Embed Size (px)
Citation preview

APPLIED AND ENvIRONMENTAL MICROBIOLOGY, Jan. 1994, p. 187-1940099-2240/94/$04.00+0Copyright © 1994, American Society for Microbiology
Inhibition of the Lignin Peroxidase of Phanerochaetechrysosporium by Hydroxylamino-Dinitrotoluene, an EarlyIntermediate in the Degradation of 2,4,6-Trinitrotoluene
JOCHEN MICHELS AND GERHARD GOTTSCHALK*Institut fir Mikrobiologie, Georg-August-Universitat Gottingen,
D-37077 Gottingen, Federal Republic of Germany
Received 11 June 1993/Accepted 21 October 1993
The ability of the white rot fungus Phanerochaete chrysosporium to mineralize 2,4,6-trinitrotoluene (TNT)was studied in the concentration range of 0.36 to 20.36 mg/liter. The initial rate of 14CO2 formation was 30%Yoin 4 days at 0.36 mg of [14C]TNT per liter and decreased to 5% in 4 days at 20.36 mg of [14C]TNT per liter.Such a pronounced inhibition was not observed when a mixture of [14C]2-amino-4,6-dinitrotoluene and["4C]4-amino-2,6-dinitrotoluene was used as a substrate. 2-Hydroxylamino-4,6-dinitrotoluene and its isomer4-hydroxylamino-2,6-dinitrotoluene were identified as the first detectable degradation products of TNT. Theirtransient accumulation correlated with the inhibition of TNT degradation and of the veratryl alcohol oxidaseactivity of lignin peroxidase. With purified lignin peroxidase H8, it could be shown that the two isomers ofhydroxylamino-dinitrotoluene were oxidized by lignin peroxidase. The corresponding nitroso-dinitrotoluenesapparently were formed, as indicated by the formation of azoxy-tetranitrotoluenes.
2,4,6-Trinitrotoluene (TNT) is a predominant contaminantof soil and groundwater at many sites ofTNT production andprocessing in Germany and other countries. Its presence isdue to contaminated wastewaters released into the environ-ment and to the destruction of ammunition plants at the endof World War II. TNT is recalcitrant and has been shown tobe toxic to gram-positive bacteria, yeasts, and fungi (13) andgreen algae (25). It is also mutagenic, according to the Amestest (36).The wood-rotting fungus Phanerochaete chrysosporium
secretes nonspecific extracellular peroxidases for lignin deg-radation. The fungus has been reported to degrade a varietyof recalcitrant pollutants, such as chlorinated xenobiotics,polycyclic aromatic hydrocarbons, and polymers. The min-eralization of TNT was first reported by Fernando et al.using [14C]TNT and ligninolytic cultures (9). The intoleranceof the fungus for high concentrations of TNT (>20 ppm) andthe ability of nonligninolytic cultures to degrade smallamounts of TNT were observed by Spiker et al. (26). Aprocess for the cleanup of TNT-contaminated wastewaterwith P. chrysosporium immobilized on rotating discs was
proposed by Sublette et al. (29). The pathway of degradationand the participation of ligninolytic enzymes have beenstudied so far only for 2,4-dinitrotoluene, not for TNT (34).Recently, Stahl and Aust reported a plasma membrane-dependent reduction of TNT by P. chrysosponum (27). Therate of reduction was independent of the ligninolytic system,and the amino-dinitrotoluene isomers were identified as
reduction products (28).In this report, we examine the negative effect of increasing
concentrations of TNT on its rate of degradation by ligni-nolytic cultures of P. chrysosporium and show that hydrox-ylamino-dinitrotoluene, an early intermediate of TNT degra-
* Corresponding author. Mailing address: Institut fur Mikrobiol-ogie, Georg-August-Universitat Gottingen, Grisebachstrasse 8,D-37077 Gottingen, Federal Republic of Germany. Phone: 49-551-393781. Fax: 49-551-393793.
dation, causes the decrease in mineralization rates byaffecting lignin peroxidase.
MATERIALS AND METHODS
Chemicals. TNT was obtained from Dynamit Nobel AG(Troisdorf, Federal Republic of Germany [FRG]). 2-Amino-4,6-dinitrotoluene and 4-amino-2,6-dinitrotoluene were ob-tained from the Institute of Organic Chemistry of Georg-August-University (Gottingen, FRG). U-'4C-ring-labeledtoluene was obtained from Sigma Chemie GmbH, Deisen-hofen, FRG, and m-chloroperoxybenzoic acid was obtainedfrom E. Merck AG, Darmstadt, FRG.
U-14C-ring-labeled TNT. U-14C-ring-labeled toluene (1.25mCi) was diluted with 250 ,u of toluene to a specificradioactivity of 535 nCi/,mol. Nitration of toluene was
performed in three steps as described by Seidenfaden andPawellek (21). Neither purification of intermediates, e.g., theisomers of mononitrotoluene and dinitrotoluene, nor separa-tion of nitrating acid (see below) was carried out between thenitration steps. H20 formed during the nitration reactionswas bound by fuming sulfuric acid. A total of 390 ,ul ofnitrating acid I (160 ,ul of HNO3 [65%, vol/vol], 230 RI ofHXSO4 [96%, vol/vol]) was added slowly to the stirred[1 C]toluene at room temperature. After 2 h of incubation,the temperature was adjusted to 60°C, and 920 ,ul of nitratingacid II (150 ,ul of HNO3 [98%, vol/vol], 300 ,ul of H2SO4[96%, vol/vol], 470 pI of fuming sulfuric acid [65% SO3]) wasadded to the stirred mixture. The temperature was raised to85°C and maintained for at least 1 h. Afterwards, another 920pu1 of nitrating acid II was added, and the temperature was
raised to 103°C. Four hours later, the nitration reaction wasstopped by the addition of ice water (5 ml). TNT was thenextracted three times with 5 ml of ethyl acetate. Purificationof the final product was done by multiple recrystallizationfrom CC14 and additional analytical thin-layer chromatogra-phy (TLC) (toluene-methanol [9:1, vol/vol]). The yield ofU-_4C-ring-labeled TNT was 59%.
U-_4C-ring-labeled amino-dinitrotoluenes. Both isomers of
187
Vol. 60, No. 1
on May 17, 2018 by guest
http://aem.asm
.org/D
ownloaded from
188 MICHELS AND GOTFSCHALK
30-
20-
10
0 3 6 9 12 15 18 21 24
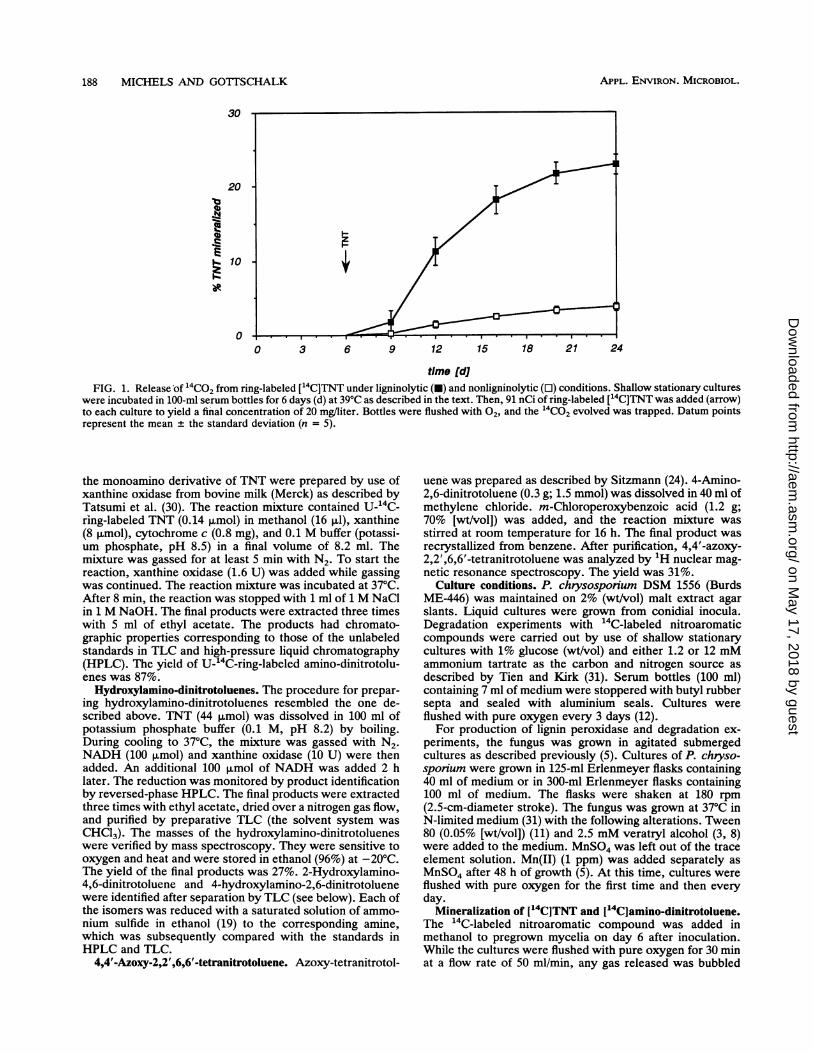
time [dlFIG. 1. Release of "4Co2 from ring-labeled ['4C]TNT under ligninolytic (-) and nonligninolytic (0) conditions. Shallow stationary cultures
were incubated in 100-ml serum bottles for 6 days (d) at 39°C as described in the text. Then, 91 nCi of ring-labeled ["4C]TNT was added (arrow)to each culture to yield a final concentration of 20 mg/liter. Bottles were flushed with 02, and the "4CO2 evolved was trapped. Datum pointsrepresent the mean + the standard deviation (n = 5).
the monoamino derivative of TNT were prepared by use ofxanthine oxidase from bovine milk (Merck) as described byTatsumi et al. (30). The reaction mixture contained U-'4C-ring-labeled TNT (0.14 ,umol) in methanol (16 RI), xanthine(8 ,umol), cytochrome c (0.8 mg), and 0.1 M buffer (potassi-um phosphate, pH 8.5) in a final volume of 8.2 ml. Themixture was gassed for at least 5 min with N2. To start thereaction, xanthine oxidase (1.6 U) was added while gassingwas continued. The reaction mixture was incubated at 37°C.After 8 min, the reaction was stopped with 1 ml of 1 M NaClin 1 M NaOH. The final products were extracted three timeswith 5 ml of ethyl acetate. The products had chromato-graphic properties corresponding to those of the unlabeledstandards in TLC and high-pressure liquid chromatography(HPLC). The yield of U-14C-ring-labeled amino-dinitrotolu-enes was 87%.
Hydroxylamino-dinitrotoluenes. The procedure for prepar-ing hydroxylamino-dinitrotoluenes resembled the one de-scribed above. TNT (44 ,mol) was dissolved in 100 ml ofpotassium phosphate buffer (0.1 M, pH 8.2) by boiling.During cooling to 37°C, the mixture was gassed with N2.NADH (100 pmol) and xanthine oxidase (10 U) were thenadded. An additional 100 ,umol of NADH was added 2 hlater. The reduction was monitored by product identificationby reversed-phase HPLC. The final products were extractedthree times with ethyl acetate, dried over a nitrogen gas flow,and purified by preparative TLC (the solvent system wasCHC13). The masses of the hydroxylamino-dinitrotolueneswere verified by mass spectroscopy. They were sensitive tooxygen and heat and were stored in ethanol (96%) at -20°C.The yield of the final products was 27%. 2-Hydroxylamino-4,6-dinitrotoluene and 4-hydroxylamino-2,6-dinitrotoluenewere identified after separation by TLC (see below). Each ofthe isomers was reduced with a saturated solution of ammo-nium sulfide in ethanol (19) to the corresponding amine,which was subsequently compared with the standards inHPLC and TLC.
4,4'-Azoxy-2,2',6,6'-tetranitrotoluene. Azoxy-tetranitrotol-
uene was prepared as described by Sitzmann (24). 4-Amino-2,6-dinitrotoluene (0.3 g; 1.5 mmol) was dissolved in 40 ml ofmethylene chloride. m-Chloroperoxybenzoic acid (1.2 g;70% [wt/vol]) was added, and the reaction mixture wasstirred at room temperature for 16 h. The final product wasrecrystallized from benzene. After purification, 4,4'-azoxy-2,2',6,6'-tetranitrotoluene was analyzed by 114 nuclear mag-netic resonance spectroscopy. The yield was 31%.
Culture conditions. P. chrysosporium DSM 1556 (BurdsME-446) was maintained on 2% (wt/vol) malt extract agarslants. Liquid cultures were grown from conidial inocula.Degradation experiments with 14C-labeled nitroaromaticcompounds were carried out by use of shallow stationarycultures with 1% glucose (wt/vol) and either 1.2 or 12 mMammonium tartrate as the carbon and nitrogen source asdescribed by Tien and Kirk (31). Serum bottles (100 ml)containing 7 ml of medium were stoppered with butyl rubbersepta and sealed with aluminium seals. Cultures wereflushed with pure oxygen every 3 days (12).For production of lignin peroxidase and degradation ex-
periments, the fungus was grown in agitated submergedcultures as described previously (5). Cultures of P. chryso-sporium were grown in 125-ml Erlenmeyer flasks containing40 ml of medium or in 300-ml Erlenmeyer flasks containing100 ml of medium. The flasks were shaken at 180 rpm(2.5-cm-diameter stroke). The fungus was grown at 37°C inN-limited medium (31) with the following alterations. Tween80 (0.05% [wt/vol]) (11) and 2.5 mM veratryl alcohol (3, 8)were added to the medium. MnSO4 was left out of the traceelement solution. Mn(II) (1 ppm) was added separately asMnSO4 after 48 h of growth (5). At this time, cultures wereflushed with pure oxygen for the first time and then everyday.
Mineralization of []4CJTNT and ['4C]amino-dinitrotoluene.The 14C-labeled nitroaromatic compound was added inmethanol to pregrown mycelia on day 6 after inoculation.While the cultures were flushed with pure oxygen for 30 minat a flow rate of 50 ml/min, any gas released was bubbled
APPL. ENvIRON. MICROBIOL.
on May 17, 2018 by guest
http://aem.asm
.org/D
ownloaded from
DEGRADATION OF TNT BY P. CHRYSOSPORIUM 189
45
30
15
045
30N~~~~~
15
00 3 6 9 12 15 18 21 24
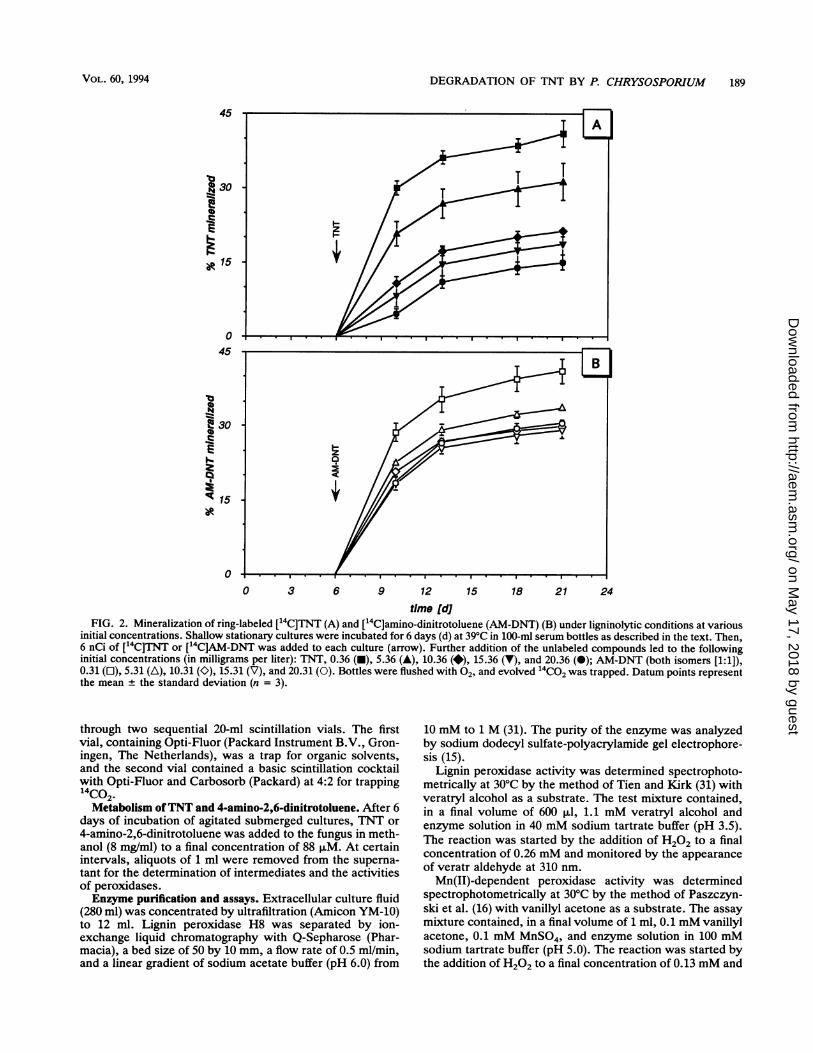
time [djFIG. 2. Mineralization of ring-labeled ['4C]TNT (A) and ["4C]amino-dinitrotoluene (AM-DNT) (B) under ligninolytic conditions at various
initial concentrations. Shallow stationary cultures were incubated for 6 days (d) at 39°C in 100-ml serum bottles as described in the text. Then,6 nCi of ["4C]TNT or ['4C]AM-DNT was added to each culture (arrow). Further addition of the unlabeled compounds led to the followinginitial concentrations (in milligrams per liter): TNT, 0.36 (-), 5.36 (A), 10.36 (*), 15.36 (V), and 20.36 (-); AM-DNT (both isomers [1:1]),0.31 (L), 5.31 (A), 10.31 (O), 15.31 (V), and 20.31 (0). Bottles were flushed with 02, and evolved '4CO2 was trapped. Datum points representthe mean + the standard deviation (n = 3).
through two sequential 20-ml scintillation vials. The firstvial, containing Opti-Fluor (Packard Instrument B.V., Gron-ingen, The Netherlands), was a trap for organic solvents,and the second vial contained a basic scintillation cocktailwith Opti-Fluor and Carbosorb (Packard) at 4:2 for trapping14c02.Metabolism ofTNT and 4-amino-2,6-dinitrotoluene. After 6
days of incubation of agitated submerged cultures, TNT or4-amino-2,6-dinitrotoluene was added to the fungus in meth-anol (8 mg/ml) to a final concentration of 88 ,uM. At certainintervals, aliquots of 1 ml were removed from the superna-tant for the determination of intermediates and the activitiesof peroxidases.Enzyme purification and assays. Extracellular culture fluid
(280 ml) was concentrated by ultrafiltration (Amicon YM-10)to 12 ml. Lignin peroxidase H8 was separated by ion-exchange liquid chromatography with Q-Sepharose (Phar-macia), a bed size of 50 by 10 mm, a flow rate of 0.5 ml/min,and a linear gradient of sodium acetate buffer (pH 6.0) from
10 mM to 1 M (31). The purity of the enzyme was analyzedby sodium dodecyl sulfate-polyacrylamide gel electrophore-sis (15).
Lignin peroxidase activity was determined spectrophoto-metrically at 30°C by the method of Tien and Kirk (31) withveratryl alcohol as a substrate. The test mixture contained,in a final volume of 600 ,ul, 1.1 mM veratryl alcohol andenzyme solution in 40 mM sodium tartrate buffer (pH 3.5).The reaction was started by the addition of H202 to a finalconcentration of 0.26 mM and monitored by the appearanceof veratr aldehyde at 310 nm.
Mn(II)-dependent peroxidase activity was determinedspectrophotometrically at 30°C by the method of Paszczyn-ski et al. (16) with vanillyl acetone as a substrate. The assaymixture contained, in a final volume of 1 ml, 0.1 mM vanillylacetone, 0.1 mM MnSO4, and enzyme solution in 100 mMsodium tartrate buffer (pH 5.0). The reaction was started bythe addition of H202 to a final concentration of 0.13 mM and
VOL. 60, 1994
on May 17, 2018 by guest
http://aem.asm
.org/D
ownloaded from
190 MICHELS AND GOTTSCHALK
120
1100
80
le60
40
20
0
80
60
.~40
20
00 12 24 36 48
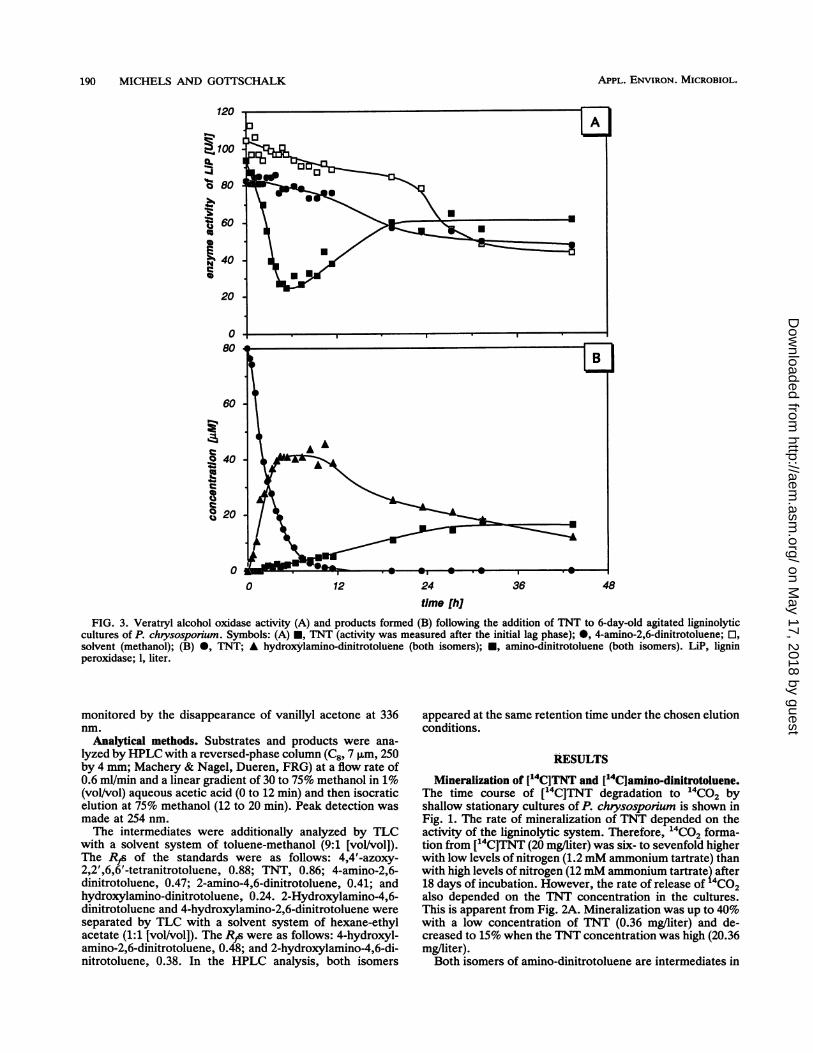
time [hiFIG. 3. Veratryl alcohol oxidase activity (A) and products formed (B) following the addition of TNT to 6-day-old agitated ligninolytic
cultures of P. chrysosporium. Symbols: (A) *, TNT (activity was measured after the initial lag phase); 0, 4-amino-2,6-dinitrotoluene; El,solvent (methanol); (B) 0, TNT; A hydroxylamino-dinitrotoluene (both isomers); *, amino-dinitrotoluene (both isomers). LiP, ligninperoxidase; 1, liter.
monitored by the disappearance of vanillyl acetone at 336nm.
Analytical methods. Substrates and products were ana-lyzed by HPLC with a reversed-phase column (C8, 7 ,um, 250by 4 mm; Machery & Nagel, Dueren, FRG) at a flow rate of0.6 m/min and a linear gradient of 30 to 75% methanol in 1%(vol/vol) aqueous acetic acid (0 to 12 min) and then isocraticelution at 75% methanol (12 to 20 min). Peak detection wasmade at 254 nm.The intermediates were additionally analyzed by TLC
with a solvent system of toluene-methanol (9:1 [vol/vol]).The R1s of the standards were as follows: 4,4'-azoxy-2,2',6,6'-tetranitrotoluene, 0.88; TNT, 0.86; 4-amino-2,6-dinitrotoluene, 0.47; 2-amino-4,6-dinitrotoluene, 0.41; andhydroxylamino-dinitrotoluene, 0.24. 2-Hydroxylamino-4,6-dinitrotoluene and 4-hydroxylamino-2,6-dinitrotoluene wereseparated by TLC with a solvent system of hexane-ethylacetate (1:1 [vol/vol]). The Rfs were as follows: 4-hydroxyl-amino-2,6-dinitrotoluene, 0.48; and 2-hydroxylamino-4,6-di-nitrotoluene, 0.38. In the HPLC analysis, both isomers
appeared at the same retention time under the chosen elutionconditions.
RESULTS
Mineralization of [14C]TNT and [14Clamino-dinitrotoluene.The time course of [14C]TNT degradation to 14CO2 byshallow stationary cultures of P. chrysosporium is shown inFig. 1. The rate of mineralization of TNT depended on theactivity of the ligninolytic system. Therefore, 14CO2 forma-tion from [14C]TNT (20 mg/liter) was six- to sevenfold higherwith low levels of nitrogen (1.2mM ammonium tartrate) thanwith high levels of nitrogen (12mM ammonium tartratel after18 days of incubation. However, the rate of release of 4CO2also depended on the TNT concentration in the cultures.This is apparent from Fig. 2A. Mineralization was up to 40%with a low concentration of TNT (0.36 mg/liter) and de-creased to 15% when the TNT concentration was high (20.36mg/liter).Both isomers of amino-dinitrotoluene are intermediates in
APPL. ENvIRON. MICROBIOL.
on May 17, 2018 by guest
http://aem.asm
.org/D
ownloaded from
DEGRADATION OF TNT BY P. CHRYSOSPORIUM 191
0,4
0,3
~~)0,2
0, 1
0,00 1 2 3 4 5
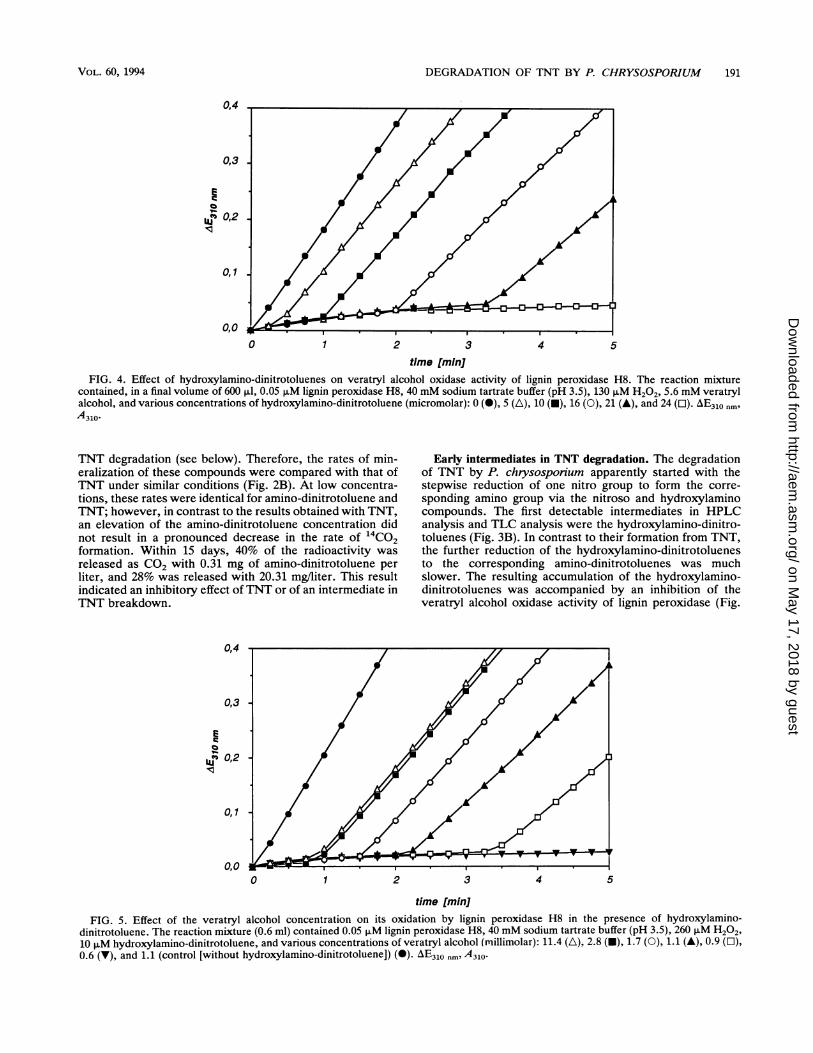
time [min]FIG. 4. Effect of hydroxylamino-dinitrotoluenes on veratryl alcohol oxidase activity of lignin peroxidase H8. The reaction mixture
contained, in a final volume of 600 ,ul, 0.05 ,uM lignin peroxidase H8, 40 mM sodium tartrate buffer (pH 3.5), 130 ,uM H202, 5.6 mM veratrylalcohol, and various concentrations of hydroxylamino-dinitrotoluene (micromolar): 0 (0), 5 (A), 10 (-), 16 (0), 21 (A), and 24 (O). AE310 nmA310.
TNT degradation (see below). Therefore, the rates of min-eralization of these compounds were compared with that ofTNT under similar conditions (Fig. 2B). At low concentra-tions, these rates were identical for amino-dinitrotoluene andTNT; however, in contrast to the results obtained with TNT,an elevation of the amino-dinitrotoluene concentration didnot result in a pronounced decrease in the rate of "4Co2formation. Within 15 days, 40% of the radioactivity wasreleased as CO2 with 0.31 mg of amino-dinitrotoluene perliter, and 28% was released with 20.31 mg/liter. This resultindicated an inhibitory effect of TNT or of an intermediate inTNT breakdown.
0,4
0,3
, 0,2
°0-0
Early intermediates in TNT degradation. The degradationof TNT by P. chrysosponum apparently started with thestepwise reduction of one nitro group to form the corre-sponding amino group via the nitroso and hydroxylaminocompounds. The first detectable intermediates in HPLCanalysis and TLC analysis were the hydroxylamino-dinitro-toluenes (Fig. 3B). In contrast to their formation from TNT,the further reduction of the hydroxylamino-dinitrotoluenesto the corresponding amino-dinitrotoluenes was muchslower. The resulting accumulation of the hydroxylamino-dinitrotoluenes was accompanied by an inhibition of theveratryl alcohol oxidase activity of lignin peroxidase (Fig.
5
FIG. 5. Effect of the veratryl alcohol concentration on its oxidation by lignin peroxidase H8 in the presence of hydroxylamino-dinitrotoluene. The reaction mixture (0.6 ml) contained 0.05 ,uM lignin peroxidase H8, 40 mM sodium tartrate buffer (pH 3.5), 260 ,uM H202,10 pLM hydroxylamino-dinitrotoluene, and various concentrations of veratryl alcohol (millimolar): 11.4 (A), 2.8 (-), 1.7 (0), 1.1 (A), 0.9 (Li),0.6 (V), and 1.1 (control [without hydroxylamino-dinitrotoluene]) (0). AE310 nm3, A310.
0 1 2 3 4
time [min]
VOL. 60, 1994
on May 17, 2018 by guest
http://aem.asm
.org/D
ownloaded from
192 MICHELS AND GOTTSCHALK
X0,66-
oo06 ..0,4
0,2-
0,00 5 10 15 20 25
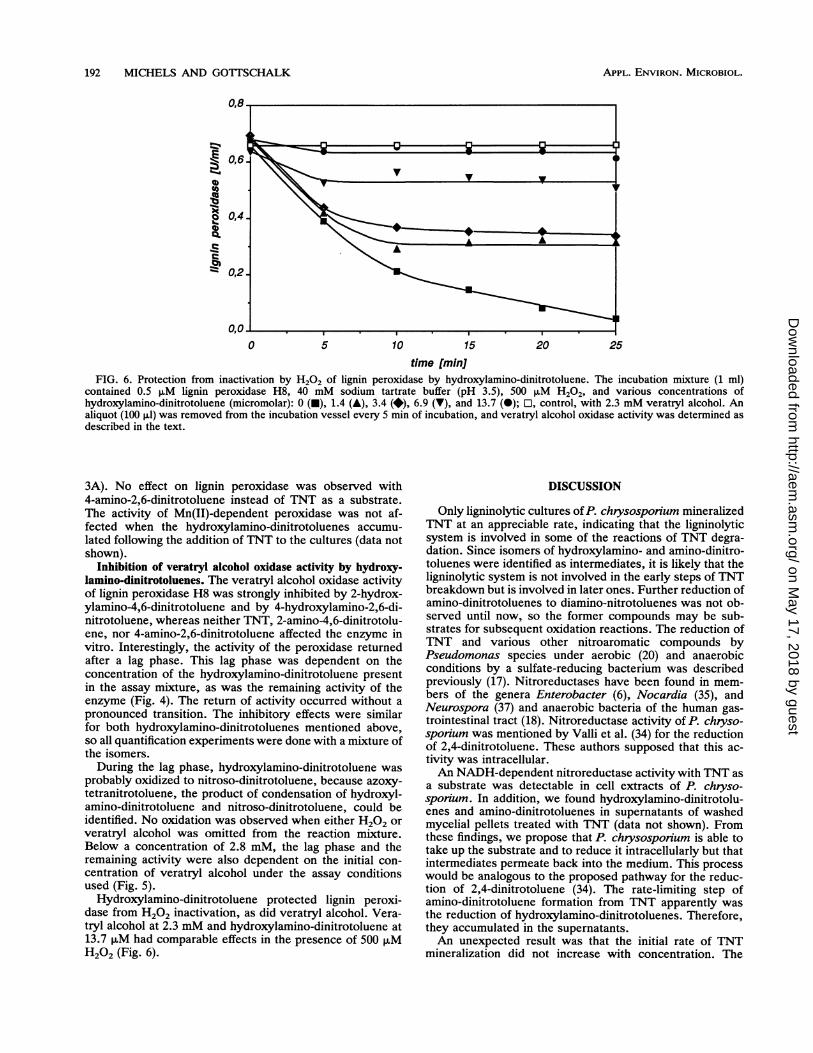
time [min]FIG. 6. Protection from inactivation by H202 of lignin peroxidase by hydroxylamino-dinitrotoluene. The incubation mixture (1 ml)
contained 0.5 ,uM lignin peroxidase H8, 40 mM sodium tartrate buffer (pH 3.5), 500 ,uM H202, and various concentrations ofhydroxylamino-dinitrotoluene (micromolar): 0 (-), 1.4 (A), 3.4 (*), 6.9 (V), and 13.7 (-); O, control, with 2.3 mM veratryl alcohol. Analiquot (100 ,ul) was removed from the incubation vessel every 5 min of incubation, and veratryl alcohol oxidase activity was determined asdescribed in the text.
3A). No effect on lignin peroxidase was observed with4-amino-2,6-dinitrotoluene instead of TNT as a substrate.The activity of Mn(II)-dependent peroxidase was not af-fected when the hydroxylamino-dinitrotoluenes accumu-lated following the addition of TNT to the cultures (data notshown).
Inhibition of veratryl alcohol oxidase activity by hydroxy-lamino-dinitrotoluenes. The veratryl alcohol oxidase activityof lignin peroxidase H8 was strongly inhibited by 2-hydrox-ylamino-4,6-dinitrotoluene and by 4-hydroxylamino-2,6-di-nitrotoluene, whereas neither TNT, 2-amino-4,6-dinitrotolu-ene, nor 4-amino-2,6-dinitrotoluene affected the enzyme invitro. Interestingly, the activity of the peroxidase returnedafter a lag phase. This lag phase was dependent on theconcentration of the hydroxylamino-dinitrotoluene presentin the assay mixture, as was the remaining activity of theenzyme (Fig. 4). The return of activity occurred without apronounced transition. The inhibitory effects were similarfor both hydroxylamino-dinitrotoluenes mentioned above,so all quantification experiments were done with a mixture ofthe isomers.
During the lag phase, hydroxylamino-dinitrotoluene wasprobably oxidized to nitroso-dinitrotoluene, because azoxy-tetranitrotoluene, the product of condensation of hydroxyl-amino-dinitrotoluene and nitroso-dinitrotoluene, could beidentified. No oxidation was observed when either H202 orveratryl alcohol was omitted from the reaction mixture.Below a concentration of 2.8 mM, the lag phase and theremaining activity were also dependent on the initial con-centration of veratryl alcohol under the assay conditionsused (Fig. 5).
Hydroxylamino-dinitrotoluene protected lignin peroxi-dase from H202 inactivation, as did veratryl alcohol. Vera-tryl alcohol at 2.3 mM and hydroxylamino-dinitrotoluene at13.7 puM had comparable effects in the presence of 500 puMH202 (Fig. 6).
DISCUSSION
Only ligninolytic cultures of P. chrysosponum mineralizedTNT at an appreciable rate, indicating that the ligninolyticsystem is involved in some of the reactions of TNT degra-dation. Since isomers of hydroxylamino- and amino-dinitro-toluenes were identified as intermediates, it is likely that theligninolytic system is not involved in the early steps of TNTbreakdown but is involved in later ones. Further reduction ofamino-dinitrotoluenes to diamino-nitrotoluenes was not ob-served until now, so the former compounds may be sub-strates for subsequent oxidation reactions. The reduction ofTNT and various other nitroaromatic compounds byPseudomonas species under aerobic (20) and anaerobicconditions by a sulfate-reducing bacterium was describedpreviously (17). Nitroreductases have been found in mem-bers of the genera Enterobacter (6), Nocardia (35), andNeurospora (37) and anaerobic bacteria of the human gas-trointestinal tract (18). Nitroreductase activity of P. chryso-sporium was mentioned by Valli et al. (34) for the reductionof 2,4-dinitrotoluene. These authors supposed that this ac-tivity was intracellular.An NADH-dependent nitroreductase activity with TNT as
a substrate was detectable in cell extracts of P. chryso-sporium. In addition, we found hydroxylamino-dinitrotolu-enes and amino-dinitrotoluenes in supernatants of washedmycelial pellets treated with TNT (data not shown). Fromthese findings, we propose that P. chrysosponum is able totake up the substrate and to reduce it intracellularly but thatintermediates permeate back into the medium. This processwould be analogous to the proposed pathway for the reduc-tion of 2,4-dinitrotoluene (34). The rate-limiting step ofamino-dinitrotoluene formation from TNT apparently wasthe reduction of hydroxylamino-dinitrotoluenes. Therefore,they accumulated in the supernatants.An unexpected result was that the initial rate of TNT
mineralization did not increase with concentration. The
APPL. ENvIRON. MICROBIOL.
on May 17, 2018 by guest
http://aem.asm
.org/D
ownloaded from

DEGRADATION OF TNT BY P. CHRYSOSPORIUM 193
same amounts were degraded in the concentration range of 5to 20 mg of TNT per liter, when the 14Co2 release wasexamined four days after the addition of the explosive. Thiswas not the case when amino-dinitrotoluene were used as asubstrate. This result strongly indicated that an intermediatewas inhibitory. This intermediate was identified as hydrox-ylamino-dinitrotoluene, as shown by inhibition studies of theveratryl alcohol oxidase activity of purified lignin peroxi-dase. The oxidation of veratryl alcohol was fully repressedfor a short period of time when hydroxylamino-dinitrotolu-ene was present in the reaction mixture. Thereafter, theactivity reappeared without any transition. To study theseresults in more detail, we tested the inhibition by usingvarious concentrations of the substrate and the inhibitor.Increasing the inhibitor concentration led to an increase inthe lag phase and some loss of the reappearing activity.Interestingly, veratryl alcohol was able to reduce the lagphase and to prevent the loss of activity. Known inhibitorsof the veratryl alcohol oxidase activity of lignin peroxidaseare often substrates of the enzyme. Sodium azide, a knowninhibitor, is oxidized (7, 33). 3-Amino-1,2,4-triazole, whichis mineralized by the fungus (32), is also an inhibitor of theveratryl alcohol oxidase activity of lignin peroxidase. EDTAis decarboxylated and H202 is consumed during inhibition ofthe veratryl alcohol oxidase activity (23). Also, like veratrylalcohol, oxalate, a secondary metabolite of P. chryso-sporium (4), is an inhibitor of the veratryl alcohol oxidaseactivity (1). The effect of cyanide, another inhibitor of ligninperoxidase (14), resembled that of hydroxylamino-dinitrotol-uene; after a short lag phase, the activity of the enzymeappeared again (23). Hydroxylamino-dinitrotoluene was ableto protect lignin peroxidase H8 from inactivation by H202, atypical effect for oxidizable substrates (2). The compoundwas only oxidized by the complete peroxidase system,consisting of lignin peroxidase, H202, and veratryl alcohol.The latter could function as a cation radical mediator in theoxidation of hydroxylamino-dinitrotoluene, as proposed forthe decarboxylation of oxalate and EDTA (22, 23). This rolewas proposed for free radical-catalyzed oxidations of chem-icals which are poor substrates for lignin peroxidase (10).The data presented here are important for the design of
bioremediation processes for TNT-contaminated soil with P.chrysosporium. Conditions under which hydroxylamino-di-nitrotoluenes do not accumulate must be found. If they do,corresponding nitroso compounds will be formed, as willazoxy compounds generated by condensation of the nitroso-and hydroxylamino-dinitrotoluenes. This result would be asevere disadvantage, because azoxy compounds are muchless soluble than TNT and are probably more resistant todegradation.
ACKNOWLEDGMENT
This work was supported by the Fonds der Chemischen Industrie.
REFERENCES
1. Akamatsu, Y., D. B. Ma, T. Higuchi, and M. Shimada. 1990. Anovel enzymatic decarboxylation of oxalic acid by the ligninperoxidase system of white-rot fungus Phanerochaete chryso-sponum. FEBS Lett. 269:261-263.
2. Arnao, M. B., M. Acosta, J. A. del Rio, and F. Garcia-Canovas.1990. Inactivation of peroxidase by hydrogen peroxide and itsprotection by a reductant agent. Biochim. Biophys. Acta 1038:85-89.
3. Aust, S. D., H. Tuisel, C.-W. Chang, and J. A. Bumpus. 1990.Oxidation of environmental pollutants by lignin peroxidases
from white rot fungi, p. 453-464. In C. A. Reddy, G. A.Hamilton, and K. M. Madyastha (ed.), Biological oxidationsystems, vol. I. Academic Press, Inc., New York.
4. Barr, D. P., M. M. Shah, T. A. Grover, and S. D. Aust. 1992.Production of hydroxyl radical by lignin peroxidase from Pha-nerochaete chrysosporium. Arch. Biochem. Biophys. 298:480-485.
5. Bonnarme, P., and T. W. Jeffries. 1990. Mn(II) regulation oflignin peroxidases and manganese-dependent peroxidases fromlignin-degrading white rot fungi. Appl. Environ. Microbiol.56:210-217.
6. Bryant, C., and M. DeLuca. 1991. Purification and characteriza-tion of an oxygen-insensitive NAD(P)H nitroreductase fromEnterobacter cloacae. J. Biol. Chem. 266:4119-4125.
7. DePillis, G. D., H. Wariishi, M. H. Gold, and P. R. Ortiz deMontellano. 1990. Inactivation of lignin peroxidase by phenyl-hydrazine and sodium azide. Arch. Biochem. Biophys. 280:217-223.
8. Faison, B. D., and T. K. KirL 1985. Factors involved in theregulation of a ligninase activity in Phanerochaete chryso-sponum. Appl. Environ. Microbiol. 49:299-304.
9. Fernando, T., J. A. Bumpus, and S. D. Aust. 1990. Biodegrada-tion of TNT (2,4,6-trinitrotoluene) by Phanerochaete chryso-sponum. Appl. Environ. Microbiol. 56:1666-1671.
10. Harvey, P. J., H. E. Schoemaker, and J. M. Palmer. 1986.Veratryl alcohol as a mediator and the role of radical cations inlignin biodegradation by Phanerochaete chrysosporium. FEBSLett. 195:242-246.
11. Jager, A., S. Croan, and T. K. Kirk. 1985. Production ofligninases and degradation of lignin in agitated submergedcultures of Phanerochaete chrysosporium. Appl. Environ. Mi-crobiol. 50:1274-1278.
12. Kirk, T. K., E. Schultz, W. J. Connors, L. F. Lorenz, and J. G.Zeikus. 1978. Influence of culture parameters on lignin metabo-lism by Phanerochaete chrysosponum. Arch. Microbiol. 117:277-285.
13. Klausmeier, R. E., J. L. Osmon, and D. R. Walls. 1973. Theeffect of trinitrotoluene on microorganisms. Dev. Ind. Micro-biol. 15:309-317.
14. Kuwahara, M., J. K. Glenn, M. A. Morgan, and M. H. Gold.1984. Separation and characterization of two extracellularH202-dependent oxidases from ligninolytic cultures of Phanero-chaete chrysosporium. FEBS Lett. 169:247-250.
15. Laemmli, U. K. 1970. Cleavage of structural proteins during theassembly of the head of bacteriophage T4. Nature (London)277:580-585.
16. Paszczynski, A., V.-B. Huynh, and R. Crawford. 1986. Compar-ison of ligninase-1 and peroxidase-M2 from the white-rot fungusPhanerochaete chrysosporium. Arch. Biochem. Biophys. 244:750-765.
17. Preuss, A., J. Fimpel, and G. Diekert. 1993. Anaerobic transfor-mation of 2,4,6-trinitrotoluene (TNT). Arch. Microbiol. 159:345-353.
18. Rafii, F., W. Franklin, R. H. Heflich, and C. E. Cerniglia. 1991.Reduction of nitroaromatic compounds by anaerobic bacteriaisolated from the human gastrointestinal tract. Appl. Environ.Microbiol. 57:962-968.
19. Ruggli, P., and H. Zaeslin. 1936. Uber zwei neue Dichlor-o-nitro-benzoesauren. Helv. Chim. Acta 19:434-439.
20. Schackmann, A., and R. Muller. 1991. Reduction of nitroaro-matic compounds by different Pseudomonas species underaerobic conditions. Appl. Microbiol. Biotechnol. 34:809-813.
21. Seidenfaden, W., and D. Pawellek 1971. Methoden zur Herstel-lung und Umwandlung von aromatischen Nitroverbindungen.Nitrierung von Alkyl-aromaten, p. 515-527. In E. Muller (ed.),Methoden der organischen Chemie (Houben-Weyl), 4th ed.,vol. X/I. Georg Thieme, Stuttgart, Federal Republic of Ger-many.
22. Shah, M. M., T. A. Grover, and S. D. Aust. 1991. Metabolism ofcyanide by Phanerochaete chrysosponum. Arch. Biochem.Biophys. 290:173-178.
23. Shah, M. M., T. A. Grover, D. P. Barr, and S. D. Aust. 1992. Onthe mechanism of inhibition of the veratryl alcohol oxidase
VOL. 60, 1994
on May 17, 2018 by guest
http://aem.asm
.org/D
ownloaded from

194 MICHELS AND GOTTSCHALK
activity of lignin peroxidase H2 by EDTA. J. Biol. Chem.267:21564-21569.
24. Sitzmann, M. E. 1974. Chemical reduction of 2,4,6-trinitrotolu-ene-initial products. J. Chem. Eng. Data 19:179-181.
25. Smock, L. A., D. L. Stoneburner, and J. R. Clark. 1976. Thetoxic effects of trinitrotoluene (TNT) and its primary degrada-tion products on two species of algae and fathead minnow.Water Res. 10:537-543.
26. Spiker, J. K., D. L. Crawford, and R. L. Crawford. 1992.Influence of 2,4,6,-trinitrotoluene (TNT) concentration on thedegradation of TNT in explosive-contaminated soils by thewhite rot fungus Phanerochaete chrysosporium. Appl. Environ.Microbiol. 58:3199-3202.
27. Stahl, J. D., and S. D. Aust. 1993. Plasma membrane dependentreduction of 2,4,6-trinitrotoluene by Phanerochaete chryso-sporium. Biochem. Biophys. Res. Commun. 192:471-476.
28. Stahl, J. D., and S. D. Aust. 1993. Metabolism and detoxificationof TNT by Phanerochaete chrysosporium. Biochem. Biophys.Res. Commun. 192:477-482.
29. Sublette, K. L., E. V. Ganapathy, and S. Schwartz. 1992.Degradation of munition wastes by Phanerochaete chryso-sporium. Appl. Biochem. Biotechnol. 34/35:709-723.
30. Tatsumi, K., A. Inoue, and H. Yoshimura. 1981. Mode of
APPL. ENVIRON. MICROBIOL.
reactions between xanthine oxidase and aromatic nitro com-pounds. J. Pharm. Dyn. 4:101-108.
31. Tien, M., and T. K. Kirk. 1988. Lignin peroxidase of Phanero-chaete chrysosporium. Methods Enzymol. 161:238-249.
32. Tuisel, H., T. A. Grover, J. A. Bumpus, and S. D. Aust. 1992.Inhibition of veratryl alcohol oxidase activity of lignin peroxi-dase H2 by 3-amino-1,2,4-triazole. Arch. Biochem. Biophys.293:287-291.
33. Tuisel, H., T. A. Grover, J. R. Lancaster, J. A. Bumpus, andS. D. Aust. 1991. Inhibition of lignin peroxidase H2 by sodiumazide. Arch. Biochem. Biophys. 288:456-462.
34. Valli, K., B. J. Brock, D. K. Joshi, and M. H. Gold. 1992.Degradation of 2,4-dinitrotoluene by the lignin-degrading fungusPhanerochaete chrysosporium. Appl. Environ. Microbiol. 58:221-228.
35. Villanueva, J. R. 1964. The purification of a nitroreductase fromNocardia V. J. Biol. Chem. 239:773-776.
36. Won, W. D., L. H. Disalvo, and J. Ng. 1976. Toxicity andmutagenicity of 2,4,6-trinitrotoluene and its microbial metabo-lites. Appl. Environ. Microbiol. 31:576-580.
37. Zucker, M., and A. Nason. 1955. Nitroaryl reductase fromNeurospora crassa. Methods Enzymol. 2:406-411.
on May 17, 2018 by guest
http://aem.asm
.org/D
ownloaded from





























