Embed Size (px)
Citation preview

Proc. Natl. Acad. Sci. USAVol. 88, pp. 4494-4497, May 1991Population Biology
Intraspecific variation in population gene diversity and effectivepopulation size correlates with the mating system in plants
(inbreeding species/genetic conservation/neutral model/maximnum likelihood/isozyme)
DANIEL J. SCHOEN* AND ANTHONY H. D. BROWNCommonwealth Science and Industry Research Organization, Division of Plant Industry, GPO Box 1600, Canberra, ACT 2601, Australia
Communicated by Michael T. Clegg, February 19, 1991 (received for review August 20, 1990)
ABSTRACT Published data on allele frequencies atisozyme loci in inbreeding and outbreeding plant species wereanalyzed to examine intraspecific variation in gene diversityand effective population size (Ne). Compared with outbreeders,inbreeding species showed markedly greater variation amongpopulations in average values of Nei's gene diversity statistic.Effective population size was estimated by assuming that thevariation observed at isozyme loci is selectively neutral. In-bieeding species showed greater levels of variation in Ne thandid outbreoders, although the upper range ofNe was similar inthe two classes of species. The results suggest that there may beconsiderable genetic variation and potential for evolutionarychange in some but not all populations of inbreeders. More-over, these findings are important with respect to the conser-vation of genetic resources. In particular, that the amount ofintraspecific variation in population genetic diversity and Nediffers between inbreeding and outbreeding species should betaken into account in sampling efforts designed to optimize thediversity of germplasm collections.
Studies ofgenetic variation in plants have typically used Nei'sgenetic diversity statistics or Wright's F statistics as tools fordescribing the extent of differentiation among populations(1-3). While informative, these statistics do not provide directinformation about the amount of among-population variationin levels of polymorphism and gene diversity, a feature thatcan be of key importance in efforts to conserve the geneticresources of a species. In this regard, a survey of isozymevariation in several plant populations conducted by Brown (4)suggested that inbreeding species will tend to show greatervariation among populations in the level of genetic diversitycompared with outbreeding ones. Brown (4) proposed that, ininbreeders, among-population variation in the average value ofNei's gene diversity statistic will often exhibit an L-shaped orbimodal distribution. Published data for the predominantlyself-pollinating plants Hordeum spontaneum and Lycopersi-con pimpinellifolium supported this view (4).
In this investigation, we analyze additional publishedisozyme data from inbreeding and outbreeding plant speciesin order to explore in detail the nature and implications ofvariation in genetic diversity among populations of inbreed-ing and outbreeding species. This is done by examining thelevel of among-population variation in gene diversity. Wehave also asked what the observed patterns of variation indiversity imply about the amount of variation in effectivepopulation size in species with contrasting breeding systems.
MATERIALS AND METHODSPublished tables of allozyme frequency data were used in allanalyses described below. These data sets were selectedaccording to the following criteria: (i) the mating system of
the species in question was known either through progenytesting of open-pollinated seed parents (5), from analysis ofgenotype frequencies (6), or based on the observation ofself-incompatibility-furthermore, to preserve an extremecontrast in the mating system of the taxa compared, weincluded only those species shown to be either predominantselfers or outcrossers; (ii) there were no apparent problemsin the genetic interpretation of the isozyme phenotypes (e.g.,we excluded from consideration species having polysomicinheritance where no formal Mendelian analysis had beenconducted); and (iii) isozyme data were available for aminimum of seven populations, allowing a representativerange of intraspecific variation in gene diversity and Ne to besampled. The species studied are listed in Table 1, togetherwith information on their mating system and the numbers ofpopulations and loci surveyed in the allozyme studies. Wenoted the existence of additional studies of outcrossing plantspecies, particularly of forest trees, but the results fromanalyses ofthese data are not included below for two reasons.First, as much as possible, we wished to balance our overallcomparison by considering data from species with a widevariety of different growth habits and ecologies. Second, datafrom additional forest tree species gave results very similar tothose for the three tree species presented below.Data on allele frequencies and numbers of individuals
sampled, obtained from the references listed in Table 1, wereused to calculate the average population value of Nei's genediversity statistic for each population (hj) according to theformula
[1]
where Pa. is the frequency of the ath allele at the ith locus (i= 1, .. ., I) in the jth population (j = 1,... , J) (1). Thedistribution of these hj values was compared for each in-breeding and outbreeding species after angular transforma-tion.To estimate effective population size (Ne), we used Chak-
raborty and Neel's (29) procedure based on Kimura andCrow's model of selectively neutral variation (30). For thismodel, Ewens (31) has shown that when a population is atequilibrium under mutation-drift balance, the likelihood (L)ofobserving k alleles in a sample ofn genes from a given locusand population is
[2]ikln.ok r(6)r(n + 0)
where the coefficient l(k) is the Stirling number ofthe first kind(32) corresponding to k and n, 6 = 4UNe, u is the mutationrate at the locus in question, and J7( ) is the gamma function(33). With I loci and J populations, there are I + J parameters
Abbreviation: MLE, maximum likelihood estimate.*Present address: Department of Biology, McGill University, 1205Avenue Dr. Penfield, Montreal, Quebec H3A IB1, Canada.
4494
The publication costs of this article were defrayed in part by page chargepayment. This article must therefore be hereby marked "advertisement"in accordance with 18 U.S.C. §1734 solely to indicate this fact.
h.j = EA EaP2VI = Ei hifl,
Dow
nloa
ded
by g
uest
on
Feb
ruar
y 18
, 202
1

Proc. Natl. Acad. Sci. USA 88 (1991) 4495
Table 1. Mating systems, numbers of populations, and numbers of loci for species used toestimate Ne
Outcrossing rate Allozyme dataor compatibility from I loci, J
Species reaction populations Ref(s).Self-fertilizing speciesArabidopsis thaliana 0* 7, 16 7Avena barbata (California) 0.02* 7, 14 8-10Avena barbata (Spain) 0.02* 8, 12 8, 9, 11Avena canariensis Ot 8, 19 12H. spontaneum 0.02* 25, 28 13, 14L. pimpinellifolium 0.14* 13, 41 15Phlox cuspidata 0.22* 5, 43 16Plantago major 0.17t 7, 7 17Polygala vulgaris 0.13t 10, 53 18
Outcrossing speciesEchium plantagineum 0.97* 16, 8 19, 20Helianthus debilis Self-incompatible 5, 17 21Phlox drummondii Self-incompatible 5, 73 16Phlox roemariana Self-incompatible 4, 15 16Picea abies 0.89* 4, 8 22, 23Pinus sylvestris 0.90* 10, 8 24, 25Plantago lanceolata Self-incompatible 14, 7 17Pseudotsuga menziesii 0.90* 18, 11 26, 27Stephanomeria exigua Self-incompatible 8, 11 28
*Outcrossing rate estimated by progeny-testing parents.tOutcrossing rate estimated from genotype frequency data in population.
(u... . . ,u1, Nei... .. Ne,). Using only those loci that segre-gate in at least one of the J populations, Chakraborty andNeel (29) have shown that the appropriate joint-likelihoodfunction is
L= [
I (k +Hi.(,J)k.(i j nij+Oj
I' J (nj 1)'!61ijF(6ij)Ht(1Hlr(+8[3]
Maximum likelihood estimates (MLEs) of the model param-eters were found by iterating the set of normal equations forthe ui and Nej (obtained through partial differentiation of Eq.3 with respect to the ui and Nej and subject to the constraintthat Oij = 4ui Nej) (29). For simplicity, we assumed that thenumber of genes sampled, n, in outcrossing populationsequals 2-Z, where Z is the number of individuals sampledfrom the population (29). With inbreeding populations, weassumed n = Z; i.e., to account for the high probability thatthe two genes sampled from single individuals are identical bydescent (2), as expected, given the high levels of inbreedingcharacteristics of most of the selfing species in our study(Table 1).Because the ui and Nej appear only in product form, it is not
possible to obtain estimates ofthem without first assuming anoverall mean value for either one of these two sets ofparameters. In our work, we assume that _u = 10-5 (31). Thisallows for estimation of the absolute values of the Nej.Importantly, while the estimates of the absolute values of theNej are dependent on the choice of u, the pattern of variabilityof the Nej is not.
RESULTS
Table 2 summarizes several characteristics on the distribu-tions ofpopulation gene diversity in eight inbreeding and nineoutbreeding plant species. Gene diversity in the inbreeders is
lower (mean = 0.12) than in the outbreeders (mean = 0.26),a result that is in accord with the larger survey conducted byHamrick and Godt (3). Moreover, apparent from this table isthe greater range and variability of h*j in the inbreeders. Theaverage range of h.1 in the inbreeding species is 0.29, com-
pared with 0.15 for the outbreeding species. The averagecoefficient of variation of h.1 is 64% in the inbreeding speciesand 12% in the outbreeding species. All but one of the
Table 2. Summary of variation in average population values ofNei's diversity statistic (h.)
Mean Min Max Range CV ofSpecies hi hi hj of hi h.j,* %
Self-fertilizing speciesA. thaliana 0.137 0 0.385 0.385 53A. barbata
(California) 0.041 0 0.268 0.268 162A. barbata (Spain) 0.068 0 0.228 0.228 94H. spontaneum 0.110 0 0.197 0.197 35L. pimpinellifoliumt 0.137 0 0.271 0.271 38P. cuspidata 0.050 0 0.252 0.252 95P. major 0.165 0.056 0.336 0.280 55P. vulgaris 0.145 0 0.264 0.264 36
Mean 0.125 0.008 0.294 0.286 64SE 0.024 0.008 0.026 0.021 15
Outcrossing speciesE. plantagineum 0.337 0.281 0.399 0.118 6H. debilis 0.130 0.022 0.228 0.206 25P. drummondii 0.178 0.023 0.390 0.367 30P. roemariana 0.275 0.108 0.372 0.264 16P. abies 0.336 0.297 0.363 0.066 4P. sylvestris 0.320 0.269 0.343 0.074 4P. lanceolata 0.355 0.290 0.373 0.083 5P. menziesii 0.193 0.164 0.218 0.054 5S. exigua 0.187 0.115 0.268 0.153 14
Mean 0.257 0.174 0.328 0.154 12SE 0.028 0.038 0.024 0.035 3
Min, minimum; Max, maximum.*Angular transform of data.tPopulations of fewer than 20 plants not included.
Population Biology: Schoen and Brown
Dow
nloa
ded
by g
uest
on
Feb
ruar
y 18
, 202
1

4496 Population Biology: Schoen and Brown
inbreeding species had some populations that were mono-morphic at all loci tested, and, therefore, the minimum hvalues in these species were 0. On the other hand, themaximum hj values in the inbreeding species were onlyslightly lower than those in the outbreeding species.
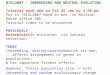
Estimates of Nej are shown in Fig. 1 for inbreeders and inFig. 2 for outbreeders. The mean values of Nej for inbreedingand outbreeding species were 3557 and 6990, respectively.Apparent from the two figures is the generally greater rangeof Nej values for the inbreeding versus outbreeding species.On average, the range of Nej in the inbreeding species was8530 (SE = 1277), while in the outbreeding species it was 4444(SE = 782). The coefficient of variation of Nej in the inbreed-ing species was 84%, on average, while in the outbreedingspecies it was 24%. The average minimum Nej in the inbreed-ing species was 86 (SE = 86), while in the outbreeding speciesit was 4668 (SE = 1119). The average maximum Nej in theinbreeding species was 8721 (SE = 1331), while in theoutbreeding species it was 9143 (SE = 1354). Significantvariation among the Nej values of the selfing species, but notthose of the outcrossing species, is further indicated byexamination of the confidence intervals of the MLEs ob-tained from the large sample variance formula (29, 31) (whereNej values of 0 have been set equal to 1). For all outcrossingspecies, the 95% confidence limit around any single estimateof relative Nej includes all other estimates, whereas for allselfing species the 95% confidence limits of the largest andsmallest Nej values do not overlap.The estimation of Ne via allele counts (29) is sensitive to
sampling effects when populations have low levels of poly-morphism, as occurs in some populations of inbreedingspecies. In particular, if a population is scored as monomor-phic for all loci tested, but is in fact polymorphic for one or
Araboh)psi5tiIXixahll Ai 'l bur/'ihienaVO Wharfala4
-Ca iforn11i i; SpI il11
81 4ni
I. vqler()p 'e suis'uIiuu
pllif7ilIII/IfiIi!Ift?
-.
C-
-r
J
. (I thl Ca'tIt At 7 (V'IN.\ I}tt{L l e ./t111 [eOiW II IM,~
0 2468 1012 0123456789
I
02 4 6 8 1 0 1 2 0 1 23 45 6 7 8 9O 0
Phiox cuspidali Plantagonll/fir
Ne x l ()0
FIG. 1. Distribution ofestimates ofNr in eight inbreeding speciesof plants.
4.,'E
}{(1/ii1t1[/l|s.t elf Fiji;
;- or-
/
Ar
4
;: g Xi j_ ^ C In <;.. . . . ....... .f ;; v1 be, he .\
1irununiJ.
HN ;
--l
frl
x
-C
Zz
rL
~~~~fE
Ne \ 1 NO
FIG. 2. Distribution of estimates of N, in nine outbreedingspecies of plants.
more loci, then Ne for that population will be erroneouslyestimated as 0. This is due to the result, shown by Ewens (31),that the MLE of 6 = 4uNe = 0 whenever loci are scored asmonomorphic. To rectify such sampling problems, a conser-vative approach to the estimation of Ne is to assume thatpopulations scored as monomorphic for all loci tested insteadcontain two alleles at the (globally) most polymorphic locus.When this is done, the distributions of N<, values for theinbreeding species remain qualitatively similar to those illus-trated in Fig. 1. For instance, the mean Nj becomes 3731,while the average range of Nj values within inbreedingspecies is slightly reduced to 7950 (SE = 1307). The averagecoefficient of variation of Nej is 66%, and the averageminimum and maximum values of Nej are 834 (SE = 145) and8723 (SE = 1362), respectively. This procedure, then, doesnot qualitatively alter the contrast in variability of Ne ob-served between inbreeding and outbreeding species.
DISCUSSIONAnalysis of published allele frequencies at isozyme loci inplants has shown that not only do inbreeding and outbreedingplant species differ in overall levels of genetic diversity, butthey also differ in the amount of among-population varianceof gene diversity. Inbreeding species exhibit markedlygreater population to population variation in Nei's genediversity statistic. In parallel with this trend, estimates of Nrwithin inbreeding species are more variable than those withinthe outbreeding species. Thus, two different analyses, one ofallele frequencies and one of allele numbers, both point to theconclusion that inbreeding species are more variable thanoutbreeding species in how genetic variation is structured.
Proc. Natl. Acad. Sci. USA 88 (1991)
P/ant age~ ) If} m .(,I /It11I a / ,, ( -I? ?ih F.. u c I ?7 I -- ", f .., i ;' N ! t ) . Li . 1 1 . ? It ? "t " "
Polv ala I u''l'.
') r-, ;. "-.
Dow
nloa
ded
by g
uest
on
Feb
ruar
y 18
, 202
1

Proc. Natl. Acad. Sci. USA 88 (1991) 4497
These results are of interest with respect to questions aboutthe amount of genetic variation in populations of inbreedingplants, their potential for evolutionary change, and strategiesfor preserving genetic resources in such species, many ofwhich are close crop relatives. Since self-pollination (inparticular, uniparental reproduction) may more readily allowfor colonization from single propagules compared with out-crossing, self-pollination has been linked with the founding ofisolated populations and with population recovery followingdisturbance, a relationship known as Baker's law (34). Oneexpected consequence of such population bottlenecks isreduction in Ne (2, 35). Using an island model, Maruyama andKimura (36) have shown that when local extinction of pop-ulations and recolonization occur frequently, genetic diver-sity and Ne are greatly reduced. Following population recov-ery from such events, Ne may be much smaller than theapparent population size.
Selection may also play a role in increasing the variance ofNe among populations of inbreeding species. Strong selectionfor particular ecotypes in some, but not all, geographicregions of a species range would contribute to variation indiversity among populations. Inbreeding is known to facili-tate selection among homozygous lines (37), and, for exam-ple, in the case of A. barbata it has been argued that the lowdiversity valley or inland populations have been selected forgrowth and survival in semiarid conditions (10).One implication of these results is that the evolution of
self-fertilization need not be thought of as an "evolutionaryblind alley" (38). Instead, our results suggest that it is moreappropriate to view selfing species as composed of a numberof ancient and genetically diverse "source" populations ofrelatively large Ne that have given rise to a subset of marginal"sink" populations of limited genetic diversity and evolu-tionary potential. Our results thus provide a genetic confir-mation of Baker's law. Moreover, they suggest an extensionof Pulliam's (39) concept of source and sink populations froman ecological to an evolutionary time scale. Such a viewpointhas relevance to the conservation of genetic resources inplants. Many species that are related to crop plants areself-pollinating (e.g., A. barbata, A. canariensis, H. sponta-neum, L. pimpinellifolium). That such species consist of hotspots of genetic variation is important for the design ofsampling strategies intended to conserve a representativesample of their genetic diversity. Clearly such genetically richpopulations, when known, deserve priority in conservationprograms. For inbreeding species, a useful sampling strategywould perhaps begin by first conducting surveys of popula-tion diversity before making final decisions about how pop-ulations will be represented in germplasm collections (40).Such surveys may be less important in outbreeding species,where there is comparatively less variation among popula-tions in gene diversity and Ne.For some of the inbreeding species studied, there appears
to be a connection between low levels of diversity and Ne andthe type of site occupied. For example, in A. barbata, inlandpopulations in both California and in Spain tend to be oflowerdiversity and Ne compared with coastal populations (10, 11).In H. spontaneum, populations occupying high elevationshad the lowest diversity and Ne values, while in L. pimpinel-lifolium, the smallest populations sampled corresponded tothose of lowest diversity and Ne. Recent signs of coloniza-tion, such as location along roadsides, also tend to correlatewith reduced diversity in some instances, as seen in A.barbata (41) and P. vulgaris (18). Unfortunately, there areother species in which ecological predictors of genetic diver-sity and Ne levels are lacking. For example, no clear corre-lates have emerged from the studies of A. thaliana, A.
canariensis, P. cuspidata, and P. major. In such cases,information about the genetic diversity of individual popu-lations, as derived directly from isozyme surveys, may becritical in guiding germplasm collections.
This paper is dedicated to Herbert G. Baker. We thank J. J.Burdon, 0. H. Frankel, R. H. Groves, D. R. Marshall, G. F. Mo-ran, R. N. Oram, and J. G. West for comments on the manuscript.D.J.S. acknowledges support from the Natural Sciences and Engi-neering Research Council of Canada and from the CommonwealthScience and Industry Research Organization, Plant Industry Divi-sion.
1. Nei, M. (1973) Proc. Natl. Acad. Sci. USA 70, 3321-3323.2. Wright, S. (1969) Evolution and Genetics of Populations, The
Theory of Gene Frequencies (Univ. Chicago Press, Chicago), Vol.2.
3. Hamrick, J. L. & Godt, M. J. W. (1990) in Plant Population Ge-netics, Breeding and Genetic Resources, eds. Brown, A. H. D.,Clegg, M. T., Kahler, A. L. & Weir, B. S. (Sinauer, Sunderland,MA), pp. 42-63.
4. Brown, A. H. D. (1978) Theor. Appl. Genet. 52, 145-157.5. Clegg, M. T. (1980) BioScience 30, 814-818.6. Fyfe, J. L. & Bailey, N. T. J. (1951) J. Agric. Sci. 41, 371-378.7. Abbott, R. J. & Gomes, M. F. (1989) Heredity 62, 411-418.8. Hamrick, J. L. & Allard, R. W. (1972) Proc. Natl. Acad. Sci. USA
69, 2100-2104.9. Marshall, D. R. & Allard, R. W. (1970) Genetics 66, 393-399.
10. Clegg, M. T. & Allard, R. W. (1972) Proc. Natl. Acad. Sci. USA 69,1820-1824.
11. Jain, S. K. & Singh, R. S. (1979) Bot. Gaz. (Chicago) 140, 256-362.12. Morikawa, T. & Leggett, J. M. (1990) Heredity 64, 403-411.13. Brown, A. H. D., Zohary, D. & Nevo, E. (1978) Heredity 41,
49-62.14. Nevo, E., Zohary, D., Brown, A. H. D. & Haber, M. (1979)
Evolution 33, 815-833.15. Rick, C., Fobes, J. F. & Holle, M. (1977) Plant Syst. Evol. 127,
133-170.16. Levin, D. A. (1978) Evolution 32, 245-263.17. Van Dijk, H., Wolff, K. & DeVries, A. (1988) Theor. Appl. Genet.
75, 518-528.18. Lack, A. J. & Kay, Q. 0. N. (1988) Biol. J. Linn. Soc. 34, 119-147.19. Brown, A. H. D. & Burdon, J. J. (1983) Aust. J. Biol. Sci. 36,
503-509.20. Burdon, J. J., Jarosz, A. M. & Brown, A. H. D. (1988) Biol. J.
Linn. Soc. 34, 81-92.21. Wain, R. P. (1982) Am. J. Bot. 69, 1573-1578.22. Lundkvist, K. (1979) Hereditas 90, 127-143.23. Lundkvist, K. & Rudin, D. (1977) Hereditas 85, 67-74.24. Stem, K. & Roche, L. (1974) Genetics of Forest Ecosystems
(Springer, Berlin).25. Gullberg, U., Yazdani, R., Rudin, D. & Ryman, N. (1985) Silvae
Genet. 34, 193-199.26. Shaw, D. V., Kahler, A. L. & Allard, R. W. (1981) Proc. Natl.
Acad. Sci. USA 78,1298-1302.27. Yeh, F. C. & O'Malley, D. 0. (1980) Silvae Genet. 29, 83-92.28. Gottlieb, L. D. (1975) Evolution 29, 213-225.29. Chakraborty, R. & Neel, J. V. (1989) Proc. Natl. Acad. Sci. USA
86, 9407-9411.30. Kimura, M. & Crow, J. F. (1964) Genetics 49, 725-738.31. Ewens, W. J. (1972) Theor. Popul. Biol. 3, 87-112.32. David, F. N. & Barton, D. E. (1962) Combinatorial Choice (Griffin,
London).33. Abramowitz, M. & Stegin, I. A. (1972) Handbook ofMathematical
Functions (Dover, New York).34. Baker, H. G. (1955) Evolution 9, 347-349.35. Nei, M., Maruyama, T. & Chakraborty, R. (1975) Evolution 29,
1-10.36. Maruyama, T. & Kimura, M. (1980) Proc. Natl. Acad. Sci. USA 77,
6710-6714.37. Haldane, J. B. S. (1924) Proc. Cambridge Philos. Soc. 1, 158-163.38. Stebbins, G. L. (1957) Am. Nat. 91, 337-354.39. Pulliam, H. R. (1988) Am. Nat. 132, 652-661.40. Jain, S. K. (1975) in Crop Genetic Resources for Today and To-
morrow, eds. Frankel, 0. H. & Hawkes, J. G. (Cambridge Univ.Press, Cambridge), pp. 15-36.
41. Jain, S. K., Rai, K. N. & Singh, R. S. (1981) Genetica 56,213-215.
Population Biology: Schoen and Brown
Dow
nloa
ded
by g
uest
on
Feb
ruar
y 18
, 202
1