Embed Size (px)
Citation preview

Inhibition of the Growth of Premalignant and Malignant Human OralCell Lines by Extracts and Components of Black Raspberries
ChunHua Han, Haiming Ding, Bruce Casto, Gary D. Stoner, and Steven M. D’Ambrosio
Abstract: Black raspberries are a rich natural source ofchemopreventive phytochemicals. Recent studies have shownthat freeze-dried black raspberries inhibit the development oforal, esophageal, and colon cancer in rodents, and extractsof black raspberries inhibit benzo(a)pyrene-induced celltransformation of hamster embryo fibroblasts. However, themolecular mechanisms and the active components responsi-ble for black raspberry chemoprevention are unclear. In thisstudy, we found that 2 major chemopreventive components ofblack raspberries, ferulic acid and �-sitosterol, and a frac-tion eluted with ethanol (RO-ET) during silica column chro-matography of the organic extract of freeze-dried black rasp-berries inhibit the growth of premalignant and malignant butnot normal human oral epithelial cell lines. Another fractioneluted with CH2Cl2/ethanol (DM:ET) and ellagic acid inhib-ited the growth of normal as well as premalignant and malig-nant human oral cell lines. We investigated the molecularmechanisms by which ferulic acid and �-sitosterol and theRO-ET fraction selectively inhibited the growth ofpremalignant and malignant oral cells using flow cytometryand Western blotting of cell cycle regulatory proteins. Therewas no discernable change in the cell cycle distribution fol-lowing treatment of cells with the RO-ET fraction.Premalignant and malignant cells redistributed to the G2/Mphase of the cell cycle following incubation with ferulic acid.�-sitosterol treated premalignant and malignant cells accu-mulated in the G0/G1 and G2/M phases, respectively. TheRO-ET fraction reduced the levels of cyclin A and cell divi-sion cycle gene 2 (cdc2) in premalignant cells and cyclin B1,cyclin D1, and cdc2 in the malignant cell lines. This fractionalso elevated the levels of p21waf1/cip1 in the malignant cellline. Ferulic acid treatment led to increased levels of cyclinB1 and cdc2 in both cell lines, and p21waf1/cip1 was induced inthe malignant cell line. �-sitosterol reduced the levels ofcyclin B1 and cdc2 while increasing p21waf1/cip1 in both thepremalignant and malignant cell lines. These results showfor the first time that the growth inhibitory effects of blackraspberries on premalignant and malignant human oral cells
may reside in specific components that target aberrant sig-naling pathways regulating cell cycle progression.
Introduction
The consumption of fruits and vegetables has been associ-ated with a reduced risk for many different types of humancancer (1–3). A high consumption of fruits is associated witha large reduction in the risk for cancers of the oral cavity andpharynx (1). The protective effect of fruits and vegetables isrelated to multiple anticancer components including dietaryfiber, carotenoids, vitamins C and E, selenium,glucosinolates, indoles, isothiocyanates, flavonoids,polyphenols, protease inhibitors, and plant sterols. Amongfruits, black raspberries have a relatively large content ofthese chemopreventive agents (4–12). Freeze-dried blackraspberries inhibit N-nitrosomethylbenzylamine (NMBA)induced tumorigenesis in the rat esophagus (13,14), reducingby 40–60% esophageal tumor multiplicity, when adminis-tered at dietary concentrations of 5% and 10% before, dur-ing, and after treatment of the rats with NMBA (15). Anotherstudy showed that black raspberries given postinitiation in-hibited azoxymethane-induced tumors in the rat colon, indi-cating that berries are effective in inhibiting the progressionstages of cancer (16). Recently, the feeding of lyophilizedblack raspberries to hamsters significantly reduced the num-ber of 7,12-dimethylbenz(a)anthracene-induced tumors inthe cheek pouch (5). These animal studies have indicated thatthe phytochemicals in berries can inhibit multiple types ofcancer during both the initiation and promotion stages. In vi-tro studies using a Syrian hamster embryo transformation as-say showed that a methanol extract of black raspberries in-hibited benzo(a)pyrene diol-epoxide-induced celltransformation (17). The same organic fraction of black rasp-berries inhibited benzo(a)pyrene diol-epoxide-inducedtransactivation of AP-1 and NFκB in mouse epidermal JB6CI 41 cells (18). These studies have suggested that selective
NUTRITION AND CANCER, 51(2), 207–217Copyright © 2005, Lawrence Erlbaum Associates, Inc.
C. Han, H. Ding, and S. M. D’Ambrosio are affiliated with the Department of Radiology, The Ohio State University, Columbus, OH 43210. B. Casto, G. D.Stoner, and S. M. D’ Ambrosio are affiliated with The Comprehensive Cancer Center, The Ohio State University, Columbus, OH 43210. B. Casto is also affili-ated with the Division of Environmental Health, School of Public Health, The Ohio State University, Columbus, OH 43210. G. Stoner is also affiliated with theDepartment of Internal Medicine, The Ohio State University, Columbus, OH 43210. S. M. D’Ambrosio is also affiliated with the Department of Pharmacology,College of Medicine, The Ohio State University, Columbus, OH 43210.

phytochemicals may represent the active chemopreventivecomponents of the berries. They also provide a rationale forcombining identified phytochemicals to mimic active berryextracts. Although it is clear that whole berries and berry ex-tracts elicit significant chemopreventive effects, the compo-nents and mechanisms by which the berries and their compo-nents inhibit multiple stages of cancer are not wellunderstood.
In the United States, cancer of the oral cavity is one of the10 most common cancers representing 2–3% of all cancers inmen and women (19,20). Of these cancers, 90% aresquamous cell carcinomas, and the prognosis is poor, with anoverall mortality rate of approximately 50%. Because currenttherapeutic protocols are relatively ineffective for oral can-cer, it is important to identify and develop chemopreventivestrategies to prevent this disease. Although most chemo-preventive studies in the oral cavity have focused on demon-strating the inhibition of tumor formation, few studies havedetermined the effects in normal cell phenotypes and themechanisms of chemopreventive activity in premalignantand malignant oral cell types. It is important to include nor-mal human cells in these studies to better identify and under-stand molecular mechanisms leading to agents that selec-tively target premalignant and malignant cells.
To determine whether black raspberries and their compo-nents are effective chemopreventive agents in human oralcancer, we used a previously described human oral cancercell model consisting of primary normal cells andpremalignant and malignant human oral cell lines. Our re-sults show that ferulic acid and β-sitosterol and a fraction offreeze-dried berries (eluted from a silica gel column with eth-anol) inhibit the growth of both premalignant and malignantbut not normal human oral cell lines. Growth inhibition wasdue to inhibition of cell cycle regulatory proteins leading tothe blocking of cells in the G0/G1 (β-sitosterol inpremalignant cell line) and G2/M (ferulic acid inpremalignant and malignant and β-sitosterol in malignantcell lines) phases of the cell cycle.
Materials and Methods
Cell Lines and Culture
The previously described (21) in vitro model consisting ofthe normal (TE1177), premalignant (SCC-83-01-82), andmalignant (83-01-82CA) human oral epithelial cell lines wasused in this study. The characteristics of the premalignantand malignant oral cell lines (both kindly provided by Dr G.Milo of The Ohio State University) have been previously de-scribed (22). The cell lines were maintained in modified min-imal essential medium (MEM) supplemented with 10% fetalbovine serum, 5 U/ml penicillin, and 5 µg/ml streptomycin(23). TE1177 cells were grown in a modified alpha MEMmedium supplemented with epithelial growth factor, insulin,transferrin, hydrocortisone, and 10% fetal bovine serum.
Black Raspberry Extract, Fractions,and Components
Black raspberries [Rubus occidentalis (RO), Jewel vari-ety) were supplied by the Stokes Raspberry Farm(Wilmington, OH) and shipped frozen to Van Drunen Farms(Momence, IL) for freeze-drying. The composition of theberry powder was determined by Covance Laboratories(Madison, WI) and has been previously described (4,5).Black raspberry extracts were prepared and fractionated intoRO-F001, RO-F003, RO-F004, RO-DM, and RO-ET frac-tions as previously described (17,18) except that ethanol wassubstituted for methanol in the extraction protocol. Ellagicacid, ferulic acid, β-sitosterol, and 2-hydroxypropyl-β-cyclodextrin (HBC) were purchased from Sigma (St.Louis, MO). The β-sitosterol-cyclodextrin complex was pre-pared according to the method of Greenberg et al. (24). Themolar ratio of β-sitosterol to cyclodextrin was maintained at1:300.
Growth Inhibition Assay
Normal human oral cells were seeded at 300 cells, andboth premalignant and malignant cells were seeded at 200cells per well in 96-well plates 24 h prior to treatment. Blackraspberry fractions, ellagic acid, ferulic acid, and β-sitosteroland their respective solvents were added to the culture at theindicated concentrations. The concentrations of water,dimethyl sulfoxide (DMSO; <0.2%), ethanol (<0.4%), andHBC (<4.5 mM) used as solvents did not inhibit the growthof the cell lines. On Day 5, the medium was replenished withfresh medium-containing agent. On Day 7, the medium wasremoved and the cells washed twice with phosphate-bufferedsaline (PBS), air dried, and fixed with 4% paraformaldehyde.A 100 µl solution containing 1% methylene blue was addedfor 30 min to stain the cells. The dye was extracted with 100µl of methanol/glacial acetic acid/water (100:1:99) solution,and the optical density was measured at 660 nm using aBioTex microplate reader (Houston, TX). The number ofcells following treatment was compared to the solvent andcontrol groups and used to determine growth inhibition asfollows: GI = (Abtreated/Abcontrol) × 100, where Ab representsthe mean absorbance. Mean values ± standard error of mea-surement (SEM) were calculated from 4–6 experiments. Stu-dent’s 2-tailed t-test was used to determine significance.Cytotoxicity was assayed following 24 h of treatment withagent using a 3-(4,5-dimethylthiazol-2-yl)-2,5-diphenylte-trazolium bromide (MTT) assay. In this assay, normal humanoral cells were seeded at 7,000, and both premalignant andmalignant cells were seeded at 4,000 cells per well in 96-wellplates. MTT was added at a final concentration of 0.5 mg/mland incubated during the 20th to 24th h of treatment. Thesupernatant was removed, and the cells were washed withPBS twice, dried in air, and 100 µl DMSO was added. Theoptical density was measured at 570 nm. A decrease in MTTactivity compared to solvent and untreated controls was usedas an indicator and measure of cytotoxicity.
208 Nutrition and Cancer 2005

Cell Cycle Analyses Using Flow Cytometry
Cells (5–10 × 104 per 75 cm2 flask) were treated on the2nd and 5th days with 200 µg/ml of either the berry RO-ETfraction or with DMSO (solvent control). Cells were treatedwith 200 µg/ml ferulic acid, ethanol (solvent control), 15 µMβ-sitosterol, and HBC (solvent control) for 3 days. At the endof treatment, cells were harvested with a 0.01% trypsin/0.1mM ethyleneglycotetraacetic acid (EGTA) solution, fixed in70% ethanol and stored at –20°C as previously described(25). Prior to fluorescence-activated cell sorter analyses, thefixed cells were washed with PBS and incubated with a solu-tion containing 20 µg/ml propidium iodide, 200 µg/mlribonuclease A, 0.1% Triton-x100 for 30 min in the dark.Flow cytometry analysis was performed by the AnalyticalCytometry Laboratory of The Ohio State University Com-prehensive Cancer Center using a Beckman-Coulter EPICSElite flow Cytometer (BD FACSCalibur, Bedford, MA)equipped with a 488 nm 15 mW air-cooled Argon laser col-lecting at least 30,000 events. Multicycle DNA modelingsoftware and Modfit software (ver. 3.0, Verity SoftwareHouse, Topsham, ME) were used to estimate cell cycle distri-bution. The DNA profile indicated the relative abundance ofthe cell population in subG0, G0/G1, S, and G2/M phases. Thecoefficient of variance and χ2 value were used to determinesignificance of the distribution of cells in each phase of thecell cycle.
Western Blot Analyses
Cells were treated with RO-ET, ferulic acid, β-sitosterol,and their corresponding solvent controls in the same concen-trations and times described previously for flow cytometry.Immediately following treatment, cells were washed twicewith PBS and removed from the surface of the flask using a0.01% Trypsin/0.1 mM EGTA solution. Cells were collectedby centrifugation, washed twice with PBS, and lysed by boil-ing for 10 min in a buffer containing 2% sodium dodecyl sul-fate (SDS), 10% glycerol, 10 mM dithio-DL-threitol, 62 mMTris-HCl pH 6.8, 10 µg/ml pepstatin, and 1 mg/ml leupeptin.Cell debris was removed by centrifugation and protein con-centration determined using the Bio-Rad assay (Hercules,CA). Equal amounts (30–70 µg) of protein were subjected togel electrophoresis in 12% polyacrylamide slab gels(InVitrogen, Carlsbad, CA) in the presence of SDS. The pro-tein was electrophoretically transferred to a nitrocellulosemembrane and dried. The membrane was preincubated withblocking buffer (5% nonfat milk in TBST buffer; 150 mMNaCl, 10 mM Tris, and 0.1% Tween-20) for 2 h and then in-cubated with primary antibody at 4°C overnight. The pri-mary antibodies used were rabbit polyclonal anti-cyclin D1(Cell Signaling Technology, Beverly, MA; 1:500), mousemonoclonal anti-tubulin (Oncogene Research Products, SanDiego, CA; 1:1000), rabbit polyclonal anti-cyclin A andanti-p21waf1/cip1, and mouse monoclonal anti-cyclin B1(Santa Cruz Biotechnology, Santa Cruz, CA; 1:1000). Afterwashing the membrane three times with TBST for 10 min,
the membrane was incubated with mouse or rabbit secondaryantibody labeled with horseradish peroxidase for 1 h at 37°C.The Western blots were developed using achemiluminescence kit (Amersham Pharmacia Biotech,Uppsala, Sweden). The membrane was then exposed toX-ray film and the image documented and the relative den-sity measured using AlphaEase software from AlphaInnotech Corporation (San Leandro, CA). α-Tubulin or actin(used as loading controls) was detected on the same mem-brane following incubation with a stripping buffer (20 mMTris-HCl at pH 7.4, 2% SDS, 500 mM NaCl, 100 mM2-mercaptoethanol) for 1 h at 37°C.
Statistical Analyses
Each experiment was performed three to four times as in-dicated in the legends. Where appropriate, the data are pre-sented as the mean ± SEM. Student’s t-test was used to assessthe statistical significance of the differences between thecontrol and treatment groups. A statistically significant dif-ference was considered to be P < 0.05.
Results
Black Raspberry Fractionsand Components Selectively Inhibitthe Growth of Premalignantand Malignant Oral Cell Lines
To identify the active chemopreventive components inblack raspberries, previously described fractions of raspberryextracts and identified components of lyophilized berrieswere tested for growth inhibition of normal, premalignant,and malignant oral cell lines (4,17,18). The RO-F001 frac-tion, isolated following overnight extractions of thefreeze-dried berries with ethanol three times, was not growthinhibitory to the oral cell lines at its highest soluble concen-tration for 6 days. The water soluble RO-F003 fraction andthe CH2Cl2 soluble RO-F004 fraction isolated following par-titioning of the RO-F001 fraction with CH2Cl2 and waterwere also inactive. Following chromatography of theRO-F001 fraction on a silica gel column, the RO-DM(CH2Cl2:ethanol, 1:1 eluate) and RO-ET (ethanol eluate)fractions were obtained. At the 200 µg/ml concentration, theRO-DM fraction inhibited 80% of normal, premalignant, andmalignant cell growth (Table 1). In contrast to the unselectivegrowth inhibition by the RO-DM fraction, the RO-ET frac-tion selectively inhibited the growth of the premalignant andmalignant cell lines 45% (P < 0.001) and 27% (P < 0.005),respectively, at 200 µg/ml and 35% (P < 0.05) and 25% (P <0.05), respectively, at 100 µg/ml and the premalignant cellline by 23% (P < 0.05) at 50 µg/ml (Table 1). There was nodiscernable growth inhibition of the normal cell line. TheRO-DM and RO-ET fractions were not cytotoxic to the celllines as determined by MTT and trypan blue dye exclusionassays (data not shown).
Vol. 51, No. 2 209

The chemical compositions of cultivars of black raspber-ries used in previously published animal chemopreventionstudies indicate that varying levels of sterols, polyphenols,and carotenoids are present that have demonstratedchemopreventive activity (5,15). Among these, ellagic acid(166–200 mg/100g), ferulic acid (18–21 mg/100g), andβ-sitosterol (72–89 mg/100g) were found in high quantities.To determine the contribution of these components to thegrowth-inhibitory activity of the berries, the oral cell lineswere treated with various concentrations of these three com-ponents over a 6-day period for growth inhibition. Ellagicacid (as ellagitannins in the insoluble residue after the initialethanol extraction) inhibited the growth of all three cell lines,producing 50% growth inhibition at the 25 µg/ml concentra-tion (Fig. 1A). Ellagic acid was also observed to be cytotoxicto the cell lines, which presumably accounts for its growth in-hibition. Ferulic acid was a selective growth inhibitor of boththe premalignant and malignant oral cell lines (Fig. 1B). Atthe highest concentration (200 µg/ml) tested, ferulic acid sig-nificantly (P < 0.05) reduced the growth of premalignant andmalignant oral cells by 40% and 26%, respectively, with nodiscernable effect on the normal cell line. Ferulic acid wasnot cytotoxic to the cell lines. There was a significant (P <0.05) difference in the growth inhibition of the normal (26%)and malignant (42%) oral cell lines following treatment with15 µM β-sitosterol (Fig. 1C). β-Sitosterol caused a concen-tration dependent growth inhibition of the premalignant cellline, with the 15 µM concentration yielding 80% growth in-hibition. The difference between the premalignant and thenormal and malignant cell line was significant at the 15, 10,and 5 (P < 0.001) and 2.5 and 0.5 (P < 0.05) µM concentra-tions. β-Sitosterol was not cytotoxic to the cells.
Ferulic Acid and β-Sitosterol Induce CellCycle Arrest in Premalignantand Malignant Cells
Flow cytometry was used to gain further insight into themechanisms of black raspberry chemoprevention and the
selective growth inhibiting effects of the RO-ET fraction,ferulic acid, and β-sitosterol. Figure 2 compares the cell cy-cle distribution patterns in the premalignant and malignantoral cell lines following incubation with the RO-ET fractionfor 6 days and either ferulic acid or β-sitosterol for 3 days.Although the RO-ET fraction induced significant growthinhibition of both the premalignant and malignant celllines, there was no significant difference in the cell cycledistribution between the DMSO control and RO-ET frac-tion (Fig. 2A). Although there was no difference in the cellcycle distribution following incubation with DMSO andHBC solvent, ethanol, used as the solvent for ferulic acid,increased the number of cells in the G2/M phase. Comparedto the ethanol solvent control, treatment of the cell lineswith 200 µg/ml ferulic acid led to a further accumulation of
210 Nutrition and Cancer 2005
Table 1. Growth Inhibitory Effects of the RO-DM (DM)and RO-ET (ET) Berry Fractions on Normal, Premalignant,and Malignant Oral Cell Linesa,b
Fraction Normal Premalignant Malignant
DMc
50 17.0 ± 0.6 4.7 ± 0.2 –0.3 ± 0.0100 57.2 ± 8.3 14.4 ± 0.1** 21.4 ± 1.1**200 78.2 ± 1.1 79.4 ± 4.2 79.2 ± 3.8
ETb
50 3.1 ± 0.1 23.3 ± 0.9# 4.7 ± 0.2100 –4.5 ± 0.2 35.2 ± 0.8# 24.7 ± 1.2#
200 –7.3 ± 0.3 45.1 ± 2.2* 27.4 ± 0.9**
a: Data are expressed as the percent mean ± standard error of measurementof solvent control from four to six replicates.
b: Statistical significance is as follows: *, P < 0.001; **, P < 0.005; and #, P< 0.05; indicates significant differences from the normal cell line.
c: µg/ml, treated twice over 6 days as described in Materials and Methods.
Figure 1. Effect of the ellagic acid, ferulic acid, and β-sitosterol on thegrowth of normal, premalignant, and malignant human oral cell lines. Cellswere treated with various concentration of A: ellagic acid in water, B: ferulicacid in ethanol, and C: β-sitosterol-2-hydroxypropyl-β-cyclodextrin com-plex and the corresponding solvents for 6 days. � (solid line), normal; �
(dash line), premalignant; and � (dot line), malignant cell lines. The relativenumber of cells was determined using a methylene blue assay as described inMaterials and Methods. Results are expressed as a percentage of growth,with 100% representing control cells. Data represent the mean ± SEM offour to six experiments. *, P < 0.001; **, P < 0.005; and #, P < 0.05 indicatessignificant difference from the normal cell line.

Vol. 51, No. 2 211
Figure 2. Cell cycle distribution of premalignant and malignant oral cell lines following incubation with the RO-ET berry fraction, ferulic acid (FA), andβ-sitosterol (SIT). Flow cytometry analyses and cell cycle distribution of cells treated with A: 0.2% dimethyl sulfoxide (DMSO) and 200 µg/ml RO-ET (ET) for6 days, B: 0.4% ethanol and 200 µg/ml FA for 72 h, and C: 4.5 mM 2-hydroxypropyl-β-cyclodextrin (HBC) and 15 µM SIT) for 72 h. Data are representative ofthree independent experiments. Dotted shading indicates cells in G1, hatched shading denotes cells in S, and solid shading are cells in the G2/M (G2) phase of thecell cycle. #P < 0.05 indicates significant increase from solvent control.

both the premalignant (58.6% vs 24.7%) and malignant(75.0% vs 30.2%) cell lines in the G2/M phase of the cellcycle (Fig. 2B). There was a corresponding decrease in thenumber of cells in the G0/G1 phase (16.8% vs 28.6% inpremalignant cells and 7.6% vs 29.9% in malignant cells).These results suggest that ferulic acid arrests both thepremalignant and malignant oral cell lines in the G2/Mphase of the cell cycle. Compared to the HBC-treated con-trol, 15 µM β-sitosterol treatment led to a moderate accu-mulation of cells in the G0/G1 phase of the cell cycle in thepremalignant cell line (56.3% vs 47.7%; Fig. 2C). How-ever, treatment of the malignant cell lines with β-sitosterolled to a marked depletion of cells in the G0/G1 (43.7% vs26.2%) and corresponding increases in the G2/M (18.2% vs34.3%) phases of the cell cycle.
Effect of the RO-ET Berry Fraction,Ferulic Acid, and β-Sitosterolon Cell Cycle Regulatory Proteinsin the Human Oral Cell Lines
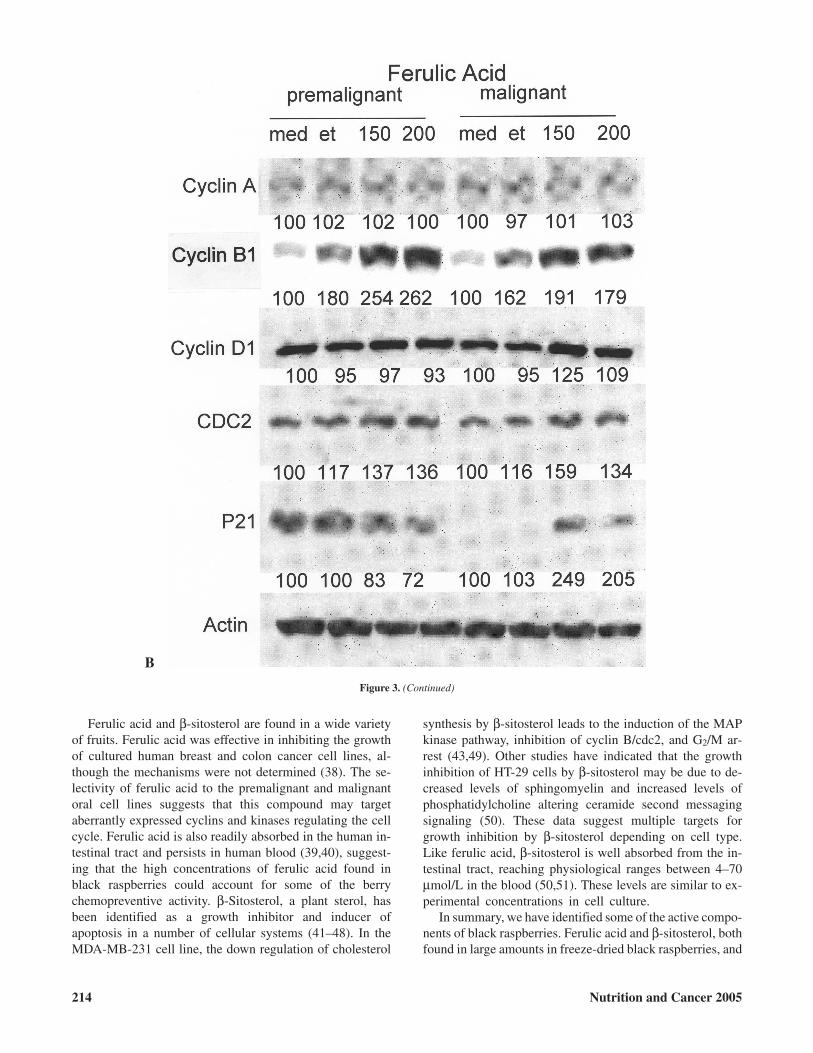
Because ferulic acid and β-sitosterol redistributed thepremalignant and malignant cells in either the G0/G1 orG2/M phases of the cell cycle, we evaluated their effectsand the effects of the RO-ET fraction on proteins that regu-late progression through the cell cycle. Although there wasno discernible change in the cell cycle distribution ofpremalignant and malignant cells following treatment for 6days with 200 µg/ml of the berry RO-ET fraction, a numberof changes were noted in the levels of cyclins, cell divisioncycle gene 2 (cdc2), and p21waf1/cip1 (Fig. 3A). In thepremalignant cell line, cdc2 protein levels were reduced30% by the RO-ET fraction, and cyclin A was slightly(17%) reduced. There were increases in the levels of cyclinB1 (11%) and cyclin D1 (48%), whereas p21waf1/cip1 re-mained unchanged. When the malignant cell line wastreated with the RO-ET fraction, the levels of cyclin A ap-peared unchanged, whereas there were decreases in the lev-els of cyclin B1 (33%), cyclin D1 (28%), and cdc2 (30%)and increased (24%) levels of p21waf1/cip1. The changes inthe levels of these cell cycle regulatory proteins were muchlarger in the cell lines treated with ferulic acid (Fig. 3B)and β-sitosterol (Fig. 3C) than with the RO-ET fraction.This may be expected due to the higher concentrations ofthese two chemicals used in these experiments compared tothat found in the black raspberry extracts. As described pre-viously, treatment of both the premalignant and malignantcell lines with ferulic acid led to an accumulation of cells inthe G2/M phase of the cell cycle. In both the premalignantand malignant cell lines, ferulic acid increased the levels ofthe G2/M proteins, cyclin B1 (45%, 19%), and cdc2 (16%,37%; Fig. 3B). The levels of cyclin A, induced during the Sphase, and cyclin D1, required for the transition of cellsinto the G1 phase, were not affected by ferulic acid.p21waf1/cip1, an inhibitor of the cdk/cyclin complex, was re-duced 28% in the premalignant and increased twofold inthe malignant cell line. β-Sitosterol, which increased the
number of premalignant and malignant cells in the G0/G1
and G2/M phase of the cell cycle, respectively, inhibited theexpression of cyclin B1 (38%, 73%) and cdc2 (39%, 40%)and increased p21waf1/cip1 (39%, 287%) levels in both thepremalignant and malignant cell lines (Fig. 3C). Cyclin D1was 36% lower in the premalignant and 45% higher in themalignant cell lines following treatment. Cyclin A was notaffected by β-sitosterol.
Discussion
Theconsumptionof fruits andvegetableshasbeenstronglyassociated with the reduced risk of many different types of hu-man cancers including the oral cavity and pharynx (1–3,26).The identification of fruits and vegetables and their compo-nents with chemopreventive activity provides a “food-based”approach toward chemoprevention. Among the fruits, blackraspberries have demonstrated chemopreventive effects in an-imal models, and clinical trials are currently being initiated forprevention of oral, esophageal, and colon cancer(4,5,14–16,18). Black raspberries contain large amounts ofcompounds with demonstrated chemopreventive activity in-cluding vitamins, selenium, anthocyanins, β-sitosterol,ellagic acid, and ferulic acid (5,15). However, there is little in-formation as to the active components and mechanisms ofchemoprevention. In this study, we observe that among the sixfractions isolated from freeze-dried berries, only the fractioneluted with ethanol (RO-ET) from the silica gel chromatogra-phy of the F001 fraction exhibited selective growth inhibitionof premalignant and malignant human oral cell lines. Of themany components identified in lyophilized black raspberries,ferulic acid and β-sitosterol were selective inhibitors ofpremalignant and malignant oral cell line growth. Althoughtheseare found inhighquantities in the freezedriedberries (5),only small amounts (<5 µg/g) are observed in the RO-ET frac-tion.Growth inhibitionbyferulicacidandβ-sitosterolwasdueto their modulation of key signaling proteins regulating theprogression of cells through the cell cycle. Ferulic acid treat-ment led to increased levels of cyclin B1 and cdc2 in both celllines and higher levels of p21waf1/cip1 in the malignant cell line.β-Sitosterol reduced the levels of cyclin B1 and cdc2 while in-creasing p21waf1/cip1 in both the premalignant and malignantcell lines. These results indicate that the chemopreventive ef-fect of black raspberries may reside in specific componentstargeting cell cycle regulatory signaling pathways in humanpremalignant and malignant oral cells.
The lack of observable cell cycle delay by the RO-ET ex-tract may account for its modest effect on growth inhibition ofthe premalignant and malignant cell lines. However, we wereable to observe RO-ET induced reductions in cdc2 protein inboth the premalignant and malignant cell lines, cyclin B1 andcyclin D1 proteins in the malignant cell line, and elevation ofp21waf1/cip1 protein in the malignant cell line. These effects areindependent of ferulic acid and β-sitosterol, as these two com-pounds are found in very low (<5 µg/g) quantities in theRO-ET extract. Studies are ongoing to identify the active com-
212 Nutrition and Cancer 2005

ponentsof theRO-ETextract.The lackofgrowth inhibitionbythe first ethanol extraction, RO-F001, and the other organicand aqueous fractions suggests that these may containsuboptimal levels of growth-inhibiting chemicals or the pres-ence of growth-stimulatory compounds. This may be revealedwith further chemical, biological, and molecular analyses ofthe fractions.
The delays in the progression of the premalignant and ma-lignant cells through the cell cycle by ferulic acid andβ-sitosterol were correlated with changes in the levels of pro-teins regulating cell cycle progression. Our previous study andthis study have indicated that cyclin A, cyclin B1, and cdc2 areexpressed at high and p21waf1/cip1 at low levels in both thepremalignant and malignant oral cell lines compared to thenormal oral epithelial cell line (25). These are consistent with
other studies that have shown elevated levels of cyclins andcyclin kinases and low levels of cyclin kinase inhibitors inmost head and neck tumor cells (27–32). The treatment ofpremalignant and malignant oral cell lines with ferulic acid in-creased the levels of cdc2 and cyclin B1 proteins, which mayfacilitate the movement of cells from S into the G2 phase of thecell cycle. However, they were not able to go through mitosis.The accumulation of cells in either the G0/G1 or G2/M phasesfollowing β-sitosterol treatment may be due to increases in thelevel of p21waf1/cip1 inhibiting the cdk2/cyclin E complex forexiting G1 and cdc2/cyclin B complex for progression throughG2 into mitosis (33–37). Further studies are needed to deter-mine possible inhibitory interactions of these compounds di-rectly or indirectly via upstream signals regulating cyclins andcyclin-associated kinases.
Vol. 51, No. 2 213
Figure 3. Expression of cell cycle regulatory proteins in premalignant and malignant human oral cell lines treated with the RO-ET berry fraction, ferulic acid,and β-sitosterol. Cells were treated with A: 0.2% dimethyl sulfoxide (DMSO) and 200 µg/ml RO-ET (ET) for 6 days; B: medium (med), 0.4% ethanol (et), and150 and 200 µg/ml ferulic acid for 72 h; and C: med, 4.5 mM 2-hydroxypropyl-β-cyclodextrin (HBC), and 15 µM β-sitosterol (sit) for 72 h. Cell lysates wereevaluated for the levels of cyclin A, cyclin B1, cyclin D1, cell division cycle gene 2 (cdc2), and p21waf1/cip1 by Western blotting as described in Materials andMethods. The blots are representative of three independent experiments. The blots were scanned using Scan Maker 5900 (Alpha Innotech, San Leandro, CA),and the density was quantified using AlphaEase software (ver. 3.0). The values for the medium control were set to 100.
A
Ferulic acid and β-sitosterol are found in a wide varietyof fruits. Ferulic acid was effective in inhibiting the growthof cultured human breast and colon cancer cell lines, al-though the mechanisms were not determined (38). The se-lectivity of ferulic acid to the premalignant and malignantoral cell lines suggests that this compound may targetaberrantly expressed cyclins and kinases regulating the cellcycle. Ferulic acid is also readily absorbed in the human in-testinal tract and persists in human blood (39,40), suggest-ing that the high concentrations of ferulic acid found inblack raspberries could account for some of the berrychemopreventive activity. β-Sitosterol, a plant sterol, hasbeen identified as a growth inhibitor and inducer ofapoptosis in a number of cellular systems (41–48). In theMDA-MB-231 cell line, the down regulation of cholesterol
synthesis by β-sitosterol leads to the induction of the MAPkinase pathway, inhibition of cyclin B/cdc2, and G2/M ar-rest (43,49). Other studies have indicated that the growthinhibition of HT-29 cells by β-sitosterol may be due to de-creased levels of sphingomyelin and increased levels ofphosphatidylcholine altering ceramide second messagingsignaling (50). These data suggest multiple targets forgrowth inhibition by β-sitosterol depending on cell type.Like ferulic acid, β-sitosterol is well absorbed from the in-testinal tract, reaching physiological ranges between 4–70µmol/L in the blood (50,51). These levels are similar to ex-perimental concentrations in cell culture.
In summary, we have identified some of the active compo-nents of black raspberries. Ferulic acid and β-sitosterol, bothfound in large amounts in freeze-dried black raspberries, and
214 Nutrition and Cancer 2005
Figure 3. (Continued)
B

the RO-ET fraction selectively inhibit the growth ofpremalignant and malignant oral cells by targeting cell cycleregulatory proteins. The further identification and character-ization of these and other active components of black rasp-berries should help to better establish the nutritional compo-nents responsible for food-based chemopreventive activity.
Acknowledgments and Notes
Supported by grants from the National Institute of Dental andCraniofacial Research, P01–DE12704. We express our special gratitude toDale Stokes of Dale Stokes Berry Farm (Wilmington, OH) for cultivationand preparation of black raspberries and his continuing interest in our re-search. Address correspondence to S. M. D’Ambrosio, PhD, Division ofRadiobiology, Department of Radiology, Room 103 Wiseman Hall, 400
West 12th Avenue, Columbus, OH 43210. Phone: 614–292–9375. FAX:614–292–7237. E-mail: [email protected].
Submitted 14 July 2004; accepted in final form 5 January 2005.
References
1. La V, Altieri A, and Tavani A: Vegetables, fruit, antioxidants and can-cer: a review of Italian studies. Eur J Nutr 40, 261–267, 2001.
2. Vainio H and Bianchini F: Evaluation of cancer-preventive agents andstrategies: a new program at the International Agency for Research onCancer. Ann N Y Acad Sci 952, 177–180, 2001.
3. Byers TE: Nutrition and cancer: ten lessons from the 20th century. Nu-trition 16, 561–563, 2000.
4. Stoner GD, Weghorst CM, D’Ambrosio SM, Kresty LA, Harris GK, etal.: Prevention of cancer by berries and berry components. PharmacolTher, 2004. Forthcoming.
Vol. 51, No. 2 215
Figure 3. (Continued)
C

5. Casto BC, Kresty LA, Kraly CL, Pearl DK, Knobloch TJ, et al.:Chemoprevention of oral cancer by black raspberries. Anticancer Res22, 4005–4015, 2002.
6. Wada L and Ou B: Antioxidant activity and phenolic content of Ore-gon caneberries. J Agric Food Chem 50, 3495–3500, 2002.
7. Wang SY and Jiao H: Scavenging capacity of berry crops onsuperoxide radicals, hydrogen peroxide, hydroxyl radicals, and singletoxygen. J Agric Food Chem 48, 5677–5684, 2000.
8. Wang SY and Lin HS: Antioxidant activity in fruits and leaves ofblackberry, raspberry, and strawberry varies with cultivar and develop-mental stage. J Agric Food Chem 48, 140–146, 2000.
9. Park EJ and Pezzuto JM: Botanicals in cancer chemoprevention. Can-cer Metastasis Rev 21, 231–255, 2002.
10. Wang H, Cao G, and Prior RL: Total antioxidant capacity of fruits. JAgric Food Chem 44, 701–705, 1996.
11. Kähkönen MP, Hopia AI, Vuorela HJ, Pihlaja K, Kujala TS, et al.: An-tioxidant activity of plant extracts containing phenolic compounds. JAgric Food Chem 47, 3954–3962, 2002.
12. Katsube N, Iwashita K, Tsushida T, Yamaki K, and Kobori M: Induc-tion of apoptosis in cancer cells by Bilberry (Vaccinium myrtillus) andthe anthocyanins. J Agric Food Chem 51, 68–75, 2003.
13. Stoner GD, Kresty LA, Carlton PS, Siglin JC, and Morse MA:Isothiocyanates and freeze-dried strawberries as inhibitors of esopha-geal cancer. Toxicol Sci 52, 95–100, 1999.
14. Carlton PS, Kresty LA, Siglin JC, Morse MA, Lu J, et al.: Inhibition ofN-nitrosomethylbenzylamine-induced tumorigenesis in the rat esoph-agus by dietary freeze-dried strawberries. Carcinogenesis 22,441–446, 2001.
15. Kresty LA, Morse MA, Morgan C, Carlton PS, Lu J, et al.:Chemoprevention of esophageal tumorigenesis by dietary administra-tion of lyophilized black raspberries. Cancer Res 61, 6112–6119,2001.
16. Harris GK, Gupta A, Nines RG, Kresty LA, Habib SG, et al.: Effects oflyophilized black raspberries on azoxymethane-induced colon cancerand 8-hydroxy-2′-deoxyguanosine levels in the Fischer 344 rat. NutrCancer 40, 125–133, 2001.
17. Xue H, Aziz RM, Sun N, Cassady JM, Kamendulis LM, et al.: Inhibi-tion of cellular transformation by berry extracts. Carcinogenesis 22,351–356, 2001.
18. Huang C, Huang Y, Li J, Hu W, Aziz R, et al.: Inhibition ofbenzo(a)pyrene diol-epoxide-induced transactivation of activated pro-tein 1 and nuclear factor kappaB by black raspberry extracts. CancerRes 62, 6857–6863, 2002.
19. Walker DM, Boey G, and McDonald LA: The pathology of oral can-cer. Pathology 35, 376–383, 2003.
20. Neville BW and Day TA: Oral cancer and precancerous lesions. CACancer J Clin 52, 195–215, 2002.
21. D’Ambrosio SM, D’Ambrosio REG, Milo GE, Casto BC, Kelloff GJ,et al.: Differential growth responses of normal, premalignant, and ma-lignant human oral epithelial cells by chemopreventive agents.Anticancer Res 20, 2273–2280, 2000.
22. Lee H, Li D, Prior T, Casto BC, Weghorst CM, et al.: Ineffectiveness ofthe presence of h-ras/p53 combination mutations in squamous cell car-cinoma cells to induce the conversion of a nontumorigenic to atumorigenic tumor type. Cell Biol Toxicol 13, 419–434, 1997.
23. Milo GE, Shuler C, Kurian P, French BT, Mannix DG, et al.:Nontumorigenic squamous cell carcinoma line converted totumorigenicity with methyl methanesulfonate without activation ofHRAS or MYC. Proc Natl Acad Sci USA 87, 1268–1272, 1990.
24. Greenberg O, Terespolosky Y, Kahane I, and Bar R: Cyclodextrins ascarriers of cholesterol and fatty acids in cultivation of mycoplasmas.Appl Environ Microbiol 59, 547–551, 1993.
25. Ding H, Han C, Gibson-D’Ambrosio R, Steele VE, and D’AmbrosioSM: Piroxicam selectively inhibits the growth of premalignant andmalignant human oral cell lines by limiting their progression throughthe S phase and reducing the levels of cyclins and AP-1. Int J Cancer107, 830–836, 2003.
26. Gerber M, Boutron R, Hercberg S, Riboli E, Scalbert A, et al.: [Foodand cancer: state of the art about the protective effect of fruits and veg-etables]. Bull Cancer 89, 293–312, 2002.
27. Dong Y, Sui L, Tai Y, Sugimoto K, and Tokuda M: The overexpressionof cyclin-dependent kinase (CDK) 2 in laryngeal squamous cell carci-nomas. Anticancer Res 21, 103–108, 2001.
28. Quon H, Liu FF, and Cummings BJ: Potential molecular prognosticmarkers in head and neck squamous cell carcinomas. Head Neck 23,147–159, 2001.
29. Liu SC and Klein-Szanto AJ: Markers of proliferation in normal andleukoplakic oral epithelia. Oral Oncol 36, 145–151, 2000.
30. Hassan K, Ang K, El-Naggar A, Story M, Lee J, et al.: Cyclin B1overexpression and resistance to radiotherapy in head and necksquamous cell carcinoma. Cancer Res 62, 6414–6417, 2002.
31. Bukholm I, Bukholm G, and Nesland J: Coexpression of cyclin A andbeta-catenin and survival in breast cancer patients. Int J Cancer 94,148–149, 2001.
32. Opitz O, Harada H, Suliman Y, Rhoades B, Sharpless N, et al.: Amouse model of human oral-esophageal cancer. J Clin Invest 110,761–769, 2002.
33. Cazzalini O, Perucca P, Valsecchi F, Stivala LA, Bianchi L, et al.:Intracellular localization of the cyclin-dependent kinase inhibitorp21(CDKN1A)-GFP fusion protein during cell cycle arrest.Histochem Cell Biol 121, 377–381, 2004.
34. Dupont J, Karas M, and LeRoith D: The Cyclin-dependent kinase in-hibitor p21CIP/WAF is a positive regulator of insulin-like growthfactor induced cell proliferation in MCF-7 human breast cancer cells.J Biol Chem 278, 37256–37264, 2003.
35. Liu WK, Sze SC, Ho JC, Liu BP, and Yu MC: Wheat germ lectin in-duces G2/M arrest in mouse L929 fibroblasts. J Cell Biochem 91,1159–1173, 2004.
36. Kim OH, Lim JH, Woo KJ, Kim YH, Jin IN, et al.: Influence of p53and p21Waf1 expression on G2/M phase arrest of colorectal carcinomaHCT116 cells to proteasome inhibitors. Int J Oncol 24, 935–941,2004.
37. Kobayashi T, Nakata T, and Kuzumaki T: Effect of flavonoids on cellcycle progression in prostate cancer cells. Cancer Lett 176, 17–23,2002.
38. Hudson EA, Dinh PA, Kokubun T, Simmonds MSJ, and Gescher A:Characterization of potentially chemopreventive phenols in extractsof brown rice that inhibit the growth of human breast and coloncancer cells. Cancer Epidemiol Biomarkers Prev 9, 1163–1170,2000.
39. Bourne LC and Rice-Evans C: Bioavailability of ferulic acid. BiochemBiophys Res Commun 253, 222–227, 1998.
40. Zhang K and Zuo Y: GC-MS determination of flavonoids and phenolicand benzoic acids in human plasma after consumption of cranberryjuice. J Agricult Food Chem 52, 222–227, 2004.
41. Awad AB, Roy R, and Fink CS: Beta-sitosterol, a plant sterol, inducesapoptosis and activates key caspases in MDA-MB-231 human breastcancer cells. Oncol Rep 10, 497–500, 2003.
42. Park KY, Cho EJ, Rhee SH, Jung KO, Yi SJ, et al.: Kimchi and an ac-tive component, beta-sitosterol, reduce oncogenic H-Ras(v12)-in-duced DNA synthesis. J Med Food 6, 151–156, 2003.
43. Awad AB, Williams H, and Fink CS: Phytosterols reduce in vitro met-astatic ability of MDA-MB-231 human breast cancer cells. Nutr Can-cer 40, 157–164, 2001.
44. Awad AB, Downie AC, and Fink CS: Inhibition of growth and stimula-tion of apoptosis by beta-sitosterol treatment of MDA-MB-231 humanbreast cancer cells in culture. Int J Mol Med 5, 541–545, 2000.
45. Awad AB, Gan Y, and Fink CS: Effect of beta-sitosterol, a plant sterol,on growth, protein phosphatase 2A, and phospholipase D in LNCaPcells. Nutr Cancer 36, 74–78, 2000.
46. Awad AB, von Holtz RL, Cone JP, Fink CS, and Chen YC:Beta-sitosterol inhibits growth of HT-29 human colon cancer cells byactivating the sphingomyelin cycle. Anticancer Res 18, 471–473,1998.
216 Nutrition and Cancer 2005

47. von Holtz RL, Fink CS, and Awad AB: Beta-sitosterol activates thesphingomyelin cycle and induces apoptosis in LNCaP human prostatecancer cells. Nutr Cancer 32, 8–12, 1998.
48. Awad AB, Chen YC, Fink CS, and Hennessey T: Beta-sitosterol inhib-its HT-29 human colon cancer cell growth and alters membrane lipids.Anticancer Res 16, 2797–2804, 1996.
49. Awad AB, Williams H, and Fink CS: Effect of phytosterols on choles-terol metabolism and MAP kinase in MDA-MB-231 human breastcancer cells. J Nutr Biochem 14, 111–119, 2003.
50. Awad AB and Fink CS: Phytosterols as anticancer dietary compo-nents: evidence and mechanism of action. J Nutr 130, 2127–2130,2000.
51. Ling WH and Jones PJH: Minireview dietary phytosterols: a review ofmetabolism, benefits and side effects. Life Sci 57, 195–206, 1995.
Vol. 51, No. 2 217










![Morphological and histopathological analysis of cutaneous horn · tumor base or a low height-to-base ratio and the tendency to be malignant or premalignant in nature [5,6]. On the](https://img.pdfslide.us/doc/110x75/5e44a85990fdf211f81f84e2/morphological-and-histopathological-analysis-of-cutaneous-horn-tumor-base-or-a-low.jpg)



















