Embed Size (px)
Citation preview

Spring 2002290
Influenza - History
a. Oldest record of epidem
ic probably caused by influenza was recorded by H
ippocrates in 412 BC.
b. Looking back at recorded epidemics prior to 1800s:
1. Epidemics occurred relatively frequently but at irregular intervals
2. Sometim
es the disease appeared as if had disappeared for significant periods of time
3. Epidemics varied in severity but usually caused death in the very young and elderly.
4. Epidemics appeared to radiate from
specific locationsa. Ex: 1781 epidem
ic that spread across Russia from
Asia.
5. Influenza has killed untold millions throughout the centuries
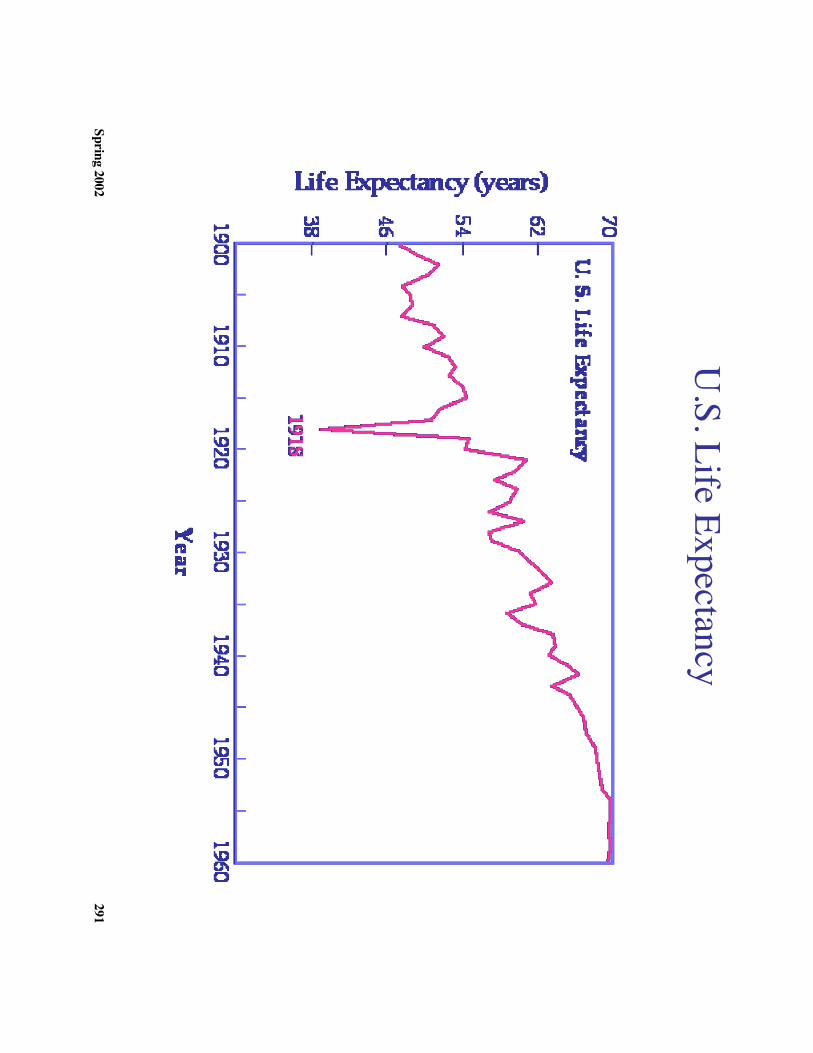
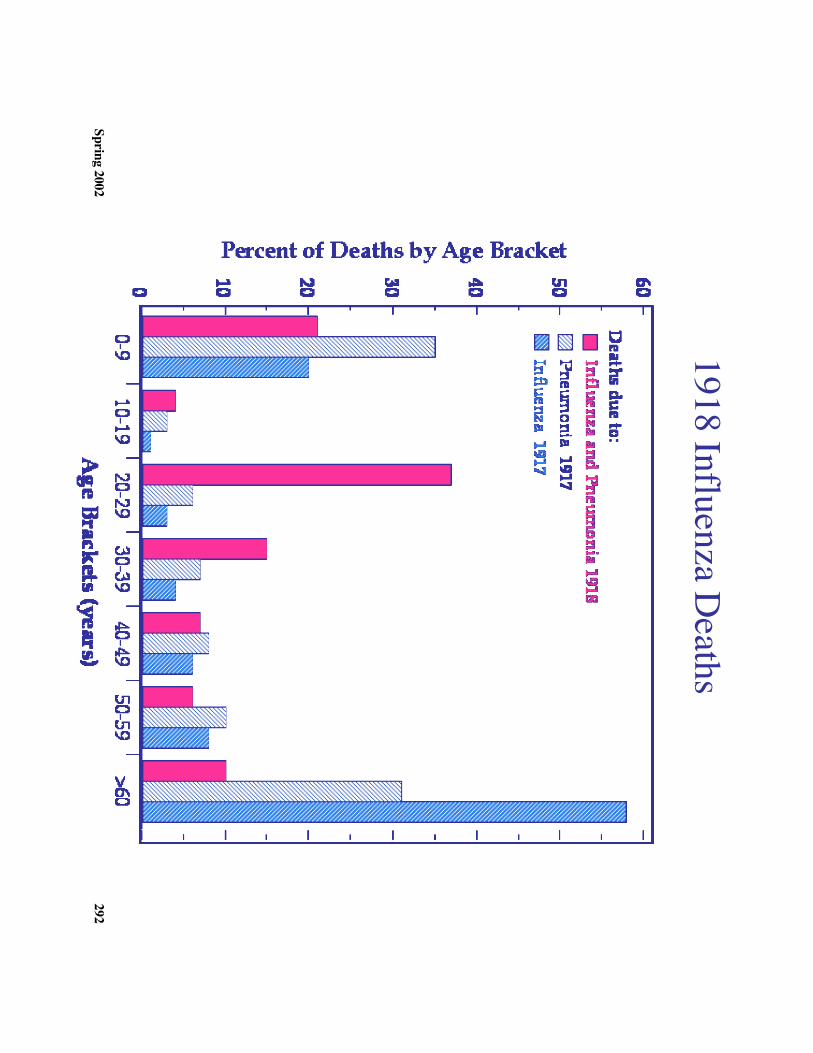
c. Influenza in the 20th century1. 1918-1919 epidem
ic was particularly severe
2. 40 million people died: Significantly m
ore than died in World W
ar I.3. 80%
of US W
.W.I deaths w
ere due to influenza4. A
significant factor in the Germ
an loss was influenza
5. First human influenza virus w
as isolated in 1933.6. R
ecent outbreak in Hong K
ong (1997) H5N
1
d. Different strains cause different epidem
ics, but human strains can recirculate
Spring 2002291
U.S. Life Expectancy
Spring 2002292
1918 Influenza Deaths

Spring 2002293
Influenza N
omenclature
a. Influenza divided into three types: A,B, C
1. Type A: a. Found in m
any animal species, in addition to hum
ansb. C
losely related to Type B but not Type Cc. M
ain Type found in human epidem
icsd. D
emonstrates the greatest antigenic variability (“antigenic drift”)
2. Types B and C:
a. Found almost exclusively in hum
ansb. Type C
can also infect swine
c. Clear m
orphological and antigenic differences between Types B
and Cb. D
esignation of Type A strains: type A
subdivided by:a. H
ost of originb. G
eographic locationc. Strain num
berd. Y
ear of isolatione. D
escription of hemagglutinin (H
A) and neuram
inidase (NA
) virion sufaceglycoproteins. Ex: A
/swine/Iow
a/15/30 (H1N
1)

Spring 2002294
Pathogenesisa. Transm
ission is by person-to-person spread via aerosol route1. H
ighly infectious & contagious
b. Virus replicates throughout the respiratory track
c. Disease tim
e course1. V
irus peak 24-48 Hr. post infection
2. Virus triggers both a host interferon and im
mune response
Antibodies m
ade HA
, NA
, NP, and M
proteins
d. Influenza A virus induces pathological changes throughout the entire respiratory
tract.1. M
ost significant pathology is in the lower respiratory tract.
2. Clinical sym
ptoms can range from
asymptom
atic infection to a primary viral
pneumonia (fatal outcom
e)3. Serious low
er respiratory tract complications include:
Primary viral pneum
onia which can lead to death in 1-4 days.
Com
bined viral-bacterial pneumonia: 3x m
ore comm
on than primary
pneumonia: Influenza infection follow
ed by bacterial pneumonia

Spring 2002295
Pathogenesis (cont’d)4. Extra pulm
onary manifestation of influenza:
a. Infection in the blood (rare)b. M
uscle and cardiac involvement
c. Reye’s Syndrom
e: neurological and metabolic disease of children and adolescents
d. Central nervous system
: encephalitise. Pregnant w
omen have an increased risk of developing fatal influenza disease
5. Influenza B and C virus infections
a. Influenza B: Less comm
on than influenza A (4x less serious)
Effects predominately adolescents and school-age children
b. Influenza C:
Seldom causes sever disease.
By early adulthood, most individuals (96%
) have antibodies to influenza C

Spring 2002296
Epidemiology
The virus undergoes significant antigenic variation which results in epidem
icsa. Tw
o types of antigenic variation:1. A
ntigenic drift: Involves minor changes in H
A and N
A polypetides
2. Antigenic shift: Involves m
ajor antigenic changes in HA
and NA
polypeptides.
b. Nature of A
ntigenic drift1. H
A is the m
ajor surface antigen of influenza2. M
Abs to H
A are used to determ
ine antigenic drift3. Type A
virus undergoes pronounce antigenic drift4. A
ntigenic drift in HA
occurs by point mutations
5. Antigenic drift can also occur in N
A at approxim
ately the same rate as H
A6. M
echanism of antigenic drift:
i. Clearly results from
the accumulation of point m
utations, but multiple point m
utations m
ust occur before the virus is able to escape neutralization by antibodies ii. M
utations are thought to occur during the spread of virus from person-to-person
c. Nature of antigenic shift1. Thus far, all m
ajor antigenic shifts occurred in China
i. Ex: 1957: H2N
2 subtype replaced the H1N
1 1968 H
3N2 appeared to be replaced by H
1N1
ii. Ex: Since the 1890s: three different HA
subtypes have been introduced
There has been a cyclical appearance of three human subtypes:
H2: 1890, 1957
H3: 1900, 1968
H1: 1918, 1977

Spring 2002297
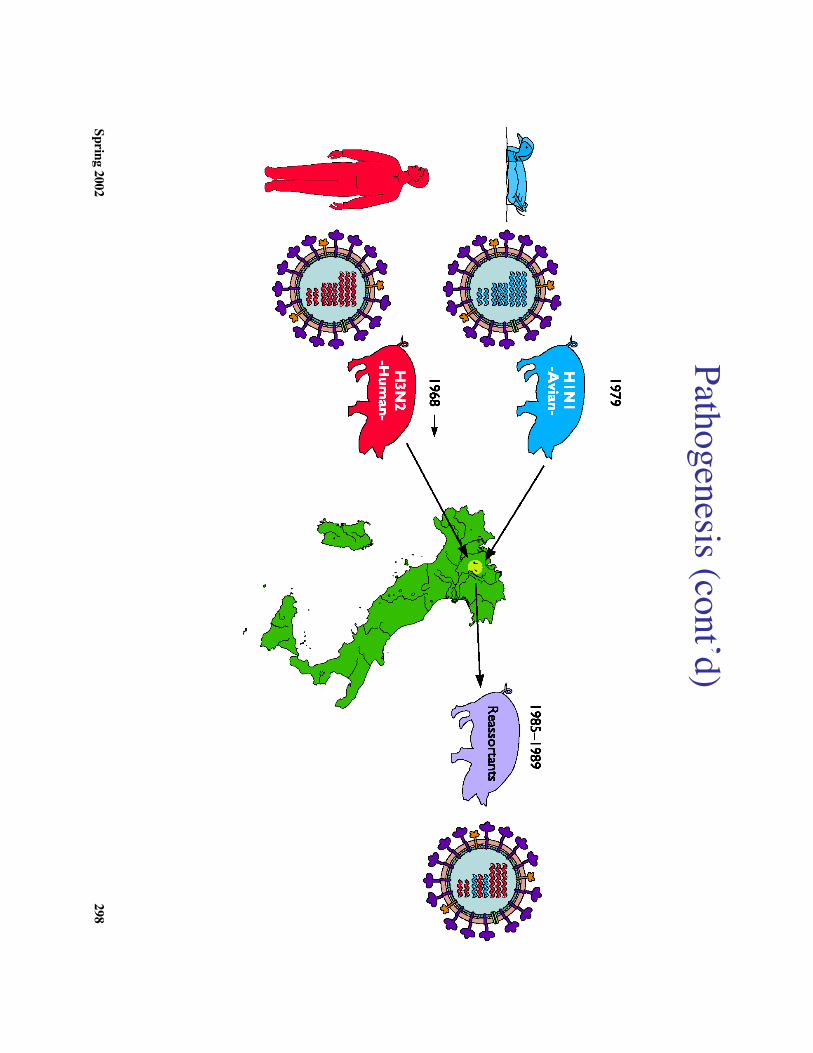
Transmission M
odels2. First M
odel: Antigenic shift m
ay result from genetic reassortm
ent between hum
anand anim
al influenza A viruses in vivo (see diagram
). i. Ex: 1968 a shift occurred from
H2N
2 to H3N
2 H
uman H
3 was 98%
identical on the nucleic acid level to avian influenza A.
ii. Little antigenic drift observed in influenza B and C because they have
principally human hosts
3. Second model: The ‘new
” form of virus m
ay have been hidden.Ex: R
ussian influenza H1N
1 appeared 1977 in northern China that w
as identical tosam
e virus that caused epidemics in 1950.
4. Third model: N
ew virus could appear in the hum
an population because an animal or
bird form of the virus becam
e infectious for humans.
Ex: Virus from
pigs to man on the sam
e farm in W
isconsin
d. The virus is maintained in the hum
an population by person-to-person spread duringacute infection1. School-age children are the m
ajor vectors in the transmission of the virus in
comm
unities2. Increase in m
ortality seen with introduction of new
strain
Spring 2002298
Pathogenesis (cont’d)
Spring 2002299
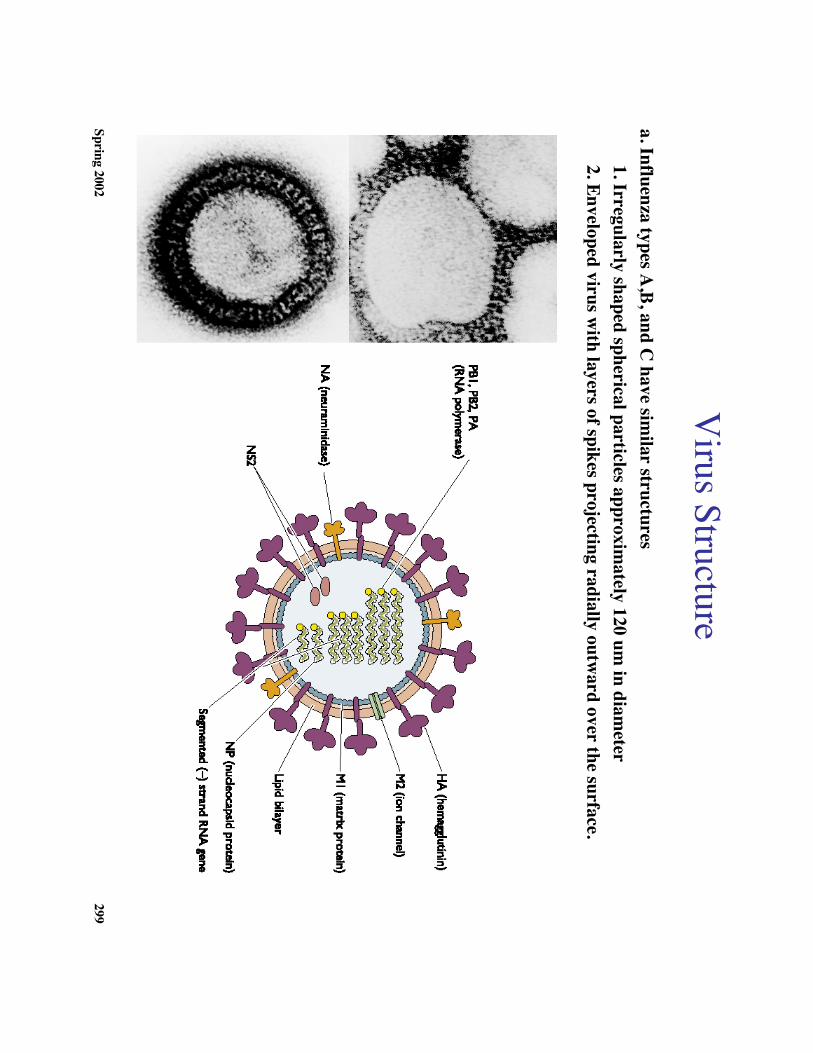
Virus Structure
a. Influenza types A,B, and C
have similar structures
1. Irregularly shaped spherical particles approximately 120 um
in diameter
2. Enveloped virus with layers of spikes projecting radially outw
ard over the surface.
Spring 2002300
Viral Proteins
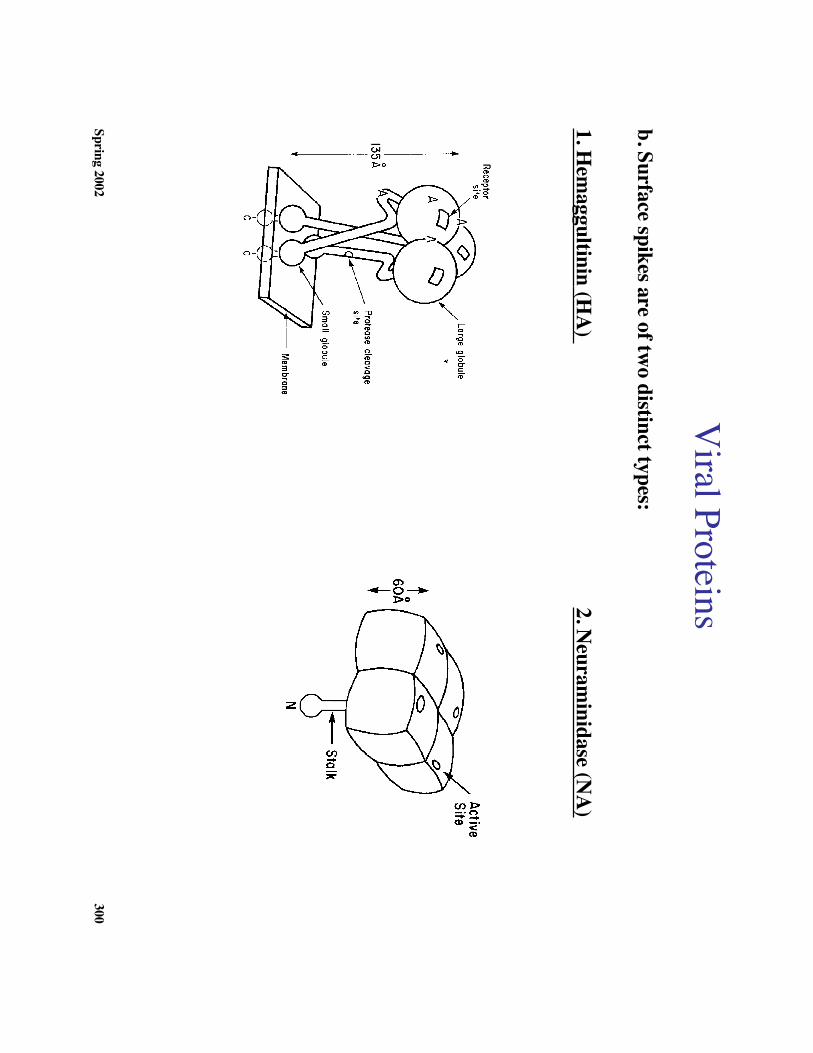
b. Surface spikes are of two distinct types:
1. Hem
aggultinin (HA
) 2. N
euraminidase (N
A)

Spring 2002301
Viral Proteins
•c. W
ithin the lipid envelope lies the mem
braneprotein M
1 (see diagram)
•d. Spanning the lipid envelope is the sm
all mem
braneprotein, M
2.
•e. W
ithin the matrix shell are eight single stranded
RN
A m
olecules of the negative sense each associatedw
ith the nucleocapsid protein (NP).
•f. Three large proteins (PB1, PB2, and PA
)responsible for R
NA
replication and transcriptionare clustered at the 3' end of each R
NA
as if poisedto initiate R
NA
synthesis.
•g. O
ne additional virus coded nonstructual proteins(N
S2) isfound in the virion. It is responsible fortransport of nucleocapsids out of the nucleus.

Spring 2002302
Genom
e Organization
a. Eight ss (-) RN
A segm
ents, separately encapsidated by NP, w
hich encode for 10 proteinproducts (except Type C
which only encodes for 8 or 9)
b. Nucleocapsid proteins:1. 3 polym
erase proteins (PB1, PB2, PA)
2. Nucleoprotein (N
P)3. A
ll nucleocapsid proteins function in the cell nucleus and contain specific nucleartransport sequences for transport from
their cytoplasmic site of synthesis.
c. Envelope proteins1. H
emm
agglutinin (HA
):i. M
ediates the initial attachment of virus particle its cellular receptor (sialic acid)
ii. HA
is cleaved post-translationally which enables the virus envelope to fuse
with an intracellular m
embrane so that the virion nucleocapsid can be
delivered into the cell cytoplasm.
2. Neuram
inidase (NA
):i. A
ppears to have an exclusively enzymatic function: rem
oval of sialic acidresidues from
any gylcoconjugate.ii. A
ppears to play a role in virion assembly and release
Spring 2002303
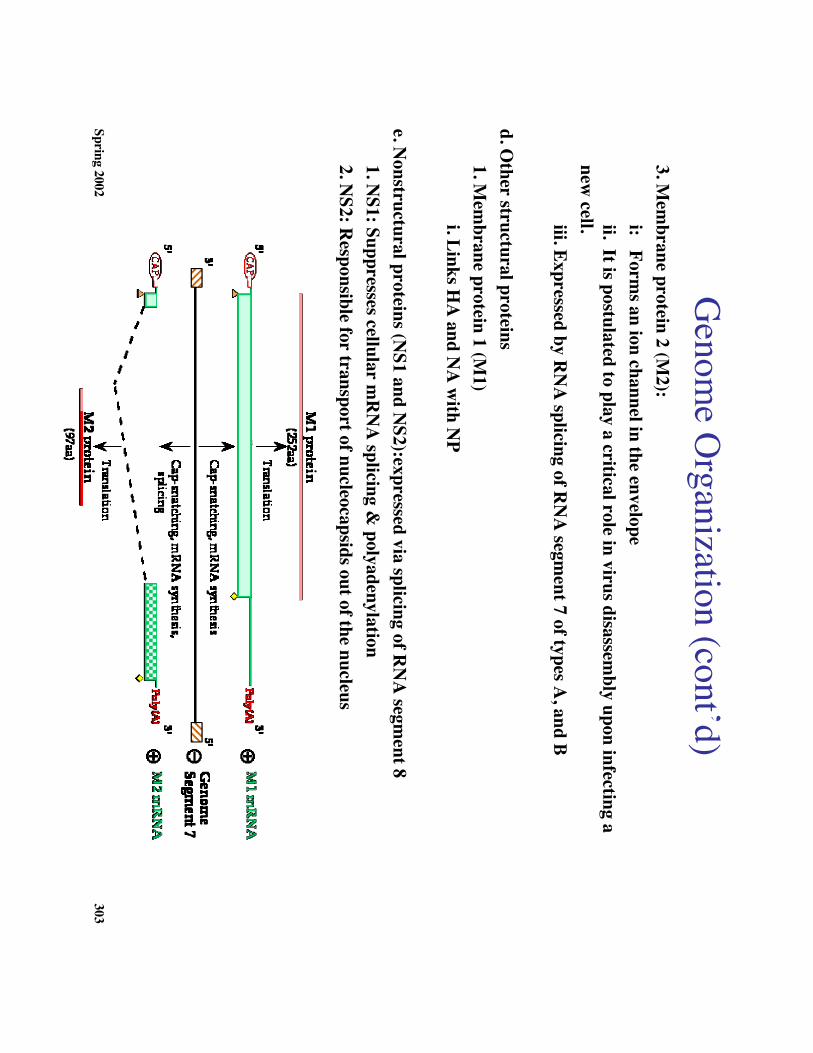
Genom
e Organization (cont’d)
3. Mem
brane protein 2 (M2):
i: Forms an ion channel in the envelope
ii. It is postulated to play a critical role in virus disassembly upon infecting a
new cell.iii. Expressed by R
NA
splicing of RN
A segm
ent 7 of types A, and B
d. Other structural proteins1. M
embrane protein 1 (M
1)i. Links H
A and N
A w
ith NP
e. Nonstructural proteins (N
S1 and NS2):expressed via splicing of R
NA
segment 8
1. NS1: Suppresses cellular m
RN
A splicing &
polyadenylation2. N
S2: Responsible for transport of nucleocapsids out of the nucleus
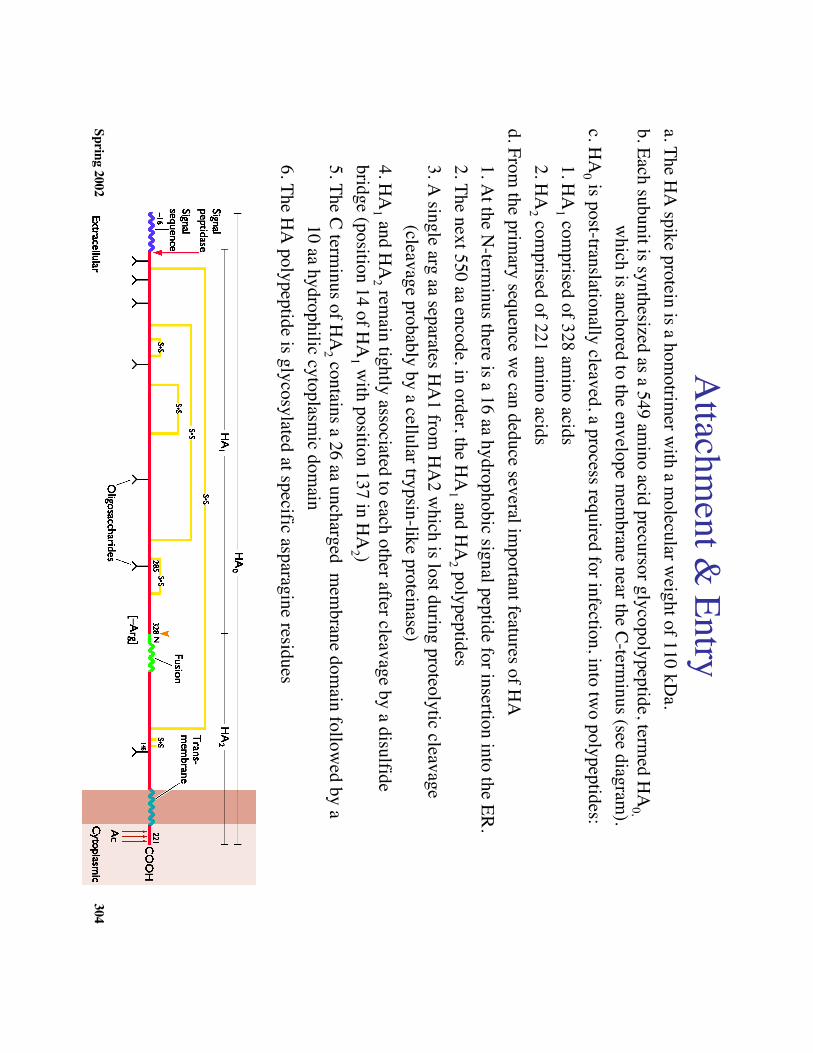
Spring 2002304
Attachm
ent & Entry
a. The HA
spike protein is a homotrim
er with a m
olecular weight of 110 kD
a.b. Each subunit is synthesized as a 549 am
ino acid precursor glycopolypeptide, termed H
A0,
which is anchored to the envelope m
embrane near the C-term
inus (see diagram).
c. HA
0 is post-translationally cleaved, a process required for infection, into two polypeptides:
1. HA
1 comprised of 328 am
ino acids2. H
A2 com
prised of 221 amino acids
d. From the prim
ary sequence we can deduce several im
portant features of HA
1. At the N
-terminus there is a 16 aa hydrophobic signal peptide for insertion into the ER.
2. The next 550 aa encode, in order, the HA
1 and HA
2 polypeptides3. A
single arg aa separates HA
1 from H
A2 w
hich is lost during proteolytic cleavage (cleavage probably by a cellular trypsin-like proteinase)
4. HA
1 and HA
2 remain tightly associated to each other after cleavage by a disulfide
bridge (position 14 of HA
1 with position 137 in H
A2 )
5. The C terminus of H
A2 contains a 26 aa uncharged m
embrane dom
ain followed by a
10 aa hydrophilic cytoplasmic dom
ain6. The H
A polypeptide is glycosylated at specific asparagine residues
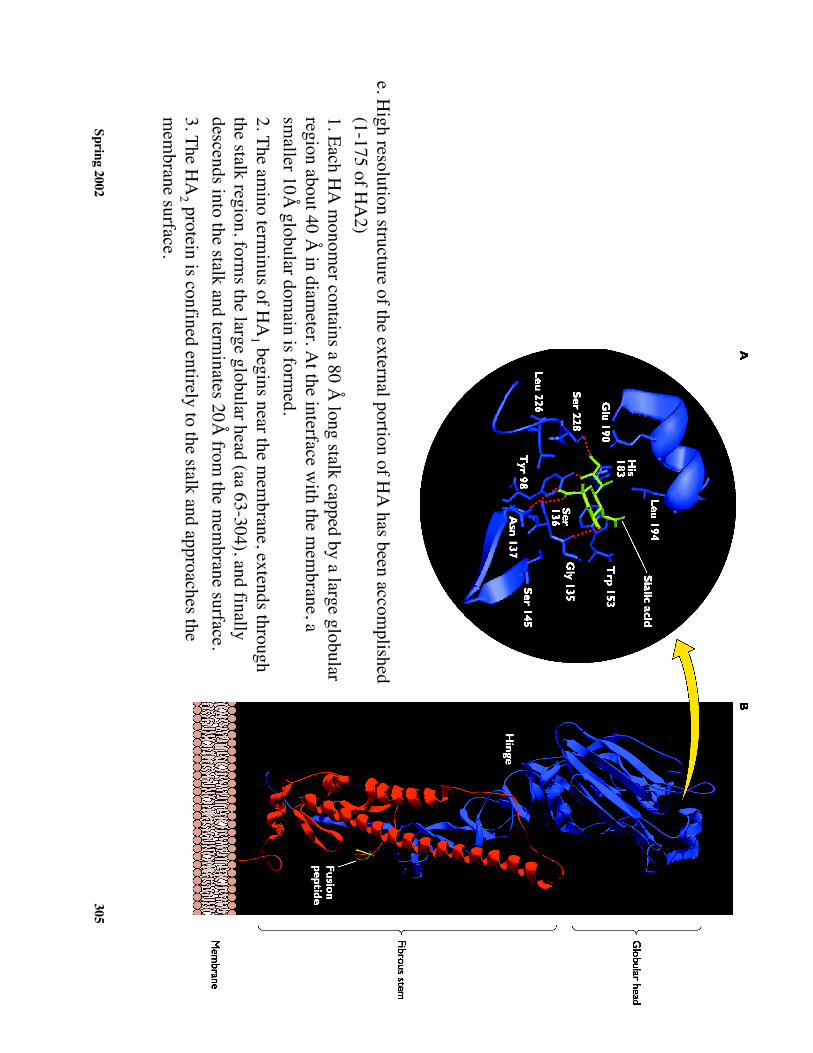
Spring 2002305
e. High resolution structure of the external portion of H
A has been accom
plished(1-175 of H
A2)
1. Each HA
monom
er contains a 80 Å long stalk capped by a large globular
region about 40 Å in diam
eter. At the interface w
ith the mem
brane, asm
aller 10Å globular dom
ain is formed.
2. The amino term
inus of HA
1 begins near the mem
brane, extends throughthe stalk region, form
s the large globular head (aa 63-304), and finallydescends into the stalk and term
inates 20Å from
the mem
brane surface.3. The H
A2 protein is confined entirely to the stalk and approaches the
mem
brane surface.

Spring 2002306
4. The HA
trimer is stabilized by the form
ation of a hydrophobic core formed betw
een the three stalkregions.
5. Attachm
ent sites for the cellular receptors are located near the top of each large globular region6. The large globule also contains all the antigenic sites required for virus neutralization.
a. Sites occur in surface loops which do not contribute significantly to the H
A quater-
nary structureb. These sites are free to m
utate extensively which contributes greatly to the observed
antigenic variability
f. Receptor binding
1. The exact glycoprotein(s) that serve as host cell surface receptors has not been identified, but it is know
n to contain sialic acid2. A
shallow binding pocket has been defined
Attachm
ent & Entry

Spring 2002307
HA
-mediated m
embrane fusion
a.After binding H
A to the cell surface receptor(s) the virus is internalized by
endocytosis.
b. The low pH
of endosomes ( pH
5.0-6.0) activates the mem
brane fusion potential ofH
A.
c. The subsequent fusion of viral and endosomal m
embranes allow
s the release of theviral genom
e into the cellular cytoplasm
d. How
is mem
brane fusion accomplished?
The low pH
results in a irreversible conformational change in H
A w
hich is required form
embrane fusion activity
1. At low
pH there is an extrusion of the highly conserved hydrophobic am
inoterm
inus of HA
2 from its position in the native protein
2. This region can promote m
embrane fusion (term
ed the ‘’fusion peptide’).3. The m
echanism of how
the ‘fusion peptide’ promotes m
embrane fusion is
unknown
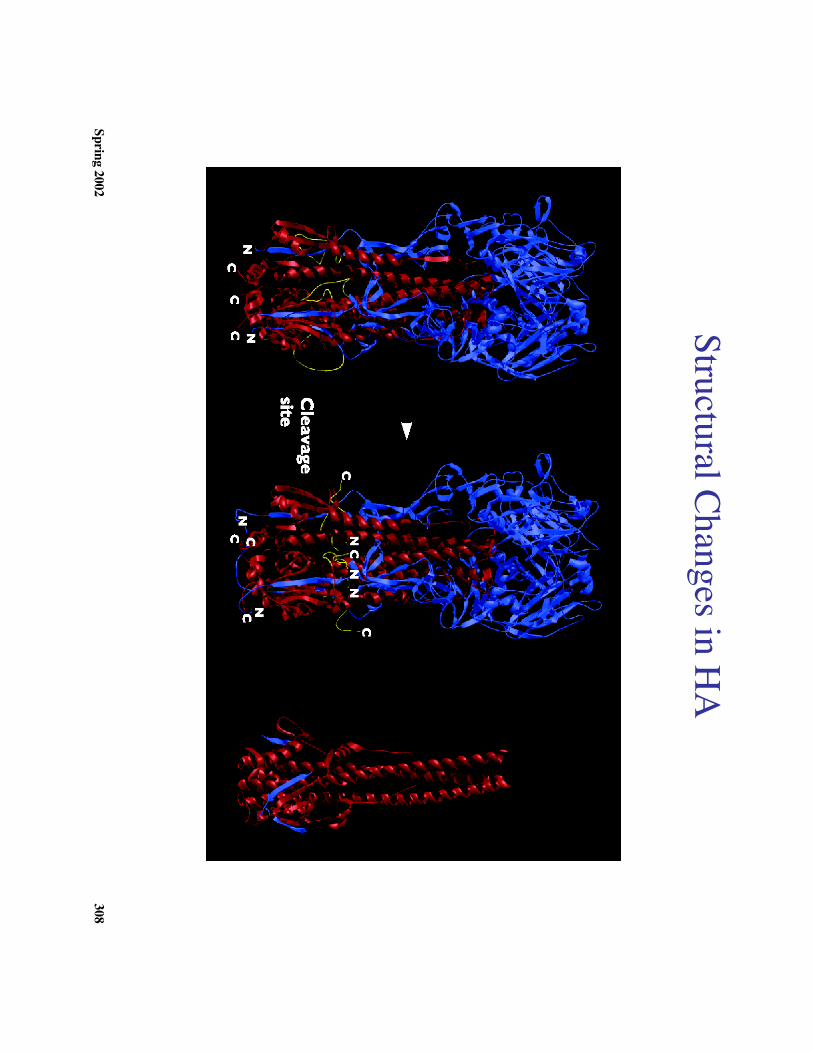
Spring 2002308
Structural Changes in HA
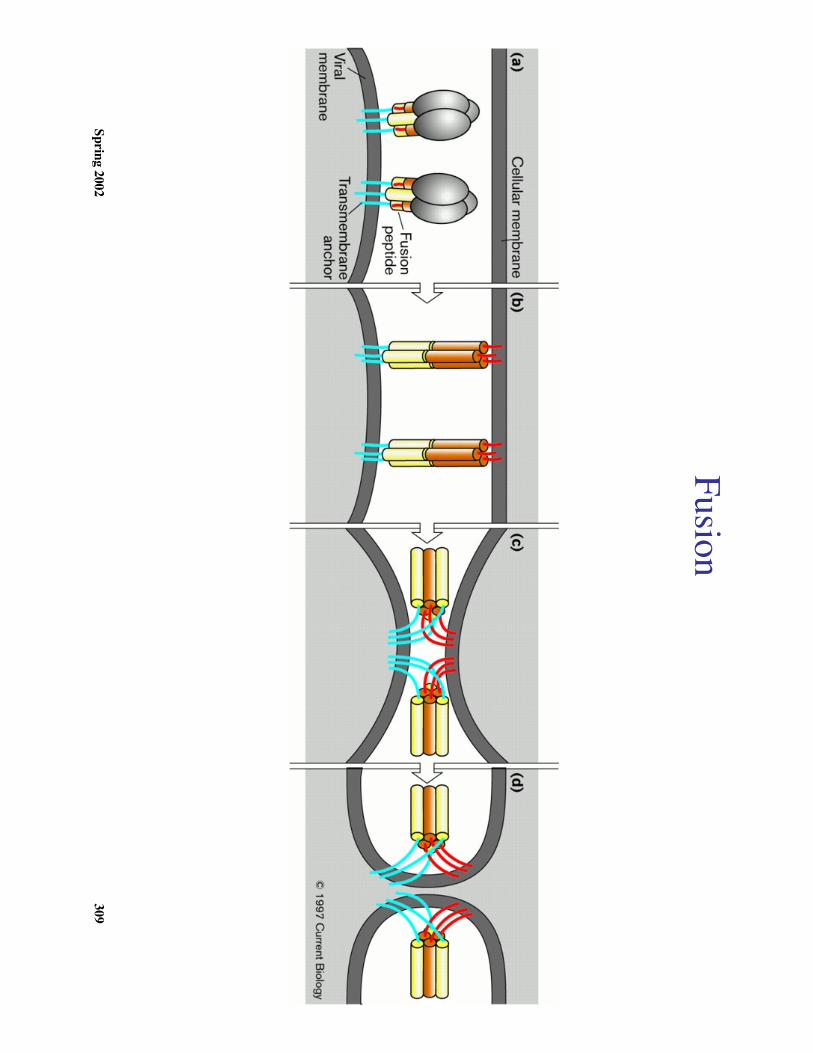
Spring 2002309
Fusion

Spring 2002310
M2 Protein
III. Function of the M2 protein
a. M2 is a 97 residue, non-glycosylated transm
embrane protein
b. A tetramer of M
2 appears to functions as a transmem
brane ion channelc. The ion channel appears to play tw
o important roles:
1. During virus entry it may facilitate release of free nucleocapsid into
the cella. How
?:1. It is known that the drug am
antadine acts early ininfection and effects M
2 function.2. The drug does not block virus binding, internalizationby endocytosis, or m
embrane fusion-nucleocapsid
release into the cytoplasm3. Apparently the drug inhibits nucleocapsid uncoatingand/or nuclear transport4. H
ow is a m
embrane protein (M
2) important in N
Pdisassem
bly?a. There is a 25 m
in. lag between virus
appearance in endosomes and m
embrane fusion.
b. During this lag, the M2 protein m
ay allow an
influx of H+ w
hich may induce conform
ationalchanges in the nucleocapsid w
hich allow release
of NP, and M
12. Late in infection it m
ay regulate the pH of G
olgi vesicles involved intransport of viral glycoproteins to the cell surface.
a. How
?: Clue came after analyzing the effect of the drug
amantadine on influenza.
1. All drug resistant mutants m
apped to the M2
polypeptide2. In the presence of the drug, W
T influenza displayedH
A on the surface of the cell in its low-pH
conformation
3. This suggests that in the absence of the drug the M2
polypeptide may alter the pH
of the acidic Golgi vesicles
(to a more basic pH
)4. In the presence of the drug, M
2 function is blocked,and the G
olgi vesicles remain acidic.
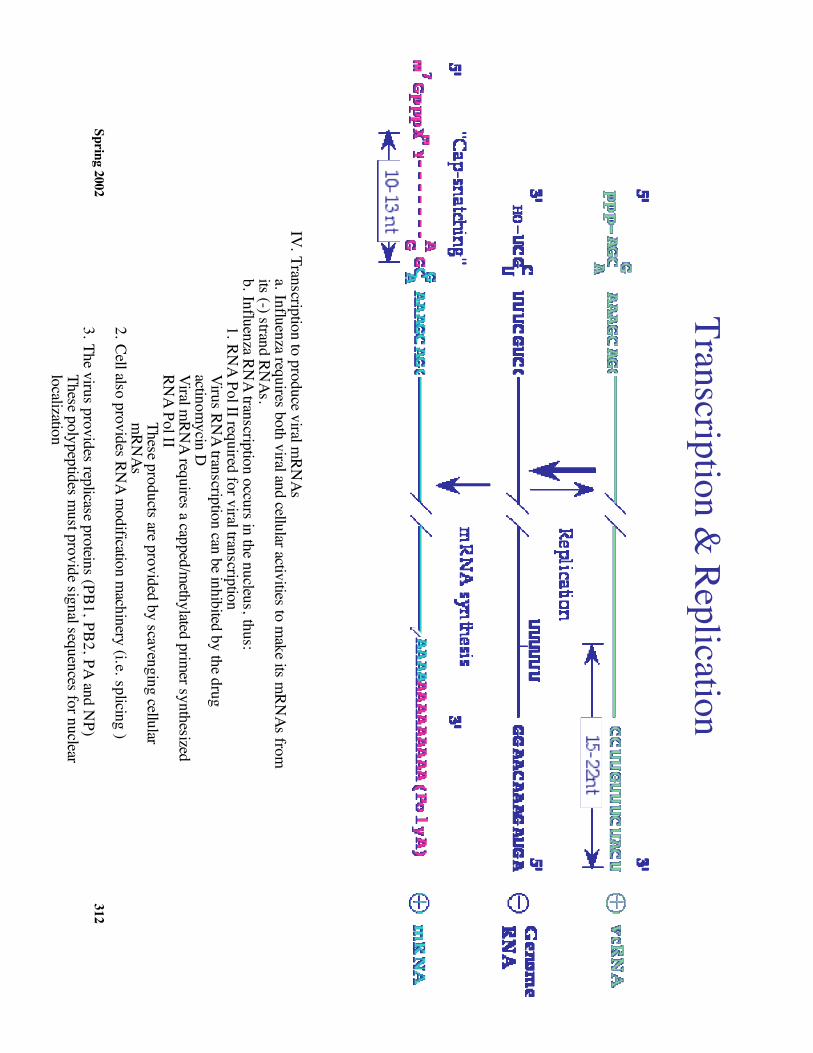
IV. Transcription to produce viral mRN
Asa. Influenza requires both viral and cellular activities to m
ake its mRN
As fromits (-) strand RN
As.b. Influenza RN
A transcription occurs in the nucleus, thus:1. RN
A Pol II required for viral transcriptionVirus RN
A transcription can be inhibited by the drugactinom
ycin DViral m
RNA requires a capped/m
ethylated primer synthesized
RNA Pol II
These products are provided by scavenging cellularm
RNAs
2. Cell also provides RNA m
odification machinery (i.e. splicing )
3. The virus provides replicase proteins (PB1, PB2, PA and NP)
These polypeptides must provide signal sequences for nuclear
localization4. M
odel for initiation of viral mRN
A synthesis (see diagram)

Spring 2002311
M2 (cont’d)
Spring 2002312
III. Function of the M2 protein
a. M2 is a 97 residue, non-glycosylated transm
embrane protein
b. A tetramer of M
2 appears to functions as a transmem
brane ion channelc. The ion channel appears to play tw
o important roles:
1. During virus entry it may facilitate release of free nucleocapsid into
the cella. How
?:1. It is known that the drug am
antadine acts early ininfection and effects M
2 function.2. The drug does not block virus binding, internalizationby endocytosis, or m
embrane fusion-nucleocapsid
release into the cytoplasm3. Apparently the drug inhibits nucleocapsid uncoatingand/or nuclear transport4. H
ow is a m
embrane protein (M
2) important in N
Pdisassem
bly?a. There is a 25 m
in. lag between virus
appearance in endosomes and m
embrane fusion.
b. During this lag, the M2 protein m
ay allow an
influx of H+ w
hich may induce conform
ationalchanges in the nucleocapsid w
hich allow release
of NP, and M
12. Late in infection it m
ay regulate the pH of G
olgi vesicles involved intransport of viral glycoproteins to the cell surface.
a. How
?: Clue came after analyzing the effect of the drug
amantadine on influenza.
1. All drug resistant mutants m
apped to the M2
polypeptide2. In the presence of the drug, W
T influenza displayedH
A on the surface of the cell in its low-pH
conformation
3. This suggests that in the absence of the drug the M2
polypeptide may alter the pH
of the acidic Golgi vesicles
(to a more basic pH
)4. In the presence of the drug, M
2 function is blocked,and the G
olgi vesicles remain acidic.
IV. Transcription to produce viral mRN
Asa. Influenza requires both viral and cellular activities to m
ake its mRN
As fromits (-) strand RN
As.b. Influenza RN
A transcription occurs in the nucleus, thus:1. RN
A Pol II required for viral transcriptionVirus RN
A transcription can be inhibited by the drugactinom
ycin DViral m
RNA requires a capped/m
ethylated primer synthesized
RNA Pol II
These products are provided by scavenging cellularm
RNAs
2. Cell also provides RNA m
odification machinery (i.e. splicing )
3. The virus provides replicase proteins (PB1, PB2, PA and NP)
These polypeptides must provide signal sequences for nuclear
localization4. M
odel for initiation of viral mRN
A synthesis (see diagram)
Transcription & Replication

Spring 2002313
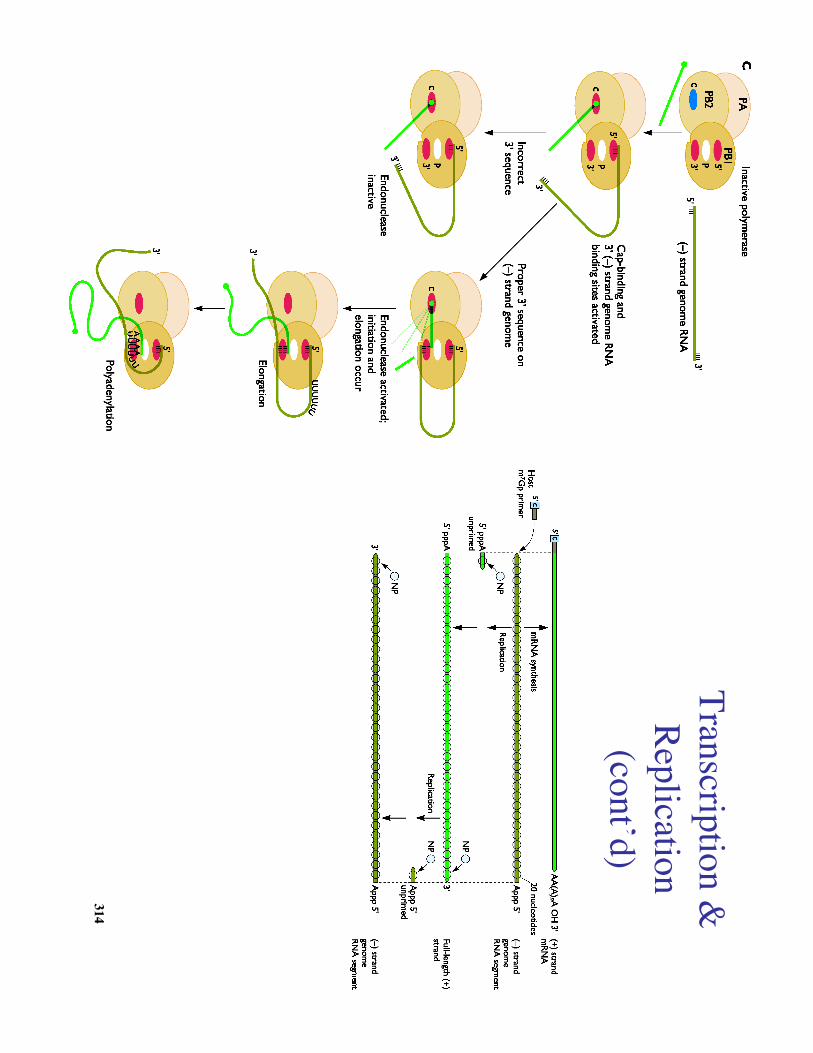
a. A viral encoded endonuclease cleaves the donor mRN
A 10-13nucleotides from
its cap, leaving a 3'OH1. The endonuclease requires a cap and usually cleaves3' to a A w
hich can base pair with the 3' U
found at theend of each viral RN
Ab. The resulting oligonucleotide acts directly as a prim
er, thefirst base added is a G
(penultimate in each viral RN
A is a C)c. Elongation of the transcript is then m
ediated by the viraltranscriptase (PB1, PB2, PA)d Transcription term
inates 15-22 nt from the ends of their
template w
here oligo U"s are used as a tem
plate for synthesis ofa 3' term
inal poly A tail
V. V
iral RN
A genom
e replicationa. N
uclear extracts from infected cells can synthesize full-length (+)
viral RNA
in the absence of capped primers
i. Indicates that 5' cap structure regulates termination in viral m
RNA
s ii. N
P is required along with PB1, PB2, and PA
b. The full-length (+) serve as a template for production of full-length
progeny (-) RNA
c. All viral RN
As have identical 3' and 5' term
ini, so each RNA
segment is capable of being efficiently replicated.
d. Both viral mRN
As and (-) full-length progeny are quickly exported
to the cytoplasm, w
hereas (+) full-length progeny remain localized to
the nucleus.
Spring 2002314
Transcription &Replication
(cont’d)

Spring 2002315
Assem
bly
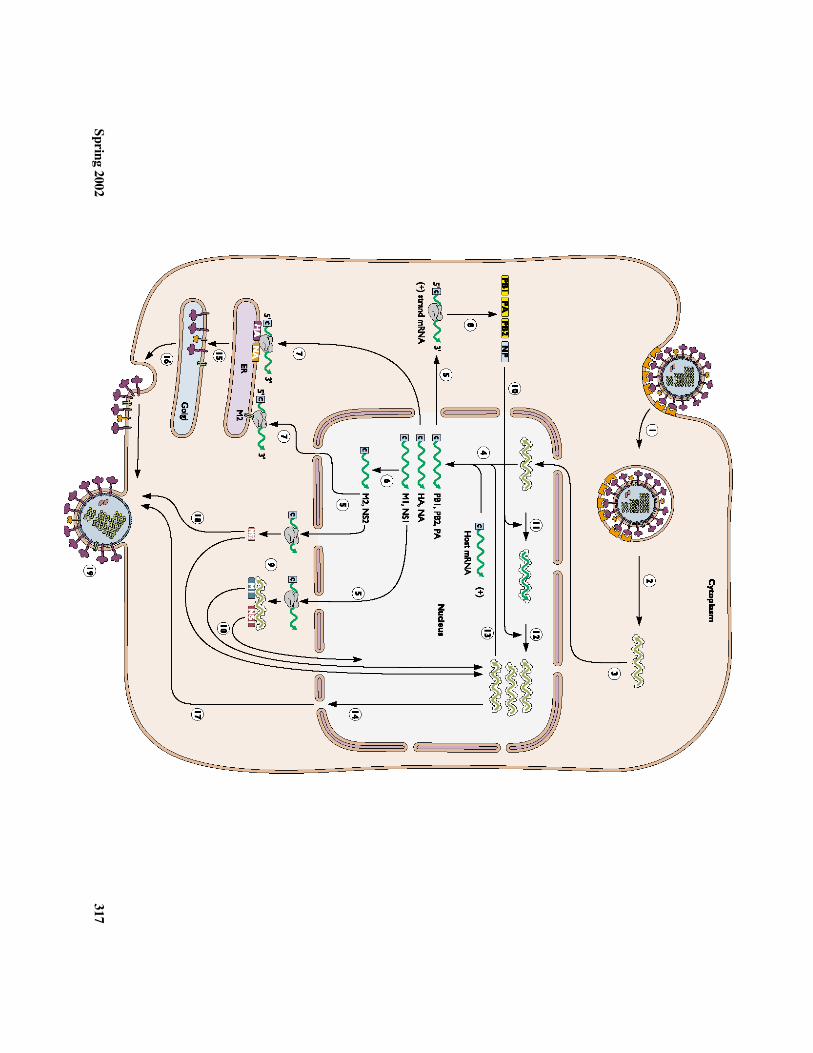
VI. Virion assembly and release
a. Complete virion assem
bly requires two assem
bly modules assem
bled indifferent subcellular locations:
1. The nucleocapsids are assembled in the nucleus
a. The RNA assem
bly sequences for initiating formation of
nucelocapsids are located in the termini of each full-length viral
RNA (but not in viral m
RNA w
hich lack the 5'-termini)
2. The envelope is assembled at the cell-surface m
embrane
b. The M1 protein m
ay serve to bind the nucleocapsid to the C-terminus
cytoplasmic dom
ains of HA and N
Ac. Rem
ember: M
2 may play a critical role in regulating G
olgi vesicle pH.
d. There must be som
e mechanism
(yet to be elucidated) to assure that eachvirion receives a full com
plement of nucleocapsids
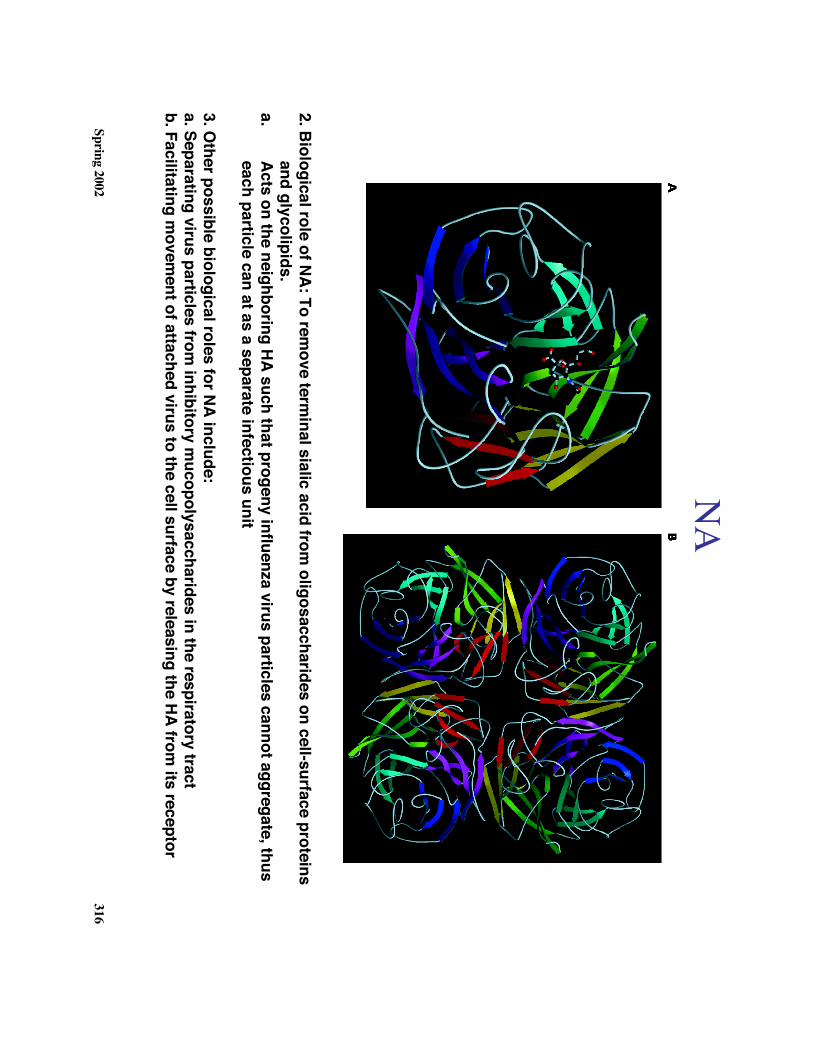
e. The role of NA
1. Structure (see diagram):
a. 469 aa glycoprotein which form
s a tetrameric spike on the
virus envelope surfaceb. The catalytic site for neuram
inic acid esterase activity islocated on top of each head ( 4 sites per spike)
Spring 2002316
NA
2. Biological role of NA: To remove term
inal sialic acid from oligosaccharides on cell-surface proteins
and glycolipids.a.
Acts on the neighboring HA such that progeny influenza virus particles cannot aggregate, thuseach particle can at as a separate infectious unit
3. Other possible biological roles for NA include:
a. Separating virus particles from inhibitory m
ucopolysaccharides in the respiratory tractb. Facilitating m
ovement of attached virus to the cell surface by releasing the HA from
its receptor
Spring 2002317

Spring 2002318
Prevention & Control
a. Antivirals:1. D
rugs (amatadine, ribavirin);
new recent round of drugs target N
A (see box 19.6 p 700)
2. Interferon
b. Inactivated vaccines (inactivated with form
alin)1. 60-80 %
effective in most individuals
2. Swine Flu V
accine (1976)