Embed Size (px)
Citation preview

Inhibitory Roles of Prohibitin and Chemerin inFSH-Induced Rat Granulosa Cell Steroidogenesis
Qi Wang, Arthur Leader, and Benjamin K. Tsang
Departments of Cellular and Molecular Medicine (Q.W., B.K.T.) and Obstetrics and Gynaecology(A.L., B.K.T.), University of Ottawa, and Chronic Disease Program (Q.W., A.L., B.K.T.), Ottawa HospitalResearch Institute, Ottawa, Ontario, Canada K1H 8L6; Ottawa Fertility Centre (A.L.), Ottawa, Ontario,Canada K2C 3V4; and World Class University Biomodulation Major (B.K.T.), Department of AgriculturalBiotechnology, College of Agriculture and Life Sciences, Seoul National University, Seoul 151-742,Republic of Korea
Follicular differentiation is a tightly regulated process involving various endocrine, autocrine, andparacrine factors. The biosynthesis of progesterone and estradiol in response to FSH involves theregulation of multiple steroidogenic enzymes, such as p450 cholesterol side-chain cleavage enzymeand aromatase. Here we demonstrated that prohibitin (PHB), a multifunctional protein, inhibitsFSH-induced progesterone and estradiol secretion in rat granulosa cells. The mRNA abundances ofcyp11a (coding p450 cholesterol side-chain cleavage enzyme) and cyp19 (coding aromatase) werealso suppressed by PHB in a time-dependent manner. It is known that a novel adipokine chemerinsuppresses FSH-induced steroidogenesis in granulosa cells. Chemerin up-regulates the content ofPHB, and PHB knockdown attenuates the suppressive role of chemerin on steroidogenesis. Inaddition, inhibition of phosphatidylinositol 3-kinase/Akt pathway enhances the suppressive actionof PHB, whereas expression of constitutively active Akt attenuates this response. These findingssuggest that PHB is a novel negative regulator of FSH-induced steroidogenesis, and its action withchemerin may contribute to the dysregulation of steroidogenesis in the pathogenesis of polycysticovarian syndrome. (Endocrinology 154: 956–967, 2013)
Follicular growth and differentiation are tightly regu-lated by endocrine, autocrine, and paracrine factors
(1). The coordinated biosynthesis of steroids in the ovaryis critical for reproductive cycle, successful ovulation, andeventual pregnancy (2). FSH-induced synthesis of proges-terone and estradiol involves the regulation of varioussteroidogenic enzymes, such as steroidogenic acute reg-ulatory protein (StAR), p450 cholesterol side-chaincleavage enzyme (p450scc), hydroxysteroid dehydro-genase-3� (HSD-3�), and aromatase (3–5). p450scc isthe first and rate-limiting step in the steroidogenesispathway in human granulosa cells, and aromatase con-verts androstenedione and testosterone to estrone andestradiol, respectively (6, 7).
Chemerin is a chemoattractant and a recently discov-ered adipokine associated with obesity and the metabolic
syndrome in women (8, 9). Chemerin is present in humanplasma in an inactive state, and its active form is releasedby the proteolytic cleavage of the C terminus (9–11). Inaddition to promoting lipogenesis, adipogenesis, and an-giogenesis (12–14), chemerin has been reported to act asa pro- as well as antiinflammatory factor, which regulatesimmunity (9, 15–17). Serum chemerin levels are higher inobese women and in polycystic ovary syndrome (PCOS)subjects, and metformin decreases the serum chemerin level(8, 18), implying an association between chemerin andPCOS. We have previously reported that chemerin sup-pressed FSH-induced steroidogenesis in granulosa cells viadown-regulation of the expression of nuclear receptorsNR5a1/2 (19). However, the mediators and mechanism by
ISSN Print 0013-7227 ISSN Online 1945-7170Printed in U.S.A.Copyright © 2013 by The Endocrine Societydoi: 10.1210/en.2012-1836 Received August 10, 2012. Accepted November 28, 2012.First Published Online December 18, 2012
Abbreviations: A-Akt, active Akt; CTL, control; DES, diethylstilbestrol; DHT, dihydrotes-tosterone; DMSO, dimethylsulfoxide; DN, dominant negative; eCG, equine chronic go-nadotropin; EIA, enzyme immunoassay; GAPDH, glyceraldehyde-3-phosphate dehydro-genase; HA, hemagglutinin; HSD-3�, hydroxysteroid dehydrogenase-3�; LY, LY294002;MEK, MAPK kinase; MOI, multiplicity of infection; PCOS, polycystic ovary syndrome; PHB,prohibitin; PI3K, phosphatidylinositol 3-kinase; PKA, protein kinase A; p450scc, p450 cho-lesterol side-chain cleavage enzyme; StAR, steroidogenic acute regulatory protein; TBST,Tween-20 in Tris and NaCl (pH 7.4) (Tris buffered saline); TCN, triciribine.
R E P R O D U C T I O N - D E V E L O P M E N T
956 endo.endojournals.org Endocrinology, February 2013, 154(2):956–967

which chemerin suppresses FSH-induced steroidogenesis re-main unknown.
Prohibitin (PHB) is a multifunctional protein associ-ated with many cellular processes such as cell cycle, pro-liferation, apoptosis, senescence, cellular immortaliza-tion, and differentiation (20-23). Recently PHB has beenlinked to adipogenesis and obesity because its expressionis up-regulated in high-fat diet-induced obese mice (24),and knockdown of PHB significantly reduces the expres-sion of adipogenic markers and the accumulation of lipids(25). In the ovary, PHB is widely expressed and its expres-sion is age and follicular stage regulated (26, 27). Al-though it is antiapoptotic in granulosa cells (28) and si-lencing of PHB induces granulosa cell shape changes (29),our knowledge on the role and contribution of PHB togranulosa cell steroidogenesis is incomplete.
In this study, we first examined the expression of PHBand chemerin and their regulation by gonadotropins invitro and in vivo. We also investigated the role of PHB onFSH-induced steroidogenesis in granulosa cells. We in-tended to demonstrate that PHB is antidifferentiativeduring follicular development and that it mediates the sup-pressive action of chemerin on FSH-induced steroidogen-esis, which would suggest that PHB, together withchemerin, may contribute to the dysregulation of steroidsproduction in the pathogenesis of ovarian diseases, such aspolycystic ovaries.
Materials and Methods
Antibodies and reagentsCell culture media (M199), fetal bovine serum, penicillin and
streptomycin, L-glutamine, sodium pyruvate, and trypsin werepurchased from Invitrogen (Burlington, Canada). HEPES andequine chronic gonadotropin (eCG) were purchased from Sigma(St Louis, MO). Recombinant mouse chemerin was obtainedfrom R&D Systems (Minneapolis, MN) and recombinant hu-man FSH was purchased from National Hormone and PeptideProgram (Harbor-UCLA Medical Center, Torrance, CA). Anti-aromatase antibody was purchased from AbD Serotec (Oxford,UK), anti-p450scc antibody from United States Biological(Swampscott, MA), anti-PHB and anti-�-actin antibodies fromAbcam (Cambridge, MA), and antihemagglutinin (HA) anti-body from Roche Applied Science (Québec, Canada). Horserad-ish peroxidase-conjugated secondary antibodies and reagents forSDS-PAGE were supplied by Bio-Rad (Mississauga, Canada).Enhanced chemiluminescent reagent was from Thermo FisherScientific (Rockford, IL). QIAShredder and RNeasy minikitwere purchased from QIAGEN (Mississauga, Canada). Randomdecamer primers were from Ambion (Austin, TX). Ribonucleaseinhibitor and deoxynucleotide triphosphate were from Fermen-tas (Burlington, Canada). Moloney murine leukemia virus re-verse transcriptase was from Promega (Madison, WI). Quanti-Tect SYBR Green PCR kit and phosphatase/protease inhibitors
were from Roche Applied Science. The PCR primers were fromInvitrogen. Adenoviral-PHB and adenoviral-short hairpin RNAtargeting PHB were obtained from Dr. Winston Thompson(Morehouse School of Medicine, Atlanta, GA). Dominant neg-ative (DN) Akt and constitutively active Akt (A-Akt) adenovi-ruses were generous gifts from Dr. Kenneth Walsh (Boston Uni-versity School of Medicine, Boston, MA). All chemical inhibitorswere purchased from Calbiochem (Gibbstown, NJ). All otherchemicals were of the highest analytical grade available fromSigma.
Animal preparationSprague Dawley rats (Charles River, Montréal, Canada) were
maintained on 12-h light, 12-h dark cycles and given food andwater ad libitum. All procedures were carried out in accordancewith the Guidelines for the Care and Use of Laboratory Animals,Canadian Council on Animal Care, and were approved by theUniversity of Ottawa and the Ottawa Hospital Research Insti-tute Animal Care Committee.
Primary culture of rat granulosa cells andadenoviral infection
Ovaries from diethylstilbestrol (DES)-primed immature rats(d 21, 1 mg/d, sc, for 3 consecutive days) were preincubated with6 mM EGTA and 0.5 M sucrose (30), and granulosa cells werereleased by follicular puncture with a 26.5-gauge needle,washed, and centrifuged (900 � g, 5 min). Cell clumps andoocytes were removed by filtering the cell suspensions through a40-�m nylon cell strainer (BD Biosciences, Mississauga, On-tario, Canada). The number of viable granulosa cells was deter-mined by trypan blue exclusion. Granulosa cells (0.9 � 106 perwell in 6 well plate) were plated overnight in M199 with 10%fetal bovine serum under a humidified atmosphere of 95% airand 5% CO2. After culture overnight in serum-free medium,granulosa cells were treated with FSH (0–100 ng/mL) orchemerin (0-100 ng/mL) for a designated duration. Unless spec-ified, granulosa cells used in the present study were collectedfrom large preantral and early antral follicles of DES-treated rats.
To harvest granulosa cells from follicles at different stages ofdevelopment, 21-d-old immature rats were injected (sc) witheCG (10 IU), and granulosa cells were collected from the ovariesprimed with eCG at 0 h (predominantly preantral and early an-tral follicles), 24 h (predominantly early antral follicles), or 48 h(predominantly large antral and preovulatory follicles), as pre-viously described (31).
For adenoviral infection, granulosa cells were cultured in se-rum-free M199 medium containing adenoviral particles for 24 hfollowed by medium change. Multiplicity of infection (MOI) andduration of infection are detailed in the figures. Equal amountsof adenovirus in each experimental group were achieved by theadjustment with an appropriate amount of adenoviral-LacZ(negative control for adenoviral-PHB, DN-Akt, and A-Akt) oradenoviral-shNeg (negative control for shPHB).
Preparation of dihydrotestosterone (DHT)-treatedrats
The DHT-filled capsules were prepared as previously de-scribed (32). In brief, SILASTIC brand tubing (Dow Corning Inc,Midland, MI; inner diameter 1.98 mm � outer diameter 3.18mm), cut to an appropriate length to achieve a desired surface
Endocrinology, February 2013, 154(2):956–967 endo.endojournals.org 957

area of 300 mm2, was filled with DHT powder (Steraloids Inc,Newport) from a 1-mL syringe. The tubing was closed at eachend (3 mm) with a sealant (Silicone type A; Dow Corning), en-suring that adhesive is in contact with tubing walls and that thereare no air bubbles. Control (CTL) capsules were empty withsealant on both ends. After being left overnight to dry, the cap-sules were rinsed for 2 days in 3% BSA in PBS with 0.1% NaN3
solution, washed with PBS, and sterilized by dipping briefly in70% ethanol before use.
Female rats at 21 days of age were divided into two groups(DHT vs. CTL) and implanted with a DHT-filled silicone cap-sules continuous-releasing (83 �g per day, empty SILASTICbrand capsule as control; Dow Corning) for 12 weeks to mimicthe hyperandrogenic state in women with PCOS, whose plasmaDHT levels are approximately 1.7-fold higher than those ofhealthy control (33, 34). Rats were monitored twice daily in thefirst 3 d and once daily thereafter. Animals were weighed weeklyto monitor weight gain and euthanized at 12 weeks afterimplantation.
Isolation of granulosa cells from CTL or DHT ratsRat ovaries were collected after rat euthanization and gran-
ulosa cells from CTL or DHT rats were recovered in a similar butmore precise method than that mentioned above. Granulosa cellsfrom preantral/early antral follicle stages in the ovary were re-leased by follicular puncture, but the preovulatory follicles andcorpora lutea were spared. A pool of granulosa cells collectedfrom 5 rats of each group was used for mRNA and proteinanalysis.
Reverse transcription-polymerase chain reactionTotal RNAs of granulosa cells were extracted according to
the manufacturer’s instruction, using the QIAGEN RNeasyminikit. Then 0.2 �g total RNAs was used to reverse transcribecDNAs, and the mRNA abundance of target genes was analyzedby real-time PCR and normalized to glyceraldehyde-3-phos-phate dehydrogenase (GAPDH). Specific primer pairs used in theexperiments are listed in Table 1. Data were analyzed by the2���CT method (35).
Protein extraction and Western blotTotal protein extracts were prepared by adding 100 �l hot
(100°C) Tris buffer (10 mM Tris, pH 7.4; 1% sodium dodecylsulfate; 1 mM sodium orthovanadate) to each well. The extractswere obtained by rapidly scraping, transferred to a microcentri-fuge tube, and boiled for 5 minutes. Protein concentrations ineach sample were determined by a Bradford assay (Bio-Rad Lab-oratories). Twenty-microgram cell lysates were subjected toSDS-PAGE with 4.5% stacking and 10% separating gels. Pro-teins were electrophoretically transferred to nitrocellular mem-brane (Bio-Rad Laboratories), blocked at room temperaturewith 5% skim milk in TBST [0.05% Tween-20 in 10 mM Tris,0.15 M NaCl (pH 7.4) (Tris buffered saline)] for 1 h, and thenincubated overnight at 4°C with diluted primary antibodies (1:1000 to 1:10,000 according to the manufacturer’s instruction) inTBST with constant agitation. The membranes were then treatedwith a secondary antibody (1:5000 to 1:50,000 based on differ-ent primary antibody). After washing 3 times with TBST, im-munoreactive bands were visualized with an enhanced chemilu-minescent agent according to the manufacturer’s instruction.Intensity of bands of the exposed X-ray film was determined bydensitometrically scanning, quantified using AlphaEaseFC (Al-pha Innotech, San Leandro, CA), and normalized with �-actin.
Steroids secretion analysisSpent medium of cultured granulosa cells were collected, cen-
trifuged (900 � g, 5 min) and kept in �80°C for hormone anal-ysis. 17�-Estradiol and progesterone concentrations in spent me-dium were measured using enzyme immunoassay (EIA) kit (EnzoLife Sciences, Farmingdale, NY) according to the manufacturer’sinstruction. The detection limitation of 17�-estradiol was 28pg/mL, and the intra- and interassay coefficients of variationwere 8% and 6%, respectively. The detection limitation of pro-gesterone was 8.5 pg/mL, and the intra- and interassay coeffi-cients of variation were 7% and 6%, respectively.
Statistical analysisAll data were analyzed using GraphPad Prism 5.0 statistical
software (San Diego, CA). Results are expressed as mean � SEMof at least 3 independent experiments as detailed in the figures.Unpaired t test was used for comparing the mRNA abundance of
TABLE 1. Primer pairs used for quantitative real-time PCR
Gene Primer Sequence (5�–3�) Accession no.
Cyp19 Forward GGCATGCACGAGAATGGCATCATA NM_017085Reverse CAGCCTGTCCAAATGCTGCTTGAT
Cyp11a Forward ACACGACCTCCATGACTCTGCAAT NM_017286Reverse TCAGTGTCTCCTTGATGCTGGCTT
StAR Forward TGTTAAGGACTGCCCACCACATCT NM_031558Reverse TGTCCTTGGCTGAAGGTGAACAGA
HSD-3� Forward AGATCTGGGCTATGTGCCACTTGT NM_001007719Reverse ACCTGGTAACACCCAGAACCACAT
PHB Forward ACCGATTCTGTGGCATACAGGACA XM_001053030Reverse TGGTCGAGAGCAGCAGTCAAAGAT
Chemerin Forward GGCACCTTTGTGAGGCTGGAATTT NM_001013427Reverse ACCCTGTCCAGGGCTTATTTGGAT
GAPDH Forward TGACTCTACCCACGGCAAGTTCAA NM_017008Reverse ACGACATACTCAGCACCAGCATCA
958 Wang et al Prohibitin and Chemerin Suppress Steroidogenesis Endocrinology, February 2013, 154(2):956–967

target genes in granulosa cells treated with FSH or mRNA/pro-tein level of targets in granulosa from CTL/DHT rats. One-wayand two-way ANOVA were used to assess the effects and inter-actions of 1 or 2 variables, and multiple comparisons wereachieved by a Bonferroni post hoc test. Significant difference wasdefined at P � .05 (* or #).
Results
Gonadotropin down-regulates the mRNAabundance of chemerin and PHB in granulosa cells
It has been reported that chemerin and PHB are ex-pressed in ovarian cells (9, 27). To determine whether theirexpression is dependent on the stage of follicular devel-opment, granulosa cells were collected from the ovariesprimed with eCG at 0 h (predominantly preantral andearly antral follicles), 24 h (predominantly early antralfollicles), or 48 h (predominantly large antral and preovu-latory follicles), and mRNA abundance of chemerin andPHB were analyzed by real-time PCR. As shown in Figure1A, mRNA expression of both chemerin (P � .001) andPHB (P � .001) in granulosa cells significantly decreasedat late stages of follicular development (gonadotropin 24and 48 h).
We also examined whether FSH down-regulates theexpression of chemerin and PHB in vitro. Figure 1B indi-cated that both chemerin (P � .001) and PHB (P � .01)mRNA levels were decreased by FSH. Because gonado-
tropin similarly regulates the expression of chemerin andPHB, we then examined whether chemerin regulates PHBexpression. As shown in Figure 1C, treatment withchemerin significantly increased PHB content in granulosacells (P � .05).
Exogenous PHB suppressed FSH-induced estradioland progesterone secretion
Our previous results indicated that chemerin sup-pressed FSH-induced granulosa cell steroidogenesis (19).Because chemerin up-regulates PHB content in granulosacells, it is possible that the suppressive role of chemerinmay be mediated by PHB. To test this possibility, we firstexamined whether PHB overexpression suppresses FSH-induced progesterone and estradiol production. As ob-served in Figure 2A, FSH markedly stimulated progester-one secretion in granulosa cells, a response significantlysuppressed by exogenous PHB (FSH, P � .001; PHB, P �
.05; FSH � PHB, P � .05). Similarly, PHB also suppressedFSH-induced estradiol secretion (FSH, P � .001; PHB, P�
.01; FSH � PHB, P � .01). In addition, the increase of aro-matase and p450scc contents induced by FSH was sup-pressed by exogenous PHB (Figure 2B; for p450scc: FSH,P � .001; PHB, P � .001; FSH � PHB, P � .001; for aro-matase: FSH,P� .001;PHB,P� .001;FSH�PHB,P� .01).
We then examined whether PHB knockdown attenu-ates the suppression of steroidogenesis by chemerin. Gran-
Figure 1. Expression of chemerin and PHB and their regulation by gonadotropin. A, Granulosa cells were isolated from follicles at different stagesof development in immature rats injected with eCG (0 h, preantral; 24 h, early antral; 48 h, late antral and preovulatory follicles). The mRNAabundances of chemerin and PHB were analyzed by real-time PCR and normalized to the GAPDH mRNA. B, Granulosa cells from DES-treated ratswere cultured with FSH (100 ng/mL) for designated time point (6 and 24 h), and the mRNA abundance of chemerin and PHB was analyzed by real-time PCR and normalized to GAPDH mRNA. C, Granulosa cells from DES-treated rats were treated with chemerin (0–100 ng/mL, 24 h), and thecontent of PHB was assessed by Western blot. Shown are representative immunoblots and data are presented as mean � SEM of threeindependent experiments. A and C, One-way ANOVA followed by Bonferroni post hoc test; B, unpaired t test. *, P � .05; **, P � .01;***, P � .001 compared with eCG at 0 h or untreated control (FSH � 0 or chemerin � 0).
Endocrinology, February 2013, 154(2):956–967 endo.endojournals.org 959

ulosa cells were infected with adenoviral-shPHB (MOI �10, shNeg as control) to knock down PHB and thencultured with chemerin (0–100 ng/mL) in the presence ofFSH (100 ng/mL, 24 h). As shown in Figure 2C, the sup-pression of p450scc and aromatase expression bychemerin was attenuated by PHB knockdown (forp450scc: chemerin, P � .05; shPHB, P � .05; chemerin �
shPHB, P � .05; for aromatase:chemerin, P � .05; shPHB, P � .05;chemerin � shPHB, P � .05).
PHB suppressed FSH-inducedsteroidogenesis in atime-dependent manner ingranulosa cells
Next we examined whether the sup-pression of FSH-induced steroidogene-sis by PHB is time dependent andwhether steroidogenic enzymes expres-sion is regulated at the transcriptionallevel. As shown in Figure 3A, FSH grad-ually increased estradiol production,starting at 6 h and reaching significanceafter 18 h. Exogenous PHB suppressedFSH-induced estradiol production in atime-dependent manner (PHB, P �.001; time, P � .0001; PHB � time, P �.0001). PHB had no effect on FSH-in-duced estradiol secretion at an earliertime point but showed inhibition actionat 18 and 24 h. The mRNA abundanceof cyp19 rapidly increased 50–350 fold(3–12 h) and reached significance at18 h with an elevated level of about2200-fold in the presence of FSH (Fig-ure 3A). The effect of PHB on cyp19expression showed a similar trend asthat of estradiol production, with dra-matically down-regulation at 18 and24 h (PHB, P � .001; time, P � .001;PHB � time, P � .001).
In contrast to FSH-stimulated estra-diol secretion, FSH rapidly increasedprogesterone secretion at 3 h and main-tained a higher level until 12 h and thena greater increase in progesterone pro-duction at 18 and 24 h (Figure 3B). Al-though PHB had minimum or no effecton progesterone levels at an early timepoint (0-18 h), it was inhibitory at 24 h(PHB, P � .05; time, P � .001; PHB �time, P � .05). Although progesteroneproductionquicklyand significantly in-
creased at 3 h in response to FSH, there was only a slightlyincrease of cyp11a expression until 12 h (Figure 3B,3.03 � 0.24-fold at 3 h and 7.76 � 1.01-fold at 12 h vs.1.03 � 0.03 at 0 h). Obvious elevation of cyp11a ex-pression induced by FSH occurred at 18 h (51.66 �5.54-folds) and 24 h (60.99 � 5.45-fold), which was
Figure 2. Role of PHB in FSH-induced, chemerin-mediated steroidogenesis. A, Granulosacells were infected with adenoviral-PHB (MOI � 40, adenoviral-LacZ as control) for 24 h andthen cultured with FSH (100 ng/mL, 24 h). Testosterone (0.5 �M) was added to serve assubstrate of aromatase during culture. Progesterone and estradiol secreted into the mediumwas measured by EIA. Data are presented as mean � SEM of 4 independent experiments,analyzed by 2-way ANOVA and subsequently by Bonferroni post hoc test. #, P � .05, ##, P �.01 compared with LacZ. B, Granulosa cells were infected with adenoviral-PHB (MOI � 40,adenoviral-LacZ as control) for 24 h and then cultured with FSH (100 ng/mL, 24 h). Thecontents of p450scc and aromatase were examined by Western blot. Shown arerepresentative immunoblots. Data are presented as mean � SEM of 4 independentexperiments and analyzed by 2-way ANOVA and subsequently by Bonferroni post hoc test.###, P � .001 compared with LacZ. C, Granulosa cells were infected with adenoviral-shPHB(MOI � 10, adenoviral-shNeg as control) for 48 h and then treated with chemerin (0-100 ng/mL, 24 h) in the presence of FSH (100 ng/mL). The contents of p450scc and aromatase wereexamined by Western blot. Data are presented as mean � SEM of 4 independent experimentsand analyzed by 2-way ANOVA and subsequently by Bonferroni post hoc test. *, P � .05compared with control (chemerin � 0); #, P � .05 compared with shNeg. ns, Not significant
960 Wang et al Prohibitin and Chemerin Suppress Steroidogenesis Endocrinology, February 2013, 154(2):956–967

significantly suppressed by PHB (PHB, P � .001; time,P � .0001; PHB � time, P � .0001). However, PHB hadno significant effect on FSH-induced StAR and HSD-3�
expression (Supplemental Figure 1, published on TheEndocrine Society’s Journals Online web site athttp://endo.endojournals.org).
Phosphatidylinositol 3-kinase (PI3K)/Akt pathwaynegatively regulates the expression of PHB
Akt activation is an essential for granulosa cell differ-entiation (36–38). We therefore determined whether thePI3K/Akt pathway modulates the expression and functionof PHB. A specific inhibitor of PI3K [LY294002 (LY), 10
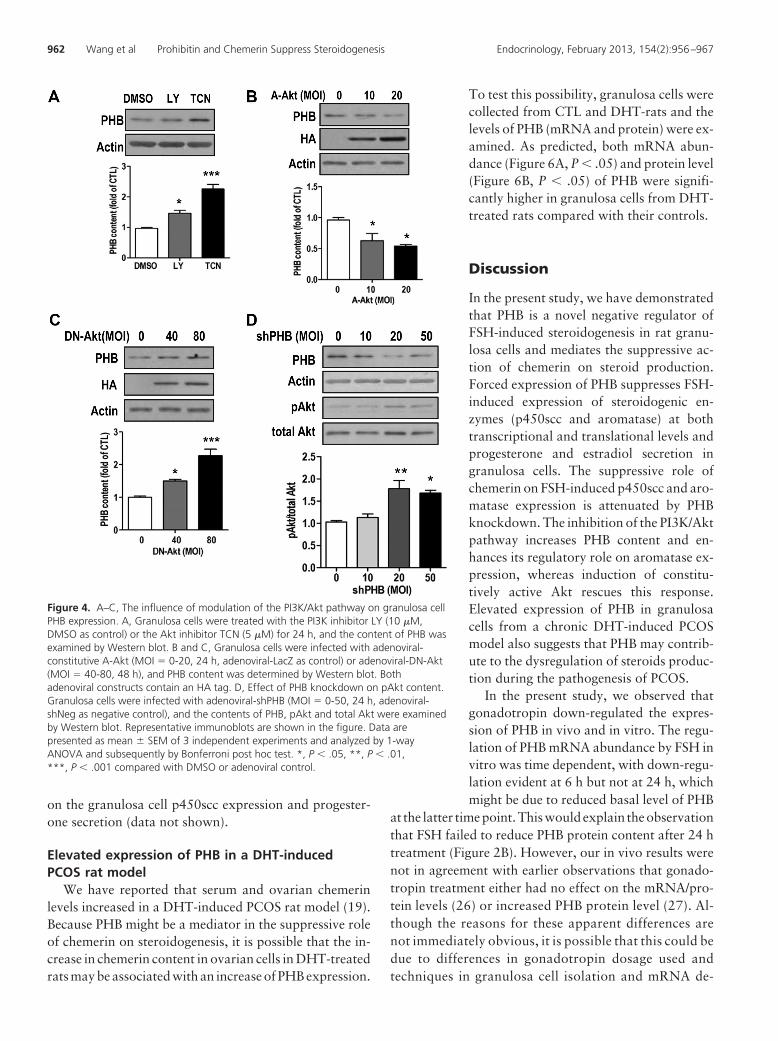
�M; dimethylsulfoxide (DMSO) as control] and Akt[triciribine (TCN), 5 �M; DMSO as control] was added tothe granulosa cell cultures, and Western blot resultsindicated that inhibition of the PI3K/Akt pathway sig-nificantly increased the expression of PHB (Figure 4A,P � .001).
We further examined whether activation of Akt affectsthe level of PHB using adenoviral constitutively A-Akt(containing a HA tag) and adenoviral DN-Akt. As shownin Figure 4, B and C, PHB content was down-regulated bythe activation of Akt (P � .05) and up-regulated when Aktfunction was down-regulated by DN-Akt (P � .001).
PHB knockdown up-regulates Akt phosphorylationWe next examined whether PHB regulates Akt content
and phosphorylation in granulosa cells. Granulosa cellswere infected with adenoviral-shPHB, and the contents ofphosphorylated and total Akt were examined. As shownin Figure 4D, knockdown of PHB significantly increasedthe phosphorylated Akt levels (P � .01) at higher doses(MOI � 20 and 50).
Inhibition of PI3K/Akt pathway enhanced thesuppressive role of PHB on steroidogenesis
Next we assessed whether modulation of PI3K/Aktpathway alters the steroidogenic response to PHB. Asshown in Figure 5A, PHB alone suppressed FSH-inducedaromatase expression (22%) and estradiol secretion(31%). The effect of PHB was enhanced in the presence ofLY, exhibiting approximate 75% suppression of aroma-tase expression and 82% inhibition of estradiol secretion.Although pretreatment of LY alone suppressed FSH-in-duced steroidogenesis, there was significant interactionbetween PHB and LY treatment (treatment, P � .001;PHB, P � .001; PHB � treatment, P � .01). PHB was lesseffective on TCN treatment group because the Akt inhib-itor alone blocked greater than 95% steroidogenesis in-duced by FSH. These inhibitors had similar effect onp450scc expression and secretion of progesterone (datanot shown).
Activation of PI3K/Akt pathway attenuated thesuppressive role of PHB in steroidogenesis
In addition, the experiments using adenoviral A-Aktwere also performed to examine whether the expression ofconstitutively active Akt attenuates the action of PHB onFSH-induced steroidogenesis. Figure 5B indicated thatforced expression of the A-Akt in granulosa cells prior totreatment with FSH and LY significantly attenuated thePHB-induced down-regulation of aromatase expressionand estradiol secretion (A-Akt, P � .05; PHB, P � .001;PHB � A-Akt, P � .01). These inhibitors had similar effect
Figure 3. Influence of PHB on FSH-induced steroid production andsteroidogenic enzyme expression. Granulosa cells were infected withadenoviral-PHB (MOI � 40, adenoviral-LacZ as control) for 24 h andthen cultured with FSH (100 ng/mL) at a designated duration (0–24 h).Testosterone (0.5 �M) was added to serve as substrate of aromataseduring culture. Estradiol (A) and progesterone (B) secreted into themedium were measured by EIA. The mRNA abundance of cyp19 (A)and cyp11a (B) was determined by real-time PCR. Data are presentedas mean � SEM of 4 independent experiments and analyzed by 2-wayANOVA and subsequently by Bonferroni post hoc test. #, P � .05,##, P � .01, ###, P � .001 compared with LacZ
Endocrinology, February 2013, 154(2):956–967 endo.endojournals.org 961
on the granulosa cell p450scc expression and progester-one secretion (data not shown).
Elevated expression of PHB in a DHT-inducedPCOS rat model
We have reported that serum and ovarian chemerinlevels increased in a DHT-induced PCOS rat model (19).Because PHB might be a mediator in the suppressive roleof chemerin on steroidogenesis, it is possible that the in-crease in chemerin content in ovarian cells in DHT-treatedrats may be associated with an increase of PHB expression.
To test this possibility, granulosa cells werecollected from CTL and DHT-rats and thelevels of PHB (mRNA and protein) were ex-amined. As predicted, both mRNA abun-dance (Figure 6A, P � .05) and protein level(Figure 6B, P � .05) of PHB were signifi-cantly higher in granulosa cells from DHT-treated rats compared with their controls.
Discussion
In the present study, we have demonstratedthat PHB is a novel negative regulator ofFSH-induced steroidogenesis in rat granu-losa cells and mediates the suppressive ac-tion of chemerin on steroid production.Forced expression of PHB suppresses FSH-induced expression of steroidogenic en-zymes (p450scc and aromatase) at bothtranscriptional and translational levels andprogesterone and estradiol secretion ingranulosa cells. The suppressive role ofchemerin on FSH-induced p450scc and aro-matase expression is attenuated by PHBknockdown. The inhibition of the PI3K/Aktpathway increases PHB content and en-hances its regulatory role on aromatase ex-pression, whereas induction of constitu-tively active Akt rescues this response.Elevated expression of PHB in granulosacells from a chronic DHT-induced PCOSmodel also suggests that PHB may contrib-ute to the dysregulation of steroids produc-tion during the pathogenesis of PCOS.
In the present study, we observed thatgonadotropin down-regulated the expres-sion of PHB in vivo and in vitro. The regu-lation of PHB mRNA abundance by FSH invitro was time dependent, with down-regu-lation evident at 6 h but not at 24 h, whichmight be due to reduced basal level of PHB
at the latter timepoint.Thiswouldexplain theobservationthat FSH failed to reduce PHB protein content after 24 htreatment (Figure 2B). However, our in vivo results werenot in agreement with earlier observations that gonado-tropin treatment either had no effect on the mRNA/pro-tein levels (26) or increased PHB protein level (27). Al-though the reasons for these apparent differences arenot immediately obvious, it is possible that this could bedue to differences in gonadotropin dosage used andtechniques in granulosa cell isolation and mRNA de-
Figure 4. A–C, The influence of modulation of the PI3K/Akt pathway on granulosa cellPHB expression. A, Granulosa cells were treated with the PI3K inhibitor LY (10 �M,DMSO as control) or the Akt inhibitor TCN (5 �M) for 24 h, and the content of PHB wasexamined by Western blot. B and C, Granulosa cells were infected with adenoviral-constitutive A-Akt (MOI � 0-20, 24 h, adenoviral-LacZ as control) or adenoviral-DN-Akt(MOI � 40-80, 48 h), and PHB content was determined by Western blot. Bothadenoviral constructs contain an HA tag. D, Effect of PHB knockdown on pAkt content.Granulosa cells were infected with adenoviral-shPHB (MOI � 0-50, 24 h, adenoviral-shNeg as negative control), and the contents of PHB, pAkt and total Akt were examinedby Western blot. Representative immunoblots are shown in the figure. Data arepresented as mean � SEM of 3 independent experiments and analyzed by 1-wayANOVA and subsequently by Bonferroni post hoc test. *, P � .05, **, P � .01,***, P � .001 compared with DMSO or adenoviral control.
962 Wang et al Prohibitin and Chemerin Suppress Steroidogenesis Endocrinology, February 2013, 154(2):956–967

tection. A high dose (50 IU) of gonadotropin could haveminimized its effect on PHB expression via receptordown-regulation/desensitization.
Although the multifunctional role of PHB has beendemonstrated in various cell types, how the PHB gene isregulated is not completely understood. Here we show, inaddition to FSH, chemerin may be a novel regulator ofPHB in granulosa cells. However, the mechanism bywhich chemerin up-regulates PHB expression is unknown.In addition to the putative binding sites for the CCAAT/enhancer-binding protein and insulin response element inthe promoter of PHB (39), there are other elements such asperoxisomal proliferator-activated receptor, activatingprotein 2, estrogen receptor, and specificity protein-1based on the prediction using the transcription elementsearch system. Whether chemerin regulates the expressionof PHB via modulating these transcriptional factors re-quires further investigation.
The observations that chemerin andPHB were regulated by FSH in a similarpattern and that chemerin up-regulatesPHB content in granulosa cells suggestthat PHB may act as a downstreammolecule in granulosa cells. We havedemonstrated that chemerin plays aninhibitory roleonFSH-induced steroid-ogenesis in follicle and granulosa cellcultures (19) and that exogenous PHBsuppressed FSH-induced steroidogene-sis (present study). These findings arefurther supported by the loss-of-func-tion study showing that PHB knock-down attenuated chemerin-suppressedgranulosa cell steroidogenesis. How-ever, the mechanism by which PHB reg-ulates steroidogenesis is unclear. Be-cause chemerin suppresses the nuclearreceptor NR5a1/2-mediated steroido-genesis (19) and PHB is shown to act asa coactivator or corepressor of varioustranscriptional factors (e.g., E2F tran-scriptional factor 1 or p53) (40, 41), itis possible that PHB acts as a corepres-sor of NR5a1/2 to regulate granulosacell steroidogenesis.
PCOS is commonly associated withobesity, insulin resistance, dysregula-tion of steroids, and polycystic ovaries(42). Using a chronic DHT-inducedPCOS rat model as previously reported(43), we have demonstrated that serumand ovarian chemerin levels are ele-
vated in DHT-treated rats compared with controls (19).The observations that serum chemerin levels are elevatedin obese women and in PCOS subjects (8), that chemerinis important in adipogenesis, and that it suppresses FSH-induced steroidogenesis (19) are consistent with a possibleinvolvement of chemerin in the pathogenesis of PCOS. Inaddition, our finding that both mRNA and protein levelsof PHB increase in DHT-treated rats, together with itssuppressive role on steroidogenesis, also raises the inter-esting possibility of an association between PHB andPCOS. Recently PHB has been linked to adipogenesis andobesity because its expression is up-regulated in high-fatdiet-induced obese mice, and PHB knockdown signifi-cantly reduces the expression of adipogenic markers (24,25, 39). Both chemerin and PHB could regulate insulinsignaling because chemerin induces insulin resistance inskeletal muscle in vivo (44), metformin treatment reducesserum chemerin level (18), and PHB attenuates insulin-
Figure 5. The modulatory role of the PI3K/Akt pathway on the suppression of FSH-inducedsteroidogenesis by PHB. A, Granulosa cells were infected with adenoviral-PHB (MOI � 40,adenoviral-LacZ as control) for 24 h and treated with LY (10 �M) or the Akt inhibitor TCN (5�M) for 1 h prior to culture with FSH (100 ng/mL, 24 h). B, Granulosa cells were coinfectedwith adenoviral-PHB (MOI � 40) and constitutively A-Akt (with HA tag, MOI � 10) for 24 hand then treated with LY in the presence of FSH. Testosterone (0.5 �M) was added to serveas substrate of aromatase during culture. Estradiol secreted into the medium (EIA), and thecontents of aromatase, PHB, and HA (Western blot) were analyzed. Shown are representativeimmunoblots. The 2 blots for aromatase show the signals with shorter (top panel) and longer(bottom panel) exposure time. Data are presented as mean � SEM of 3 independentexperiments and analyzed by 2-way ANOVA and subsequently by Bonferroni post hoc test.*, P � .05, **, P � .01, ***, P � .001 compared with DMSO; #, P � .05, ##, P � .01,###, P � .001 compared with adenovirus control (LacZ or Neg).
Endocrinology, February 2013, 154(2):956–967 endo.endojournals.org 963

stimulated glucose transport (45, 46). The up-regulationof PHB by chemerin, as well as their suppressive role onsteroidogenesis and insulin signaling, raises a possibilitythat chemerin may contribute to the pathogenesis ofPCOS, particularly in the phenotype of obesity, insulinresistance, and dysregulation of steroidogenesis in a PHB-mediated way.
In our culture system, granulosa cells have a good ste-roidogenic response to FSH, which is in good agreementwith well-established observations that FSH stimulates
cyp19/cyp11a expression and steroid production (5, 47,48). Testosterone was added in the culture as the substrateof aromatase, which is commonly used in the studies ofFSH-induced estrogen production (38, 48, 49). We alsoobserved that the presence of testosterone augmented theeffect of FSH on the production of progesterone and es-tradiol in granulosa cells (Supplemental Figure 2), whichwas in agreement with the literature (38, 50, 51). It isnoteworthy that exogenous PHB suppresses FSH-inducedsteroidogenic enzyme expression at the transcriptionaland translational levels. The inhibitory role of PHB oncyp19 expression and estradiol production occurs at 18and 24 h after an FSH challenge. Although there is a rapidincrease of progesterone production at 3 h, exogenousPHB suppresses cyp11a expression and progesterone se-cretion only at 24 h under current experimental condi-tions. The increase of progesterone secretion is likely dueto rapid increase in StAR expression (42-fold) known tofacilitate cholesterol transport into the inner mitochon-drial membrane for steroidogenesis (48, 52). HoweverPHB has no suppression on StAR and HSD-3� expression.Because PHB is reported to serve as a coactivator or core-pressor of distinct transcriptional factors (estrogen recep-tor-�, p53, E2F family of DNA-binding transcription fac-tors, etc,) (40, 53, 54), we hypothesize that PHBsuppresses FSH-induced steroidogenesis via acting as acorepressor of transcriptional factors targeting steroido-
genic enzymes, such as NR5a1, NR5a2,CCAAT/enhancer-binding protein, et al. (55–58). The different effect of PHB on cyp11a,cyp19, StAR, and HSD-3� may result from theaffinity or specificity of PHB to various tran-scriptional factors. Further studies on the pro-moter activity and loss in function by mutagen-esis are needed to test this possibility and toexplore the interaction region between thesemolecules.
The PI3K/Akt signaling pathway plays anessential role in the regulation granulosa cellsurvival and differentiation during folliculardevelopment (36–38). Using specific inhibi-tors and dominant negative Akt construct, weobserved that active Akt negatively regulatesPHB expression and its suppressive role ingranulosa cells. Although PHB is reported to bea substrate of Akt and the insulin receptor (59,60), its phosphorylation status is not affectedby PI3K inhibitor in granulosa cells (29). Nev-ertheless, recent data show that overexpressionof PHB decreases phospho-Akt (S473 and T308)levels in adipocytes and MCF-7 cells (59, 61),suggesting an inverse relationship between
Figure 7. A hypothetical model depicting PHB and chemerin as negative regulatorsof FSH-induced steroidogenesis. In the absence of chemerin (A), FSH stimulates PI3K/Akt signaling, induces the expression of steroidogenic enzymes (e.g., cyp19), andpromotes steroid production. Akt negatively regulates PHB expression and function;NR5a1 and NR5a2 are the main transcriptional factors involved in this process. Whenextracellular chemerin level increases (B), chemerin up-regulates PHB expression,which suppresses FSH-induced steroidogenesis via modulating the expression andactivity of NR5a1/2. Inhibition of PI3K/Akt signaling enhances the suppressive role ofPHB due to reduced feedback inhibition.
Figure 6. Expression of PHB in granulosa cells from CTL and DHT-treated rats. Granulosa cells were collected from 5 rats of the CTL orDHT group and pooled for mRNA and protein analysis. The mRNAabundance (A) and protein content (B) of PHB were measured by real-time PCR and Western blot, respectively. Data are presented asmean � SEM of 3 independent experiments and analyzed by 1-wayANOVA and subsequently by Bonferroni post hoc test. *, P � .05compared with CTL.
964 Wang et al Prohibitin and Chemerin Suppress Steroidogenesis Endocrinology, February 2013, 154(2):956–967

PHB and phosphorylated Akt. We have observed thatknockdown of PHB increased the level of phospho-Akt(S473) in granulosa cells, suggesting that a negative feed-back regulation on the Akt phosphorylation by PHB. An-other example of a negative regulation loop is reportedbetween the up-regulation of PHB by insulin and an in-hibitory effect of PHB overexpression on insulin-inducedadipogenesis (39). In addition, a positive regulatory loopis observed between PHB and MAPK kinase (MEK)-1,demonstrating that PHB is required for MEK1 activity,while being a potential target of MEK1 (29). These regu-latory loops, whether positive or negative, may representa common mechanism for the precise control of PHB func-tion and regulation of granulosa cell differentiation.
In addition to regulating the PI3K/Akt pathways, FSHis known to activate the protein kinase A (PKA) pathwayin ovarian cells (62-64). We have demonstrated thatchemerin suppressed forskolin-induced steroidogenesis,possible via decreasing the transcriptional activity ofNR5a1/2 (19) and that PHB inhibited Akt phosphoryla-tion. Whether chemerin/PHB interacts with the PKA path-way is unknown. Besides the regulation of NR5a1/2, it isalso possible that chemerin binds to its Gi protein-coupledreceptor and negatively regulates PKA signaling; however,this possibility needs to be further investigated.
In this study, the presence of the PI3K inhibitor LYdramatically reduced FSH-induced steroidogenesis. Al-though constitutively active Akt amplified FSH-inducedgranulosa cell differentiation (38), it alone had no signif-icant influence on the action of FSH in the presence of thePI3K inhibitor LY (Figure 5B). A possible reason is that thedownstream molecules of PI3K other than Akt (such asPKA and protein kinase C) are also required in the regu-lation of steroidogenesis. Apart from phosphorylatingAkt at Thr308, 3�-phosphoinositide-dependent kinase-1(PDK1) phosphorylates the equivalent residues on PKA,protein kinase C, and p70-S6 kinase, kinases in the sameAGC family as Akt (65–68). Suppression of PI3K activitymay affect PKA signaling, and constitutively active Aktmay be unable to compensate for the inhibition of PKAsignaling by inhibitors on FSH-induced steroidogenesis, asreported previously that constitutively active Akt does notovercome the inhibitory effect of PKA inhibitor H89 onFSH-induced estradiol production (38).
In conclusion, our findings demonstrate that PHB is anovel negative regulator on FSH-induced steroidogenesisand mediates the action of chemerin on steroid produc-tion. As shown in the hypothetical model (Figure 7),chemerin up-regulates PHB expression, which inhibitsFSH-induced steroidogenesis. Knockdown of PHB atten-uates the suppressive effect of chemerin on FSH-inducedp450scc and aromatase expression. PI3K/Akt activation
negatively regulates the expression and function of PHB,whereas inhibition of PI3K pathway enhances the sup-pressive role of PHB. The increase of PHB expression ingranulosa cells from DHT-treated rats suggests that, as inthe case of chemerin, PHB may be involved in the dys-regulation of steroids production in PCOS. This study sig-nificantly improves our understanding of the role of PHBin ovarian cells and its potential contribution to the patho-genesis of PCOS.
Acknowledgments
Address all correspondence and requests for reprints to:Benjamin K. Tsang, PhD, Ottawa Hospital Research Institute,The Ottawa Hospital, General Campus, 501 Smyth Road, MailBox 511, Ottawa, Ontario, Canada K1H 8L6. E-mail:[email protected].
This work is supported by grants from the CanadianInstitutes of Health Research (CIHR; MOP-119381) andthe World Class University (WCU) program through theMinistry of Education, Science and Technology andfunded by the National Research Foundation of Korea(R31-10056), Human Reproduction Research Fund of theOttawa Fertility Centre, and Canadian Institutes ofHealth Research-Quebec Training Network in PerinatalResearch (CIHR-QTNPR) graduate scholarship (toQ.W.).
Disclosure Summary: The authors declare no conflict ofinterest.
References
1. McGee EA, Hsueh AJ. Initial and cyclic recruitment of ovarian fol-licles. Endocr Rev. 2000;21(2):200–214.
2. Jamnongjit M, Hammes SR. Ovarian steroids: the good, the bad,and the signals that raise them. Cell Cycle. 2006;5(11):1178–1183.
3. Yong EL, Hillier SG, Turner M, Baird DT, Ng SC, Bongso A, Rat-nam SS. Differential regulation of cholesterol side-chain cleavage(P450scc) and aromatase (P450arom) enzyme mRNA expression bygonadotrophins and cyclic AMP in human granulosa cells. J MolEndocrinol. 1994;12(2):239–249.
4. Sher N, Yivgi-Ohana N, Orly J. Transcriptional regulation of thecholesterol side chain cleavage cytochrome P450 gene (CYP11A1)revisited: binding of GATA, cyclic adenosine 3�,5�-monophosphateresponse element-binding protein and activating protein (AP)-1 pro-teins to a distal novel cluster of cis-regulatory elements potentiatesAP-2 and steroidogenic factor-1-dependent gene expression in therodent placenta and ovary. Mol Endocrinol. 2007;21(4):948–962.
5. Eimerl S, Orly J. Regulation of steroidogenic genes by insulin-likegrowth factor-1 and follicle-stimulating hormone: differential re-sponses of cytochrome P450 side-chain cleavage, steroidogenicacute regulatory protein, and 3�-hydroxysteroid dehydrogenase/isomerase in rat granulosa cells. Biol Reprod. 2002;67(3):900–910.
6. Irving-Rodgers HF, Harland ML, Sullivan TR, Rodgers RJ. Studies
Endocrinology, February 2013, 154(2):956–967 endo.endojournals.org 965

of granulosa cell maturation in dominant and subordinate bovinefollicles: novel extracellular matrix focimatrix is co-ordinately reg-ulated with cholesterol side-chain cleavage CYP11A1. Reproduc-tion. 2009;137(5):825–834.
7. Hong Y, Yu B, Sherman M, Yuan YC, Zhou D, Chen S. Molecularbasis for the aromatization reaction and exemestane-mediated irre-versible inhibition of human aromatase. Mol Endocrinol. 2007;21(2):401–414.
8. Bozaoglu K, Bolton K, McMillan J, Zimmet P, Jowett J, Collier G,Walder K, Segal D. Chemerin is a novel adipokine associated withobesity and metabolic syndrome. Endocrinology. 2007;148(10):4687–4694.
9. Wittamer V, Franssen JD, Vulcano M, et al. Specific recruitment ofantigen-presenting cells by chemerin, a novel processed ligand fromhuman inflammatory fluids. J Exp Med. 2003;198(7):977–985.
10. Zabel BA, Allen SJ, Kulig P, et al. Chemerin activation by serineproteases of the coagulation, fibrinolytic, and inflammatory cas-cades. J Biol Chem. 2005;280(41):34661–34666.
11. Zhao L, Yamaguchi Y, Sharif S, et al. Chemerin158K protein is thedominant chemerin isoform in synovial and cerebrospinal fluids butnot in plasma. J Biol Chem. 2011;286(45):39520–39527.
12. Bozaoglu K, Curran JE, Stocker CJ, et al. Chemerin, a novel adi-pokine in the regulation of angiogenesis. J Clin Endocrinol Metab.2010;95(5):2476–2485.
13. Huang J, Zhang J, Lei T, et al. Cloning of porcine chemerin,ChemR23 and GPR1 and their involvement in regulation of lipo-genesis. BMB Rep. 2010;43(7):491–498.
14. Goralski KB, McCarthy TC, Hanniman EA, et al. Chemerin, a noveladipokine that regulates adipogenesis and adipocyte metabolism.J Biol Chem. 2007;282(38):28175–28188.
15. Luangsay S, Wittamer V, Bondue B, et al. Mouse ChemR23 is ex-pressed in dendritic cell subsets and macrophages, and mediates ananti-inflammatory activity of chemerin in a lung disease model.J Immunol. 2009;183(10):6489–6499.
16. Bondue B, Wittamer V, Parmentier M. Chemerin and its receptorsin leukocyte trafficking, inflammation and metabolism. CytokineGrowth Factor Rev. 2011;22(5–6):331–338.
17. Cash JL, Hart R, Russ A, et al. Synthetic chemerin-derived peptidessuppress inflammation through ChemR23. J Exp Med. 2008;205(4):767–775.
18. Tan BK, Chen J, Farhatullah S, et al. Insulin and metformin regulatecirculating and adipose tissue chemerin. Diabetes. 2009;58(9):1971–1977.
19. Wang Q, Kim JY, Xue K, Liu JY, Leader A, Tsang BK. Chemerin,a novel regulator of follicular steroidogenesis and its potential in-volvement in polycystic ovarian syndrome. Endocrinology. 2012;153(11):5600–5611.
20. Nuell MJ, Stewart DA, Walker L, et al. Prohibitin, an evolutionarilyconserved intracellular protein that blocks DNA synthesis in normalfibroblasts and HeLa cells. Mol Cell Biol. 1991;11(3):1372–1381.
21. McClung JK, Jupe ER, Liu XT, Dell’Orco RT. Prohibitin: potentialrole in senescence, development, and tumor suppression. ExpGerontol. 1995;30(2):99–124.
22. Rastogi S, Joshi B, Dasgupta P, Morris M, Wright K, Chellappan S.Prohibitin facilitates cellular senescence by recruiting specific core-pressors to inhibit E2F target genes. Mol Cell Biol. 2006;26(11):4161–4171.
23. Roskams AJ, Friedman V, Wood CM, et al. Cell cycle activity andexpression of prohibitin mRNA. J Cell Physiol. 1993;157(2):289–295.
24. Oh TS, Kwon EY, Choi JW, Choi MS, Yun JW. Time-dependenthepatic proteome analysis in lean and diet-induced obese mice. JMicrobiol Biotechnol. 2011;21(12):1211–1227.
25. Liu D, Lin Y, Kang T, et al. Mitochondrial dysfunction and adipo-genic reduction by prohibitin silencing in 3T3-L1 cells. PLoS One.2012;7(3):e34315.
26. Thompson WE, Powell JM, Whittaker JA, Sridaran R, Thomas KH.
Immunolocalization and expression of prohibitin, a mitochondrialassociated protein within the rat ovaries. Anat Rec. 1999;256(1):40–48.
27. Thompson WE, Asselin E, Branch A, et al. Regulation of prohibitinexpression during follicular development and atresia in the mam-malian ovary. Biol Reprod. 2004;71(1):282–290.
28. Chowdhury I, Xu W, Stiles JK, et al. Apoptosis of rat granulosa cellsafter staurosporine and serum withdrawal is suppressed by adeno-virus-directed overexpression of prohibitin. Endocrinology. 2007;148(1):206–217.
29. Chowdhury I, Garcia-Barrio M, Harp D, Thomas K, Matthews R,Thompson WE. The emerging roles of prohibitins in folliculogen-esis. Front Biosci (Elite Ed). 2012;4:690–699.
30. Campbell KL. Ovarian granulosa cells isolated with EGTA and hy-pertonic sucrose: cellular integrity and function. Biol Reprod. 1979;21(4):773–786.
31. Wang H, Jiang JY, Zhu C, Peng C, Tsang BK. Role and regulationof nodal/activin receptor-like kinase 7 signaling pathway in the con-trol of ovarian follicular atresia. Mol Endocrinol. 2006;20(10):2469–2482.
32. Brawer J, Schipper H, Robaire B. Effects of long term androgen andestradiol exposure on the hypothalamus. Endocrinology. 1983;112(1):194–199.
33. Silfen ME, Denburg MR, Manibo AM, et al. Early endocrine, met-abolic, and sonographic characteristics of polycystic ovary syn-drome (PCOS): comparison between nonobese and obese adoles-cents. J Clin Endocrinol Metab. 2003;88(10):4682–4688.
34. Fassnacht M, Schlenz N, Schneider SB, Wudy SA, Allolio B, Arlt W.Beyond adrenal and ovarian androgen generation: Increased periph-eral 5�-reductase activity in women with polycystic ovary syn-drome. J Clin Endocrinol Metab. 2003;88(6):2760–2766.
35. Livak KJ, Schmittgen TD. Analysis of relative gene expression datausing real-time quantitative PCR and the 2[���C(T)] method.Methods. 2001;25(4):402–408.
36. Mani AM, Fenwick MA, Cheng Z, Sharma MK, Singh D, WathesDC. IGF1 induces up-regulation of steroidogenic and apoptotic reg-ulatory genes via activation of phosphatidylinositol-dependent ki-nase/AKT in bovine granulosa cells. Reproduction. 2010;139(1):139–151.
37. Alam H, Maizels ET, Park Y, et al. Follicle-stimulating hormoneactivation of hypoxia-inducible factor-1 by the phosphatidylinositol3-kinase/AKT/Ras homolog enriched in brain (Rheb)/mammaliantarget of rapamycin (mTOR) pathway is necessary for induction ofselect protein markers of follicular differentiation. J Biol Chem.2004;279(19):19431–19440.
38. Zeleznik AJ, Saxena D, Little-Ihrig L. Protein kinase B is obligatoryfor follicle-stimulating hormone-induced granulosa cell differenti-ation. Endocrinology. 2003;144(9):3985–3994.
39. Ande SR, Xu Z, Gu Y, Mishra S. 2011 Prohibitin has an importantrole in adipocyte differentiation. Int J Obes (Lond). 2012;36(9):1236–1244.
40. Fusaro G, Dasgupta P, Rastogi S, Joshi B, Chellappan S. Prohibitininduces the transcriptional activity of p53 and is exported from thenucleus upon apoptotic signaling. J Biol Chem. 2003;278(48):47853–47861.
41. Joshi B, Rastogi S, Morris M, et al. Differential regulation of humanYY1 and caspase 7 promoters by prohibitin through E2F1 and p53binding sites. Biochem J. 2007;401(1):155–166.
42. Yildiz BO, Azziz R. Ovarian and adipose tissue dysfunction in poly-cystic ovary syndrome: report of the 4th special scientific meeting ofthe Androgen Excess and PCOS Society. Fertil Steril. 2010;94(2):690–693.
43. Manneras L, Cajander S, Holmang A, et al. A new rat model ex-hibiting both ovarian and metabolic characteristics of polycysticovary syndrome. Endocrinology. 2007;148(8):3781–3791.
44. Becker M, Rabe K, Lebherz C, et al. Expression of human chemerininduces insulin resistance in the skeletal muscle but does not affect
966 Wang et al Prohibitin and Chemerin Suppress Steroidogenesis Endocrinology, February 2013, 154(2):956–967

weight, lipid levels, and atherosclerosis in LDL receptor knockoutmice on high-fat diet. Diabetes. 2010;59(11):2898–2903.
45. Brasaemle DL, Dolios G, Shapiro L, Wang R. Proteomic analysis ofproteins associated with lipid droplets of basal and lipolytically stim-ulated 3T3-L1 adipocytes. J Biol Chem. 2004;279(45):46835–46842.
46. Vessal M, Mishra S, Moulik S, Murphy LJ. Prohibitin attenuatesinsulin-stimulated glucose and fatty acid oxidation in adipose tissueby inhibition of pyruvate carboxylase. FEBS J. 2006;273(3):568–576.
47. Miller WL. Molecular biology of steroid hormone synthesis. EndocrRev. 1988;9(3):295–318.
48. Wayne CM, Fan HY, Cheng X, Richards JS. Follicle-stimulatinghormone induces multiple signaling cascades: evidence that activa-tion of Rous sarcoma oncogene, RAS, and the epidermal growthfactor receptor are critical for granulosa cell differentiation. MolEndocrinol. 2007;21(8):1940–1957.
49. Escamilla-Hernandez R, Little-Ihrig L, Zeleznik AJ. Inhibition of ratgranulosa cell differentiation by overexpression of G�q. Endocrine.2008;33(1):21–31.
50. Armstrong DT, Dorrington JH. Androgens augment FSH-inducedprogesterone secretion by cultured rat granulosa cells. Endocrinol-ogy. 1976;99(5):1411–1414.
51. Harlow CR, Shaw HJ, Hillier SG, Hodges JK. Factors influencingfollicle-stimulating hormone-responsive steroidogenesis in marmo-set granulosa cells: effects of androgens and the stage of follicularmaturity. Endocrinology. 1988;122(6):2780–2787.
52. Pescador N, Houde A, Stocco DM, Murphy BD. Follicle-stimulatinghormone and intracellular second messengers regulate steroidogenicacute regulatory protein messenger ribonucleic acid in luteinizedporcine granulosa cells. Biol Reprod. 1997;57(3):660–668.
53. He B, Feng Q, Mukherjee A, et al. A repressive role for prohibitin inestrogen signaling. Mol Endocrinol. 2008;22(2):344–360.
54. Choi D, Lee SJ, Hong S, Kim IH, Kang S. Prohibitin interacts withRNF2 and regulates E2F1 function via dual pathways. Oncogene.2008;27(12):1716–1725.
55. Boerboom D, Pilon N, Behdjani R, Silversides DW, Sirois J. Ex-pression and regulation of transcripts encoding two members of theNR5A nuclear receptor subfamily of orphan nuclear receptors, ste-roidogenic factor-1 and NR5A2, in equine ovarian cells during theovulatory process. Endocrinology. 2000;141(12):4647–4656.
56. Hinshelwood MM, Repa JJ, Shelton JM, Richardson JA, Mangels-dorf DJ, Mendelson CR. Expression of LRH-1 and SF-1 in the mouseovary: localization in different cell types correlates with differingfunction. Mol Cell Endocrinol. 2003;207(1-2):39–45.
57. Reinhart AJ, Williams SC, Stocco DM. Transcriptional regulation ofthe StAR gene. Mol Cell Endocrinol. 1999;151(1-2):161–169.
58. Andrieu T, Pezzi V, Sirianni R, et al. cAMP-dependent regulation ofCYP19 gene in rabbit preovulatory granulosa cells and corpus lu-teum. J Steroid Biochem Mol Biol. 2009;116(1-2):110–117.
59. Ande SR, Mishra S. Prohibitin interacts with phosphatidylinositol3,4,5-triphosphate (PIP3) and modulates insulin signaling. BiochemBiophys Res Commun. 2009;390(3):1023–1028.
60. Han EK, McGonigal T, Butler C, Giranda VL, Luo Y Character-ization of Akt overexpression in MiaPaCa-2 cells: prohibitin is anAkt substrate both in vitro and in cells. Anticancer Res. 2008;28(2A):957–963.
61. Ande SR, Gu Y, Nyomba BL, Mishra S. Insulin induced phosphor-ylation of prohibitin at tyrosine 114 recruits Shp1. Biochim BiophysActa. 2009;1793(8):1372–1378.
62. Salvador LM, Park Y, Cottom J, et al. Follicle-stimulating hormonestimulates protein kinase A-mediated histone H3 phosphorylationand acetylation leading to select gene activation in ovarian granulosacells. J Biol Chem. 2001;276(43):40146–40155.
63. DeManno DA, Cottom JE, Kline MP, Peters CA, Maizels ET, Hun-zicker-Dunn M. Follicle-stimulating hormone promotes histone H3phosphorylation on serine-10. Mol Endocrinol. 1999;13(1):91–105.
64. Hunzicker-Dunn M, Maizels ET. FSH signaling pathways in im-mature granulosa cells that regulate target gene expression: branch-ing out from protein kinase A. Cell Signal. 2006;18(9):1351–1359.
65. Vanhaesebroeck B, Alessi DR. The PI3K-PDK1 connection: morethan just a road to PKB. Biochem J. 2000;(346 Pt 3):561–576.
66. Le Good JA, Ziegler WH, Parekh DB, Alessi DR, Cohen P, ParkerPJ. Protein kinase C isotypes controlled by phosphoinositide 3-ki-nase through the protein kinase PDK1. Science. 1998;281(5385):2042–2045.
67. Pullen N, Dennis PB, Andjelkovic M, et al. Phosphorylation andactivation of p70s6k by PDK1. Science. 1998;279(5351):707–710.
68. Cheng X, Ma Y, Moore M, Hemmings BA, Taylor SS. Phosphor-ylation and activation of cAMP-dependent protein kinase by phos-phoinositide-dependent protein kinase. Proc Natl Acad Sci USA.1998;95(17):9849–9854.
Endocrinology, February 2013, 154(2):956–967 endo.endojournals.org 967





























