Embed Size (px)
DESCRIPTION
inmunologia
Citation preview

A
cr©
K
1
ybdmcctHmadomoq1crn[eig
1d
Seminars in Immunology 19 (2007) 94–105
Review
What immunologists should know about bacterial communitiesof the human bowel
Gerald W. Tannock ∗Department of Microbiology and Immunology, University of Otago, P.O. Box 56, 720 Cumberland Street, Dunedin, New Zealand
bstract
The human bowel is home to a bacterial community of much complexity. This article summarizes current bacteriological knowledge of theommunity and highlights topics of potential interest to innovative immunologists. The role of the bacterial community in the development andegulation of the immune system of neonates seems likely to be a particularly important area of future research.
2006 Elsevier Ltd. All rights reserved.
gF0ttmabtbsbacgiaTaig(b
eywords: Bacteria; Bowel; Microbiota; Gut; Microbial ecology
. The company that you keep
Homo sapiens, it seems, first walked the Earth about 130,000ears ago. Our distant ancestors presumably carried within theirowels a large and complex community of microbes just aso their primate relations, modern descendants and other ani-al species represented in nature today. Whether this microbial
ommunity has the same composition in humans of the 21stentury as it did when associated with our ancient progeni-ors, somewhere in Africa, will perhaps never be determined.umans inhabiting developed countries today are, however,uch less likely to be colonized by parasitic worms (helminthes)
nd it is possible that the more hygienic lifestyles of recentecades have resulted in additional alterations to the compositionf the bowel community because exposure to certain bacteriaay be less common [1,2]. Even so, extant bowel communities
f humans are impressive both qualitatively (biodiversity) anduantitatively. Large bowel contents and feces contain about011 bacterial cells per gram (wet weight) and bacterial cellsomprise about 50% of fecal mass [3]. Four bacterial phyla areepresented (Firmicutes [Gram-positive], Bacteroidetes [Gram-egative], Actinobacteria [Gram-positive], and ProteobacteriaGram-negative]) and three groups, still phyogenetically broad,
ach containing many genera and species, are numerically dom-nant in the feces of healthy humans (Clostridium coccoidesroup, Clostridium leptum subgroup, Bacteroides-Prevotella∗ Tel.: +64 3 479 7713; fax: +64 3 479 8540.E-mail address: [email protected].
atidsii
044-5323/$ – see front matter © 2006 Elsevier Ltd. All rights reserved.oi:10.1016/j.smim.2006.09.001
roup) [4,5]. Bacteria, then, dominate the bowel community.ungi and Archaea may also be resident, but comprise less than.05% and 1% of the total inhabitants, respectively [6,7]. Fur-her information concerning the phylogenetic composition ofhe bowel community is provided in Fig. 1. Much of this infor-
ation has been generated through the application of nucleiccid-based methodologies, most of which target the nucleotidease sequence of small ribosomal subunit RNA (16S rRNA inhe case of bacteria) which provides a cornerstone of micro-ial taxonomy. An earlier estimate of the number of bacterialpecies that might be resident in the human large bowel wasased on bacteriological culture. Four hundred species seemedlikely number by extrapolation from what had already been
ultured [12,13]. Nucleic acid-based methods of detection sug-est that about 50% of the bacterial cells seen microscopicallyn feces cannot yet be cultured in the laboratory, even whenccounting for the fact that some of the bacteria are dead [14,15].his phenomenon, manifest even more dramatically in terrestrialnd aquatic ecosystems was, based on traditional bacteriolog-cal experience, totally unexpected and has been called “thereat plate count anomaly” [16]. “Operational taxonomic units”OTU; molecular species) never encountered in culture-basedacteriology are detectable by the molecular methods, revealingnew world remaining to be investigated by bacteriologists of
he future. Estimates of biodiversity now seem to continuallynflate, probably because many of the 16S rRNA sequences in
atabanks differ by one nucleotide base. These are likely to beequences representing the same OTU but containing sequenc-ng errors. Curiously, therefore, we do not really know what,n any detail, our modern bowel community is composed of let
G.W. Tannock / Seminars in Imm
F
aap
2
guvrotcgahpglttbaioi
fdwhNfchtmtacdcatn
3
afgobppoioseogwrcplotgwtatrcaob
ig. 1. Where and what types of bacteria are detected in the human gut [8–11].
lone what that of ancient humans contained. What we do know,nd all that matters really, is that these bowel inhabitants haverofound effects upon us.
. Unseen and unfelt
A chemostat is a culture apparatus that is used to maintain therowth of bacterial cultures continuously and at a constant ratender laboratory conditions. Culture medium is fed into a cultureessel containing the bacterial cells at a rate controlled by a flowegulator. The culture volume is held constant by means of anverflow tube that enables spent medium (effluent) to pass fromhe culture vessel. The human large bowel is the equivalent of ahemostat. It is fed with culture medium derived from the undi-ested components of the diet that pass from the small bowel,s well as substances produced endogenously by the humanost. Hence this culture medium is particularly rich in com-lex carbohydrates derived from plant cell walls and complexlycoproteins from mucus [17,18]. The ileo-cecal valve regu-ates the flow of culture medium into the large bowel; feces arehe effluent. Under these circumstances, a continuous fermen-ation of exogenous and endogenous substrates by consortia ofacteria proceeds, resulting in the formation of short chain fatty
cids (mainly acetic, propionic, and butyric), amines, phenols,ndoles, and gases as the major products [17]. The absorptionf the fatty acids from the bowel lumen once probably providedmportant caloric and carbon sources for humans, just as they dowtti
unology 19 (2007) 94–105 95
or other animal species even today [19]. Well-fed New Zealan-ers and North Americans, as examples, can doubtless manageithout this nutritional contribution much better than their pre-istoric forebears whose diet was impoverished by comparison.evertheless, nutritionists invoke butyric acid as an important
uel for colonocytes [20]. One would imagine that the bacterialulture responsible for the fermentation would be the same fromuman to human since bowel biochemistry follows the same pat-ern from one person to the next [5]. Yet, again on the basis ofolecular methods of analysis, it appears that bowel communi-
ies are about 30% dissimilar between humans (Fig. 2). Thoughsurprising fact when first encountered, the uniqueness of bowelommunities is likely to be due to extensive functional redun-ancy in the bacterial World: more than one bacterial speciesan carry out a particular catabolic process. Thus, just as there isnormal range of blood chemistry values in the human popula-
ion, there is also a range of inhabitants that can fill the ecologicaliches responsible for the overall bowel fermentation.
. It’s a wise man that knows his own father
According to Sophocles, Oedipus did not know hisntecedents and, because of this, in all innocence, murdered hisather and married his mother. A colossal tragedy resulting fromenealogical ignorance. We do not know where the phylogeneticrigins of the members of bacterial communities in the bowel lie,ut an educated guess would indicate that their ancestors werelant-associated, and regularly ingested by mammals. Cellulose,ectins, arabino-xylans, starch and beta-glucans, all substancesf plant origin, figure prominently as substrates for the predom-nant bowel bacteria [22–24]. Genomic sequences of residentsf the human bowel (Bacteroides and Bifidobacterium species)how that these bacteria devote much of their genomes to genesncoding hydrolytic enzymes, often contained in self-regulatedperons, that catalyze the degradation of oligosaccharides andlycoproteins [22,23]. There are also many genes associatedith signal transduction by which the bacteria can detect and
eact to changes in the nutrient milieu. Responses to qualitativehanges in substrate availability, and well-regulated metabolicathways that conserve cellular energy, help to explain the eco-ogical fitness of these bacteria and the phylogenetic stabilityf bowel communities [25]. The numerically predominant bac-eria of bowel communities are obligate anaerobes. This sug-ests a long-distant temporal origin when Earth’s atmosphereas anoxic, but their biochemical evolution must have con-
inued after the appearance of plants with complex cell wallsnd vascular systems that provide their nutritional substratesoday. Perhaps they were decomposers in sediments and natu-al composts, or similar anaerobic environments rich in complexarbohydrates? Some of these environmental bacteria must havedapted to the bowel environment so that residence, still on theutside (bowel lumen), but sequestered inside, of the humanody was established. The human bowel is daily contaminated
ith bacterial cells present in food and water. Some of these bac-eria survive and transit the digestive tract and are detectable inhe feces [26]. The same applies to “probiotic” bacteria that arententionally consumed in self-care health products [27]. These

96 G.W. Tannock / Seminars in Immunology 19 (2007) 94–105
al com
“msotoa[acbhw
atihmvtit
Fig. 2. The composition of the fecal bacteri
allochthonous” bacteria do not become part of the bowel com-unity because they are no longer detected in feces once the
ource of their ingestion is removed. “Autochthonous” bacteria,n the other hand, have a habitat in the bowel. They have long-erm associations with the human host, form stable populationsf characteristic size, and have demonstrable means of earningliving (an ecological function or niche) in the distal bowel
28]. Researchers interested in the bowel community need to beware of the Oedipus tragedy; would not it be embarrassing to
laim that the digestive tract was colonized by a vast array ofacteria when, in fact, only the DNA of allochthonous microbesad been detected? Unraveling the secrets of life in the bowelould be impossible if an allochthonous fecal isolate was chosentbas
munity varies from human to human [21].
s the model organism for study! Snapshots of the composi-ion of bowel communities are only the beginning of ecologicalnvestigations and DNA-based methods reveal only “who mightave been there”. Temporal (quantitative sequential measure-ents) and function-based (transcriptomics, proteomics) obser-
ations are necessary to define bacterial communities. We needo know “what the bowel residents are doing and how they dot”. It would also be very interesting to know how the transi-ion from allochthonous to autochthonous status in relation to
he bowel was achieved. The highly competitive nature of theowel community may have represented, and perhaps still does,hotbed of evolution where novel attributes developed and wereelected. The molecular (genetic) processes by which bacteria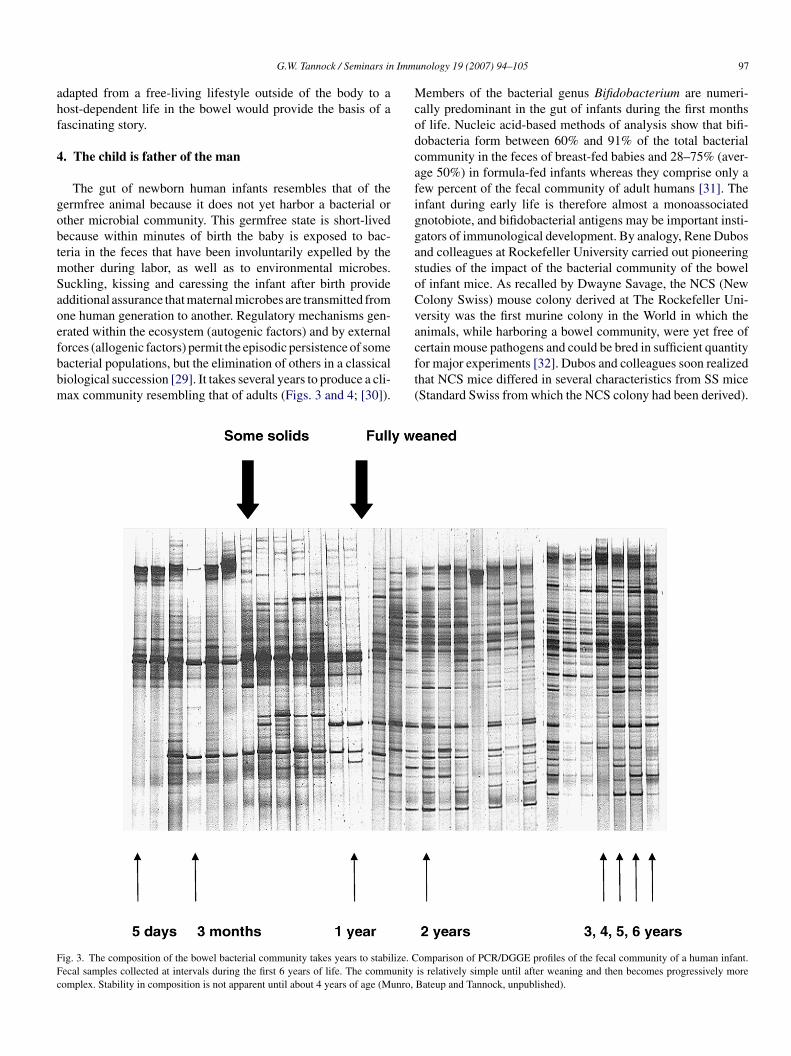
Imm
ahf
4
gobtmSaoefbbm
McodcafiggasoCva
FFc
G.W. Tannock / Seminars in
dapted from a free-living lifestyle outside of the body to aost-dependent life in the bowel would provide the basis of aascinating story.
. The child is father of the man
The gut of newborn human infants resembles that of theermfree animal because it does not yet harbor a bacterial orther microbial community. This germfree state is short-livedecause within minutes of birth the baby is exposed to bac-eria in the feces that have been involuntarily expelled by the
other during labor, as well as to environmental microbes.uckling, kissing and caressing the infant after birth providedditional assurance that maternal microbes are transmitted fromne human generation to another. Regulatory mechanisms gen-rated within the ecosystem (autogenic factors) and by external
orces (allogenic factors) permit the episodic persistence of someacterial populations, but the elimination of others in a classicaliological succession [29]. It takes several years to produce a cli-ax community resembling that of adults (Figs. 3 and 4; [30]).cft(
ig. 3. The composition of the bowel bacterial community takes years to stabilize. Cecal samples collected at intervals during the first 6 years of life. The communityomplex. Stability in composition is not apparent until about 4 years of age (Munro,
unology 19 (2007) 94–105 97
embers of the bacterial genus Bifidobacterium are numeri-ally predominant in the gut of infants during the first monthsf life. Nucleic acid-based methods of analysis show that bifi-obacteria form between 60% and 91% of the total bacterialommunity in the feces of breast-fed babies and 28–75% (aver-ge 50%) in formula-fed infants whereas they comprise only aew percent of the fecal community of adult humans [31]. Thenfant during early life is therefore almost a monoassociatednotobiote, and bifidobacterial antigens may be important insti-ators of immunological development. By analogy, Rene Dubosnd colleagues at Rockefeller University carried out pioneeringtudies of the impact of the bacterial community of the bowelf infant mice. As recalled by Dwayne Savage, the NCS (Newolony Swiss) mouse colony derived at The Rockefeller Uni-ersity was the first murine colony in the World in which thenimals, while harboring a bowel community, were yet free of
ertain mouse pathogens and could be bred in sufficient quantityor major experiments [32]. Dubos and colleagues soon realizedhat NCS mice differed in several characteristics from SS miceStandard Swiss from which the NCS colony had been derived).omparison of PCR/DGGE profiles of the fecal community of a human infant.is relatively simple until after weaning and then becomes progressively moreBateup and Tannock, unpublished).
98 G.W. Tannock / Seminars in Imm
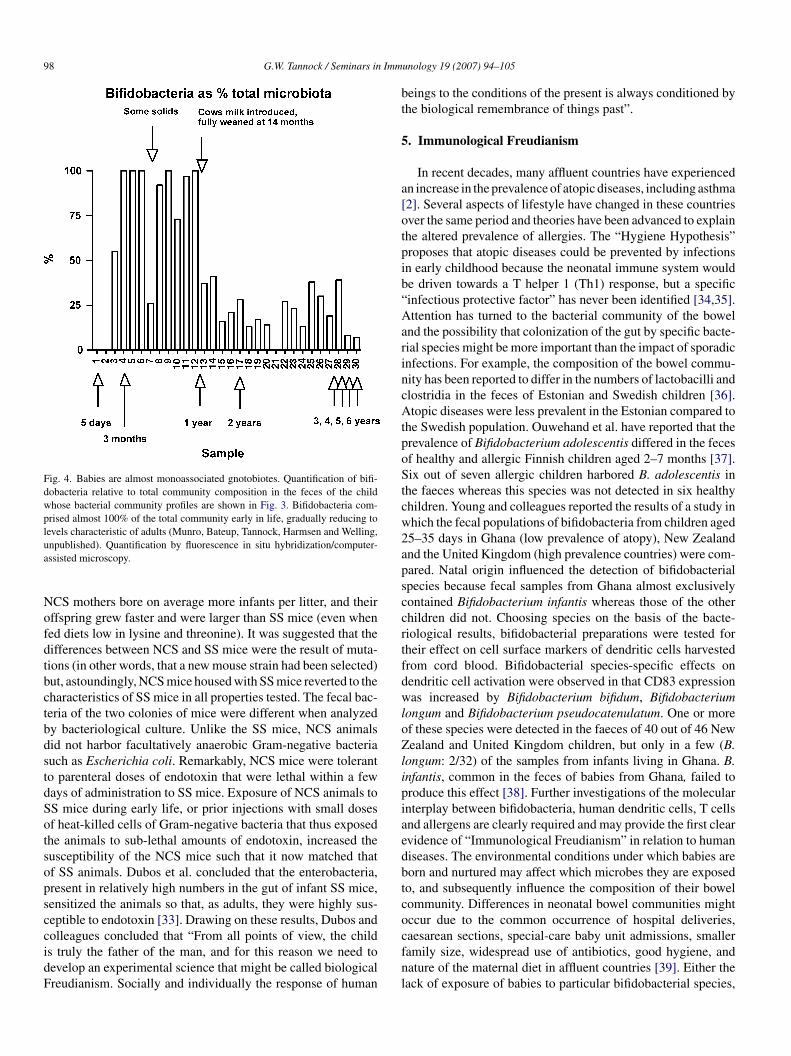
Fig. 4. Babies are almost monoassociated gnotobiotes. Quantification of bifi-dobacteria relative to total community composition in the feces of the childwhose bacterial community profiles are shown in Fig. 3. Bifidobacteria com-prised almost 100% of the total community early in life, gradually reducing tolevels characteristic of adults (Munro, Bateup, Tannock, Harmsen and Welling,ua
NofdtbctbdstdSotsopsccidF
bt
5
a[otpib“AarincAtpoStcw2apsccrtfdwloZlipiaedbtcocaesarean sections, special-care baby unit admissions, smaller
npublished). Quantification by fluorescence in situ hybridization/computer-ssisted microscopy.
CS mothers bore on average more infants per litter, and theirffspring grew faster and were larger than SS mice (even whened diets low in lysine and threonine). It was suggested that theifferences between NCS and SS mice were the result of muta-ions (in other words, that a new mouse strain had been selected)ut, astoundingly, NCS mice housed with SS mice reverted to theharacteristics of SS mice in all properties tested. The fecal bac-eria of the two colonies of mice were different when analyzedy bacteriological culture. Unlike the SS mice, NCS animalsid not harbor facultatively anaerobic Gram-negative bacteriauch as Escherichia coli. Remarkably, NCS mice were toleranto parenteral doses of endotoxin that were lethal within a feways of administration to SS mice. Exposure of NCS animals toS mice during early life, or prior injections with small dosesf heat-killed cells of Gram-negative bacteria that thus exposedhe animals to sub-lethal amounts of endotoxin, increased theusceptibility of the NCS mice such that it now matched thatf SS animals. Dubos et al. concluded that the enterobacteria,resent in relatively high numbers in the gut of infant SS mice,ensitized the animals so that, as adults, they were highly sus-eptible to endotoxin [33]. Drawing on these results, Dubos andolleagues concluded that “From all points of view, the child
s truly the father of the man, and for this reason we need toevelop an experimental science that might be called biologicalreudianism. Socially and individually the response of humanfnl
unology 19 (2007) 94–105
eings to the conditions of the present is always conditioned byhe biological remembrance of things past”.
. Immunological Freudianism
In recent decades, many affluent countries have experiencedn increase in the prevalence of atopic diseases, including asthma2]. Several aspects of lifestyle have changed in these countriesver the same period and theories have been advanced to explainhe altered prevalence of allergies. The “Hygiene Hypothesis”roposes that atopic diseases could be prevented by infectionsn early childhood because the neonatal immune system woulde driven towards a T helper 1 (Th1) response, but a specificinfectious protective factor” has never been identified [34,35].ttention has turned to the bacterial community of the bowel
nd the possibility that colonization of the gut by specific bacte-ial species might be more important than the impact of sporadicnfections. For example, the composition of the bowel commu-ity has been reported to differ in the numbers of lactobacilli andlostridia in the feces of Estonian and Swedish children [36].topic diseases were less prevalent in the Estonian compared to
he Swedish population. Ouwehand et al. have reported that therevalence of Bifidobacterium adolescentis differed in the fecesf healthy and allergic Finnish children aged 2–7 months [37].ix out of seven allergic children harbored B. adolescentis in
he faeces whereas this species was not detected in six healthyhildren. Young and colleagues reported the results of a study inhich the fecal populations of bifidobacteria from children aged5–35 days in Ghana (low prevalence of atopy), New Zealandnd the United Kingdom (high prevalence countries) were com-ared. Natal origin influenced the detection of bifidobacterialpecies because fecal samples from Ghana almost exclusivelyontained Bifidobacterium infantis whereas those of the otherhildren did not. Choosing species on the basis of the bacte-iological results, bifidobacterial preparations were tested forheir effect on cell surface markers of dendritic cells harvestedrom cord blood. Bifidobacterial species-specific effects onendritic cell activation were observed in that CD83 expressionas increased by Bifidobacterium bifidum, Bifidobacterium
ongum and Bifidobacterium pseudocatenulatum. One or moref these species were detected in the faeces of 40 out of 46 Newealand and United Kingdom children, but only in a few (B.
ongum: 2/32) of the samples from infants living in Ghana. B.nfantis, common in the feces of babies from Ghana, failed toroduce this effect [38]. Further investigations of the molecularnterplay between bifidobacteria, human dendritic cells, T cellsnd allergens are clearly required and may provide the first clearvidence of “Immunological Freudianism” in relation to humaniseases. The environmental conditions under which babies areorn and nurtured may affect which microbes they are exposedo, and subsequently influence the composition of their bowelommunity. Differences in neonatal bowel communities mightccur due to the common occurrence of hospital deliveries,
amily size, widespread use of antibiotics, good hygiene, andature of the maternal diet in affluent countries [39]. Either theack of exposure of babies to particular bifidobacterial species,

Imm
atctia
6
b1aeesuiiwTimcttiaclcfbrtti[tbNtbtvgdagwoapwnrn
[tcadeeoocttii
7
atrbTiabmasmoco(lIfgbsbsIcmbo(tgtAi
G.W. Tannock / Seminars in
nd/or elimination of bifidobacterial species from the bowelhrough the use of antibiotics, might reduce the exposure ofhildren in early life to important bacterial antigens at a criticalime in the maturation of the immune system, for examplen removing (immune deviation) the T helper 2 (Th2) skewpparently characteristic of the newborn [40,41].
. Retained but contained
The antigenic burden associated with approximately 2 × 1013
acterial cells (exceeding the human population of Earth about0,000-fold) present at any moment in the large bowel is prob-bly incalculable. Dendritic cells associated with the bowelpithelium may sample this antigenic landscape regularly byxtending dendrites between enterocytes to reach the mucosalurface [42]. M cells of Peyer’s Patches are also sites of antigenptake [43]. Relative to germfree animals, the lamina proprian the gut of animals harboring bacterial communities is mildlynflamed in terms of histopathology [44]. Antibodies reactiveith bowel bacterial antigens are present in human sera [45].hus bacterial antigens are detected by the innate and adaptive
mmune systems, but inflammation is somehow kept at mini-al levels. How “tolerance” (hypo-responsiveness) to the bowel
ommunity is achieved mechanistically should be an interestingopic of immunological research. It can be postulated, though,hat sequestration of the bacterial community away from themmune system in a “virtual tube” within the bowel may belarge part of the answer. The mucosal surface of the bowel is
overed with a blanket of mucus that flows away from the epithe-ium and is continuously replaced by fresh mucus from the gobletells [46]. This moving, sticky blanket cleanses the mucosal sur-ace keeping bacterial cells at bay. The tight junctions formedetween enterocytes render the epithelium a formidable bar-ier to bacterial penetration [47]. Moreover, bacterial cells inhe bowel are coated with sIgA, possibly counteracting effec-ive use of bacterial adhesins [48]. Defensins liberated in thentestinal crypts by Paneth cells complete the “wall” of the tube49]. Everything is designed to stop bacteria associating withhe mucosal surface. Contained within the virtual tube in theowel lumen, the bacterial community can be retained safely.evertheless, it does not seem to be a totally efficacious sys-
em since pathogens can reach the mucosal surface and attachy means of adhesins to specific receptors prior to prolifera-ion or invasion. Even in health, small temporary cracks in theirtual walls of the tube must occur which allow bacterial anti-ens to seep from the tube and make contact with enterocytes,endrites of dendritic cells, and M cells. Although enterocytesre not professional immune cells, contact with bacterial anti-ens may cause a transient induction of “immune responses”ithin them. For example, Ruiz et al. have described inductionf phosphorylation/activation of the NF-�B transcriptionallyctive subunit RelA and the mitogen-activated protein kinase38 in enterocytes by an innocuous bifidobacterial species,
hich resulted in interleukin-6 gene expression [50]. The cor-erstone of innate signaling between bacteria and enterocytesests on pattern recognition receptors (Toll-like receptors [TLR],ucleotide-binding oligomerization domain protein receptorsitmd
unology 19 (2007) 94–105 99
NOD]) that recognize molecular patterns associated with bac-erial cells regardless of whether they originate in the bowelommunity or are pathogens [51]. However, pathogens inducen acute inflammatory response, harmless bacteria do not. Thisifferential effect may be due to the relative location of differ-nt TLR (apical membrane, intracellular, basal membrane ofnterocytes), the rapid turnover of enterocytes at villous tips,r to particular properties of pathogens (virulence factors) thatther bacteria lack. Negative regulators exerted by the entero-ytes themselves might also mediate the differential responseo signals from bowel inhabitants and pathogens [52]. Defininghese differential/regulatory systems is important immunolog-cal research because it impinges on the etiology of chronicmmune inflammatory bowel diseases.
. Surrogate pathogens
Failure to demonstrate convincingly an association betweenspecific pathogen and chronic immune inflammatory condi-
ions of the bowel (Crohn’s disease and ulcerative colitis) hasesulted in the concept that the inflammation is fueled by theacterial bowel community, or at least some members of it [53].his is demonstrably true in experimental animal models of col-
tis in which gene deletion or transgenic manipulation producesdysfunctional immune system that reacts aggressively with
acterial antigens [53]. Under germfree conditions, these ani-als have minimal disease but develop colitis when exposed tospecific-pathogen-free collection of bowel bacteria, or selectedtrains of bowel bacteria. Human patients suffering from inflam-atory bowel diseases (IBD) have a broken tolerance to their
wn bowel bacteria as has been clearly demonstrated by thelassical studies of Duchmann et al. [54]. Familial clusteringf cases and association with specific chromosomal aberrationsat least in a proportion of patients) and with particular humaneukocyte antigen genotypes indicate a genetic predisposition toBD [55,56]. The bowel community may indeed be the antigenicuel for chronic inflammation (in this sense acting as a surro-ate pathogen), but how and what initiates the process? It coulde postulated, perhaps, that the enterocytes and, concurrently orubsequently, immune cells are exposed to excessive amounts ofacterial antigens because the virtual, multi-component defen-ive wall of the bowel of humans predisposed genetically toBD is defective in at least one factor. For example, the alteredhemical composition of the mucus of ulcerative colitis patientsay be linked to the larger number of bacteria associated with
iopsies collected from them (Fig. 5). Genetically based causesf breaches in the wall, possibly occurring very early in lifediagnosis of IBD is characteristically made in young adults),ogether with broken immunological tolerance to bacterial anti-ens, could result in a sensitization to bacterial antigens andhe progression of poorly controlled inflammatory reactions.lthough the initiating factors may well occur early in life, stud-
es of the composition of the distal bowel community represented
n feces have mostly been conducted with adult IBD patients inhe hope that they would reveal specific groups of bacteria thatight be associated with inflammation. These studies have pro-uced extremely variable results (Table 1). In retrospect, this
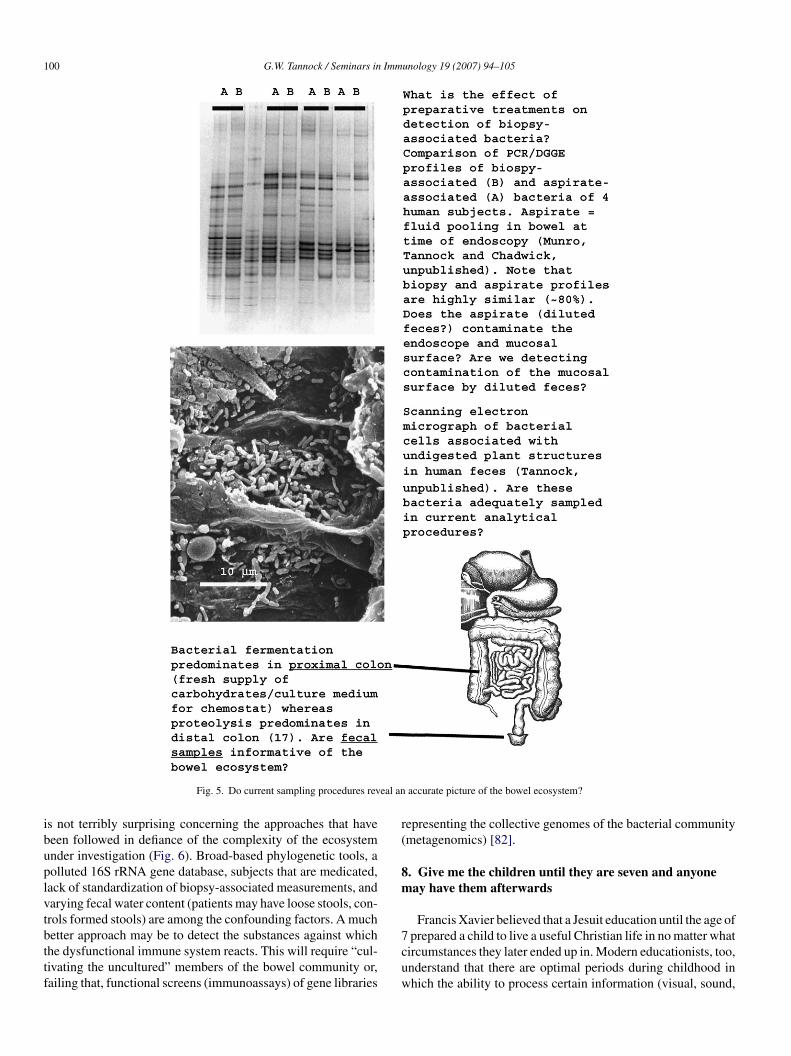
100 G.W. Tannock / Seminars in Immunology 19 (2007) 94–105
eal an
ibuplvtbttf
r(
8m
Fig. 5. Do current sampling procedures rev
s not terribly surprising concerning the approaches that haveeen followed in defiance of the complexity of the ecosystemnder investigation (Fig. 6). Broad-based phylogenetic tools, aolluted 16S rRNA gene database, subjects that are medicated,ack of standardization of biopsy-associated measurements, andarying fecal water content (patients may have loose stools, con-rols formed stools) are among the confounding factors. A much
etter approach may be to detect the substances against whichhe dysfunctional immune system reacts. This will require “cul-ivating the uncultured” members of the bowel community or,ailing that, functional screens (immunoassays) of gene libraries7cuw
accurate picture of the bowel ecosystem?
epresenting the collective genomes of the bacterial communitymetagenomics) [82].
. Give me the children until they are seven and anyoneay have them afterwards
Francis Xavier believed that a Jesuit education until the age of
prepared a child to live a useful Christian life in no matter whatircumstances they later ended up in. Modern educationists, too,nderstand that there are optimal periods during childhood inhich the ability to process certain information (visual, sound,
G.W. Tannock / Seminars in Immunology 19 (2007) 94–105 101
Table 1Summary of investigations of the bowel bacterial community of humans in relation to inflammatory bowel diseases
Investigation Patients medicated forIBD or administeredantimicrobial drugs?
Observations Reference
Biopsies were collected from five colonic sites as well asthe terminal ileum of nine healthy subjects. Fluorescencein situ hybridization/microscopy was used to target fecalbacterial groups
None Bacterial cells were seen associated with the luminalsurface of mucus and seldom in the mucus itself
[57]
Colonic biopsies from 21 ulcerative colitis patients, 14Crohn’s disease patients, and 24 controls were collectedand treated with dithiothreitol to remove mucus, thenmucosa-associated and intramucosal bacteria (released byhypotonic lysis and gentamicin treatment) were cultured
Yes, IBD treatment Bacteria were cultured more commonly fromsamples from Crohn’s disease patients compared tothose from ulcerative colitis or controls. AdherentEscherichia coli were more commonly detected insamples from Crohn’s disease patients compared tocontrols
[58]
DNA extracted from paraffin-embedded tissue samplescollected from 16 Crohn’s disease, 11 ulcerative colitisand 18 colon cancer patients was used to enumerateMycobacterium species, Bacteroides vulgatus, and E. coliusing real time quantitative PCR
IBD treatment notstated
Mycobacteria were not detected but B. vulgatus andE. coli were detected more frequently and in greaternumbers in samples obtained from Crohn’s diseaseand ulcerative colitis patients compared to coloncancer subjects
[59]
Colonic biopsies were washed and used for culture ofbacteria, and detection of bacteria by electronmicroscopy, fluorescence in situ hybridization, and PCR.The results from 28 patients with self-limiting colitis, 104with indeterminate colitis, 119 with ulcerative colitis, 54with Crohn’s disease, and 40 asymptomatic subjects werecompared
IBD treatment notstated
There were more bacterial cells associated withbiopsies collected from patients than fromasymptomatic controls. The numbers of bacteriaincreased in association with increasedinflammation (colitis)
[60]
Rectal biopsies and feces were collected from 12 Crohn’sdisease patients during periods with or withoutantimicrobial (metronidazole, cotrimoxazole) treatment.Culture-based studies were conducted
Yes, antimicrobialdrugs during part ofthe study
Eight of the 12 patients showed clinicalimprovement when administered antimicrobialdrugs but this could not be correlated withbacteriological results
[61]
Biopsies collected from four sites (ileum, right and leftcolon, rectum) were collected from 20 Crohn’s disease,11 ulcerative colitis, and 4 control subjects. Temporaltemperature gradient gel electrophetic fingerprints ofbiopsy-associated bacteria were prepared
IBD treatment notstated
Similar profiles were obtained from sampling sitesintra-individual, but varied inter-individually.Profiles did not discriminate between subject groups
[62]
Rectal biopsies collected from 9 ulcerative colitis patientsand 10 control subjects were examined by culture andfluorescence in situ hybridization/microscopy
IBD treatment notstated
The bacterial types that were detected variedbetween subjects making it impossible to assign apathogenic role to specific bacteria
[63]
Rectal biopsies collected from 33 ulcerative colitis patients,6 Crohn’s disease, and 14 control subjects were examinedby fluoresence in situ hybridization/microscopy
Yes, IBD treatment Bifidobacterial numbers were lower in ulcerativecolitis patients; E. coli numbers were higher inulcerative colitis and Crohn’s disease patients
[64]
Tissue sections were prepared from resected intestine(terminal ileum, colon) and examined by fluorescence insitu hybridization/microscopy. Samples were collectedfrom 12 ulcerative colitis, 12 Crohn’s disease, and 14control subjects
IBD treatment notstated. Antimicrobialdrugs givenintravenously justprior to surgery
There were higher numbers of bacteria associatedwith tissue collected from ulcerative colitis andCrohn’s disease subjects and there was moreevidence of bacterial invasion of tissue. Samplesfrom Crohn’s disease patients had mostlyproteobacteria and Bacteroides-Prevotellaassociated with the tissue, whereas a more diversecollection of bacteria was present in ulcerativecolitis samples
[65]
Biopsies collected endoscopically or surgically from theterminal ileum of 11 Crohn’s disease and 11 controlsubjects were examined by PCR
IBD treatment notstated. Antimicrobialsprior to surgery
16S rRNA gene sequences similar to those ofHelicobacter species, Mycobacterium aviumsubspecies paratuberculosus, Listeriamonocytogenes and E. coli were detected in three offive biopsies collected surgically from Crohn’sdisease patients. Biospies collected from all subjectsby endoscopy had associated bacteria, but biopsiescollected at surgery from controls were devoid ofbacteria
[66]
Colonic biopsies collected from 13 ulcerative colitispatients and 61 controls were examined by PCRcombined with enrichment culture to detect the presenceof sulphate-reducing bacteria
IBD treatment notstated
All of the biopsies contained sulphate-reducingbacteria
[67]

102 G.W. Tannock / Seminars in Immunology 19 (2007) 94–105
Table 1 (Continued )
Investigation Patients medicated forIBD or administeredantimicrobial drugs?
Observations Reference
Colonic biopsies from 26 Crohn’s disease, 31 ulcerativecolitis, 15 controls with inflammation, and 31non-inflamed control subjects were examined by 16SrRNA gene profiling, the preparation of 16S rRNA geneclone libraries, and real time quantitative PCR
Yes, IBD treatment The diversity of bacterial types was less in samplesfrom Crohn’s disease and ulcerative colitis patientscompared to controls. This was due to a reduction indetection of Bacteroides species, Eubacteriumspecies, and Lactobacillus species
[68]
Rectal biopsies were collected from 19 IBD patients (notdifferentiated) and 14 controls. The biopsies wereexamined by RNA in situ hybridization/microscopy
Yes, IBD treatment Bacterial cells were not observed in the case of 10control and 6 IBD patient biopsies. In the remainingsamples, there were more bacteria associated withIBD biopsies compared to those of controls. Thebacteria were seen within the mucus but not incontact with enterocytes, nor within the laminapropria
[69]
16S rRNA gene clone libraries were prepared from 16biopsies collected from Crohn’s disease patients(aphthous ulcers), 15 surgical samples from Crohn’sdisease patients, and biopsies from controls
Yes, IBD treatment Specific phylogenetic groups of bacteria could notbe associated with Crohn’s disease lesions
[70]
Ileal and colonic tissues from Crohn’s disease patients andcontrols were studied to determine the prevalence ofadherent-invasive strains of E. coli
IBD treatment notstated
Adherent-invasive strains of E. coli were morecommonly detected in association with tissues,particularly from the ileum, of Crohn’s diseasepatients compared to those of controls
[71]
Tissue samples collected at surgery from 20 Crohn’s diseasepatients and 6 controls, and examined by nested PCR andfluorescence in situ hybridization/microscopy for thepresence of Mycobacterium avium subspeciesparatuberculosis
Yes, IBD treament M. avium subsp. paratuberculosis DNA wasdetected more commonly in samples from Crohn’sdisease patients than from those of controls.
[72]
Archival tissue obtained at surgery from 15 Crohn’s diseasepatients and 10 non-Crohn’s granulomatous boweldisease patients were examined. Granulomas weremicrodissected from the tissue and DNA was extracted.Nested PCR was used to detect sequences characteristicof E. coli.
Treatments not stated. E. coli DNA was detected more frequently insamples from Crohn’s disease patients than fromnon-Crohn’s disease patients. The authors concludedthat, since a similar result with respect to M. aviumsubsp. paratuberculosis had been obtained withthese same samples in a previous study, the findingsmay reflect non-specific association of bacteria withgranulomas, rather than cause and effect
[73,74]
Colonic biopsies were collected from 4 ulcerative colitispatients and 2 controls. 16S rRNA gene clone librarieswere prepared and selected libraries were probed with alabeled oligonucleotide probe that detectedBacteroides-Prevotella
Yes, IBD treatment There were more clones representingBacteroides-Prevotella bacteria in libraries fromulcerative colitis patients than from controls
[75]
Pediatric patients with Crohn’s disease (12 subjects),ulcerative colitis [7], other intestinal abnormalities [16],and 7 controls were studied. Biopsies were collectedfrom ileum, cecum and rectum and examined bybacteriological culture and qualitative and quantitativePCR
None Higher numbers of facultatively anaerobic bacteriawere associated with samples from Crohn’s diseaseand ulcerative colitis patients compared to controls.The numbers of Bacteroides vulgatus were lower inthe case of Crohn’s disease and ulcerative colitispatients relative to controls
[76]
16S rRNA gene clone libraries were prepared from biopsiescollected from 20 Crohn’s disease, 15 ulcerative colitispatients, and 14 controls. Patient biopsies were collectedfrom both inflamed and non-inflamed mucosa of eachsubject. Denaturing gradient gel electrophoretic (DGGE)profiles were prepared, and total bacterial numbersassociated with biopsies were enumerated by real timequantitative PCR
None. Patients werenewly diagnosed anduntreated.
DGGE profiles were similar for all sites sampledintra-individually but differed inter-individually.LIBSHUFF statistical comparison showed that 16SrRNA gene clone libraries prepared from Crohn’sdisease, ulcerative colitis and control subjects haddifferent compositions. Collections of bacteriaassociated with inflamed tissue did not differ fromthose of non-inflamed tissue. There were morebacterial cells associated with biopsies collectedfrom ulcerative colitis patients compared to those ofCrohn’s disease and control subjects. Unclassifiedmembers of the Class Bacteroidetes were moreprevalent in the samples from Crohn’s diseasepatients compared to the other groups.
[77]
The fecal bacterial communities of Crohn’s disease (fivepatients) and healthy controls (five) were compared bybacteriological culture
None The number of bacteria belonging to the Bacteroidesfragilis group was higher, and these bacteria weredetected more frequently, in patients than controls
[78]
G.W. Tannock / Seminars in Immunology 19 (2007) 94–105 103
Table 1 (Continued )
Investigation Patients medicated forIBD or administeredantimicrobial drugs?
Observations Reference
Two metagenomic libraries were prepared from fecalsamples obtained from 6 Crohn’s disease patients and 6controls, respectively. 16S rRNA genes detected in thelibraries were sequenced
Yes, IBD treatment Forty three OTU belonging to the phylumFirmicutes were detected in the control library butonly 13 in the Crohn’s disease library. This reduceddiversity in Crohn’s disease subjects was confirmedby fluorescence in situ hybridization/flow cytometryexamination of feces in which a reduction in theClostridum leptum subgroup as a proportion of thetotal bacterial community was observed
[79]
Biopsies were collected from the intestines of 15 Crohn’sdisease patients during flare-ups of disease. Inflamed andnon-inflamed mucosal samples were obtained andexamined by PCR/TTGE
IBD treatment notstated
The bacterial profiles did not differ betweeninflamed and non-inflamed tissues
[80]
Radioactive probes were used to quantify bacterial groupsin the feces of 17 Crohn’s disease patients and 16controls. Community profiles were prepared by temporaltemperature gradient gel electrophoresis (TTGE)
Yes, IBD treatment Enterobacteria were more numerous in the feces ofCrohn’s disease patients, but Bacteroides andClostridium coccoides group numbers weredecreased compared to those of controls. There was
[81]
ntitspsefhichibr
FucwnaFr
(dlactadbt4cw
umbers, language) can be acquired. Comparisons of the charac-eristics of germfree, monoassociated or conventionalized micen short-term experiments show that the presence of bowel bac-eria influences gene expression in the bowel mucosa. Mostpectacular are the impacts on the transcription of genes whoseroducts influence epithelial integrity, angiogenesis, and depo-ition of fat in adipocytes [83–85]. The results of gnotobioticxperiments are certainly dramatic and point to the potentialor cross-talk between bowel bacteria and the tissues of theirost. These studies do not tell us if the same interactions occurn humans but, if they do, they are most likely to occur inhildhood. Gnotobiotic animals mimic the bowel conditions ofuman neonates that are initially bacteria-free but then are, to all
ntents and purposes, monoassociated with bifidobacteria, laterecoming associated with a progressively more complex bacte-ial collection that seems to climax at about 4 years after birthig. 6. Comparison of numbers of bacteria associated with biopsies. Mean val-es and standard errors are shown. Bacterial numbers were the same for samplesollected from Crohn’s disease (CD) and healthy subjects, whereas numbersere about doubled (P <0.01) for ulcerative colitis patients (UC). This wasot due to inflammation (I) of the tissue since noninflamed (NI) mucosa wasssociated with similar numbers of bacteria (77; reproduced with permission).ourteen, 11 and 33 biopsies were examined for the CD, UC and healthy groups,espectively.
lotoatita(tfino
R
temporal variation in TTGE profiles of patientscompared to stability in those of controls
Figs. 3 and 4). The neonatal bowel is physiologically immatureuring early life and programming of the immune system seemsikely to be a continuous process during the first years of lifes appropriate windows of opportunity with regards to dietaryomposition and bacterial exposure are opened [86]. If the vir-ual walls of the tube that sequester bacteria and their antigensnd inhibit them from making mucosal contact are not yet fullyeveloped in the first few years of life, one could postulate thatacteria-host conversations, such as occurs in experimental gno-obiotes, might have long-lasting, biological repercussions. By
years of age, the education of the bowel tissue and its asso-iated immune phenomena might all be over [87]. All of theindows may be closed and shuttered. We are unlikely to ever
earn much about the composition of the bacterial communityf antediluvian humans. We do, however, have the opportunityo investigate the developing bowel community and maturationf the immune system of modern children. In Freudianism, thedult conscious is unknowingly influenced by the subconscioushat has developed in early life due to the influences of variousnteractive circumstances (social and environmental). Exposureo a succession of bacterial populations in early life may result inphysiological version of the Freudian iceberg in which the tip
the discernible result) is but a small portion of a gigantic struc-ure, most of which is unseen. Here lies a potentially productiveeld of research for bacteriologists, physiologists and immu-ologists who are interested in neonatal development, allergy,r inflammatory bowel diseases.
eferences
[1] Gerrard JW, Geddes CA, Reggin PL, Gerrard CD, Horne S. Serum IgElevels in white and metis communities in Saskatchewan. Ann Allergy
1976;37:91–100.[2] Hopkin JM. Mechanisms of enhanced prevalence of asthma and atopy indeveloped countries. Curr Opin Immunol 1997;9:788–92.
[3] Suau A, Bonnet R, Sutren M, Godon J-J, Gibson GR, Collins MD, et al.Direct analysis of genes encoding 16S rRNA from complex communities

1 Imm
[[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
04 G.W. Tannock / Seminars in
reveals many novel molecular species within the human gut. Appl EnvironMicrobiol 1999;65:4799–807.
[4] Eckburg PB, Bik EM, Bernstein CN, Purdom E, Dethlefsen L, Sar-gent M, et al. Diversity of the human intestinal microbial flora. Science2005;308:1635–8.
[5] Tannock GW. The intestinal microflora. In: Fuller R, Perdigon G, editors.Gut flora. Nutrition, immunity and health. Oxford: Blackwell Press; 2003.p. 1–23.
[6] Miller TL, Wolin MJ. Stability of Methanobacter smithii populations inthe microbial flora excreted from the human large bowel. Appl EnvironMicrobiol 1983;45:317–8.
[7] Simon GL, Gorbach SL. Intestinal flora in health and disease. Gastroen-terology 1984;86:174–93.
[8] Bik EM, Eckburg PB, Gill SR, Nelson KE, Purdom EA, Francois F, et al.Molecular analysis of the bacterial microbiota in the human stomach. ProcNatl Acad Sci USA 2006;103:732–7.
[9] Hayashi H, Takahashi R, Nishi T, Sakamoto M, Benno Y. Molecular anal-ysis of jejunal, ileal, caecal and recto-sigmoidal human colonic microbiotausing 16S rRNA gene libraries and terminal restriction fragment lengthpolymorphism. J Med Microbiol 2005;54:1093–101.
10] Drasar BS, Hill MJ. Human intestinal flora. London: Academic Press; 1974.11] Lay C, Rigottier-Gois L, Holmstrom K, Rajilic M, Vaughan EE, de Vos
WM, et al. Colonic microbiota signatures across five northern Europeancountries. Appl Environ Microbiol 2005;71:4153–5.
12] Moore WEC, Holdeman LV. Special problems associated with the isolationand identification of intestinal bacteria in fecal flora studies. Am J Clin Nutr1974;27:1450–5.
13] Finegold SM, Attebury R, Sutter VL. Effect of diet on human fecalflora: comparison of Japanese and American diets. Am J Clin Nutr1974;27:1456–69.
14] Ben-Amor K, Heilig H, Smidt H, Vaughan EE, Abee T, de Vos WM.Genetic diversity of viable, injured, and dead fecal bacteria assessed byfluorescence-activated cell sorting and 16S rRNA gene analysis. Appl Env-iron Microbiol 2005;71:4679–89.
15] Tannock GW, Munro K, Harmsen HJM, Welling GW, Smart J, Gopal PK.Analysis of the fecal microflora of human subjects consuming a probi-otic containing Lactobacillus rhamnosus DR20. Appl Environ Microbiol2000;66:2578–88.
16] Amann RI, Ludwig W, Schleifer K-H. Phylogenetic identification and insitu detection of individual microbial cells without cultivation. MicrobiolRev 1995;59:143–69.
17] Cummings JH, Macfarlane GT. The control and consequences of bacterialfermentation in the human colon. J Appl Bacteriol 1991;70:443–59.
18] Roberton AM, Corfield AP. Mucin degradation and its significance ininflammatory conditions of the gastrointestinal tract. In: Tannock GW, edi-tor. Medical importance of the normal microflora. Dordrecht, The Nether-lands: Kluwer Academic Publishers; 1999. p. 222–61.
19] McBee RH. Fermentation in the hindgut. In: Clarke RTJ, Bauchop T, edi-tors. Microbial ecology of the gut. London: Academic Press; 1977. p.185–222.
20] Roediger WE. Role of anaerobic bacteria in the metabolic welfare of thecolonic mucosa in man. Gut 1980;21:793–8.
21] Stebbings S, Munro K, Simon MA, Tannock G, Highton J, Harmsen H,et al. Comparison of the faecal microflora of patients with ankylosingspondylitis and controls using molecular methods of analysis. Rheuma-tology 2002;41:1395–401.
22] Gill SR, Pop M, DeBoy RT, Eckburg PB, Turnbaugh PJ, Samuel BS, etal. Metagenomic analysis of the human distal gut microbiome. Science2006;312:1355–9.
23] Schell MA, Karmirantzou M, Snel B, Vilanova D, Berger B, Pessi G, etal. The genome sequence of Bifidobacterium longum reflects its adap-tation to the human gastrointestinal tract. Proc Natl Acad Sci USA2002;99:14422–7.
24] Xu J, Bjursell MK, Himrod J, Deng S, Carmichael LK, Chiang HC, et al.A genomic view of the human-Bacteroides thetaiotaomicron symbiosis.Science 2003;299:2074–6.
25] Zoetendal EG, Akkermans AD, de Vos WM. Temperature gradient gel elec-trophoresis analysis of 16S rRNA from human fecal samples reveals stable
[
[
unology 19 (2007) 94–105
and host specific communities of active bacteria. Appl Environ Microbiol1998;64:3854–9.
26] Walter J, Hertel C, Tannock GW, Lis CM, Munro K, Hammes WP. Detectionof Lactobacillus, Pediococcus, Leuconostoc, and Weisella species in humanfeces by using group-specific PCR primers and denaturing gradient gelelectrophoresis. Appl Environ Microbiol 2001;67:2578–85.
27] Tannock GW. Probiotics: time for a dose of realism. Curr Issues IntestMicrobiol 2003;4:33–42.
28] Tannock GW. A special fondness for lactobacilli. Appl Environ Microbiol2004;70:3189–94.
29] Cooperstock MS, Zedd AJ. Intestinal flora of infants. In: Hentges DJ, editor.Human intestinal microflora in health and disease. New York: AcademicPress; 1983. p. 79–99.
30] Norin KE, Gustafsson BE, Lindblad BS, Midtvedt T. The establishmentof some microflora associated biochemical characteristics in feces fromchildren during the first years of life. Acta Paediatr Scand 1985;74:207–12.
31] Harmsen HJM, Wildeboer ACM, Raangs GC, Wagendorp AA, Klijn N,Bindels JG, et al. Analysis of intestinal flora development in breast-fedand formula-fed infants by using molecular identification and detectionmethods. J Pediatr Gastroenterol Nutr 2000;30:61–7.
32] Savage DC. Microbial biota of the human intestine: a tribute to some pio-neering scientists. Curr Issues Intest Microbiol 2001;2:1–15.
33] Dubos R, Savage D, Schaedler R, Biological, Freudianism. Lasting effectsof early environmental influences. Pediatrics 1966;38:789–800.
34] Murray CS, Woodcock A. Gut microflora and atopic disease. In: TannockGW, editor. Probiotics and prebiotics: where are we going? Wymondham,United Kingdom: Caister Academic Press; 2006. p. 239–61.
35] Strachan DP. Hay fever, hygiene, and household size. Brit Med J1989;299:1259–60.
36] Sepp E, Julge K, Vasar M, Naaber P, Bjorksten B, Mikelsaar M. Intestinalmicroflora of Estonian and Swedish infants. Acta Pediatr 1997;86:856–961.
37] Ouwehand AC, Isolauri E, He F, Hashimoto H, Benno Y, Salminen S.Differences in Bifidobacterium flora composition in allergic and healthyinfants. J Allergy Clin Immunol 2001;108:144–5.
38] Young SL, Simon MA, Baird MA, Tannock GW, Bibiloni R, Spencely K,et al. Bifidobacterial species differentially affect expression of cell surfacemarkers and cytokines of dendritic cells harvested from cord blood. ClinDiag Lab Immunol 2004;11:686–90.
39] Brandtzaeg P. Current understanding of gastrointestinal immuno-regulationand its relation to food allergy. Ann N Y Acad Sci 2002;964:13–45.
40] Prescott SL, Macaubas C, Holt BJ, Nisini R, Smallacombe TB, Loh R, etal. Transplacental priming of the human immune system to environmentalallergens: universal skewing of initial T cell responses toward the Th2cytokine profile. J Immunol 1998;160:4730–7.
41] Yabuhara A, Macaubas C, Prescott SL, Venaille TJ, Holt BJ, HabreW, et al. TH2-polarized immunological memory to inhalant allergens inatopics is established during infancy and early childhood. Clin Exp Allergy1997;27:1237–9.
42] Rescigno M, Urbano M, Valzasina B, Francolini M, Rotta G, Bonasio R, etal. Dendritic cells express tight junction proteins and penetrate gut epithelialmonolayers to sample bacteria. Nature Immunology 2001;2:361–7.
43] MacDonald TT, Monteleone G. Immunity, inflammation, and allergy in thegut. Science 2005;307:1920–5.
44] Gordon HA, Pesti L. The gnotobiotic animal as a tool in the study of hostmicrobial relationships. Bacteriol Rev 1971;35:390–429.
45] Kimura K, McCartney AL, McConnell MA, Tannock GW. Analysis offecal populations of bifidobacteria and lactobacilli and investigations of theimmunological responses of their human hosts to the predominant strains.Appl Environ Microbiol 1997;63:3394–8.
46] Lievin-Le Moal V, Servin AL. The front line of enteric host defence againstunwelcome intrusion of harmful microorganisms: mucins, antimicrobialpeptides, and microbiota. Clin Microbiol Rev 2006;19:315–37.
47] Laukoetter MG, Bruewer M, Nusrat A. Regulation of the intestinal epithe-lial barrier by the apical junctional complex. Curr Opin Gastroenterol2006;22:85–9.
48] Van der Waaij LA, Limburg PC, Mesander G, van der Waaij D. In vivo IgAcoating of anaerobic bacteria in human faeces. Gut 1996;38:354–84.

Imm
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
G.W. Tannock / Seminars in
49] Salzman NH, Ghosh D, Huttner KM, Paterson Y, Bevins CL. Protectionagainst enteric salmonellosis in transgenic mice expressing a human intesti-nal defensin. Nature 2003;422:522–6.
50] Ruiz PA, Hoffman M, Szcesny S, Blaut M, Haller D. Innate mecha-nisms for Bifidobacterium lactis to activate transient pro-inflammatory hostresponses in intestinal epithelial cells after the colonization of germ-freerats. Immunology 2005;115:441–50.
51] Cario E. Bacterial interactions with cells of the intestinal mucosa: toll-likereceptors and NOD2. Gut 2005;54:1182–93.
52] Haller D. Intestinal epithelial cell signaling and host-derived negativeregulators under chronic inflammation: to be or not to be activated deter-mines the balance towards commensal bacteria. Neurogastroenterol Motil2006;18:184–99.
53] Sartor RB. Microbial influences in inflammatory bowel disease: role inpathogenesis and clinical implications. In: Sartor RB, Sandborn WJ, edi-tors. Kirstner’s inflammatory bowel diseases. London: Elsevier Publishers;2004. p. 138–62.
54] Duchmann R, Kaiser I, Hermann E, Mayet W, Ewe K, Meyer zumBuschenfelde K-H. Tolerance exists towards resident intestinal flora butis broken in active inflammatory bowel disease (IBD). Clin Exp Immunol1995;102:448–55.
55] Bouma G, Strober W. The immunological and genetic basis of inflammatorybowel disease. Nat Rev Immunol 2003;3:521–33.
56] Podolsky DK. Inflammatory bowel disease. N Engl J Med 2002;347:417–28.
57] Van der Waaij L, Harmsen HJM, Madjipour M, Kroese FGM, Zwiers M,van Dullemen HM, et al. Bacterial population analysis of human colon andterminal ileum biopsies with 16S rRNA-based fluorescent probes: commen-sal bacteria live in suspension and have no direct contact with epithelialcells. Inflamm Bowel Dis 2005;11:865–71.
58] Martin HM, Campbell BJ, Hart CA, Mpofu C, Nayar M, Singh R, et al.Enhanced Escherichia coli adherence and invasion in Crohn’s disease andcolon cancer. Gastroenterology 2004;127:80–93.
59] Fujita H, Eishi Y, Ishige I, Saitoh K, Takizawa T, Arima T, et al. Quantitativeanalysis of bacterial DNA from Mycobacteria spp., Bacteroides vulgatus,and Escherichia coli in samples from patients with inflammatory boweldiseases. J Gastroenterol 2002;37:509–16.
60] Swidsinski A, Ladhoff A, Pernthaler A, Swidsinski S, Loening-Baucke V,Ortner M, et al. Mucosal flora in inflammatory bowel disease. Gastroen-terology 2002;122:44–54.
61] Hudson MJ, Hill MJ, Elliot PR, Berghouse LM, Burnham WR, Lennard-Jones JE. The microbial flora of the rectal mucosa and faeces of patientswith Crohn’s disease before and during antimicrobial chemotherapy. J MedMicrobiol 1984;18:335–45.
62] Lepage P, Seksik P, Sutren M, de al Cochetiere M-F, Jian R, Marteau P,et al. Biodiversity of the mucosa-associated microbiota is stable along thedistal digestive tract in healthy individuals and patients with IBD. InflammBowel Dis 2005;11:473–80.
63] Macfarlane S, Furrie E, Cummings JH, Macfarlane GT. Chemotaxonomicanalysis of bacterial populations colonizing the rectal mucosa in patientswith ulcerative colitis. Clin Infect Dis 2004;38:1690–9.
64] Mylonaki M, Rayment NB, Rampton DS, Hudspith BN, Brostoff J. Molec-ular characterization of rectal mucosa-associated bacterial flora in inflam-matory bowel disease. Inflamm Bowel Dis 2005;11:481–7.
65] Kleeson B, Kroesen AJ, Buhr HJ, Blaut M. Mucosal and invading bacteriain patients with inflammatory bowel disease compared with controls. ScandJ Gastroenterol 2002;37:1034–41.
66] Tiveljung A, Soderholm JD, Olaison G, Jonasson J, Monstein H-J. Presenceof eubacteria in biopsies from Crohn’s disease inflammatory lesions as
determined by 16S rRNA gene-based PCR. J Med Microbiol 1999;48:263–8.67] Zinkevich V, Beech IB. Screening of sulfate-reducing bacteria incolonoscopy samples from healthy and colitic human gut mucosa. FEMSMicrobiol Ecol 2000;34:147–55.
[
unology 19 (2007) 94–105 105
68] Ott SJ, Musfeldt M, Wenderoth DF, Hampe J, Brant O, Folsch UR, et al.Reduction in diversity of the colonic mucosa associated bacterial microflorain patients with active inflammatory bowel disease. Gut 2004;53:685–93.
69] Schultsz C, van den Berg F, Ten Kate FW, Tytgat NJ, Dankert J. Theintestinal mucus layer from patients with inflammatory bowel disease har-bors high numbers of bacteria compared with controls. Gastroenterology1999;117:1089–97.
70] Prindiville T, Cantrell M, Wilson KH. Ribosomal DNA sequence analy-sis of mucosa-associated bacteria in Crohn’s disease. Inflamm Bowel Dis2004;10:824–33.
71] Darfeuille-Michaud A, Boudeau J, Bulois P, Neut C, Glasser A-L, BarnichN, et al. High prevalence of adherent-invasive Escherichia coli associ-ated with ileal mucosa in Crohn’s disease. Gastroentrology 2004;127:412–21.
72] Romero C, Hamdi A, Valentine JF, Naser SA. Evauation of surgical tissuefrom patients with Crohn’s disease for the presence of Mycobacteriumavium subspecies paratuberculosis DNA by in situ hybridization and nestedpolymerase chain reaction. Inflamm Bowel Dis 2005;11:116–25.
73] Ryan P, Bennett MW, Aarons S, Lee G, Collins JK, O’Sullivan GC, etal. PCR detection of Mycobacterium paratuberculosis in Crohn’s diseasegranulomas isolated by laser capture microdissection. Gut 2002;51:665–70.
74] Ryan P, Kelly RG, Lee G, Collins JK, O’Sullivan GC, O’ConnellJ, et al. Bacterial DNA within granulomas of patients with Crohn’sdisease—detection by laser capture microdissection and PCR. Am J Gas-troenterol 2004;99:1539–43.
75] Lucke K, Miehlke S, Jacobs E, Schuppler M. Prevalence of Bacteroidesand Prevotella spp. in ulcerative colitis. J Med Microbiol 2006;55:617–24.
76] Conte MP, Schippa S, Zamboni I, Penta M, Chiarini F, Seganti L, Osborn J,Falconieri P, Borrelli O, Cucchiara S. Gut-associated bacterial microbiotain paediatric patients with inflammatory bowel disease. Gut 2006; Epub.
77] Bibiloni R, Mangold M, Madsen KL, Fedorak RN, Tannock GW. The bac-teriology of biopsies differs between newly diagnosed, untreated. Crohn’sdisease and ulcerative colitis patients. J Med Microbiol 2006;55:1141–9.
78] Ruseler-van Embden JGH, Both-Patoir HC. Anaerobic Gram-negative fae-cal flora in patients with Crohn’s disease and healthy subjects. Anton vanLeeuwen 1983;49:125–32.
79] Manichanh C, Rigottier-Gois L, Bonnaud E, Gloux K, Pelletier E, FrangeulL, et al. Reduced diversity of faecal microbiota in Crohn’s disease revealedby a metagenomic approach. Gut 2005;55:205–11.
80] Seksik P, Lepage P, de la Cochetiere M-F, Bourreille A, Sutren M, GalmicheJ-P, et al. Search for localized dysbiosis in Crohn’s disease ulcerations bytemporal temperature gradient gel electrophoresis of 16S rRNA. J ClinMicrobiol 2005;43:4654–8.
81] Seksik P, Rigottier-Gois L, Gramet G, Sutren M, Pochart P, Marteau P,et al. Alterations of the dominant faecal bacterial groups in patients withCrohn’s disease of the colon. Gut 2003;52:237–42.
82] Handelsman J. Metagenomics: application of genomics to unculturedmicroorganisms. Microbiol Mol Biol Rev 2004;68:669–85.
83] Backhed F, Ding H, Wang T, Hooper LV, Koh GY, Nagy A, et al. The gutmicrobiota as an environmental factor that regulates fat storage. Proc NatlAcad Sci USA 2004;101:15718–23.
84] Hooper LV, Wong MH, Thelin A, Hansson L, Falk PG, Gordon JI. Molec-ular analysis of commensal host-microbial relationships in the intestine.Science 2001;291:881–4.
85] Stappenbeck TS, Hooper LV, Gordon JI. Developmental regulation ofintestinal angiogenesis by indigenous microbes via Paneth cells. Proc NatlAcad Sci 2002;99:15451–5.
86] Boudry G, Peron V, Le Huerou-Luron I, Lalles JP, Seve B. Weaning induces
both transient and long-lasting modifications of absorptive, secretory, andbarrier properties of piglet intestine. J Nutr 2004;134:2256–62.87] Prescott SL, Holt PG. Abnormalities in cord blood mononuclear cytokineproduction as a predictor of later atopic disease in childhood. Clin ExpAllergy 1998;28:1313–6.