallel paths to the same endpoint. Forexample, gene disruption of many immunologicallyrelevant proteins has revealed significant redundancyin the generation of functional immune responses.Within the class II antigen presentation pathway theconcept is emerging that although there may be onepredominant mechanism for class II–peptide loading,it is not the only pathway. Many divergent pathwaysappear to exist that allow for class II to gain access topeptides that are generated through unusual process-ing events or in different intracellular compart-ments.32-35 In combination with the sensitivity of T-cellantigen recognition, this allows the immune system tosample a wide variety of antigens in the context ofclass II antigens. This potential diversity in thepathways that can be used for class II-restrictedantigen presentation adds an extra level of complexityin understanding the cell biology of antigen presenta-tion in the context of class II molecules.
References
1. Ziegler K, Unanue E (1981) Identification of a macrophageantigen-processing event required for I-region-restricted anti-gen presentation to T lymphocytes. J Immunol 127:1869-1875
2. Ziegler K, Unanue E (1982) Decrease in macrophage antigencatabolism caused by ammonia and chloroquine is associatedwith inhibition of antigen presentation to T cells. Proc NatlAcad Sci USA 79:175-178
3. Streicher H, Berkower I, Busch M, Gurd F, Berzofsky J (1984)Antigen conformation determines processing requirements forT cell activation. Proc Natl Acad Sci USA 81:6831-6835
4. Shimonkevitz R, Kappler J, Marrack P, Grey H (1983) Antigenrecognition by H-2 restricted T cells: cell free antigen process-ing. J Exp Med 158:303-316
5. Babbitt R, Allen P, Matseuda G, Huber E, Unanue E (1985)Binding of immunogenic peptides to Ia histocompatibilitymolecules. Nature 317:359-361
6. Buus S, Sette A, Colon S, Jenis D, Grey H (1986) Isolation andcharacterization of antigen-Ia complexes involved in T cellrecognition. Cell 47:1071-1077
7. Watts T, Brian A, Kappler J, Marrack P, McConnell H (1984)Antigen presentation by supported planar membranes contain-ing affinity-purified I-Ad. Proc Natl Acad Sci USA81:7564-7568
8. Morrison L, Lukacher A, Braciale V, Fan D, Braciale T (1986)Differences in antigen presentation to MHC class I- and class II-restricted influenza virus-specific cytolytic T lymphocyte clones.J Exp Med 163:903-921
Introduction
345

9. Townsend A, Rothbard J, Gotch F, Bahadur G, Wraith D,McMichael A (1986) The epitopes of influenza nucleoproteinrecognized by cytotoxic T lymphocytes can be defined withshort synthetic peptides. Cell 44:959-968
10. Townsend A, Ohlen C, Bastin J, Ljunggren H, Foster L, Karre K(1989) Association of class I major histocompatibility heavy andlight chains induced by viral peptides. Nature 340:443-448
11. Howard J (1995) Support and transport of peptides presentedby class I molecules. Curr Opin Immunol 7:69-76
12. Ortmann B, Androlewicz M, Cresswell P (1994) MHC classI/beta 2-microglobulin complexes associate with TAP trans-porters before peptide binding. Nature 368:864-867
13. Suh W, Cohen-Doyle M, Fruh K, Wang K, Peterson P, WilliamsD (1994) Interaction of MHC class I molecules with thetransporter associated with antigen processing. Science264:1322-1326
14. Brown M, Driscoll J, Monaco J (1991) Structural and sero-logical similarity of MHC-linked LMP and proteasome (multi-catalytic proteinase) complexes. Nature 353:355-357
15. Cresswell P (1985) Intracellular class II HLA antigens areaccessible to transferrin-neuraminidase conjugates internalizedby receptor-mediated endocytosis. Proc Natl Acad Sci USA82:8188-8192
16. Neefjes J, Stollorz V, Peters P, Geuze H, Ploegh H (1990) Thebiosynthetic pathway of MHC class II but not class I moleculesintersects the endocytic route. Cell 61:171-183
17. Peters P, Neefjes J, Oorschot V, Ploegh H, Geuze H (1991)Segregation of MHC class II molecules from MHC class Imolecules in the Golgi complex for transport to lysosomalcompartments. Nature 349:669-676
18. Sant A, Miller J (1994) MHC class II antigen processing:Biology of invariant chain. Curr Opin Immunol 6:57-63
19. Denzin L, Robbins N, Carboy-Newcomb C, Cresswell P (1994)Assembly and intracellular transport of HLA-DM and correc-tion of the class II antigen-processing defect in T2 cells.Immunity 1:595-606
20. Fling S, Arp B, Pious D (1994) HLA-DMA and HLA-DMB genesare both required for MHC class II/peptide complex formationin antigen presenting cells. Nature 368:554-558
21. Morris P, Shaman J, Attaya M, Amaya M, Goodman S, BergmanC, Monaco J, Mellins E (1994) An essential role for HLA-DM inantigen presentation by class II MHC molecules. Nature368:551-554
22. Sanderson F, Kleijmeer M, Kelly A, Verwoerd D, Tulp A, NeefjesJ, Geuze H, Trowsdale J (1994) Accumulation of HLA-DM aregulator of antigen presentation, in MHC class II compart-ments. Science 266:1566-1569
23. Stebbins C, Loss G, Elias C, Chervonsky A, Sant A (1994) Therequirement for DM in class II restricted antigen presentationand SDS-stable dimer formation is allele and species depend-ent. J Exp Med 181:223-234
24. Falk K, Rotzschke O, Stevanovic S, Jung G, Rammensee H-G(1991) Allele-specific motifs revealed by sequencing of self-peptides eluted from MHC molecules. Nature 353:290-296
25. Hunt D, Henderson R, Shabanowitz J, Sakaguchi K, Michel H,Sevilir N, Cox A, Appella E, Engelhard V (1992) Character-ization of peptides bound to the class I MHC molecule HLA-A2.1 by mass spectrometry. Science 255:1261-1263
26. Rudensky A, Preston-Hurlburt P, Hong S, Barlow A, Janeway C(1991) Sequence analysis of peptides bound to MHC class IImolecules. Nature 353:622-627
27. Hunt D, Michel H, Dickinson T, Shabanowitz J, Cox A,Sakaguchi K, Appella E, Grey H, Sette A (1992) Peptidespresented to the immune system by the murine class II majorhistocompatibility complex molecule I-Ad. Science256:1817-1820
28. Chicz R, Urban R, Gorga J, Vignali D, Lane W, Strominger J(1993) Specificity and promiscuity among naturally processedpeptides bound to HLA-DR alleles. J Exp Med 178:27-47
29. Harding C, Unanue E (1990) Quantitation of antigen present-ing cell MHC class II/peptide complexes necessary for T cellstimulation. Nature 346:574-576
30. Demotz S, Grey H, Sette A (1990) The minimal number of classII MHC-antigen complexes needed for T cell activation.Science 249:1028-1030
31. Valitutti S, Muller S, Cella M, Padovan E, Lanzavecchia A(1995) Serial triggering of many T-cell receptors by a fewpeptide-MHC complexes. Nature 375:148-151
32. Malnati M, Marti M, LaVaute T, Jaraquemada D, Biddison W,DeMars R, Long E (1992) Processing pathways for presentationof cytosolic antigen to MHC class II restricted T cells. Nature357:702-704
33. Loss G, Elias C, Fields P, Ribaudo R, McKisic M, Sant A (1993)MHC class II–restricted presentation of an internally synthe-sized antigen displays cell type variability and segregates fromthe exogenous class II and endogenous class I antigenpresentation pathways. J Exp Med 178:73-85
34. Bodmer H, Viville S, Benoist C, Mathis D (1994) Diversity ofendogenous epitopes bound to MHC class II molecules limitedby invariant chain. Science 263:1284-1286
35. Bijlmakers M, Benafoch P, Ploegh H (1994) Assembly of HLADR1 molecules translated in vitro: binding of peptide in theendoplasmic reticulum precludes association with invariantchain. EMBO J 13:2699-2707
Introduction
346