Embed Size (px)
Citation preview

Vol. 23, No. 1 1
VOLUME 23 NUMBER1 1998
InternationalRice Research
Notes
INTERNATIONAL RICE RESEARCH INSTITUTE

2 IRRN 1998
International Rice Research Notes
The International Rice Research Notes (IRRN) expeditescommunication among scientists concerned with thedevelopment of improved technology for rice and rice-based systems.
The IRRN is a mechanism to help scientists keep eachother informed of current rice research findings. Theconcise scientific notes are meant to encourage ricescientists to communicate with one another to obtaindetails on the research reported.
The IRRN is published three times a year in April,August, and December by the International RiceResearch Institute.
IRRN production team
Editor: Bill HardyAssistant editor: Tess RolaLayout and design: Erlie PutunganArtwork: Erlie Putungan
○ ○ ○ ○ ○ ○ ○ ○ ○ ○ ○ ○ ○ ○ ○
ContentsVol. 23, No. 1, 1998
Germplasm improvement
Genetic resourcesInternational testing of rice hybrids for yield and adaptability by
INGER: prospects and problems 4Screening rice hybrids for quality traits 4Studying comparative suitability of CMS lines 5Identifying a new long-duration CMS line (APMS 5A) for coastal
regions 5Wide hybridization for diversification of CMS in rice 5Krishna CMS lines in the background of four different cytoplasmic
sources in rice 6Characterizing thermosensitive genic male sterile lines of rice 6Identifying and characterizing thermosensitive cytoplasmic male
sterile lines 7Screening rice germplasm for floral attributes that influence
outcrossing 7
GeneticsPhysical localization of phyA and rbcS genes in rice 8Fertility restoration studies in four WA CMS lines of rice 9Effect of minor genes in restoration of fertility in CMS lines
of rice 9Breeding for male fertility restoration incorporating Basmati rice
quality 9Genetics of thermosensitive genic male sterile lines in rice 10Mitochondrial gene expression in cytoplasmic male sterile and
fertile lines of rice 10RAPD and PBR analysis of CMS maintainer and restorer lines
in rice 11Transferring wild abortive cytoplasmic male sterility through
asymmetric fusion of protoplasts 11
Breeding methods 11Adaptability and yield of rice hybrids 11Yield stability in rice hybrids 12Exploiting heterosis using TGMS lines in intra- and intersubspecific
hybridization 12Developing CMS and restorer lines of tropical rice hybrids 12Developing Pusa 3A, a Basmati CMS line 13Screening rice genotypes for thermosensitive genic male sterility
reaction 13Identifying parental lines in hybrid rice by RAPD fingerprinting 13Exploiting the in vitro ovary culture technique to breed rice
hybrids 14
Pest resistance—diseasesA variety with durable resistance to rice blast 14Multilocational evaluation of promising advanced breeding lines for
resistance to rice tungro viruses 15
Focus on hybrid riceAchieving self-sufficiency in rice production and maintainingprice stability are important in countries where rice providesfood security and generates employment and income forpeople. In the past three decades, most rice-growingcountries, particularly in Asia, have done remarkably well inmeeting their rice needs. But by 2030, the world must produce60% more rice than it produced in 1995 to meet the demandcreated by increasing population and rising income. Thisincrease must be achieved using less land, less labor, lesswater, and fewer pesticides, and it must be sustainable. Tomeet this challenge, increasing the yield potential of ricebeyond that of the semidwarf varieties is an importantstrategy.
Hybrid rice is a technology for meeting this challenge. Thistechnology has enabled China to increase its rice productionsignificantly during the past 20 years. IRRI, in collaborationwith several national agricultural research systems, hasdeveloped rice hybrids for the tropics and helped India,Vietnam, and the Philippines to begin commercializing them.By the 21st century, about 3 million hectares are expected to becovered with hybrid rice, which should produce about 3million tons of extra rice (worth $450 million) annually.
The Third International Symposium on Hybrid RiceResearch was held 14-16 November 1996, at the Directorate ofRice Research in Hyderabad, India. The key papers presentedare being published as an IRRI-Indian Council of AgriculturalResearch book. The posters displayed at the symposiumappear as notes (in a modified format) throughout this issueof the IRRN. They are denoted by the symbol.
We hope that you find these notes to be a valuable sourceof information.

Vol. 23, No. 1 3
Pest resistanceAdvanced breeding lines with resistance to rice tungro
viruses 17
Stress tolerance—adverse soilsSeasonal differences in iron toxicity tolerance of lowland rice
cultivars 18Correlation of some varietal characteristics with grain yield and
stress tolerance index under saline conditions 19
Integrated germplasm improvement—uplandAltamirano, Mandisovi, and Quebracho: new rice cultivars from
Argentina 20Four promising medium-duration upland rice breeding lines for
the République Démocratique du Congo 21
Seed technologyA seed production system for backstopping a hybrid rice
breeding program 22Standardizing hybrid rice seed production practices 22Evaluating hybrid seed production technology along with the
new growth hormone Mangiferin 23Synchronization studies in hybrid rice seed production 23Effect of seedling age on flowering time in A, B, and
R lines 23Storage potential of parental lines of hybrid rice 24Dormancy of seeds of some rice hybrids and their parents 24Significance of stigma exsertion in enhancing outcrossing in
male sterile rice lines 25
Crop and resource management
Fertilizer managementEffect of N levels and time of application on grain yield of hybrid
rice 25
Crop management 25Three-drying cultivation of rice: a new technology for rice produc-
tion in drought-prone areas 25Agronomic package for hybrid rice cultivation 27
Integrated germplasm improvement—weedsChemical weed control in rice nurseries of the hill zone of
Karnataka 27Weed control in dry-seeded lowland rice 28
Research methodology
The relationship between experimental plot research and Texasrice producers’ ratoon field results 29
Comparing methods of stability analysis in rice hybrids 30
Education and communication
Role of voluntary agencies in popularizing hybrid rice production inIndia 30
Socioeconomic impact
Economics of hybrid rice seed production in India 31Impact of hybrid rice on India’s food economy: implications and
policy issues 31
The International Rice Research Institute (IRRI) was established in 1960 by the Ford and Rockefeller Foundations with thehelp and approval of the Government of the Philippines. Today IRRI is one of 16 nonprofit international research centerssupported by the Consultative Group on International Agricultural Research (CGIAR). The CGIAR is cosponsored by the Foodand Agriculture Organization of the United Nations (FAO), the International Bank for Reconstruction and Development (WorldBank), the United Nations Development Programme (UNDP), and the United Nations Environment Programme (UNEP). Itsmembership comprises donor countries, international and regional organizations, and private foundations.
As listed in its most recent Corporate Report, IRRI receives support, through the CGIAR, from a number of donorsincluding UNDP, World Bank, European Union, Asian Development Bank, and Rockefeller Foundation, and the internationalaid agencies of the following governments: Australia, Belgium, Canada, People’s Republic of China, Denmark, France,Germany, India, Indonesia, Islamic Republic of Iran, Japan, Republic of Korea, The Netherlands, Norway, Philippines, Spain,Sweden, Switzerland, United Kingdom, and United States.

4 IRRN 1998
Germplasm improvementGenetic resources
International testing of ricehybrids for yield andadaptability by INGER:prospects and problems
R. C. Chaudhary and S. S. Virmani, IRRI
Because of the potential of hybrid riceand its commercial success in China,many countries in Asia, Africa, LatinAmerica, and the Caribbean showed akeen interest in developing and usingthis technology. The first InternationalHybrid Rice Observational Nursery(IRHON) was put together in 1994 with44 hybrids (H), 36 restorers (R), and 5maintainers (B) developed at IRRI. Thisnursery was evaluated in Bangladesh(Gazipur), China (Changsha, Hang-zhou), India (Coimbatore, Faizabad,Hyderabad, Kapurthala, Karnal,Mandya, Maruteru, Pantnagar), Myan-mar (Yangon, Yezin), the Philippines(Los Baños, Maligaya), and Sri Lanka(Batalagoda). A number of hybrids that
could be adapted in most of thesecountries were identified. HybridsIR67693H, IR69672H, IR69684H,IR69686H, and IR69689H were identi-fied as superior and widely adapted. Inthe second IRHON conducted at 29locations in 13 countries, 33 hybrids, 3 Blines, and 25 R lines developed in IRRI,China, India, and Myanmar weretested. Hybrids IR67693H, IR64616H,IR68284H, IR68877H, IR69679H, andIR70402H were identified as superior.In the third IRHON, three hybrids froma private company were also included.This nursery is now undergoing testingat 45 locations in 16 countries in Asia,Africa, and Latin America.
Superior hybrids in earlier testsproduced up to 3 t ha-1 more grain thanthe best locally adapted check varieties.Genotype x environment (G × E) inter-action analysis and superiority analysishelped identify the stability of thehybrids and forecast their adaptability
to untested locations. The G × Eanalyses did not show a wider adapt-ability of the hybrids when comparedwith their parents or checks. INGER,through its global access to hybrids,some parental material, and test sites,can support the testing and use ofhybrids and available parents in inte-rested national agricultural systems.INGER can thus help realize the dreamof interdependence, exchange, recipro-city, and sharing in a network where nocountry is too poor to give and nocountry is too rich to receive. Butreservations of some breeders insharing parental lines and seedborneproblems like kernel smut (Tilletia) andnematodes impose restrictions. Thecost of testing, breeders’ rights, andintellectual property rights are someproblems that still loom over the scien-tific community. All these problemscould affect the evaluation of hybridsthrough the INGER mechanism. ■
Screening rice hybrids forquality traits
N. S. Rani, M. I. Ahmed, and B. Krishnaveni,Directorate of Rice Research, Rajendranagar,Hyderabad 500030, India
In rice hybrids, characters such aslemma and palea, seed size and shape,and pericarp color do not segregatebecause they are inherited throughmaternal tissue. Endosperm trans-lucency, chalkiness, and cooking andeating quality traits generally showgenetic segregation. Rice is the onlycereal consumed as unprocessed wholegrain and consumers value specificappearance, taste, and cooking qualitytraits. Therefore, the effect of hetero-zygosity of F1 hybrids on grain qualityis more important.
We evaluated 27 hybrids in tworeplications for 13 physicochemical
characteristics in the 1995 wet seasonharvest and using standard proce-dures, including physical parameterssuch as grain shape and size, endo-sperm appearance, milling, and headrice recovery (HR%). Among cookingand eating quality traits, we studiedamylose content (AC%), gelatinizationtemperature (GT), water uptake (WU),volume expansion ratio (VER), kernellength after cooking (KLAC), andelongation ratio (ER).
Of the 27 hybrids tested, 17 belongedto long slender, 4 to long bold, 5 tomedium slender, and 1 to short boldgrain type. Four hybrids—URH1,IR58025A/IR54742, IR58025A/IR34686, and IR58025A/IR32809—possessed extra long grains. Except forURH1, the other three hybrids showedintermediate AC (23.6 and 25.4%).IR58025A/IR34686 exhibited a highVER (5.3). Six hybrids—IR58025A/
IR29723, 3RI-086, MTURH2020, 2RI158, MTURH2015, and MPH517—recorded a high HR (ranging from 60.3to 63.9%).
Eleven hybrids were in the mostdesirable AC range (20-25%) preferredin the Indian subcontinent. IR58025A/IR48751 and IR58025A/IR21567 hadtypical intermediate AC values (24.9and 23.3%) with long slender andattractive translucent grains. Otherhybrids that possessed an intermediateAC and long slender grains, but withoccasional chalkiness, were PA112,HKRH1002, and IR58025A/IR48749.
Hybrids MPH517, 3RI 160, andPMS1OA/IR48725 possessed endo-sperm translucency and high HR.Hybrids with desirable starch proper-ties (intermediate AC) and translucentgrains with moderate HR wereIR58025A/IR29723, IR58025A/IR34686, IR58025A/IR55838, and

Vol. 23, No. 1 5
IR58025A/IR21567. But the last twohybrids also possess long slender andattractive grains, moderate HR, and ahigh VER, a good combination ofquality traits. ■
Studying comparativesuitability of CMS lines
M. I. Ahmed, S. Singh, B. C. Viraktamath, M. S.Ramesha, and C. H. M. Vijayakumar, Directorateof Rice Research, Rajendranagar, Hyderabad500030, India
Parental lines constitute the first andforemost step in a hybrid breedingprogram. In this context, developing acommercially viable cytoplasmic malesterile (CMS) line is considered to be ahighly cumbersome process. In fact, thebelated success of hybrid rice technol-ogy in India was basically due to thenonavailability of a CMS line suited tothe tropics. Over the years, a largenumber of male sterile lines withdifferent cytosterility sources havebeen developed in India and elsewhere.Only a few—such as IR58025A andIR62829A—are now used commer-cially in India. They possess all theessential traits—complete and stablemale sterility, high outcrossing rate,better grain quality, easy restorability,good combining ability, and adaptabi-lity—of a commercially viable CMSline. Some 64 CMS lines from India,China, Malaysia, and IRRI were evalu-ated along with IR58025A, IR62829A,and standard checks in the wet seasonof 1995 and 1996 and dry season of 1994and 1995 to assess their comparativesuitability for commercial use. TheCMS lines, along with their corre-sponding maintainers, were grown formaintenance and evaluation for theirfloral traits such as pollen sterility,panicle and stigma exsertion, out-crossing rate, duration and angle ofglume opening, growth duration,number of effective tillers, spikeletspanicle-1, grain type, adaptability, andpigmentation (if any). Each CMS linewas characterized for all the traitsindividually. The final value was
obtained by adding together theweighted score allotted to a CMS linefor all these traits. This single finalvalue on a 1 to 9 scale indicated theline’s practical utility.
Only 11 lines were found to beequally better for all the charactersstudied than IR58025A and IR62829A:four CMS lines (IR68280A, IR68897A,IR68899A, and IR69628A) from IRRI,two (DRR2A and DRR3A) from theDirectorate of Rice Research, and oneeach from China (9601A), Malaysia(MH-841A), IARI (Pusa 5A), Cuttack(CRMS 31A), and Faizabad (NDCMS7A). Some of the promising CMS lineswere IR67684A, IR68890A, IR68902A,IR68279A, IR68895A, and CRMS 6A.About 31 lines possess one or two goodcharacters and can be used for specificpurposes. All Chinese lines, for exam-ple, are useful as cytosterility sources toconvert promising maintainers intonew CMS lines. The remaining 16 lineshave one or more drawbacks and willneed further improvement. Thepromising CMS lines are now beingstudied for their combining ability anduse in breeding programs. ■
Identifying a new long-duration CMS line (APMS5A) for coastal regions
R. V. Kumar, P. V. Satyanarayana, and M. S. Rao,Andhra Pradesh Agricultural University,Agricultural Research Station, Maruteru 534122,India
Several male sterile lines developed inIndia belong to the early- to medium-duration group. Hybrids developedusing these male sterile lines are also ofearly to medium growth duration.These hybrids are not suitable forcultivation during the wet season,particularly in the coastal areas ofAndhra Pradesh, where long-durationvarieties are predominantly grown. Weneed to develop long-duration ricehybrids to increase rice productivityand fit them into the cropping system.An effort was made to develop long-duration (145-150 d) and stable local
cytoplasmic male sterile (CMS) lines.Several testcrosses were made usinglocal long-duration elite lines and IRRICMS lines to screen the elite varietiesfor their maintaining or restoringability. A few long-duration maintainerlines were identified based on pollenand spikelet sterility. MTU4870 wastherefore selected and successfullyconverted into a local cytosterile line inthe background of a wild abortivesource of cytoplasm through thebackcross breeding technique. This linewas designated as APMS5A.
APMS5A is a long-duration (145-150 d) line with tolerance for brownplanthopper, bacterial leaf blight, ricetungro virus, and sheath blight. It alsopossesses a sturdy culm. It has acomparable angle of spikelet openingduration (190 min), angle of spikeletopening (31°C), stigma exsertion, andnatural outcrossing potential (12%)with the popular CMS line IR58025A.APMS5A, a 100% sterile line, is the firstlong-duration CMS line developed inIndia with desirable floral andagronomic traits. A few effectiverestorers were also identified. APMS5Awill facilitate the development of long-duration rice hybrids suitable forcultivation in the coastal areas. ■
Wide hybridization fordiversification of CMSin rice
N. T. Hoan, N. P. Sarma, and E. A. Siddiq,Directorate of Rice Research, Rajendranagar,Hyderabad 500030, India
To identify new sources of malesterility-inducing cytoplasm within theA genome of genus Oryza, 132 inter-specific crosses involving accessions offour wild (O. rufipogon, O. nivara, O.barthii, and O. longistaminata) and twocultivated species (O. sativa and O.glaberrima) were effected. Accessionspossessing sterility-inducingcytoplasm were identified followingreciprocal and F2 backcross methodsand advanced through substitutionbackcrossing to develop cytoplasmic

6 IRRN 1998
male sterile (CMS) lines. Two indiceswere used: (1) pollen morphology andstaining pattern and (2) type ofinteraction with a set of maintainersand restorers of wild abortive (WA)cytosterile stock. The newly developedCMS lines are grouped using theseindices as follows: I. RPMS1 (O.rufipogon; VNI) and RPMS2 (O. nivara;DRW21039) are gametophytic types;II. RPMS3 (O. nivara; DRW21030) issimilar to MS577A in pollen stainabil-ity, but the restoration and main-tenance reactions are similar to those of
Krishna CMS lines in thebackground of fourdifferent cytoplasmicsources in rice
S. B. Pradhan and P. J. Jachuck, Central RiceResearch Institute, Cuttack 753006, India
Four cytoplasmic male sterile (CMS)lines of Krishna with sterile cytoplasmfrom four different sources (wildabortive [WA], O. perennis, Kalinga 1,and Lalruma from Mizoram) have beendeveloped. The two CMS lines—Krish-na A with WA and Krishna A with O.perennis as a source—were developedthrough conversion with CMS linesV20A (WA) and IR66707A (O. perennis),respectively, by subsequent backcross-ing with common isonuclear main-
tainer Krishna A. But Krishna A withKalinga 1 and Krishna A with Lalrumacytoplasmic sources were developedthrough indica/indica crosses. Thesefour CMS lines are all dwarf in stature,are of medium duration, and havereduced white anthers that exhibit 80-90% unstained withered pollen grains.
To understand the nature of thesefour cytosterility sources, they weretestcrossed with four maintainers—V20B (maintainer of WA), IR66707B(maintainer of O. perennis), Yar-Ai-ZhaoB (maintainer of Gambiaca), andMS577B (maintainer of O. sativa f.spontanea)—and the elite inbred linesSPR7210-1-3, IR48725-B-B-120-1,NDR30074, TTB150-61, and R657-93-869. The sterility of the four CMS lineswas maintained by the four main-
the WA type; III. RPMS4 (O. nivara;DRW21018) is a sporophytic type, butrestorers and maintainers are differentfrom WA; IV. RPMS5 and RPMS6 (O.nivara; RPW21111) are sporophytictypes with restorer and maintainerreactions similar to those of WA. All thenew CMS lines possessed completepanicle exsertion essential for enhanc-ing outcrossed seed yield.
For the two stable CMS lines,MS577A and IR66707A, no restorers areavailable in cultivated rice germplasm.A search for restorer sources for these
CMS lines was made among the wildaccessions of the A genome species.Although none of the accessions couldrestore fertility in IR66707A, threeaccessions of O. rufipogon (VN2,DRW22016, DRW220175) and one eachfrom O. sativa f. spontanea (RPW20001)and O. glaberrima (DRGL 30090) didrestore fertility in MS577A. Studies onthe genetics of fertility restoration inthese cross combinations indicated thattwo dominant genes act in an additivemanner to restore fertility. ■
tainers. The restorer SPR7210-1-3 ofKrishna A (WA) was found to be amaintainer of the other three CMSlines—Krishna A (O. perennis), KrishnaA (Kalinga 1), and Krishna A(Lalruma)—whereas the restorerIR48725-B-B-120-1 of Krishna A (WA)behaved as partial restorer of Krishna A(Kalinga 1) and Krishna A (Lalruma)and as a maintainer of Krishna A (O.perennis). Variety R657-93-869 wasfound to be a partial restorer of KrishnaA (WA) and Krishna A (Lalruma) and amaintainer of Krishna A (Kalinga 1)and Krishna A (O. perennis). Therestorer TTB150-61 for Krishna A(Lalruma) behaved as a partial restorerof Krishna A (WA) and Krishna A(Kalinga 1), and as a maintainer ofKrishna A (O. perennis). ■
Characterizing thermo-sensitive genic male sterilelines of rice
B. C. Viraktamath, M. T. Lopez, and S. S. Virmani,IRRI
The cytoplasmic male sterility (CMS)system is widely used to develop ricehybrids. Although effective, thissystem is quite cumbersome to prac-tice. Certain genic male sterile lines thatrevert to fertility under specifictemperature are called thermosensitivegenic male sterile (TGMS) lines. De-ployment of the TGMS system todevelop two-line hybrids has several
advantages over the conventional CMSsystem, as it requires neither main-tainers for seed multiplication norrestorers for producing hybrids. Inaddition, two-line hybrids may notsuffer from possible negative effects ofthe sterility-inducing cytoplasm. For aneffective use of TGMS lines, they haveto be characterized for their criticalsterility and fertility points. Thisinformation is essential for deciding ona suitable location and/or period forseed multiplication and hybrid seedproduction. Two IRRI-bred indicaTGMS lines-IR68945-4-33-4-14 andIR68949-11-5-31-along with theirjaponica TGMS donor, were charac-
terized under controlled conditions inthe IRRI phytotron. These lines weresubjected to eight constant tempe-ratures (20-32 °C) and four day-nightcombination temperatures (27/21 °C-32/24 °C) for a period of 3 wk duringthe sensitive stage. The critical sterilitypoint for both lines was 30 °C andabove, whereas it was only 27 °C andabove for Norin PL 12. The criticalfertility points for the indica TGMSlines ranged between 24-28 °C(constant temperate) and 27/21 °C-28/22 °C (day-night combination tempera-ture). Norin PL 12 was found to be verysensitive to higher temperature com-pared with the indica TGMS lines.

Vol. 23, No. 1 7
Comparative effects from treatmentsrevealed that the maximum tempera-ture in day-night combination tempe-ratures was crucial in determining thesterility or fertility of TGMS lines. Theeffects of intervening fertility-inducingtemperature (27 °C) during the sterilityphase (32/24 °C) were studied inTGMS lines IR68945-4-33-4-14, ID24,and Norin PL 12. Even an exposure for2 h at 27 °C could induce fertility inIR68945-4-33-4-14, whereas ID24remained unaffected even after 10 hexposure at 27 °C. The behavior ofNorin PL 12 was intermediate in thisrespect as 4 h of interruption couldinduce fertility. ■
Screening rice germplasmfor floral attributes thatinfluence outcrossing
S. K. Sahoo, R. Singh, L. C. Prasad, R. M. Singh,D. K. Singh, Genetics and Plant BreedingDepartment, Institute of Agricultural Sciences,Banaras Hindu University, Varanasi 221005; andS. K. Agrawal, Directorate of Rice Research,Rajendranagar, Hyderabad 500030, India
A cytosterile line with high outcrossingpotential would economize the cost ofhybrid seed production in rice. Wescreened rice germplasm to identifygenotypes with useful floral traits thatcould influence outcrossing. We usedprospective maintainers and restorersin a testcross nursery program. Isola-tion of prospective maintainers withuseful floral characteristics could leadto the development of cytosterile lineswith high outcrossing potential
Identifying and charac-terizing thermosensitivecytoplasmic male sterilelines
A. J. Ali, Crop Improvement Department,Agriculture College and Research Institute,Navalur Kuttapattu, Trichy 620009; E. A. Siddiq,Indian Council of Agricultural Research, NewDelhi 110001; F. U. Zaman, Indian AgriculturalResearch Institute, New Delhi 110012; and M. I.Ahmed, Directorate of Rice Research, Hyderabad500030, India
In wheat, an alloplasmic line of Norin26 (Triticum aestivum) with Aegilopscrassa cytoplasm has been found toshow male sterility under long-day(15.0 h) conditions and fertility undershort-day (14.5 h) conditions with noinfluence of temperature. In the presentstudy, a similar influence of tempera-ture rather than photoperiod was ob-served in wild abortive (WA) cyto-plasmic male sterile (CMS) line J24A(Pusa 33-16A). It was studied alongwith its maintainer line J24B (Pusa 33-16B) under different temperatureregimes for fertility-sterility alterations.The pattern of their fertility-sterilitytransformation revealed them to be ofhigh critical sterility point (CSP) andhigh critical fertility point (CFP) type.Yet they differed in their degree oftransformation; the CSP was 33.8 °C inJ24A and 35.1 °C in J24B. Although the
CFPs of both fall in the same range(24␣ °C), a comparative study of the Aand B lines for their temperature-influenced sterility-fertility behaviorrevealed the thermosensitive genicmale sterility (TGMS) gene to beprobably linked to the nuclear sterilitygene (rf). Fertile cytoplasm (C gene)also possessed some degree ofinfluence on fertility as well as onTGMS genes located in the nucleargenome. This was evident from the Bline, which showed a relatively higherCSP and CFP than the A line. To have astable CMS line, the nuclear genomeshould have the fertility gene Rf withthe environment-sensitive malesterility gene (EGMS) in a dominantstate, so that neither temperature nordaylength would affect its stability forpollen sterility. A CMS line having the
EGMS gene in a recessive state can bedeployed under high-temperatureconditions for hybrid seed production.The A line in this state will be morestable for sterility because of the com-bined effect of both fertility and EGMSgenes. Seed multiplication of the CMSline can be done with ease, takingadvantage of fertility transformationunder low-temperature conditions. Tominimize the risk of temperature fluc-tuations, the CMS line can be planted inalternate rows with its B line. If hightemperatures prevail during the seedmultiplication phase and make theCMS line completely sterile, the B lineas a maintainer can provide abundantpollen for outcrossing. If selfing doesoccur, it would help to further increaseseed yield of the thermosensitive CMSline. ■
through a backcrossing program. Weevaluated 75 genotypes for floral andrelated traits: duration of opening offlorets, angle of opened florets, per-centage of exserted stigma, spikeletlength, anther length, stigma length,panicle length, number of effectivetillers plant-1, grain yield plant-1, andplant height. The analysis of variancerecorded highly significant differencesfor all the traits, indicating the presenceof a sufficient degree of variation forthese traits for selection among culti-vars. We identified genotypes Aditya,VL Dhan 63, IET10119, IET10115,IET10119, Shankar, VL Dhan 8,Pratibha, Kunti, Surajmukhi, and Vikasas possessing multiple floral traits thatwould enhance seed set. All these geno-types can therefore be used as parentallines to develop hybrids and enhanceoutcrossing. ■
Review of notes. The IRRN editor will send an acknowledgment card or an e-mailmessage when a note is received. An IRRI scientist, selected by the editor, reviewseach note. Reviewer names are not disclosed. Depending on the reviewer’s report, anote will be accepted for publication, rejected, or returned to the author(s) forrevision.
Comments. If you have comments or suggestions about the IRRN, please write to theeditor.

8 IRRN 1998
Genetics
Physical localization of phyAand rbcS genes in rice
Xuezhi Bi, Nan Ren, Lihua Liu, Kecheng Cheng,Yunchun Song, College of Life Science, WuhanUniversity, Wuhan 430072, People's Republic ofChina
Phytochrome is an important photo-receptor and regulates many growthand development processes in plants(Thompson and White 1991, Furuya1993, Terzaghi and Cashmore, 1995).Ribulose-1.5-bisphosphate carboxylase(Rubisco) small subunit genes (rbcS) areregulated developmentally by light(Tobin and Silverthorne 1985, Gilmar-tin et al 1990). Both genes (phyA andrbcS) coding for phytochrome andRubisco SSU are functionally related.We tried to physically map onto thechromosomes by in situ hybridizationrice genes related to photoperiod/lightreception, signal transduction, andgene expression regulated by photo-period, aiming to understand the rela-tionship between the structure andfunction of the rice genome and be-tween the genetic map and the physicalmap. The physical locations of CaMand Ca2+-ATPase genes were reported(Bi et al 1996). The results of those ofphyA and rbcS genes follow.
The rice phyA gene clone, pPHY1,and rbcS gene clone, pRR1, are genomicclones of 6.6 kb and 1.1 kb in size, kind-ly provided by Prof. Nam-Hai Chua ofRockefeller University and Prof. RayWu of Cornell University, USA. Oryzasativa L. subsp. indica, cv. Guang-lu-ai 4,was used for chromosome prepara-tions. Clones pPHY1 and pRR1 weretransformed into E. coli TGI. Plasmidamplification, extraction, labeling withbiotin, in situ hybridization, and detec-tion were performed with the proce-dures followed by Bi et al (1996). Chro-mosomes were identified according todata from Kurata (1986).
The results showed that the hybridi-zation detection ratio was about 29.79%for phyA and 21.56% for rbcS. The phyA
gene was located near the centromereof the long arm, at the end of the shortarm, and in the middle of the long armon chromosome 3. The rbcS gene wasmapped near the centromere on thelong arm of chromosome 7, at the endof the long arm of chromosome 5, andat the 2/3 distance of the long arm fromthe centromere in chromosome 6 (seetable, figure).
The phyA gene was near RZ575 onchromosome 3 of the genetic map(Causse et al 1994), and RZ575 was onthe long arm (Singh et al 1996). We alsolocated the phyA gene on chromosome3 and found three loci. This suggeststhat fragments for the phyA gene aredispersed on chromosome 3.
Kuozuka et al (1993) proved that therbcS sequence of pRR1 (Xie and Wu1988) showed 98.8% homology withthe rice rbcS cDNA clone OSRUBPC1(Matsuoka et al 1988). Wu et al (1986)indicated that two genes coding for ricerbcS share more than 95% DNA
sequence homology in a sequenced 230bp segment, and one of the two clonedgenes (pOSrbcS-1) was probably loca-ted on three chromosomes. Matsuokaet al (1988) also thought that a fewgenes encoded rbcS in rice from theSouthern blot results, with thepOSSS1139 cDNA insert as a probe.Therefore, our result of physicallymapping multipositions of rbcS onchromosomes is consistent with theabove results.
Rice chromosomes are small andvery difficult to identify individually,and all chromosomes have deeplystained regions on both sides of thecentromere (Kurata and Omura 1978).This different Giemsa staining patternmakes it difficult to locate the true endsof the chromosomes and the relativechromosomal location of probes (Jianget al 1995). We are going to continue tomap the phyA and rbcS genes bymulticolor fluorescence in situhybridization with both gene clonesand centromere-specific randomfragment length polymorphismmarkers for special chromosomes.
Cited referencesBi XZ, Song YC, Ren N, Liu LH. 1996.
Physical mapping of CaM and Ca2+-ATPase genes in rice by in situhybridization. Rice Genet. Newsl. 13:85-89.
Causse MA, Fulton TM, Cho YG, Ahn SN,Chun-wongse J, Wu K, Xiao J, Yu Z,Ronald PC. 1994. Saturated molecularmap of the rice genome based on aninterspecific backcross population.Genetics 138:1251-1274.
Chromosome locations and detection ratios of phyA and rbcS genes in rice.
Clone name Size (kb) Chromosome locationa Average detection ratio (%)
3L near centromere 14.89phyA 6.6 3S end 12.77
3L middle 2.13Total 29.79
7L near centromere 8.62rbcS 1.1 5L end 6.90
6L 2/3 distance 6.04from centromere
Total 21.56
aL = long arm, S = short arm.
3 5 6 7
rbcS
rbcS
rbcSphyA
Chromosome maps of rice phyA and rbcSgenes showing hybridized positions. Shortarm is at the top, constriction represents thecentromere, and the long arm is under theconstriction.

Vol. 23, No. 1 9
Furuya M. 1993. Phytochromes: theirmolecular species, gene families, andfunctions. Annu. Rev. Plant Physiol.Plant Mol. Biol. 44:617-645.
Gilmatin PM, Sarokin L, Memelink J, ChuaNH. 1990. Molecular light switches forplant genes. Plant Cell 2:369-378.
Jiang J, Gill B S, Wang G L, Ronald PC, WardDC. 1995. Metaphase and interphasefluorescence in situ hybridizationmapping of the rice genome withbacterial artificial chromosomes. Proc.Natl. Acad. Sci. USA 92:4487-4491.
Kurata N.1986. Chromosome analysis ofmitosis and meiosis in rice. In: Ricegenetics. International Rice ResearchInstitute, Manila, Philippines. p 143-152.
Kurata N, Omura T. 1978. Karyotypeanalysis in rice. I. A new method for
identifying all chromosome pairs. Jpn. J.Genet. 53:251-255.
Kyozuka J, McElroy D, Hayakawa T, Xie Y,Wu R, Shimamoto K. 1993. Light-regulated and cell-specific expression ofTomoto rbcS-gusA rice rbcS-gusA fusiongenes in transgenic rice. Plant Physiol.102:991-1000.
Matsuoka M, Kano-Murakami Y, Tanaka Y,Ozeki Y, Yamamoto N. 1988. Classifica-tion and nucleotide sequence of cDNAencoding the small subunit of ribulose-1,5-bisphosphate carboxylase from rice.Plant Cell Physiol. 29(6):1015-1022.
Singh K, Ishii T, Parco A, Huang N, Brar DS,Khush GS. 1996. Centromere mappingand orientation of the molecular linkagemap of rice (Oryza sativa L.). Proc. Natl.Acad. Sci. USA 93:6163-6168.
Terzaghi WB, Cashmore AR. 1995. Light-regulated transcription. Annu. Rev.Plant Physiol. Plant Mol. Biol. 46:445-474.
Tobin EM, Silverthorne J. 1985. Light regu-lation of gene expression. Annu. Rev.Plant Physiol. Plant Mol. Biol. 36:569-593.
Tompson W F, White M J. 1991. Physio-logical and molecular studies of light-regulated nuclear genes in higher plants.Annu. Rev. Plant Physiol. Plant Mol.Biol. 42:423-460.
Xie Y, Wu R. 1988. Nucleotide sequence of aribulose-1,5-bisphosphate carboxylase/oxygenase small subunit gene (rbcS) inrice. Nucleic Acids Res. 16:7749.
Wu H-K, Chung M-C, Xie Y, Wu R. 1986.Chromosomal localization of rice rbcSgenes. Rice Genet. Newsl. 3:117-119. ■
Fertility restoration studiesin four WA CMS lines ofrice
S. L. Kumari, Regional Agricultural ResearchStation, Pattambi; M. Mahadevappa, Universityof Agricultural Sciences (UAS), Dharwad; andR. S. Kulkarni, UAS, Bangalore, India
Fertility restoration studies involvingfour wild abortive (WA) cytoplasmicmale sterile (CMS) lines—V20A,IR58025A, IR62829A, and IR54752A—resulted in the identification of 20restorers. We studied segregation forfertility in the F2 and backcross pro-genies of these CMS lines with fourrestorer lines—ARC11353, IR30864,IR13419-113-3, and IR9761-19-1. Theresults indicated that fertility restora-tion in WA CMS lines was controlled bytwo additive major genes. Their effectwas sporophytic and they displayeddifferential gene interactions such asepistasis with complete dominance(12:3:1) or epistasis with incompletedominance (9:6:1) depending on theparents involved in the cross.
We noticed differential segregationof the F2 and backcross progenies in thecrosses involving IR30864 and IR9761-19-1 depending on the female parent.This suggested that the performanceand expressivity of the R genes variedaccording to the nuclear background of
the female parent. The pattern of segre-gation in crosses involving IR54752Awas different from that of other CMSlines, suggesting the involvement ofcertain minor genes for fertility resto-ration carried by this CMS line indeciding the fertility of the progenies. ■
Effect of minor genes inrestoration of fertility inCMS lines of rice
K. N. Ganesan, K. Thiyagarajan, and M.Rangaswamy, School of Genetics, Tamil NaduAgricultural University, Coimbatore 641003,India
We evaluated 128 hybrids in the 1994dry season and 135 hybrids in the 1995wet season. The hybrids, involvingvarieties C29 and PMK2, with differentwild abortive (WA) sources of cyto-plasmic male sterile (CMS) lines—IR58025A, IR62829A, and PMS3A—showed marked differences in pollenand spikelet fertility. When C29 wascrossed with IR62829A and PMS3A, itbehaved as a partial restorer (21-90%pollen and spikelet fertility). But whencrossed with IR58025A, it behaved as amaintainer (100% pollen and spikeletsterility). All three CMS lines, however,had WA cytoplasm. Similarly, PMK2behaved as a restorer (>90% pollen and
spikelet fertility) with PMS3A, while itmaintained sterility (100%) withIR62829A. The minor or modifier genespresent in the pollinator might havereacted with the CMS lines used andresulted in this type of variation. ■
Breeding for male fertilityrestoration incorporatingBasmati rice quality
F. U. Zaman, M. J. Abraham, U. S. Natarajan,A. Mahendru, and F. Mohammad, Division ofGenetics, Indian Agricultural Research Institute(IARI), New Delhi 110012, India
Basmati is the most highly valued ricecommodity in the world agriculturaltrade market. Pusa Basmati 1, releasedin India, combines both high yield andideal Basmati quality. In Pakistan, semi-tall Pak 385 and Pak 386 are reported tobe better yielding than Basmati 370.Results at IARI have demonstrated thepossibility of developing hybrids withBasmati grain quality and a high levelof heterosis. Such Basmati hybridswould raise yields further.
A major limitation to developingBasmati rice hybrids is the nonavaila-bility of satisfactory restoration amongBasmati cultivars or breeding lines.Some 90% of Basmati materials showedimperfect maintaining ability, 3%

10 IRRN 1998
showed good maintenance, and 7%showed partial restoration. But fewshowed 60-70% restoration ability. Thissituation led us to begin a systematicrestorer breeding program incorpora-ting ideal Basmati quality.
Testcrosses were made using cyto-plasmic male sterile (CMS) lines Pusa1127A ( a new cytosterility source fromPusa 743), Pusa 3A wild abortive (WA),and IR58025A (WA) with the Basmaticultivar or breeding line that hadshown high F1 fertility (60-70%) inprevious testcrosses. All the F1s wereraised. Seeds from the best fertile plantswere harvested and subjected to grainand cooking quality tests. Only thosehybrids that exhibited Basmati cookingcharacteristics in an appreciable pro-portion of grains were raised to the F2generation. Selections were made onthe basis of spikelet fertility, grainshape, and grain appearance. Theywere subjected to cooking quality testsand only those possessing idealBasmati characteristics were raised tothe F3. In the F3, diverse-looking plantswere selectively intermated and theentire process was repeated. Simul-taneously, some of these selected plantswere testcrossed with Pusa 3A. Pollendonor plants of hybrids showing im-proved restoration and ideal Basmatiquality were advanced to the F4. In theF4 and later generations, the entireprocess was again repeated as in the F3.Likewise, the material was advanced tothe F6 generation. The intermated lineswere also advanced to further gene-rations.
Testcrosses made between Pusa 3Aand the selected lines revealed that ninelines—Sps95-694-2, Sps95-694-3, Sps95-81-1, Sps95-81-3, Sps95-81-4, Sps95-81-5, Sps95-81-6, Sps95-768, and Sps95-167-1—showed normal restoration.The grain quality of these hybrids wascomparable with that of Pusa Basmati 1and Karnal Local. Three of these linesare now used for Basmati hybrid seedproduction with Pusa 3A (BasmatiCMS line). ■
Genetics of thermo-sensitive genic male sterilelines in rice
O. U. K. Reddy, Directorate of Rice Research(DRR), Rajendranagar, Hyderabad 500030;E. A. Siddiq, Indian Council of AgriculturalResearch, New Delhi 110001; J. Ali, CropImprovement Department, Agriculture Collegeand Research Institute, Navalur Kuttapattu,Trichy 620009; A. J. Hussain and M. I. Ahmed,DRR, Rajendranagar, Hyderabad 500030, India
Understanding the inheritance patternand studying allelic relations amongdiverse thermosensitive genic malesterile lines (TGMS) were the objectivesof this investigation. Lines F61, SA2,JP1, JP8-8-1s, ID24, and IC10, devel-oped at the DRR, Hyderabad, and linesIR32364, IR68292, IR68294, IR68945,and IR68949, received from IRRI,Philippines, were crossed with a set ofnormal pollen parents, to study segre-gation patterns in the F2, F3, andbackcross generations. The resultsindicated that sterility caused by theTGMS trait was governed by a singlerecessive gene. In segregating popu-lations, however, individual TGMSprogenies from the same F2 showeddifferent fertility levels. This indicatedthat some other genes modify the majorgene and affect fertility.
Allelic relations were examined bystudying direct crosses and backcrossesinvolving two different TGMS linesover seasons. The lines developed fromNorin PL 12 (tms2 ) (IR68292, IR68294,IR68945, and IR68949) at IRRI wereallelic to ID24 and JP1. The linesIR32364 (tms3), JP8-8-1s, and IC10 werealso allelic. Lines SA2 and F61 wereobserved to be nonallelic to all otherlines, and may possess different TGMSgenes, including tms1 from China. ■
Mitochondrial geneexpression in cytoplasmicmale sterile and fertilelines of rice
S. Venmadhi and K. K. Narayanan, Center forBiotechnology, SPIC Science Foundation, 110Mount Road, Guindy 600032, India
In various plant species, rearrange-ments in the mitochondrial genomehave been associated with cytoplasmicmale sterility (CMS). These rearrange-ments could modify genome expres-sion and thereby play a role in pollenabortion. In rice, investigations carriedout in our laboratory have establishedsome rearrangements in the geneticorganization of mitochondrial DNAbetween male sterile and fertile lines.For example, the cox1 locus was foundrearranged in the CMS line comparedwith the fertile lines.
To establish the correlation, if any, inDNA organizational pattern with theexpression of the male sterile pheno-type, we conducted a Northern analy-sis on total RNA from the sterile andfertile lines. The blot, when probedwith atp6, showed a single transcript inall the lines tested, whereas atp9 andcox3/orf25 showed five and two trans-cripts, respectively. On probing withatp9, we observed multiple transcriptsof sizes 3.2 kb, 1.7 kb, 1.3 kb, 0.8 kb, and0.6 kb, and 0.3 kb was the most abun-dant of the transcripts, probably repre-senting the mature functional trans-cript. The cotranscribed genetic locus ofcox3/orf25 when used as a probe,yielded two transcripts of 2 kb and 1 kb.Even though atp6 in Chinsurah Boro-type cytoplasm has been found to bedifferentially transcribed and edited,and hence implicated in CMS, we couldnot detect different transcripts of atp6 inthe rice lines used in our study bearingWA type cytoplasm. Work on puttingmore probes on the Northern blots andstudying the gene-specific pattern ofexpression is in progress. ■

Vol. 23, No. 1 11
RAPD and PBR analysis ofCMS maintainer andrestorer lines in rice
K. K. Jena and M. Anil, MAHYCO ResearchFoundation, Biotechnology Center, 245-IIKamalapuri Colony, Hyderabad 500073, India
Recently developed molecular toolssuch as randomly amplified poly-morphic DNA (RAPD) and polymerasechain reaction-based random fragment
Breeding methods
Transferring wild abortivecytoplasmic male sterilitythrough asymmetric fusionof protoplasts
B. Bhattacharjee and H. S. Gupta, Division ofPlant Breeding, ICAR Research Complex forNEH Region, Umroi Road, Barapani 793103,Meghalaya; and A. P. Sane, Center for PlantMolecular Biology, National Botanical ResearchInstitute, Lucknow 226001, India
We attempted a one-step transfer ofwild abortive cytoplasmic male sterility(CMS) through asymmetric fusionbetween the protoplasts of CMS andfertile maintainer lines. We establishedembryogenic cell suspensions from thecalli obtained from mature seed scu-tella of a CMS line (V20A), fertile
V20A/V20B. Pollen grains from all thecybrid lines were sterile.
Analyses of mitochondrial DNA ofthe fusion partners and cybrids wereconducted by using orf 156 as a probe.This revealed differences in polymor-phism between the mitochondrial DNAof the CMS line and fertile maintainers.In the case of HindIII-digested DNA, theorf 156 gene was located on a 1.3-kbfragment in the male sterile line, V20A,and on a 12-kb fragment in its fertilemaintainers, RCPL1-2C and V20B. ADNA blot analysis of the HindIII-digested DNA of all five cybridsshowed hybridization of orf 156 to the1.3-kb fragment. This demonstrated thetransfer of mitochondrial DNA from theCMS line to the cybrids. ■
length polymorphism (PBR) are avail-able for characterizing genetic variationand DNA typing of rice genotypes.DNA analysis of three cytoplasmicmale sterile (CMS) lines with therespective maintainers (B) and tworestorer lines (R) using 12 decamerprimers from Operon (OPA-01 to OPA-12) revealed polymorphism only withprimers OPA-03, OPA-07, OPA-09, andOPA-12. CMS lines IR67684A,IR58025A, and IR62829A have distinct
RAPD markers in contrast to their Band R lines. DNA polymorphism of A,B, and R lines was also confirmed withPBR in 15 primer/enzyme combina-tions. PBR reaction amplified a 546-bp(OPA-12) fragment in IR58025A and an872-bp (OPA-12) fragment in IR62829Alines in a genotype-specific manner.These markers are being developed foridentifying genotype-specific DNAmarkers and mapping desirable genesfor hybrid rice research. ■
maintainers (RCPL1-2C and V20B),and restorer IR36 lines. We isolated andcultured protoplasts to produce proto-calli that differentiated into plants.Pollen grains of the protoplast-derivedplants of the CMS line were sterile,whereas those derived from the main-tainer and restorer lines were fertile. Toaccomplish asymmetric fusion, theprotoplasts of the CMS line and fertilemaintainer lines were inactivated with30 krad gamma ray and 10 mM iodo-acetamide, respectively. Electricallyfused protoplasts divided on a culturemedium and formed micro-coloniesthat developed into calli. These callialso differentiated on transfer to aregeneration medium. We recovered 27cybrid lines from the fusion product ofV20A/RCPL1-2C and 23 lines from
Adaptability and yield ofrice hybrids
M. I. Ahmed, C. H. M. Vijayakumar, S. Singh,M. S. Ramesha, and B. C. Viraktamath,Directorate of Rice Research, Rajendranagar,Hyderabad 500030, India
Heterosis in rice was reported in 1926.But commercial exploitation tookalmost 5 decades mainly because ofproblems in hybrid seed production.China was the only country to commer-cially exploit heterosis before 1994.Recently, a few rice hybrids werereleased in India for commercial culti-vation. Although the yield advantage
of rice hybrids is well documented,doubts regarding their yield gains andadaptability over locations still persist.Therefore, we present here the averageyield gains obtained from hybrids andtheir adaptability to locations in India.We used data from 1991 to 1995 onvarious hybrids tested under differenttrials over 12 locations in the wetseason (WS) and at six locations duringthe dry season (DS). The yearwisemean yields of hybrids over locationsand over trials were calculatedseparately for both the WS and DS. Weidentified a hybrid as widely adaptableif it had an overall yield advantage ofmore than 0.7 t ha-1 over the high-
yielding national check variety Jaya,and we calculated the frequency ofsuch hybrids for each season and year.Percentage heterosis was computed foreach year based on the highest yieldinghybrid vs Jaya.
The results showed that the overallmean of experimental hybridsincreased gradually during the WSfrom 4.5 t ha-1 in 1991 to 5.5 t ha-1 in1995; during the DS, yields increasedsimilarly from 5.0 to 6.0 t ha-1. Duringthe same period, the percentage ofwidely adaptable hybrids in the WSincreased from zero to 17%. Similarly,in the DS, it increased from 1.75%(1991-92) to 17% (1994-95). The

12 IRRN 1998
maximum yield gain obtained duringthe WS derived from the performanceof hybrids vs Jaya was 544 kg ha-1 in1991 and 1,522 kg ha-1 in 1995. Similarly,in DS, the yield gain in hybrid rice was846 kg ha-1 in 1991-92 and 1,634 kg ha-1
in 1993-94. The heterosis for grain yieldincreased—from 14% (1991) to 31%(1995) in the WS, and from 8% (1991-92)to 27% (1993-94) in the DS. The resultsindicate that hybrids undoubtedlyhave a yield advantage over high-yielding varieties such as Jaya. Themargin of yield gain increased over theyears. The diversity of material addedover time may be the reason for suchgains. The hybrids are also as widelyadaptable as Jaya. ■
Yield stability in ricehybrids
M. I. Ahmed, C. H. M. Vijayakumar,B. C. Viraktamath, M. S. Ramesha, and S. Singh,Directorate of Rice Research, Rajendranagar,Hyderabad 500030, India
Hybrid rice technology is considered tohave the most potential and be readilyexploitable to further raise the yieldceiling in the irrigated ecosystem. Butsuccessful exploitation over a large areadepends on identifying stable hybrids.Yield heterosis is reported to be a varia-ble trait, which depends not only onparent combinations but also on envi-ronmental conditions. Stability foryield of rice hybrids is therefore ana-lyzed. We examined the stability ofhybrids for yield in two sets of environ-ments categorized as low and high.Grain yield data of 10 trials conductedbetween the 1994 wet season (WS) and1995 WS were used. Locations with amean yield lower than the grand meanyield were termed low environments,whereas those with a mean yield higherthan the grand mean yield were identi-fied as high environments. We ana-lyzed grain yield data on eight hybridsin the 1994-95 WS and dry seasontested along with inbred Jaya, Rasi, andIR36 at five locations. The mean yieldand coefficient of variation over
seasons and locations were calculatedfor both hybrids and inbreds to findenvironment-dependent heterosis.
The mean grain yield advantage ofthe best-yielding hybrid over Jaya wascalculated for each environment. Theresults indicated that the stability ofrice hybrids for grain yield is compa-rable with that of the best inbred checkvariety, Jaya. Some of the hybrids, suchas IR58025A/IR34686 and IR58025A/IR29723, showed better yield stabilityover seasons compared with Jaya. Butthe yield heterosis of the hybrids wasbetter expressed in environmentscategorized as high. ■
Exploiting heterosis usingTGMS lines in intra- andintersubspecific hybridi-zation
O. U. K. Reddy, Directorate of Rice Research(DRR), Rajendranagar, Hyderabad 500030;E. A. Siddiq, Indian Council of AgriculturalResearch, Krishi Bhavan, New Delhi 110001;J. Ali, Crop Improvement Department, Agri-culture College and Research Institute, NavalurKuttapattu, Trichy 620009; A. J. Hussain, P. Arti,M. I. Ahmed, and N. P. Sarma, DRR,Rajendranagar 500030, India
The performance of various thermo-sensitive genic male sterile (TGMS)lines was assessed in order to use themin two-line intra- and intersubspecifichybridization with wide compatibilitylines. Some 26 lines comprising TGMSlines, indica accessions, and japonicawith wide compatibility (WC) gene(s)were used to make 65 hybrids: 31 wereindica/indica and 34 were indica/japo-nica (WC). Standard heterosis was re-corded up to 58%. Seed yield (g plant-1)ranged from 2.3 to 31.3. The number ofheterotic combinations was higher inintersubspecific cross combinations.Spikelet fertility ranged from 2.7 to88.4% in intersubspecific crosses andfrom 3.6 to 96.0% in intrasubspecificcrosses. The WC lines were not univer-sally compatible in all combinations.The TGMS line ID/24 was completely
compatible with all WC as well as withother indica accessions.
A molecular phylogenetic map wasconstructed involving all 26 lines basedon the polymorphism generated by 10random primers—OPD3, OPU7,OPU14, OPZ18, OPZ19, OPZ20, OPA7+ OPA6, and OPA12 + OPD5. By usingthe un-weighted pair-group method,arithmetic average (UPGMA), adendrogram was made, taking intoconsideration the distance calculatedbased on Jaccard similarity indices. Thegenetic distances were from 0.2 to 0.88.The height in hybrids increased as thegenetic distance between the parentswidened. With genetic distance > 0.70among parents, spikelet fertility andgrain yield plant-1 decreased consi-derably. Heterotic combinations wereobserved when the genetic distanceranged from 0.4 to 0.7 among parents. ■
Developing CMS andrestorer lines of tropicalrice hybrids
C. L. Casal, Jr. and S. S. Virmani, IRRI
The successful development andcultivation of hybrid rice in China in1976 encouraged IRRI scientists andcollaborating countries to intensify thisresearch in 1980. Because the Chinesehybrids and parental lines were notadapted to the tropics, IRRI begancollaborating with several tropical rice-growing countries to develop suitableparental lines and hybrids. By 1989,IRRI developed two commerciallyusable cytoplasmic male sterile (CMS)lines that, when combined with easilyavailable restorers among elite tropicalindica rice cultivars, producedhundreds of experimental rice hybrids.Some of these yielded about 1 t ha-1
more than inbred checks in nationaltrials under irrigated conditions. By1994, India, Vietnam, and the Philip-pines had released some promising ricehybrids for commercial cultivation byfarmers. New and better CMS lines arenow available in the genetic back-ground of irrigated, rainfed, boro, and

Vol. 23, No. 1 13
aromatic rice cultivars. Thecytoplasmic base has been diversifiedto protect future rice hybrids fromoutbreaks of disease or insect pests.Breeding indica/tropical japonicahybrids has begun with thedevelopment of CMS lines in thebackground of tropical japonica. ■
Developing Pusa 3A, aBasmati CMS line
F. U. Zaman, M. J. Abraham, U. S. Natarajan,A. Mahendru, and F. Mohammad, Division ofGenetics, Indian Agricultural Research Institute,New Delhi 110012, India
Basmati rice occupies a significantplace in India’s rice economy onaccount of its high export potential.With the release of Pusa Basmati 1, thefirst high-yielding semidwarf Basmativariety meeting quality standards,India leaped ahead in Basmati riceproduction. Further improvement inBasmati yield may be possible with theadoption of hybrid rice technology.
Various Basmati rice lines weretestcrossed with IR58025A. PusaBasmati 1 showed complete sterilityand proved to be a perfect maintainerof wild abortive (WA) cytosterility. Thepollen sterility in the F1 hybrid was100%. The F1 was backcrossed to PusaBasmati 1 and a BC1 population ofabout 600 plants was raised. All theplants showed 100% pollen sterility.Segregation was observed for variousplant characters. Twenty BC1 plantsthat were closer to Pusa Basmati 1 forvarious characters were backcrossed toPusa Basmati 1. About 30 seedlings ineach of the 20 BC2 progenies showed100% pollen sterility. Cooking qualityof the 20 pollen donors was analyzed.The best five donors showing goodcooking quality were selected on thebasis of their similarity to Pusa Basmati1 for various agronomic characterssuch as plant height, panicle length,spikelet morphology, and days to 50%flowering. These plants were againbackcrossed to the selected five pollendonors, thus maintaining the identity
of the pollen donors for the respectivepopulation. The BC3 plants raised fromthe above crosses were screened forpollen sterility; all the plants were 100%sterile. Three plants were selected fromeach BC3 progeny row and these wereagain backcrossed to their respectivepollen parent from Pusa Basmati 1.Progenies from pollen plants No. 1 and3 (Pusa Basmati 1-1 and Pusa Basmati1-3) had all the morphological charac-ters of Pusa Basmati 1. They werefurther backcrossed to their respectivepollen plants—Pusa Basmati 1-1 andPusa Basmati 1-3, both with excellentcooking quality traits. The CMS linederived from Pusa Basmati 1-3 hasbeen named Pusa 3A. This line showsstable sterility under different environ-mental conditions at New Delhi andAduthurai, Tamil Nadu, during thetwo different rice seasons. This line isnow being used extensively for test-crossing with both Basmati and non-Basmati male parents. A few promisingcombinations with Basmati restorershave been identified. It was also foundthat heterosis with non-Basmati re-storers was far greater than withBasmati restorers for various charac-ters, including yield. ■
Screening rice genotypesfor thermosensitive genicmale sterility reaction
R. V. Kumar, P. V. Satyanarayana, and M. S. Rao,Andhra Pradesh Agricultural University,Agricultural Research Station, Maruteru 534122,India
Two-line rice hybrids were found to bemore heterotic than three-line hybrids.Constraints of the three-line system canalso be avoided in the two-line system.Any rice variety can be used as a pollenparent in the two-line system irrespec-tive of the presence of a restorer gene.The two-line system also offers theadvantage of easy seed production. Forphotoperiod-sensitive genic malesterility (PGMS) vs thermosensitivegenic male sterility (TGMS), the TGMSsystem is more useful for the weather
conditions that prevail in southernIndia. We therefore screened somediverse genotypes to identify potentiallines. Staggered sowings and plantingswere done during the 1995-96 wet anddry seasons. We carried out pollenstudies on these lines and also re-corded spikelet fertility. Data on pollensterility were correlated with mini-mum, maximum, and daily meantemperatures during the critical periodof sensitivity of the lines, that is, 10 dfrom the third day after panicleinitiation.
During the 1995 wet season, oneTGMS line—IR68949-5-31-34—exhi-bited complete pollen and spikeletsterility for a period of 51 consecutive d.The mean daily temperatures duringthis 51-d period were between 27.1 and28.4 °C. The same line transformed topartial fertility when the averagetemperatures reached 26 °C and below.
During the 1996 dry season, asecond TGMS line, IR68945-33-4-14-40,was found to be completely sterile for aperiod of 15 d. The mean daily tempe-rature regimes at the critical period ofsensitivity ranged from 23.5 to 26.5 °C.Later, this line transformed to partial orcomplete fertility. The two TGMS linesidentified in this study may be used todevelop new two-line rice hybrids forenhancing heterosis and thus produc-tion and productivity of the rice crop. ■
Identifying parental linesin hybrid rice by RAPDfingerprinting
P. Arti, V. V. Shenoy, and N. P. Sarma, Directorateof Rice Research, Rajendranagar, Hyderabad500030, India
The commercial production of hybridrice hinges on three-line breeding, malesterile, maintainer, and restorer lines.Developing and producing newparental lines are essential for thesustainability of hybrid rice. Charac-terizing these lines based on morpho-logical and phenological charactersconsumes time, lacks sufficientresolving power, and is influenced by

14 IRRN 1998
the environment. Therefore, we need tocharacterize parental lines, especiallyrestorers, using biotechnological tools.
DNA was extracted from commonlyused parental stocks (CMS linesIR58025A and IR62829A; their isonu-clear maintainer (B) lines IR58025B andIR62829B; and elite restorer (R) linesIR40750, IR9761, IR34686, and IR10198.These extracted DNA samples wereamplified by polymerase chain reactionusing 20 arbitrary oligo-nucleotideprimers. The amplified products wereanalyzed on agarose gel and scored forthe presence or absence of bands. Withselected primers (OPA-7, OPA-12,OPA-20, OPB-19, OPU-17, and OPW-4),sufficient polymorphism could bedetected to allow identification of indi-vidual stocks. The isonuclear main-tainer and the CMS lines were, how-ever, indistinguishable. The randomlyamplified polymorphic DNA (RAPD)analysis is a potentially simple, rapid,and reliable DNA finger-printingmethod for identifying parental stocksand determining the parentage of ricehybrids. As this molecular techniqueverifies the degree of dissimilaritybetween the parental lines, the RAPDanalysis may also help in identifyingnew potential combina-tions based ongenetic divergence. ■
Exploiting the in vitroovary culture technique tobreed rice hybrids
Li Rongbai, M. P. Pandey, S. K. Pandey,D. K. Dwivedi, and Ashima, Genetics and PlantBreeding Department, G. B. Pant University ofAgriculture and Technology, Pantnagar 263145,India
The thermosensitive genic male sterility(TGMS) system has a high potential todevelop two-line hybrids, which aremore economical for seed production.The ovary culture technique can alsohelp to improve or develop new linesfor several economic traits. Wetherefore cultured unpollinated ovariesfrom plants of five crosses involvingTGMS UPRI 95-140 (P1), the good
ideotype UPRI 95-117 (P2), and main-tainers UPRI 95-139 (P3) and UPRI 95-130 (P4). Quality rices—Basmati 385(P5), Haryana Basmati 1 (P6), and UPRI95-145 (P7)—were also cultured. Thecultured ovary from hybrid plants P1/P2 pretreated at 8 °C for 14 d on an N6medium supple-mented with 500 mglactoprotein hydrolysate (LH) L-1, 4 mg2,4-D L-1, 2 mg NAA L-1, and 1 mg BA L-1
produced 0.5% calli. Regeneration ofthese calli on an MS medium contain-ing 500 mg casein acid hydrolysate L-1,0.5 mg NAA L-1, and 1.5 mg BA L-1
produced four clumps of completelygreen plantlets and one partially greenplantlet. One of these green plantletsshowed the TGMS trait. It was com-pletely spikelet sterile during thepanicle heading period between 15 Junand 5 Sep, but was partially fertile andset seed after 18 Sep. Seed set was 0.5,1.7, 4.8, 12.3, and 6.4% at heading on 18Sep, 1 Oct, 12 Oct, 26 Oct, and 1 Nov1995, respectively. The ovary culture-derived line was dwarf, with an inter-mediate plant type, and flag leaf and
Pest resistance — diseases
panicle length similar to those of thefemale parent. But this line producedmore spikelets.
The unpollinated ovaries of F1 plantsderived from maintainer/quality ricescultured on an N6 medium supple-mented with 0.5 mg 2,4-D L-1, 4 mgNAA L-1, and 1.0 mg BA L-1 producedcalli. Callus induction was 3.33, 2.67,1.33, and 2.00% for P5/P3, P3/P5, P6/P4, and P7/P4, respectively. But plant-lets were regenerated only from P5/P3(46.7%) and P3/P5 (50.0%) on an N6regeneration medium supplementedwith 500 mg LH L-1, 0.5 mg NAA L-1,and 2 mg kinetin L-1. The performanceof seven H1 clones of the former crossshowed four clones with long, boldgrain and a nontranslucent endosperm,whereas clones 6 and 7 had long,slender grains and a translucent endo-sperm free of an opaque area and werecomparable in quality with Basmati385. Exploitation of these lines to im-prove quality and yield potential in ahybrid rice breeding program is inprogress. ■
A variety with durableresistance to rice blast
Shao-Chuan Zhou, Xiao-Yuan Zhu, and Qi-YunYang, Guangdong Academy of AgriculturalSciences, Guangzhou 510640, China
San-Huang-Zhan No. 2 (SHZ), a ricevariety developed by the GuangdongAcademy of Agricultural Sciences, wascultivated on 1,533 ha in 1986. It thenspread from 8,000 ha in 1987 to 10,977ha in 1995 and showed excellent resis-tance to blast in an environment favora-ble to blast in Guangdong, China.Therefore, we assumed that SHZ is avariety with durable resistance to blast.
Looking at the resistance per-formance of SHZ, we found that itsresistance spectrum (RS) was morethan 95% from 1985 to 1995 (Table 1). In1995, the RS of SHZ was 99.5% underartificial inoculation with 220 isolates inGuangdong and 95.2% with 124
isolates of Pyricularia grisea from eightrice-growing zones in China. Further-more, we screened a stable isolate (GD-V1) that is compatible to SHZ toinvestigate the reaction of SHZ com-pared with the reaction of the resistantcheck IR36 and the susceptible checkB40. Entries were inoculated at the 5-leaf stage. Lesion density (LD) andlesion size (LS) were scored by themethod of Roumen and diseased leafarea (DLA) was measured by themethod of Notteghem. To make a bettercomparison, we converted the ob-served data (OD) of LD, LS, and DLAinto relative value (RV) on the basis ofB40, and computed the mean of LD, LS,and DLA. At the same time, resistanceof the varieties was assessed in fields ineight rice-growing zones in China bythe Standard of the National Blast Co-research Group. Results (Table 2)indicated that the resistance of SHZand IR36 is different because SHZ
Vol. 23, No. 1 15
Table 1. Resistance spectrum (%) of San-Huang-Zhan No. 2 from 1985 to 1995 in Guangdong, China.
Year 1985 1986 1987 1989 1991 1992 1993 1994 1995
RS 100 100 98 95 96.5 98 98 98 99.5
Table 3. Reaction to ZB13 blast isolate of F1 and F2 plants from crosses of San-Huang-Zhan No. 2 (SHZ).
Cross F1 F2 reactionreaction
Ra S Expected ratio χ2 P
CO 39/SHZ R 331 10 63:1 3.31 0.05-0.1B40/SHZ R 228 8 63:1 0.54 0.25-0.5SHZ/CO 39 R 401 0SHZ/B40 R 601 0
aR = resistant, S = susceptible.
Table 2. Qualitative and quantitative resistances of San-Huang-Zhan No. 2 (SHZ).
Resistance LD (lesions LS DLA Field observationCultivar spectrum (%) cm–1) (mm) (%)
LBe PBf
Guangdonga Chinab ODc RVd OD RV OD RVMean Maximum Mean Maximum
SHZ 99.5 95.2 0.3 4 1.3 19 8.5 10 1.3 4 1 3IR36 (check) 58.2 - 0.4 6 1.4 20 24.6 30 3.0 9 3.2 9B40 (check) - - 6.6 100 6.9 100 82.0 100 - - - -
aIncluding 220 isolates in Guangdong. bIncluding 124 isolates in China. cOD = observed data = mean of three experiments with 6replications. dRV = relative value = tested data of cultivar/tested data of B40 × 100. eLB = leaf blast. fPB = panicle blast.
exhibited a wide qualitative RS and ahigh quantitative resistance for threefactors—LD, LS, and DLA—whereasIR36 appeared to have a narrow quali-tative RS but high quantitative resis-tance under a wide range of popula-tions of P. grisea in different latitudes ofChina. Quantitative resistance plays akey role in the durability of resistance.
The genetic experiment showed thatthree independently dominant genescontrol the blast resistance of SHZ andthere is another gene present in thecytoplasm of SHZ as well (Table 3).
IR36 represents an indica varietywith durable blast resistance under tro-pical irrigated conditions. Some re-searchers concluded that the partialresistance of IR36 is associated withblast durability. Other researchersshowed that IR36 had at least threemajor genes for complete resistance totwo Philippine blast isolates. The inhe-ritance of partial blast resistance in IR36is most likely polygenic. SHZ seemedto have three major genes for completeresistance to blast, but some questionsremain. Is the durability of SHZ inChina mainly due to major genes ornot? What is the function of anothergene present in the cytoplasm for blastdurability? Further research is neededto answer these questions. ■
Multilocational evaluation ofpromising advanced breedinglines for resistance to rice tungroviruses
R. C. Cabunagan, E. R. Angeles, E. R. Tiongco(current address: Philippine Rice Research Insti-tute, Maligaya, Muñoz, Nueva Ecija), S. Villareal,O. Azzam, P. S. Teng, and G. S. Khush, IRRI;T. C. B. Chancellor, Natural Resources Institute,University of Greenwich, U.K.; X. H. Truong andS. Mancao, Philippine Rice Research Institute,Maligaya, Muñoz, Nueva Ecija, Philippines;I.G.N. Astika, Food Crop Protection Centre VII,Denpasar, Bali, Indonesia; A. Muis, MarosResearch Institute for Maize and Other CerealCrops, Maros, South Sulawesi, Indonesia;A. K. Chowdhury, Bidhan Chandra KrishiViswavidyalaya University, Kalyani, WestBengal, India; T. Ganapathy and N. Subra-
manian, Tamil Nadu Agricultural University,Coimbatore, Tamil Nadu, India
An important objective of IRRI’sbreeding program for irrigated riceecosystems is to develop varieties withresistance to rice tungro viruses in orderto achieve durable resistance to tungrodisease. We conducted this study tomonitor tungro virus and leafhoppervector variability between test locations,based on the reaction of selected varie-ties and advanced breeding lines withdifferent sources of resistance. Theadvanced breeding lines were IR68305-18-1 (IR64*4/Balimau Putih); IR71026-3-2-4-3-5-2 (IR1561-228-3-3*3/Oryzalongistaminata); IR69705-1-1-3-2-1(IR1561-228-3-3*2/Utri Merah);
IR71030-2-3-2-1 (IR1561-228-3-3*6/ARC11554). IR64 was used as a suscepti-ble check and IR62 as a vector-resistantcheck with field resistance to tungro.
The table shows locations, seasons,and years of tests. At each location, weused a randomized complete blockdesign with four replications. The plotsize was 8 m × 8 m with 2 m betweenplots. We transplanted 21-d-oldseedlings at 20 cm × 20 cm spacing, with2-3 seedlings hill-1. The plants wereexposed to natural infection with tungroviruses in the field. We assessed plantsfor disease symptoms and sampledleaves to detect tungro viruses byenzyme-linked immunosorbent assay(ELISA) at 30-35 and 55-60 d aftertransplanting (DAT). We recorded
16 IRRN 1998
disease and sampled leaves in sixquadrats of 4 × 4 hills arranged in a“W” pattern in each plot. We collectedvector leafhoppers using 10 sweeps of a30-cm-diameter insect net in each ploton the same dates as for recordingdisease.
Tungro disease incidence was low inMaros, Indonesia, as evidenced by thereaction of IR64, but was high enoughat all other locations to allow the per-
Averagea percentage infection with rice tungro bacilliform virus (RTBV), rice tungro spherical virus (RTSV),visual disease incidence, and number of green leafhoppers (GLH) in different varieties and advanced breedinglines planted in field trials in the Philippines, Indonesia, and India in 1995-96.
Infection (%)Test location/season Varieties/lines GLH
BBb SS Visual (no.)c
PhilippinesMaligaya, IR62 0.8 34.2 0.3 24.6Nueva Ecija IR64 27.0 67.3 23.1 169.51995 WSd, 1996 WS IR68305-18-1 2.0 55.2 0.1 145.4
IR71026-3-2-4-3-5-2 0.9 28.5 0.1 134.8IR69705-1-1-3-2-1 2.3 3.4 0.2 174.8IR71030-2-3-2-1 0.6 11.1 0.1 52.1
Midsayap, IR62 8.9 48.8 17.8 10.2North Cotabato IR64 80.8 89.1 92.2 100.71995 WS, 1996 DS, IR68305-18-1 18.0 72.3 21.2 42.01996 WS IR71026-3-2-4-3-5-2 38.8 74.1 41.6 44.8
IR69705-1-1-3-2-1 6.4 6.0 1.8 35.1IR71030-2-3-2-1 6.1 13.4 20.9 12.8
IndonesiaCeluk, IR62 3.8 9.1 2.9 2.0Bali IR64 48.0 64.8 64.2 5.61995 DS, 1996 WS IR68305-18-1 18.2 41.4 15.3 1.4
IR71026-3-2-4-3-5-2 29.7 52.1 31.3 5.2IR69705-1-1-3-2-1 3.7 2.0 1.2 2.7IR71030-2-3-2-1 1.8 4.4 0.7 1.5
Maros, IR62 3.3 6.1 0.6 10.4South Sulawesi IR64 8.2 13.4 3.5 15.21995 DS, 1996 WS IR68305-18-1 2.2 5.8 0.2 14.1
IR71026-3-2-4-3-5-2 3.4 5.6 0.2 13.8IR69705-1-1-3-2-1 4.4 4.3 0.0 11.0IR71030-2-3-2-1 2.2 2.6 0.0 12.0
IndiaChakdaha, IR62 1.5 7.8 9.8 26.6West Bengal IR64 4.3 18.0 20.0 27.91996 WS IR68305-18-1 1.1 5.1 10.8 35.9
IR71026-3-2-4-3-5-2 0.8 6.0 23.8 20.5IR69705-1-1-3-2-1 0.3 1.1 9.5 24.9IR71030-2-3-2-1 6.1 31.1 16.9 26.0
Tirur, IR62 2.3 14.5 2.5 10.4Tamil Nadu IR64 10.0 24.9 16.8 11.31996 DS, 1996 WS IR68305-18-1 1.1 14.0 2.2 14.4
IR71026-3-2-4-3-5-2 1.4 16.6 2.2 10.0IR69705-1-1-3-2-1 0.6 8.6 2.3 15.1IR71030-2-3-2-1 5.9 15.8 7.7 12.4
aAverage across all seasons and computed from the mean of two samplings at 30-35 and 55-60 d after transplanting. bBB = totalRTBV from BS and B alone, SS = total RTSV from BS and S alone. Infection with tungro viruses was assessed by enzyme-linkedimmunosorbent assay, visual incidence was assessed based on characteristic tungro symptoms of stunting and yellowing. cCountbased on 10 sweeps of a 30-cm-diameter insect net in each plot. dWS = wet season, DS = dry season.
formance of the test entries to beevaluated effectively (see table). Tungroincidence was the highest in Celuk,Indonesia, and Midsayap, Philippines.Locational differences in the reaction ofthe test lines and varieties were ob-served only in IR71030-2-3-2-1, whichhad a relatively high infection withtungro viruses both in Chakdaha andTirur, India, compared with otherlocations (see table).
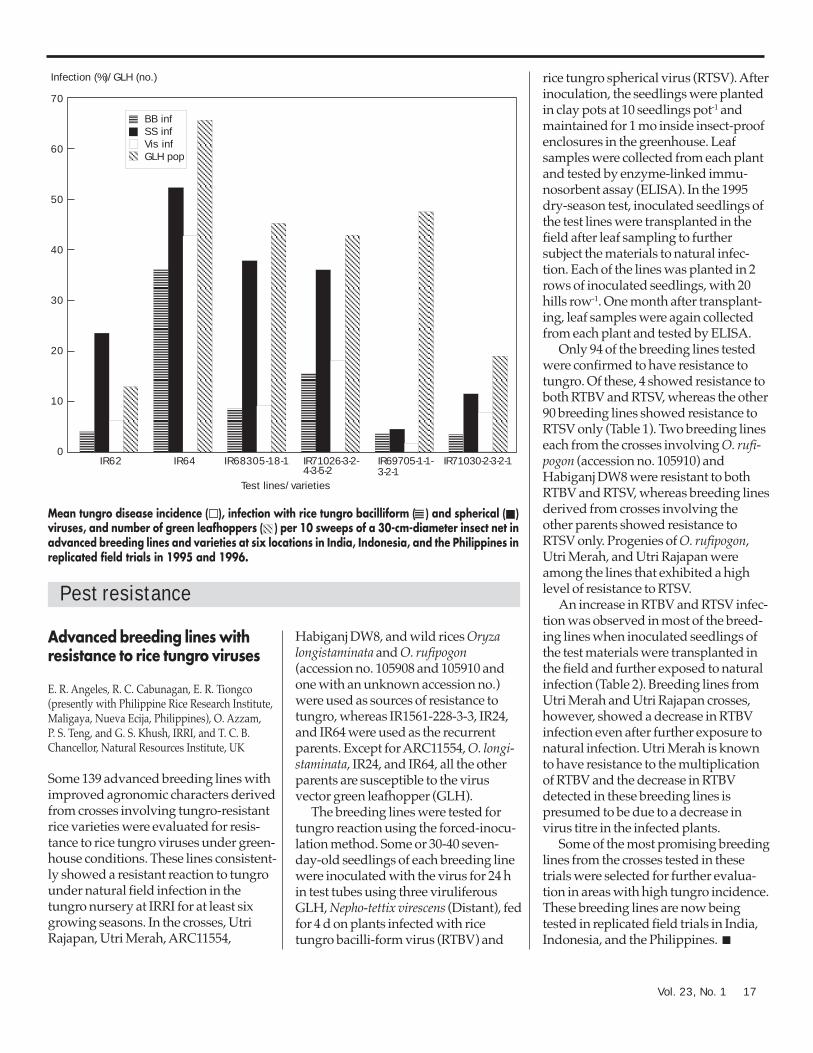
The figure shows the averagereactions of the test lines and varieties,regardless of test location and season.Infection with rice tungro bacilliformvirus (RTBV) was the lowest inIR69705-1-1-3-2-1, IR71030-2-3-2-1, andIR62 at 3-4%. In contrast, a mean RTBVinfection of 16% was recorded inIR71026-3-2-4-3-5-2. Mean infectionwith rice tungro spherical virus (RTSV)was the lowest in IR69705-1-1-3-2-1 andIR71030-2-3-2-1 at 5% and 12%,respectively. RTSV infection in IR71026-3-2-4-3-5-2 and IR68305-18-1 was 36%and 38%, respectively. Tungroincidence scored visually was 2% inIR69705-1-1-3-2-1 and was less than10% in IR62, IR71030-2-3-2-1, andIR68305-18-1 (see figure). Tungroincidence on the susceptible check,IR64, was 44%. Green leafhoppernumbers were the lowest on IR62 andIR71030-2-3-2-1 and were similar tothose on the susceptible IR64 on theother lines.
The most promising advancedbreeding line was IR69705-1-1-3-2-1,which showed tolerance for RTBV andresistance to RTSV at all test locations.This indicates that the virus resistancewas successfully transferred from itsparent, Utri Merah, and that thisresistance is likely to be effectiveagainst tungro disease at a wide rangeof locations. IR71030-2-3-2-1 showedresistance to both RTBV and RTSV atmost sites, but infection was relativelyhigh in Chakdaha and Tirur. The virus-resistant parent of this line, ARC11554,originates from India. Further evalu-ation needs to be done to confirm theresults. ■
Multiple submissions. Normally, onlyone report for a single experiment willbe accepted. Two or more items aboutthe same work submitted at the sametime will be returned for merging.Submitting at different times multiplenotes from the same experiment ishighly inappropriate. Detection willresult in the rejection of allsubmissions on that research.
Vol. 23, No. 1 17
70
60
50
40
30
20
10
0
Infection (%)/GLH (no.)
BB infSS infVis infGLH pop
IR62 IR64 IR68305-18-1 IR71026-3-2-4-3-5-2
IR69705-1-1-3-2-1
IR71030-2-3-2-1
Test lines/varieties
Pest resistance
Advanced breeding lines withresistance to rice tungro viruses
E. R. Angeles, R. C. Cabunagan, E. R. Tiongco(presently with Philippine Rice Research Institute,Maligaya, Nueva Ecija, Philippines), O. Azzam,P. S. Teng, and G. S. Khush, IRRI, and T. C. B.Chancellor, Natural Resources Institute, UK
Some 139 advanced breeding lines withimproved agronomic characters derivedfrom crosses involving tungro-resistantrice varieties were evaluated for resis-tance to rice tungro viruses under green-house conditions. These lines consistent-ly showed a resistant reaction to tungrounder natural field infection in thetungro nursery at IRRI for at least sixgrowing seasons. In the crosses, UtriRajapan, Utri Merah, ARC11554,
Mean tungro disease incidence ( ), infection with rice tungro bacilliform ( ) and spherical ( )viruses, and number of green leafhoppers ( ) per 10 sweeps of a 30-cm-diameter insect net inadvanced breeding lines and varieties at six locations in India, Indonesia, and the Philippines inreplicated field trials in 1995 and 1996.
Habiganj DW8, and wild rices Oryzalongistaminata and O. rufipogon(accession no. 105908 and 105910 andone with an unknown accession no.)were used as sources of resistance totungro, whereas IR1561-228-3-3, IR24,and IR64 were used as the recurrentparents. Except for ARC11554, O. longi-staminata, IR24, and IR64, all the otherparents are susceptible to the virusvector green leafhopper (GLH).
The breeding lines were tested fortungro reaction using the forced-inocu-lation method. Some or 30-40 seven-day-old seedlings of each breeding linewere inoculated with the virus for 24 hin test tubes using three viruliferousGLH, Nepho-tettix virescens (Distant), fedfor 4 d on plants infected with ricetungro bacilli-form virus (RTBV) and
rice tungro spherical virus (RTSV). Afterinoculation, the seedlings were plantedin clay pots at 10 seedlings pot-1 andmaintained for 1 mo inside insect-proofenclosures in the greenhouse. Leafsamples were collected from each plantand tested by enzyme-linked immu-nosorbent assay (ELISA). In the 1995dry-season test, inoculated seedlings ofthe test lines were transplanted in thefield after leaf sampling to furthersubject the materials to natural infec-tion. Each of the lines was planted in 2rows of inoculated seedlings, with 20hills row-1. One month after transplant-ing, leaf samples were again collectedfrom each plant and tested by ELISA.
Only 94 of the breeding lines testedwere confirmed to have resistance totungro. Of these, 4 showed resistance toboth RTBV and RTSV, whereas the other90 breeding lines showed resistance toRTSV only (Table 1). Two breeding lineseach from the crosses involving O. rufi-pogon (accession no. 105910) andHabiganj DW8 were resistant to bothRTBV and RTSV, whereas breeding linesderived from crosses involving theother parents showed resistance toRTSV only. Progenies of O. rufipogon,Utri Merah, and Utri Rajapan wereamong the lines that exhibited a highlevel of resistance to RTSV.
An increase in RTBV and RTSV infec-tion was observed in most of the breed-ing lines when inoculated seedlings ofthe test materials were transplanted inthe field and further exposed to naturalinfection (Table 2). Breeding lines fromUtri Merah and Utri Rajapan crosses,however, showed a decrease in RTBVinfection even after further exposure tonatural infection. Utri Merah is knownto have resistance to the multiplicationof RTBV and the decrease in RTBVdetected in these breeding lines ispresumed to be due to a decrease invirus titre in the infected plants.
Some of the most promising breedinglines from the crosses tested in thesetrials were selected for further evalua-tion in areas with high tungro incidence.These breeding lines are now beingtested in replicated field trials in India,Indonesia, and the Philippines. ■

18 IRRN 1998
Table 1. Average percentage infection of rice tungro bacilliform virus (RTBV) and rice tungro spherical virus(RTSV) in advanced breeding lines with different sources of resistance.
Resistant Infectiona (%)Cross lines (no.)
BS B S BB SS
1995 dry seasonb
IR1561-228-3-3*3/O. longistaminata 16 8.3 57.2 2.0 65.5 10.3IR1561-228-3-3*2/Utri Rajapan 15 14.8 72.7 0.2 87.5 15.0IR1561-228-3-3*2/Utri Merah 10 8.6 56.1 2.1 64.7 10.7IR1561-2l28-3-3*4/Utri Merah 10 5.7 53.1 1.7 58.8 7.4IR1561-228-3-3*6/ARC11554 5 5.0 50.0 1.6 55.0 6.6IR1561-228-3-3*4/Habiganj DW8 2 1.5 15.5 1.5 17.0 3.0IR1561-228-3-3*6/Habiganj DW8 2 6.0 61.5 0.0 67.5 6.0
1996 wet seasonIR1561-228-3-3*2/O. longistaminata 2 1.5 69.0 1.5 70.5 3.0IR1561-228-3-3*3/O. longistaminatac 2 1.5 71.5 1.5 73.0 3.0IR1561-228-3-3*2/O. longistaminata//IR24 2 1.5 82.5 0.0 84.0 1.5IR1561-228-3-3*4/O. longistaminata//3*IR24 4 1.5 59.0 0.0 60.5 1.5IR1561-228-3-3*2/Utri Merahc 3 1.0 30.0 0.0 31.0 1.0IR1561-228-3-3*4/Utri Merahc 2 0.0 40.5 0.0 40.5 0.0IR1561-228-3-3*2/Utri Merah//IR24 3 9.0 48.3 0.0 57.3 9.0IR1561-228-3-3*6/ARC 11554c 5 2.4 43.6 0.6 46.0 3.0IR1561-228-3-3*3/Habiganj DW8 1 3.0 47.0 0.0 50.0 3.0IR1561-228-3-3*4/Habiganj DW8c 2 4.0 24.5 4.5 28.5 8.5IR1561-228-3-3*6/Habiganj DW8 1 9.0 53.0 0.0 62.0 9.0IR1561-228-3-3*3/Habiganj DW8//4*IR64 5 3.0 39.8 0.0 42.8 3.0IR1561-228-3-3*2/Utri Rajapanc 2 5.0 77.5 0.0 82.5 5.0IR1561-228-3-3*2/Utri Rajapan//IR24 3 1.0 68.7 0.0 69.7 1.0IR1561-228-3-3/O. rufipogon (Acc. 105910) 1 0.0 76.0 0.0 76.0 0.0IR1561-228-3-3/O. rufipogon (Acc. 105908)//IR24 5 0.0 69.4 0.0 69.4 0.0IR24/O. rufopogon (Acc. 105910)//3*IR64 2 0.0 26.0 0.0 26.0 0.0IR1561-228-3-3//IR42/O. rufipogon (unknown Acc. no.) 5 0.0 60.2 0.8 60.2 0.8
aAv of resistant lines. Infection of tungro viruses was assessed by enzyme-linked immunosorbent assay. BS = RTBV + RTSV, B =RTBV alone, S = RTSV alone, BB = total RTBV (BS + B), SS = total RTSV (BS + S). bTwo inoculation trials conducted. Percentageinfection is av of two trials. cAdvanced progenies of breeding lines identified as resistant in 1995 dry season trial.
Table 2. Average percentage infection of rice tungro bacilliform virus (RTBV) and rice tungro spherical virus(RTSV) in advanced breeding lines inoculated in the greenhouse and subjected to natural infection in the field.
Lines Infection (%)bCross testeda
(no.) BS B S BB SS
IR1561-228-3-3*3/O. longistaminata 4 18.8 (4.6) 55.3 (68.4) 2.8 (0.5) 74.1 (73.0) 21.6 (5.1)IR1561-228-3-3*2/Utri Rajapan 4 32.6 (20.3) 52.8 (71.4) 4.5 (0.1) 85.4 (91.7) 37.1 (20.4)IR1561-228-3-3*2/Utri Merah 4 5.7 (4.3) 43.0 (71.8) 3.2 (0.6) 48.7 (76.1) 8.9 (4.9)IR1561-228-3-3*6/ARC11554 4 8.8 (2.8) 64.0 (59.2) 1.8 (0) 72.8 (62.0) 10.6 (2.8)IR1561-228-3-3*6/Habiganj DW8 4 9.2 (5.0) 59.8 (47.4) 1.6 (0) 69.0 (52.4) 10.8 (5.0)
aTest lines were inoculated with RTBV + RTSV in the greenhouse at 7d after sowing (DAS) and transplanted in the field at 38 DAS.Values represent infection with tungro viruses at 69 DAS. Values in parentheses represent infection with tungro viruses at 38 DAS.bInfection of tungro viruses was assessed by enzyme-linked immunosorbent assay. BS = RTBV + RTSV, B = RTBV alone, S = RTSValone, BB = total RTBV (BS + B), SS = total RTSV (BS + S).
Stress tolerance —adverse soils
Routine research. Reports of screening trials of varieties, fertilizer, croppingmethods, and other routine observations using standard methodologies to establishlocal recommendations are not ordinarily accepted. Examples are single-season,single-trial field experiments. Field trials should be repeated across more than oneseason, in multiple seasons, or in more than one location as appropriate. Allexperiments should include replications and an internationally known check orcontrol treatment.
Seasonal differences in irontoxicity tolerance of lowland ricecultivars
L. Sahrawat and B. N. Singh, West Africa RiceDevelopment Association (WARDA), 01 BP 2551Bouake 01, Côte d’Ivoire, West Africa
Iron toxicity is a major stress and yield-reducing factor in irrigated and rainfedlowlands in West Africa. Varietaltolerance is the most practical and cost-effective means of reducing irontoxicity in iron-toxic soils. We havebeen evaluating lowland rice cultivarsfor iron toxicity tolerance at Korhogo,Côte d’Ivoire, a site with a consistentand high iron toxicity pressure onplants. Because irrigation water isavailable, it was possible to evaluategermplasm in both the wet and dryseasons.
We have observed that iron toxicitypressure is higher in the dry seasonthan in the wet season at Korhogo. Inthe 1994 wet season (Jul-Nov 1994) and1995 dry season (Dec 1994-Apr 1995),we evaluated the performance of 12elite selections (based on evaluationsmade in 1992 and 1993). To determinethe yield potential of these cultivars,they were also grown at Mbe, whichprovides a lower level of stress.
Experiments at both sites used a ran-domized complete block design withfour replications. The plot size was 12m2 at Mbe and 24 m2 at Korhogo. Allplots received 20-36-36 kg NPK ha-1 as abasal application. A total of 80 kg N ha-1
was added to each plot. The soil at Mbewas an Alfisol (pH 6.2; organic carbon,1.8%; extractable Fe, 93 mg kg-1) withsandy clay-loam texture; the soil atKorhogo was an Ultisol (pH 5.7;organic carbon, 1.02%; extractable Fe,154 mg kg-1) with sandy loam texture.
The cultivars evaluated at Korhogohad large differences in yield duringthe wet and dry seasons (see table). Irontoxicity scores at Korhogo, based onbronzing symptoms on the rice plant
Vol. 23, No. 1 19
Performance of 12 lowland rice cultivars evaluated for grain yield (t ha–1) and iron toxicity score (ITS) at siteswith high iron toxicity (Korhogo) and low iron toxicity (Mbe), Côte d'Ivoire, 1994 wet season and 1995 dryseason.
KorhogoMbe
Cultivar 1994 wet season 1994 dry season 1994wet season
Grain ITSa Grain ITS Grainyield yield yield
TOX3118-6-E2-3-2 (WITA 1) 6.7 1 5.0 2 7.6CK4 6.1 1 5.0 2 8.3CK73 4.9 2 4.9 3 4.2b
TOX3052-41-E1-2 6.3 1 4.8 3 8.1TOX3100-32-2-3-5 (WITA 3) 4.3 3 4.7 3 7.8TOX3050-46-E3-3 6.0 1 4.6 3 6.2TOX3081-36-2-3 6.2 1 4.4 3 6.0TOX3027-43-1-E3-1-1-1 5.5 2 4.1 5 7.5TOX3093-35-2-3-3 (WITA 2) 4.8 2 3.7 5 7.6Suakoko 8 3.7 2 3.3 3 5.3Bouake 189 4.7 3 2.9 7 6.5TOX 3069-66-2-1-6 4.2 5 2.8 7 7.7
LSD (0.05) 1.1 0.8 0.8
aBased on the Standard evaluation system for rice with a scale of 1-9, where 1 = growth and tillering nearly normal and 9 = almostall plants dead or dying. bYield was low because of bird damage.
foliage, were higher (indicating higheriron toxicity) in the dry season than inthe wet season. Iron toxicity in thegrowing season was not observed toany significant extent at Mbe and therice plants grew normally there.
WITA 2, and WITA 3 were clearlyexpressed in the 1994 wet seasonexperiment at Mbe (see table). The iron-toxicity susceptible cultivars Bouake189 and TOX 3069-66-2-1-6 yielded 6.5and 7.7 t ha-1, respectively, at Mbe.
These results indicate that irontoxicity, coupled with dry-seasonenvironmental conditions at Korhogo,reduces the grain yield of both iron-toxicity-tolerant and -susceptiblecultivars. The high temperatures in thedry season affect crop physiology,especially during grain maturity. Hightemperature may also enhance trans-piration rates of rice plants, whichcould cause a higher uptake of iron. It isalso known that in a growing mediumwhere iron in solution is high (which isexpected in the dry season because ofthe high prevailing temperatures), ironuptake increases with higher transpira-tion rates because of a passive uptakemechanism. In addition, watersupplied by rainfall in the rainy seasonwould dilute the iron in a soil solutionand could provide some relief fromiron toxicity. ■
Seasonal differences in yield werelarger for susceptible cultivars such asBouake 189 and TOX 3069-66-2-1-6than for iron-tolerant cultivars such asCK 4, Suakoko 8, and WITA 3. The yieldpotentials of cultivars CK 4, WITA 1,
Correlation of some varietalcharacteristics with grain yieldand stress tolerance index undersaline conditions
L. M. González and R. Ramírez, NuclearLaboratory of the Agricultural Research Institute“Jorge Dimitrov,” Gaveta Postal 2360, Bayamo85100, Granma, Cuba
Rice production in saline soils could beincreased if salt-tolerant genotypeswere developed, but progress towardcombining superior tolerance withgood yield potential is slow, largelybecause of the nonavailability ofreliable and efficient selection criteria.Selecting directly for grain yield undersaline conditions may be limited by thelow heritability of observed variationand failure to distinguish betweenyield potential and capacity to copewith adverse stress factors.
We evaluated the correlation of 26characteristics with grain yield andstress tolerance index (average grainyield under stress in relation to thecorresponding controls taken as one) inrice under saline conditions to ascertainthe role of these traits in indirect selec-tion for yielding ability under theseconditions and in measuring varietalratings.
We studied 20 rice varieties underlaboratory conditions and in the field.In petri dishes with NaCl at electricalconductivity of 12 dS m-1 (stress) anddistilled water (0.02 dS m-1 as a control),30 seeds of each genotype were placedin petri dishes with filter paper, withfour replications. After 48 h, seed wateruptake, peroxidase, catalase, alpha,beta, and total amylase isozyme activi-ty in germinating seeds, seed respira-tion, and seed redox potential wereevaluated. After 7 d, seed germination,seedling height, root length, fresh and
dry weight of seedlings, leaf respira-tion, total water content, relative leafwater content, transpiration, sap cellconcentration, proline content, andtotal chlorophyll content were mea-sured. In the field, the genotypes weresown in specially designed 3 m × 3 m ×1 m test plots in saline soil (9 dS m-1)and in nonstress productive soil overthree seasons. The experiment was laidout in a randomized block design withfour replications. Normal cultural prac-tices were followed. We collected dataon agronomic attributes (plant height,panicles plant–1, panicle length andweight, filled grains panicle–1, andgrain yield) from 10 plants selected atrandom in each replication. We com-puted the correlation coefficient of alltraits estimated.
Seed water uptake, peroxidaseactivity, seed germination, seedlingheight, root length, seedling dryweight, total water content, and

20 IRRN 1998
relative leaf water content were highlycorrelated with the saline stresstolerance index (see table), indicatingthat measuring varietal ratings for salttolerance at the early stage of growthvia these traits was likely to be effective.
At the late stage of growth, plantheight and panicle weight correlatedhighly and significantly with grainyield under saline conditions, as well aswith the saline stress tolerance index,showing that varietal evaluation andindirect selection for grain yield insaline soil via these traits would be aneven better method. ■
Integrated germplasm improvement — uplandCorrelation of some characteristics of rice varietiesgrown in saline conditions with grain yield and salinestress index.
Correlation withCharacteristic
Grain Saline stressyield tolerance index
Seed water uptake 0.47* 0.51*Peroxidase 0.42* 0.69*Catalase 0.15 0.36Alpha amylase 0.10 0.11Beta amylase 0.02 0.15Total amylase 0.47* 0.15Seed respiration –0.18 0.28Seed redox potential 0.02 0.15Seed germination 0.68** 0.57*Seedling height 0.71** 0.72**Root length 0.62** 0.78**Seedling fresh weight 0.05 0.26Seedling dry weight 0.63** 0.66**Leaf seedling respiration 0.29 0.30Total water content 0.46* 0.65**Relative leaf water content 0.57* 0.62**Transpiration 0.05 0.04Sap cell concentration 0.02 0.01Proline content 0.21 0.39Total chlorophyll content 0.25 0.29Plant height 0.80*** 0.83***Panicles plant–1 0.30 0.29Panicle length 0.32 0.40Panicle weight 0.81*** 0.83***Filled grains panicle–1 0.33 0.301000-grain weight 0.73** 0.38
* = significant at P=0.05, ** = significant at P=0.01, and ***= significant at P=0.001.
Altamirano, Mandisovi, andQuebracho: new rice cultivarsfrom Argentina
J. J. Marassi, A. A. Vidal, and R. Bezus, RiceProgram, Facultad de Ciencias Agrarias yForestales, Universidad Nacional de La Plata,C.C. 31 (1900), Buenos Aires, Argentina
Altamirano P.A. is, because of itsgrowth duration, tolerance for low tem-peratures, and good performance onsaline and alkaline soils, recommendedfor sowing in temperate zones or underthese soil limitations, according to thenew aims of our rice program.
This variety is 0.98 m tall, resistslodging, and has intermediate thresh-ability. It reaches maturity in 133 d andits yield potential is 8,000 kg ha-1. Thecaryopsis is 7.47 mm long and 2.79 mmwide and the ratio of length to width is2.67, matching that of the commercialtype.
Its endosperm has a medium amy-lose content (21%) and an intermediategelatinization temperature. Its proteincontent is 8.35%. It is moderatelyresistant to Pyricularia oryzae Cav.
This variety originated from thecross of experimental line H-115-19-1and variety Yerua P.A. to obtain thehighest quality of grain, yield, and P.grisea Cav. resistance. Line H-115-19-1came from the cross of varieties Dawnand Zenith. Crossing was carried outby the rice program at the Ing. Agr.Julio Hirschhorn Experiment Station. Acombination of mass and pedigreeselection was used.
Because of its earliness, Altamiranowas released for cultivation in areasnear 30 °S in Buenos Aires Province.
Mandisovi P.A. was bred to satisfythe demand for aromatic rices, which,unlike La Calendaria FA, has a caryop-sis that corresponds to the long-widetype. It can be consumed both aspolished or integral rice (this alone ormixed with red pericarp rice).
Mandisovi is 0.83 m tall, resistslodging, and reaches maturity in 135 d.Its yield potential is 6,000 kg ha-1 and ithas intermediate threshability. Mandi-sovi’s caryopsis is 7.64 mm long and 2.7mm wide and its ratio of length to widthis 2.82.
The endosperm contains an interme-diate amylose content (23%) and lowgelatinization temperature. Its proteincontent is 8.80% and it has a weakaroma. Mandisovi has moderateresistance to P. oryzae Cav.
This variety was obtained by crossingAfrican cultivar Chocoto and H161-18//desc./80-17. Line H161-18 was devel-oped by crossing IR1103-15-10 andCalady 40 sel. FA to obtain earliness andhigh protein content. The crossing wascarried out by the rice program. Acombination of mass and pedigreeselection was used. The target area fordiffusion is 32 °S near Villaguay City.
Quebracho P.A. is the first Argentinecultivar of the long-wide type, and it isaromatic and has a red pericarp. Thisrelease aimed to satisfy an increasinginterest in the consumption of specialrices. It is consumed as integral grain,alone or combined with similar rices ofnormal pericarp.
Quebracho is 0.89 m tall and resistslodging. It reaches maturity in 145 d andyields 7,000 kg ha-1. It has intermediatethreshability. The caryopsis is 8.1 mmlong and 2.75 mm wide, and its ratio oflength to width is 2.94.
For quality, it is important to mentionits medium amylose content (22%) andits intermediate gelatinization tempera-ture. Its protein content is 9.68% and it isstrongly aromatic. Quebracho isresistant to P. oryzae Cav.
This cultivar was developed bycrossing Chocoto and H161-18//desc./80-17, in a crossing carried out by therice program. A combination of massand pedigree selection was used. Theprobable target area is 32 °S nearVillaguay City. ■
Multiple submissions. Normally, onlyone report for a single experiment willbe accepted. Two or more items aboutthe same work submitted at the sametime will be returned for merging.Submitting at different times multiplenotes from the same experiment ishighly inappropriate. Detection willresult in the rejection of allsubmissions on that research.
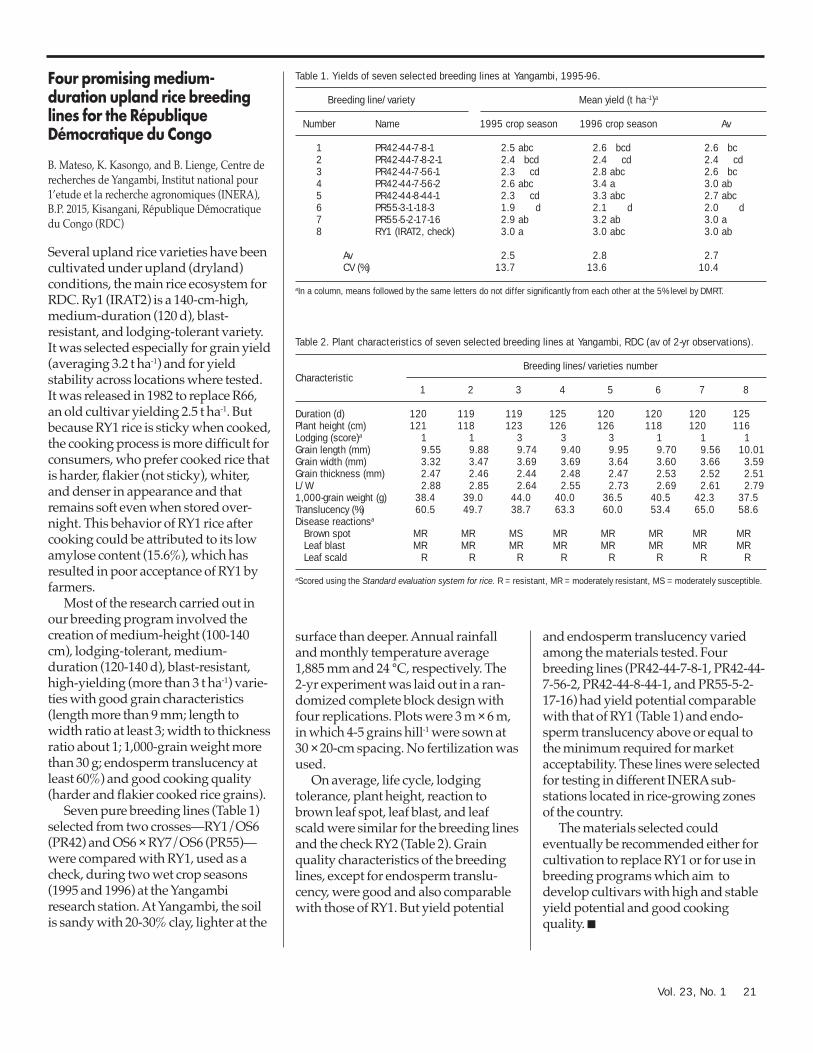
Vol. 23, No. 1 21
Table 1. Yields of seven selected breeding lines at Yangambi, 1995-96.
Breeding line/variety Mean yield (t ha–1)a
Number Name 1995 crop season 1996 crop season Av
1 PR42-44-7-8-1 2.5 abc 2.6 bcd 2.6 bc2 PR42-44-7-8-2-1 2.4 bcd 2.4 cd 2.4 cd3 PR42-44-7-56-1 2.3 cd 2.8 abc 2.6 bc4 PR42-44-7-56-2 2.6 abc 3.4 a 3.0 ab5 PR42-44-8-44-1 2.3 cd 3.3 abc 2.7 abc6 PR55-3-1-18-3 1.9 d 2.1 d 2.0 d7 PR55-5-2-17-16 2.9 ab 3.2 ab 3.0 a8 RY1 (IRAT2, check) 3.0 a 3.0 abc 3.0 ab
Av 2.5 2.8 2.7CV (%) 13.7 13.6 10.4
aIn a column, means followed by the same letters do not differ significantly from each other at the 5% level by DMRT.
Table 2. Plant characteristics of seven selected breeding lines at Yangambi, RDC (av of 2-yr observations).
Breeding lines/varieties numberCharacteristic
1 2 3 4 5 6 7 8
Duration (d) 120 119 119 125 120 120 120 125Plant height (cm) 121 118 123 126 126 118 120 116Lodging (score)a 1 1 3 3 3 1 1 1Grain length (mm) 9.55 9.88 9.74 9.40 9.95 9.70 9.56 10.01Grain width (mm) 3.32 3.47 3.69 3.69 3.64 3.60 3.66 3.59Grain thickness (mm) 2.47 2.46 2.44 2.48 2.47 2.53 2.52 2.51L/W 2.88 2.85 2.64 2.55 2.73 2.69 2.61 2.791,000-grain weight (g) 38.4 39.0 44.0 40.0 36.5 40.5 42.3 37.5Translucency (%) 60.5 49.7 38.7 63.3 60.0 53.4 65.0 58.6Disease reactionsa
Brown spot MR MR MS MR MR MR MR MR Leaf blast MR MR MR MR MR MR MR MR Leaf scald R R R R R R R R
aScored using the Standard evaluation system for rice. R = resistant, MR = moderately resistant, MS = moderately susceptible.
Four promising medium-duration upland rice breedinglines for the RépubliqueDémocratique du Congo
B. Mateso, K. Kasongo, and B. Lienge, Centre derecherches de Yangambi, Institut national pour1’etude et la recherche agronomiques (INERA),B.P. 2015, Kisangani, République Démocratiquedu Congo (RDC)
Several upland rice varieties have beencultivated under upland (dryland)conditions, the main rice ecosystem forRDC. Ry1 (IRAT2) is a 140-cm-high,medium-duration (120 d), blast-resistant, and lodging-tolerant variety.It was selected especially for grain yield(averaging 3.2 t ha-1) and for yieldstability across locations where tested.It was released in 1982 to replace R66,an old cultivar yielding 2.5 t ha-1. Butbecause RY1 rice is sticky when cooked,the cooking process is more difficult forconsumers, who prefer cooked rice thatis harder, flakier (not sticky), whiter,and denser in appearance and thatremains soft even when stored over-night. This behavior of RY1 rice aftercooking could be attributed to its lowamylose content (15.6%), which hasresulted in poor acceptance of RY1 byfarmers.
Most of the research carried out inour breeding program involved thecreation of medium-height (100-140cm), lodging-tolerant, medium-duration (120-140 d), blast-resistant,high-yielding (more than 3 t ha-1) varie-ties with good grain characteristics(length more than 9 mm; length towidth ratio at least 3; width to thicknessratio about 1; 1,000-grain weight morethan 30 g; endosperm translucency atleast 60%) and good cooking quality(harder and flakier cooked rice grains).
Seven pure breeding lines (Table 1)selected from two crosses—RY1/OS6(PR42) and OS6 × RY7/OS6 (PR55)—were compared with RY1, used as acheck, during two wet crop seasons(1995 and 1996) at the Yangambiresearch station. At Yangambi, the soilis sandy with 20-30% clay, lighter at the
surface than deeper. Annual rainfalland monthly temperature average1,885 mm and 24 °C, respectively. The2-yr experiment was laid out in a ran-domized complete block design withfour replications. Plots were 3 m × 6 m,in which 4-5 grains hill-1 were sown at30 × 20-cm spacing. No fertilization wasused.
On average, life cycle, lodgingtolerance, plant height, reaction tobrown leaf spot, leaf blast, and leafscald were similar for the breeding linesand the check RY2 (Table 2). Grainquality characteristics of the breedinglines, except for endosperm translu-cency, were good and also comparablewith those of RY1. But yield potential
and endosperm translucency variedamong the materials tested. Fourbreeding lines (PR42-44-7-8-1, PR42-44-7-56-2, PR42-44-8-44-1, and PR55-5-2-17-16) had yield potential comparablewith that of RY1 (Table 1) and endo-sperm translucency above or equal tothe minimum required for marketacceptability. These lines were selectedfor testing in different INERA sub-stations located in rice-growing zonesof the country.
The materials selected couldeventually be recommended either forcultivation to replace RY1 or for use inbreeding programs which aim todevelop cultivars with high and stableyield potential and good cookingquality. ■

22 IRRN 1998
Seed technology
A seed production systemfor backstopping a hybridrice breeding program
R. S. Toledo, C. L. Casal, and S. S. Virmani, IRRI
For a hybrid rice breeding program tosucceed, it should have a system formass producing parental lines anddeveloped hybrids. The parental linesshould be multiplied efficiently with-out compromising purity. Sufficienthybrid seeds of acceptable purityshould be made available for thevarious stages of evaluation. At IRRI,we employ a system for mass produ-cing parental lines and hybrids for in-house use and sharing with colla-borators.
The scheme for increasing seed ofpromising parental lines starts withnucleus seed production and is fol-lowed by breeder seed production.These stages of seed multiplication arecarried out under strict isolation condi-tions. Experimental hybrids are pro-duced in quantities depending on theircurrent stage of yield evaluation. Inproducing seeds for trials requiring afew thousand seeds, we use the “isola-tion-free” system. This allows us to pro-duce seeds of several hybrids in smalladjacent plots. To produce a few kilo-grams of hybrids for multilocationaltesting and sharing with collaborators,we use 100-500-m2 partially isolatedplots. Within this seed production sys-tem, we assess the outcrossing poten-tial of the seed parent and determinethe ease of production of particularhybrid combinations. We determinedthe quality of output of the various seedproduction steps through grow-outsconducted after each season. ■
Standardizing hybrid riceseed production practices
S. Singh, B. C. Viraktamath, C. H. M. Vijaya-kumar, M. S. Ramesha, and M. I. Ahmed,Directorate of Rice Research, Rajendranagar,Hyderabad 500030, India
Seed yield in hybrid rice seed produc-tion plots depends, among otherthings, on the area under the femaleparent and the extent of seed set.Factors influencing seed set percentageon the female parent include row ratio,row direction, height of the pollenparent, weather conditions, particu-larly at the time of flowering such aswind velocity, pollen load, nature of thehybrid combination, and the adoptionof practices that promote out-pollination such as proper applicationof GA3 and supplementary pollination.
We conducted an experiment to findout the optimum row ratio of the pollenparent to the seed parent and theappropriate season for maximizingseed yields in male sterile lines andhybrid rice. The material for the studyconsisted of IR58025A, with itsmaintainer line in cytoplasmic malesterile (CMS) line multiplication, andIR58025A, with restorer line IR40750.Only in the 1993 wet season (WS) wasIR46 used as a restorer in hybrid riceseed production. Using these experi-mental materials, a trial involving bothCMS multiplication and hybrid seedproduction was conducted in a fixedrow ratio of 2 male:16 female parentsduring 1993-95 WS and dry season (DS)with six replications. The spacingadopted was 30 cm between two malerows, 20 cm from male to female row,and 15 cm each for female to femalerow and plant to plant. Seeding of thefemale parent was done only once,whereas the male parent was sowntwice at an interval of 3-5 d except in the1995 WS, when only one seeding wasdone. Seedlings of appropriate agewere transplanted at the rate of a singleseedling hill-1 for the male and female
in the 1993 WS and 2-3 seedlings for themale and 2 seedlings for the femaleduring the remaining seasons. Seedproduction practices such as GA3application at 50 g ha-1 and supple-mentary pollination were commonlyadopted. Leaf clipping, however, waspracticed only during the 1993 WS.
Besides days to flowering and extentof synchronization, observations wererecorded on percentage seed set andseed yield of individual female parentrows in each plot and in all replications.By using the yield of individual rows,the seed yield for different row ratiossuch as 2:2, 2:4, ..., and 2:16 were com-puted by adding the yield of the 2nd,3rd, ..., and 8th row from both sides at atime. The results showed a constantdecline in seed set in CMS multiplica-tion and hybrid seed production, with agradual increase in row ratio from 2:2 to2:16.
The optimum row ratio in hybridseed production varied from 2:8 (1993and 1994) to 2:12 (1995) in the WS,whereas in the 1995 DS it was 2:10. Seedyield obtained at optimum row ratioswas the highest in the 1995 WS (1,636kg ha-1) and the lowest in the 1993 WS(720 kg ha-1). The low mean yields in1993 and 1994 in the WS were mainlydue to nonsynchrony in floweringbetween the male and female parents.The mean seed yield was also maxi-mum in the 1995 WS (1,427 kg ha-1),which was attributed to higher pollenload by virtue of perfect synchronyresulting from one-time sowing andplanting of the male parent seedlings.Similarly, in CMS multiplication ofIR58025A, seed yield was the highestwith the ratio of 2:6 (1994 and 1995 WS)and 2:8 (1993 WS and 1995 DS). Themain reason for the very low yield (515kg ha-1) in the 1995 WS was incessantrain during flowering. The mean seedyield obtained in different seasonsindicated DS was more favorable forsuccessful commercial seedproduction. ■

Vol. 23, No. 1 23
Evaluating hybrid seedproduction technologyalong with the newgrowth hormoneMangiferin
R. Singh and S. K. Sahoo, Department of Geneticsand Plant Breeding, Institute of AgriculturalSciences, Banaras Hindu University, Varanasi221005, India
Doubts have been raised about the eco-nomic viability of hybrid seed produc-tion as rice is predominantly a self-pollinated crop. An efficient seed pro-duction package is as important asimproving cytosterile lines and develo-ping and evaluating hybrids. Weattempted to evaluate hybrid seedproduction technology using the newgrowth hormone Mangiferin becausethe cost of gibberellic acid (GA3) washigh. Besides Mangiferin, glycine wasalso used to economize hybrid seedproduction in rice. Using GA3, boricacid, urea, Mangiferin, and glycine indifferent concentrations, we studiedtheir effect on several characters inhybrids, such as duration of opening offlorets, stigma exsertion, angle ofopened florets, plant height before andafter spraying, panicle exsertion at 5%,at 50% flowering, and at maturity, grainyield ha-1, 1000-grain weight, andspikelet length. Analysis of varianceshowed highly significant differencesfor all the characters except panicle ex-sertion at 50% flowering, which wassignificant only at 5%. In general, treat-ment combinations of GA3 andMangiferin produced a positiveresponse for most floral and associatedtraits. GA3 at 40-60 ppm or GA3 +1.5%boric acid influenced more than onecharacter: duration of opening offlorets, stigma exsertion, grain yieldplant-1, plant height, and panicleexsertion. The treatment with GA3+1.5% boric acid was one of the mosteffective combinations as it influencedalmost all the characters studied.Mangiferin in different combinationsand concentrations also responded likeGA3. Mangiferin alone (60 ppm) or incombination with boric acid influenced
many traits: duration of opening offlorets, stigma exsertion, angle ofopened florets, grain yield ha-1, plantheight, and panicle exsertion. All thesetraits recorded higher values than thecontrol. The treatment containing GA3or Mangiferin (60 ppm) along with flagleaf clipping and rope pulling hadmuch higher values for duration ofopening of florets and stigma exsertionthan the control. Another importantfinding was the use of glycine inenhancing outcrossing in rice. GA3 andglycine (40 ppm each) applied togetherresulted in 23% more yield than withthe control. ■
Synchronization studies inhybrid rice seedproduction
S. Singh, B. C. Viraktamath, M. S. Ramesha,C. H. M. Vijayakumar, and M. I. Ahmed,Directorate of Rice Research, Rajendranagar,Hyderabad 500030, India
The two parents of a hybrid combina-tion differ in their growth duration andtherefore flower at different times evenwhen sown on the same date. To pro-duce hybrid seed successfully on acommercial scale, male and femaleparents planted side by side in a fixedrow ratio should flower simultaneous-ly, which is called synchronization.Synchronized flowering between twoparental lines of different growthduration can be achieved by seedingthem on different dates which is termedseeding interval (SI). The method ofarriving at an SI using growth durationis simple to practice. In this method, thedifference between parental lines in thenumber of days to initial heading or50% flowering from seeding is used todetermine SI. The growth duration of avariety, however, is known to be influ-enced by seasonal and weather condi-tions. Therefore, it is essential to deter-mine an SI between the two parents of ahybrid for each season and location.The present study aimed at under-standing the effect of different seedingdates in a season on growth duration
and its implications in hybrid seedproduction. The material consisted of19 restorers belonging to differentmaturity groups—early (6), medium(5), and late (8)—and 3 cytoplasmicmale sterile (CMS) lines with theirmaintainers. All were sown on fivedifferent dates from 11 Jun 1994 at 5-dintervals. The seedlings were uprooted30 d after sowing and planted in arandomized block design with threereplications at a spacing of 20 × 15 cm.Observations on initial heading anddays to 50% flowering were recordedfor each entry and in each replication.The first and last sowings increasedgrowth duration irrespective of thematerial used; the last sowing on 1 Julincreased growth duration by 5-8 d inrestorers, 4 d in CMS lines, and 12 d inmaintainers compared with the firstsowing. Other sowings did notinfluence growth duration markedly.
The results clearly show that there isa definite time period for sowingwithin which growth duration is notaffected significantly. Therefore,growth duration could safely be usedto determine SIs to ensure synchro-nized flowering in parental lines.Studies should be conducted for agiven hybrid combination at each seedproduction location and for eachseason. Seeding interval determined bygrowth duration would then become amuch more reliable parameter for long-term use. ■
Effect of seedling age onflowering time in A, B, andR lines
B. C. Viraktamath, S. Singh, C. H. M. Vijaya-kumar, M. S. Ramesha, and M. I. Ahmed,Directorate of Rice Research, Rajendranagar,Hyderabad 500030, India
Flowering time in parental lines plays avital role in hybrid rice seed produc-tion. Transplanting rice seedlings at theright age assumes special significancein obtaining higher yields in cultivationas well as in hybrid seed production. In

24 IRRN 1998
conditions where timely planting is notpossible because of unavoidablecircumstances, one is bound totransplant aged seedlings. In hybridrice seed production, in spite ofstaggered seeding, simultaneousplanting of male and female parents ispreferred to avoid inconvenience andwasted labor. An experiment wastherefore conducted during the 1994wet season to find out the effect ofseedling age at transplanting onflowering time in A, B, and R lines. Theexperiment was laid out in arandomized block design using fivecytoplasmic male sterile lines alongwith their corresponding maintainers,six restorer lines, and one check varietyin three replications at a spacing of 20 ×15 cm. Sequential transplanting wasdone at an interval of 5 d using 15-50-d-old seedlings. Observations wererecorded on days to 50% flowering.
The results showed that seedling ageexerted a significant and positive effecton flowering; older seedlings at trans-planting delayed flowering in all thelines studied. Instead of using seed-lings of the optimum age (20 d), trans-planting 15-d-old seedlings advancedflowering by 1 d in all the lines. When25-d-old seedlings were used for trans-planting, flowering delay was negligi-ble in the R and B lines, but delay was1 d in the A line. Flowering in the Rlines was delayed markedly (3 d),followed by A and B lines (2 d), when30-d-old seedlings were transplanted.
Restorer lines, which generally havea longer growth duration than theirfemale counterparts, are sown first toproduce hybrid rice seed. Therefore, atthe time of transplanting, R line seed-lings would be older than A line ones.Recent studies have indicated that theoptimum standard of synchronizationis that the female parent should flower1 or 2 d earlier than the male parent.
Our study indicated that floweringwas advanced by 1 d in A lines when15-d-old seedlings were transplanted; itremained the same in B and R lines if20-25-d-old seedlings were trans-planted. In a hybrid combinationinvolving a male parent with 10 d
longer growth duration than the femaleparent, simultaneous transplantingusing 15-d-old A line seedlings and 20-25-d-old seedlings of B and R lines canbe done without affecting the expectedtime of flowering. Transplantingseedlings aged more than 30 d showedpositive and similar effects in delayingflowering in A, B, and R lines. Theresults in our studies indicate thatsimilar information for the parents ofcommercial hybrids must be obtainedin both seasons in the target area priorto large-scale seed production. ■
Storage potential ofparental lines of hybridrice
S. R. Prabagaran, Center for Biotechnology,Pondicherry University, Pondicherry 605104 andA. S. Ponnuswamy, Seed Technology Depart-ment, Tamil Nadu Agricultural University,Coimbatore, India
To elucidate the storability of parentallines of hybrid rice, we studied seedquality parameters (such as germi-nability and vigor index) and somebiochemical traits. Seeds of differentparental lines of hybrid rice—A lines(IR62829A and IR58025A), B lines(IR62829B and IR58025B), and R lines(IR10198-66-2R, AS89044, Pusa 150R,C20, and C22) having superiorgerminability—were stored in clothand 400-gauge polyethylene bags. Theseeds were evaluated for seed qualityparameters at bimonthly intervals for12 mo. Finally, they were subjected toaccelerated aging for 9 d in an agingchamber (40 °C and 100% relativehumidity).
IR10198-66-2R and AS89044 (R lines)maintained prolonged viability,whereas deterioration was faster inC20. A lines usually recorded highgerminability initially but deterioratedfaster than their respective maintainersduring storage. A similar trend wasnoticed in dry matter production andvigor index. Biochemical analysisshowed a steady decrease in dehydro-genase activity and an increase in both
free sugars and electrical conductivityof seed leachates as the storage periodincreased. Among the containerstested, the polyethylene bag proved itssuperiority over the cloth bag in main-taining vigor and viability, probablybecause of less influence from externalfactors such as relative humidity ortemperature. It is obvious from thestudy that, besides container andgenotype influence, the male sterilitysystem also has a key role in seedstorability. ■
Dormancy of seeds ofsome rice hybrids andtheir parents
V. Bhaskar, A. H. Krishnamurthy, R. M. Radha-krishna, and B. V. Chandra, Regional ResearchStation, VC Farm, Mandya 571405, Karnataka,India
The extent of seed dormancy in hybridsand their parents was investigatedduring the 1994 wet season. Harvestedseeds from parents, hybrids, and checkvarieties were stored in paper bags andtested for germination at 5-d intervals.The study included the hybridsIR58025A/IR9761-19-R and IR58025A/IR29723-143-3-2-1R, male parentIR62829A, and check varieties MandyaVijaya and IR20.
The results indicated that bothcytoplasmic male sterile (CMS) lines,IR58025A and IR62829A, breakdormancy 25 d after maturity andrecord more than 90% germination. Thehybrid IR58025A/IR9761-19-1Rrecorded 72% germination after 25 dand 88% after 30 d. Its restorer took 30 dto break dormancy, with 84%germination. The other hybrid,IR58025A/IR29723-143-3-2-1R,recorded 92% germination after 30 d,whereas IR29723-143-3-2-1R did notgerminate at all even after 30 d. It took45 d after harvest to record more than85% germination. Check varietiesMandya Vijaya and IR20 brokedormancy after 20 d. The resultsdemonstrated that CMS lines IR58025Aand IR62829A can be used for sowing

Vol. 23, No. 1 25
Significance of stigmaexsertion in enhancingoutcrossing in male sterilerice lines
C. R. Elsy, M. Rangaswamy, and K. N. Ganesan,School of Genetics, Tamil Nadu AgriculturalUniversity, Coimbatore 641003, India
An investigation undertaken to studythe floral characteristics of male sterilelines will lead to better outcrossing andenhanced seed set. Eight cytoplasmicmale sterile (CMS) lines were used forthe study. Floral characters such as rateof stigma exsertion, stigma length and
25 d after maturity. Restorer linesIR9761-19-1R and IR29723-143-3-2-1Rcan be sown only 30 and 45 d afterharvest, respectively. It is safe to marketthe hybrid seeds 30 d after harvest. ■
width, stigmatic area, style length, andnatural seed set of these lines wereobserved during the 1995 wet season.
The natural seed set in these CMSlines ranged between 1.86% inIR68275A and 10.6% in IR58025A, witha mean value of 5.59%. Among thefloret characteristics studied, stigmaexsertion showed a predominantinfluence on seed set. The mean stigmaexsertion rate ranged between 6% inIR68275A and 48% in IR58025A.IR58025A, IR68281A, and IR67684Ashowed a significantly higher percent-age of stigma exsertion. The significant-ly higher seed set (10.6%) in IR58025Awas attributed to its higher stigmaexsertion rather than to any other floralcharacteristics. The genotype with theleast stigma exsertion (IR68275A)recorded the lowest natural seed set(1.86%). Genotypes with a higher rate
of stigma exsertion recorded a higherstyle length, as observed in IR58025A,IR68281A, and IR67684A. Even thoughstigma exsertion and style length inIR67684A were higher than in othergenotypes, seed set was low (4.1%),indicating the linkage factor involvedwith cytoplasm for male sterility. InIR64707A, multiple stigmas wererecorded in 3% of the florets.
Our results indicated that, amongfloral traits, better stigma exsertionshould be given maximum importancewhen developing desirable CMS lines.IR58025A showed maximum seed set(47.68%) and maximum style length(1.22 mm). Although genotypeIR67684A showed many favorablefloral characteristics for high seed set, itis undesirable because of the inter-action of genome and cytoplasm onseed set. ■
Crop and resource managementFertilizer management — inorganic sourcesFertilizer management
Effect of N levels and timeof application on grainyield of hybrid rice
S. P. Singh, B. S. Devi, and S. V. Subbiah,Directorate of Rice Research, Rajendranagar,Hyderabad 500030, India
During the 1995-96 wet season and dryseason, we studied the performance ofthree hybrids—KHR1, ProAgro 103,and MGR1—using Jaya and Rasi asstandard checks. This study included
growth and grain yield performance atfour levels of N (0, 60, 120, and 180 kgha-1). Three schedules of N applicationwere followed: three splits (50:25:25%at basal, tillering and panicle initiation),four splits (25% N at basal, tillering,panicle initiation, and 50% flowering),and five splits (25% N at basal, tillering,and panicle initiation and the remain-der applied in two equal splits at 50%flowering and 10 d later). Grain yielddifferences among the test hybrids andvarieties were significant during both
seasons. The maximum grain yieldswere recorded for ProAgro 103, Vikas,KHR1, and MGR1. Rasi produced 5-14% less grain yield than ProAgro 103.Varieties responded linearly to appliedN levels up to 120 kg ha-1. Grain yielddifferences were not significant amongdifferent schedules of N application,indicating that more than three splits ofN application are not required forhigher grain production by hybrid ricein Vertisol soils. ■
Crop management
Three-drying cultivation of rice: anew technology for rice produc-tion in drought-prone areas
Zhou Jie, Zhou Long, Rice Research Institute,Agriculture Bureau, Linyi, Shandong, China
Shandong, China, has a dry spring andrainy summer. In Linyi district, for
example, mean annual rainfall is 872-949 mm, of which 125-150 mm (15%)occur in spring and 560-600 mm (65%)in summer. Because of waterlogging inthe wet summer period, grain yields ofrainfed crops such as maize, soybean,or sweet potato were often poor in low-lying farming regions. When rice wasdry-seeded late in spring after fall
wheat, the medium-duration rice varie-ties could not ripen because of theshortened growing period. It was diffi-cult to grow rice seedlings because ofthe lack of water in spring. Therefore,only a fall wheat crop could be grownin the region.
This paper reports research exami-ning the ability to raise rice seedlings,
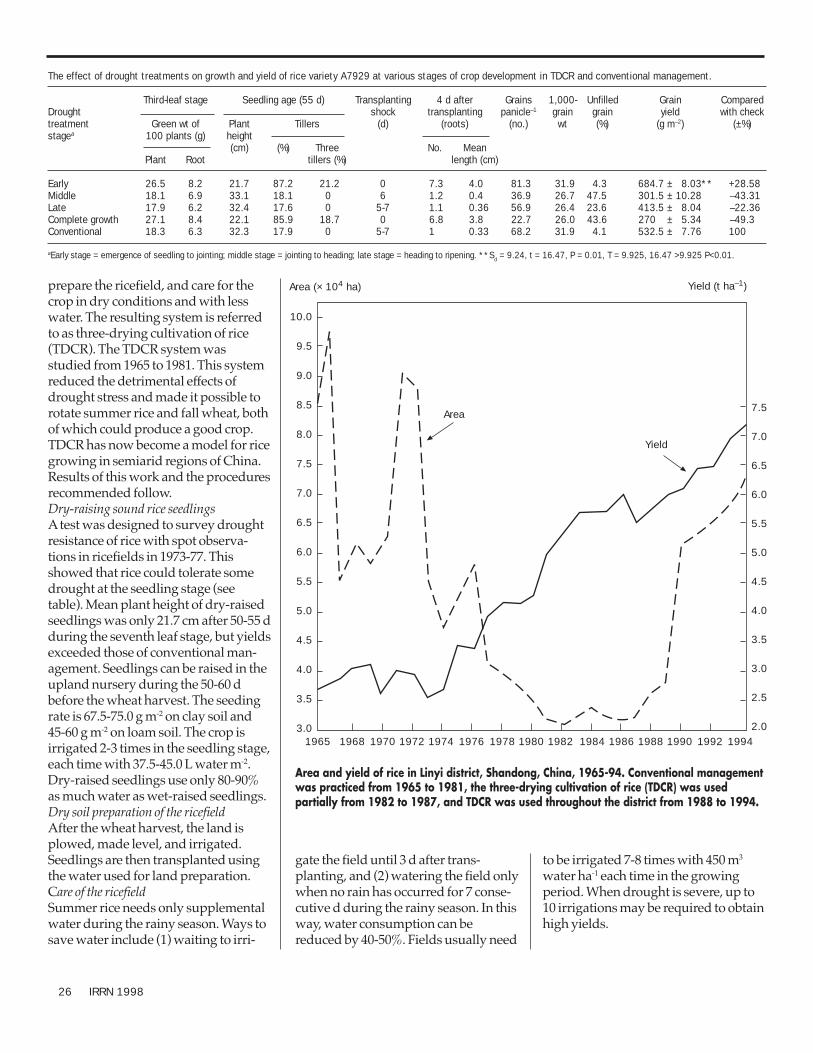
26 IRRN 1998
prepare the ricefield, and care for thecrop in dry conditions and with lesswater. The resulting system is referredto as three-drying cultivation of rice(TDCR). The TDCR system wasstudied from 1965 to 1981. This systemreduced the detrimental effects ofdrought stress and made it possible torotate summer rice and fall wheat, bothof which could produce a good crop.TDCR has now become a model for ricegrowing in semiarid regions of China.Results of this work and the proceduresrecommended follow.Dry-raising sound rice seedlingsA test was designed to survey droughtresistance of rice with spot observa-tions in ricefields in 1973-77. Thisshowed that rice could tolerate somedrought at the seedling stage (seetable). Mean plant height of dry-raisedseedlings was only 21.7 cm after 50-55 dduring the seventh leaf stage, but yieldsexceeded those of conventional man-agement. Seedlings can be raised in theupland nursery during the 50-60 dbefore the wheat harvest. The seedingrate is 67.5-75.0 g m-2 on clay soil and45-60 g m-2 on loam soil. The crop isirrigated 2-3 times in the seedling stage,each time with 37.5-45.0 L water m-2.Dry-raised seedlings use only 80-90%as much water as wet-raised seedlings.Dry soil preparation of the ricefieldAfter the wheat harvest, the land isplowed, made level, and irrigated.Seedlings are then transplanted usingthe water used for land preparation.Care of the ricefieldSummer rice needs only supplementalwater during the rainy season. Ways tosave water include (1) waiting to irri-
The effect of drought treatments on growth and yield of rice variety A7929 at various stages of crop development in TDCR and conventional management.
Third-leaf stage Seedling age (55 d) Transplanting 4 d after Grains 1,000- Unfilled Grain ComparedDrought shock transplanting panicle–1 grain grain yield with checktreatment Green wt of Plant Tillers (d) (roots) (no.) wt (%) (g m–2) (±%)stagea 100 plants (g) height
(cm) (%) Three No. MeanPlant Root tillers (%) length (cm)
Early 26.5 8.2 21.7 87.2 21.2 0 7.3 4.0 81.3 31.9 4.3 684.7 ± 8.03** +28.58Middle 18.1 6.9 33.1 18.1 0 6 1.2 0.4 36.9 26.7 47.5 301.5 ± 10.28 –43.31Late 17.9 6.2 32.4 17.6 0 5-7 1.1 0.36 56.9 26.4 23.6 413.5 ± 8.04 –22.36Complete growth 27.1 8.4 22.1 85.9 18.7 0 6.8 3.8 22.7 26.0 43.6 270 ± 5.34 –49.3Conventional 18.3 6.3 32.3 17.9 0 5-7 1 0.33 68.2 31.9 4.1 532.5 ± 7.76 100
aEarly stage = emergence of seedling to jointing; middle stage = jointing to heading; late stage = heading to ripening. **Sd = 9.24, t = 16.47, P = 0.01, T = 9.925, 16.47 >9.925 P<0.01.
gate the field until 3 d after trans-planting, and (2) watering the field onlywhen no rain has occurred for 7 conse-cutive d during the rainy season. In thisway, water consumption can bereduced by 40-50%. Fields usually need
10.0
9.5
9.0
8.5
8.0
7.5
7.0
6.5
6.0
5.5
5.0
4.5
4.0
3.5
3.0
7.5
7.0
6.5
6.0
5.5
5.0
4.5
4.0
3.5
3.0
2.5
2.01965 1968 1970 1972 1974 1976 1978 1980 1982 1984 1986 1988 1990 1992 1994
Area (× 104 ha)
Area
Yield
Yield (t ha–1)
Area and yield of rice in Linyi district, Shandong, China, 1965-94. Conventional managementwas practiced from 1965 to 1981, the three-drying cultivation of rice (TDCR) was usedpartially from 1982 to 1987, and TDCR was used throughout the district from 1988 to 1994.
to be irrigated 7-8 times with 450 m3
water ha-1 each time in the growingperiod. When drought is severe, up to10 irrigations may be required to obtainhigh yields.
Vol. 23, No. 1 27
Impact on rice productionSince the late 1970s, drought has causeddrastic cuts in rice area in Linyi (seefigure). After 1982, TDCR became morepopular and by 1988 was used by allfarmers. Ricefields multiplied andrough rice yield increased year afteryear. By 1994 there was a 33% and 26%increase in per unit area yield com-pared with before 1982 and up to 1988,respectively. Yields of wheat followingrice also increased from 1.5 to 5.4 t ha-1.Therefore, profit increased by 22%.Yield increased because the dry-raisedseedling itself was healthy and mid-laterice cultivars would be planted in timeto use their 20-d-longer growth period.Pests and diseases are insignificant androtation cropping is improved.
TDCR has several advantages: (1) itis economical in water, labor, seeds, andenergy; (2) it reduces costs and labor;and 3) it ensures a good harvest despitedrought or waterlogging. ■
Agronomic package forhybrid rice cultivation
S. V. Subbaiah, R. M. Kumar, and S. P. Singh,Directorate of Rice Research (DRR),Rajendranagar, Hyderabad 500030, India
Yield stagnation or even a decline hasbeen observed in many rice-growingareas, particularly in potentially highlyproductive areas. To increase rice pro-duction, a few rice hybrids have beendeveloped and released in recent yearsin India. To exploit the crop’s fullheterotic potential, it is essential todevelop a package of optimum produc-tion practices for hybrid rice cultiva-tion. We therefore attempted to studynursery management, seeding densi-ties, seedling ratios, transplantingdates, N management, and hybridresponse at several locations under thecoordinated rice improve-mentprogram.
The studies revealed that the opti-mum seed density of 15-20 kg in a1,000-m2 nursery and planting in thesecond fortnight of July (1 seedling
hill-1) are ideal practices for obtainingsuperior grain yield in hybrids. Theresults also indicated that different ricehybrids performed well at differentlocations: Hybrid ProAgro at DRR andFaizabad; ProAgro and CRH1 at theCentral Rice Research Institute andChinsurah. The local standard HKR46performed well at Kaul. These hybridsrecorded superior grain yields andshowed higher N use efficiency at theselocations. Grain yield differences in thetested hybrids were not significant atAduthurai, Mandya, and Pantnagar. As
the tests were made in the wet season,the hybrids may not have exhibitedtheir full yield potential at Cuttack andMaruteru because of rain. The responseto N was limited up to 100 kg N ha-1 atCuttack, Chinsurah, and Pantnagar,and up to 150 kg N ha-1 at Aduthurai,Hyderabad, Mandya, and Faizabad.Hybrids failed to respond beyond 150kg N ha-1 at most test locations. Apply-ing the recommended N in three splits(50% basal + 25% tillering + 25% boot-ing) was the best management practicefor hybrids in Vertisols at DRR. ■
Integrated germplasm improvement — weeds
Chemical weed control in ricenurseries of the hill zone ofKarnataka
M. Syed Anwarulla, NARP, Regional ResearchStation, Mudigere 577132, Karnataka, India
Weeds pose a threat to rice cultivationfrom the nursery stage in the hill zoneof Karnataka (south India), which has182,000 ha under rainfed rice (the solefood and field crop of the zone). Thisarea is located between 12-14° N and
74-76° E. Average annual rainfall is2,460 mm, of which 83% occurs be-tween June and September (with theremaining 17% from October to De-cember). Rice nurseries are sown fromthe first week of June to the secondweek of July depending upon thetoposequence and rainfall pattern forthe wet season and during the secondweek of December to the second weekof January for the summer (dry) season.
Rice nurseries are infested by grasssedges and broadleaf weeds that
Table 1. Effect of herbicides on germination of rice and dry weight of rice seedlings in the hill zone of Karnataka,India.
Germination (%) recorded Mean Dry weight (av) of 10 MeanTreatment at 10 DASa of seedlings at 25 DAS (g) of
2 yr 2 yrKharif Summer Kharif Summer Kharif Summer Kharif Summer
(1993-94) (1994-95) (1993-94) (1994-95)
1. Control, unweeded check 92 94 94 95 94 1.06 1.13 1.12 1.27 1.192. Butachlor preemergence 92 94 88 92 91 2.39 2.43 2.43 2.67 2.55 50 EC (1.25 L ha–1)3. Benthiocarb-30 EC 74 76 68 72 72 1.84 1.94 1.96 2.01 1.98 (1.25 kg ai ha–1) preemergence4. Pendimethaline 30 EC 72 73 68 64 69 1.92 1.82 2.01 1.98 1.99 (preemergence) (1.0 kg ai ha–1)5. Butachlor (5%) granules 73 74 77 79 75 2.12 1.93 2.20 2.08 2.14 (25 kg ha–1)6. Anilophos 30 EC 89 93 90 88 90 2.26 2.33 2.32 2.46 2.39 @300 mL ha–1) (3 DAS) Postemergence7. Hand weeding at 92 93 93 94 93 1.94 2.03 2.10 2.19 2.14 10 and 20 DAS LSD (P = 0.05) 0.03 0.08 0.12 0.27 0.16
aDAS = days after sowing.
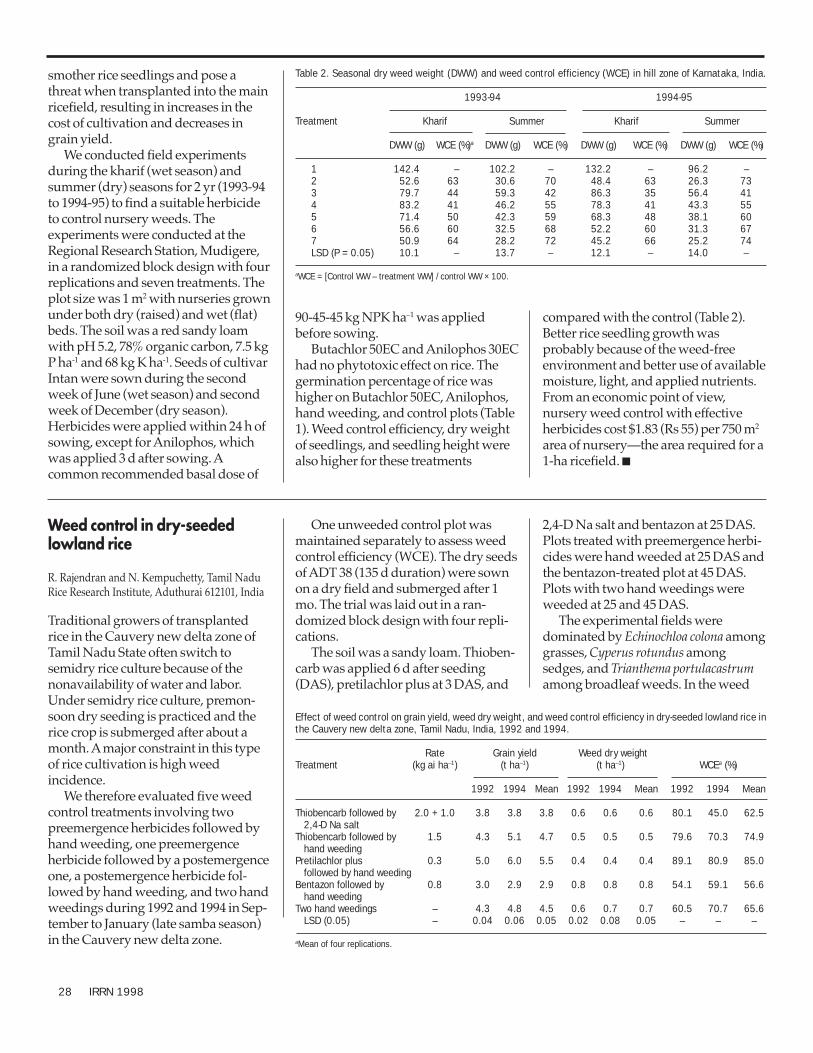
28 IRRN 1998
Weed control in dry-seededlowland rice
R. Rajendran and N. Kempuchetty, Tamil NaduRice Research Institute, Aduthurai 612101, India
Traditional growers of transplantedrice in the Cauvery new delta zone ofTamil Nadu State often switch tosemidry rice culture because of thenonavailability of water and labor.Under semidry rice culture, premon-soon dry seeding is practiced and therice crop is submerged after about amonth. A major constraint in this typeof rice cultivation is high weedincidence.
We therefore evaluated five weedcontrol treatments involving twopreemergence herbicides followed byhand weeding, one preemergenceherbicide followed by a postemergenceone, a postemergence herbicide fol-lowed by hand weeding, and two handweedings during 1992 and 1994 in Sep-tember to January (late samba season)in the Cauvery new delta zone.
smother rice seedlings and pose athreat when transplanted into the mainricefield, resulting in increases in thecost of cultivation and decreases ingrain yield.
We conducted field experimentsduring the kharif (wet season) andsummer (dry) seasons for 2 yr (1993-94to 1994-95) to find a suitable herbicideto control nursery weeds. Theexperiments were conducted at theRegional Research Station, Mudigere,in a randomized block design with fourreplications and seven treatments. Theplot size was 1 m2 with nurseries grownunder both dry (raised) and wet (flat)beds. The soil was a red sandy loamwith pH 5.2, 78% organic carbon, 7.5 kgP ha-1 and 68 kg K ha-1. Seeds of cultivarIntan were sown during the secondweek of June (wet season) and secondweek of December (dry season).Herbicides were applied within 24 h ofsowing, except for Anilophos, whichwas applied 3 d after sowing. Acommon recommended basal dose of
Table 2. Seasonal dry weed weight (DWW) and weed control efficiency (WCE) in hill zone of Karnataka, India.
1993-94 1994-95
Treatment Kharif Summer Kharif Summer
DWW (g) WCE (%)a DWW (g) WCE (%) DWW (g) WCE (%) DWW (g) WCE (%)
1 142.4 – 102.2 – 132.2 – 96.2 –2 52.6 63 30.6 70 48.4 63 26.3 733 79.7 44 59.3 42 86.3 35 56.4 414 83.2 41 46.2 55 78.3 41 43.3 555 71.4 50 42.3 59 68.3 48 38.1 606 56.6 60 32.5 68 52.2 60 31.3 677 50.9 64 28.2 72 45.2 66 25.2 74LSD (P = 0.05) 10.1 – 13.7 – 12.1 – 14.0 –
aWCE = [Control WW – treatment WW] /control WW × 100.
One unweeded control plot wasmaintained separately to assess weedcontrol efficiency (WCE). The dry seedsof ADT 38 (135 d duration) were sownon a dry field and submerged after 1mo. The trial was laid out in a ran-domized block design with four repli-cations.
The soil was a sandy loam. Thioben-carb was applied 6 d after seeding(DAS), pretilachlor plus at 3 DAS, and
2,4-D Na salt and bentazon at 25 DAS.Plots treated with preemergence herbi-cides were hand weeded at 25 DAS andthe bentazon-treated plot at 45 DAS.Plots with two hand weedings wereweeded at 25 and 45 DAS.
The experimental fields weredominated by Echinochloa colona amonggrasses, Cyperus rotundus amongsedges, and Trianthema portulacastrumamong broadleaf weeds. In the weed
Effect of weed control on grain yield, weed dry weight, and weed control efficiency in dry-seeded lowland rice inthe Cauvery new delta zone, Tamil Nadu, India, 1992 and 1994.
Rate Grain yield Weed dry weightTreatment (kg ai ha–1) (t ha–1) (t ha–1) WCEa (%)
1992 1994 Mean 1992 1994 Mean 1992 1994 Mean
Thiobencarb followed by 2.0 + 1.0 3.8 3.8 3.8 0.6 0.6 0.6 80.1 45.0 62.5 2,4-D Na saltThiobencarb followed by 1.5 4.3 5.1 4.7 0.5 0.5 0.5 79.6 70.3 74.9 hand weedingPretilachlor plus 0.3 5.0 6.0 5.5 0.4 0.4 0.4 89.1 80.9 85.0 followed by hand weedingBentazon followed by 0.8 3.0 2.9 2.9 0.8 0.8 0.8 54.1 59.1 56.6 hand weedingTwo hand weedings – 4.3 4.8 4.5 0.6 0.7 0.7 60.5 70.7 65.6 LSD (0.05) – 0.04 0.06 0.05 0.02 0.08 0.05 – – –
aMean of four replications.
90-45-45 kg NPK ha–1 was appliedbefore sowing.
Butachlor 50EC and Anilophos 30EChad no phytotoxic effect on rice. Thegermination percentage of rice washigher on Butachlor 50EC, Anilophos,hand weeding, and control plots (Table1). Weed control efficiency, dry weightof seedlings, and seedling height werealso higher for these treatments
compared with the control (Table 2).Better rice seedling growth wasprobably because of the weed-freeenvironment and better use of availablemoisture, light, and applied nutrients.From an economic point of view,nursery weed control with effectiveherbicides cost $1.83 (Rs 55) per 750 m2
area of nursery—the area required for a1-ha ricefield. ■

Vol. 23, No. 1 29
control treatments, pretilachlor plus(0.3 kg ai ha-1) with one hand weedingsignificantly reduced total weed dryweight at 60 DAS, with a maximum
WCE of 85%. This treatment alsoproduced the best yield—5.49 t ha-1 (seetable). Thiobencarb (1.5 kg ai ha-1) withone hand weeding was as effective as
Research methodologySummary statistics for the ratoon crop models, 1987-89 Texas ratoon crop survey.
Ratoon crop modelsa
ItemRYb RYMc RHYd RGe
F value 9.8350 2.6570 2.5010 1.6730Prob > F 0.0001 0.0001 0.0001 0.0047R square 0.6397 0.3413 0.4057 0.0203Adj R square 0.5746 0.2129 0.2898 0.0816CV 1.6388 2.6347 5.2959 71.1072Observations 413 381 381 380 (no.)
aEach model has approximately 40-50 variables that werehypothesized to affect yield and quality according to riceresearch and extension scientists. The variables include thosethat are (a) known at the beginning of the production period(predetermined variables such as soil type), (b) controlled bythe decision maker within the production period (decisionvariables (such as fertilizer management), and (c) beyond thedecision maker's control (uncertain variables, such as climate).bRY = ratoon crop yield. cRMY = ratoon crop milling yield. dRHY= ratoon crop head yield. eRG = ratoon crop grade.
The relationship betweenexperimental plot research andTexas rice producers’ ratoonfield results
T. N. Thompson, M. E. Rister, W. R. Grant,Department of Agricultural Economics, TexasA&M University, College Station, Texas 77843,USA; A. D. Klosterboer, G. N. McCauley,J. W. Stansel, Agricultural Research Center,Beaumont, Texas 77713, USA; J P. Krausz,Department of Plant Pathology and Micro-biology, Texas A&M University; B. M. Drees,Department of Entomology; F. T. Turner,C. N. Bollich, M. A. Marchetti, M. O. Way,B. D. Webb, N. G. Whitney, S. R. M. Pinson,Agricultural Research Center, Beaumont, Texas;and C. R. Schubert, Department of AgriculturalEconomics, Texas A&M University
At several recent Texas (USA) riceindustry meetings, rice consultants andproducers mentioned observing inputresponses different from those reportedby extension and research specialists.This study documents the correlationbetween scientists’ recommendationsbased on existing theory drawn fromrice plot research and actual resultsfrom commercial fields obtained fromproducers’ surveys during 1987-89.Scientists’ expert ex ante opinions onthe effects of several predetermined,decision, and uncertain factors onratoon crop rice (Oryza sativa L.) fieldyield and quality (milling yield, headyield, and grade) were documented. Amodified Delphi procedure, wherebyscientists come to a consensus agree-ment, was used to determine an exten-sive set of independent variableshypothesized to either positively ornegatively affect ratoon crop yield andquality. The scientists’ identifiedvariables provided the basis for a mailsurvey of Texas rice producers’ culturalpractices and yield-quality responses
during 1987-89. Following collection ofthe survey data, multiple regressionprocedures and associated statisticaltests were used to identify statisticallysignificant ratoon yield and quality-influencing variables.
Table presents summary regressionresults. The predetermined variablesincluded management (3 variables),soils (2), water management (2), landpreparation (4), and rotation (7). Thedecision variables were varieties (2),fertilization (10), emergence/seeding(3), water management (8), diseases (3),pests (4), and harvesting (10). Theuncertainty variables consisted ofweather (6) and diseases (2).
Low coefficients of determinationindicate that the model formulationsand the hypotheses identified for theratoon crop yield and quality modelsdo not fully account for the variabilityin actual ratoon crop field yields andquality. In many cases, scientists’ hypo-theses matched producers’ ratoon yieldand quality responses; however, some-times the apparent direction of effect in
the data set was in an opposite direc-tion than hypothesized. For example,1988 was hypothesized to have been amore favorable year for ratoon fieldyields and quality. Statistical resultsindicate that the scientists’ hypothesisof higher field yields was supported,but their hypothesis regarding higherquality (milling yield) was rejected.
Comparison of the number ofstatistically significant variablesbetween the yield and quality modelsindicates that 42% of the scientists’hypothesized yield variables weresignificant, whereas 45% of thescientists’ hypothesized qualityvariables were significant. Hypothe-sized signs for the statistically signifi-cant variables were then comparedwith calculated signs for alternativefarm scenarios based on the regressionresults. Agreement with scientists’hypotheses occurs if the calculated signis the same as the hypothesized sign.Such “in agreement” results suggestthat scientists well recognize andunderstand the effect of these factors onTexas rice farmers’ ratoon crop yieldsand quality. Comparison between theyield and quality models indicates that,for the yield model, 36% of the statisti-cally significant hypotheses were inagreement, whereas 24% were in agree-ment for the quality models. In sum-mary, scientists were less likely todetermine the input impacts on qualityrather than yield. These results indicatethe importance of determining directand interactive effects of productionvariables on rice quality.
The use of producer surveys pro-vides opportunities for researchers toextensively evaluate numerous pro-ducer circumstances beyond the capa-bility of experimental plot research andverification trials. This type of evalua-
two hand weedings. No post-emergence herbicide was as effectivefor dry-seeded lowland rice. ■

30 IRRN 1998
tion can lead to the identification ofinteraction variables and constraints toproduction. In instances where newproduction practices evolve (such asthe introduction of high-yieldingsemidwarf rice varieties), producersurveys may point to the need for arefinement of recommendations.
Comparing methods ofstability analysis in ricehybrids
C. H. M. Vijayakumar, M. I. Ahmed,B. C. Viraktamath, S. Singh, and M. S. Ramesha,Directorate of Rice Research, Rajendranagar,Hyderabad 500030, India
Yield heterosis in rice is reported tointeract with environment. Identifyingrice hybrids with stable performance isa prerequisite to successfully exploitingthis technology over a large area. Thepopular parameter for selection ofstable entries used by many breeders isthe varietal mean averaged over loca-tions. Another method is to rank entriesfor each location and then use meanperformance of the rank to assess theirpotential. It is well known that geno-type × environment (G × E) interactions
Producers can use the results tocompare their farm responses withthose of others. Identification of theprincipal sources of variation will allowproducers to focus on information thatcan provide a basis for refining theirproduction practices. Producers’knowledge of the relationship between
production inputs and rice quality canbe improved if research and extensionspecialists report yield and qualityresults from research and experimentaltrials and if more research is devoted toproducer-controlled impacts such aspostharvest handling. ■
in multilocational yield trials and thecomparison of cultivar means are notvery useful. In addition, the mean andits ranks do not indicate the location-wise magnitude of differences betweenthe test entry and the standard checkagainst which selection is to be made.In recent years, many new methodshave been proposed for stability analy-sis. We compared the following threemethods with selection based on theexperimental mean: 1) a superioritymeasure of a cultivar, 2) nonparametricmeasures of phenotypic stability, and 3)relative yield as a measure.
We used the grain yield data of ahybrid rice trial conducted over 12 loca-tions in the 1994 wet season for thestudy, and calculated the rank correla-tion (rs) between methods. The resultsshowed that selection based on means
of entries was closer to method 3, rs =0.99, followed by the superioritymeasure (rs = 0.95). The ranking basedon nonparametric measure was notrelated to the mean. Of the threemethods used, the superiority measurewas found to be useful for identifyingstable hybrids. We analyzed the yielddata from six trials conducted during1994-95 to ascertain the usefulness ofthe superiority measure for identifyingstable hybrids. The best-yieldinghybrid was found to be the most stable.The superiority measure—as amethod—is simple, convenient, andquite useful, because it uses only oneparameter, which makes comparisonamong test genotypes easier. Thismethod also indicates the highestyielding geno-type, one that could berecommended for the whole region. ■
Role of voluntary agenciesin popularizing hybrid riceproduction in India
G. G. Reddy and G. Satyanarayana, KrishiVigyan Kendra, Sri Aurobindo Institute of RuralDevelopment, Gaddipalli, Nalgonda 508201; andM. I. Ahmed and K. Krishnaiah, Directorate ofRice Research, Rajendranagar, Hyderabad500030, India
The Sri Aurobindo Institute of RuralDevelopment, Gaddipalli, a nonprofit-oriented, nonsectarian, voluntary, andautonomous organization wasestablished in 1973. A Krishi VigyanKendra (KVK) was started in 1985 with
funds from the Indian Council ofAgricultural Research (ICAR). ThisKVK was judged as the best among 261KVKs in the country during 1994-95.Nalgonda District is in the southernTelangana zone of Andhra Pradesh.This is a rural district and 88% of thepopulation depends on agriculture.Hybrid rice in China on a large scaleproved to have a more than 25% yieldadvantage over conventional purelinevarieties. The wide adaptability of ricehybrids has produced increased riceproduction and productivity in Chinasince the mid-1980s. This successprompted ICAR to accelerate researchon hybrid rice, through an ICAR-
UNDP-funded project in India. Thisresearch network project involved 12research centers (two lead, seven asso-ciate, and three strategic centers) andstate seed production agencies locatedin the target environment. Through thisnetwork, several rice hybrids weredeveloped and released for commercialcultivation. To popularize hybrid riceproduction on a large scale, the SriAurobindo Institute developed anintegrated project that involved adap-tive research at the institute farm; on-farm testing in farmers’ fields; layoutdemonstrations on hybrid rice seedproduction; production and distri-bution of good quality hybrid rice
Education and communication

Vol. 23, No. 1 31
seeds; training farmers, farm women,rural youth, and science graduates;organizing demonstrations in F1commercial seed production; populari-zing hybrid rice technology throughextension services; and producingmultimedia training materials onhybrid rice technology. The importantlesson we learned was that for eachlocation and season, we needed todetermine the seeding interval for theparental lines. During the past threeseasons (1995-96), hybrid rice seed
production was carried out on 164 ha infarmers’ fields. A total yield of 195 t ofhybrid seeds was produced and sup-plied to various companies and indivi-dual farmers. This seed was enough forthe commercial production of hybridrice on 9,750 ha. During the 1996 wetseason, hybrid rice seed productionwas carried out on 90 ha.
Training progressive farmers, youth,and farm women in hybrid rice seedproduction and F1 commercial cultiva-tion promotes hybrid rice cultivation
and increases rice yields. Training wasorganized in hybrid rice seed produc-tion for various categories of clients—farmers, farm women, farm youth,extension officers, and representativesof seed companies. More than 33,400man-days of training were impartedduring 1995-96. With the assistance ofthe United Nations DevelopmentProgramme, this KVK organized alarge-scale training program for farmwomen. ■
Impact of hybrid rice onIndia’s food economy:implications and policyissues
A. Janaiah, M. I. Ahmed, and K. Krishnaiah,Directorate of Rice Research, Rajendranagar,Hyderabad 500030, India
Basic data from the literature on thenature and impact of the adoption ofhybrid rice in China and experiences inother research centers were gathered.Additional supporting data werecollected from farmers’ fields in themajor rice-growing states in India
purchase costly hybrid rice seed everycrop season. A long-term use of thistechnology, however, would benefit theentire rice-farming community andurban population as well. Hybrid ricetechnology would generate a moremarketable surplus. As a consequence,higher domestic prices of rice would beprevented in the consumer market,which would benefit urban consumers,especially low and middle class groups.Hybrid rice would also add to centralbuffer stocks and increase the supply ofrice through the public distributionsystem at cheaper prices. Becausehybrid rice technology is labor-
Economics of hybrid riceseed production in India
A. Janaiah and M. I. Ahmed, Directorate of RiceResearch, Rajendranagar, Hyderabad 500030,India
Input-output data on hybrid seed pro-duction were obtained from seedgrowers’ farms under private compa-nies and a nongovernment organiza-tion (KVK, Gaddipally, AndhraPradesh) from 1992-93 to 1994-95.Using these data, we studied the cost-return profile of F1 seed production.The F1 seed yield was only 560 kg ha-1
when it was produced for the first timein 1992-93, but it increased to 1,385 kgha-1 in 1994-95. The average total cost of
F1 seed production was Rs 23,840 ha-1.Costs involved 45% for labor, 24% formanure and fertilizers, and 8% forgibberellic acid. The cost of 1 kg of seedproduced dropped from Rs 33(US$0.92) in 1992-93 to Rs 17 (US$0.48)during 1994-95, primarily because ofincreased yield. Because hybrid riceseed production is labor-intensive, 280-300 additional man-days of employ-ment could be generated in the ruralarea, especially for women. In addition,gross and net returns to F1 seed produc-tion are Rs 42,710 (US$1,186) andRs 18,870 (US$524) ha-1, respectively, atRs 25 (US$0.69) as the procurementprice kg-1. At the rate of the procure-ment price paid to seed growers by theseed industry, however, the computed
eco-nomically viable threshold seedyield was 1,900 kg ha-1. If F1 seed pro-duction is to be made economicallyviable to seed growers at 1,385 kg ha-1 ofseed yield, the procurement priceshould be Rs 34 (US$0.95) kg-1. The ratioof seed (F1) area to commercial cultiva-tion area of hybrid rice is estimated at1:37.6. Thus, close to 53,000 ha areneeded for an F1 seed production pro-gram to meet the seed requirement of 2million ha under hybrid rice by the year2000. Our results stress the need tofurther improve F1 seed yields. We alsoneed to formulate an incentive-oriented and forward-contract baseprice policy for F1 seed to induce seedgrowers to produce the required hybridrice seed. ■
where rice hybrids were releasedrecently. All these data formed the basisfor this study. Hybrid rice outyieldsexisting conventional high-yieldingvarieties by 1-2 t ha-1 in irrigatedenvironments. The adoption of hybridrice would shift the productionfunction farther upward and theincreased rice output in irrigated areaswould result in a higher domesticsupply. In the short run, this could leadto a further widening of the incomeinequalities between irrigated andunirrigated regions. In addition, hybridrice would benefit large farmers, whoare early adopters with an ability to
Socioeconomic impact

32 IRRN 1998
intensive, particularly seed production,it generates additional employmentopportunities, especially for women(nearly 280-300 man-days ha-1) in ruralIndia. Thus, the purchasing power oflandless labor would increase throughan increase in earnings from labor.
Evidence from China, a centrallyplanned economy, shows that directgovernment intervention throughincentive policies, apart from yieldadvantage, played a significant role inpopularizing hybrid rice on a large
scale. These policies involved anincentive output price for hybrid ricegrains despite poor grain quality, on apar with that of other popular varieties;the assured procurement of surplusoutput at an incentive price; a supply ofhybrid seed free of cost during the ini-tial stage; a supply of other inputs suchas fertilizer at concessional rates forfarmers who adopt hybrid rice; and awell-organized seed productionsystem. Experiences in the UnitedStates and Japan market economies
during the 1980s, however, indicatethat farmers did not accept hybrid ricebecause of its poor grain quality. NowIndia has emerged as the second coun-try after China, in the development andrelease of high-yielding rice hybrids.The fixing of a support price for hybridrice grains by the government, on a parwith that of popular varieties, wouldcertainly induce rice farmers in irri-gated regions to adopt hybrid rice on alarge scale. ■

Vol. 23, No. 1 33
Instructions for contributorsNOTES
General criteria. Scientificnotes submitted to the IRRN forpossible publication should● be original work,● have international or pan-national relevance,● be conducted during theimmediate past three years orbe work in progress,● have rice environmentrelevance,● advance rice knowledge,● use appropriate researchdesign and data collectionmethodology,● report pertinent, adequatedata,● apply appropriate statisticalanalysis, and● reach supportable conclu-sions.
Routine research. Reports ofscreening trials of varieties,fertilizer, cropping methods,and other routine observationsusing standard methodologiesto establish local recommenda-tions are not ordinarily ac-cepted. Examples are single-season, single-trial fieldexperiments. Field trials shouldbe repeated across more thanone season, in multipleseasons, or in more than onelocation as appropriate. Allexperiments should includereplications and an internation-ally known check or controltreatment.
Multiple submissions. Nor-mally, only one report for asingle experiment will beaccepted. Two or more itemsabout the same work submittedat the same time will bereturned for merging. Submit-ting at different times multiplenotes from the same experi-ment is highly inappropriate.Detection will result in therejection of all submissions onthat research.
IRRN categories. Specify thecategory in which the notebeing submitted should appear.Write the category in the upperright-hand corner of the firstpage of the note.
GERMPLASM IMPROVEMENTgenetic resourcesgeneticsbreeding methodsyield potentialgrain qualitypest resistance
diseasesinsectsother pests
stress tolerancedroughtexcess wateradverse temperatureadverse soilsother stresses
integrated germplasm improve-ment
irrigatedrainfed lowlanduplandflood-prone (deepwater and tidal wetlands)
seed technology
CROP AND RESOURCEMANAGEMENT
soilssoil microbiologyphysiology and plant nutritionfertilizer management
inorganic sourcesorganic sources
crop managementintegrated pest management
diseasesinsectsweedsother pests
water managementfarming systemsfarm machinerypostharvest technologyeconomic analysis
ENVIRONMENTSOCIOECONOMIC IMPACTEDUCATION AND COMMUNI-
CATIONRESEARCH METHODOLOGY
Manuscript preparation.Arrange the note as a briefstatement of research objec-tives, a short description ofproject design, and a succintdiscussion of results. Relateresults to the objectives. Do notinclude abstracts. Do not citereferences or include abibliography. Restrain acknowl-edgments.
Manuscripts must be inEnglish. Limit each note to nomore than two pages of double-spaced typewritten text. Submitthe original manuscript and aduplicate, each with a clear copyof all tables and figures. Authorsshould retain a copy of the noteand of all tables and figures.
Apply these rules, asappropriate, in the note:
● Specify the rice productionecosystems as irrigated, rainfedlowland, upland, and flood-prone(deepwater and tidal wetlands).● Indicate the type of riceculture (transplanted, wetseeded, dry seeded).● If local terms for seasons areused, define them by character-istic weather (wet season, dryseason, monsoon) and bymonths.● Use standard, internationallyrecognized terms to describerice plant parts, growth stages,and management practices. Donot use local names.● Provide genetic background fornew varieties or breeding lines.● For soil nutrient studies,include a standard soil profiledescription, classification, andrelevant soil properties.● Provide scientific names fordiseases, insects, weeds, andcrop plants. Do not use commonnames or local names alone.● Quantify survey data, such asinfection percentage, degree ofseverity, and sampling base.● When evaluating susceptibility,resistance, and tolerance,report the actual quantificationof damage due to stress, whichwas used to assess level orincidence. Specify the measure-ments used.
● Use generic names, not tradenames, for all chemicals.● Use the International Systemof Units for measurements. Forexample, express yield data inmetric tons per hectare (t ha–1)for field studies. Do not uselocal units of measure.● Express all economic data interms of the US$. Do not uselocal monetary units. Economicinformation should be pre-sented at the exchange rateUS$:local currency at the timedata were collected.● When using acronyms orabbreviations, write the namein full on first mention, followedby the acronym or abbreviationin parentheses. Use theabbreviation thereafter.● Define any nonstandardabbreviations or symbols usedin tables or figures in a foot-note, caption, or legend.
Each note can have nomore than two tables and/orfigures (graphs, illustrations, orphotos). All tables and figuresmust be referred to in the text;they should be grouped at theend of the note, each on aseparate page. Tables andfigures must have clear titlesthat adequately explain thecontents.
Review of notes. The IRRNeditor will send an acknowledg-ment card or an e-mailmessage when a note isreceived. An IRRI scientist,selected by the editor, reviewseach note. Reviewer names arenot disclosed. Depending onthe reviewer’s report, a notewill be accepted for publication,rejected, or returned to theauthor(s) for revision.
Comments. If you have com-ments or suggestions about theIRRN, please write to the editor.
Mailing address. Send notesand correspondence to theIRRN Editor, IRRI, P.O. Box 933,Manila 1099, Philippines.Fax: (63-2) 845-0606E-mail: [email protected] page: http://www.cgiar.org/irriRiceweb: http://www.riceweb.orgRiceworld: http://www.riceworld.org

INTERNATIONAL RICE RESEARCH INSTITUTE
P. O. Box 933, 1099 Manila, Philippines
Printed Matter
ISSN 0115-0944





























