Embed Size (px)
Citation preview

Vol.:(0123456789)1 3
Plant Molecular Biology (2018) 96:245–263 https://doi.org/10.1007/s11103-017-0692-7
Interaction network of core ABA signaling components in maize
Ying‑Ge Wang1 · Feng‑Ling Fu1 · Hao‑Qiang Yu1 · Tao Hu2 · Yuan‑Yuan Zhang1 · Yi Tao1 · Jian‑Kang Zhu2 · Yang Zhao2 · Wan‑Chen Li1
Received: 14 September 2017 / Accepted: 6 December 2017 / Published online: 17 January 2018 © Springer Science+Business Media B.V., part of Springer Nature 2018
AbstractKey message We defined a comprehensive core ABA signaling network in monocot maize, including the gene expres-sion, subcellular localization and interaction network of ZmPYLs, ZmPP2Cs, ZmSnRK2s and the putative substrates.Abstract The phytohormone abscisic acid (ABA) plays an important role in plant developmental processes and abiotic stress responses. In Arabidopsis, ABA is sensed by the PYL ABA receptors, which leads to binding of the PP2C protein phosphatase and activation of the SnRK2 protein kinases. These components functioning diversely and redundantly in ABA signaling are lit-tle known in maize. Using Arabidopsis pyl112458 and snrk2.2/3/6 mutants, we identified several ABA-responsive ZmPYLs and ZmSnRK2s, and also ZmPP2Cs. We showed the gene expression, subcellular localization and interaction network of ZmPYLs, ZmPP2Cs, and ZmSnRK2s, and the isolation of putative ZmSnRK2 substrates by mass spectrometry in monocot maize. We found that the ABA dependency of PYL-PP2C interactions is contingent on the identity of the PP2Cs. Among 238 candidate substrates for ABA-activated protein kinases, 69 are putative ZmSnRK2 substrates. Besides homologs of previously reported putative AtSnRK2 substrates, 23 phosphoproteins have not been discovered in the dicot Arabidopsis. Thus, we have defined a comprehensive core ABA signaling network in monocot maize and shed new light on ABA signaling.
Keywords Maize · ABA signaling · ZmPYLs · ZmPP2Cs · ZmSnRK2s · Signaling network
Introduction
Abscisic acid (ABA) is an important hormone that regu-lates plant growth and development, and responses to abi-otic stresses, including drought, high salinity and low tem-perature (Cutler et al. 2010; Zhu 2002, 2016). Although the biosynthesis, physiological effects and signal transduction of ABA were extensively investigated from 1960s (Cutler et al. 2010; Nambara and Marion-Poll 2005; Ohkuma et al. 1963), the perception of ABA had remained unclear until the identification of pyrabactin resistance 1 (PYR1) and PYR1-like (PYL)/regulatory component of ABA recep-tor proteins (RCARs) in Arabidopsis (Ma et al. 2009; Park et al. 2009). The ABA-bound PYLs interact with clade A protein phosphatases type 2Cs (PP2Cs) and prevent them
Ying-Ge Wang and Feng-Ling Fu have contributed equally to this work.
Accession numbers The corresponding accession numbers of these ZmPP2Cs at maizeGDB were GRMZM2G102255 (ZmPP2C1), GRMZM2G134628 (ZmPP2C2), GRMZM2G059453 (ZmPP2C3), GRMZM2G177386 (ZmPP2C4), GRMZM2G437575 (ZmPP2C5), GRMZM2G166297 (ZmPP2C6), GRMZM2G383807 (ZmPP2C7), GRMZM2G019819 (ZmPP2C8), GRMZM2G122228 (ZmPP2C9), GRMZM2G001243 (ZmPP2C10), GRMZM2G308615 (ZmPP2C11), GRMZM5G818101 (ZmPP2C12), GRMZM2G159811 (ZmPP2C13), GRMZM2G010855 (ZmPP2C14), GRMZM2G300125 (ZmPP2C15) and GRMZM2G082487 (ZmPP2C16), respectively.
Electronic supplementary material The online version of this article (https://doi.org/10.1007/s11103-017-0692-7) contains supplementary material, which is available to authorized users.
* Yang Zhao [email protected]
* Wan-Chen Li [email protected]
1 Maize Research Institute, Sichuan Agricultural University, Chengdu 611130, Sichuan, China
2 Shanghai Center for Plant Stress Biology, and CAS Center for Excellence in Molecular Plant Sciences, Chinese Academy of Sciences, Shanghai 200032, China

246 Plant Molecular Biology (2018) 96:245–263
1 3
from inhibiting the sucrose non-fermenting 1-related pro-tein kinase 2 s (SnRK2s) (Fujii et al. 2009). The activated SnRK2s promote stomatal closure by phosphorylating ion channels such as KAT1, SLAC1 and SLAH3, and induce ABA-responsive gene expression by phosphorylating tran-scription factors such as ABA-responsive element-binding factors (ABFs), and may regulate many other processes through phosphorylating other putative substrates (Umezawa et al. 2013; Wang et al. 2013).
Except PYL13, the remaining 13-members of the PYL family in Arabidopsis are able to activate ABA-responsive gene expression in protoplasts (Fujii et al. 2009). Although PYLs have functional redundancy in mediating ABA-responsive gene expression, some of them have distinct properties. For example, the Arabidopsis pyl8 mutants show a reduced sensitivity to ABA on primary root growth (Antoni et al. 2013), and an increased sensitivity to ABA-mediated inhibition on lateral root elongation (Zhao et al. 2014). AtPYL8 and AtPYL9 directly interact with MYB77, MYB44 and MYB73 transcription factors to enhance auxin signaling (Xing et al. 2016; Zhao et al. 2014), while AtPYL6 interact with MYC2 to regulate JA signaling (Aleman et al. 2016). AtPYL9 plays a major role in ABA-induced leaf senescence (Zhao et al. 2016). Two orthologs OsPYL12 and AtPYL13 constitutively interact with and inhibit sev-eral PP2Cs in an ABA-independent manner (He et al. 2014; Li et al. 2013; Zhao et al. 2013). Furthermore, ABA cannot bind to OsPYL12 (He et al. 2014). OsPYL12 shares simi-larity in the key residues Gln48/Phe76 with the correspond-ing Gln38/Phe71 in AtPYL13 (He et al. 2014), which are crucial for their selective and ABA-independent inhibitory activities on PP2Cs (He et al. 2014; Li et al. 2013; Zhao et al. 2013). Except AtPYL7, AtPYL4–AtPYL10 are puri-fied as monomers in solution and partially inhibit PP2Cs in the absence of ABA (Hao et al. 2011). By contrast, AtPYR1 and AtPYL1-2 are dimers in solution and inhibit PP2Cs only in the presence of ABA. The dimeric PYLs interact with PP2Cs in an ABA-dependent manner, while these mono-meric PYLs bind to PP2Cs mainly in an ABA-independent manner (Hao et al. 2011; Park et al. 2009). Moreover, these PYLs show different binding affinities to ABA, especially between dimeric and monomeric PYLs (Hao et al. 2011).
There are nine clade A PP2Cs in Arabidopsis, which are negative regulators of ABA signaling (Cutler et al. 2010).These PP2Cs include ABA-insensitive 1 (ABI1), ABI2, PP2CA, ABA-hypersensitive germination 1 (AHG1), hyper-sensitive to ABA 1 (HAB1), HAB2, highly ABA induced PP2C gene1 (HAI1), HAI2, and HAI3. PP2Cs bind to SnRK2s and dephosphorylate a serine residue in their activation loop in the absence of ABA, thus inhibiting the kinase activity of the SnRK2s (Soon et al. 2012). In hos-tile environments, the ABA-bound PYLs bind to the cata-lytic site of PP2Cs and inhibit their phosphatase activities,
thus mimicking SnRK2–PP2C interactions and allowing the SnRK2s activated by a combination of auto- and trans-phos-phorylation (Boudsocq et al. 2007; Soon et al. 2012; Xie et al. 2012). These PP2Cs selectively interact with and are inhibited by PYLs in an ABA-dependent manner or ABA-enhanced manner, or ABA-independent manner (Antoni et al. 2012; Hao et al. 2011; He et al. 2014; Li et al. 2013; Tischer et al. 2017; Zhao et al. 2013). ABI1, ABI2 and HAB1 were used as baits in yeast two-hybrid or immunoprecipitation assays for the identification of PYLs (Ma et al. 2009; Nishimura et al. 2010; Santiago et al. 2009). The interaction between PYL and PP2C is required for the transduction of ABA signal-ing. The ABA-bound PYLs do not interact with or inhibit ABI1G180D (Park et al. 2009). The dominant/hypermorphic ABA-insensitive (ABI) mutants abi1-1 and abi2-1 were iden-tified in 1984 (Koornneef et al. 1984). The mutations bear the same amino acid substitution when aligned at the coincidental glycine (ABIG180D, ABI2G168D) (Leung et al. 1994; Leung et al. 1997; Meyer et al. 1994; Rodriguez et al. 1998). There is a functional specialization of different PYLs to interact with and inhibit certain PP2Cs (Antoni et al. 2012; Hao et al. 2011; Tischer et al. 2017). The binding affinity of PP2Cs and PYLs is modulated by the VxGPhiL motif of PP2Cs and a unique tyrosine residue in PYLs (Y169 in PYL9, and F154 in PYL10) (Han et al. 2017; Tischer et al. 2017). The func-tional diversity of ABA receptor PYLs and co-receptor PP2Cs help plant to generate an accurate and sensitive ABA sensing complex. The comprehensive interactions between all PYLs and all PP2Cs need to be performed to explore the underlying mechanism controlling the ABA sensing and signaling.
Among the ten members of SnRK2 protein kinases in Arabidopsis, five of them have been reported to be acti-vated by ABA. SnRK2.2/SRK2D, SnRK2.3/SRK2I and SnRK2.6/SRK2E are major regulators of ABA signaling and are strongly activated by ABA (Fujii and Zhu 2009; Fujita et al. 2009; Furihata et al. 2006; Nakashima et al. 2009). SnRK2.7/SRK2F and SnRK2.8/SRK2C are also acti-vated by ABA and participate in the regulation of expres-sion of drought-induced ABA-responsive genes (Mizoguchi et al. 2010). The ABA-activates SnRK2s promote lots of ABA responses by phosphorylating lots of substrate pro-teins. Phosphoproteomic analysis has been performed in snrk2.2/3/6 triple mutant and pyr1/pyl1/pyl2/pyl4 quadru-ple mutant in Arabidopsis and identified lots of putative substrates for SnRK2 protein kinase (Minkoff et al. 2015; Umezawa et al. 2013; Wang et al. 2013). Although these analyses have been widely performed in Arabidopsis, they need to be performed in maize for more putative substrates for ABA-activated protein kinase including ZmSnRK2s.
Most of the studies of ABA receptor PYLs together with PP2Cs and SnRK2s have been done in Arabidopsis, with a few reports in other species such as rice (He et al. 2014; Kim et al. 2012; Tian et al. 2015), soybean (Bai et al. 2013),

247Plant Molecular Biology (2018) 96:245–263
1 3
maize (Fan et al. 2016; Wang et al. 2014; Wei and Pan 2014), strawberry (Chai et al. 2011) and beechnut (Saavedra et al. 2010). Maize is one of the most important cereal plants in the world. Highly repeated and rearranged sequences make the genome of maize much more complex than those of Arabidopsis and rice (Schnable et al. 2009; Zhou et al. 2009). This might lead to more diverse components for ABA signaling (Hauser et al. 2011). Based on sequence similar-ity analysis, 13 putative maize PYL homologs (ZmPYLs), 16 putative maize clade A PP2C homologs (ZmPP2Cs) and 11 putative maize SnRK2 homologs (ZmSnRK2s) were retrieved from maize genome sequence database (Huai 2008a, b; Wang et al. 2014). Up to now, only ZmPYL3 and ZmPP2C16 were verified as ABA receptor and co-receptor, respectively (Wang et al. 2014). ZmSnRK2.8, a member of clade III of the SnRK2 subfamily, interacts with ZmPP2Cs in an ABA-independent manner (Vilela et al. 2012). Puta-tive substrates of ZmSnRK2s have been identified, such as ZmSNAC1 (NAC domain-containing transcription factors) (Hu et al. 2015; Vilela et al. 2013). In this study, we eluci-dated the redundant and diversified function of the putative ZmPYLs, the clade A ZmPP2Cs, and the ZmSnRK2s in ABA signaling.
Results
PYLs in maize
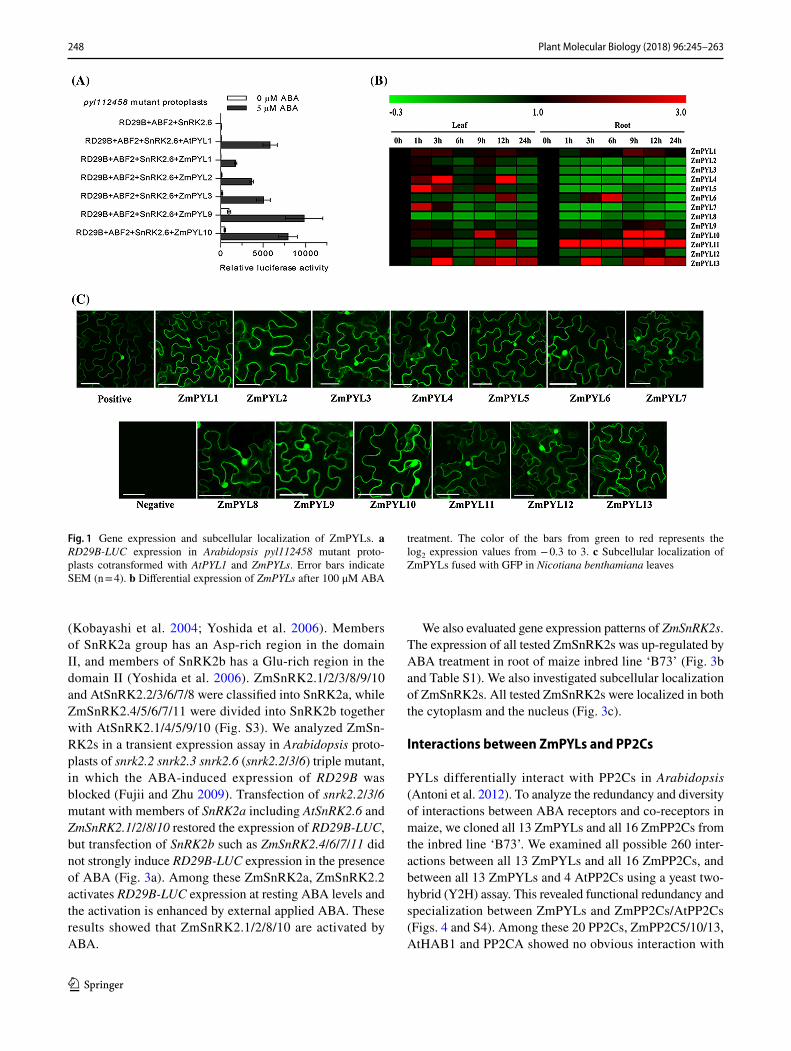
Based on sequence similarity to PYL genes in Arabidop-sis and rice, 13 putative maize PYL homologs (ZmPYLs) were retrieved from maize genome sequence database (Wang et al. 2014). These PYLs can be categorized into three subfamilies (Fig. S1a). Subfamily I is comprised of ZmPYL9-13, AtPYL7-10, and OsPYL7-12. Subfam-ily II contains ZmPYL4-8, AtPYL4-6 and AtPYL11-13, and OsPYL4-6. Subfamily III consists of ZmPYL1-3, which are closely clustered with AtPYR1 and AtPYL1-3, and OsPYL1-3. The ABA-binding pocket of PYLs com-prises of four highly conserved regions named CL1-4 (Yin et al. 2009). ZmPYL13 differs from other PYLs in CL1, CL2 and CL4 regions (Fig. S1b). We evaluated ZmPYLs in a transient expression assay in Arabidopsis protoplasts of pyr1 pyl1 pyl2 pyl4 pyl5 pyl8 (112458) sextuple mutant, in which the ABA-induced expression of RD29B was blocked (Fujii et al. 2009; Gonzalez-Guzman et al. 2012). Trans-fection of 112458 mutant with ABF2 and SnRK2.6 did not induce RD29B-LUC expression in the presence of ABA, but co-transfection with AtPYL1 and ZmPYL1/2/3/9/10 restored the ABA-induced expression of RD29B-LUC (Fig. 1a). Among these ZmPYLs, ZmPYL1-3 activates RD29B-LUC expression in the presence of ABA, while ZmPYL9-10 can enhance RD29B-LUC expression in the absence of external
applied ABA. These results showed that ZmPYLs function as ABA receptors.
We also evaluated gene expression patterns of ZmPYLs in maize inbred line ‘B73’ with ABA treatment using quantitative real-time PCR (qRT-PCR). The expression of ZmPYL2/3/8/9/12 in leaf and ZmPYL2/3/4/5/7/8/9/12 in root was down-regulated after ABA treatment, but the expression of ZmPYL1/4/5/6/7/10/11/13 in leaf and ZmPYL1/6/10/11/13 in root was up-regulated after ABA treatment (Fig. 1b and Table S1). We investigated subcellular localization of ZmPYLs by expressing green fluorescent protein (GFP)-tagged ZmPYL constructs in Nicotiana benthamiana leaves through agroinfiltration. ZmPYL1-12 were localized in both cytoplasm and nucleus, while ZmPYL13 was localized in the cytoplasm but not nucleus (Fig. 1c).
Clade A PP2Cs in maize
Among the 130 members of ZmPP2Cs in maize, 16 of them are categorized into clade A and denominated as ZmPP2C1-16. These clade A PP2Cs can be categorized into several branches (Fig. S2). ZmPP2C1/2/4/5/7/10 are in the same branch with AtABI1/2 and AtHAB1/2. ZmPP2C8 and ZmPP2C13 have higher similarity with AtAHG1. ZmPP2C3/6/9/11/12/14/15/16 are much closer with AtP-P2CA and AtHAI1/2/3. We evaluated ZmPP2Cs in a tran-sient expression assay in Arabidopsis Col-0 wild-type pro-toplasts. Transfection of Col-0 with ABF2 and AtSnRK2.6 induced RD29B-LUC expression in the presence of ABA. Co-transfection with ZmPP2C4/7/16 blocked the ABA-induced expression of RD29B-LUC (Fig. 2a). These results showed that ZmPP2C can inhibit the activity of AtSnRK2.6 and function as negative regulator of ABA signaling.
We evaluated gene expression patterns of ZmPP2Cs in maize inbred line ‘B73’ with ABA treatment. The expres-sion of ZmPP2C2/3/5/6/8/9/10/11/13/14/15/16 in leaf and ZmPP2C2-16 in root was up-regulated by ABA treat-ment, while the expression of ZmPP2C1/4/7/12 in leaf and ZmPP2C1 in root was not obviously changed by ABA treat-ment (Fig. 2b and Table S1). We also investigated subcel-lular localization of ZmPP2Cs. ZmPP2C3/6/8/12/14, which have higher similarity with AtHAI1/2/3, AtPP2CA and AtAHG1, were distributed in the nucleus (Fig. 2c). Similar to ZmPYL13, ZmPP2C1 was localized in the cytoplasm. The other ZmPP2Cs were found in both cytoplasm and nucleus.
SnRK2s in maize
SnRK2s have 11 members in maize and 10 members in Arabidopsis (Fujii et al. 2011; Huai 2008a, b). The domain II in the C-terminal region of the SnRK2 protein is critical for PP2C interaction and ABA responsiveness
248 Plant Molecular Biology (2018) 96:245–263
1 3
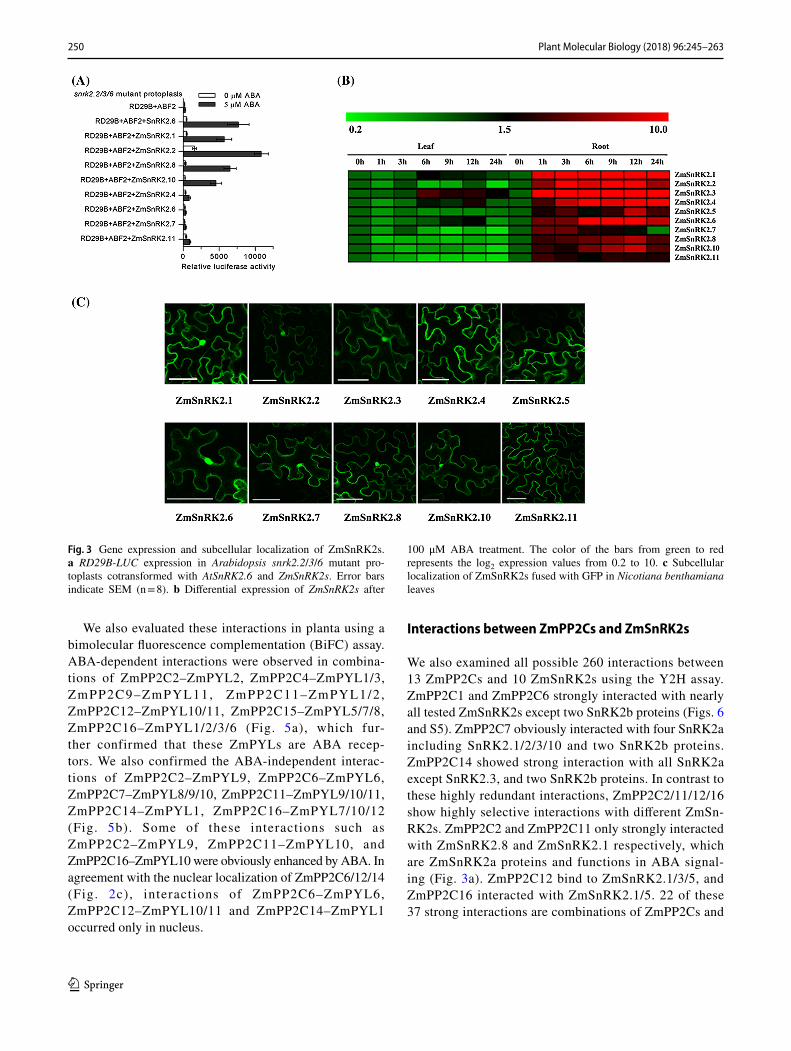
(Kobayashi et al. 2004; Yoshida et al. 2006). Members of SnRK2a group has an Asp-rich region in the domain II, and members of SnRK2b has a Glu-rich region in the domain II (Yoshida et al. 2006). ZmSnRK2.1/2/3/8/9/10 and AtSnRK2.2/3/6/7/8 were classified into SnRK2a, while ZmSnRK2.4/5/6/7/11 were divided into SnRK2b together with AtSnRK2.1/4/5/9/10 (Fig. S3). We analyzed ZmSn-RK2s in a transient expression assay in Arabidopsis proto-plasts of snrk2.2 snrk2.3 snrk2.6 (snrk2.2/3/6) triple mutant, in which the ABA-induced expression of RD29B was blocked (Fujii and Zhu 2009). Transfection of snrk2.2/3/6 mutant with members of SnRK2a including AtSnRK2.6 and ZmSnRK2.1/2/8/10 restored the expression of RD29B-LUC, but transfection of SnRK2b such as ZmSnRK2.4/6/7/11 did not strongly induce RD29B-LUC expression in the presence of ABA (Fig. 3a). Among these ZmSnRK2a, ZmSnRK2.2 activates RD29B-LUC expression at resting ABA levels and the activation is enhanced by external applied ABA. These results showed that ZmSnRK2.1/2/8/10 are activated by ABA.
We also evaluated gene expression patterns of ZmSnRK2s. The expression of all tested ZmSnRK2s was up-regulated by ABA treatment in root of maize inbred line ‘B73’ (Fig. 3b and Table S1). We also investigated subcellular localization of ZmSnRK2s. All tested ZmSnRK2s were localized in both the cytoplasm and the nucleus (Fig. 3c).
Interactions between ZmPYLs and PP2Cs
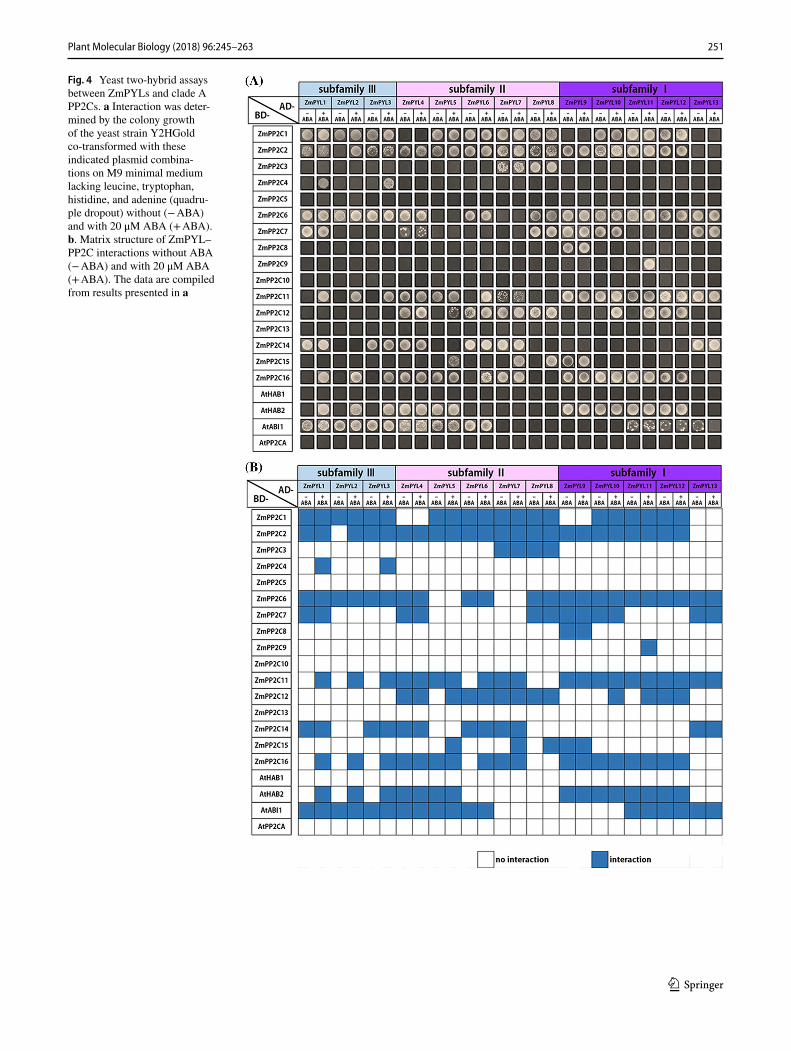
PYLs differentially interact with PP2Cs in Arabidopsis (Antoni et al. 2012). To analyze the redundancy and diversity of interactions between ABA receptors and co-receptors in maize, we cloned all 13 ZmPYLs and all 16 ZmPP2Cs from the inbred line ‘B73’. We examined all possible 260 inter-actions between all 13 ZmPYLs and all 16 ZmPP2Cs, and between all 13 ZmPYLs and 4 AtPP2Cs using a yeast two-hybrid (Y2H) assay. This revealed functional redundancy and specialization between ZmPYLs and ZmPP2Cs/AtPP2Cs (Figs. 4 and S4). Among these 20 PP2Cs, ZmPP2C5/10/13, AtHAB1 and PP2CA showed no obvious interaction with
Fig. 1 Gene expression and subcellular localization of ZmPYLs. a RD29B-LUC expression in Arabidopsis pyl112458 mutant proto-plasts cotransformed with AtPYL1 and ZmPYLs. Error bars indicate SEM (n = 4). b Differential expression of ZmPYLs after 100 μM ABA
treatment. The color of the bars from green to red represents the log2 expression values from − 0.3 to 3. c Subcellular localization of ZmPYLs fused with GFP in Nicotiana benthamiana leaves

249Plant Molecular Biology (2018) 96:245–263
1 3
any of the 13 ZmPYLs even in the presence of ABA accord-ing to the yeast growth on medium deprived of Trp, His, Leu and Ade. Four ZmPP2Cs including ZmPP2C1/3/8/9 were strongly interacted with only one or two ZmPYLs. For example, the ABA-dependent interactions were observed in combinations of ZmPP2C4–ZmPYL1/3 and ZmPP2C9–ZmPYL11, and ABA-independent interactions were exhibited in combinations of ZmPP2C3–ZmPYL7/8 and ZmPP2C8–ZmPYL9. Three ZmPP2Cs interacted with 4–6 different ZmPYLs. In comparison with the ABA-inde-pendent interactions of ZmPP2C7–ZmPYL1/4/8/9/10/13 and ZmPP2C14–ZmPYL1/3/4/6/7/13, ZmPP2C15 inter-acted with ZmPYL5/7/8/9 in an ABA-induced manner. The two closely related PP2Cs ZmPP2C8 and ZmPP2C13 have very similar interaction profile with all ZmPYLs except ZmPYL9.
In contrast to these highly selective interactions, ZmPP2C1/2/6/11/12/16, AtABI1 and AtHAB2 interacted with most of these ZmPYLs (Figs. 4 and S4). Among them, ZmPP2C1 interacted with all ZmPYLs except ZmPYL4/9/13 in an ABA-independent manner. Similar
to the ABA-independent interactions between ZmPP2C1 and ZmPYLs, ZmPP2C6 bind to all ZmPYLs except ZmPYL5/7, and AtABI1 interacted with ZmPYL1-6 and ZmPYL11-13. AtHAB2 and the other four ZmP-P2Cs showed both ABA-dependent and ABA-inde-pendent interactions with different ZmPYLs. Except ZmPYL2, ZmPP2C2 interacted with ZmPYL1-12 in an ABA-independent manner. In contrast with the ABA-dependent interactions of ZmPP2C11–ZmPYL1/2/3/6, ZmPP2C12–ZmPYL5/10/11, ZmPP2C16–ZmPYL1/2/3/6, and AtHAB2–ZmPYL1,2,3, we also noticed ABA-independ-ent interactions of ZmPP2C11–ZmPYL4/5/7/9/10/11/12/13, ZmPP2C12–ZmPYL4/6/7/8/12, ZmPP2C16–Zm PYL4/5/7/9/10/11/12, and HAB2–ZmPYL4/5/9/10/11/12. The two closely related PP2Cs ZmPP2C11 and ZmPP2C16 have very similar interaction profile with all ZmPYLs except ZmPYL13. The ABA-induced interactions between ZmPYLs and ZmPP2Cs further confirmed that ZmPYL1/2/3/5/6/7/8/9/10/11 are ABA receptors and ZmP-P2Cs are ABA coreceptors.
Fig. 2 Gene expression and subcellular localization of ZmPP2Cs. a RD29B-LUC expression in Arabidopsis Col-0 wild-type protoplasts cotransformed with AtSnRK2.6 and ZmPP2Cs. Error bars indicate SEM (n = 6). b Differential expression of ZmPP2Cs after 100 μM
ABA treatment. The color of the bars from green to red represents the log2 expression values from 0.5 to 18. c Subcellular localization of ZmPP2Cs fused with GFP in Nicotiana benthamiana leaves
250 Plant Molecular Biology (2018) 96:245–263
1 3
We also evaluated these interactions in planta using a bimolecular fluorescence complementation (BiFC) assay. ABA-dependent interactions were observed in combina-tions of ZmPP2C2–ZmPYL2, ZmPP2C4–ZmPYL1/3, ZmPP2C9–ZmPYL11, ZmPP2C11–ZmPYL1/2 , ZmPP2C12–ZmPYL10/11, ZmPP2C15–ZmPYL5/7/8, ZmPP2C16–ZmPYL1/2/3/6 (Fig. 5a), which fur-ther confirmed that these ZmPYLs are ABA recep-tors. We also confirmed the ABA-independent interac-tions of ZmPP2C2–ZmPYL9, ZmPP2C6–ZmPYL6, ZmPP2C7–ZmPYL8/9/10, ZmPP2C11–ZmPYL9/10/11, ZmPP2C14–ZmPYL1, ZmPP2C16–ZmPYL7/10/12 (Fig. 5b). Some of these interactions such as ZmPP2C2–ZmPYL9, ZmPP2C11–ZmPYL10, and ZmPP2C16–ZmPYL10 were obviously enhanced by ABA. In agreement with the nuclear localization of ZmPP2C6/12/14 (Fig. 2c), interactions of ZmPP2C6–ZmPYL6, ZmPP2C12–ZmPYL10/11 and ZmPP2C14–ZmPYL1 occurred only in nucleus.
Interactions between ZmPP2Cs and ZmSnRK2s
We also examined all possible 260 interactions between 13 ZmPP2Cs and 10 ZmSnRK2s using the Y2H assay. ZmPP2C1 and ZmPP2C6 strongly interacted with nearly all tested ZmSnRK2s except two SnRK2b proteins (Figs. 6 and S5). ZmPP2C7 obviously interacted with four SnRK2a including SnRK2.1/2/3/10 and two SnRK2b proteins. ZmPP2C14 showed strong interaction with all SnRK2a except SnRK2.3, and two SnRK2b proteins. In contrast to these highly redundant interactions, ZmPP2C2/11/12/16 show highly selective interactions with different ZmSn-RK2s. ZmPP2C2 and ZmPP2C11 only strongly interacted with ZmSnRK2.8 and ZmSnRK2.1 respectively, which are ZmSnRK2a proteins and functions in ABA signal-ing (Fig. 3a). ZmPP2C12 bind to ZmSnRK2.1/3/5, and ZmPP2C16 interacted with ZmSnRK2.1/5. 22 of these 37 strong interactions are combinations of ZmPP2Cs and
Fig. 3 Gene expression and subcellular localization of ZmSnRK2s. a RD29B-LUC expression in Arabidopsis snrk2.2/3/6 mutant pro-toplasts cotransformed with AtSnRK2.6 and ZmSnRK2s. Error bars indicate SEM (n = 8). b Differential expression of ZmSnRK2s after
100 μM ABA treatment. The color of the bars from green to red represents the log2 expression values from 0.2 to 10. c Subcellular localization of ZmSnRK2s fused with GFP in Nicotiana benthamiana leaves
251Plant Molecular Biology (2018) 96:245–263
1 3
Fig. 4 Yeast two-hybrid assays between ZmPYLs and clade A PP2Cs. a Interaction was deter-mined by the colony growth of the yeast strain Y2HGold co-transformed with these indicated plasmid combina-tions on M9 minimal medium lacking leucine, tryptophan, histidine, and adenine (quadru-ple dropout) without (− ABA) and with 20 μM ABA (+ ABA). b. Matrix structure of ZmPYL–PP2C interactions without ABA (− ABA) and with 20 μM ABA (+ ABA). The data are compiled from results presented in a

252 Plant Molecular Biology (2018) 96:245–263
1 3
Fig. 5 Bimolecular fluorescence complementation between ZmPYLs and ZmPP2Cs. In planta interactions was determined by the fluores-cence signal in the epidermal cells of N. benthaminana transiently expressing ZmPYL1-13-YFPn and ZmPP2C1-16-YFPc with or with-
out 100 μM (±) ABA. The white bars represent 50 μm. a Seventeen combinations with ABA-dependent interactions. b Twelve combina-tions with ABA-independent interactions
Fig. 6 Yeast two-hybrid assays between the ZmSnRKs and ZmPP2Cs. a Interaction was determined by the colony growth of the yeast strain Y2HGold co-transformed with these indicated plasmid combi-nations on M9 minimal medium lacking leucine, tryptophan, histidine, and adenine (quadru-ple dropout). b. Matrix structure of ZmSnRK–PP2C interactions. The data are compiled from results presented in a

253Plant Molecular Biology (2018) 96:245–263
1 3
ZmSnRK2a proteins, which participate in the regulation of ABA signaling.
Among these 13 tested ZmPP2Cs, ZmPP2C3/4/8/9/15 showed weak interaction with ZmSnRK2s according to the yeast growth on medium deprived of Trp, His, Leu and Ade. However, these combinations may show interactions in Y2H assay with less stringent conditions.
Candidate substrate proteins phosphorylated by SnRK2s
To analyze the ABA-regulated phosphoproteome in maize, we applied a liquid chromatography-tandem mass spectrom-etry approach. Previous studies have tried to identify sub-strates of ABA-activated SnRK2s using quantitative phos-phoproteomics in snrk2.2/2.3/2.6 and pyr1/pyl1/pyl2/pyl4 mutants grown on plates treated with 50 μM ABA for 5–90 min (Kline et al. 2010; Minkoff et al. 2015; Umezawa et al. 2013; Wang et al. 2013). To analysis substrates of protein kinases directly or indirectly activated by long-term ABA treatment, we treated the three-leaf stage maize seedlings grown in hydroponic culture with 100 μM ABA by soaking root for 6 h. We identified 5028 phosphopep-tides representing 2795 phosphoproteins from the phos-phoproteomic analysis by using label-free quantitation method (Table S2-1). The phosphopeptide abundance of 117 proteins was down-regulated by ABA treatment, while that of 238 proteins was up-regulated by ABA treatment (Table S2-2). These ABA-upregulated phosphoproteins were considered putative substrates of ABA-responsive protein kinase, including SnRK2a proteins, group B3 Raf-like MAP kinase kinase kinases, MPK1/2/3/6/9/12 and CIPK9/27 (Jammes et al. 2009; Lu et al. 2002; Saruhashi et al. 2015; Umezawa et al. 2013; Wang et al. 2013; Zhu 2016). Forty-six of the ABA-upregulated phosphoproteins were homologs of previously identified substrates or putative substrates of SnRK2s, including the well characterized ABA responsive element binding factor 2 (ABF2), the validated substrates such as FBH3 and TOC159 (Table 1) (Umezawa et al. 2013; Wang et al. 2013).
A conserved phosphor motif -RxxS- was found in the SnRK2 substrate proteins (Umezawa et al. 2013; Wang et al. 2013). Therefore, we applied online tool Motif X (http://motif-x.med.harvard.edu) to identify the -RxxS- motif from the 238 up-regulated proteins with the central character as S#, width as 15, and occurrence as 10 (Chou and Schwartz 2011; Schwartz and Gygi 2005). Thirty-four phosphopro-teins were found containing the -RxxS- motif, and predicted to be possible ZmSnRK2 substrates. Eleven of them has been listed in Table 1, and the other 23 and their protein description are listed in Table 2.
We classify the protein descriptions of all the 69 phos-phoproteins (Tables 1, 2) into six categories (Fig. 7). The major function categories are post-transcriptional regulation (32%), protein kinase (22%), transcriptional regulation and epigenetic modification (19%), metabolism (10%) and anion channels (3%).
Discussion
To survive unfavorable abiotic stress conditions, plants have evolved ABA signaling cascades to activate stress responses. The core ABA signaling components have been widely studied in the dicot model plant Arabidopsis, but not in the monocot maize. In this study, we analyzed 13 ZmPYLs, 16 ZmPP2Cs and 11 ZmSnRK2s comprehen-sively. Based on the ABA-dependent interaction between PYLs and PP2Cs (Figs. 4 and S4), and the transient expression assay in pyl112458 sextuple mutant protoplasts (Fig. 1a), we found out at least ten ZmPYLs including ZmPYL1/2/3/5/6/7/8/9/10/11 are functional ABA receptors. According to sequence alignment (fig. S3), we proposed that five ZmSnRK2a proteins including ZmSnRK2.1/2/3/8/10 are ABA-activated SnRK2s. We further verified that at least four ZmSnRK2s including ZmSnRK2.1/2/8/10 are responsive to ABA based on the transient expression assay in snrk2.2/3/6 triple mutant protoplasts (Fig. 3a). Although the core ABA signaling components function redundantly in a wide range of ABA responses (Gonzalez-Guzman et al. 2012), they also generate specific signalosomes controlling the fine-tuning ABA signaling (Antoni et al. 2012; Bhaskara et al. 2012; Fujii et al. 2007; Li et al. 2013; Mustilli et al. 2002; Zhao et al. 2013, 2014). The formation of ABA signalosomes might be determined by the structural properties affecting preference and selection of protein interactions, and also might be related to the spatio-temporal profiles of these com-ponents. We systemically analyzed multiple parameters of ZmPYLs, ZmPP2Cs and ZmSnRK2s, including the in vitro and in vivo protein interactions, subcellular localization, and gene expression. We also investigated putative substrates for ABA-activated protein kinase such as SnRK2s using a large-scale phosphoproteomic analysis. These data are valuable for the future genetic modification of crop plants.
The subcellular localization and gene expression of core signaling components are related to spatio-temporal limitations for the formation of ABA signalosome. Most GFP fused ZmPYLs, ZmPP2Cs and ZmSnRK2s are local-ized to the cytoplasm and nucleus, which is consistent with previous reports of the subcellular localization of PYLs, SnRK2s (Bai et al. 2013; Fujita et al. 2009; Ma et al. 2009; Mizoguchi et al. 2010; Umezawa et al. 2009), suggested that these proteins regulate multiple ABA responses. However, ZmPP2C3/6/8/12/14 and their close

254 Plant Molecular Biology (2018) 96:245–263
1 3
Table 1 Forty-six candidate substrates for ZmSnRK2s
Maize Arabidopsis
Accession number Description GO terms Accession number Description References
XP_008654950.1 Putative DEAD-box ATP-dependent RNA helicase 24-like isoform X2
A AT1G54270 ATP-dependent helicase EIF2A-2
Umezawa et al. (2013), Wang et al. (2013)
XP_008659638.1 Putative DEAD-box ATP-dependent RNA helicase 31-like
A AT1G54270 ATP-dependent helicase EIF2A-2
Umezawa et al. (2013), Wang et al. (2013)
NP_001147574.1 ATP-dependent RNA heli-case DRS1
A AT3G13920 ATP-dependent helicase EIF4A-1
Umezawa et al. (2013), Wang et al. (2013)
XP_008656586.1 DEAD-box ATP-dependent RNA helicase 40-like isoform X2
A AT3G13920 ATP-dependent helicase EIF4A-1
Umezawa et al. (2013), Wang et al. (2013)
XP_008673170.1 Uncharacterized protein LOC100383248
A AT3G13920 ATP-dependent helicase EIF4A-1
Umezawa et al. (2013), Wang et al. (2013)
XP_008652144.1 Glycine-rich RNA-binding protein
A AT3G07810 RNA-binding (RRM/RBD/RNP motifs)
Umezawa et al. (2013), Wang et al. (2013)
XP_008652160.1 Heterogeneous nuclear ribo-nucleo protein 1-like
A AT3G07810 RNA-binding (RRM/RBD/RNP motifs)
Umezawa et al. (2013), Wang et al. (2013)
XP_008673161.1 Putative isoform X1 A AT3G07810 RNA-binding (RRM/RBD/RNP motifs)
Umezawa et al. (2013), Wang et al. (2013)
NP_001131976.1 Splicing factor, arginine/serine-rich 2
A AT3G55460 SC35-like splicing factor 30, SCL30
Umezawa et al. (2013)
NP_001150206.1 SR repressor protein A AT3G55460 SC35-like splicing factor 30, SCL30
Umezawa et al. (2013)
NP_001146195.1 Uncharacterized protein LOC100279765
A AT3G61860 Arginine/serine-rich splicing factor 31
Umezawa et al. (2013)
XP_008659408.1 Putative MEI2-like 2 iso-form X1
A AT3G61860 Arginine/serine-rich splicing factor 31
Umezawa et al. (2013)
NP_001142276.1 26S protease regulatory subunit 8
A AT4G02480 AAA-type ATPase family protein
Wang et al. (2013)
XP_008656079.1 Spastin-like A AT4G02480 AAA-type ATPase family protein
Wang et al. (2013)
XP_008672906.1 Katanin p60 ATPase-con-taining subunit
A AT4G02480 AAA-type ATPase family protein
Wang et al. (2013)
XP_008664485.1 Putative translocase of chlo-roplast 159
A AT4G02510 Translocase of chloroplast 159, TOC159
Wang et al. (2013)
NP_001131184.1 Putative casein kinase B AT1G01140 Calcineurin B-like protein kinase 9, CIPK9
Umezawa et al. (2013), Wang et al. (2013)
NP_001167741.1 Uncharacterized protein LOC100381429
B AT1G01140 Calcineurin B-like protein kinase 9, CIPK9
Umezawa et al. (2013), Wang et al. (2013)
NP_001167855.1 Putative calcium-dependent protein kinase
B AT1G01140 Calcineurin B-like protein kinase 9, CIPK9
Umezawa et al. (2013), Wang et al. (2013)
XP_008665285.1 Uncharacterized protein LOC103643892
B AT1G01140 Calcineurin B-like protein kinase 9, CIPK9
Umezawa et al. (2013), Wang et al. (2013)
XP_008651641.1 Putative protein LOC100382036 isoform X1
B AT2G43790 Mitogen-activated protein kinase 6, MPK6
Umezawa et al. (2013), Wang et al. (2013)
XP_008651696.1 Leucine-rich transmembrane protein kinase1
B AT2G43790 Mitogen-activated protein kinase 6, MPK6
Umezawa et al. (2013), Wang et al. (2013)
XP_008674779.1 Putative protein kinase isoform X1
B AT1G59580 Mitogen-activated protein kinase 2, MPK2
Umezawa et al. (2013)
NP_001132279.1 Uncharacterized LOC100193715
B AT1G69220 Putative serine/threonine kinase SIK1
Umezawa et al. (2013), Wang et al. (2013)
NP_001146379.1 Putative DUF26-domain receptor-like protein kinase
B AT1G69220 Putative serine/threonine kinase SIK1
Umezawa et al. (2013), Wang et al. (2013)

255Plant Molecular Biology (2018) 96:245–263
1 3
relatives AtAHG1 and AtPP2CA (also named as AtAHG3) were distributed in the nucleus (Fig. 2c) (Umezawa et al. 2009), indicated that ZmPP2C3/6/8/12/14 mainly regulate
gene expression in the nucleus. Our quantitative RT-PCR analysis indicated that nearly all ZmPP2Cs were highly induced by ABA treatment (Fig. 2a), which is consistent
The GO terms A/B/C/D/E/F represents post-transcriptional regulation (A), protein kinase (B), transcriptional regulation and epigenetic modifica-tion (C), metabolism (D), Anion channels (E) and others (F) respectively
Table 1 (continued)
Maize Arabidopsis
Accession number Description GO terms Accession number Description References
NP_001147286.1 Serine/threonine-protein kinase NAK
B AT1G69220 Putative serine/threonine kinase SIK1
Umezawa et al. (2013), Wang et al. (2013)
XP_008651654.1 Uncharacterized protein LOC100382164
B AT1G69220 Putative serine/threonine kinase SIK1
Umezawa et al. (2013), Wang et al. (2013)
NP_001143071.1 Uncharacterized protein LOC100275543
B AT5G14720 Protein kinase Wang et al. (2013)
NP_001168730.1 Putative ACT-domain con-taining protein kinase
B AT5G14720 Protein kinase Wang et al. (2013)
XP_008649802.1 Probable leucine-rich repeat receptor-like protein kinase At5g49770
B AT5G14720 Protein kinase Wang et al. (2013)
XP_008659538.1 Serine/threonine-protein kinase BLUS1
B AT5G14720 Protein kinase Wang et al. (2013)
NP_001105291.1 Putative methyl-CpG DNA binding protein
C AT1G15340 Methyl-CpG-binding domain 10, MBD10
Umezawa et al. (2013)
NP_001132507.1 Uncharacterized protein LOC100193967
C AT1G45249 Abscisic acid responsive elements-binding factor 2, ABF2, AREB1
Furihata et al. (2006), Umezawa et al. (2013), Wang et al. (2013)
NP_001151233.1 Helix-loop-helix DNA-binding domain
C AT1G51140 Flowering BHLH 3, FBH3, AKS1
Umezawa et al. (2013), Wang et al. (2013)
XP_008672965.1 Putative isoform helix-loop-helix DNA-binding domain X2
C AT1G51140 Flowering BHLH 3, FBH3, AKS1
Umezawa et al. (2013), Wang et al. (2013)
XP_008658790.1 Putative SAP DNA-binding domain family protein isoform 1
C AT4G39680 SAP domain-containing protein
Wang et al. (2013)
XP_008679859.1 Putative coatomer subunit beta’-2-like
C AT5G25150 TATA-binding-protein asso-ciated factor TAF5
Wang et al. (2013)
NP_001149188.1 Seed specific protein Bn15D14A
D AT2G35880 TPX2 (targeting protein for Xklp2) protein family
Umezawa et al. (2013), Wang et al. (2013)
XP_008643899.1 Putative protein isoform X1 D AT2G35880 TPX2 (targeting protein for Xklp2) protein family
Umezawa et al. (2013), Wang et al. (2013)
NP_001141489.1 Putative protein D AT3G09200 60S acidic ribosomal protein P0
Wang et al. (2013)
NP_001183913.1 Putative protein E AT4G17970 Quickly-activating anion channel 1, QUAC1, ALMT12
Imes et al. (2013)
XP_008644266.1 Chloride channel E AT5G40890 Chloride channel AtCLCa Wege et al. (2014)NP_001170284.1 Uncharacterized protein
LOC100384247F AT1G13030 Atcoilin Wang et al. (2013)
NP_001145910.1 Uncharacterized protein LOC100279429
F AT1G80680 Modifier of snc1,3, MOS3, NUP96, SAR3
Wang et al. (2013)
XP_008669404.1 Uncharacterized protein At4g26450-like isoform X2
F AT4G26450 Uncharacterized protein At4g26450
Wang et al. (2013)
XP_008675107.1 Uncharacterized protein LOC103651267
F AT1G42550 Plastid movement impaired 1, PMI1
Wang et al. (2013)

256 Plant Molecular Biology (2018) 96:245–263
1 3
Table 2 Twenty-three of ABA up-regulated phosphoproteins containing -RXXS- motif
The GO terms A/B/C/D/E/F represents post-transcriptional regulation (A), transcriptional regulation and epigenetic modification (C), metabo-lism (D) and others (F) respectively
Maize Arabidopsis
Accession number Description GO terms Accession number Description
NP_001150768.1 Ubiquitinating enzyme A AT3G17000 Ubiquitin-conjugating enzyme 32, UBC32XP_008662247.1 Uncharacterized protein LOC100384278 A AT1G70320 Ubiquitin-protein ligase 2, UPL2XP_008664038.1 Type II inositol 1,4,5-trisphosphate 5-phos-
phatase FRA3-likeA AT1G65580 Fragile fiber3, FRA3
XP_008667844.1 Putative protein isoform X1 A AT1G16860 Ubiquitin-specific protease family C19-related protein
XP_008673837.1 Nipped-B-like protein B isoform X2 A AT3G23900 RNA recognition motif-containing proteinNP_001169415.1 Uncharacterized protein LOC100383284 A AT1G79380 Ring domain ligase 4, RGLG4NP_001130313.1 Hypothetical protein C AT2G42580 Tetratricopetide-repeat thioredoxin-like 3,
TTL3, BRL2/VH1-interacting TPR con-taining protein, VIT
XP_008656995.1 Ninja-family protein 6-like C AT3G29575 Ninja-family protein, ABI five binding protein 3, AFP3
NP_001105612.1 Eukaryotic translation initiation factor 4E-1 C AT4G18040 Eukaryotic translation initiation factor 4E1, EIF4E1, CUM1, EIF4E
NP_001105687.1 Opaque2 heterodimerizing protein1 (OHP1) C AT5G28770 Basic leucine zipper 63, BZO2H3NP_001150760.1 DNA binding protein C AT3G53710 ARF-GAP domain 6, AGD6XP_008646287.1 Probable pectin methyltransferase QUA2 C AT1G78240 Tumorous shoot development 2, TSD2,
Quasimodo2, QUA2, OSU1XP_008652162.1 Transcriptional regulator ATRX-like C AT1G18950 DDT domain superfamilyXP_008646114.1 Nitrate transporter 1/peptide transporter D AT2G02040 Peptide transporter 2, PTR2, NPF8.3, nitrate
transporter 1, NTR1XP_008659018.1 Sucrose synthase 2 isoform X1 D AT3G43190 Sucrose synthase 4, SUS4XP_008660863.1 Cell division cycle and apoptosis regulator
protein 1-likeD AT2G03150 Short root in salt medium 1, RSA1,
EMB1579, embryo defective 1579XP_008666965.1 Auxin-repressed protein D AT2G33830 Dormancy associated gene 2NP_001145931.1 Hypothetical protein F AT5G56980 Unknown proteinNP_001152904.1 Hypothetical protein F AT5G11970 Protein of unknown functionNP_001183680.1 Uncharacterized protein LOC100502274 F AT4G38360 LAZ1, LAZARUS 1XP_008652643.1 Uncharacterized protein DDB_G0283357-
LikeF AT5G07980 Dentin sialophosphoprotein-related
XP_008676659.1 DENN (AEX-3) domain-containing protein F AT5G35560 DENN (AEX-3) domain-containing proteinXP_008665591.1 Stem-specific protein TSJT1 F AT3G22850 Aluminium induced protein with YGL and
LRDR motifs
Fig. 7 Classification of the 69 putative ZmSnRK2 substrates identified by phosphoproteomic analysis in maize

257Plant Molecular Biology (2018) 96:245–263
1 3
with the reported expression profile of AtPP2Cs (Bhaskara et al. 2012). Conversely, expression of many ZmPYLs was down-regulated by ABA treatment (Fig. 1b), indicated a feedback regulation of ABA signaling. We also noticed that expression of ZmSnRK2s was upregulated by ABA (Fig. 3b), suggested a self-amplification machinery of ABA signal transduction.
The protein interactions determine the preference of signalosome formation. In this study, interactions were detected in 86 ZmPYL–ZmPP2C combinations out of all 208 possible pairs, and were also identified in 18 ZmPYL–AtPP2C combinations out of 52 pairs (Figs. 4 and S4). Moreover, interactions of ZmPP2C–ZmSnRK2 were found in 37 pairs out of 130 combinations (Fig. 6 and S5). These interactions form 311 signalosomes of ABA and osmotic stress signaling in maize. ZmPP2C1/6/7/14 widely interacted with most ZmSnRK2s, and bound to 6–11 dif-ferent ZmPYLs, indicated that ZmPP2C1/6/7/14 are major components of ABA signalosomes in maize. ZmPP2C6 and ZmPP2C14 were localized only in nucleus and inter-acted with ZmPYLs in nucleus (Fig. 5b), suggested they are main negative regulators for activation of ABA respon-sive genes. ZmSnRKs also interacted in a highly selective mode with ZmPP2C2/11/12/16, which widely bound to most of ZmPYLs. 4 out of 13 tested ZmPP2Cs including ZmPP2C3/4/8/9/15 did not show strong interaction with ZmSnRK2s, and only interacted with a few ZmPYLs, indi-cate that these ZmPP2Cs may function in specific cellular events and may participate in maintaining a background ABA signaling. ZmPP2C5/10/13 could not interact with any of the ZmPYLs. There might be more interactions among ZmPYLs, ZmPP2Cs and ZmSnRK2s since we detected interactions based on the yeast growth on medium deprived of Trp, His, Leu and Ade. All the 13 ZmPYL members and five ZmSnRK2a proteins were implicated in ABA sig-nal transduction, but 8 of the 16 clade A members of the ZmPP2C family were not obviously involved. This could be explained by the putative functions of the ZmPP2C mem-bers in many other physiological processes. The interactions between five ZmSnRK2b proteins and ZmPP2Cs implicated that ZmPP2Cs may also affect ABA-independent activation of ZmSnRK2b proteins under osmotic stress conditions.
Current model derived from Arabidopsis work argues that ABA is required for interactions between dimeric PYLs and PP2Cs because the binding of ABA will dissoci-ate the dimeric PYLs, permitting each moiety to interact with a PP2C (Hao et al. 2011). However, all 13 ZmPYLs were involved in the ABA-independent interactions. In maize, ZmPYL1-3 are supposed to be dimers since they are closest homologs for AtPYR1, AtPYL1-2 and OsPYL1-3, which are dimers in solution (Hao et al. 2011; He et al. 2014). However, ZmPYL1-3 interact with ZmPP2Cs and AtPP2Cs either in ABA-dependent and ABA-independent
manners, which mainly depend on PP2Cs. In particular, ZmPYL1-3 interact with ZmPP2C1/2/6/7/14 and AtABI1 in an ABA-independent manner except the combination of ZmPYL2–ZmPP2C2, and bind to ZmPP2C4/11/16 and AtHAB2 in an ABA-dependent manner (Figs. 4 and S4). Moreover, we found that ZmPYL1-3 mainly interact with ZmPP2Cs in an ABA-independent manner. To our surprise, ZmPP2C4 and ZmPP2C9 showed only ABA-dependent interactions with both dimeric and monomeric ZmPYLs. ZmPP2C5, 10, and 13 were not involved in any interactions. Our result suggested that the ABA-independent PYL–PP2C interactions is not only determined by the oligomer status of PYLs, but also the structure of PP2Cs. Consistent with this finding, the dissociation of dimeric SIPYL1 and CsPYL1 cannot be promoted by ABA (Moreno-Alvero et al. 2017). These results indicated that the dissociation of dimeric PYLs can be defined into three types: (i) ABA promotes the dis-sociation of dimeric PYLs (e.g. AtPYLs); (ii) PP2Cs pro-mote the dissociation of dimeric PYLs without ABA (e.g. ZmPYLs); (iii) PP2Cs promote the dissociation of ABA-bounded dimeric PYLs (e.g. CsPYLs and SlPYLs).
The binding between SnRK2.6 and PP2Cs has been char-acterized in genetic and structural level (Belin et al. 2006; Kobayashi et al. 2004; Soon et al. 2012; Yoshida et al. 2006). SnRK2.6 interact with HAB1 both through its kinase domain and its highly acidic C-terminal ABA-box (Soon et al. 2012). Other studies found that ABI1 bind to the C-terminals of the ABA-responsive SnRK2a proteins, but not SnRK2b proteins (Yoshida et al. 2006; Zhang et al. 2016). Our data suggested that PP2Cs also bind to SnRK2b proteins (Figs. 6 and S5). Among SnRK2a and SnRK2b proteins, the kinase domain is conserved. Moreover, the Glu-rich C-terminal of SnRK2b proteins is also highly acidic, which may contribute for the charge-mediated interactions with PP2Cs (Soon et al. 2012). Thus, SnRK2b proteins may interact with PP2Cs in the simi-lar manner as SnRK2a. Our data indicated that PP2Cs also participate in the regulation of kinase activity of SnRK2b. In this case, the ABA-independent activation of SnRK2b should also require the release of SnRK2b from interacting PP2Cs.
Although phosphoproteomic analysis was widely used to identify substrates of ABA-activated protein kinases in dicot Arabidopsis (Chen et al. 2010; Kline et al. 2010; Minkoff et al. 2015; Umezawa et al. 2013; Wang et al. 2013), these study covers neither any monocots nor any crops. We found phosphorylation of 238 proteins in maize was up-regulated by ABA treatment. Among these proteins, 69 of them were putative substrates of SnRK2s. The largest function catego-ries are post-transcriptional regulation (31%), protein kinase (21%), transcriptional regulation and epigenetic modifica-tion (20%), suggest that ABA regulated abiotic responses through multiple strategies, including transcription regu-lation, chromatin modification, pre-mRNA splicing, and

258 Plant Molecular Biology (2018) 96:245–263
1 3
protein phosphorylation modification (Fujii and Zhu 2012; Yoshida et al. 2014; Zhan et al. 2015). Most of these puta-tive ZnSnRK2 substrates are homologs of previously iden-tified putative SnRK2 substrates by large scale phospho-proteomics (Umezawa et al. 2013; Wang et al. 2013). For example, abscisic acid responsive elements-binding factor 2 (NP_001132507.1) is a member of ABF family bZIP class transcription factors, which are important for transcription regulation of many ABA responsive genes (Yoshida et al. 2014, 2010). Reversible phosphorylation catalyzed by pro-tein kinase and phosphatase works as a switch to activate proteins and regulate various cellular processes (Fujii and Zhu 2012; Zhu 2016).
Besides these previously identified putative SnRK2 sub-strates (Umezawa et al. 2013; Wang et al. 2013), we found more putative substrates including the quickly-activating anion channel AtQUAC1 and 23 other proteins (Tables 1, 2). The anion channels SLAC1 together with QUAC1 regu-late stomatal closure (Meyer et al. 2010; Negi et al. 2008; Sasaki et al. 2010; Vahisalu et al. 2008). It is well known that SLAC1 is phosphorylated by SnRK2.6 (Geiger et al. 2009; Lee et al. 2009). However, whether QUAC1 directly phosphorylated by SnRK2.6 is still not clear. We identified an ABA-up-regulated phosphopeptide “_S(ph)FLANS(ph)FLSR_” representing ZmQUAC1 (NP_001183913.1) (Tables 1 and S2-2), which suggested that SnRK2 phospho-rylates ZmQUAC1 at Ser505 and Ser510. These two sites are conserved in maize and Arabidopsis and are in the C-ter-minus cytoplasmic region, which regulates the activity of QUAC1 (Mumm et al. 2013). Several newly identified sub-strates function on the ubiquitination of proteins, including homologs of Ubiquitin-conjugating enzyme 32 (UBC32), Ubiquitin-protein ligase 2 (UPL2), Ubiquitin-specific pro-tease family C19-related protein (AT1G16860), and Ring domain ligase 4 (RGLG4), some of which are also reported regulating salt stress responses and programmed cell death (Cui et al. 2012; Zhang et al. 2015). Some newly identi-fied substrates may participate in the regulation of growth in response to abiotic stresses, including the Type II inositol 1,4,5-trisphosphate 5-phosphatase (XP_008664038.1), dor-mancy/auxin associated family protein (XP_008666965.1), Ninja-family protein 6-like (XP_008656995.1), Opaque2 heterodimerizing protein1 (OHP1), and homologs of Quasi-modo2 (QUA2) and Short root in salt medium 1 (RSA1) (Chen et al. 2013; Fuentes et al. 2010; Guan et al. 2013; Huang and Wu 2007; Zhong et al. 2004). Together with the phosphoproteomic data from dicot Arabidopsis, our work in monocot maize shed new light on ABA signaling network.
Methods
Genomic search and multiple alignment
ZmPYL, ZmPP2C and ZmSnRK2 protein sequences with high similarity to these components in Arabidopsis and rice were gained from Maize GDB database (http://www.maizegdb.org) using BLAST tools. The redundant sequences were eliminated with a custom Perl program, and the con-served domains were identified by using the hidden Markov model from the Pfam database (http://pfam.xfam.org). Mul-tiple alignment among ZmPYLs, AtPYLs and OsPYLs, and among ZmPP2Cs and AtPP2Cs were conducted using ClustalX 2.1 software (http://mac.softpedia.com/get/Math-Scientific/ClustalX.shtml). The phylogenetic tree was con-structed by using software MEGA5.1 (http://mega.software.informer.com/5.1b/).
ABA treatment and RNA extraction
The seeds of the model maize inbred line B73 were steri-lized, germinated in petri dishes, and transplanted into a plastic mesh grid for hydroponic culture at 28 °C under illumination of 200 μmol/m−2 s−1 for a photoperiod of 12 h light/12 h dark. The nutrient solution was replaced every three days. At the three-leaf stage, 100 identical seedlings were divided into three replicates, and treated with ABA by soaking root in nutrient solution with 100 μM (±) ABA (Sigma, USA) (Huai et al. 2008a, b; Ying et al. 2011). After 0 (control), 1, 3, 6, 9, 12 and 24 h, the leaves and roots were sampled, and ground into powder in liquid nitrogen. Total RNA was extracted with RNAiso Plus (TaKaRa, Japan), measured for RNA concentration and integrity, and used for reverse transcription of cDNA with PrimeScript™II1st Strand cDNA synthesis Kit (Takara, Japan).
Quantitative real‑time PCR
Three sets of specific primers were designed by the Primer-BLAST software (http://www.ncbi.nlm.nih.gov/tools/primer-blast), to amplify a 80–200 bp fragment of each members of the ZmPYL gene family, the clade A of the ZmPP2C gene family, and the ZmSnRK2 gene subfam-ily, respectively (Table S3-1). The cDNA samples prepared above were used as templates for qRT-PCR. Referring to Hu et al. (2009) (Hu et al. 2009), the ZmEF1α, ZmTUB, ZmUBQ9 and ZmCYP genes were used as internal con-trols (Table S3-1). Each reaction contained 5 μl of SYBR® Premix Ex TaqTM II (Tli RNaseH Plus) (Takara, Japan), 1 μl cDNA, and 0.4 μl of 10 μmol/L specific primers in a final volume of 10 μl. Three technical replicates were per-formed for each of the cDNA samples. The thermal cycles

259Plant Molecular Biology (2018) 96:245–263
1 3
were conducted in CFX96™ Real-Time System (Bio-Rad, USA) as follows: 95 °C for 30 s; 39 cycles of 95 °C for 5 s, 63 °C or 57.5 °C for 30 s; 95 °C for 10 s. Then, the temperature was increased by 0.5 °C s−1 to 95 °C, so that a melting curve could be calculated and used to differ-entiate between specific and non-specific amplicons. The 2−ΔΔCT method of the CFX Manger™ software version 2.0 (Bio-Rad, USA) was used to normalize the differen-tial gene expression among the multiple internal controls and the target genes(Livak and Schmittgen 2001). The statistical significance among the three biological repli-cates was tested by the method of the Student’s t test with IBM-SPSS software (http://www-01.ibm.com/software/analytics/spss/).
Subcellular localization
Three sets of primers were designed to introduce the appropriate restriction sites (Table S3-2), and used to amplify the coding sequences of the 13 ZmPYLs, the 16 clade A ZmPP2Cs, and the ten ZmSnRK2s without the ter-minal codons from the cDNA samples. The products were inserted into dicotyledonous expression plasmid pC2300-eGFP, and used to transform Agrobacterium tumefaciens strain GV3101. The transformed A. tumefaciens strains harbouring the loaded expression plasmids, together with p19 silencing plasmid, were used to infiltrated into the abaxial air space of 3-week-old Nicotiana benthamiana leaves. The epidermal cell layers of the tobacco leaves near the infiltrated sites were observed for fluorescence after 24 h of the infiltration under confocal laser scanning microscope model A1 (Nikon, Japan).
Yeast two‑hybrid assay
The complete coding sequences of the 13 ZmPYL genes, the 16 clade A genes of the ZmPP2C family, and the ten ZmSnRK2 genes were amplified from the cDNA samples by using the specific primers listed in Table S3-3, and inserted into plasmids pGADT7, pGBDT7, and pGADT7, respectively. With the lithium acetate method follow-ing the protocol provided by Yeastmaker™ Yeast Trans-formation System 2 (Clotech, Japan), the yeast strain Y2HGold was co-transformed by each pair of the loaded prey plasmids pGADT7–ZmPYL1-13 and bait plasmids pGBDT7–ZmPP2C1-5, 7–16, 6, each pair of the loaded prey plasmids pGADT7–ZmPYL1-13 and bait plasmid pGBDT7–ZmPYL13, and each pair of the loaded bait plas-mids pGBDT7–ZmPP2C1-5, 7–16, 6 and prey plasmids pGADT7–ZmSnRK2.1-8, 10, 11, respectively. The possi-ble autoactivation was excluded by the control experiments
co-transformed by respective pairs of the empty prey and the loaded bait plasmids, and of the empty bait and the loaded prey plasmids. After screening on the auxotroph medium SD/-Trp-Leu at 30 °C for 3 days, the transformed monoclones were inoculated liquid auxotroph medium SD/-Trp-Leu, and incubated under the same conditions until OD600 = 0.8–1.0. Then, the cultures were diluted by ten-fold serial to 1:102 with the same medium, inoculated onto the auxotroph medium SD/-Trp/-Leu/-Ade/-His containing 0 (control) and 20 μmol L−1 (±) ABA, and incubated at 30 °C for 5 days.
Bimolecular fluorescence complementation
Two sets of primers were designed to introduce the appro-priate restriction sites (Table S3-4), and used to amplify the coding sequences of the 13 ZmPYL genes and the 16 clade A genes of the ZmPP2C family without the terminal codons from the cDNA samples. The products were recombined into dicotyledonous expression plasmids 35S::pSPYNE fused the N-terminal 155 aa with yellow fluorescence pro-tein (YFP) and 35::pSPYCE fused with the C-terminal 84 aa of YFP, and transformed into Agrobacterium tumefa-ciens strain GV3101. As described by Walter et al. (Wal-ter et al. 2004), each pair of the transformed A. tumefa-ciens strains harbouring the loaded expression plasmids 35::pSPYCE–ZmPYL1-13 and 35::pSPYNE–ZmPP2C1-5, 7–16, 6 was co-infiltrated into the abaxial air space of 3-week-old Nicotiana benthamiana leaves, together with p19 silencing plasmid at OD600 = 0.7:0.7:1.0. The infiltrated leaves were sprayed with 100 μM ABA after 36 h of the infiltration. The epidermal cells of the tobacco leaves near the infiltrated sites were observed for fluorescence after 24 h of the co-infiltration, and 12 h after the ABA spraying under confocal laser scanning microscope model A1 (Nikon, Japan).
Protoplast experiment in vivo
The Arabidopsis thaliana Col-0 wild type, pyl112458 mutant and snrk2.2/3/6 mutant were used as plant materials for protoplast preparation. The CDS of ZmPYL1/2/3/9/10, ZmPP2C7/16 and ZmSnRK2.1/2/4/6/7/8/10/11 were con-structed into pHBT95, primers used listed in Supplemen-tary Table S3-5. pHBT95–AtPYL1/AtSnRK2.6 were used as control vectors. All these constructs were confirmed by sequencing and purified by QIAGEN Plasmid Maxi Kit.
The protoplasts for transient expression were performed as described (Zhao et al. 2013). Co-transfected AtSnRK2.6, ABF2 with ZmPYL1/2/3/9/10 respectively to reconstitute the ABA signaling pathway in pyl112458 mutant protoplast. RD29B-LUC was used as the ABA-responsive reporter and ZmUBQ-GUS was used as the internal control. After

260 Plant Molecular Biology (2018) 96:245–263
1 3
transfection, the protoplasts were incubated in the absence/presence of 5 μM ABA for 4.5 h under light. The ABF2 and AtSnRK2.6/ZmSnRK2.1/2/4/6/7/8/10/11 respectively were transfected into snrk2.2/3/6 mutant protoplasts as above. Using the same method, AtSnRK2.6 and ABF2 were co-transfected with ZmPP2C7/16 to reconstitute the ABA sign-aling pathway in Col-0.
Phosphoproteomics analysis
Total protein was extracted from the three independent biological replicates of the cultured seedlings treated with ABA by soaking root in nutrient solution with 100 μM (±) ABA for 6 h and the negative control by using metha-nol–chloroform precipitation (Huai et al. 2008a, b; Ying et al. 2011), and digested by using trypsin (Sep-PaktC18, Waters). Phosphopeptides were enriched by using TiO2 par-ticles (GL Sciences Inc.) as described by Sugiyama et al. (2007). The liquid chromatography-tandem mass spec-trometry was conducted in a nanoACQUITY UPLC system (Waters) coupled with a hybrid Quadrupole-Orbitrap mass spectrometer (Thermo Q Exactive Orbitrap). The mass spec-trometry raw files were searched by using MaxQuant 1.4.1.2 search engine for protein identification and quantification. Quantification was performed through label-free quantifica-tion, using intensity-based absolute quantification (iBAQ), in which protein content is normalized to the total number of potential peptides (Mann K, Mann M). The proteome of the calcified layer organic matrix of turkey (Meleagris gal-lopavo) eggshell.
Acknowledgements We thank the technical support from Key Labo-ratory of Biology and Genetic Improvement of Maize in Southwest Region and the operation of Confocal Fluorescence Microscope pro-vided form Wenming Wang lab, pSPYNE and pSPYCE vector provided from Shigui Li lab and technical support of bimolecular fluorescence complementation directed by Xuewei Chen lab of Rice Research Institute.
Author contributions Y-GW, F-LF, H-QY, TH, YYZ preformed the experiments, Y-GW and YT preformed the phosphoproteomics data analysis; YZ and J-KZ provided technical support; Y-GW and YZ wrote the manuscript; YZ and W-CL directed the research.
Funding This project was supported by the National Nature Science Foundation of China (31071433).
References
Aleman F et al (2016) An ABA-increased interaction of the PYL6 ABA receptor with MYC2 transcription factor: a putative link of ABA and JA. Signal Sci Rep 6:28941. https://doi.org/10.1038/srep28941
Antoni R, Gonzalez-Guzman M, Rodriguez L, Rodrigues A, Piz-zio GA, Rodriguez PL (2012) Selective inhibition of clade A phosphatases type 2C by PYR/PYL/RCAR abscisic acid
receptors. Plant Physiol 158:970–980. https://doi.org/10.1104/pp.111.188623
Antoni R et al (2013) PYRABACTIN RESISTANCE1-LIKE8 plays an important role for the regulation of abscisic acid signaling in root. Plant Physiol 161:931–941. https://doi.org/10.1104/pp.112.208678
Bai G et al (2013) Interactions between soybean ABA receptors and type 2C protein phosphatases. Plant Mol Biol 83:651–664. https://doi.org/10.1007/s11103-013-0114-4
Belin C et al (2006) Identification of features regulating OST1 kinase activity and OST1 function in guard cells. Plant Physiol 141:1316–1327. https://doi.org/10.1104/pp.106.079327
Bhaskara GB, Nguyen TT, Verslues PE (2012) Unique drought resistance functions of the highly ABA-induced clade A protein phosphatase 2. Cs Plant physiology 160:379–395. https://doi.org/10.1104/pp.112.202408
Boudsocq M, Droillard M-J, Barbier-Brygoo H, Laurière C (2007) Dif-ferent phosphorylation mechanisms are involved in the activation of sucrose non-fermenting 1 related protein kinases 2 by osmotic stresses and abscisic acid. Plant Mol Biol 63(4):491–503
Chai YM, Jia HF, Li CL, Dong QH, Shen YY (2011) FaPYR1 is involved in strawberry fruit ripening. J Exp Bot 62:5079–5089. https://doi.org/10.1093/jxb/err207
Chen Y, Hoehenwarter W, Weckwerth W (2010) Comparative analy-sis of phytohormone-responsive phosphoproteins in Arabidop-sis thaliana using TiO2-phosphopeptide enrichment and mass accuracy precursor alignment. Plant J 63:1–17. https://doi.org/10.1111/j.1365-313X.2010.04218.x
Chen Z, Huang J, Muttucumaru N, Powers SJ, Halford NG (2013) Expression analysis of abscisic acid (ABA) and metabolic sig-nalling factors in developing endosperm and embryo of barley. J Cereal Sci 58:255–262. https://doi.org/10.1016/j.jcs.2013.06.009
Chou MF, Schwartz D (2011) Biological sequence motif dis-covery using motif-x. Curr Protoc Bioinform. https://doi.org/10.1002/0471250953.bi1315s35
Cui F et al (2012) Arabidopsis ubiquitin conjugase UBC32 is an ERAD component that functions in brassinosteroid-mediated salt stress tolerance. Plant Cell 24:233–244. https://doi.org/10.1105/tpc.111.093062
Cutler SR, Rodriguez PL, Finkelstein RR, Abrams SR (2010) Abscisic acid: emergence of a core signaling network Annu. Rev Plant Biol 61:651–679. https://doi.org/10.1146/annurev-arplant-042809-112122
Fan W, Zhao M, Li S, Bai X, Li J, Meng H, Mu Z (2016) Contrast-ing transcriptional responses of PYR1/PYL/RCAR ABA recep-tors to ABA or dehydration stress between maize seedling leaves and roots. BMC Plant Biol 16:99. https://doi.org/10.1186/s12870-016-0764-x
Fuentes S, Pires N, Ostergaard L (2010) A clade in the QUASIMODO2 family evolved with vascular plants and supports a role for cell wall composition in adaptation to environmental changes. Plant Mol Biol 73:605–615. https://doi.org/10.1007/s11103-010-9640-5
Fujii H, Zhu JK (2009) Arabidopsis mutant deficient in 3 abscisic acid-activated protein kinases reveals critical roles in growth, reproduc-tion, and stress. Proc Natl Acad Sci USA 106:8380–8385. https://doi.org/10.1073/pnas.0903144106
Fujii H, Zhu JK (2012) Osmotic stress signaling via protein kinases. Cell Mol Life Sci 69:3165–3173. https://doi.org/10.1007/s00018-012-1087-1
Fujii H, Verslues PE, Zhu JK (2007) Identification of two protein kinases required for abscisic acid regulation of seed germina-tion, root growth, and gene expression in. Arabidopsis Plant Cell 19:485–494. https://doi.org/10.1105/tpc.106.048538
Fujii H et al (2009) In vitro reconstitution of an abscisic acid sig-nalling pathway. Nature 462:660–664. https://doi.org/10.1038/nature08599

261Plant Molecular Biology (2018) 96:245–263
1 3
Fujii H, Verslues PE, Zhu JK (2011) Arabidopsis decuple mutant reveals the importance of SnRK2 kinases in osmotic stress responses in vivo. Proc Natl Acad Sci USA 108:1717–1722. https://doi.org/10.1073/pnas.1018367108
Fujita Y et al (2009) Three SnRK2 protein kinases are the main positive regulators of abscisic acid signaling in response to water stress in Arabidopsis. Plant Cell Physiol 50:2123–2132. https://doi.org/10.1093/pcp/pcp147
Furihata T, Maruyama K, Fujita Y, Umezawa T, Yoshida R, Shino-zaki K, Yamaguchi-Shinozaki K (2006) Abscisic acid-dependent multisite phosphorylation regulates the activity of a transcrip-tion activator AREB1. Proc Natl Acad Sci USA 103:1988–1993. https://doi.org/10.1073/pnas.0505667103
Geiger D et al (2009) Activity of guard cell anion channel SLAC1 is controlled by drought-stress signaling kinase-phosphatase pair. Proc Natl Acad Sci USA 106:21425–21430. https://doi.org/10.1073/pnas.0912021106
Gonzalez-Guzman M et al (2012) Arabidopsis PYR/PYL/RCAR receptors play a major role in quantitative regulation of stoma-tal aperture and transcriptional response to abscisic. Acid Plant Cell 24:2483–2496. https://doi.org/10.1105/tpc.112.098574
Guan Q, Wu J, Yue X, Zhang Y, Zhu J (2013) A nuclear calcium-sensing pathway is critical for gene regulation and salt stress tolerance in. Arabidopsis PLoS Genet 9:e1003755. https://doi.org/10.1371/journal.pgen.1003755
Han S et al (2017) Modulation of ABA signaling by altering VxG-PhiL motif of PP2Cs in Oryza sativa. Mol Plant 10:1190–1205. https://doi.org/10.1016/j.molp.2017.08.003
Hao Q et al (2011) The molecular basis of ABA-independent inhibi-tion of PP2Cs by a subclass of PYL proteins. Mol Cell 42:662–672. https://doi.org/10.1016/j.molcel.2011.05.011
Hauser F, Waadt R, Schroeder JI (2011) Evolution of abscisic acid synthesis and signaling mechanisms. Curr Biol 21:R346–R355. https://doi.org/10.1016/j.cub.2011.03.015
He Y, Hao Q, Li W, Yan C, Yan N, Yin P (2014) Identification and characterization of ABA receptors in Oryza sativa. PLoS ONE 9:e95246. https://doi.org/10.1371/journal.pone.0095246
Hu R, Fan C, Li H, Zhang Q, Fu YF (2009) Evaluation of putative reference genes for gene expression normalization in soybean by quantitative real-time RT-PCR. BMC Mol Biol 10:93. https://doi.org/10.1186/1471-2199-10-93
Hu X et al (2015) Quantitative iTRAQ-based proteomic analysis of phosphoproteins and ABA-regulated phosphoproteins in maize leaves under osmotic stress. Sci Rep 5:15626. https://doi.org/10.1038/srep15626
Huai J et al (2008a) Cloning and characterization of the SnRK2 gene family from Zea mays. Plant cell Rep 27:1861–1868. https://doi.org/10.1007/s00299-008-0608-8
Huai J, He MW,J, Zheng J, Dong Z, Lv H, Zhao J, Wang G (2008b) Cloning and characterization of the SnRK2 gene family from maize. Plant Cell Rep 27:1861–1868
Huang MD, Wu WL (2007) Overexpression of TMAC2, a novel negative regulator of abscisic acid and salinity responses, has pleiotropic effects in Arabidopsis thaliana. Plant Mol Biol 63:557–569. https://doi.org/10.1007/s11103-006-9109-8
Imes D, Mumm P, Böhm J, Al-Rasheid KAS, Marten I, Geiger D, Hedrich R (2013) Open stomata 1 (OST1) kinase controls R-type anion channel QUAC1 in Arabidopsis guard cells. Plant J 74(3):372–382
Jammes F et al (2009) MAP kinases MPK9 and MPK12 are prefer-entially expressed in guard cells and positively regulate ROS-mediated ABA signaling. Proc Natl Acad Sci USA 106:20520–20525. https://doi.org/10.1073/pnas.0907205106
Kim H et al (2012) A rice orthologue of the ABA receptor, OsPYL/RCAR5, is a positive regulator of the ABA signal transduction
pathway in seed germination and early seedling growth. J Exp Bot 63:1013–1024. https://doi.org/10.1093/jxb/err338
Kline KG, Barrett-Wilt GA, Sussman MR (2010) In planta changes in protein phosphorylation induced by the plant hormone absci-sic acid. Proc Natl Acad Sci USA 107:15986–15991. https://doi.org/10.1073/pnas.1007879107
Kobayashi Y, Yamamoto S, Minami H, Kagaya Y, Hattori T (2004) Differential activation of the rice sucrose nonfermenting1-related protein kinase2 family by hyperosmotic stress and abscisic acid. Plant Cell 16:1163–1177. https://doi.org/10.1105/tpc.019943
Koornneef M, Reuling G, Karssen CM (1984) The isolation and characterization of abscisic acid-insensitive mutants of Arabidopsis thaliana. Physiol Plant 61:377–383. https://doi.org/10.1111/j.1399-3054.1984.tb06343.x
Lee SC, Lan W, Buchanan BB, Luan S (2009) A protein kinase-phos-phatase pair interacts with an ion channel to regulate ABA sign-aling in plant guard cells. Proc Natl Acad Sci USA 106:21419–21424. https://doi.org/10.1073/pnas.0910601106
Leung J, Bouvier-Durand M, Morris P, Guerrier D, Chefdor F, Giraudat J (1994) Arabidopsis ABA response gene ABI1: fea-tures of a calcium-modulated protein phosphatase. Sci 264 (5164):1448–1452
Leung J, Merlot S, Giraudat J (1997) The Arabidopsis ABSCISIC ACID-INSENSITIVE2 (ABI2) and ABI1 genes encode homolo-gous protein phosphatases 2C involved in abscisic acid signal transduction. Plant Cell 9:759–771. https://doi.org/10.1105/tpc.9.5.759
Li W et al (2013) Molecular basis for the selective and ABA-inde-pendent inhibition of PP2CA by PYL 13. Cell Res 23:1369–1379. https://doi.org/10.1038/cr.2013.143
Livak KJ, Schmittgen TD (2001) Analysis of relative gene expres-sion data using real-time quantitative PCR and the 2(-Delta Delta C(T). Method Methods 25:402–408. https://doi.org/10.1006/meth.2001.1262
Lu C, Han MH, Guevara-Garcia A, Fedoroff NV (2002) Mitogen-activated protein kinase signaling in postgermination arrest of development by abscisic acid. Proc Natl Acad Sci USA 99:15812–15817. https://doi.org/10.1073/pnas.242607499
Ma Y, Szostkiewicz I, Korte A, Moes D, Yang Y, Christmann A, Grill E (2009) Regulators of PP2C phosphatase activity func-tion as abscisic acid sensors. Science 324:1064–1068. https://doi.org/10.1126/science.1172408
Meyer K, Leube M, Grill E (1994) A protein phosphatase 2C involved in ABA signal transduction in Arabidopsis thaliana. Sci 264(5164):1452–1455
Meyer S et al (2010) AtALMT12 represents an R-type anion channel required for stomatal movement in Arabidopsis guard cells. Plant J 63:1054–1062. https://doi.org/10.1111/j.1365-313X.2010.04302.x
Minkoff BB, Stecker KE, Sussman MR (2015) Rapid phosphoprot-eomic effects of ABA on wildtype and ABA receptor-deficient A. thaliana mutants. Mol Cell Proteom. https://doi.org/10.1074/mcp.M114.043307
Mizoguchi M et al (2010) Two closely related subclass II SnRK2 pro-tein kinases cooperatively regulate drought-inducible gene expres-sion. Plant Cell Physiol 51:842–847. https://doi.org/10.1093/pcp/pcq041
Moreno-Alvero M et al (2017) Structure of ligand-bound intermedi-ates of crop ABA receptors highlights PP2C as necessary ABA Co-receptor. Mol Plant 10:1250–1253. https://doi.org/10.1016/j.molp.2017.07.004
Mumm P, Imes D, Martinoia E, Al-Rasheid KA, Geiger D, Marten I, Hedrich R (2013) C-terminus-mediated voltage gating of Arabi-dopsis guard cell anion channel QUAC1. Mol Plant 6:1550–1563. https://doi.org/10.1093/mp/sst008

262 Plant Molecular Biology (2018) 96:245–263
1 3
Mustilli AC, Merlot S, Vavasseur A, Fenzi F, Giraudat J (2002) Arabi-dopsis OST1 protein kinase mediates the regulation of stomatal aperture by abscisic acid and acts upstream of reactive oxygen species production. Plant Cell 14:3089–3099
Nakashima K et al (2009) Three Arabidopsis SnRK2 protein kinases, SRK2D/SnRK2.2, SRK2E/SnRK2.6/OST1 and SRK2I/SnRK2.3, involved in ABA signaling are essential for the control of seed development and dormancy. Plant Cell Physiol 50:1345–1363. https://doi.org/10.1093/pcp/pcp083
Nambara E, Marion-Poll A (2005) Abscisic acid biosynthesis and catabolism. Annu Rev Plant Biol 56:165–185. https://doi.org/10.1146/annurev.arplant.56.032604.144046
Negi J et al (2008) CO2 regulator SLAC1 and its homologues are essen-tial for anion homeostasis in plant cells. Nature 452:483–486. https://doi.org/10.1038/nature06720
Nishimura N et al (2010) PYR/PYL/RCAR family members are major in-vivo ABI1 protein phosphatase 2C-interacting proteins in Arabidopsis. Plant J 61:290–299. https://doi.org/10.1111/j.1365-313X.2009.04054.x
Ohkuma K, Lyon JL, Addicott FT, Smith OE (1963) Abscisin II, an abscission-accelerating substance from young cotton fruit. Science 142:1592–1593. https://doi.org/10.1126/science.142.3599.1592
Park SY et al (2009) Abscisic acid inhibits type 2C protein phos-phatases via the PYR/PYL family of START proteins. Science 324:1068–1071. https://doi.org/10.1126/science.1173041
Rodriguez PL, Benning G, Grill E (1998) ABI2, a second protein phosphatase 2C involved in abscisic acid signal transduction in Arabidopsis. FEBS Lett 421(3):185–190
Saavedra X, Modrego A, Rodriguez D, Gonzalez-Garcia MP, Sanz L, Nicolas G, Lorenzo O (2010) The nuclear interactor PYL8/RCAR3 of Fagus sylvatica FsPP2C1 is a positive regulator of abscisic acid signaling in seeds and stress. Plant Physiol 152:133–150. https://doi.org/10.1104/pp.109.146381
Santiago J et al (2009) Modulation of drought resistance by the abscisic acid receptor PYL5 through inhibition of clade A PP2Cs. Plant J 60:575–588. https://doi.org/10.1111/j.1365-313X.2009.03981.x
Saruhashi M et al (2015) Plant Raf-like kinase integrates abscisic acid and hyperosmotic stress signaling upstream of SNF1-related pro-tein kinase2. Proc Natl Acad Sci USA 112:E6388–E6396. https://doi.org/10.1073/pnas.1511238112
Sasaki T et al (2010) Closing plant stomata requires a homolog of an aluminum-activated malate transporter. Plant Cell Physiol 51:354–365. https://doi.org/10.1093/pcp/pcq016
Schnable PS et al (2009) The B73 maize genome: complexity, diversity and dynamics. Science 326:1112–1115. https://doi.org/10.1126/science.1178534
Schwartz D, Gygi SP (2005) An iterative statistical approach to the identification of protein phosphorylation motifs from large-scale data sets. Nat Biotechnol 23:1391–1398. https://doi.org/10.1038/nbt1146
Soon FF et al (2012) Molecular mimicry regulates ABA signaling by SnRK2 kinases and PP2C phosphatases. Science 335:85–88. https://doi.org/10.1126/science.1215106
Sugiyama N, Masuda T, Shinoda K, Nakamura A, Tomita M, Ishihama Y (2007) Phosphopeptide enrichment by aliphatic hydroxy acid-modified metal oxide chromatography for nano-LC-MS/MS in proteomics applications. Mol Cell Proteom 6:1103–1109. https://doi.org/10.1074/mcp.T600060-MCP200
Tian X et al (2015) Characterization and functional analysis of pyra-bactin resistance-like abscisic acid receptor family in rice. Rice 8:28. https://doi.org/10.1186/s12284-015-0061-6
Tischer SV, Wunschel C, Papacek M, Kleigrewe K, Hofmann T, Christmann A, Grill E (2017) Combinatorial interaction net-work of abscisic acid receptors and coreceptors from Arabidop-sis thaliana. Proc Natl Acad Sci USA. https://doi.org/10.1073/pnas.1706593114
Umezawa T et al (2009) Type 2C protein phosphatases directly regu-late abscisic acid-activated protein kinases in Arabidopsis. Proc Natl Acad Sci USA 106:17588–17593. https://doi.org/10.1073/pnas.0907095106
Umezawa T, Sugiyama N, Takahashi F, Anderson JC, Ishihama Y, Peck SC, Shinozaki K (2013) Genetics and phosphoproteomics reveal a protein phosphorylation network in the abscisic acid signaling pathway in Arabidopsis thaliana. Sci Signal 6:rs8. https://doi.org/10.1126/scisignal.2003509
Vahisalu T et al (2008) SLAC1 is required for plant guard cell S-type anion channel function in stomatal signalling. Nature 452:487–491. https://doi.org/10.1038/nature06608
Vilela B, Moreno A, Capellades M, Pagès M, Lumbreras V (2012) ZmSnRK2.8 responds to ABA through the SnRK2-PP2C complex Maydica 57:11–18
Vilela B, Moreno-Cortes A, Rabissi A, Leung J, Pages M, Lumbreras V (2013) The maize OST1 kinase homolog phosphorylates and regulates the maize SNAC1-type transcription factor. PLoS ONE 8:e58105. https://doi.org/10.1371/journal.pone.0058105
Walter M et al (2004) Visualization of protein interactions in living plant cells using bimolecular fluorescence complementation. Plant J 40:428–438. https://doi.org/10.1111/j.1365-313X.2004.02219.x
Wang P et al (2013) Quantitative phosphoproteomics identifies SnRK2 protein kinase substrates and reveals the effectors of abscisic acid action. Proc Natl Acad Sci USA 110:11205–11210. https://doi.org/10.1073/pnas.1308974110
Wang Y-G, Yu H-Q, Zhang Y-Y, Lai C-X, She Y-H, Li W-C, Fu F-L (2014) Interaction between abscisic acid receptor PYL3 and protein phosphatase type 2C in response to ABA signal-ing in maize. Gene 549:179–185 doi. https://doi.org/10.1016/j.gene.2014.08.001
Wei K, Pan S (2014) Maize protein phosphatase gene family: identifica-tion and molecular characterization. BMC Genom 15:773. https://doi.org/10.1186/1471-2164-15-773
Wege S, De Angeli A, Droillard M-J, Kroniewicz L, Merlot S, Cornu D, Gambale F, Martinoia E, Barbier-Brygoo H, Thomine S, Leon-hardt N, Filleur S (2014) Phosphorylation of the vacuolar anion exchanger AtCLCa is required for the stomatal response to absci-sic acid. Sci Signal 7(333):ra65
Xie T et al (2012) Molecular mechanism for inhibition of a critical component in the Arabidopsis thaliana abscisic acid signal trans-duction pathways, SnRK2.6, by protein phosphatase ABI1. J Biol Chem 287:794–802. https://doi.org/10.1074/jbc.M111.313106
Xing L, Zhao Y, Gao J, Xiang C, Zhu JK (2016) The ABA receptor PYL9 together with PYL8 plays an important role in regulating lateral root growth. Sci Rep 6:27177. https://doi.org/10.1038/srep27177
Yin P et al (2009) Structural insights into the mechanism of abscisic acid signaling by PYL proteins. Nat Struct Mol Biol 16:1230–1236. https://doi.org/10.1038/nsmb.1730
Ying S et al (2011) Cloning and characterization of a maize SnRK2 protein kinase gene confers enhanced salt tolerance in trans-genic Arabidopsis. Plant cell Rep 30:1683–1699. https://doi.org/10.1007/s00299-011-1077-z
Yoshida R, Umezawa T, Mizoguchi T, Takahashi S, Takahashi F, Shinozaki K (2006) The regulatory domain of SRK2E/OST1/SnRK2.6 interacts with ABI1 and integrates abscisic acid (ABA) and osmotic stress signals controlling stomatal closure in Arabi-dopsis. J Biol Chem 281:5310–5318. https://doi.org/10.1074/jbc.M509820200
Yoshida T et al (2010) AREB1, AREB2, and ABF3 are master tran-scription factors that cooperatively regulate ABRE-depend-ent ABA signaling involved in drought stress tolerance and require ABA for full activation. Plant J 61:672–685. https://doi.org/10.1111/j.1365-313X.2009.04092.x

263Plant Molecular Biology (2018) 96:245–263
1 3
Yoshida T, Fujita Y, Maruyama K, Mogami J, Todaka D, Shinozaki K, Yamaguchi-Shinozaki K (2014) Four Arabidopsis AREB/ABF transcription factors function predominantly in gene expres-sion downstream of SnRK2 kinases in abscisic acid signalling in response to osmotic stress. Plant Cell Environ. https://doi.org/10.1111/pce.12351
Zhan X et al (2015) An Arabidopsis PWI and RRM motif-containing protein is critical for pre-mRNA splicing and ABA responses. Nat Commun 6:8139. https://doi.org/10.1038/ncomms9139
Zhang X et al (2015) Hijacking of the jasmonate pathway by the myco-toxin fumonisin B1 (FB1) to initiate programmed cell death in Arabidopsis is modulated by RGLG3 and RGLG4. J Exp Bot 66:2709–2721. https://doi.org/10.1093/jxb/erv068
Zhang H, Li W, Mao X, Jing R, Jia H (2016) Differential activation of the wheat SnRK2 family by abiotic stresses. Front Plant Sci 7:420. https://doi.org/10.3389/fpls.2016.00420
Zhao Y et al (2013) The unique mode of action of a divergent member of the ABA-receptor protein family in ABA and stress signaling. Cell Res 23:1380–1395. https://doi.org/10.1038/cr.2013.149
Zhao Y et al (2014) The ABA receptor PYL8 promotes lateral root growth by enhancing MYB77-dependent transcription of
auxin-responsive. genes Sci Signal 7:ra53. https://doi.org/10.1126/scisignal.2005051
Zhao Y et al (2016) ABA receptor PYL9 promotes drought resistance and leaf senescence. Proc Natl Acad Sci USA 113:1949–1954. https://doi.org/10.1073/pnas.1522840113
Zhong R, Burk DH, Morrison WH 3rd, Ye ZH (2004) FRAG-ILE FIBER3, an Arabidopsis gene encoding a type II inositol polyphosphate 5-phosphatase, is required for secondary wall syn-thesis and actin organization in fiber cells. Plant Cell 16:3242–3259. https://doi.org/10.1105/tpc.104.027466
Zhou S et al. (2009) A single molecule scaffold for the maize genome. PLoS Genet 5:e1000711 https://doi.org/10.1371/journal.pgen.1000711
Zhu JK (2002) Salt and drought stress signal transduction in plants. Annu Rev Plant Biol 53:247–273. https://doi.org/10.1146/annurev.arplant.53.091401.143329
Zhu JK (2016) Abiotic stress signaling and responses in plants. Cell 167:313–324. https://doi.org/10.1016/j.cell.2016.08.029





























