Embed Size (px)
Citation preview

THE JOURNAL OF BIOLOGICAL CHEM~STRV
Vol 252, No. 11, Issue of June 10, pp. 3995-4001, 197’7
Prmted in U S.A.
Intestinal Basement Membrane of Ascaris suum ANALYSIS OF POLYPEPTIDE COMPONENTS*
(Received for publication, October 4, 1976)
CHUNG-HO HUNG, MIKIO OHNO, J. WILLIAM FREYTAG, AND BILLY G. HUDSON+
From the Department of Biochemistry, University of Kansas Medical Center, Kansas City, Kansas 66103
The Ascaris suum intestinal basement membrane was sol-
ubilized on reduction with P-mercaptoethanol in 1% sodium
dodecyl sulfate and 1% sodium dodecyl sulfate, 8 M urea to
the extent of 90% and lOO%, respectively. The reduced mem-
brane consists of at least 17 polypeptides ranging in molecu-
lar weight from 22,500 to greater than 400,000, as deter-
mined by sodium dodecyl sulfate-polyacrylamide gel elec-
trophoresis and gel filtration chromatography using 6 M urea
as eluent. Six of these polypeptides contain carbohydrate, as
determined by periodic acid-Schiff stain. Certain polypep-
tides, representing about 8% of the membrane by weight, are
soluble in sodium dodecyl sulfate solutions without reduc-
tion and are not interlinked with each other by disulfide
bonds; these interact in the membrane exclusively through
noncovalent forces. Amino acid and carbohydrate analyses
of the column fractions revealed that certain polypeptides
are collagen-like, being greatly enriched in 4-hydroxypro-
line, hydroxylysine and glycine, and disaccharide units;
whereas others are less collagen-like, being characterized by
the absence of 4-hydroxyproline, low contents of hydroxyly-
sine and glycine, and large contents of glutamic acid and
proline, and enriched in oligosaccharide units. These results
extend the observation from vertebrates to invertebrates
that basement membranes consist of multiple polypeptides
which have compositions of varying degrees of relatedness
to collagen, suggesting a conservation of this structural
characteristic during evolutionary development.
The intestinal basement membrane of the helminth Ascaris suum is classified as a specialized member of the collagen
family of proteins. It exhibits a wide angle x-ray diffraction
pattern characteristic of collagen (2) and contains appreciable
amounts of the amino acids characteristic of collagen (i.e.
hydroxyproline, hydroxylysine, and glycine) (3). The Ascaris membrane is related in amino acid and carbohydrate composi-
tion to that of vertebrate membranes, but it is distinctly
different. These differences are due to a decrease in the con-
tent of hydroxyproline, hydroxylysine, and glycine; an in-
crease in acidic amino acids; and the absence of S-hydroxypro-
line and sialic acid in the Ascaris membrane as compared to
* This work was supported by Grant AM18381 from the National Institutes of Health. A preliminary report of this work was published
(1). $ To whom all correspondence should be addressed.
vertebrate membranes, such as renal glomerular basement
membrane (3).
No information is available regarding the number, molecu-
lar weight, composition, and nature of the interaction of the
polypeptide components of this invertebrate basement mem-
brane. Information of this type has been reported for bovine
renal glomerular basement membrane and shows that it con-
sists of many polypeptide components of varying molecular
weight and composition (4-9) and that certain of these poly-
peptides are cross-linked by disulfide bonds (4, 10) and alde-
hyde-derived collagen cross-links (11).
The purpose of the present research concerning the Ascaris basement membrane was to determine the overall polypeptide
composition, the molecular weight of the polypeptides, and
their mode of interaction; and further, to determine which
polypeptides contain carbohydrate and which contain the
amino acids characteristic of collagen. In a succeeding paper
the chemical nature of the carbohydrate units is reported (12).
EXPERIMENTAL PROCEDURES
Materinls
All electrophoresis chemicals were obtained from Bio-Rad. Schiff s reagent was obtained from Fisher Scientific and P-mercaptoethanol from Eastman. P-Galactosidase, bovine serum albumin, ovalbumin, chymotrypsinogen, calf skin collagen, iPr,PF,l MalNEt, EDTA, and urea were obtained from Sigma. Myosin was prepared by the method of Perry (13). All other chemicals were the best commercially availa- ble and used without further purification.
Preparation of Basement Membrane
The membrane was prepared by the method previously described (3) and by a modification of this method. This modification involves (a) placing the dissected intestines directly into an ice-cold solution of 0.85% NaCl containing 10 rnM EDTA, 10 rnM iPr,PF, 25 rnM MalNEt followed by sonic disruption in this solution; and (6) wash- ing the insoluble membrane released on sonication with the same ice-cold solutions.
Electrophoresis
Two systems of sodium dodecyl sulfate-polyacrylamide gel electro- phoresis were employed for the determination of membrane polypep- tide composition and molecular weight.
Systmn 1 - Electrophoresis was performed essentially as previ- ously described (4) on 5% acrylamide gels (9 cm) containing 0.1% sodium dodecyl sulfate. Gel columns were stored at room tempera- ture for 12 to 16 h and prerun for 30 min prior to the application of
’ The abbreviations used are: iPr,PF, diisopropyl fluorophosphate; MalNEt, N-ethylmaleimide; PAS, periodic acid-Schiff; TEMED, N,N,N’,N’-tetramethylethylene diamine.
3995
by guest on Novem
ber 11, 2018http://w
ww
.jbc.org/D
ownloaded from

3996 Basement Membrane Pol.ypeptides
protein samples. Electrophoresis was performed with a current of 8 mA/tube for 4l12 h, at which time the tracking dye had migrated about 8.5 cm.
Gels were stained for protein by fixing in 40% isopropyl alcohol for 1 h and then staining in a solution of 0.05% Coomassie blue, 10% acetic acid, 10% isopropyl alcohol for 6 h. Gels were then washed with several changes of 10% acetic acid over a period of 2 to 3 days. Duplicate gels were stained for carbohydrate using PAS reagents as described by Fairbanks et nl. (14). Spectrophotometric scanning of the Coomassie blue-stained gels and the PAS-stained gels was per- formed at 650 nm and 550 nm, respectively, using a Gilford spectro- photometer equipped with a scanning accessory.
Samples of membrane were prepared for electrophoresis by incu- bating 3 mg of membrane in 1 ml of 1% sodium dodecyl sulfate, 2% p- mercaptoethanol, 0.1 M phosphate buffer (pH 7.0) overnight at 37” The insoluble material was removed by centrifugation and the su- pernatant solution adjusted to 15% (v/v) glycerol, and then 25 ~1 of 0.05% bromphenol blue was added. Aliquots (10 to 50 ~1) of this solution were then applied to gel columns.
System 2 - Electrophoresis was performed on 5% acrylamide gels as described for System 1 except that 8 M urea was included in the gel columns and membrane samples. In order to obtain good polymeriza- tion of gels, freshly prepared urea solutions (no longer than 3 h after preparation) were absolutely required. After the addition of urea to ihe acrylamide solution, the pH was readjusted to 7.0 with concen- trated phosphoric acid and the mixture was deaerated. Conditions for electrophoresis and staining procedures were identical to those described in System 1 above.
For the molecular weight determination of membrane polypep- tides observed by electrophoresis using Systems 1 and 2, the posi- tions of the protein bands were measured and their mobilities were calculated relative to the mobility of bromphenol blue. Their appar- ent molecular weights were estimated by comparison of relative mobility with a calibration line of log molecular weight versus rela- tive mobility. A calibration line was constructed for both Systems 1 and 2 using the following standard proteins of known molecular weights: myosin (212,000) (151, P-galactosidase (135,000) (161, bovine serum albumin (69,000) (171, ovalbumin (43,000) (181, and chymo- trypsinogen (25,700) (17).
Soluhility ofBasement Membrane
Membrane suspension (1 mgiml) was incubated in 1% sodium dodecyl sulfate, 0.1 M phosphate, pH 7.0, with and without 2% p- mercaptoethanol overnight at 37” with shaking. The insoluble mate- rial was removed by centrifugation and several aliquots of the super- natant solution were analyzed for protein using the method of Geiger and Bessman (19). Bovine serum albumin was used as a reference to construct a standard curve. Solubility of membranes in 8 M urea, 1% sodium dodecyl sulfate, 0.1 M phosphate, pH 7.0, with and without p- mercaptoethanol was also determined by this same procedure.
Gel Filtration Chromatography ofReduced Membrane
Sepharose CL-4B was packed to a height of 85 cm in a Pharmacia K261100 glass column (2.6 x 100 cm). The column was equilibrated with deaerated and nitrogen-flushed 6 M urea, 0.1% P-mercaptoetha- nol, 0.1 M PO,, pH 6.0. The membrane sample for column fractiona- tion was prepared by incubating 55 mg of whole membrane in 11.0 ml of deareated 8 M urea, 0.1 M PO, (pH 6.01, 4% P-mercaptoethanol for 24 h at 37” with constant shaking under a nitrogen barrier. The sample (10 ml) was diluted to 6 M urea prior to application to the column and eluted with equilibrating buffer under a hydrostatic pressure head of 25 cm at a flow rate of 10 ml/h. Fractions of 2.2 ml were collected. The protein content of the column fractions was measured as a function of the absorbance at 650 nm, using the calorimetric procedure of Geiger and Bessman (191, and by measure- ment of the absorbance at 280 nm. The polypeptides in the column fractions were identified by sodium dodecyl sulfate-polyacrylamide gel electrophoresis. The amount of a given polypeptide in a fraction, measured in terms of its absorbance at 650 nm, was calculated by multiplying the absorbance value for total protein content by the relative amount (%) of that polypeptide in the fraction. The relative amount was determined from the electrophoresis results in which the polyacrylamide gels were scanned spectrophotometrically and the relative peak areas measured from the recording.
Fractions containing certain polypeptides were pooled, dialyzed exhaustively against distilled water at 4”, lyophilized, and analyzed for amino acids and neutral monosaccharides. Fractions eluting
before void volume of column and after column volume were treated in the same manner and used as controls in carbohydrate determina- tions. Electrophoresis was conducted by incubating 50 ~1 of sample fractions with 50 ~1 of 2% sodium dodecyl sulfate, 2% P-mercaptoeth- anal, 0.1 M PO, (pH 7.0) overnight at 27”. All 100 pl of sample was applied to 5% polyacrylamide gels as previously described (System 1).
The column was calibrated with S-carboxymethylated standard proteins using the same experimental conditions. Serum albumin (5 mg), ovalbumin (4 mg), and acid-soluble calf skin collagen (10 mg) were reduced and alkylated in 10 ml of 8 M urea, 0.5 M Tris, pH 8.5, by the method of Mann and Fish (20). The molecular weights of collagen components are: u chain 95,000 (21, 22), p component (di- mer, 190,000), and y component (trimer, 285,000).
Chemical Analyses
For amino acid analyses, samples were hydrolyzed with glass- distilled 6 N HCl under reduced pressure in sealed tubes at 105” for 24 h. The amino acid contents of the hydrolysates were determined on a Beckman 121 HP analyzer by the procedure of Guire et al. (23).
Neutral monosaccharides were identified and quantitated as alditol acetates by gas-liquid chromatography essentially as described by Kim et al. (241, but a different column packing was used. Sugars were released from the protein by hydrolysis with Dowex 50-X12 resin and 0.25 N H,SO, for 24 h at loo”, then reduced and acetylated. The chromatography was performed on a single column (2 mm x 6 ft) of 3% SP2340 (Supelco) on Supelcoport (80 to 120 mesh) with nitrogen as carrier gas at a flow rate of 20 mlimin in a Hewlett-Packard 5931A gas chromatograph. Temperature was programmed from 180 to 225” at a rate of l”/min. This column packing is far more stable than the 3% ECNSS-M customarily used in this analysis and gives excellent resolution of alditol acetates.
RESULTS
Solubility - The membrane is soluble to the extent of 8% in
1% sodium dodecyl sulfate, 0.1 M phosphate buffer (pH 7.0)
with or without 8 M urea. On reduction with 2% P-mercapto-
ethanol, its solubility is increased to 90% in this buffer without
urea and to 100% with 8 M urea.
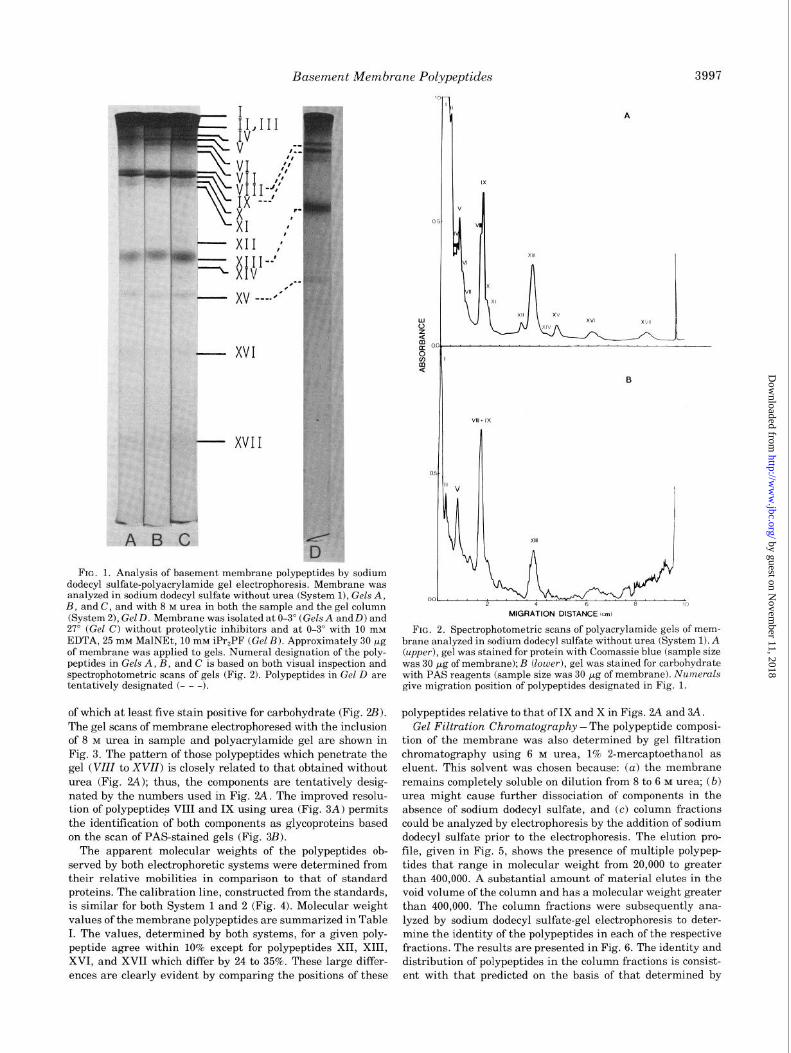
Electrophoresis ~ Membrane was analyzed by polyacryl-
amide gel electrophoresis using a sodium dodecyl sulfate/phos-
phate buffer (System 1). It consists of several polypeptide
components as shown by the electrophoretic pattern presented
in Fig. 1, Gel A. To assess whether this multiplicity of compo-
nents was generated artifactually by proteolytic degradation
during the isolation procedure, the membrane was isolated
with and without proteolytic inhibitors under conditions used
previously for glomerular basement membrane (9). The pat-
tern for membrane prepared at O-3” in the absence of exoge-
nous protease inhibitors is identical to that of membrane
prepared at O-3” using inhibitors (EDTA, MalNEt, and
iPr,PF), Gel B, and at 27” without these inhibitors, Gel C.
Since the solubility of the membrane is increased from 90 to
100% in the sodium dodecyl sulfate buffer with the inclusion of
8 M urea, its total polypeptide constituents could be analyzed
by sodium dodecyl sulfate electrophoresis by using 8 M urea in
both sample and polyacrylamide gels (System 2). Also, the
inclusion of urea would be expected to cause dissociation of
components which might be incompletely dissociated in so-
dium dodecyl sulfate alone. As shown in Gel D, Fig. 1, the
pattern with urea is basically the same as that without urea
(Gels A to C), except that mobilities are reduced and resolu-
tion of two polypeptides (VZZZ and IX) is greatly improved.
Polyacrylamide gels of membrane electrophoresed using
System 1 and System 2 were scanned spectrophotometrically
to detect precisely polypeptide components and to identify
those which contain carbohydrate. The gel scans of membrane
electrophoresed without urea are shown in Fig. 2. At least 17
polypeptides were detected by Coomassie blue stain (Fig. 24 1,
by guest on Novem
ber 11, 2018http://w
ww
.jbc.org/D
ownloaded from
Basement Membrane Polypeptides 3997
FIG. 1. Analysis of basement membrane polypeptides by sodium dodecyl sulfate-polyacrylamide gel electrophoresis. Membrane was analyzed in sodium dodecyl sulfate without urea (System 11, Gels A, B, and C, and with 8 M urea in both the sample and the gel column (System 21, Gel D. Membrane was isolated at O-3” (Gels A andD) and 27” (Gel C) without proteolytic inhibitors and at O-3” with 10 rnM EDTA, 25 rnM MalNEt, 10 rnM iPr,PF (Gel B). Approximately 30 Fg of membrane was applied to gels. Numeral designation of the poly- peptides in Gels A, B, and C is based on both visual inspection and spectrophotometric scans of gels (Fig. 2). Polypeptides in Gel D are tentatively designated (- - -).
of which at least five stain positive for carbohydrate (Fig. 2Z3).
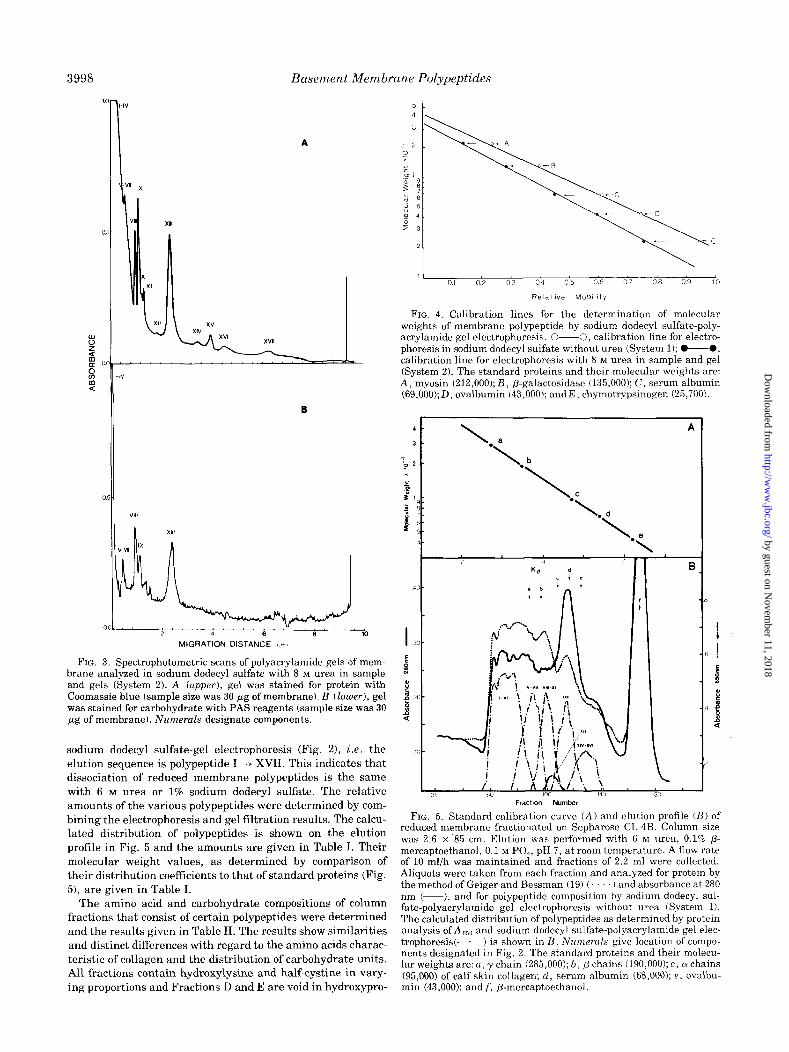
The gel scans of membrane electrophoresed with the inclusion
of 8 M urea in sample and polyacrylamide gel are shown in
Fig. 3. The pattern of those polypeptides which penetrate the
gel (VZZZ to XVZZ) is closely related to that obtained without
urea (Fig. 2A); thus, the components are tentatively desig-
nated by the numbers used in Fig. 2A. The improved resolu-
tion of polypeptideg VIII and IX using urea (Fig. 3A) permits
the identification of both components as glycoproteins based
on the scan of PAS-stained gels (Fig. 3B).
The apparent molecular weights of the polypeptides ob-
served by both electrophoretic systems were determined from
their relative mobilities in comparison to that of standard
proteins. The calibration line, constructed from the standards,
is similar for both System 1 and 2 (Fig. 4). Molecular weight
values of the membrane polypeptides are summarized in Table
I. The values, determined by both systems, for a given poly-
peptide agree within 10% except for polypeptides XII, XIII,
XVI, and XVII which differ by 24 to 35%. These large differ-
ences are clearly evident by comparing the positions of these
ool-
A
B
MIGRATION DISTANCE fcml
FIG. 2. Spectrophotometric scans of polyacrylamide gels of mem- brane analyzed in sodium dodecyl sulfate without urea (System 1). A (upper), gel was stained for protein with Coomassie blue (sample size was 30 pg of membrane); B (lower), gel was stained for carbohydrate with PAS reagents (sample size was 30 pg of membrane). Numerals give migration position of polypeptides designated in Fig. 1.
polypeptides relative to that of IX and X in Figs. 2A and 3A.
Gel Filtration Chromatography - The polypeptide composi-
tion of the membrane was also determined by gel filtration
chromatography using 6 M urea, 1% 2-mercaptoethanol as
eluent. This solvent was chosen because: (a) the membrane
remains completely soluble on dilution from 8 to 6 M urea; (b)
urea might cause further dissociation of components in the
absence of sodium dodecyl sulfate, and (c) column fractions
could be analyzed by electrophoresis by the addition of sodium
dodecyl sulfate prior to the electrophoresis. The elution pro-
file, given in Fig. 5, shows the presence of multiple polypep-
tides that range in molecular weight from 20,000 to greater
than 400,000. A substantial amount of material elutes in the
void volume of the column and has a molecular weight greater
than 400,000. The column fractions were subsequently ana-
lyzed by sodium dodecyl sulfate-gel electrophoresis to deter-
mine the identity of the polypeptides in each of the respective
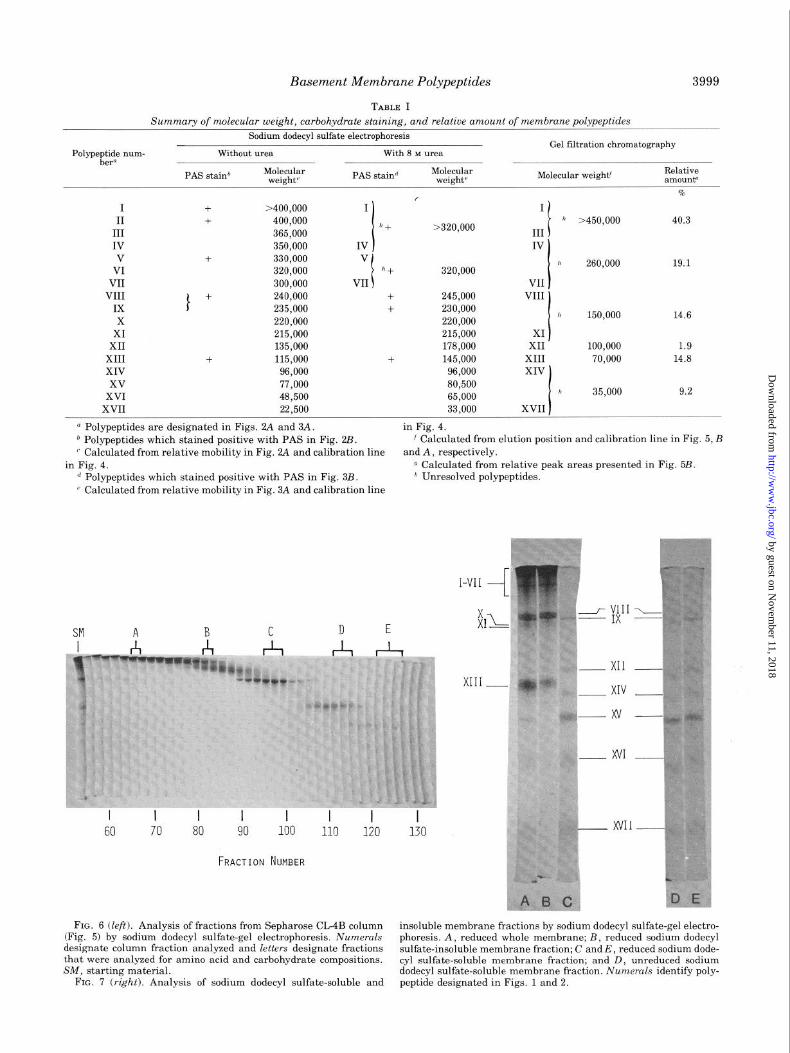
fractions. The results are presented in Fig. 6. The identity and
distribution of polypeptides in the column fractions is consist-
ent with that predicted on the basis of that determined by
by guest on Novem
ber 11, 2018http://w
ww
.jbc.org/D
ownloaded from
Basement Membrane Polypeptides
B
OOL- ”
2 4 6 8 10
MIGRATION DISTANCE r<rn,
FIG. 3. Spectrophotometric scans of polyacrylamide gels of mem- brane analyzed in sodium dodecyl sulfate with 8 M urea in sample and gels (System 2). A (upper), gel was stained for protein with Coomassie blue (sample size was 30 pg of membrane). B (loluer), gel was stained for carbohydrate with PAS reagents (sample size was 30 pg of membrane). Numerals designate components.
sodium dodecyl sulfate-gel electropnoresis (Fig. 21, i.e. the
elution sequence is polypeptide I ---) XVII. This indicates that
dissociation of reduced membrane polypeptides is the same
with 6 M urea or 1% sodium dodecyl sulfate. The relative
amounts of the various polypeptides were determined by com-
bining the electrophoresis and gel filtration results. The calcu-
lated distribution of polypeptides is shown on the elution
profile in Fig. 5 and the amounts are given in Table I. Their
molecular weight values, as determined by comparison of
their distribution coefficients to that of standard proteins (Fig.
51, are given in Table I.
The amino acid and carbohydrate compositions of column
fractions that consist of certain polypeptides were determined
and the results given in Table II. The results show similarities
and distinct differences with regard to the amino acids charac-
teristic of collagen and the distribution of carbohydrate units.
All fractions contain hydroxylysine and half-cystine in vary-
ing proportions and Fractions D and E are void in hydroxypro-
1 01 02 03 04 05 06 07 08 09 10
Relattve MoblIlly
FIG. 4. Calibration lines for the determination of molecular weights of membrane polypeptide by sodium dodecyl sulfate-poly- acrylamide gel electrophoresis. O-0, calibration line for electro- phoresis in sodium dodecyl sulfate without urea (System 1); O-0, calibration line for electrophoresis with 8 M urea in sample and gel (System 2). The standard proteins and their molecular weights are: A, myosin (212,000); B, P-galactosidase (135,000); C, serum albumin (69,000); D, ovalbumin (43,000); and E, chymotrypsinogen (25,700).
1.. A
Lb
\
‘L. d
\
‘T
-2
Fraction Number
FIG. 5. Standard calibration curve (A) and elution profile (B) of reduced membrane fractionated on Sepharose CL4B. Column size was 2.6 x 85 cm. Elution was performed with 6 M urea, 0.1% /3- mercaptoethanol, 0.1 M PO,, pH 7, at room temperature. A flow rate of 10 ml/h was maintained and fractions of 2.2 ml were collected. Aliquots were taken from each fraction and analyzed for protein by the method of Geiger and Bessman (19) (. ) and absorbance at 280 nm (-), and for polypeptide composition by sodium dodecyl sul- fate-polyacrylamide gel electrophoresis without urea (System 1). The calculated distribution of polypeptides as determined by protein analysis of A (i5,, and sodium dodecyl sulfate-polyacrylamide gel elec- trophoresis(-.-) is shown in B. Numerals give location of compo- nents designated in Fig. 2. The standard proteins and their molecu- lar weights are: a, y chain (285,000); b, p chains (190,000); c, 01 chains (95,000) of calf skin collagen; d, serum albumin (68,000); e, ovalbu- min (43,000); and f, P-mercaptoethanol.
r B
by guest on Novem
ber 11, 2018http://w
ww
.jbc.org/D
ownloaded from
Basement Membrane Polypeptides 3999
TABLE I
Summary of molecular weight, carbohydrate staining, and relative amount of membrane polypeptides
Sodium dodecyl sulfate electrophoresis Gel filtration chromatography
Polypqi$ *urn- Without urea With 8 M urea (I
I - -
PAS stain” Molecular weight’ PAS staind Molecular
weight’ Molecular weight’ Relative amountu
I II
III
IV V
VI VII
VIII IX
X
XI XII
XIII XIV
+ >400,000 + 400,000
365,000
350,000 + 330,000
320,000 300,000
I + 240,000 235,000
220,000
215,000 135,000
+ 115,000 96,000
r I
” + >320,000
IV
*+ 320,000
+ 245,000 + 230,000
220,000
215,000 178,000
+ 145,000 96,000
I
t
h >450,000 40.3
III
IV
I
R 260,000 19.1
VII VIII
h 150,000 14.6
XI XII 100,000 1.9
XIII 70,000 14.8 XIV \
xv 77,000 80,500 XVI 48,500 65,000 h 35,000 9.2
XVII 22,500 33,000 XVII
o Polypeptides are designated in Figs. 2A and 3A. in Fig. 4. b Polypeptides which stained positive with PAS in Fig. 2B. f Calculated from elution position and calibration line in Fig. 5, B
(- Calculated from relative mobility in Fig. 2A and calibration line and A, respectively.
in Fig. 4. o Calculated from relative peak areas presented in Fig. 5B.
d Polypeptides which stained positive with PAS in Fig. 3B. h Unresolved polypeptides.
( Calculated from relative mobility in Fig. 3A and calibration line
SM A B C n E x
I I I I I I I 60 70 80 90 100 110 120
FRACTION NUMBER
D E
FIG. 6 (left). Analysis of fractions from Sepharose CL-4B column insoluble membrane fractions by sodium dodecyl sulfate-gel electro- (Fig. 5) by sodium dodecyl sulfate-gel electrophoresis. Numerals phoresis. A, reduced whole membrane; B, reduced sodium dodecyl designate column fraction analyzed and letters designate fractions sulfate-insoluble membrane fraction; C and E, reduced sodium dode- that were analyzed for amino acid and carbohydrate compositions. cyl sulfate-soluble membrane fraction; and D, unreduced sodium SM, starting material. dodecyl sulfate-soluble membrane fraction. Numerals identify poly-
FIG. 7 (right). Analysis of sodium dodecyl sulfate-soluble and peptide designated in Figs. 1 and 2.
by guest on Novem
ber 11, 2018http://w
ww
.jbc.org/D
ownloaded from

4000 Basement Membrane Pol.ypeptides
line. Fraction C, consisting mainly of polypeptides VIII and
IX, is most collagen-like in that it has the largest proportion of
hydroxyproline, hydroxylysine, and glycine. In contrast, Frac-
tion D, consisting of mainly XIII, and E, consisting mainly of
XIV to XVII, are least collagen-like in that they contain the
least amount of glycine and hydroxylysine, are void of hydrox-
yproline, and contain the largest proportion of glutamic acid
and proline. Both the disaccharide and oligosaccharide units of
the membrane (12) are present in all fractions, but in varying
amounts; the relative amount of disaccharide unit to oligosac-
charide unit is largest in Fraction C and lowest in Fraction E.
Mode oflnteraction between Polypeptides - The partial solu-
bility of the membrane, 8% by weight, in sodium dodecyl
sulfate in the absence of a reducing agent clearly represents a
separation of the membrane into two fractions, sodium dodecyl
sulfate-soluble and -insoluble fractions, which are held to-
gether exclusively by noncovalent forces in the native mem-
TABLE II
Amino acid and carboh,ydrate composition of column fractions
component
Amino acid 4-Hydroxy-
proline”
Aspartic acid Threonine
Serine
Glutamic acid Proline
Glycine
Alanine
Half-cystine
Valine
Methionine
Isoleucine
Leucine
Tyrosine
Phenylalanine
Hydroxylysine
Lysine
Histidine
Arginine
Monosaccharide
Fucose
Mannose Gala&se
Glucose’
Disaccharideioligo-
saccharide”
23.8 18.2 43.8
87.9 101.8 63.6
50.8 57.3 32.9
55.9 43.4 42.9
115.4 127.7 111.4
63.5 93.9 98.9
165.9 150.6 256.9
66.7 68.9 73.2
5.4 8.5 6.4 44.6 49.6 32.5
3.0 5.5 2.3
31.6 34.7 27.0
66.3 61.1 65.7
21.0 23.9 18.8
22.2 19.7 19.8
17.6 10.4 18.4
33.5 25.8 17.8
28.5 23.7 14.9
96.2 75.2 52.7
3.4 3.3 1.8
4.6 3.7 2.6
18.2 11.8 16.7
13.3 8.9 15.6
2.911 2.411 ‘31 1.611 0.511’
Column fraction”
A B C D E
reszduellOO0 amino acid rescdues
0 0
61.7 69.0
51.7 43.5
79.3 77.4
174.7 146.5
131.7 122.1
134.2 101.1
59.1 62.5
34.4 35.3
44.5 38.0
4.6 8.7
21.8 23.4
37.6 38.5
28.4 25.7
12.6 19.6
4.6 3.4
16.4 38.0
34.8 46.2
67.6 100.8
2.0 7.1
2.0 7.5
5.0 9.0
3.1 - 11
” Column fractions are those designated in Fig. 6. Their polypep-
tide compositions are: A, >99% I; B, 38% II + III and 62% IV-VII; C,
75% VIII + IX and 25% X-XII; D, 93% XIII and 7% XIV-XVII; E, 12%
XIII and 88% XIV-XVII, as determined from spectrophotometric
scans of gels in Fig. 6.
’ 3-Hydroxyproline is absent in all fractions.
’ Values are corrected for the amount of monosaccharide that
arises from column packing.
” Value is undetermined because value was small in comparison
to the correction value for the amount of glucose arising from column
packing.
” Values were calculated on the basis of 1 glucose residue per
disaccharide unit and 1 mannose residue per oligosaccharide unit
(12). ’ Value calculated assuming glucose content is equal to hydroxyly-
sine content.
brane. The identity of the polypeptides comprising these two
membrane fractions was determined by sodium dodecyl sul-
fate-polyacrylamide gel electrophoresis. As shown in Fig. 7,
the sodium dodecyl sulfate-insoluble fraction consists of only
12 (I to XI, XIII) of the 17 polypeptides found in the whole
membrane (compare Gel A with R); whereas, the soluble
fraction consists of the remaining 5 polypeptides (XII, XIV to
XVII) and is devoid of all those comprising the insoluble
fraction except for a proportion of polypeptides VIII and IX
(compare Gels A and C).
The polypeptides comprising the sodium dodecyl sulfate-
insoluble fraction appear to be held together in part by inter-
chain disulfide bonds because this fraction is only soluble in
sodium dodecyl sulfate on reduction with 2-mercaptoethanol.
In contrast, those in the sodium dodecyl sulfate-soluble frac-
tion are not cross-linked by disulfide bonds because the pat-
terns (Fig:. 7) obtained by electrophoresis of the unreduced
(Gel E) and reduced (Gel D) fractions are very similar.
Thus, polypeptides VIII, IX, XII, and XIV to XVII compris-
ing the sodium dodecyl sulfate-soluble fraction, are held to-
gether with polypeptides I to XI and XIII, comprising the
sodium dodecyl sulfate-insoluble fraction, exclusively through
noncovalent interactions. Certain, if not all, of the polypep-
tides in the sodium dodecyl sulfate-insoluble fraction are held
together by interchain disulfide bonds.
DISCUSSION
The Ascaris intestinal basement membrane is solubilized on
reduction with P-mercaptoethanol in 1% sodium dodecyl sul-
fate and 1% sodium dodecyl sulfate, 8 M urea to the extent of
90% and lOO%, respectively. Determination of its polypeptide
composition and molecular weight by sodium dodecyl sulfate-
polyacrylamide gel electrophoresis, with and without the in-
clusion of 8 M urea in the gel system, showed it to consist of at
least 17 polypeptides ranging in molecular weight from 22,500
to greater than 400,000. Six of these polypeptides contain
carbohydrate as determined by PAS stain. The molecular
weight values determined with and without urea in the sys-
tem agree within 10% for most of the polypeptides, but certain
of those of lower molecular weights (XII, XIII, XVI, and XVII)
differ by 24 to 35%. The reason for these differences is not clear
because standard proteins gave similar calibration lines with
and without urea.
The polypeptide composition and molecular weight were
also determined by gel filtration chromatography using 6 M
urea as eluent. The reduced membrane, being completely
soluble in this solvent, consists of multiple polypeptides rang-
ing in molecular weight from 20,000 to greater than 400,000
with approximately 40% of the material having a molecular
weight in excess of 400,000. A comparison of the distribution of
polypeptides obtained from the column profile to that obtained
by sodium dodecyl sulfate-gel electrophoresis indicates that
urea did not cause further dissociation of components than was
obtained with sodium dodecyl sulfate. This conclusion was also
borne out by the similar electrophoretic patterns obtained for
membrane electrophoresed with and without urea (Figs. 2 and
3). For certain polypeptides, such as VIII, IX, and XIII, the
molecular weight values determined by gel filtration chroma-
tography differ greatly from the values determined by sodium
dodecyl sulfate-polyacrylamide gel electrophoresis (Table I).
This suggests anomalous behavior of these polypeptides in
sodium dodecyl sulfate analogous to that observed for a poly-
peptide isolated from glomerular basement membrane (7) and
collagen chains (25).
by guest on Novem
ber 11, 2018http://w
ww
.jbc.org/D
ownloaded from

Basement Membrane Polypeptides 4001
Chemical analyses of column fractions that contain certain
polypeptides revealed that most, if not all, contain hydroxyly-
sine in varying proportions. Polypeptides, VIII or IX or both,
(M,. = 150,000) are the most collagen-like, being characterized
by t,he presence of the largest content of hydroxyproline, hy-
droxylysine, and glycine. In contrast, polypeptide XIII (Mr =
70,000) and possibly XIV to XVII are least collagen-like, being
characterized by the absence of hydroxyproline and the pres-
ence of the least content of hydroxylysine and glycine and the
largest of glutamic acid and proline. The disaccharide and
oligosaccharide units are constituents of both the collagen-like
(VIII and IX) and less collagen-like polypeptide (XIII), but the
proportion of the disaccharide unit relative to the oligosaccha-
ride unit is greater in the more collagen-like polypeptides.
The more collagen-like polypeptides appear to be linked
with the less collagen-like polypeptide (XIII) and probably to
the high molecular weight components (I to VIII) by disulfide
bonds in the native membrane. This conclusion is based on the
finding that these components, representing 92% of membrane
by weight, are soluble in denaturants, sodium dodecyl sulfate
and urea, only after reduction with P-mercaptoethanol. Cer-
tain of the membrane polypeptides (XII, XIV to XVII), repre-
senting the remainder of the weight, are soluble in sodium
dodecyl sulfate without reduction and are not interlinked with
each other by disulfide bonds; therefore, these interact in the
membrane exclusively through noncovalent forces. A substan-
tial amount of the reduced membrane (40%) exists as very
high molecular weight, (>400,000, components I to III) in both
sodium dodecyl sulfate and urea. These may represent aggre-
gates of the more collagen-like and less collagen-like peptides
that are covalently cross-linked by bonds other than disulfide,
such as the lysyl-derived cross-links present in vertebrate
collagen and basement membranes (11).
The multiplicity of components observed presumably re-
flects the true nature of the membrane in which each of these
polypeptides is a distinct integral membrane component. Al-
ternatively, the multiplicity could be partly due to the associa-
tion of nonmembrane polypeptides that are derived from cellu-
lar constituents during membrane isolation. In this event,
those few polypeptides that are soluble in sodium dodecyl
sulfate or urea without cleavage of disulfide bonds would be
the likely candidates (XII, XIV to XVII). The additional possi-
bility that the multiple polypeptides are generated artifac-
tually during membrane isolation by the action of tissue-
associated proteases is unlikely because the polypeptide com-
position of membrane prepared at O-3” or 27” without specific
enzyme inhibitors is identical to that membrane prepared at
O-3” in the presence of EDTA, iPr,PF, and MalNEt (Fig. 1). A
similar result was obtained with glomerular basement mem-
brane except that a single polypeptide was degraded in the
absence of iPr,PF (9).
The results of this study extend the observation from verte-
brates to invertebrates that basement membranes consist of
multiple polypeptides which have compositions of varying de-
grees of relatedness to collagen (less collagen-like to more
collagen-like) (5, 7, 8). This suggests a conservation of this
structural characteristic during evolutionary development.
Recent studies (7, 8) are consistent with the proposal (5, 26)
that the relatedness reflects the proportion of collagen-like to
non-collagen segments in each polypeptide. The identification
of multiple collagen components of varying sizes from pepsin
digests of vertebrate basement membrane (27-29) further sup-
ports this proposal.
The less collagen-like polypeptides may be related to the
polar extension peptide of procollagen, which has a small
amount of collagen-like sequence and large amounts of glu-
tamic acid (30). It is conceivable that these are derived biosyn-
thetically by cleavage of a large procollagen-like component
and are retained as part of the membrane structure. It is
possible that the function of these more polar polypeptides is to
interact with surface components of cells positioned onto the
basement membrane or serve in the alignment of the more
collagen-like polypeptides, or both.
1.
2.
3.
4.
5.
6.
7.
Hudson, B. G., Hung, C. H., and Peczon, B. D. (1976)Fed. Proc.
3, 1519 Rudall, K. M. (1955) Symp. Sot. Exp. Biol. 9, 49-71 Peczon, B. D., Venable, J. H., Beams, C. G., Jr., and Hudson, B.
G. (1975) Biochemistry 14, 4069-4075 Hudson, B. G., and Spiro, R. G. (1972) J. Biol. Chem. 247,4229-
4238
Hudson, B. G., and Spiro, R. G. (1972) J. Biol. Chem. 247, 4239- 4247
Myers, C., and Bartlett, P. (1972) Bzochim. Bioph,ys. Acta 290,
X0-157 Ohno, M., Riquetti, P., and Hudson, B. G. (1975)5. Biol. Chem.
250. 7780-7787
8. Sato, ‘T., and Spiro, R. G. (1976)J. Biol. Chen. 251, 4062-4070 9. Frevtae. J. W., Ohno. M.. and Hudson, B. G. (1976) Biochem.
10. 11.
12.
13. 14.
15.
16. 17. 18.
Btop&s. Res: Comvk. 72, 796-802
Kefalides, N. A. (1972) Corm. Tissue Res. 1, 3-13 Tanzer. M. L.. and Kefalides. N. A. (1973) Biochem. Bioohvs.
Res. &mm&. 51, 775-780 ’ I ”
Peczon, B. D., Wegener, L. J., Hung, C.-H., and Hudson, B. G. (1977) J. Biol. Chem. 252, 4002-4006
Perry. S. V. (1975) Methods Enzymol. 2, 582-588 Fairbanks, G., Steck, T. L., and Wallack, D. F. H. (1971) Bio-
chemistry 10, 2606-2617 Gersham, L. C., Stracker, A., and Dreizen, P. (1969) J. Biol.
Chem. 244, 2726-2736
Fowler. A. V.. andzabin. I. (197015. Biol. Chem. 245,5032-5041 Tanford, C. (i969)Adu. Protein Chem. 23, 121-282
Castellino, F. J., and Barker, R. (1968) Biochemistry 7, 2207- 2217
19.
20. 21.
22.
23.
24.
25. 26.
27.
28
29
30
Geiger, P. J., and Bessman, S. P. (1972)AnaZ. Biochem. 49,467- 473
Mann, K., and Fish, W. W. (1972) Methods Enzymol. 26,28-42 Kang, A. H.. Nagai, Y.. Piez, K. A.. and Gross, J. (1966) Bio-
chkistry g, 509-515
Lenaers, A., Ansay, M., Nusgens, B. V., and Lapiere, C. M. (1971) Eur. J. Biochem. 23, 533-543
Guire, P., Riquetti, P., and Hudson, B. G. (1974) J. Chromatog. 90, 350-353
Kim, E. H., Shome, B., Liao, T., and Pierce, J. G. (1967)Anal.
B&hem. 20. 258-274 Furthmayr, H., andTimp1, R. (1971)Anal. B&hem. 41,510-516 Spiro, R. G. (1973) N. Engl. J. Med. 288, 1337-1342
Daniels, J. R., and Chu, G. H. (1975) J. Biol. Chem. 250, 3531- 3537
Kefalides, N. A. (1971) B&hem. Biophys. Res. Commun. 45, 226-234
Chune. E.. Rhodes, R. K.. and Miller, E. J. (1976) B&hem. Bio,&ys. Res. Commun. 71, 1167-1174
Becker, J., Timpl, R., Helle, O., and Prockop, D. J. (1976)
Biochemistry 15, 2853-2862
REFERENCES
by guest on Novem
ber 11, 2018http://w
ww
.jbc.org/D
ownloaded from

C H Hung, M Ohno, J W Freytag and B G Hudsoncomponents.
Intestinal basement membrane of Ascaris suum. Analysis of polypeptide
1977, 252:3995-4001.J. Biol. Chem.
http://www.jbc.org/content/252/11/3995.citation
Access the most updated version of this article at
Alerts:
When a correction for this article is posted•
When this article is cited•
to choose from all of JBC's e-mail alertsClick here
http://www.jbc.org/content/252/11/3995.citation.full.html#ref-list-1
This article cites 0 references, 0 of which can be accessed free at
by guest on Novem
ber 11, 2018http://w
ww
.jbc.org/D
ownloaded from









![[PPT]OBSTRUCCION INTESTINAL - semio2013 | This … · Web viewOBSTRUCCION INTESTINAL OBSTRUCCION INTESTINAL OBSTACULO AL TRANSITO DEL CONTENIDO INTESTINAL Adinámico o paralítico](https://img.pdfslide.us/doc/110x75/5b36ceb57f8b9a4a728b5103/pptobstruccion-intestinal-semio2013-this-web-viewobstruccion-intestinal.jpg)