Embed Size (px)
Citation preview

BIOCHIMICA ET BIOPHYSICA ACTA
BBA 96952
INTERACTION OF INFORMATIONAL MACROMOLECULES W ITH
RIBOSOMES
IV. DISSOCIATION OF RNA-BINDING ABILITY FROM RIBOSOMES
BY TREATMENT WITH SALTS
269
H. NAORA AND M. J. PRITCHARD Research School o/Biological Sciences, The A ustralian National University, Canberra, A.C.T. 26Ol ( A ustralia)
(Received April 8th, 1971)
SUMMARY
I. An at tempt has been made to dissociate the RNA-binding ability from rat liver ribosomes.
2. Dissociation of this ability from ribosomes was observed by treatment of ribosomes with high concentrations of KC1 or NH~C1.
3. Salt treatment resulted in a detachment of proteins from both large and small subunits of ribosomes without appreciable release of RNA.
4. Proteins required for specific binding of RNA by ribosomes were discussed.
INTRODUCTION
Since selective recognition of homologous RNA by bacterial and animal ribo- somes was observed 1,2, the idea of specificity of ribosomes in binding and translating particular RNA species has been supported in several ways 3-9. The question naturally arises as to which constituents of ribosomes are responsible for recognizing homolo- gous RNA. Recent technical developments have been found useful for an approach to such a problem. For example, during the past few years, the salt-treatment of ribosomes has been used for investigations of the internal organization of ribosomal particles 1°-12 and of the molecular mechanism by which protein synthesis initiates on ribosomes of prokaryotic organisms z8,14.
The present communication describes experiments involving the conditions for dissociation of RNA-binding ability from rat liver ribosomes by treatment with salts.
MATERIALS AND METHODS
Materials
F5-3H]Orotic acid (9.5 C/mmole) and uniformly labeled L-E14C]phenylalanine (37 ° mC/mmole) were purchased from Radiochemical Centre and New England Nuclear Corp., respectively. Deoxyribonuclease (ribonuclease-free) was obtained from Worthington Biochemical Corp. and Sigma Chemical Co. Lyphogel was the product of Gelman Instrument Co. Male albino rats (208-29 ° g body wt.)were used.
Biochim. Biophys. Acta, 246 (1971) 269-279

270 H. NAORA, M. J. PRITCHARD
Preparation o] 3H-labeled nuclear RNA ]rom rat liver cells 3H-labeled nuclear RNA (nRNA) was prepared from rat liver cells as described
in the previous paper 3.
Preparation o/ribosomes Ribosomes were prepared by the method reported in the previous papers 2,3,
except that the concentration of sodium deoxycholate was adjusted to 0. 7 °/o and also by a modification of the method of TASHIRO AND SIEKEVITZ ~-5. The latter procedure involves further purification by precipitating ribosomes by addition of 0. 5 M MgC12 to a final concentration of 50 mM. After standing in ice for 20 mill, the precipitates were collected by centrifugation at IOOO ×g for io min and washed with I mM Tris- HC1 buffer, pH 7.6, containing 5 ° mM MgCI 2. The ribosomes were suspended in I mM Tris-HC1 buffer, pH 7.6, containing I mM MgC12 and dialyzed against IOO vol. of tile same buffer solution. After dialysis overnight, the preparation was clarified by centrifugation at io ooo ×g for IO min. The freshly prepared ribosomes were imme- diately used for further treatment.
Treatment with salts and sucrose density gradient centri]ugation Unless otherwise noted, ribosomes were diluted to 0. 7 mg/ml by adding Tris-
HCI or triethanolamip.e-HC1 buffer, pH 7.6, containing 1. 5 mM MgC12 and concen- trated KC1 or NH4C1 to adjust to the final concentrations of I mM buffer, pH 7.6, 1. 5 mM MgCI 2 and 0.5 to 2.0 M KC1 or NH4C1, depending upon the experiments. The suspensions were kept in ice for 0.5 to 5 h and centrifuged at 15o ooo ×g for 15o min. The pellets were rinsed with I mM triethanolamine buffer, pH 7.5, containing 5 mM KC1 and finally suspended in the same buffer. In some experimentes, the ribo- somes suspended in a high concentration of salt solution were directly layered on 28 ml of a linear sucrose gradient (15-3o % or 5-20 %) in 5 or 50 mM KC1, o.oi mM MgC12 and I mM triethanolamine-HC1 or Tris-HC1 buffer, pH 7.5- The gradients were centrifuged at 22 ooo rev./min for 16 h or 25 ooo rev./min for 8 h in an SW-25.I rotor. After centrifugation the gradient was fractionated into I-ml portions with the aid of an automatic ISCO density-gradient fractionator.
Isopycnie centri/ugation CsC1 gradients were preformed in I mM triethanolamine-HC1, pH 7.5, contain-
ing I mM MgC12 from 0. 7 , 1. 4 and 1. 4 ml of CsC1 solutions of 1.6o, 1.65 and 1.75 or 1.7o, 1.75 and 1.8o g/cm 3, respectively 18.
Fractions from sucrose gradients in triethanolamine-HC1 buffer to be examined were fixed with formaldehyde as described by PERRY AND KELLY 17. The samples were then concentrated by adding sufficient amounts of lyphogel and leaving for 8 to 12 h at 5 °. 0.5 ml of the sample was layered on the gradient. Centrifugation was routinely performed for 16 h at 32 ooo rev./min in an SW-39 rotor. Extending to 48 11 did not change the banding pattern, indicating that equilibrium had been reached in 16 h. io-drop fractions, except every 5th fraction, were collected from the bot tom of the centrifuge tubes for determination of absorbance at 260 m#. 5 drops of every 5th fraction were obtained and assayed for density by weighing in a 0.05 ml cali- bl ated pipette.
Biochim. Biophys. Acta, 246 (1971) 269-279

DISSOCIATION OF RNA-BINDING ABILITY 271
Treatment with EDTA Purified ribosomes were suspended in i mM triethanolamine-HC1, pH 7-5, con-
raining o.I mM MgC1, and 50 mM KC1 at a concentration of IO mg/ml and completely dissociated into subunits by EDTA treatment at a concentration of 2.5 pmoles/mg ribosome as described by TASHIRO AND SIEKEVITZ 15. I ml of treated ribosomes was layered on a linear sucrose density gradient (5-20 %) in I mM triethanolamine-HC1, pH 7.5, containing o.I mM MgC1, and 5 ° mM KC1. The gradient was centrifuged at 25 ooo rev./min for 8 h in the SW-25.I rotor TM. The gradient was fractionated as described above. The large and small subunits thus fractionated were fixed with form- aldehyde and assayed for density.
An in vitro assay/or E14Clphenylalanine incorporation The basic incubation mixture contained in 0.25 ml: o.oI5/~mole of 20 L-
amino acids excluding phenylalanine, 0.2/,mole of ATP, o.o8pmole of GTP, 6.0 pmoles of Tris-HC1 buffer, pH 7.6, 12.5/~moles of NH,C1, 2.5/zmoles of MgC12, 1.5 /,moles of mercaptoethanol, 2.5 #moles of phosphoenolpyruvate, 5.o/zg of crystal- liue pyruvate kinase, 0.05/~C of L-E14Clphenylalanine, 0.2 mg of dry mat ter of 15o ooo ×g supernatant, 12.4 #g RNA equivalent of ribosomes and IOO/~g of poly(U) where indicated.
Incubation was carried out at 37 ° for 3 ° min with occasional agitation. The reaction was stopped by transferring the tubes to an ice bath. o.I-ml aliquots of the reaction mixture were acid-precipitated on filter paper discs according to the proce- dure of MANS AND NOVELL119. The discs were then washed twice with 5 % trichloro- acetic acid, heated at 9 °0 for 15 min with 5 % trichloroacetic acid twice and finally washed with 5 % trichloroacetic acid. The radioactivity of the discs thus prepared was detelmined in a dioxane-based scintillation fluid. The amount of the labeled phenylalanine incorporated into proteins by treated ribosomes is expressed as a per- centage of that by untreated ribosomes.
The 15o ooo ×g supernatant was obtained from post-mitochondrial fraction of rat liver homogenate by centrifugation at 15o ooo ×g for 18o rain.
Assay o/labeled RNA-binding by ribosomes Binding was assayed by the Millipore filter method described previously 1, 2. The
amount of labeled RNA bound by treated ribosomes is expressed as per cent of the untreated ribosomes.
Protein and RNA determinations Protein was measured by the method of LOWRY et al. 2°. RNA was determined
by the procedure of WEBB 21.
RESULTS
EHect o/salt treatment on RNA-binding ability In order to dissociate tile RNA-binding ability from the ribosomal particles,
an experiment was carried out in which ribosomes were treated with varied concen- trations of KC1 or NH4C1 for 30 min under the conditions described under METHODS
AND MATERIALS and the RNA-binding ability of the resultant particles was assayed.
Bioehim. Biophys. Acta, 246 (1971) 269-279
272 H. NAORA, M. J. PRITCHARD
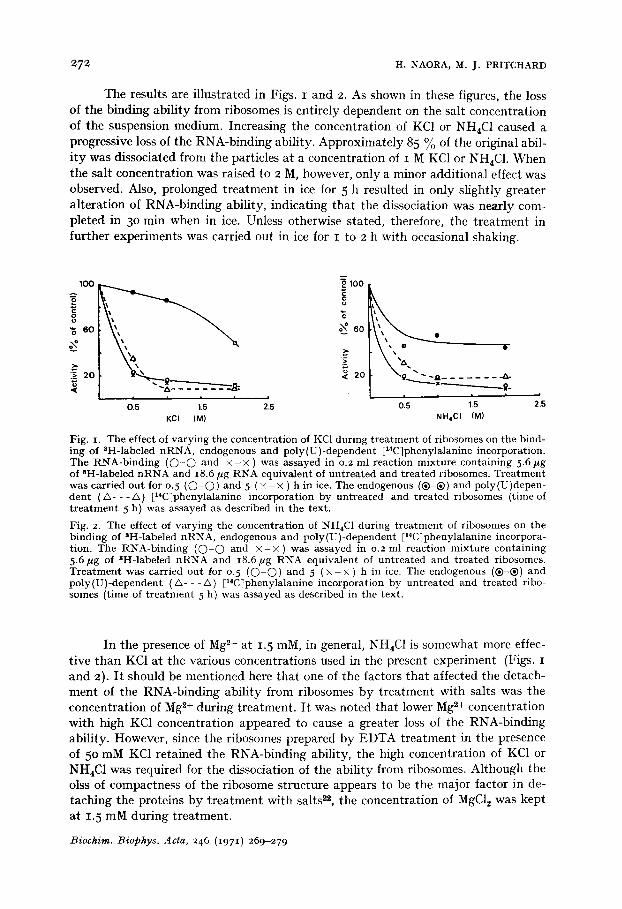
The results are illustrated in Figs. I and 2. As shown in these figures, the loss of the binding ability from ribosomes is entirely dependent on the salt concentration of the suspension medium. Increasing the concentration of KC1 or NH4C1 caused a progressive loss of the RNA-binding ability. Approximately 85 % of the original abil- i ty was dissociated from the particles at a concentration of I M KC1 or NH4C1. When the salt concentration was raised to 2 M, however, only a minor additional effect was observed. Also, prolonged treatment in ice for 5 h resulted in only slightly greater alteration of RNA-binding ability, indicating that the dissociation was nearly com- pleted in 3o min when in ice. Unless otherwise stated, therefore, the t reatment in further experiments was carried out in ice for I to 2 h with occasional shaking.
1OO
o
,~ 60
N
i~ 20 '6 .<
o 100
u
o~ 6o
2o
i i | i J
0.5 1.5 2.5 KCi (M)
o
~ - - ~ . . . . . . . .&- ~ - - - - - - . ~ ~ .
t t t =
0.5 1.5 NH4CI (M)
D
2.5
Fig. I. The effect of varying the concentration of KC1 during t rea tment of ribosomes on the bind- ing oI SH-labeled nRNA, endogenous and poly(U)-dependent [14C]phenylalanine incorporation. The RNA-binding ( O - O and × - × ) was assayed in 0.2 ml reaction mixture containing 5.6 ~g of 8H-labeled n RNA and i8.6/~g RNA equivalent of untreated and treated ribosomes. Trea tment was carried out for 0.5 ( O - O ) and 5 ( × - × ) h in ice. The endogenous (®-®) and poly(U)depen- dent ( & - - - / ' , ) [x4C]phenylalanine incorporation by untreated and treated ribosomes (time of t r ea tment 5 h) was assayed as described in the text.
Fig. 2. The effect of varying the concentration of NH4C1 during t rea tment of ribosomes on the binding of 8H-labeled nRNA, endogenous and poly(U)-dependent [14C]phenylalanine incorpora- tion. The RNA-binding ( O - O and × - × ) was assayed in 0.2 ml reaction mixture containing 5.6/~g of 3H-labeled nRNA and 18.6/~g RNA equivalent of untreated and treated ribosomes. Trea tment was carried out for 0. 5 ( O - O ) and 5 ( x - x ) h in ice. The endogenous (®-®) and poly(U)-dependent (&- - -/x) [14C]phenylalanine incorporation by untreated and treated ribo- somes (time of t r ea tment 5 h) was assayed as described in the text.
In the presence of Mg 2+ at 1.5 raM, in general, NH4C1 is somewhat more effec- tive than KC1 at the various concentrations used in the present experiment (Figs. I and 2). I t should be mentioned here that one of the factors that affected the detach- ment of the RNA-binding ability from ribosomes by treatment with salts was the concentration of Mg *+ during treatment. I t was noted that lower Mg ~+ concentration with high KC1 concentration appeared to cause a greater loss of the RNA-binding ability. However, since the ribosomes prepared by EDTA treatment in the presence of 5 ° mM KC1 retained the RNA-binding ability, the high concentration of KC1 or NH4C1 was required for the dissociation of the ability from ribosomes. Although the olss of compactness of the ribosome structure appears to be the major factor in de- taching the proteins by t reatment with salts 2., the concentration of MgC12 was kept at 1. 5 mM during treatment.
Biochim. Biophys. Acta, 246 (1971) 269-279

DISSOCIATION OF RNA-BINDING ABILITY 273
I t was observed that the concentration of ribosomes in the salt medium affects the dissociation of the RNA-binding ability. When the ribosome concentration was increased in a given medium, less dissociation was observed. This may be probably due to the changes in the total amounts of Mg ~+ in the sample. However, such a concentration dependency was found to be variable, especially at the high concen- tration of ribosomes. Therefore, the quantitat ive reproducibility in the extent of dis- sociation was somewhat poor and varied from preparation to preparation. Such a variation was more serious when ribosomes were treated with the less concentrated salt solution.
E][ect o] salt treatment on endogenous and poly (U) dependent [14C~Jphenylalanine incor- poration
The loss of the RNA-binding ability from the ribosomes treated with varied concentrations of KC1 or NH4C1 was compared with the alteration of endogenous and poly(U) dependent [14Cjphenylalanine incorporation.
Figs. I and 2 also illustrate the results obtained in these experiments. During t reatment with KC1, the activity of polyphenylalanine synthesis was lost in a manner almost similar to that of the RNA-binding ability. The same effect was observed with NH4C1, although slightly more marked changes took place in the former activity than in the latter. Since the salt- treatment also resulted in the detachment of a factor responsible for stimulation of polyphenylalanine synthesis ~3, the result observed here indicates that both factors required for binding RNA, if present, and for polyphenyl- alanine synthesis were detached from ribosomes in the similar manner.
An interesting observation was that the endogenous incorporation was decreased by low concentrations of NH4CI but not by KC1. Such an effect of NH4CI may have a positive correlation with its slightly stronger effect on the dissociation of RNA- binding ability than KC1. As shown in Fig. I, under the condition where 85 % of polyphenylalanine synthetic activity was lost by t reatment with I M KC1, most of the ribosomes already engaged in protein synthesis remained intact. This observation was in good agreement with those made with other organisms 24,~5. However, the en- dogenous incorporation was affected by 2 M KC1, suggesting that polysome structure appeared to be unstable in such a high concentration of KC1.
Sucrose densi(v gradient centri[ugation pattern o] salt-treated ribosomes The possible alteration of ribosomal particles resulting from salt t reatment was
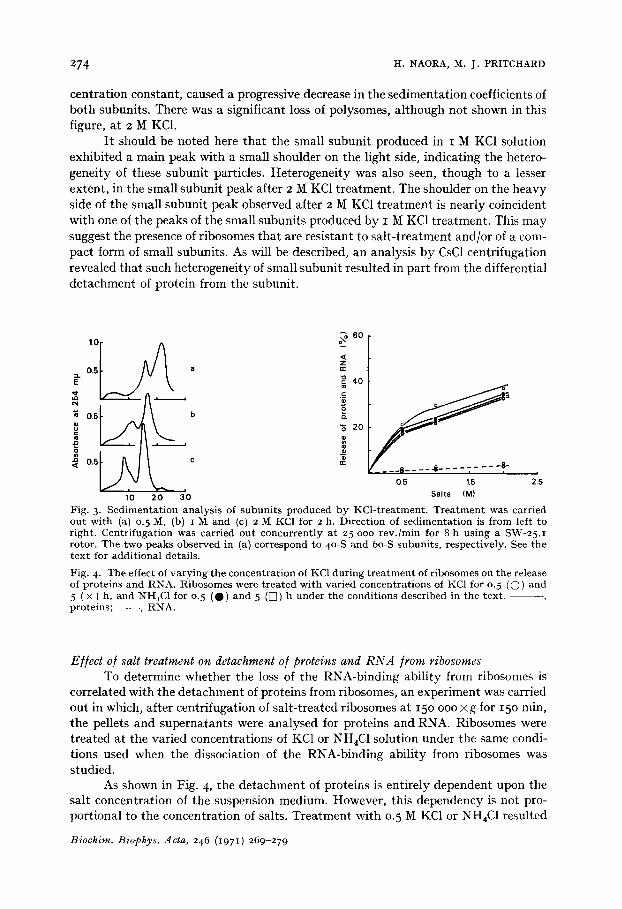
analyzed by sucrose density gradient centrifugation. As shown in Fig. 3, the addition of KC1 caused remarkable changes in the sedi-
mentation patterns of ribosomes. Suspension of rat liver ribosomes in 0.5 M KC1 under the condition described under MATERIALS AND METHODS resulted in the formation of small and large subunits. 'The complete reassociation of these subunits occurred when the subunits were exposed to low ionic strength buffer in the presence of Mg ~+ (i mM triethanolamine-HC1 or Tris-HC1, pH 7.6, 5 mM KC1 and 1.5 mM MgCI~). I t is for this reason that the concentration of MgC12 in the gradient was reduced to o.oi mM. Under the conditions used for centrifugation in Fig. 3, monosomes and polysomes which existed in the suspension were sedimentated on the bot tom and therefore were not shown in this figure. Increasing concentrations of KC1, keeping the ribosome con-
Biochim. Biophys. Acta, 246 (1971) 269-279
274 H. NAORA, M. J. PRITCHARD
centration constant, caused a progressive decrease in the sedimentation coefficients of both subunits. There was a significant loss of polysomes, although not shown in this figure, at 2 M KC1.
I t should be noted here that the small subunit produced in I M KC1 solution exhibited a main peak with a small shoulder on the light side, indicating the hetero- geneity of these subunit particles. Heterogeneity was also seen, though to a lesser extent, in the small subunit peak after 2 M KC1 treatment. The shoulder on the heavy side of the small subunit peak observed after 2 M KC1 treatment is nearly coincident with one of the peaks of the small subunits produced by I M KC1 treatment. This may suggest the presence of ribosomes that are resistant to salt- treatment and/or of a com- pact form of small subunits. As will be described, an analysis by CsC1 centrifugation revealed that such heterogeneity of small subunit resulted in part from the differential detachment of protein from the subunit.
tN
c
<
1.0
0.5 a
0.5 b
0.5 c
10 20 30
~ 60 v
z <
4o
c
~ 20
s • 8 . . . . . . . . . . . . "j . . . . . -
0 , 5 1.5 2'.5 Salts (M)
Fig. 3. S e d i m e n t a t i o n ana lys i s of s u b u n i t s p roduced by KCl - t r ea tmen t . T r e a t m e n t was carr ied ou t wi th (a) 0. 5 M, (b) i !V[ and (c) 2 M KC1 for 2 h. Direct ion of s ed imen ta t i on is f rom left to r ight . Cen t r i fuga t ion was carr ied ou t concur ren t ly a t 25 ooo r ev . /m in for 8 h us ing a SW-25. I rotor. The two peaks observed in (a) cor respond to 4o-S and 6o-S subun i t s , respect ively . See t he t ex t for add i t iona l details .
Fig. 4. The effect of va ry i ng t he concen t r a t ion of KC1 dur ing t r e a t m e n t of r ibosomes on t he release of pro te ins and RNA. R ibosomes were t r ea t ed wi th var ied concen t ra t ions of KC1 for 0. 5 (O) and 5 ( × ) h, and NHtC1 for o. 5 (O) and 5 ([[]) h unde r t he condi t ions descr ibed in t he tex t . - - p r o t e i n s ; - - - , RNA.
E[]ect o/salt treatment on detachment o/ proteins and RNA /rom ribosomes To determine whether the loss of the RNA-binding ability from ribosomes is
correlated with the detachment of proteins from ribosomes, an experiment was carried out in which, after centrifugation of salt-treated ribosomes at 15o ooo >~'g for 15o rain, the pellets and supernatants were analysed for proteins and RNA. Ribosomes were treated at the varied concentrations of KC1 or NH4C1 solution under the same condi- tions used when the dissociation of the RNA-binding ability from ribosomes was studied.
As shown in Fig. 4, the detachment of proteins is entirely dependent upon the salt concentration of the suspension medium. However, this dependency is not pro- portional to the concentration of salts. Treatment with 0.5 M KC1 or NH4C1 resulted
Biochim. Biophys. Acta, 246 (1971) 269-279

DISSOCIATION OF RNA-BINDING ABILITY 275
in an approx. 17 to 20 °//o lelease of proteins into supernatant fraction. However, this does not necessarily mean that all of the proteins found in the supernatant fraction were detached from particles themselves. Since the ribosomes used in this experiment were not highly purified, proteins contaminated the ribosome preparations used. In- deed, the ratios of protein :RNA of the ribosome preparations were o.82-1.22 :I. Addi- tion of more KC1 or NH4C1 to the ribosome suspension resulted in a release of additi- onal proteins into the supernatant fraction. This was probably due to the detachment of proteins from ribosomal particles. The total amount of protein removed increased when the salt concentration was raised. When the purified and crude preparations of ribosomes were treated under identical conditions, the respective protein contents of treated ribosomes had almost similar values, indicating that the detachment of proteins from particles was not significantly affected by the presence of proteins con- taminating the preparations.
As in the case of dissociation of the RNA-binding ability from ribosomes, the release of protein from ribosomes by the given concentration of both KC1 and NH4C1 was almost completed within 30 min in ice. Prolonged treatment for 5 h caused an additional release of only small amounts of protein although the effect of NH4C1 appeared to be slightly less instantaneously (Fig. 4).
Another factor that affected the detachment of proteins was the ribosome con- centration. When the higher concentration of ribosomes was used, less protein was detached. For example, when ribosomes were treated with I M KC1 at the concen- tration of 0. 7 mg/ml ribosomes for 3 h, 29.0 % of protein was detached. At 2.2 mg/ml, however, 19. 9 % of protein was removed. Such a dependency oil the concentration of ribosomes is almost similar to that in the loss of the RNA-binding ability from ribosomes.
It was of interest that the salt treatment did not result in the removal of sig- nificant amounts of RNA from ribosomes as compared with the loss of protein and the RNA-binding ability (Fig. 4)- As described earlier, treatment with I M KCI or NH4C1 caused the dissociation of more than 80 % of the RNA-binding ability. Under these conditions, only 2-3 % of RNA was released from the particles. An analysis of this released RNA by sucrose density gradient centrifugation showed low molecular species to be present, probably tRNA and/or 5-S RNA.
Taken altogether with the results on the dissociation of the RNA-binding ability from ribosomes, it appears likely that KC1 or NH4C1 removed proteins required for binding of RNA by ribosomes.
E//ect o~ salt-treatment on buoyant densi~, o/ subunits The next question to arise concerned the detachment of proteins from large and
small subunits of ribosomes during salt treatment. As described above, salt-treatment caused the dissociation of ribosomes into their subunits without appreciable release of RNA. Since the reciprocal of the buoyant density of particles may be used empiri- cally as a measure of the relative protein content 26, isopycnic centrifugation of the resulting subunits is a sensitive way to approach this problem.
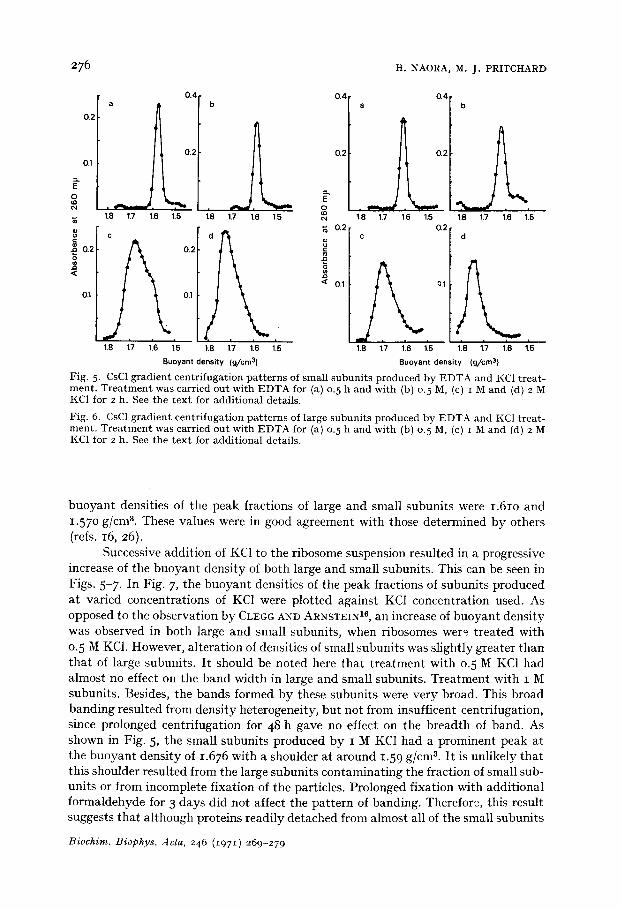
Figs. 5 and 6 showed the isopycnic banding patterns of salt-treated subunits. In these experiments, the buoyant densities of the large and small subunits produced by salt treatment were compared with those of subunits obtained by EDTA treat- ment. The patterns of banding by EDTA-treated subunits are monodisperse and the
Biochim. Biophys. Acta, 246 ~i97 I) 269-279
276 H. NAORA, M. J. PRITCHARD
0.2
0.4 a
0.;
. _ J . 1.8 1.7 1.6 1.5
1 1~8 1.7 1.6 1.5
0.4
Q2 0.1
oo .~ 0.2
< 0.1
,B ,'7 ,'6 ,:5
c
0.2
0.1
i 1.8 1.7 1.6 1.5
Buoyant density (g/cm 3)
0.1
0.4 s
, m ~ 0"2
1.8 1.7 1.6 1.5
1.8 1.7 1.6 1.5
1.8 1.7 1.6 1.5 0.2[ d
1.8 1.7 1.6 1.5 Buoyant density (g/cm 3)
Fig. 5. CsC1 g rad ien t cen t r i fuga t ion p a t t e r n s of smal l s u b u n i t s p roduced by E D T A and KC1 t r ea t - men t . T r e a t m e n t was carr ied ou t wi th E D T A for (a) o. 5 h and wi th (b) o. 5 M, (c) I M and (d) 2 M KC1 for 2 h. See t he t e x t for addi t iona l details.
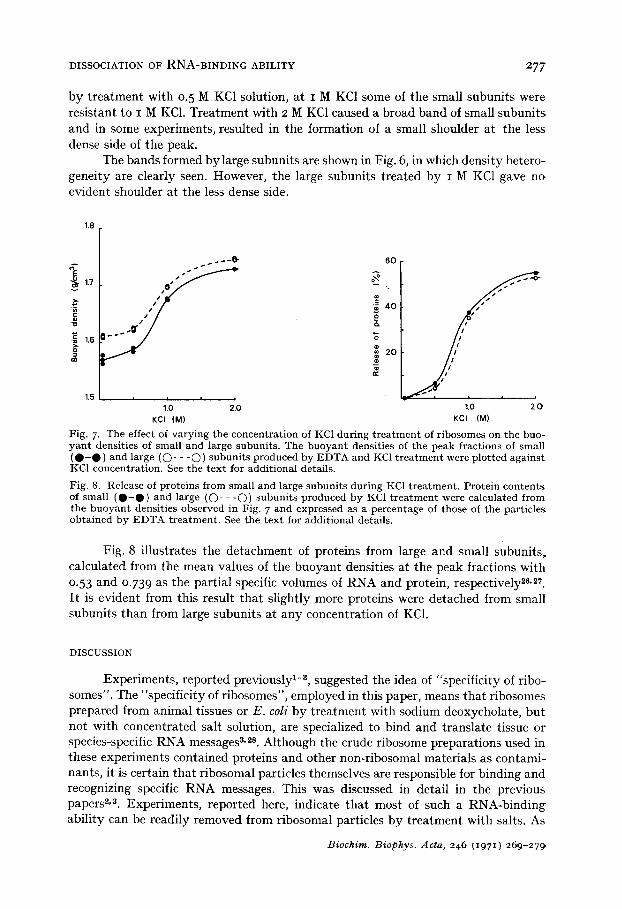
Fig. 6. CsC1 g rad ien t cen t r i fuga t ion p a t t e r n s of large s u b u n i t s p roduced by E D T A and KC1 t r ea t - men t . T r e a t m e n t was carr ied ou t wi th E D T A for (a) o. 5 h and wi th (b) o. 5 M, (c) i M and (d) 2 M KC1 for 2 h. See t he t ex t for add i t iona l details .
buoyant densities of the peak fractions of large and small subunits were 1.61o and 1.57o g/cnP. These values were in good agreement with those determined by others (refs. 16, 26).
Successive addition of KC1 to the ribosome suspension resulted in a progressive increase of the buoyant density of both large and small subunits. This can be seen in Figs. 5-7. In Fig. 7, the buoyant densities of the peak fractions of subunits produced at varied concentrations of KC1 were plotted against KC1 concentration used. As opposed to the observation by CLEGG A N D ARNSTEIN 16, an increase of buoyant density was observed in both large and small subunits, when ribosomes wer~ treated with 0. 5 M KC1. However, alteration of densities of small subunits was slightly greater than that of large subunits. I t should be noted here that t reatment with 0.5 M KC1 had almost no effect on the band width in large and small subunits. Treatment with I M subunits. Besides, the bands formed by these subunits were very broad. This broad banding resulted from density heterogeneity, but not from insufficent centrifugation, since prolonged centrifugation for 48 h gave no effect on the breadth of band. As shown in Fig. 5, the small subunits produced by I M KC1 had a prominent peak at the buoyant density of 1.676 with a shoulder at around 1.59 g/cm 8. I t is unlikely that this shoulder resulted from the large subunits contaminating the fraction of small sub- units or from incomplete fixation of the particles. Prolonged fixation with additional formaldehyde for 3 days did not affect the pattern of banding. Therefore, this result suggests that although proteins readily detached from almost all of the small subunits
Biochim. Biophys. Acta, 246 (1971) 269-279
DISSOCIATION OF R N A - B I N D I N G ABILITY 2 7 7
by t reatment with o.5 M KC1 solution, at I M KCI some of the small subunits were resistant to I M KC1. Treatment with 2 M KC1 caused a broad band of small subunits and in some experiments, resulted in the formation of a small shoulder at the less dense side of the peak.
The bands formed bylarge subunits are shown in Fig. 6, in which density hetero- geneity are clearly seen. However, the large subunits treated by I M KC1 gave no evident shoulder at the less dense side.
1.8
E 1.7
1.6
1.5
--8-
ly x
6O
III t t / t / / -'0" 40
20 /
t t t J
1.0 2.0 1.0 2.0 KCI (M) KCI (M)
Fig. 7. The effect of v a r y i n g t he concen t ra t ion of KCI du r ing t r e a t m e n t of r ibosomes on t he buo- y a n t densi t ies of smal l and large subun i t s . The b u o y a n t densi t ies of the peak f rac t ions of smal l ( • - • ) and large ((~)- - - O ) s u b u n i t s p roduced by E D T A and KC1 t r e a t m e n t were p lo t ted a g a i n s t KC1 concen t ra t ion . See t he t e x t for addi t iona l details.
Fig. 8. Release of pro te ins f rom smal l and large s u b u n i t s du r ing KC1 t r e a t m e n t . P ro te in c o n t e n t s of smal l ( • - • ) and large (O- - - O ) s u b u n i t s p roduced b y KC1 t r e a t m e n t were ca lcula ted f rom the b u o y a n t densi t ies observed in Fig. 7 and expressed as a pe rcen tage of those of t he par t ic les ob ta ined by E D T A t r e a t m e n t . See t he t ex t for addi t iona l details .
Fig. 8 illustrates the detachment of proteins from large and small subunits, calculated from the mean values of the buoyant densities at the peak fractions with 0.53 and 0.73 9 as the partial specific volumes of RNA and protein, respectively 26,27. I~ is evident from this result that slightly more proteins were detached from small subunits than from large subunits at any concentration of KC1.
DISCUSSION
Experiments, reported previously 1-~, suggested the idea of "specificity of ribo- somes". The "specificity of ribosomes", employed in this paper, means that ribosomes prepared from animal tissues or E. coli by t reatment with sodium deoxycholate, but not with concentrated salt solution, are specialized to bind and translate tissue or species-specific RNA messages 3,2s. Although the crude ribosome preparations used in these experiments contained proteins and other non-ribosomal materials as contami- nants, it is certain that ribosomal particles themselves are responsible for binding and recognizing specific RNA messages. This was discussed in detail in the previous papers 2,3. Experiments, reported here, indicate that most of such a RNA-binding ability can be readily removed from ribosomal particles by treatment with salts. As
Biochim. Biophys. Acta, 246 ~i97 I) 269-279

278 H. NAORA, M. J. PRITCHARD
described under RESULTS, the effectiveness of the salts is slightly dependent upon the nature of the particular salts used in this experiment. I M KC1 had a significant effect on the dissociation of the RNA-binding ability from ribosomes without appreciable effect on the release of RNA or the inhibition of endogenous amino acid incorporation. I t is for this reason that t reatment with I M KC1 was used for further experiments.
While a series of investigations on tile dissociation of the RNA-binding ability from ribosomes by salt t reatment have been carried out in our laboratory, reports from other laboratories provide evidence that the detachment of proteins from ribo- somes by salts is an ordered and stepwise phenomenon 1°,1e. The present experiments also revealed almost no release of RNA but marked detachment of proteins under the condition where most of the RNA-binding ability was dissociated. I t appears likely, therefore, that some of the proteins released by salts are responsible for the specific recognition of homologous RNA by intact ribosomes. Indeed, experiments to be described in the following paper 29, show that this is the case. The proteins them- selves released by treatment with KC1 or NH4C1 were highly capable of binding labeled RNA but failed to demonstrate any specific recognition of homologous RNA. How- ever, ribosomes reconstituted from KCl-treated particles and released proteins possess properties of RNA-binding specificity similar to untreated ribosomes from which proteins were isolated. This implies that the tissue specificity of ribosomes is entirely dependent upon the source of proteins which can be removed by salts, that these proteins bind correctly to their original positions on tile particles and that these positions remained intact during salt treatment. I t is also evident that electrostatic forces play a major role in binding proteins to the correct positions on the particles. Although slightly more proteins were detached from small subunits than large sub- units during KC1 treatment, no definite conclusion was reached in the present ex- periments concerning small subunit origin of the released proteins 14. Since the binding takes place on small subunit (H. NAORA, unpublished results), this possibility appears to be likely. This is now being studied. However, since a simple definition of what constitutes a ribosome is not yet possible, the question remains as to whether the proteins required for RNA-binding are the intrinsic components of ribosomes or something additional.
Finally, it should be noted here that the initiation protein factor, F 8 or B, has been purified from the proteins released from bacterial ribosomes by concentrated NH4C1 s° and appeared to be responsible for binding specific RNA messages s. A similar protein factor was also isolated from animal ribosomes by treatment with I M KC1 al. In accordance with our results ~9, such a protein was required to recognize specific RNA messages when it associated with the i M KCl-treated particle.
ACKNOWLEDGEMENT
Tile authors thank Miss Joy M. Whitelam for her skillful technical assistance.
R E F E R E N C E S
i H. NAORA. AND K. KODAIRA, Biochim. Biophys. Acta, 161 (1968) 276. 2 FI. NAORA AND K. KODAIRA, Biochim. Biophys. Acta, 182 (1969) 469. 3 H. NAORA AND K. ]5~ODAIRA, Biochim. Biophys. Acta, 209 (197 o) 196. 4 FI. F. LODISH, Nature, 224 (1969) 867.
Biochim. Biophys. Acta, 246 (1971) 269-279

DISSOCIATION OF R N A - B I N D I N G ABILITY 279
5 W. T. H s u AND S. B. WEISS, Proc. Natl. Acad. Sci. U.S., 64 (1969) 345- 6 S. K. DUDE AND P. S. RUDLAND, Nature, 226 (197 o) 82o. 7 J. A. STEITZ, S. K. DUDE, P. S. RUDLAND, Nature, 226 (197 o) 824. 8 M. RIgVEL, H. AVlV, Y. DRONER AND Y. POLLACK, F E B S Lett., 9 (197 o) 213. 9 Y. POLLACK, Y. GRONER, H. AVlV AND M. REVEL, F E B S Left., 9 (197 o) 218.
IO A. ATSMON, P. SPITNIK-ELSON AND D. ELSON, J. Mol. Biol., 45 (1969) 125. i i T. ITOH, E. OTAKA AND S. OSAWA, J. Mol. Biol., 33 (1968) lO9. 12 P. TRAUB, in Current Topics in Microbiology and Immunology, Vol. 52, Springer-VeriEr, Berlin,
197 o, p. I. 13 M. REVEL, M. HERZBERG AND H. GREENSHPAN, Cold Spring Harbor Syrup. Quant. Biol., 34
(1969) 261. I 4 A. J . WAHBA, Y. g . CHAE, K. IWASAKI, R. MAZUMDER, M. J. MILLER, S. SABOL AND M. A. G.
SILLERO, Cold Spring Harbor Syrup. Quant. Biol., 34 (1969) 285. 15 Y. TASHIEO AND P. SIEKEVITZ, J. Mol. Biol., I I (1965) 149. 16 j . c . s . CLEGG AND H. R. V. ARNSTEIN, Eur. J. Biochem., 13 (197 o) 149. 17 R. P. PERRY AND D. E. KELLEY, J. Mol. Biol., 16 (1966) 255. 18 ¥ . TASrtlRO AND T. MORI~tOTO, Biochim. Biophys. Acta, 123 (1966) 523 . 19 R. J. MANS AND G. D. NOVELLI, Arch. Bioehem. Biophys., 94 (1961) 48. 2o O. H. LOWRY, N. J. ROSEBROUGH, A. L. FARE AND R. J. RANDALL, J. Biol. Chem., 193 (1951)
265. 21 J . M. WEBB, J. Biol. Chem., 22I (1956) 635. 22 P. SPITmK-ELsoI~ AND A. ATSMON, J. Mol. Biol., 45 (1969) 113. 23 Y. W. KAN, F. GOLINI AND R. E. THACa, Proc. Natl. Acad. Sci. U.S., 67 (197 o) 1137. 24 T. E. MARXlN AND L. H. HARTWELL, J. Biol. Chem., 245 (I97 o) 15o 4. 25 E. A. ZYLBEa A~D S. PEI~MAN, Biochim. Biophys. Acta, 204 (197 o) 221. 26 M. G. HAMILTON AND g . E. RUTa, Biochemistry, 8 (1969) 851. 27 M. L. PETERMANN AND A. PAVLOVEC, Biochim. Biophys. Acta, 114 (1966) 264. 28 H. NAORA AND K. KODAIRA, Biochim. Biophys. Acta, 224 (197 o) 498. 29 H. IN'AORA, K. KODAIRA AND M. J. PRITCHARD, Biochim. Biophys. Acta, 246 (1971) 280. 3 ° S. SABOL, M. A. Cr. SILLERO, K. IWASAKI AND S. OCHOA, Nature, 228 (197 o) 1269. 31 S. M. HEYWOOD, Proc. Natl. Acad. Sci. U.S., 67 (197 o) 1782.
Biochim. Biophys. Mcta, 246 (1971) 269-279









![Pressure-Induced Dissociation Sedimenting …Pressure-Induced Dissociation of Ribosomes 1781 3 2 0C (0 w z a] 0 3 2 E EC DISTANCE FROM MENISCUS (cm) FIG. 2. Effect of length of centrifugation](https://img.pdfslide.us/doc/110x75/5fd2d4ad2ccfe1396b271cef/pressure-induced-dissociation-sedimenting-pressure-induced-dissociation-of-ribosomes.jpg)