Embed Size (px)
Citation preview

INFLUENCE OF PLANT SPECIES AND SOIL CONDITIONS ON
PLANT-SOIL FEEDBACK IN THE JARRAH FOREST
Martha Orozco Aceves
Bacteriological Parasitological Chemist, National Polytechnical Institute,
Mexico
MSc. Alternative Agriculture with mention in Ecological Agriculture,
National University of Costa Rica, Costa Rica
Thesis presented for the degree of Doctor of Philosophy
The University of Western Australia
School of Earth and Environment
Faculty of Natural and Agricultural Sciences
2015


i
ABSTRACT
Plant-soil feedback (PSF) describes how changes to soil properties are caused by plants
(via conditioning), which in turn influence the plant performance (via soil feedback).
Plant-soil feedback is driven by soil conditions and plant species involved in the
interaction, however, the influence or ‘weight’ of each one of these two components on
PSF outcomes has been scarcely examined. Studying the influence of plant species and
soil conditions on PSF is especially important in highly endemic ecosystems such as the
jarrah forest. This ecosystem displays particular features that define its structure and
function, and general ecological theories often fall short to explain those particularities.
In addition, the jarrah forest has been subjected to a strong anthropogenic pressure by
mining activity. In this case, knowing the influence of plant species and soil conditions
on PSF may contribute to the design of effective restoration strategies.
My thesis focuses on the influence of plant species and soil conditions on PSF that
operates in the jarrah forest of Western Australia. Specifically, PSF was investigated in
the contexts of unmined soils and soils restored after bauxite mining. The research is
presented in three themes: 1) a detailed study of the influence of plant species and soil
contexts on individual PSF associated with representative plant species of the jarrah
forest, described in chapter 3; 2) a study of the influence of changing soil conditions
present in a chronosequence of restoration ages on plant performance and associated
individual PSF, described in chapter 4; and 3) the influence of long-term conditioned
soil by non-native plantation species on PSF associated to native jarrah plants, described
in chapter 5.
To accomplish the aims of the research, a series of glasshouse experiments were
designed using unmined and restored soils that were conditioned in the field and the
glasshouse. To examine individual effects of PSF, the standard method of comparing

ii
plant biomass produced in soil conditioned by the same plant species (‘home soil’) with
the plant biomass produced in soil conditioned by a different plant species (‘away soil’)
was consistently used in each of the experiments. Plants used for the experiments were
representative of the jarrah forest, these being Eucalyptus marginata, Bossiaea ornata
(both long-lived species), and Acacia pulchella (a short-lived species).
Results from the first glasshouse experiment designed to study the influence of plant
species and soil contexts on individual PSF are described in Chapter 3. The main
findings from this experiment were that individual PSF associated with B. ornata and A.
pulchella was influenced by soil context (unmined, restored), and species-specific
conditioning of soil caused by plant species, influenced the PSF only in restored soil.
In unmined soil, the three species E. marginata, B. ornata and A. pulchella conditioned
the properties via changes to the chemistry (as compared with unplanted soils), but had
little effect on the biological properties measured. Eucalyptus marginata and B. ornata
conditioned the chemical properties of soil in a similar manner, and A. pulchella
conditioned soils differently. In restored soil, altered conditioning of the chemical
properties was observed for E. marginata and B. ornata and no conditioning was
observed for A. pulchella. Again, the biological properties of restored soil were little
affected by conditioning. Overall, these findings indicate that individual PSF and
conditioning capacity associated with the studied plant species is driven by soil context,
which can be critical for the restoration of native forest post-bauxite mining given the
modified properties present in restored soil.
In chapter 4, I described the results from a second glasshouse experiment designed to
study the influence of soil conditions on PSF. For the study, field-conditioned soils from
a chronosequence of restoration ages were used. These soils displayed different
properties that approached those of unmined soil as restoration age increased. The soils

iii
exerted differential soil feedback on the biomass produced by B. ornata. As a result,
different amounts of biomass were produced by the plant species. Overall, the soil
feedback of soils from the chronosequence was driven by soil age (since restoration),
not by plant species from which soils were collected in the field. From individual
properties of soil, the number of bacterial-feeding nematodes and the rate of CO2
produced by microbial respiration of phenolic compounds from CLPP were identified as
key components of the soil feedback across all soils from the chronosequence. These
properties were correlated with B. ornata biomass production. The results reinforced the
hypothesis of a PSF influenced by soil conditions more than by plant species is
operating in restored jarrah forest.
Finally described in chapter 5, a third glasshouse experiment was conducted to
investigate changes in properties of jarrah forest soils after long-term conditioning by
the non-native plantation species pine (Pinus radiata) and blue gum (Eucalyptus
saligna), and native jarrah (E. marginata). Then, the influence of conditioned soils on
growth of jarrah seedlings was tested. Pines and blue gums similarly conditioned the
physico-chemical properties of soils, which differed from soil conditioning caused by
jarrah. The two eucalypt species similarly conditioned the biological properties of soil,
and these differed from conditioning caused by pines. Species-specific conditioning of
soil did not translate into differences in the amounts of biomass produced by jarrah
seedlings, and a neutral PSF was observed. The results suggest that jarrah displays a
plastic PSF that is not strongly influenced by soil context, instead PSF associated with
this plant species is driven by the plant itself. This behavior might contribute to the
persistence and dominance of E. marginata in the jarrah forest.
The studies included in this thesis have contributed to a better understanding of PSF
operating in the endemic jarrah forest ecosystem. Specifically, this thesis contributed to

iv
understand the influence of plant species and soil conditions on PSF. Moreover, the
influence of plant species and soil conditions was examined in the contexts of unmined
soil and restored soil post-bauxite mining, as mining is a common disturbance of the
jarrah forest. Restored soil post-bauxite mining displays modified physico-chemical and
biological properties that are likely to alter individual PSF, with implications for the
restoration of the ecosystem.

v
TABLE OF CONTENTS
Abstract i
Table of contents v
Thesis declaration viii
Acknowledgements ix
List of publications xi
Chapter 1. General Introduction 1
Plant-soil feedback theory 1
Influence of plant species on plant-soil feedback: soil conditioning 2
Root deposition 3
Litter deposition 6
Influence of soil conditions on plant-soil feedback 8
Soil feedback mediated by chemical properties 9
Soil feedback mediated by biological properties 10
Empirical evidence for importance of plant-soil feedback in
vegetation succession 12
Implications of the plant-soil feedback for restoration of ecosystems 16
The jarrah forest 17
Distribution and climate 18
Soils 19
Ecological aspects 19
Evidence of potential mechanisms of soil conditioning by jarrah forest plants 21
Mining activity and restoration of the jarrah forest 22
Thesis outline and research objectives 29
Chapter 2. General methods 30
Response variables 30

vi
Soil chemical properties 30
Water content and water holding capacity 30
Soil biology and associated properties 31
Microbial biomass-carbon (C) 31
Phospholipid-fatty acid (PLFA) profiles 31
Community level physicological profiles (CLPP) 32
Nematode trophic community 33
Chapter 3. Soil conditioning and plant-soil feedbacks associated with
jarrah forest plants are soil-context dependent 34
Introduction 34
Materials and methods 36
Sampling and experimental design 36
Response variables 40 Statistical analyses 49
Results 42
Changes in properties of soils conditioned during phase one 42
Unmined soil 42
Restored soil 44
Individual PSF 45
Discussion 47
Chapter 4. Plant-soil feedback unaffected by soil condition in a
post-mining restoration chronosequence 54
Introduction 54
Materials and methods 57
Sampling and experimental design 57
Response variables 58
Statistical analyses 58

vii
Results 62
Properties of field-conditioned soils across the chronosequence
and their trajectory of change 62
Soil feedback of soils from the chronosequence 69
Individual PSF with respect to soil age 72
Discussion 73
Chapter 5. Long-term conditioning of soil by eucalypts and pines does
not alter the plant-soil feedback associated with native jarrah 83
Introduction 83
Materials and methods 86
Sampling sites and experimental design 86
Response variables 87
Statistical analyses 87
Results 88
Properties of soils conditioned in the field by pines, jarrah, and blue gums 88
Individual plant-soil feedback 94
Discussion 94
Chapter 6. Concluding discussion 101
References 106
Appendices 123
Appendix 1: Carbon substrates used for CLPP 123
Appendix 2: Publications 124

viii
THESIS DECLARATION
The thesis was substantially completed during my period of enrolment in a PhD degree
in the School of Earth and Environment at the University of Western Australia and has
not been previously accepted for a degree at this or any other institution.
The thesis is my own composition, conducted under the supervision of Mark Tibbett
and Rachel J. Standish. Supervisors assisted with establishing the direction of the
research, experimental design and planning, offered technical advice and provided
editorial input to drafts. I designed the experiments, conducted field, glasshouse, and
laboratory work, as well as data analyses.

ix
ACKNOWLEDGEMENTS
I would like to express my sincere gratitude and appreciation to my supervisors Mark
Tibbett and Rachel Standish for their support, advice, and encouragement throughout
my candidature, and to teach me the meaning of being a ‘Scientist’.
I am very appreciative to the Consejo Nacional de Ciencia y Tecnología (National
Council on Science and Technology of Mexico) for funding my PhD studies. I also
acknowledge all the staff members of the Graduate Research School for their excellent
job and for financially support my stay in the South Australian Research and
Development Institute (SARDI) in Adelaide.
Thanks to all the staff in the School of Earth and Environment and Soil Science and
Plant Nutrition department, especially Tim Morald for his help during field and
laboratory work, Michael Smirk, Evonne Walker, and Darryl Roberts for his technical
assistance during laboratory work. Robert Creasy and staff for providing the necessary
materials and equipment at the plant growth facility during glasshouse work. Yoshi
Sawada, Natasha Banning, and Suman George for their advice on methodological
aspects of my research. Margaret Pryor, Gail Ware and Karen Newham for making all
the administration work easy.
Enormous thanks to John Koch from Alcoa World Alumina Australia for his assistance
and support for field work. Jackie Nobbs from SARDI to training me on identification
of trophic groups of nematodes, and Vivien Vanstone, Sarah Collins and staff from the
Department of Agriculture and Food of Western Australia (DAFWA) for help with
nematode extraction. I thank Ken Dodds and staff from the ChemCentre, and Richard
Hobbs from the School of Plant Biology (UWA) for their support to complete the PLFA

x
analyses. Deborah Lin, Khalil Kariman and Alonso Calvo Araya for their help during
critical phases of the experiments.
I would like to thank all members of the Soil Ecology group and the Ecosystem
Restoration and Intervention Ecology Research group (ERIE), and postgraduate
students at the Soil Science and Plant Nutrition department for your advice, support, and
friendship.
Finally I would like to express gratitude and appreciation to my family and friends for
your love, support, and motivation.

xi
LIST OF PUBLICATIONS
Publications arising from this thesis (Appendix 1):
Orozco-Aceves, M., Standish, R.J., Tibbett, M., 2015. Soil conditioning and plant-soil
feedbacks in a modified forest ecosystem are soil-context dependent. Plant Soil DOI
10.1007/s11104-015-2390-z
Orozco-Aceves, M., Standish, R.J., Tibbett, M., 2015. Long-term conditioning of soil
by plantation eucalypts and pines does not affect growth of the native jarrah tree. Forest
Ecol Manage 338:92-99.
Presentations in Scientific events
Orozco-Aceves, Martha; Tibbett, Mark; Standish, Rachel J. 2011. Niche construction by
plants in soils modified by bauxite mining in the jarrah forest in Western Australia.
Student Symposium of the Australasian Plant Pathology Society. South Perth, Western
Australia, 27 October.
Orozco-Aceves, Martha; Tibbett, Mark; Standish, Rachel J. 2011. Niche construction by
plants in soils modified by bauxite mining in the jarrah forest in Western Australia.
Rhizosphere 3 Conference, Perth, Western Australia, 25 – 30 September.


1
CHAPTER 1
General Introduction
Plant–soil feedback theory
Plant-soil feedback (PSF) theory describes how plants perceive and respond to the
edaphic environment by changing the physical, chemical and biological properties of
soil, that in turn affect plant performance (Bever 1994; Bever et al. 1997; Bever 2003).
Plant-soil feedback can be divided into two processes for study and understanding, these
are: plant conditioning (Harrison & Bardgett 2010; Pernilla et al. 2010; van de Voorde
et al. 2011) and, soil feedback (van der Putten et al. 2013; Figure 1). The two processes
occur simultaneously and the resulting effects (i.e. described by their magnitude and
direction) can be positive or negative. A positive PSF has occurred when there is an
increased plant biomass produced in soil conditioned by the same plant species (often
referred to as ‘home soil’) compared with the plant biomass produced in soil
conditioned by a different plant species (often referred to as ‘away soil’; Mills & Bever
1998; van der Heijden et al. 2008). Positive or negative PSF can ultimately shape the
structure and function of the entire community if the PSF is large relative to intrinsic
growth differences among plant species to affect the outcome of competitive
interactions (Wardle et al. 2004; Bardgett et al. 2005; van der Putten et al. 2009).
The mechanistic basis by which plant performance is regulated by PSF has been
described. A negative PSF occurs due to the accumulation of parasites, pathogens, and
root herbivores in the rhizosphere that can directly remove C and nutrients from plant
tissues and reduce root uptake capacity. Examples of soil pathogens that can act as
agents of negative PSF are the members of the genus Pythium (oomycetes), the fungus
Fusarium oxysporum, and plant-parasitic nematodes (van der Putten & Peters 1997;
Mills & Bever 1998; van der Stoel et al. 2002; Mangan et al. 2010; Liu et al. 2012).

2
Plants show variable degree of susceptibility to parasites and pathogens, and so the
multiple interactions between these groups of organisms and plant species tend to be
species-specific. Negative PSF can be also associated to plant-symbiont interactions that
confer nutritional advantage to specific plant species over other species (Mills & Bever
1998; van der Putten 2000; Mangan et al. 2010; Liu et al. 2012).
A positive PSF is driven by the accumulation of beneficial organisms in soil around
roots such as those within the soil food webs, and the establishment of mutualistic
symbionts such as N-fixing bacteria and arbuscular-mycorrhizal fungi (AMF; Ferris et
al. 2001; Reynolds et al. 2003; Bardgett et al. 2005; Compan et al. 2005; Ahmad et al.
2008; Bever et al. 2012). Soil food webs unlock nutrients present in organic matter,
which improves the rates of recycling and turnover, thus increasing nutrient availability
and plant productivity (Bever 1994; Bever et al. 1997; De Deyn et al. 2004; Wardle et
al. 2004; Xiaoyun et al. 2007; van der Heijden 2008). Arbuscular-mycorrhizal fungi
increase interplant transfer of C and nutrients via hyphal links, and enhance access to
limiting nutrients such as P, thus increasing plant growth (Bever 2002; Reynolds et al.
2003; van der Heijden 2008; Bever et al. 2010).
Influence of plant species on plant-soil feedback: soil conditioning
Plant species can condition the physical, chemical and biological properties of soil
(Kyle et al. 2007; Kulmatiski et al. 2008; van der Putten et al. 2013) by inputs of
chemical compounds and organic matter such as root and litter deposition (Marschner
1995; Neumann & Römheld 1999; Uren 2001; Bertin et al. 2003; Ehrenfeld et al. 2005;
Neumann 2007; Pang et al. 2010). The conditioning of soil can be direct or indirect.
The first involves a direct plant effect on soil properties, for example root deposition
can directly improve the structure of soil by addition of mucilage. The indirect
conditioning is mediated by other soil elements, for example, C released to soil via root

3
deposition promotes an increased microbial biomass, which in turn positively impact
plant growth. Mechanisms for conditioning of soil via root and litter deposition will be
discussed in more detail in the following sections.
Figure 1. Plant-soil feedback (PSF) can be divided in two: 1) plant conditioning describing the effect of plants on soil properties by influence of root exudation and root and litter deposition, and 2) soil feedback describing the effect of physical, chemical and biological properties of soil on growth, productivity, and reproductive success of plants.
Experimentally, it has been observed that soil conditioning is influenced by the
procedure used. There are two main approaches to obtain conditioned soil; these are the
‘natural experiment approach’ where plant-conditioned soils are collected from single
plants in the field, and the ‘cultivation experiment approach’ where soils are
conditioned under controlled conditions in the glasshouse (Kulmatiski & Kardol 2008;
Pernilla et al. 2010; van de Voorde et al. 2012).
Root deposition
Root deposition comprises exudates and dead roots (Uren 2001), and accounts for 30 to
60% of the C fixed by photosynthesis that is translocated to roots, from which up to
1. P
lant
con
diti
onin
g 2. Soil feedback
Plant-soil feedback

4
70% is released to soil (Marschner 1995; Brimecombe et al. 2001; Neumann &
Römheld 2001; Figure 2). Root deposition helps plants grow, especially under
conditions of environmental stress such as mechanical impedance, anaerobiosis,
drought, presence of pollutants, and nutrient deficiency (Marschner 1995; Uren 2001;
Nautiyal et al. 2008; Martin et al. 2014). Root deposition also varies with plant species,
cultivars, and plant developmental stage (Marschner 1995; Uren 2001). This
observation implies that root deposition to soil can vary within the life span of a plant
with implications for the conditioning of soil properties at a local level and subsequent
levels of community organisation through its effects on succession of the plant
community.
Root exudates directly influence availability of nutrients such as K, Ca, Mg, P, S, Fe,
and other micronutrients by modifying the soil pH (Jones 1998; Uren 2000; Bais et al.
2008; Richardson et al. 2009). Plants excrete inorganic ions (i.e. HCO3−, OH−, H+) and
organic acids (i.e. citric, malic, malonic) that solubilise or mobilize soil nutrients to
make them readily available for uptake by plant roots (Dakora & Phillips 2002;
Ehrenfeld et al. 2005; Richardson et al. 2009; Carvalhais et al. 2011). Plant-driven
changes of soil pH help plants to overcome toxic levels of elements present in soils (i.e.
Al; Jones 1998; Dakora & Phillips 2002). Also, by altering the pH of soil, plants have
major impact on the structure of microbial communities around the root (Badri &
Vivanco 2009; Richardson et al. 2009).
Plants release to soil chemical signals to attract beneficial and harmful organisms to
colonize the rhizosphere. Among beneficial organisms there are those that tend to act as
agents of positive PSF, for example symbionts such as N-fixing bacteria and AMF.
Symbiotic organisms mobilise and transfer nutrients (i.e. N, P) to plants (Hirsch et al.
2003; Atkinson 2006; Sbrana 2006). Root exudates also serve as sources of C for free-

5
living soil organisms (Bais et al. 2008), which in turn promote plant growth, mineralise
nutrients, detoxify pollutants, modify soil structure, and regulate the abundance of plant
pests and other opportunistic species (Neher 1999; Wardle 1999; Ferris et al. 2001).
Agents of negative PSF such as oomycetes, pathogenic fungi, and plant-parasitic
nematodes (Werner 2001; Nelson 2006; Bais et al. 2008; Badri & Vivanco 2009; Perry
& Tovar-Soto 2012) are also attracted to plant roots by chemical signals released by
plants in a species-specific manner (Hirsch et al. 2003).
Some plant species can release allelochemical compounds into soil that negatively
affect soil organisms and other plants. The main group of allelochemical compounds are
phenolic acids and their derivatives: terpenoids, steroids, coumarins, flavonoids,
alkaloids, cyanohydrins and tannins (Lavelle & Spain 2003). An important group are
the flavonoids, which can directly and indirectly influence chemical and biological
properties of soil by affecting the C, N, P, S cycles (Lavelle & Spain 2003; Cesco et al.
2012; Lotina-Hennsen et al. 2006; Hadacek 2008).

6
Figure 2. Classification, quantities and release mechanisms of root deposition. Reproduced from Neumann (2007).
Litter deposition
Litter chemistry (i.e. C:N ratio, lignin and phenolic compounds content) is a species-
specific trait of plants that directly influences the physical, chemical and biological
properties of soil (Hobbie 1992; Guo & Sims 1999; Berg & McClaugherty 2008;Witt &
Setälä 2009; Uselman et al. 2012). Litter decomposition releases nutrients that alter the
nutritional status of the soil that in turn affects plant nutrition (Lugo et al. 1990;
Ganjegunte et al. 2005; Berg & McClaugherty 2008; Uselman et al. 2012). Litter
decomposition produces humus that improves soil structure (Jastrow et al. 1998; Malik
& Scullion 1998). Functional aspects of soil are also influenced by litter deposition, for
example, plants producing litter with low C:N ratios (i.e. high-quality litter) promote a
soil biota dominated by bacteria and its predators (i.e. protozoa, bacterial-feeding
ROOT DEPOSITION
LYSATES
Passive release from damaged
and sloughed-off root cells
Root turn-over
ROOT BORDER CELLS
Controlled release of intact calyptra cells
EXUDATES
Release from intact root cells
DIFFUSATES SECRETIONS EXCRETIONS
Passive release by diffussion
Controlled release of
compounds with specific
functions
Controlled release of metabolic
waste productisLow-molecular-
weight-compounds
Allelochemicalcompounds
(i.e. phenolics, phenol,
flavonoids)
PhytosiderophoresCarboxylates
PhosphohydrolasesMucilage
Lactic acid

7
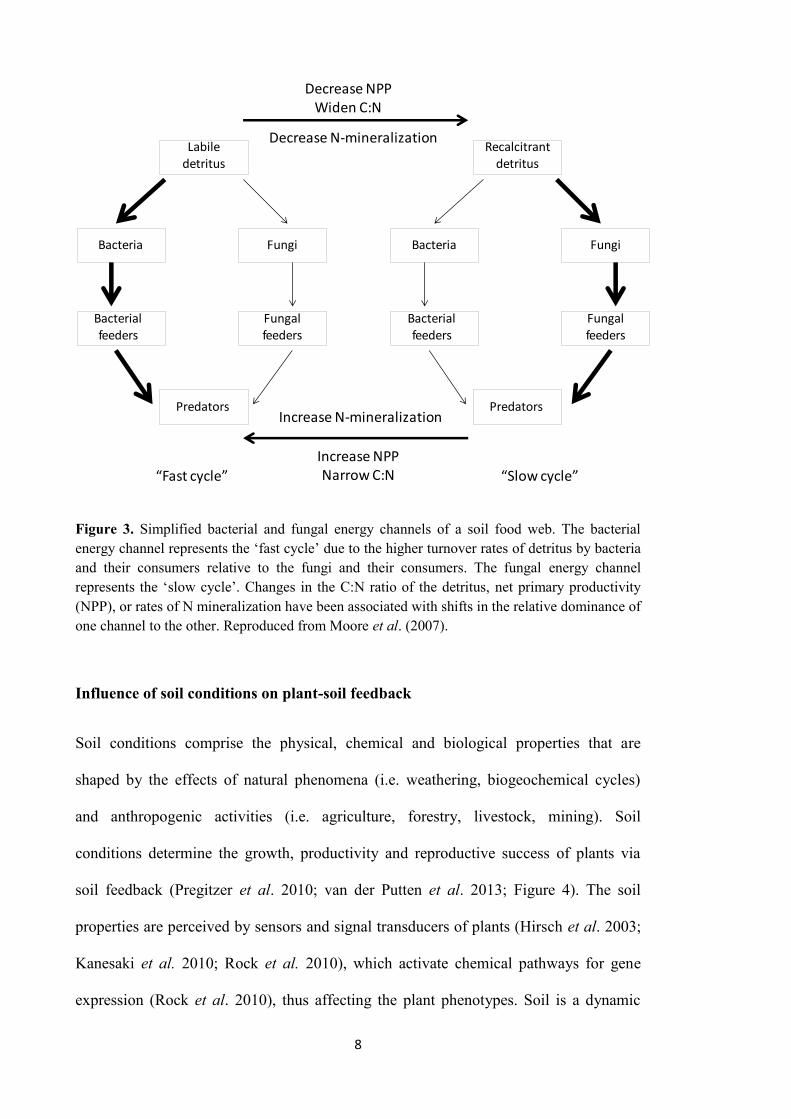
nematodes; Figure 3). As a consequence, the bacterial-based energy channel of organic
matter decomposition predominantly operates in soil. In contrast, plants producing litter
with high C:N ratios (i.e. low-quality litter) promote a soil biota dominated by fungi and
its predators (i.e. fungal-feeding nematodes, collembolans, oribatid mites), and then the
fungal-based energy channel of decomposition predominantly operates in soil (Moore &
Hunt 1988; Scheu et al. 2005). The bacterial-based energy channel is the fast track for
organic matter decomposition, in contrast to the fungal-based energy channel, which is
much slower (Moore & Hunt 1988; Figure 3).
Another mechanism by which plants can condition the soil properties via litter
deposition is the release of allelochemical compounds that are leached from fallen litter.
These allelochemical compounds produce alterations to the chemical and biological
properties of soil. For example, degradation products of litter lignin such as quinines
and phenols are involved in series of reactions that produce H2O2 (Inderjit & Malik
1997; Inderjit & Weiner 2001), which is toxic to microbes due to peroxidation of lipids
of cell membranes. Consequently, membrane permeability increases and destabilization
of cell processes occurs (Weir et al. 2004). Phenolic compounds also have inhibitory
effects on N-fixing microorganisms, symbiotic or free-living; monoterpenes negatively
affect nitrifying bacteria, and a wide variety of essential oils show antimicrobial activity
(Chalkos & Karamanoli 2006).
8
Figure 3. Simplified bacterial and fungal energy channels of a soil food web. The bacterial energy channel represents the ‘fast cycle’ due to the higher turnover rates of detritus by bacteria and their consumers relative to the fungi and their consumers. The fungal energy channel represents the ‘slow cycle’. Changes in the C:N ratio of the detritus, net primary productivity (NPP), or rates of N mineralization have been associated with shifts in the relative dominance of one channel to the other. Reproduced from Moore et al. (2007).
Influence of soil conditions on plant-soil feedback
Soil conditions comprise the physical, chemical and biological properties that are
shaped by the effects of natural phenomena (i.e. weathering, biogeochemical cycles)
and anthropogenic activities (i.e. agriculture, forestry, livestock, mining). Soil
conditions determine the growth, productivity and reproductive success of plants via
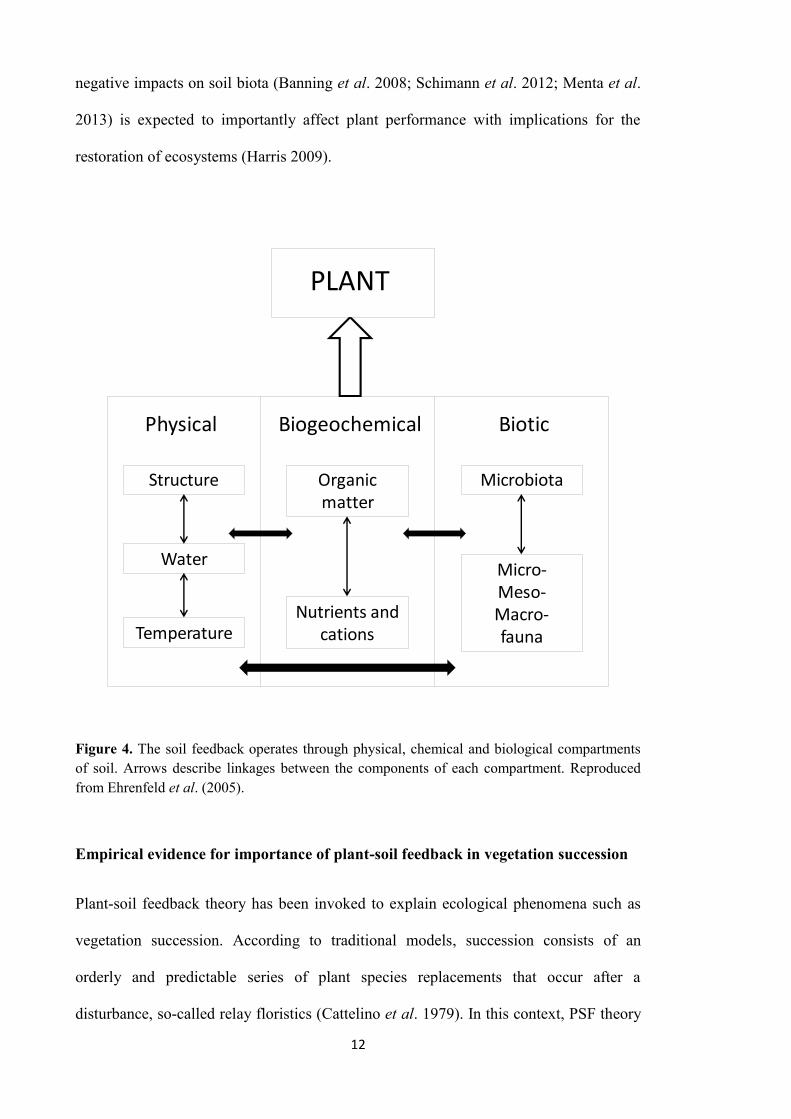
soil feedback (Pregitzer et al. 2010; van der Putten et al. 2013; Figure 4). The soil
properties are perceived by sensors and signal transducers of plants (Hirsch et al. 2003;
Kanesaki et al. 2010; Rock et al. 2010), which activate chemical pathways for gene
expression (Rock et al. 2010), thus affecting the plant phenotypes. Soil is a dynamic
Decrease NPPWiden C:N
Decrease N-mineralizationLabile
detritus
Bacteria
Bacterial feeders
Predators
Fungi
Fungal feeders
Recalcitrant detritus
Bacteria Fungi
Bacterial feeders
Fungal feeders
Predators
Increase NPPNarrow C:N
Increase N-mineralization
“Fast cycle” “Slow cycle”

9
entity and its properties experience changes over time, therefore plant species might be
subjected to differential soil feedback during ontogeny, with variable effects on
individual PSF (i.e. the PSF associated to individual plant species; Kardol et al. 2013;
van der Putten et al. 2013).
Research on soil feedback generally tests the simultaneous effects of multiple soil
properties on plant biomass production (Bonanomi & Mazzoleni 2005; Pregitzer et al.
2010; Brandt et al. 2013). However, it is likely that different soil properties act as key
drivers of the soil feedback during the life time of a plant, and so understanding which
of these drivers are operating at any one time will necessary involve detailed study of
PSF associated with plants of increasing age (Kardol et al. 2013). The soil feedback
mediated by physico-chemical properties is well documented and understood,
particularly for young plants, compared with understanding of those mediated by
biological properties and for mature plants. The knowledge of soil feedback mediated
by biological properties is rapidly developing (Miki 2012; Kardol et al. 2013; van der
Putten et al. 2013), but pieces of information are still missing. In addition, we lack
information regarding the relative contributions or ‘weight’ of physico-chemical and
biological properties on the soil feedback. In the following section soil feedback
mediated by chemical and biological properties are briefly described.
Soil feedback mediated by chemical properties
Chemical properties of soil vary according to the characteristics of the parent material
from which it originated (Buscot 2005). Through weathering process, ions are removed
from rock and mineral, given to soil particular chemical properties such as
concentrations of S, P, K, Mg, Ca, Si, Na, Fe, and a characteristic pH (Gorbushina &
Krumbein 2005). Concentrations of the nutrient elements are essential for plants to
complete their life cycle. Low concentrations of mineral elements in soil can be limiting

10
for plant growth (Foth 1990; Marschner 1995), whereas high concentration of mineral
elements can induce nutritional imbalances or toxic effects to plants (Bohn et al. 2001).
Therefore, soil nutrients and their interactions are critical components of the soil
feedback.
Soil pH directly affects the composition of plant communities, which show a clear
differentiation along gradients of pH. Soil pH is directly related to concentrations of
cations, and plant community composition is specially affected by extreme or unusual
concentrations of particular cations (Ehrenfeld et al. 2005). Soil pH also affects the
structure of the microbial community, which drives the organic matter decomposition
process and thus the availability of nutrients for plant uptake (Ehrenfeld et al. 2005;
Moore & Hunt 1988). Generally, soil pH below 5 favours the growth of fungi, whereas
neutral soil pH favours the growth of bacteria (Madigan et al. 2012). In turn, a
microbial community dominated by fungi promotes the functioning of the fungal-based
energy channel of organic matter decomposition. In contrast, a microbial community
dominated by bacteria promotes the functioning of the bacterial-based energy channel
of decomposition (Moore & Hunt 1988; Madigan et al. 2012). Thus, soil pH influences
the rates at which organic matter decomposes in soil. Moreover, changes in soil pH due
to human activities such as agriculture, forestry, and mining are likely to have
significant consequences for PSF. I have used soil pH as an example, yet there are other
chemical properties, that if modified through human activities, could lead to
modifications to PSF that usually operates in the ecosystem.
Soil feedback mediated by biological properties
Soil microbes are very abundant in soil and are the first organisms making contact with
plants, so it is probable plants are subjected to their influence during initial stages of
development through diverse mechanisms; for example, through growth mediated by

11
chemical substances such as carbon and nitrogen, solubilisation of mineral elements for
their nutrition, such as phophorus, mineralization of organic matter, soil aggregation,
regulation of abundance of pests and pathogens, N-fixation, (Neher 1999; Wardle 1999;
Altieri 1999; Buscot & Varma 2005). Plants are also subjected to the direct or indirect
influence of other soil microbes that are less abundant in soil (i.e. chemolithothrophs,
authotrophs. In addition to microbes, plants are subjected to the direct and indirect
influence of other groups of organisms within the soil food web, which are in large
numbers in soil such as protozoa, nematodes, mites, collembola, and insects (Gupta &
Yeates 1997; Griffiths et al. 2007; Yeates et al. 2009; Figure 4). For example, plants are
directly influenced by free-living nematodes (i.e. bacterial-, fungal-feeding) that
mobilize and mineralize a considerable amount of N, which is then available for plant
uptake (Freckman 1988; Postma-Blaauw et al. 2005; Yeates et al. 2009). Plants are
directly influenced by plant-parasitic nematodes that cause severe damage to plants,
specifically due to root damage (Perry & Tovar-Soto 2012). Opposite to ther organisms
in soil, nematodes are able to move in water films, for this reason nematodes are
capable to reach plant roots and influence plant development positively or negatively
(Lavelle & Spain 2003). Migration of nematodes occurs in soil and between plants only
when water is available or, soil texture adequate (Castillo & Vovlas, 2007). Protozoa
and collembola positively influence plant productivity due to ingestion of propagules of
fungal and bacterial pathogens (Lavelle & Spain 2003; Coleman et al. 2004). In
summary soil micro, meso and macrofauna influence plant growth by recycling
carbohydrates and nutrients through mineralisation, transferring energy in the detritus
food chain, providing reservoirs of nutrients, detoxifying pollutants, modifying soil
structure, and regulating the abundance of pests and other opportunistic species (Neher
1999; Ferris et al. 2001). Plants are strongly influenced by soil feedback mediated by
biological properties, hence soil modification by human activities that generally has
12
negative impacts on soil biota (Banning et al. 2008; Schimann et al. 2012; Menta et al.
2013) is expected to importantly affect plant performance with implications for the
restoration of ecosystems (Harris 2009).
Figure 4. The soil feedback operates through physical, chemical and biological compartments of soil. Arrows describe linkages between the components of each compartment. Reproduced from Ehrenfeld et al. (2005).
Empirical evidence for importance of plant-soil feedback in vegetation succession
Plant-soil feedback theory has been invoked to explain ecological phenomena such as
vegetation succession. According to traditional models, succession consists of an
orderly and predictable series of plant species replacements that occur after a
disturbance, so-called relay floristics (Cattelino et al. 1979). In this context, PSF theory
PLANT
Physical Biogeochemical Biotic
Structure
Water
Temperature
Organic matter
Nutrients and cations
Microbiota
Micro-Meso-Macro-fauna

13
has been suggested as a mechanism driving the serial species replacement through
regulation of the amounts of plant biomass produced (Bever et al. 1997; Bever 2003;
Kardol et al. 2006; Kulmatiski et al. 2008; van der Putten et al. 2009).
Negative PSF more than positive PSF, has been described as a key driver of structural
changes of plant communities during succession (van der Putten & Peters 1997; van der
Stoel et al. 2002; Bonanomi et al. 2005b; Mangan et al. 2010; Liu et al. 2012). Plant-
soils feedback tends to be negative in early-successional communities (van der Putten &
Peters 1997; Kardol et al. 2006). Early-successional plants show the greatest maximum
growth rate but the highest level of susceptibility to pathogens and parasites, which
creates adverse conditions to plants, opening gaps for the establishment of other species
(Kulmatiski et al. 2008). Therefore, negative PSF induces a greater species turnover
both in time and space, increasing in this way the rate of succession and contributing to
species coexistence (van der Putten & Peters 1997; Mills & Bever 1998; Bever 2003;
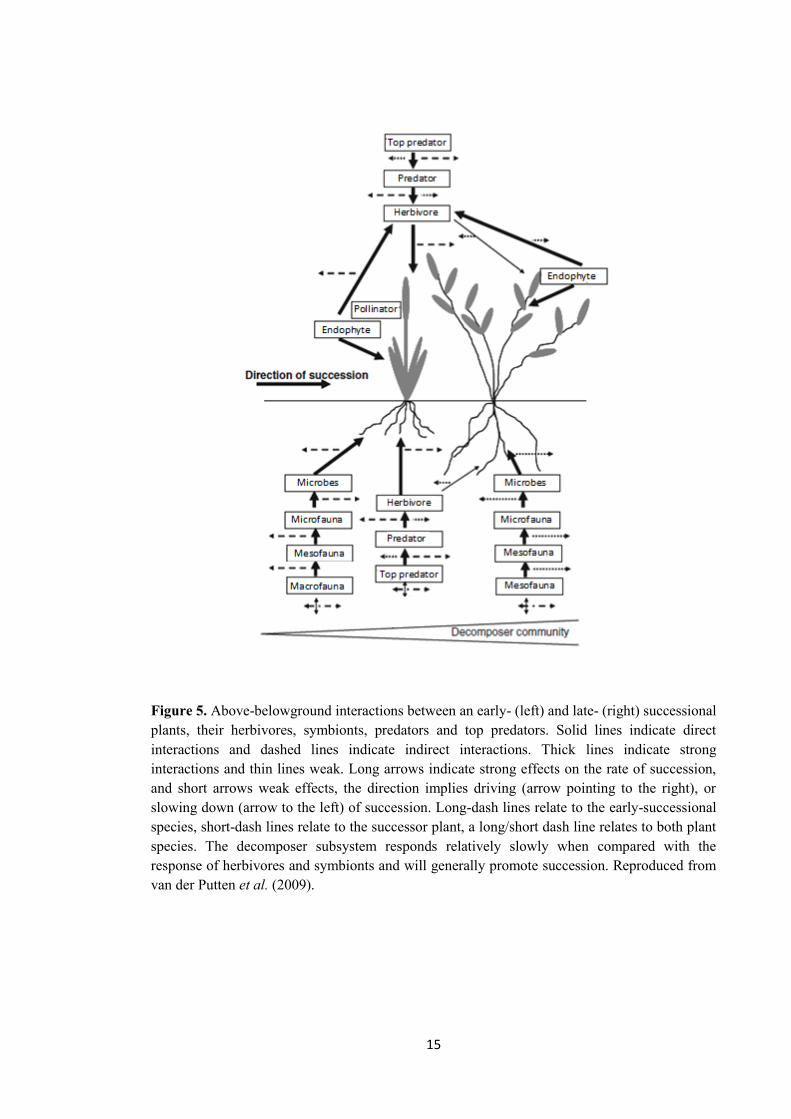
Kulmatiski et al. 2008; Figure 5).
The role of parasites, pathogens, and herbivores as agents of negative PSF has been
documented in a variety of ecosystems such as a tropical forest in Panama, a subtropical
monsoon forest in China, and a temperate black cherry forest in Europe (van der Putten
2000; Mangan et al. 2010; Liu et al. 2012). These findings suggest that negative PSF
operates across diverse ecosystems around the world, and in diverse contexts of soils
and plants (Bonanomi et al. 2005a; Bonanomi et al. 2005b; Mangan et al. 2010;
Reinhart 2012).
Positive PSF is likely to enhance plant dominance and so retard succession (De Deyn et
al. 2004). Plant-soil feedback tends to be positive in late-successional communities
(Bever 1994, Bever et al. 1997, Wardle et al. 2004), late-successional plants grow
slowly, and can establish mutualistic symbiosis (i.e. mycorrhiza, N-fixing bacteria),

14
which leads to favourable conditions for their growth, dominance, and persistence in
ecosystems (van der Heijden et al. 1998; Bever 2002; Ehrenfeld et al. 2005; Miki et al.
2010; Figure 5).
Some studies suggest that not only the successional stage of plant species influences
PSF during succession, but the interactive effects of both, plant species and soil
conditions shape PSF. For example, late-successional plants perform best in late-
successional soil (i.e. mature forest soil), and worst in early-successional soil (i.e.
cleared forest, abandoned agricultural fields). In contrast, early-successional plants can
perform bad in both early- and late-successional soils (Kardol et al. 2006). The
nutritional status of the soil can act as driver of PSF (Bezemer et al. 2006; Manning et
al. 2008; Meijer et al. 2011). Positive PSF plays a central role in early-successional
plant communities on nutrient poor soils. In these soils, positive effects of microbes on
plant productivity enhance the supply of limiting nutrients such as N and P to plants (De
Deyn et al. 2004). This microbe-mediated supply is critical in such ecosystems since up
to 90 % of P and N are provided by AMF and N-fixing bacteria (van der Heijden et al.
2008). In contrast, negative PSF is expected to contribute to species replacement and
diversification at more fertile sites. The relative importance of soil biota in suppressing
dominant plant species will increase with decreasing soil fertility (De Deyn et al. 2004).
Other hypothesis points out that plant-soil interactions are complex, during succession
this complexity enhances plant species diversity by delaying community convergence
(reduced beta diversity). Diversity enhancement occurs because the complex plant-soil
interactions lengthen the time during which local species composition change. Due to
the longer time for changes in species composition, a high level of beta (community)
and gamma (regional) diversity at early stages of succession is maintained for a long
time despite eventual community convergence (Fukami et al. 2013).
15
Figure 5. Above-belowground interactions between an early- (left) and late- (right) successional plants, their herbivores, symbionts, predators and top predators. Solid lines indicate direct interactions and dashed lines indicate indirect interactions. Thick lines indicate strong interactions and thin lines weak. Long arrows indicate strong effects on the rate of succession, and short arrows weak effects, the direction implies driving (arrow pointing to the right), or slowing down (arrow to the left) of succession. Long-dash lines relate to the early-successional species, short-dash lines relate to the successor plant, a long/short dash line relates to both plant species. The decomposer subsystem responds relatively slowly when compared with the response of herbivores and symbionts and will generally promote succession. Reproduced from van der Putten et al. (2009).

16
Implications of the plant-soil feedback for restoration of ecosystems
Plant-soil feedback has important implications for restoration of ecosystems, however,
studies on ecosystems restoration that have explicitly consider plant–soil linkages are
currently scarce (Harris 2009; Kardol & Wardle 2010; van der Putten et al. 2013). A
PSF approach considers that restoration of one of the ecosystems’ compartments (below
or aboveground) would be improved by inclusion or consideration of the other (Kardol
& Wardle 2010). For example, in temperate Australian grassy woodlands, the biomass
of exotic annual plant species (aboveground structure) was dramatically reduced by
adding to soil a C source (belowground manipulation) such as sucrose to deplete
seasonally available nitrate pool, which otherwise favours the growth of invasive
species (Prober et al. 2009).
On the other hand, successional-based restoration management is usually inefficient due
to strong feedbacks operating between above and belowground compartments, which
can make the ecosystem resilient to restorative change (Suding et al. 2004). A PSF
approach allows identifying undesirable PSF, and efficiently addressing the constraints
(Suding et al. 2004; Suding & Hobbs 2009). For example, in the Chihuahua desert,
abandoned agricultural fields have slow infiltration rates as consequence of agriculture
practice. The little rainfall that occurs is lost to runoff, creating a positive feedback
where drier conditions promote poor vegetation growth that in turn holds less water
(identification of undesirable PSF). Therefore, restoration treatments that scratch the
soil surface are advisable, once water is held grasses regenerate spontaneously, which in
turn catches more water, thereby reversing desertification (problem addressed by
breaking the undesirable PSF; Whisenant 1999).
In human-modified ecosystems the properties of soil can be importantly altered, this
implies that processes such as plant conditioning and soil feedback are likely to alter,

17
with unpredictable effects on individual PSF associated with specific plant species
(Manning et al. 2008; te Beest et al. 2009; Kardol & Wardle 2010). For example, in
many industrialized countries ex-arable soils display modified properties such as high
fertility, simplified soil communities, and seed limitation. Together, these aspects
produced negative feedbacks on native plant species, which constrained restoration of
the historic ecosystem (Kardol & Wardle 2010). In Florida, post-phosphate mining soils
have modified properties such as increased P concentration and clay, the latter lowers
air and water movement and can range from quite sticky when wet to brick-hard when
dry. These soil conditions do not promote successional change, but only the growth of
colonizing species in these areas, so that the plant community composition of mined
sites does not resemble that of the native ecosystem even after 30 years (Brown 2005).
In Italy, soils post-sand and gravel mining featured altered texture, decreased porosity,
pH and microbial population. Plant-soil feedback developing in these soils is
unfavourable for plant growth, consequently, mined sites are almost devoid of plant
cover (Menta et al. 2013). Examples like these show how soil modification can alter the
patterns of growth, dominance and abundance of plant species via PSF phenomena.
Ultimately, altered PSF can make an ecosystem resistant to restorative change, or can
promote an undesirable restoration trajectory (Suding et al. 2004; Suding & Hobbs
2008).
The jarrah forest
The jarrah (Eucalyptus marginata) forest is an ecosystem native to the south-western
corner of Western Australia. It occurs on the highly leached soils of the Darling Plateau,
which are characterized by strong limitations of nutrients such as P and N (Dell et al.
1989). To overcome nutritional limitations of soil, native plant species have developed
series of strategies to efficiently acquire nutrients (Lambers et al. 2007; Hopper 2009;

18
Sardans & Peñuelas 2013). This information suggests that strong links between plants
and soils operate in the jarrah forest (Abbott & Robson 1977; Malajczuk & McComb
1977; Malajczuk 1979; Malajczuk & McComb 1979; Grierson & Adams 2000;
Lambers et al. 2007; Bleby et al. 2009; Hopper 2009), which makes the ecosystem ideal
to study PSF phenomena. Moreover, the jarrah forest has been under strong
anthropogenic pressure due to activities such as mining for bauxite (alumina). After
mines close, the sites are restored to native forest (Koch 2007b), however, restored
jarrah forest soil displays modified properties (Ward 2000; Jasper 2007; George et al.
2010; Tibbett 2010) that are likely to alter PSF with important implications for the
restoration of the ecosystem. Restored jarrah forest offers a field laboratory to study
PSF phenomena, there is availability of restored soils of diverse types (i.e. types of
forest, types of restoration practices, types of conditioning) and ages (post-restoration)
to test the importance of soil contexts on PSF, and how PSF evolves as succession
proceeds. In the following sections relevant environmental aspects on the jarrah forest
are described.
Distribution and climate
The jarrah forest distribution extends from Toodyay in the north of Albany in the south
and its eastern limit is approximately the 750 mm rainfall isohyets (Figure 6). The
climate is Mediterranean, which is characterized by cool, wet winters and hot, dry
summers. Average summer monthly maximum temperature is 28 °C and average winter
minimum is 5 °C. Annual rainfall ranges from 750 to 1400 mm yr-1. Seasonal summer
drought may last from four to six or seven months (Churchward & Dimmock 1989;
Gentili 1989).

19
Soils
Five broad groupings of soils can be recognized in the jarrah forest, these are sandy
gravels, siliceous sands, earths, non-calcareous loams, and duplex soils (Churchward &
Dimmock 1989; Gentili 1989). Jarrah forest soils (< 2 mm fraction) have lower stores of
total and extractable forms of nutrient elements, mainly P and N. Part of the reason for
the low stores is that an average of about 75% of the whole soil consists of ferruginous
gravel (> 2 mm fraction). Although the gravel contains considerable amounts of some
nutrients, they are physically and chemically inaccessible to plant roots. Thus gravel
limits the amount of nutrients that plants can extract from a given volume of soil,
however, the soil is still the major nutrient store, followed by the aboveground biomass
and the litter (Hingston et al. 1989). For this reason soil feedback mediated by chemical
properties is likely to have strong influence on plant biomass production. In turn, plant
species of the jarrah forest might have efficient conditioning capacities to condition soil
properties in favour of their growth.
Ecological aspects of the jarrah forest
Paradoxically the jarrah forest, one of the most limiting ecosystems for plant growth in
the world, is a diversity hotspot due to its exceptional concentration of endemic species
and high biodiversity (Myers et al. 2000). Contrary to what one might think, the
environmental conditions prevalent in the ecosystem provide considerable scope for
diversity in the floristic composition. This diversity includes a vast adaptive
characteristics and structural features of the plant communities. Jarrah (E. marginata) is
the dominant tree in the forest, with long-lived resprouting species commonly
dominating the understorey, accompanied by short-lived obligatory reseeding species
which are stimulated to germinate by high soil temperatures in a fire, and a few species
which require dispersal from adjacent unburnt sites (Bell & Heddle 1989).

20
Fire, which is the main natural disturbance in the jarrah forest, has marked influence on
the vegetation composition, eliminating intolerant species and encouraging the spread of
fire-resistant species. In adapting to this environmental pressure, current members of the
flora show a variety of adaptive traits to maintain a presence in the post-fire community.
Two major fire-response syndromes are apparent in the jarrah forest: resprouters and
reseeders. Resprouting species are those species whose post-fire presence depends
predominantly on individual survival. Individuals of reseeding species die as a result of
the fire, and their continued presence in the post-fire community depends on
establishment of a new generation of seedlings from soil-stored or canopy-stored seeds
(Dell et al. 1989; Bell 2001; Keeley et al. 2012).
In the jarrah forest, plant communities contain a mix of resprouters and reseeders,
however, fire frequency influences the proportions of resprouters and reseeders in those
communities. Short intervals between fires promote the success of resprouters, which
recover quickly after fire and have the capacity to outgrow and outcompete other plant
species. Resprouter species also tend to be resilient to even highly frequent burning and,
therefore, do not have the risk of localized extinction as do reseeders (Bell 2001; Pekin
et al. 2012).
Long intervals between fires favor the reseeding strategy. With increasing time between
fires, fuel loads tend to accumulate, and the size of individuals increases with
progressive self-thinning. Fires in such long-unburned areas would be more intense,
resulting in a greater likelihood of the deaths of individual resprouters, as well as all
reseeder individuals. Also, the size of the post-fire openings may be expected to
increase with longer intervals between fires. In this situation, nutrient release following
fire may be great, and the rapid growth rate of recruits within the relatively larger

21
clearings may mean that reseeder species will eventually outcompete slower-growing
resprouter seedlings (Bell 2001; Pekin et al. 2012).
In the jarrah forest, the variability in the post-fire environment following different fires
constitutes a selection pressure that maximizes genotypic variation, plant species
diversity is greatest shortly after fires, when the vegetation simultaneously contains both
resprouting and reseeding species. An initial post-fire increase in community species
richness and diversity followed by a gradual decline is the typical pattern in most of the
world's fire-prone, Mediterranean-climate ecosystems. The decline in species richness
and diversity is predominantly a decline in living representatives (Bell 2001; Burrows
2008).
Succession in Western Australian plant communities generally follows the initial
floristic composition model (Cattelino et al. 1979; Grant 2006; Norman et al. 2006),
with a full complement of the flora represented as aboveground living individuals at the
beginning of the fire interval and a mix of live individuals and quiescent seeds at the
end of a fire cycle. Resprouter species populations in Australian communities are multi-
aged, with cohorts of seedlings surviving only if germinated early in the interfire
interval. Populations of reseeders, on the other hand, are nearly always even- and single-
aged. The high proportion of resprouters in the southwestern Western Australian plant
communities also means that post-fire plant cover and biomass recovery is very rapid,
with a leveling off of these community attributes after only 5-10 years (Dell et al. 1898;
Bell 2001).
Evidence of potential mechanisms of soil conditioning by jarrah forest plants
The native plants in the jarrah forest have the capacity to cope with low nutrient
availability by their slow growth, leaf adaptations, and recycling of nutrients within
individual plants (Hingston et al. 1989), and also due to processes occurring

22
belowground. Native plants can efficiently condition the soil properties, especially the
most limiting P and N. Conditioning of P by jarrah forest plants includes the formation
of specialized cluster roots that solubilise P by releasing root exudates, particularly
organic acids (Lambers et al. 2008; Hopper 2009; Lambers et al. 2012). Another
strategy to condition P is via formation of mycorrhizal roots to increase the uptake
efficiency of the nutrient from soil (Abbott & Robson 1977; Brundrett & Abbott 1995;
Solaiman & Abbott 2003). Conditioning of N requires the formation of symbiotic
associations between N-fixing bacteria and legumes from the jarrah forest (Lawrie
1981).
In order to establish symbioses with AMF and N-fixing bacteria, jarrah forest plants
must release chemical signals via root exudates, which is considered a conditioning
mechanism. For example, flavonoids are released to attract symbionts to the rhizosphere
(Hirsch et al. 2003; Amalesh et al. 2011; Cesco et al. 2012). Jarrah, the dominant tree in
the jarrah forest releases exudates containing a variety of sugars, amino acids and
organic acids (Malajczuk & McComb 1977), which might contribute to condition the
physical, chemical and biological properties of soil. As a result, large populations of
bacteria, actinomycetes, saprophytic fungi and AMF inhabit the rhizosphere and roots
of jarrah trees (Malajczuk 1979; Malajczuk & McComb 1979; Soilaiman & Abbott
2003).
Allelopathy comprises another mechanism by which jarrah forest plants can condition
the properties of soil (Willis 1999). Allelochemical compounds have inhibitory effects
on reproduction of soil biotic communities and seed germination, therefore the effects
of these substances can be indirectly observed as decreased microbial communities of
soil, and reduced rate of seed germination. Allelochemicals with antimicrobial action
have been detected in exudates and litter of eucalypt species and other Australian plants

23
such as Acacias (Deans 2002). For example, A. pulchella produces volatile (2- and 3-
methylbutanol, hexanol, pentanal, 2- and 3-methylbutanal, 4-methylacetophenone and
C-disulfide) and non-volatile allelochemicals (saponines; Alexander et al. 1978;
Whitfield 1981; D’Souza et al. 2004). Acacia dealbata releases allelochemical
compounds that negatively affect other plant species and soil microbes (Lorenzo et al.
2013). Acacia melanoxylon produces several phenolics and flavonoids with phytotoxic
activity (Hussain et al. 2011). Allelopathy is critical for plant persistence in ecosystems
such as the jarrah forest, where competition with other plants and soil organisms for
moisture and nutrients is intense, thus plants equipped with efficient inhibitory
chemicals raise their chances of survival (Willis 1999; Manoharachary & Mukerji
2006).
Mining activity and restoration of the jarrah forest
The jarrah forest has been mined for bauxite (alumina) since 1963. Mining is one of the
most destructive human activities to ecosystems, as a result, the physical, chemical and
biological properties of soil can be modified. Mining activity in the jarrah forest causes
alterations to soil conditions (Ward 2000; Jasper 2007; George et al. 2010; Tibbett
2010) that in turn might exert altered soil feedback on plants. In 1966 restoration
activities of the jarrah forest post-bauxite mining started. First restoration attempts
consisted in planting of fast-growing, exotic, softwoods, mainly pines (Pinus radiata)
and cypresses (Cupressus sp.), as well as eastern Australian hardwood eucalypts such as
Sydney blue gum (Eucalyptus saligna; Koch 2007a; Koch 2007b).
At the present time restoration process includes practices to improve the physical,
chemical, and biological properties of soil to facilitate vegetation succession,
encouraging the re-establishment of the original and particular characteristics of the
ecosystem (Koch 2007a; Koch 2007b). Approximately 550 ha of native forest are

24
restored per year (Bell & Hobbs 2007), however, to date there are still 3,600 ha of
plantations of non-native species in the jarrah forest that will be eventually harvested to
restore native forest for habitat preservation and exotic weed management
(Conservation Commission of Western Australia 2012).
The current mining and restoration process generally takes about 2–3 years. The
methods of restoration have changed and improved over the years as knowledge has
increased and as larger earthmoving machinery has become available. The significant
improvements have been mainly in the areas of landscaping after mining, soil return
methods, deep ripping to relieve compaction, selection of appropriate plant species for
restoration, plant propagation methods, and techniques to encourage return of fauna
(Koch 2007b).
The complete mining and restoration process includes the following steps (Koch
2007b):
Soil stripping: Soil is stripped in two layers. The process is called double
stripping. The top 15 cm (topsoil) contains the majority of the seeds, organic
material, plant nutrients, and microbial activity. It is important to return this
material on the surface during the restoration process. The second layer is from
15 cm down to the top of the lateritic duricrust layer and ranges from 10 to 80
cm deep, averaging 40 cm. This layer is called overburden and is stockpiled next
to the mined area to be returned to the pit floor during the restoration process.
Whenever possible, topsoil is direct returned. This process involves stripping
from one area that is about to be mined and immediately returning the soil to a
nearby area that is being restored. This ensures that the biological components of
the topsoil are not degraded by being stored in a stockpile.

25
Blasting: The lateritic duricrust is part of the bauxite ore deposit and averages 1
m deep. It is broken either by blasting or by ripping with a large remote control
bulldozer, a Komatsu 575. Ripping the duricrust is quieter than blasting and is
used in locations where blasting noise may impact on neighbours.
Mining: The broken duricrust and the underlying friable bauxite are loaded into
190-tonne trucks using large hydraulic excavators. The bauxite depth varies
from 2 to 10 m and averages about 4.5 m. The bauxite is trucked to a central
crusher that feeds onto a conveyor that supplies the refinery 20–25 km away.
Approximately 20 million tones of bauxite are mined each year from two mines
at Huntly and Willowdale (Figure 6).
Landscaping and pre-ripping: Following the completion of mining, bulldozers
push down the pit walls and shape the pits to a smooth rolling landscape,
ensuring that the topography created does not allow water to run from the mined
pits into adjoining unmined areas. Large rocks are buried but some are left on
the surface to provide fauna refuges. After landscaping, the pit floor is deep
ripped at 1.6-m spacing using a bulldozer fitted with a 1.5-m-long winged tine.
This breaks up the compacted pit floor, allowing the infiltration of water through
the soil profile and facilitates the growth of plant roots.
Overburden and topsoil return: Using scrapers, first the stockpiled overburden
and then the fresh topsoil (0–15 cm), usually sourced from a pit nearby that is
being prepared for mining, are spread on the area being restored. This ‘directly
returned’ topsoil contains most of the forest seed bank, nutrients, and soil
microbial activity, allowing a more rapid return of plant species and soil
processes. Where direct return is not possible, some fresh topsoil is screened
during the dry summer period to remove much of the inert gravel (50–60% of

26
the soil), and this fine fraction with its concentrated seed content is then spread
over pits, which have received stockpiled topsoil.
Return of logs and rocks for fauna habitat: Waste timber and rocks are placed in
the restored areas to provide habitat for both vertebrate and invertebrate fauna.
Without this activity, it would take hundreds of years for this type of habitat to
develop. Logs and rocks are used as shelter for foraging mammals, reptiles, and
a wide range of insects, spiders, and other invertebrates.
Contour ripping: The pre-ripping that is carried out after landscaping relieves
mining-related compaction and creates a friable rooting zone to 1.5 m depth.
Following topsoil and overburden return, the area is then ripped again to a depth
of 0.8 m on contour by a bulldozer using three tines. This final contour ripping
removes compaction caused by the soil return operation and produces contour
furrows, which allow water infiltration and provide rainfall erosion protection
until the vegetation develops
Seeding: A mixture of 78–113 plant species, including tree and understory
plants, is represented in the broadcast seed mix. Seed is collected within a
defined zone up to about 20 km from each mine to ensure that local genetic
material is used for restoration. The seeds are applied at a rate of about 1 kg/ha
via a purpose built air seeder attached to the ripping bulldozer. Seeds are sown
in the dry summer and autumn months. Appropriate germination treatments
(such as hot water or smoke) are applied to each species as required before
sowing.
Planting: Plant species that do not return from the soil seed bank or from the
broadcast seed are called ‘recalcitrant’ and are grown from cuttings, from scarce
seed quantities, or by tissue culture. Propagation is carried out at Alcoa’s
nursery, and the plants are hand planted during winter. Up to 28 species are

27
propagated. Up to 200,000 plants are grown and planted each year. Many of
these are dryland rushes and sedges that produce little viable seed. The grass-
like rushes and sedges are heavily grazed by kangaroos and need to be protected
while they are young. Small mesh bags are used to provide grazing protection.
Fertilizing: Diammonium phosphate fertilizer with potassium and micronutrients
is applied at 280 kg/ha by helicopter to newly restored areas in late winter each
year. This is a once-off application. The helicopter is used to prevent
recompaction of the ripped ground and damage to the contour furrows, which
protect against erosion. This is the last stage of the restoration operation.
Figure 6. Alcoa World Alumina Australia mining lease in the Darling Range of south-western Western Australia and the location of the bauxite mines at Jarrahdale, Huntly, and Willowdale. Reproduced from Standish et al. (2010). Soils for the experiment were collected from these three bauxite mines.
As a result of the mining and restoration process, restored soils tend to have reduced
biological activity, pH, and C, and increased P, N, K and gravel percentage (Ward 2000;
Jasper 2007; George et al. 2010; Tibbett 2010). However, many soil parameters showed

28
significant changes in the period 3.5–8.5 years after restoration. Levels of total nitrogen
in the top 10 cm of soil in restored areas increased from around 0.04–0.05% initially to
levels approaching those found in good-quality jarrah forest (0.10–0.30%) after 8.5
years. Soil pH declines in the surface layer after restoration. The rate of acidification
decreases in time, and the levels of other soil chemical parameters move towards the
range of values found in areas typically mined for bauxite (Ward 2000). Twenty-six
years post-restoration, organic matter quantity recovers to a level comparable to that of
unmined forest soil, soil N recovers faster than soil C, and recovery of microbial and
soluble organic C and N fractions is faster than total soil C and N (Banning et al. 2008;
George et al. 2010; Lin et al. 2011). Arbuscular mycorrhizal fungi recover to pre-
mining levels in bauxite restoration in five years. Ectomycorrhizal fungi are poorly
adapted to disturbance; however, they are able to reinvade through wind-blown spores.
The density of ectomycorrhizal fungi has been found to be equivalent in seven-year-old
restoration and adjacent forest, but both abundance and diversity are correlated with
development of a litter layer. Fortunately, rhizobia are known to be tolerant of soil
disturbance, and failure of N-fixation by legumes has not been reported in restoration
(Jasper 2007).
Despite a vast body of studies on restoration of jarrah forest after bauxite mining have
documented a recovery of properties of aboveground and belowground compartments of
the ecosystem, we lack information to predict how modified properties of soil will
affect the feedbacks between both compartments (i.e. PSF) since these studies have
been traditionally done on independent compartments of the ecosystem.

29
Thesis outline and research objectives
The overall aim of the studies described in this thesis was to determine the influence of
plant species (via conditioning process) and soil conditions (via soil feedback) on PSF
in the jarrah forest. Soil conditions comprise those of unmined soil and restored soil
post-bauxite mining. More specifically the research objectives were:
1. Determine the influence of plant species and soil context on PSF by examining
species-specific conditioning of soil properties by native plant species (i.e. Eucalyptus
marginata, Acacia pulchella, and Bossiaea ornata) in unmined and restored soils, as
well as individual PSF associated with these combinations of plants species and soil
contexts (Chapter 3).
2. Examine the influence of soil conditions on PSF by investigating the soil feedback
exerted by soils from a chronosequence of restoration ages (i.e. 1.5-, 7-, 22-year-old
soils post-bauxite mining, and an unmined soil) on individual PSF associated with B.
ornata plants (Chapter 4).
3. Investigate the influence of soil conditions caused by long-term conditioning (i.e. 35
years) of non-native (Pinus radiata and Eucalyptus saligna) and native species (E.
marginata) on individual PSF associated with E. marginata seedlings (Chapter 5).

30
CHAPTER 2
General methods
This chapter outlines the methods common to all experimental chapters only. Methods
specific to individual chapters, including experimental design and data analysis, are
described on a chapter by chapter basis.
Response variables
Soil chemical properties
Total N (%), total C (%), available (bicarbonate extractable) P (mg kg-1) and K
(mg kg-1), pH (CaCl2) and EC (dS m-1) were determined by the commercial laboratory
Cummings Smith British Petroleum (CSBP, Bibra Lake Western Australia). Total N
and total C were measured in a combustion analyser (LECO® USA), available P,
available K, pH, EC, were determined using standard methods described in Rayment &
Higginson (1992). Ammonium (NH4+ mg kg-1) and nitrate (NO3
- mg kg-1) analyses were
done simultaneously by the Skalar Autoanalyser (Skalar Analytical, The Netherlands) in
laboratories at the University of Western Australia.
Water content and water holding capacity of soils
To measure the water content of soil, the gravimetric method with oven drying was
used. This method consists in weighing a moist soil sample, which is then oven dried at
105 °C for 24 h and reweighed (Topp et al. 2008). To determine the water holding
capacity of soils, a known amount of soil was saturated with water in a cylinder, which
was then placed on an absorbent membrane until the excess water is drawn away by
gravity overnight. The water holding capacity was calculated based on the weight of the
water held in the sample vs. the sample dry weight (Rayment & Higginson 1992).

31
Soil biology and associated properties
Microbial biomass-carbon (C)
Microbial biomass-C was determined by chloroform-fumigation extraction technique
described in Coleman et al. (2004). Organic C in fumigated and unfumigated samples
was measured with a Total Organic-C Analyser (Shimadzu 5000A, Japan). Results from
unfumigated samples were subtracted from fumigated ones in order to obtain the
microbial biomass-C values. For calculations a kEC factor of 0.45 was used and data
were reported on a dry soil basis (µg C g-1 dry soil).
Phospholipid–fatty acid (PLFA) profiles
Phospholipid–fatty acid profiles reveal information about the community structure and relative
abundance of bacteria (Gram positive, Gram negative, actinomycetes), fungi (filamentous and
mycorrhizal), protozoa, other eukaryotic organisms including algae in soils (Cavigelli et al.
1995, Ranganayaki et al. 2001). Phospholipid-fatty acid profiles of soils were determined
as follows: total lipids were extracted from 8 g of freeze-dried soil using
chloroform:methanol:phosphate buffer at a ratio of 1:2:0.8 following the procedure
described in White et al. (1979). Then, solid phase extraction tubes (silica-bonded polar,
1 mL volume, Supelclean™) were used to separate phospholipids from neutral lipids
and glycolipids. Phospholipids were subjected to a mild alkaline hydrolysis to form
fatty acid methyl esters (FAMES) as described by Zelles & Bai (1993). Individual
PLFAs were detected using gas chromatograph with flame ionization detector (Agilent
7890A Dean Switch GCFID). Fifty μL (75μg mL-1) of methyl-nonadecanoate (Me-C19
Sigma) were added to FAMES extracts as internal standard. PLFAs were identified by
comparing their retention times with those of qualitative standards of methyl esters
(Supelco®). To calculate concentration of each FAME, the following formula was used:

32
Where PFAME and PISTD are the peak areas of fatty acid and the internal standard
respectively; μgStd is the concentration of the internal standard (μg μL-1 solvent); and W
is the dry weight of soil used (g). Finally, biomarkers for specific groups of soil
organisms such as bacteria (15:0i, 15:0a, 16:0i, 17:0i, 17:0cy), fungi (18:2ω6,9,
18:1ω9), and microeucaryotes (22:0, 24:0, 25:0; Zelles 1999; Treonis et al. 2004;
Bezemer et al. 2006) were determined.
Community level physiological profile (CLPP) of soils
To determine CLPPs of the soils the MicroResp™ methodology was used (Campbell et
al. 2003; Lalor et al. 2007). CLPP measures the functional diversity and activity of soil
microbes involved in the carbon cycle and is usually assessed by carbon substrate utilization.
The C subtrates used during the experiment were selected based on their known
occurrence in jarrah forest ecosystems, as substances produced during the thermal
oxidation of organic matter, or known plant root exudates (Banning et al. 2012). Types
of C substrates, as well as concentrations of substrates and reagents in cresol red
detector plates were the same used by Lalor et al. (2007; see Appendix 1). After a 6
hour incubation period at 25 °C, absorbance of detector plates (at 590 nm) was
determined using a microplate reader (ASYS Hitech, EXPERT 96) and headspace
%CO2 was calculated with the calibration curve equation experimentally obtained
utilizing the plate reader mentioned above: 0.6(exp(-9.95(x-0.42))+0.06. Finally the
CO2 rate (μg CO2–C g-1 soil h-1) was calculated as follows:
h
dwtsoilsoilfwt
Tvol
100%
)273(273
4412
2244
100CO2%
WPgPsoilgg
ISTD
StdFAMEacidfattyindividual
1

33
Where %CO2 is the absorbance value interpolated in the calibration curve; 44, 22, 12
and 273 are constants; T is the incubation temperature (°C); vol is the headspace volume
in the well (μl); soilfwt is the fresh weight of soil per well (g); h is the incubation time in
hours and soil%dwt is soil sample %dry weight.
Nematode trophic community
Nematodes are found in most marine, freshwater and terrestrial ecosystems. In soils, nematodes
are represented within several trophic levels of the soil food web (Yeates et al. 2009).
Nematodes can be directly influenced by plants (i.e. plant parasites), or indirectly by microbes
associated to roots (i.e. bacterial and fungal-feeders). As a result, plant conditioning is likely to
be reflected in nematode trophic community. Nematodes were extracted from 100 g of soil
(per sample) using a mistifier, where a fine mist of water at 25 ˚C is sprayed over the
soil sample for ten seconds at ten-minute intervals over a four-day period (Hooper et al.
2005). Aqueous extracts were collected and nematodes within them were sorted per1
mL extract using a counting cell (Sedgewick Rafter cell, Graticules England) observed
at 10x magnification in a compound microscope (Olympus Vanox). Nematodes of the
following trophic groups were identified and counted: bacterial feeders (BF; Rhabditida,
Enoplida), fungal feeders (FF; Tylenchida), omnivorous (Dorylaimida), predators
(Mononchida) and plant parasites (Tylenchida, Dorylaimida). Groups were assigned
based on morphological characteristics of the cephalic region, and esophageal structures
(Yeates et al. 1993). Data were expressed as the number of nematodes of each group per
100 g of dry soil. Additionally, the nematode channel ratio (FF/FF+BF) was calculated
as an indicator of the decomposition channel (bacterial or fungal) working in soils
(Yeates 2003).

34
CHAPTER 3
Soil conditioning and plant-soil feedbacks associated with jarrah forest plants
are soil-context dependent
INTRODUCTION
There is increasing interest in the restoration of ecosystems where soil has been
modified by anthropogenic activities (Harris 2009). Understanding how plant-soil
feedback (PSF) operates in soil can be a critical aspect of ecosystem restoration (van der
Putten et al. 2013). Therefore, it is timely to consider whether PSF operating in human-
modified soils function in the same way as in non-modified soils. An important
component of PSF is the conditioning of soil by plants, which is defined as changes of
soil properties that are driven in a plant species-specific manner (Harrison & Bardgett
2010; Pernilla et al. 2010; van de Voorde et al. 2011). Soil conditioning is driven by
root and litter deposition (Kulmatiski et al. 2008). Rhizodeposition and exudates can
modify the physical, chemical and biological properties of soil (Marschner 1995;
Neumann & Römheld 1999; Uren 2001; Bertin et al. 2003; Neumann 2007; Pang et al.
2010), and vary with plant species, cultivars, and plant developmental stage. As well,
rhizodeposition and exudates vary with environmental stress such as mechanical
impedance, anaerobiosis, drought, presence of pollutants, and nutrient deficiency
(Marschner 1995; Uren 2001; Nautiyal et al. 2008; Martin et al. 2014). Conditioning
might play a fundamental role in the restoration of ecosystems that have been modified
by anthropogenic activities because plant species that are capable of conditioning
modified soils may increase their chances of establishment and growth (Hobbie 1992;
Eviner 2004; Jordan et al. 2008).

35
In this chapter, I investigate the influence of plant species and soil context (i.e. unmined
and restored) on individual PSF associated with representative plant species of the
jarrah forest (i.e. Eucalyptus marginata, Acacia pulchella, and Bossiaea ornata). Native
plants have developed a variety of nutrient-acquisition strategies to allow them to
establish and persist in the impoverished soils present in the jarrah forest (Lambers et al.
2007; Hopper 2009; Sardans & Peñuelas 2013). This suggests the existence of strong
links between plants and soil, and the capability of native plants of conditioning the
physical, chemical and biological properties of soils (Abbott & Robson 1977;
Malajczuk & McComb 1977; Malajczuk 1979; Malajczuk & McComb 1979; Grierson
& Adams 2000; Lambers et al. 2007; Bleby et al. 2009; Hopper 2009).
Mining for bauxite occurs in the jarrah forest. After a mine closes, jarrah forest is
restored (Koch 2007b), however the properties of soils under restored forest (hereafter
restored soils) are altered compared with those of unmined soils. Specifically, restored
soils tend to have reduced biological activity, pH, and C, and increased P, N, K and
gravel percentage (Ward 2000; Jasper 2007; George et al. 2010; Tibbett 2010). It is
possible that soil conditioning by native plants is modified in restored soils compared
with unmined soils, which may ultimately affect the patterns of biomass production of
the restored plant community.
In the jarrah forest, plant community succession follows an initial floristic composition
model (Cattelino et al. 1979; Grant 2006). This model describes how the initial plant
community evolves only as changes in the abundance of the species present in the
landscape (Hobbs & Legg 1983; Norman et al. 2006) in response to disturbances such
as fire (Bell & Koch 1980). Therefore, it is likely that jarrah forest plants species
condition soil differently so as to secure their persistence in the community. This could
happen because the species that establish early in succession are those that are still

36
present later. The aims of this work were: 1) to examine in a detailed manner the soil
conditioning process of representative native jarrah forest plants species, and individual
PSF (i.e. magnitude and direction) associated with these plant species. 2) To determine
how conditioning process and individual PSF associated with the same plant species
differed in restored soil post-bauxite mining as compared with unmined soil.
The plant species used for this experiment were Eucalyptus marginata (i.e. long-lived
lignotuber-regenerating species), Acacia pulchella (i.e. short-lived, fire-induced species,
and Bossiaea ornata (i.e. long-lived, resprouter species). Specifically, it was
hypothesised that E. marginata, A. pulchella, and B. ornata would condition the
chemical and biological properties of unmined soil in a species-specific manner. The
second hypothesis was that modification of soils by bauxite mining and restoration
practice would differentially influence the patterns of conditioning of the plant species,
because conditioning process would differ in restored soil as compared to unmined soil.
The third hypothesis was that biomass production of jarrah forest plants and
consequently individual PSF would be influenced by plant species and soil conditions.
Positive PSF would be associated with long-lived B. ornata grown in unmined soil,
whereas negative PSF would be associated with short-lived A. pulchella in the same soil
type. However, individual PSF associated with the plant species might change in
magnitude and direction when grown in restored, compared with unmined soil.
MATERIALS AND METHODS
Sampling and experimental design
To test the hypotheses a pot experiment with two phases of plant growth (Figure 1) was
designed. Two different soil types were used, soil collected from jarrah forest with no
history of mining activity (i.e. unmined soils; 116°5' E 32°36' S) and a restored soil
collected from a site that had been cleared, mined and then restored to jarrah forest by

37
Alcoa World Alumina Australia in 2008 (i.e. restored soils; 153°21' E 27°13' S) using
practices described by Koch (2007b). The soils were collected in November 2009. Both
unmined and restored soils were gravelly sands with the following characteristics:
unmined soil presented a density of 1.05 g cm-3, 36.92% of gravel, soil fraction < 2 mm
was 89.9% sand, 5.62% silt, 5.09% clay, 0.08% and 3.05% of total N and total C
respectively, 60 and 4 mg kg-1 of available K and P respectively, pH (CaCl2) of 5.1 and
EC of 34.67 dS m-1. Restored soil presented a density of 1.3 g cm-3, soil fraction < 2
mm was 60% of gravel, 88.39% sand, 5.01% silt, 6.6% clay, 0.07% and 3.15% of total
N and total C respectively, 35 and 22 mg kg-1 of available K and P respectively, pH
(CaCl2) of 4.88 and EC of 23.3 dS m-1.
All soils were collected adjacent to roots to a depth of 15 cm (0.8 – 1.0 kg per plant).
Restored soils were sampled from beneath the plant species E. marginata (Myrtaceae),
A. pulchella (Fabaceae) and B. ornata (Fabaceae), whereas unmined soil was sampled
from the species mentioned above plus the legumes Acacia lateviticola, Gompholobium
polymorphum Bossiaea aquifolium, Labichea punctata, Hovea trisperma and Hovea
chorizemifolia since it was not possible to locate enough specimens of the three species
that were sampled at the restored site. Approximately 80 plants were sampled at each
site. All samples of unmined soils were thoroughly mixed and distributed into 48
bottom-sealed pots (1.2 L; diameter = 13.3 cm, height = 12.4 cm). This procedure was
repeated using restored soils. There were 96 pots in total and these were harvested in
two batches of 48 pots each after phases one and two respectively as described below.
The plant species grown in phase one were E. marginata A. pulchella and B. ornata.
These species were selected based on their characteristics and previous observation of
their growth behaviour in the field. Eucalyptus marginata (jarrah) is a lignotuber-
regenerating, long-lived tree (i.e. 300 years) that dominates the canopy of the forest

38
(Abbott 1984). Acacia pulchella is a fire-induced species whose population peaks after
forest burning but declines four years later (Monk et al. 1981). Bossiaea ornata is a
resprouter, long-lived bush (Bell 2001). Eucalyptus marginata is common in both
restored and unmined soils (Bell 2001), A. pulchella can be abundant in newly restored
sites but not as common in unmined jarrah forest (Monk et al. 1981), and B. ornata can
be common in both unmined and early restored mined sites but is not as prolific under
fertilisation practice typical in restoration (Bell & Heddle 1989, George et al. 2006).
The glasshouse experiment ran from November 2009 to August 2010. The 48 pots per
soil type were planted with seedlings as follows: 18 with E. marginata, 12 with A.
pulchella, 12 with B. ornata and 6 were left unplanted as controls. One seedling was
planted into each pot and dead seedlings were replaced. Within two weeks all pots had a
growing plant. To produce seedlings, seeds of A. pulchella and B. ornata were boiled
for 30 seconds to induce germination. Afterward, seeds of the three species were
surface-disinfected with ethanol (70%) and bleach (50%) to prevent pathogen growth
and germinated at 14 °C in darkness.
After phase one (4.5 months) plants from the first set of 48 pots were harvested (six
replicates × two soil types (unmined and restored soils) × three plant species (E.
marginata, A. pulchella and B. ornata) + unplanted controls) and potted soils were
sampled. After the growing phase the root system of the plants had spread throughout
the soil within the pots, and so it was assumed that a considerable amount of soil was to
some extent under the influence of roots, and thus all the potted soil was used for the
analyses. Soil from each pot was sieved (mesh = 2 mm) and stored at 4 °C pending
analyses. A soil subsample from each pot was frozen (-20 °C) and then subsequently
freeze-dried pending phospholipid-fatty acid analysis (PLFA).

39
The remaining set of 48 pots was used for phase two, shoots from plants growing in this
set of pots were removed leaving the soil intact and new seedlings of A. pulchella and
B. ornata were planted in soil previously conditioned by the same plant (home soil) or
in soil previously conditioned by E. marginata (away soil). Soil conditioned by E.
marginata was selected as ‘away soil’ because this is the dominant tree in the jarrah
forest, either in unmined or restored forest, therefore it is likely that a considerable
amount of soil in the forest has been influenced (conditioned) by this species.
Consequently, in the field other native species might grow in soil that was influenced by
jarrah at some stage. The design for phase two included six replicates × two soil types
(unmined and restored soils) × two plant species (A. pulchella and B. ornata) × two
conditioning types (home and away soils; Figure 1). Phase two ran for 4.5 months, after
which the plants were harvested; shoots and roots were oven-dried for one week at 60
°C and weighed.
Throughout the experiment pots were kept in two root cooling tanks filled with water
set to a constant temperature of 20 °C in order to maintain the roots and soil under
homogeneous conditions. Root cooling tanks were housed in a glasshouse for the
duration of the experiment. Temperatures inside the glasshouse ranged between 13 and
31 °C during phase one, and between 6 and 26 °C during phase two. Pots were rotated
between and within tanks every two weeks to negate a block effect and watered once a
week to 60% water-holding capacity.

40
Figure 1. The experiment had two phases (4.5 months each): phase one where the plants Eucalyptus marginata, Acacia pulchella, and Bossiaea ornata were planted into pots containing either unmined soils or soils from beneath jarrah forest restored after bauxite mining; and, phase two where only A. pulchella and B. ornata were planted in pots previously conditioned by the same plants (home soil) or by E. marginata (away soil). Unplanted pots were included as controls in phase one. After phase one soil properties were measured to estimate the effects of soil conditioning by plants (A). Plant biomass in home and away soils was measured after phase two to determine the magnitude and direction of individual PSF, in both unmined soils and restored soils (B).
Response variables
Refer to chapter two for methodologies used to determine response variables measured.
Statistical analysis
As evidence of soil conditioning (Figure 1A) differences in the properties of soils grown
with the experimental plants with respect to the properties of the unplanted controls
were analysed. To do this, permutational analyses of variance (PERMANOVA) of the
sets of chemical and biological properties, and of individual properties were performed.
Although PERMANOVA is a multivariate method, it can be used for univariate
analyses; to perform PERMANOVA using one response variable and Euclidean
distance yields Fisher’s traditional univariate F-statistic (Anderson 2010). The analyses
were performed independently for unmined and restored soils.

41
As well, differences in the patterns of conditioning of soil properties among the
experimental plants were determined. To do this we used PERMANOVA followed by
pair-wise tests on the chemical and biological soil properties. The total set of PLFAs
detected, the respiration rates of C-substrates from CLPPs and the total trophic groups
of nematodes were independently analysed using PERMANOVA. To visualize the
results of PERMANOVAs, scaling plots were built using principal co-ordinate analysis
(PCO; Anderson 2001). To build PCOs, variables were transformed when necessary and
normalized, and Euclidean distances were used to calculate the resemblance matrices.
Additionally, a vector showing the properties causing the major effects on the
separation of clusters along the main PCOs (i.e. PCO1 and PCO2) was included in each
plot. The vector shows the properties whose Pearson correlation coefficient with the
main PCOs was > 0.8.
To determine individual PSF (i.e. magnitude and direction; Figure 1B), univariate
PERMANOVA with plant biomass as the response variable was used. A significant
increase of the plant biomass production in home with respect to away soil was
considered a positive PSF. A significant decrease of the plant biomass production in
home soil with respect to away soil was considered a negative PSF. No difference in
biomass production in home with respect to away soil was considered a neutral PSF.
For all the statistical analyses PRIMER v6 +PERMANOVA (PRIMER-E Ltd) was
used. For the multivariate analyses, permutation of residuals under a reduced model
was selected as permutation method as well as type III (partial) sum of squares. The
unrestricted permutation of raw data as permutation method and type III (partial) sum
of squares were used for the univariate analyses. In all cases data were transformed
when necessary and Euclidean distance was used to build resemblance matrices, except
for the analyses of the trophic groups of the nematode community and CLPPs when

42
Bray-Curtis coefficient and Manhattan distance were respectively utilized to build the
similarity matrices.
RESULTS
Changes in properties of soils conditioned during phase one
Unmined soil
The chemical properties of unmined soils were conditioned by E. marginata, A.
pulchella, and B. ornata compared with the chemical properties of the unplanted
controls (t = 2.94, P = 0.002; t = 1.79, P = 0.015; t = 3.86, P = 0.003 for E. marginata,
A. pulchella, and B. ornata, respectively; Figure 2a). Also, during phase one the
chemical properties of soils grown with A. pulchella differed from the chemical
properties of soils conditioned by E. marginata (t = 2.81, P = 0.002) and B. ornata (t =
3.57, P = 0.003; Figure 2a). Eucalyptus marginata and B. ornata similarly changed the
chemical properties of soils (i.e. chemical soil properties were not significantly
different; Figure 2a). Statistical analyses of individual properties showed that E.
marginata modified the C:N ratio (t = 3.56, P = 0.002), available P (t = 17.81, P =
0.005), EC (t = 3.63, P = 0.005), pH (t = 9.48, P = 0.004) and total C (t = 4.36, P =
0.006), these properties differed from those of controls (Table 1). Acacia pulchella
altered the C:N ratio (t = 3.29, P = 0.011) and total N (t = 3.41, P = 0.022), whereas B.
ornata changed the C:N ratio (t = 4.45, P = 0.004), available P (t = 19.4, P = 0.001), EC
(t = 4.5, P = 0.009), pH (t = 9.48, P = 0.004), and total C (t = 5.05, P = 0.003; Table 1).
Significant decreases in concentrations of NH4+ in soils grown with E. marginata and B.
ornata were interpreted as an effect of plant uptake (Table 1).
The biological properties of soils planted with E. marginata, A. pulchella, and B. ornata
did not differ from the biological properties of the unplanted controls, thus there was no
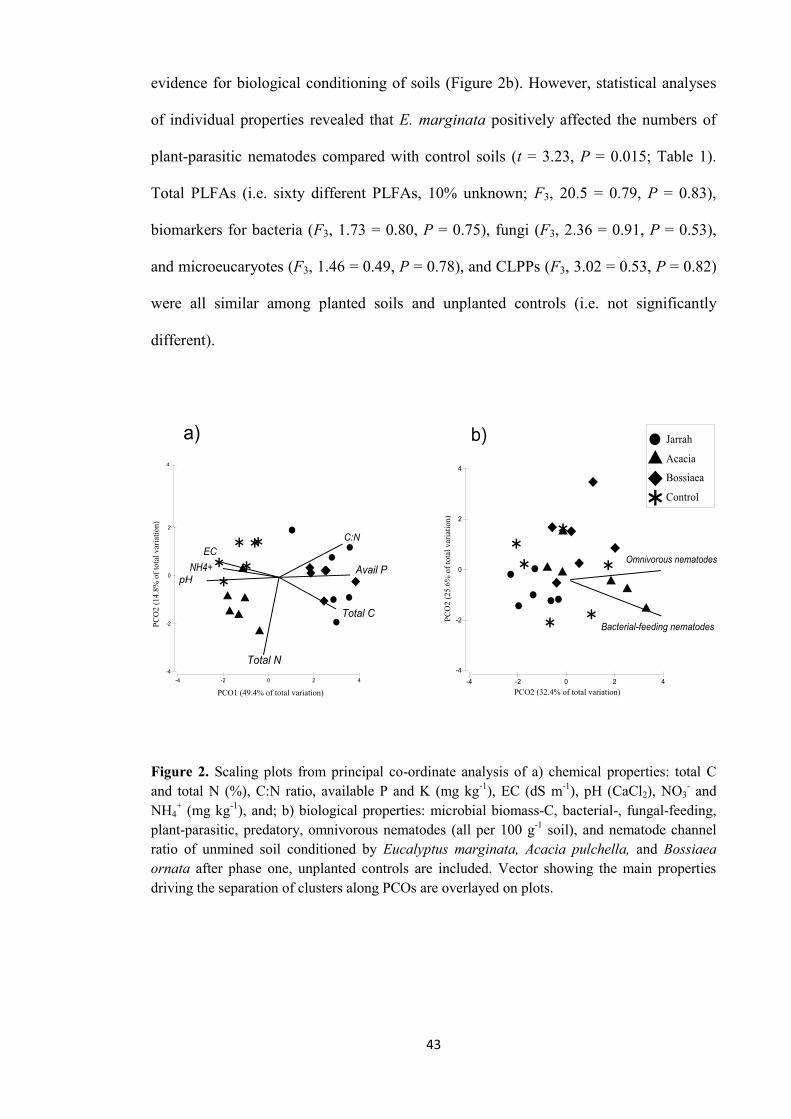
43
evidence for biological conditioning of soils (Figure 2b). However, statistical analyses
of individual properties revealed that E. marginata positively affected the numbers of
plant-parasitic nematodes compared with control soils (t = 3.23, P = 0.015; Table 1).
Total PLFAs (i.e. sixty different PLFAs, 10% unknown; F3, 20.5 = 0.79, P = 0.83),
biomarkers for bacteria (F3, 1.73 = 0.80, P = 0.75), fungi (F3, 2.36 = 0.91, P = 0.53),
and microeucaryotes (F3, 1.46 = 0.49, P = 0.78), and CLPPs (F3, 3.02 = 0.53, P = 0.82)
were all similar among planted soils and unplanted controls (i.e. not significantly
different).
Figure 2. Scaling plots from principal co-ordinate analysis of a) chemical properties: total C and total N (%), C:N ratio, available P and K (mg kg-1), EC (dS m-1), pH (CaCl2), NO3
- and NH4
+ (mg kg-1), and; b) biological properties: microbial biomass-C, bacterial-, fungal-feeding, plant-parasitic, predatory, omnivorous nematodes (all per 100 g-1 soil), and nematode channel ratio of unmined soil conditioned by Eucalyptus marginata, Acacia pulchella, and Bossiaea ornata after phase one, unplanted controls are included. Vector showing the main properties driving the separation of clusters along PCOs are overlayed on plots.

44
Table 1. Values for properties of unmined soils (mean ± SE; n = 6) after conditioning by three plant species (Eucalyptus marginata, Acacia pulchella, and Bossiaea ornata) compared with the unconditioned control soils. Only the properties that differed in conditioned soils with respect to controls are shown in the table. Values in bold font indicate significant differences between conditioned and unconditioned soils (P ≤ 0.02)
Property A. pulchella E. marginata B. ornata Control
C:N 28.99 (1) 41.15 (2.2) 39.92 (1.3) 33.23 (0.85)
Available P (mg kg-1) 4.00 (0.26) 15.67 (0.61) 16.17 (0.54) 3.67 (0.33)
EC (dS m-1) 0.035 (0.002) 0.03 (0.003) 0.03 (0.002) 0.04 (0.002)
pH 5.43 (0.02) 5.15 (0.02) 5.15 (0.02) 5.45 (0.02)
Total N (%) 0.10 (0.004) 0.08 (0.005) 0.08 (0.002) 0.08 (0.003)
Total C (%) 2.76 (0.18) 3.25 (0.14) 3.26 (0.11) 2.54 (0.09)
NH4+ (mg kg-1) 5.52 (1.14) 3.57 (1.38) 3.75 (0.88) 4.99 (0.84)
Plant-parasitic nematodes (per 100 g-1 soil)
0 39 (12) 0 4 (3)
Restored soil
The chemical properties of restored soil were modified by E. marginata and B. ornata
only, and differed from the chemical properties of controls (t = 2.63, P = 0.002; t = 3.04,
P = 0.002 for E. marginata and B. ornata respectively; Figure 3a). In contrast, the
chemical properties of soil grown with A. pulchella did not differ from controls (t =
1.36, P = 0.089). Also, the chemical properties of soil grown with the three plant
species were significantly different to each other indicating a level of interspecific
conditioning (E. marginata versus A. pulchella t = 2.85, P = 0.002; E. marginata versus
B. ornata t = 2.22, P = 0.0017; A. pulchella versus B. ornata t = 3.53, P = 0.0027;
Figure 3a). Statistical analyses of individual properties showed that A. pulchella did not
change any chemical property significantly. Eucalyptus marginata modified available P
(t = 22.6, P = 0.002), and pH (t = 6.99, P = 0.004), these properties differed from those

45
of controls. Bossiaea ornata altered available P (t = 28.36, P = 0.004), and pH (t = 12.8,
P = 0.002; Table 2). Decreased concentrations of available K in soil conditioned by B.
ornata were considered a plant uptake effect (Table 2).
The biological properties of restored soil grown with each of the three plant species did
not differ from the biological properties of controls. Also, the biological properties of
soils grown with the three plant species were all similar to each other (i.e. no significant
differences; Figure 3b). However, analyses of individual properties indicated that E.
marginata positively affected the number of fungal-feeding nematodes (t = 2.97, P =
0.01) compared with the unplanted controls (Table 2). Analyses of the total PLFAs (F3,
23.2 = 0.94, P = 0.54), biomarkers for bacteria (F3, 5.15 = 0.82, P = 0.75), fungi (F3, 2.35 =
1.28, P = 0.26), and microeucaryotes (F3, 2.78 = 1.39, P = 0.24) detected in the soils
revealed no statistical differences between planted soils and unplanted controls. The
same results were found for CLPPs that were also similar among planted soils and
controls (F3, 2.75 = 0.44, P = 0.89).
Individual PSF
Individual PSF associated with the two plant species differed in both magnitude and
direction according to soil type. In unmined soils, A. pulchella and B. ornata produced
similar amounts of biomass in home and away soils (F1, 0.01 = 2.66, P = 0.139 and F1, 0.04
= 0.43, P = 0.535 for A. pulchella and B. ornata respectively; Figure 4), thus a neutral
PSF was observed in both cases. In restored soils, both A. pulchella and B. ornata
produced less biomass in home compared with away soils (F1, 1.4 = 13.314, P = 0.004;
F1, 5.65 = 10.1, P = 0.01 for A. pulchella and B. ornata respectively; Figure 4) so a
negative PSF was observed. Also, A. pulchella produced up to three times more
biomass in restored soil than that produced in unmined soil.
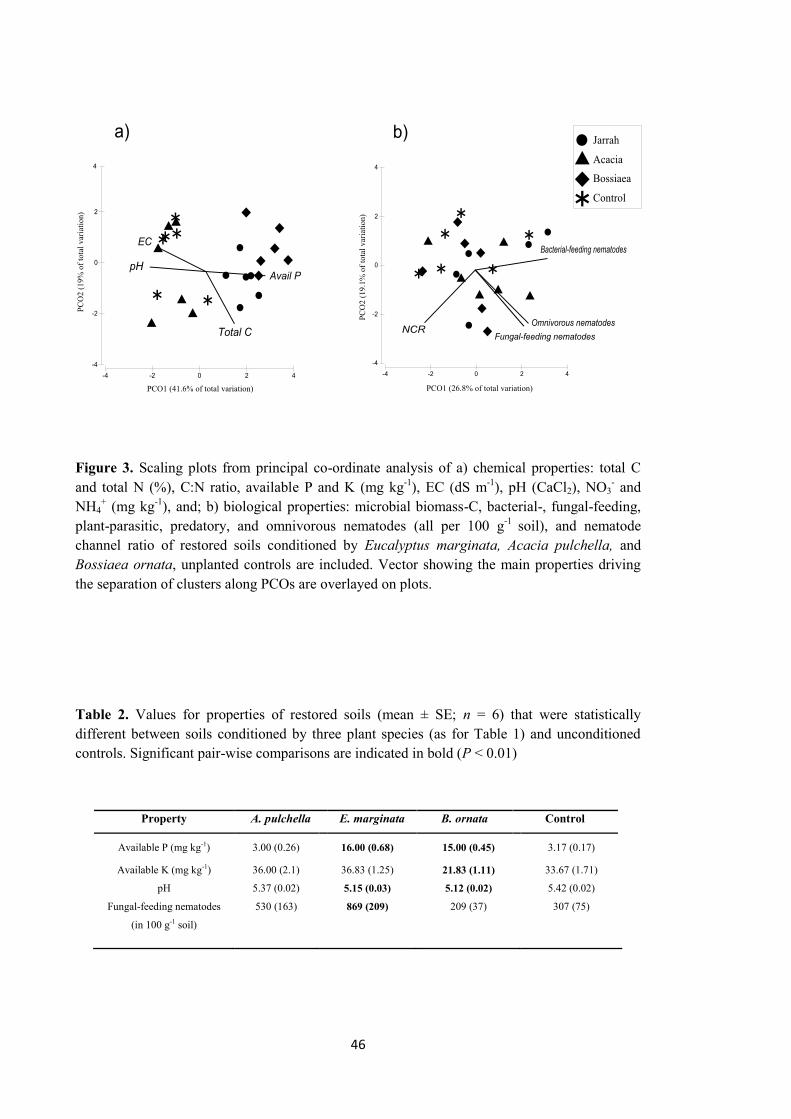
46
Figure 3. Scaling plots from principal co-ordinate analysis of a) chemical properties: total C and total N (%), C:N ratio, available P and K (mg kg-1), EC (dS m-1), pH (CaCl2), NO3
- and NH4
+ (mg kg-1), and; b) biological properties: microbial biomass-C, bacterial-, fungal-feeding, plant-parasitic, predatory, and omnivorous nematodes (all per 100 g-1 soil), and nematode channel ratio of restored soils conditioned by Eucalyptus marginata, Acacia pulchella, and Bossiaea ornata, unplanted controls are included. Vector showing the main properties driving the separation of clusters along PCOs are overlayed on plots.
Table 2. Values for properties of restored soils (mean ± SE; n = 6) that were statistically different between soils conditioned by three plant species (as for Table 1) and unconditioned controls. Significant pair-wise comparisons are indicated in bold (P < 0.01)
Property A. pulchella E. marginata B. ornata Control
Available P (mg kg-1) 3.00 (0.26) 16.00 (0.68) 15.00 (0.45) 3.17 (0.17)
Available K (mg kg-1) 36.00 (2.1) 36.83 (1.25) 21.83 (1.11) 33.67 (1.71)
pH 5.37 (0.02) 5.15 (0.03) 5.12 (0.02) 5.42 (0.02)
Fungal-feeding nematodes
(in 100 g-1 soil)
530 (163) 869 (209) 209 (37) 307 (75)
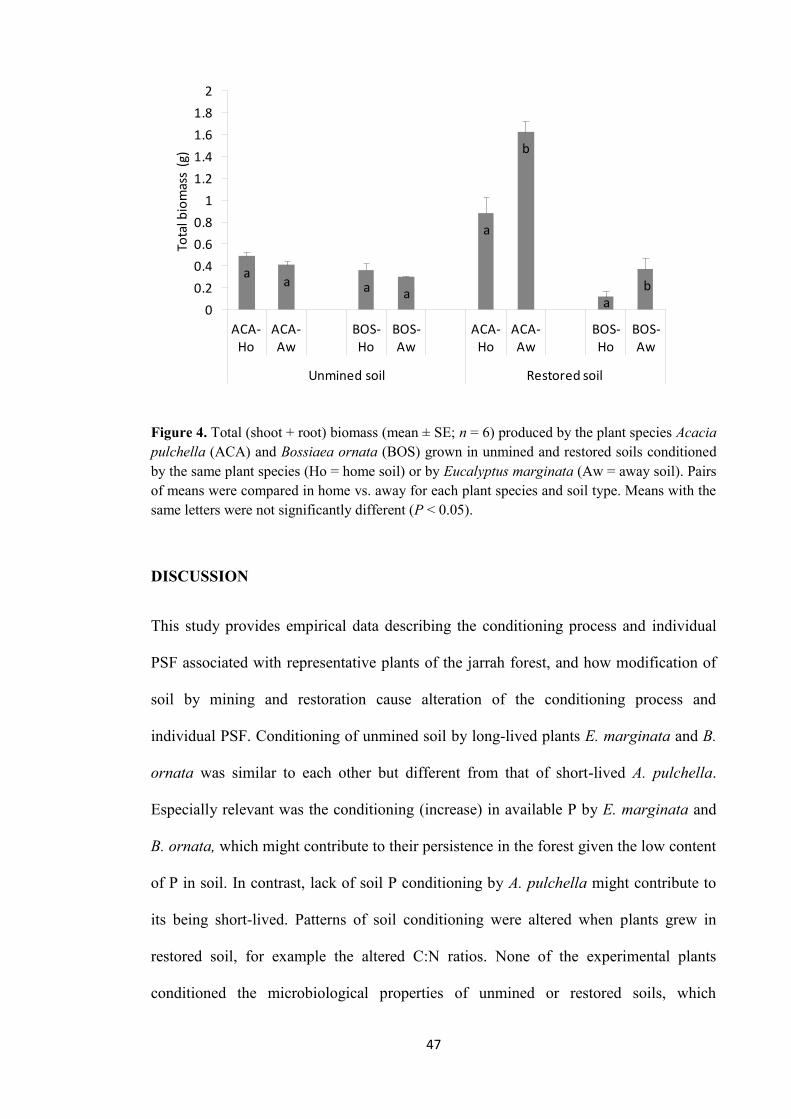
47
Figure 4. Total (shoot + root) biomass (mean ± SE; n = 6) produced by the plant species Acacia pulchella (ACA) and Bossiaea ornata (BOS) grown in unmined and restored soils conditioned by the same plant species (Ho = home soil) or by Eucalyptus marginata (Aw = away soil). Pairs of means were compared in home vs. away for each plant species and soil type. Means with the same letters were not significantly different (P < 0.05).
DISCUSSION
This study provides empirical data describing the conditioning process and individual
PSF associated with representative plants of the jarrah forest, and how modification of
soil by mining and restoration cause alteration of the conditioning process and
individual PSF. Conditioning of unmined soil by long-lived plants E. marginata and B.
ornata was similar to each other but different from that of short-lived A. pulchella.
Especially relevant was the conditioning (increase) in available P by E. marginata and
B. ornata, which might contribute to their persistence in the forest given the low content
of P in soil. In contrast, lack of soil P conditioning by A. pulchella might contribute to
its being short-lived. Patterns of soil conditioning were altered when plants grew in
restored soil, for example the altered C:N ratios. None of the experimental plants
conditioned the microbiological properties of unmined or restored soils, which
aa a a
a
b
ab
0
0.2
0.4
0.6
0.8
1
1.2
1.4
1.6
1.8
2
ACA-Ho
ACA-Aw
BOS-Ho
BOS-Aw
ACA-Ho
ACA-Aw
BOS-Ho
BOS-Aw
Unmined soil Restored soil
Tota
l bio
mas
s (g
)

48
contradicts the general finding of positive conditioning of microbial communities in the
root affected soil. Other groups of the soil biota such as the nematodes were little
affected by plant conditioning. The results indicated that individual PSF associated with
both, A. pulchella and B. ornata was affected by soil context, a neutral PSF was
observed in restored soils and a negative PSF was produced in unmined soil.
It was predicted that chemical as well as biological properties of unmined soil would be
differentially conditioned by the three plant species, however in unmined soil, species-
specific conditioning primarily occurred through changes in the chemical properties.
Eucalyptus marginata and B. ornata, both long-lived species similarly conditioned the
chemical properties of unmined soils, which differed from conditioning by short-lived
A. pulchella. The total C concentration of soil increased under the influence of the first
two plant species, likely as a result of root deposition (Marschner 1995; Brimecombe et
al. 2001; Neumann & Römheld 2001). Acacia pulchella did not affect the total C
concentration of soil, instead this species promoted an increase in the total N
concentration, which can be attributed to exudation of N-compounds such as amino
acids (Uren 2001; Bertin et al. 2003). As a result, the C:N ratios of conditioned
(unmined) soils differed from those of the unconditioned controls. Eucalyptus
marginata and B. ornata caused an increase in the C:N ratio, whereas A. pulchella
caused a decrease in the C:N ratio. The C:N ratio controls microbial-mediated soil
processes such as the decomposition of the organic matter (Adl 2003; Lavelle & Spain
2003; Coleman et al. 2004). According to the results, A. pulchella might be enhancing a
faster turnover of organic matter (i.e. by decreasing the C:N ratio), whereas E.
marginata and B. ornata may cause slower decomposition of organic matter (i.e. by
increasing the C:N ratio). Thus, the species-specific effects on C:N ratios may, in turn,
cause additional impacts on soil processes in the longer-term.

49
Root exudates play an important role in P mobilization in these P deficient (unmined)
soils and specifically, the organic acids that are released by roots to solubilise mineral
bound P, which is often accompanied by a decrease in soil pH (Jones 1998; Neumann &
Römheld 1999; Dakora & Phillips 2002; Carvalhais et al. 2011). In this study, E.
marginata and B. ornata plants appeared to actively mobilize P in unmined soil, with a
four-fold increase in the available P fraction as a result of soil conditioning by these two
species. Mobilization was accompanied by a decrease in soil pH, which suggests that
carboxylic acids or acid phosphatase enzymes could be playing an important role in this
process. Unmined soil conditioned by A. pulchella presented available P concentrations
and pH that did not differ from those of controls. Therefore mobilisation of P by this
plant might not be a critical component of the conditioning process as it appeared to be
for the other two plant species. Differential conditioning of P that is the main limiting
soil mineral, might contribute to explain the persistence of the three plant species in the
forest. Eucalyptus marginata and B. ornata are long-lived species (Abbott 1984; Bell
2001), which can be attributed to their efficient capacity to condition P. Acacia
pulchella is a short-lived species (Monk et al. 1981), a trait that could be explained by
its poor capacity to condition soil P.
There was limited evidence of conditioning of soil biological properties by the plant
species. Conditioning of the microbial community (i.e. bacteria, fungi) was not
observed, and these results contradicted the general finding of positive conditioning of
soil microbes that is mediated by root exudation (Reynolds et al. 2003; Kardol et al.
2007; Bever et al. 2010; Bever et al. 2012; Miki 2012). It is likely that plant species
used in the experiment require longer time spans to condition their own microbial
community, as previously reported for other plant species (Kulmatiski & Beard 2011).
Of the biological properties that I measured, plant-parasitic nematodes was the only
property of unmined soil to be conditioned, and just by one plant species, E. marginata.

50
Interactions between plant-parasitic nematodes and their host plants are mediated by
root exudates (Hirsch et al. 2003; Zavaleta-Mejía & Kaloshian 2012). Therefore, it is
likely E. marginata produced chemical signals to attract plant-parasitic nematodes to
the rhizosphere. Plant parasites and pathogens are agents of negative PSF, which
contribute to shaping the structure of plant communities during succession (Mills &
Bever 1998; van der Putten 2000; Bonanomi et al. 2005a; Bonanomi et al. 2005a;
Mangan et al. 2010). Information on the ecological role of nematodes in jarrah forest is
scarce, however, these organisms might contribute to counteract the dominance of
plants such as E. marginata and thus contribute towards species coexistence.
In restored soil, the patterns of conditioning of chemical properties differed from those
observed in unmined soil as predicted, but similar to unmined soils, conditioning of the
biological properties was scarcely evident. The results suggest that modified properties
initially perceived by plants induced changes to the production of rhizodeposition and
exudates, which in turn affected soil conditioning (Marschner 1995; Uren 2001;
Nautiyal et al. 2008). In contrast to the results for unmined soils, the chemical
properties of restored soils were not conditioned by A. pulchella, and chemical
properties were similarly conditioned by E. marginata and B. ornata, except by the
concentration of available K that decreased in soils grown with B. ornata, possibly due
to an increased uptake by this plant species. In contrast to unmined soils E. marginata
and B. ornata did not condition the C concentration and A. pulchella did not condition
the N concentration of restored soils. These altered patterns of conditioning are likely to
affect the C and N cycles as previously suggested by other studies (Banning et al.
2008), with important implications for the restoration of the ecosystem function.
Soil conditioning by E. marginata, A. pulchella, and B. ornata produced no change in
the C:N ratio of restored soils as compared to controls (as observed for unmined soil).

51
No change in the C:N ratios suggests that organic matter decomposition process is
differentially operating in restored soil as compared to unmined soil when these were
under the influence of the experimental plants. In contrast, conditioning of available P
was not affected by soil modification, since E. marginata and B. ornata, but not A.
pulchella, conditioned the concentration of available P of restored soil as observed in
unmined soil. This finding suggests that conditioning of P is a preserved plant trait that
might help maintaining the flux of nutrients in the ecosystem (Eviner & Hawkes 2008).
Therefore, restoration of long-lived species such as E. marginata and B. ornata might
be critical for the re-establishment of ecosystem functioning for the entire developing
forest system.
In restored soil, the numbers of fungal-feeding nematodes increased as a result of soil
conditioning by E. marginata. It is likely that rhizodeposition and exudates released to
soil by E. marginata (Malajczuk & McComb 1977) produced a positive effect on the
fungal microbial biomass. Fungi was not meaured directly but was possibly seen
indirectly in the PSF response. In restored soil, plant-parasitic nematodes were not
conditioned by E. marginata as observed in unmined soil. This result suggests that the
patterns of root exudation of the plant differed, and chemical attractants for plant-
parasitic nematodes were scarcely/ not produced. Another possibility is that plant-
parasitic nematodes were so scarce in restored soil that they were not detected during
the screening. The absence or low abundance of negative PSF agents such as plant-
parasitic nematodes could have implications on the structure and evolution of the plant
community during restoration (De Rooij-van der Goes 1995; van der Putten & Peters
1997; van der Stoel et al. 2002).
In unmined soil a neutral PSF was observed for both A. pulchella and B. ornata. These
findings contrasted with the prediction of a negative PSF associated with A. pulchella,

52
and positive PSF associated with B. ornata. The results suggested that individual PSF
was not influenced by soil properties previously conditioned by either the same (i.e.
home soil) or other plant species (i.e. away soil). In restored soil, individual PSF was
altered with respect to unmined soil, and negative PSF was observed for both A.
pulchella and B. ornata. The results indicated that individual PSF associated with the
two plant species was influenced by the modification to the soil. However, opposite to
unmined soil, in the context of restored soil, the legacy of previous plant growth (i.e. E.
marginata or B. ornata growth) seemed to influence the amounts of biomass produced
by the experimental plants (i.e. PSF that is soil-context dependent; De Deyn et al.
2004).
The notable increased biomass produced by A. pulchella in restored soil suggests a soil
feedback effect that induced a species-specific response in A. pulchella, but not in B.
ornata. Species-specific responses have been reported previously for a diversity of
couplings of soils and plants (Bret-Harte et al. 2001; Manning et al. 2008; te Beest et al.
2009; Baeten et al. 2010). These types of responses can alter the patterns of plant
abundance and dominance, and ultimately the successional trajectory of the plant
community.
In conclusion, conditioning of soil by the representative jarrah forest plants used in this
study was species-specific, the mechanism might have implications for the persistence
of plant species in the ecosystem. Modification of soil by mining and restoration
practice induced patterns of plant conditioning that differed with respect to unmined
soil. Individual PSF associated with the studied jarrah forest plants operated in a soil
context-dependent basis, as a result individual PSF observed in restored soil differed as
compared with that observed in unmined soil for both A. pulchella and B. ornata.
Alteration to conditioning process and individual PSF operating in restored soils can

53
have implications for ecosystem recovery by affecting the successional trajectory of the
plant community.

54
CHAPTER 4
Plant-soil feedback unaffected by soil condition in a post-mining
restoration chronosequence
INTRODUCTION
Ecological restoration of ecosystems that have been damaged or destroyed can result in
changes to the properties of both above and belowground compartments such that
recovery is evident over time (Ward 2000; Mummey et al. 2002; Norman et al. 2006;
Jasper 2007; Banning et al. 2008; Chodak et al. 2009). Restoration can include practices
such as ripping, seeding, planting, fertilising, and inoculation of beneficial
microorganisms to improve the physico-chemical and biological properties of soil to
accelerate successional change (Suding et al. 2004; Grant 2006; Koch 2007b; Kardol &
Wardle 2010). As a result, the concentrations of soil nutrients change due to
biogeochemical processes, C concentration increases through litter and root deposition,
the size and diversity of biotic communities increases, and the soil food chain length
grows (Schipper et al. 2001; Ferris & Matute 2003; Wardle et al. 2004; Bardgett et al.
2005; Wardle 2006; Holtkamp et al. 2008; Berg 2010; Eisenhauer & Reich 2012), all of
which ultimately contribute to ecosystem recovery. Nevertheless, there is a lack of
information as to whether the restoration of soil results in the recovery of the soil
feedback in such a way as to re-establish the plant-soil feedbacks (PSF) that are
characteristic of the undisturbed ecosystem in their magnitude and direction.
Soil feedback is the second component of PSF (after plant conditioning), and it is
important because the overall effect of physico-chemical and biological properties of
soil determines the growth, productivity, and reproductive success of individual plants
in ecosystems (van der Putten et al. 2013). In human-modified ecosystems soil

55
properties can be altered and likely their associated soil feedback, which can ultimately
modify the patterns of plant growth in the short term via PSF phenomena (Bret-Harte et
al. 2001; Baeten et al. 2010; but also see Chapter 3). In the long-term, the development
of soil properties during restoration suggests that the soil feedback and associated PSF
might experience changes over time. Empirical data indicating whether or not the soil
feedback of restored ecosystems moves toward to a soil feedback approaching that of
unmodified ecosystem are limited (Maluf & Ferreira 2004; Ribeiro et al. 2004; Shono et
al. 2005; Paul et al. 2010; Gong et al. 2013).
In the jarrah (Eucalyptus marginata) forest of south-western Western Australia, patches
of forest have been cleared for bauxite mining since 1963, and restored after mining
since 1966. However, restored soil displays modified properties such as reduced
biological activity, pH, and C, and increased P, N, K and gravel percentage (Ward 2000;
Jasper 2007; George et al. 2010; Tibbett 2010). Increased concentrations of P, N, and K
are consequence of fertiliser addition as part of the restoration process (Koch 2007b).
Increased amounts of these nutrients are expected to be significant components of the
soil feedback that in turn influence plant growth in recently-restored soil. Recently
restored jarrah forest soil also shows lower C concentration and biological activity than
native forest, but after 18 and 8 years respectively, each one of these soil properties
recovers to levels comparable to those of unmined forest soil (Jasper 2007; Banning et
al. 2008; George et al. 2010; Lin et al. 2011). Therefore, it is likely that C concentration
and biological communities of soil strongly influence plant growth via soil feedback in
the mid-, long-term, and not in the short-term. The relevance of these properties in the
soil feedback might become apparent with increasing restoration age given the
dependence of native plants on biologically-supplied nutrients such as N and P (Lawrie
1981; Brundrett & Abbott 1991; Lambers et al. 2007), and decreasing concentrations of
nutrients from fertilisers. The changing soil feedback components of restored jarrah

56
forest soil differentially influence plant biomass production (i.e. the individual PSF), as
described in the previous chapter (Chapter 3), where negative PSF was associated with
plant species grown in recently-restored soils (i.e. 1.5-year-old soils), and neutral PSF
was associated with the same plant species grown in unmined jarrah forest soils.
This investigation aimed to elucidate the influence of soil conditions on individual PSF
associated with Bossiaea ornata plants. For this purpose a glasshouse experiment was
designed to study the soil feedback produced in soils collected from a chronosequence
of restoration ages (1.5-, 7-, and 22-year-old soils) and an unmined soil that is the
benchmark for restoration. Specifically, a first aim was to determine the factors (i.e.
soil age and plant species) influencing the physico-chemical and biological properties
(i.e. the soil feedback) of soils from the chronosequence. It was predicted that properties
of soils from the chronosequence would be significantly influenced by both; age since
restoration and plant species grown on soils. Also, the properties of soil from the
chronosequence would be more similar to those of unmined soils as restoration age
increases (Jasper 2007; Banning et al. 2008; George et al. 2010; Lin et al. 2011).
There is potential for jarrah forest soils of diverse restoration age to exert differential
soil feedback on plant growth in response to variable soil properties (Ward 2000;
Banning et al. 2008; George et al. 2010; Lin et al. 2011). Therefore, the second aim of
this investigation was to examine whether or not the differential soil feedback
associated with soils from the restoration chronosequence differentially influence the
biomass produced by native B. ornata seedlings. This plant species was selected
because displays variable growth behaviour in restored as compared with unmined soils.
Bossiaea ornata is a resprouter, long-lived bush (Bell 2001) that is abundant in unmined
soils, but is not as prolific under fertilisation practice typical in restoration (Bell &
Heddle 1989; George et al. 2006). One hypothesis that could help explaining this

57
behaviour is that soil feedback exerts a strong influence on the establishment and
growth of this plant species. Therefore, variable biomass production by B. ornata was
expected in response to differential soil feedback associated to soils of variable
restoration age. Specifically, the biomass produced by B. ornata would be significantly
influenced by the effects of soil age and plant species previously grown on soils. It is
also predicted that physico-chemical properties would be significant components of the
soil feedback in recently-restored soils, whereas biological properties would be
significant components of the soil feedback in late-restored soils (Bardgett et al. 2005;
Wardle 2006; Holtkamp et al. 2008; Berg 2010; Eisenhauer & Reich 2012).
Finally, the third aim was to investigate if variable soil feedback of soils from the
chronosequence differentially influence individual PSF (i.e. its magnitude and direction)
associated with native B. ornata. A negative PSF associated with B. ornata grown in
recently-restored soil (i.e. 1.5-year-old soil) was expected, and then a transition to
neutral PSF associated to the same plant species grown in late-restored soil was
expected according to results from Chapter 3 (Kardol et al. 2006).
MATERIAL AND METHODS
Sampling and experimental design
Sites of increasing age since restoration after bauxite mining (1.5-, 7-, and 22-year-old
soils) and an unmined site that was burnt under controlled conditions in 2008 (hereafter
referred to collectively as ‘the chronosequence’) were selected within the mining lease
of Alcoa World Alumina Australia. Soils for PSF experiment were collected in June
2011. At each site, six B. ornata (Fabaceae) plants and six E. marginata (Myrtaceae)
plants were located in an area of approximately 1 km2, and sampled at random to take
two intact soil cores (0.785 L; diameter = 10 cm, depth = 10 cm) from near to the root
area of each plant. Sampled plants were some meters apart from each other. One half of

58
soil cores were used to measure the physico-chemical and biological properties of soils
to determine effects of field-based soil conditioning. Soil subsamples were sieved (2
mm-mesh), frozen (-20 °C) and then subsequently freeze-dried pending phospholipid-
fatty acid analysis (PLFA). The second half of soil cores were placed in bottom-sealed
pots (1.2 L; diameter = 13.3 cm, depth = 12.4 cm) ready for use in the pot experiment.
The experimental design included 6 replicates × four soil ages (1.5-, 7-, 22-year-old,
unmined soil) × two plant species (B. ornata or E. marginata) = 48 pots (Figure 1).
The pot experiment ran from June to October 2011. Pots containing the intact soil cores
(hereafter the field-conditioned soils) were placed into two root cooling tanks that were
filled with water set to a constant temperature of 20 °C in order to maintain the area of
contact between roots and soil under homogeneous conditions. Root cooling tanks were
housed in a glasshouse for the duration of the experiment. Temperatures inside the
glasshouse ranged between 4.5 and 27.8 °C. For the experiment seedlings of B. ornata
were used. In each pot several seedlings of B. ornata were planted, once a plant
established, the rest of seedlings were removed leaving only one growing plant per pot.
To produce the seedlings, seeds were submerged in boiling water for 30 seconds to
break dormancy, then surface-disinfected with ethanol (70%) and bleach (50%) to
prevent pathogen growth and germinated at 14 °C in darkness. Pots planted with
seedlings were rotated between and within tanks every two weeks to negate a block
effect and watered once a week to 60% water-holding capacity. After four months
plants were harvested; shoots and roots were removed, separated, oven-dried for one
week at 60 °C and weighed.

59
Figure 1. Experimental design where intact soil cores were collected from a chronosequence of restoration ages (1.5-, 7-, 22-year-old soils and unmined soils). At each site soil cores were taken from area near roots of Bossiaea ornata and Eucalyptus marginata. One half of cores were used to determine the physico-chemical and biological properties (A) to determine soil feedback components (B). The second half of the cores was placed in pots where B. ornata seedlings were planted to have B. ornata seedlings grown in soils conditioned in the field by B. ornata plants (i.e. home soils), and B. ornata seedlings grown in soils conditioned in the field by E. marginata plants (i.e. away soils). After four months all plants were harvested and plant biomass produced in each restoration age was compared in home vs. away soils (C).
Response variables
Refer to chapter two for methodologies used to determine response variables measured.
Statistical analysis
The influence of the factors soil age (with four levels; 1.5-, 7-, 22-year-old, unmined
soils) and plant species (with two levels; E. marginata and B. ornata; Figure 1A) on soil
properties was tested. To do this, a two-way permutational analysis of variance
(PERMANOVA; Anderson 2001) on the sets of physico-chemical and biological
properties of the field-conditioned soils was used. To visualize the results of the
PERMANOVA, scaling plots using principal co-ordinate analysis (PCO; Anderson
2001) were built. PCO plots showing the distances from centroids of all soil ages were
also built to determine the trajectory of multivariate change through time followed by
Properties of field-
conditioned soils
C
Intact soil cores conditioned by plant species from a chronosequence of
restoration ages
Bossiaea ornata
Eucalyptus marginata
Bossiaea ornata
grown in “home soil”
Total biomass production in
home vs. away soils
Individual Plant-soil feedback
A
Bossiaeaornata
grown in “away soil”

60
the physico-chemical and biological properties of soils from the chronosequence. The
plots allowed determining if the properties of field-conditioned soils from the
chronosequence were approaching those of unmined soils. As well, PERMANOVA
analyses were performed to determined significant effects of soil age and plant species
on total PLFAs, PLFA biomarkers for total bacteria, Gram positive bacteria, Gram
negative bacteria, fungi, and microeucaryotes, respiration rates of total C-substrates
from CLPPs, and groups of C-substrates of similar chemical nature (i.e. carboxylic
acids, aminoacids, phenolics, carbohydrates, etc. See appendix 1), and the trophic
groups of nematodes of soils within the chronosequence.
To determine differential soil feedback effect of soils from the chronosequence on
biomass produced by B. ornata seedlings (Figure 1B), two types of analyses were
performed; a univariate two-way PERMANOVA for the factors soil age (with four
levels; 1.5-, 7-, 22-year-old, unmined soils) and plant species (from which soils were
collected in the field; with two levels; E. marginata and B. ornata) on the response
variable plant biomass. Although PERMANOVA is a multivariate method, it can be
used for univariate analyses. To perform PERMANOVA using one response variable
and Euclidean distance yields Fisher’s traditional univariate F-statistic (Anderson
2010). The analysis allowed to determine significant differences in biomass produced
by B. ornata in soils from the chronosequence in response to the factors mentioned
above. The other type of analysis used was an analysis of covariance (ANCOVA) that
consisted in univariate two-way PERMANOVAs for the factors soil age and plant
species on the response variable plant biomass, but individual soil properties were
included in the model as covariables. These analyses allowed determining significant
and strong effects of the covariables on the biomass produced by B. ornata (Anderson et
al. 2008). Tests for the interactions between covariables and factors (i.e. soil age and

61
plant species) were also included in the PERMANOVA design to evidence differential
effects of the covariables across soil ages (Anderson et al. 2008).
Finally, individual PSF (i.e. magnitude and direction; Figure 1C) was determined
through univariate PERMANOVA with plant biomass as the response variable. The
effect of the factor conditioning type (with two levels; home soils - conditioned in the
field by B. ornata, and away soils – conditioned in the field by E. marginata) was
determined for each soil age. These results were interpreted as follows: significantly
increased plant biomass production in home with respect to away soils was considered
evidence of positive PSF. Significantly decreased plant biomass production in home
soils with respect to away soils was considered evidence of negative PSF. No difference
in biomass production in home with respect to away soils was considered neutral PSF.
The software PRIMER v6 +PERMANOVA (PRIMER-E Ltd) was used for all statistical
analyses. Permutation of residuals under a reduced model and type III (partial) sum of
squares were selected to perform multivariate analyses, whereas unrestricted
permutation of raw data and type III (partial) sum of squares were selected for
univariate analyses. In all cases data were transformed when necessary and normalised,
and Euclidean distance was used to build resemblance matrices, except for the analyses
of the trophic groups of the nematode community when Bray-Curtis coefficient was
utilized because individuals were counted, and for the analysis of CLPPs when the
Manhattan distance was used because it has been pointed out as an appropriate distance
for the analysis (Lalor et al. 2007; Gonzalez-Quiñones et al. 2009; Banning et al. 2012).
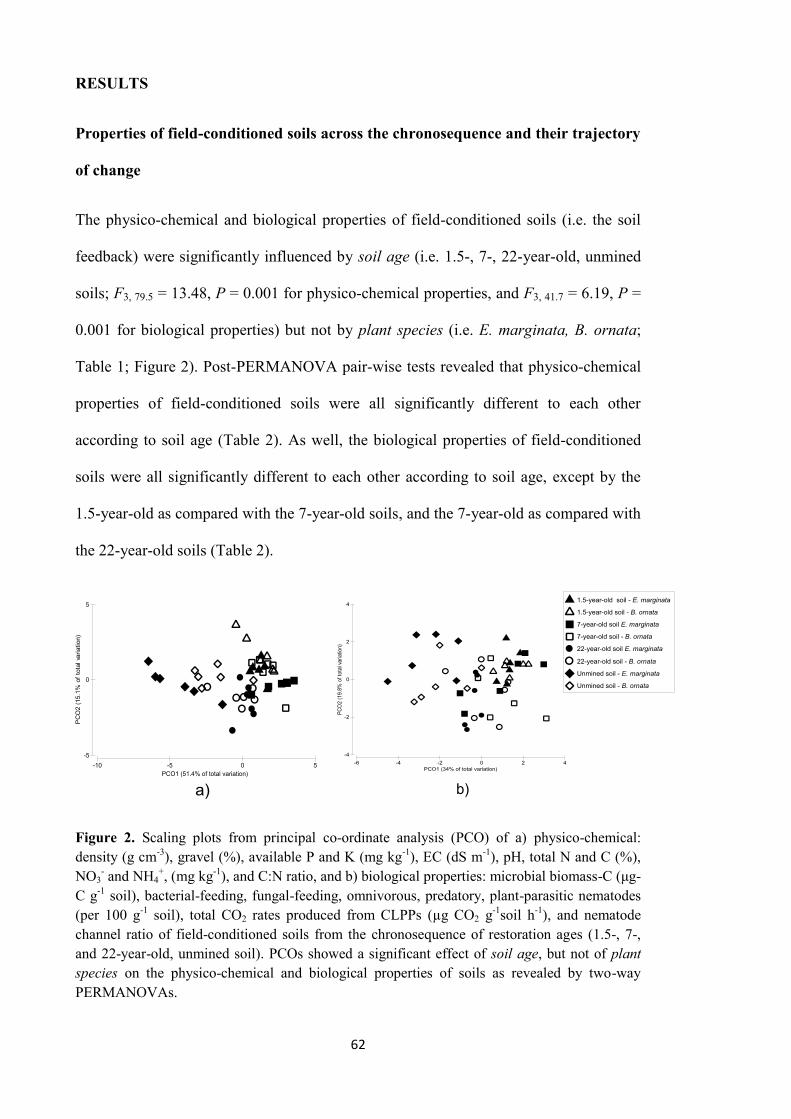
62
RESULTS
Properties of field-conditioned soils across the chronosequence and their trajectory
of change
The physico-chemical and biological properties of field-conditioned soils (i.e. the soil
feedback) were significantly influenced by soil age (i.e. 1.5-, 7-, 22-year-old, unmined
soils; F3, 79.5 = 13.48, P = 0.001 for physico-chemical properties, and F3, 41.7 = 6.19, P =
0.001 for biological properties) but not by plant species (i.e. E. marginata, B. ornata;
Table 1; Figure 2). Post-PERMANOVA pair-wise tests revealed that physico-chemical
properties of field-conditioned soils were all significantly different to each other
according to soil age (Table 2). As well, the biological properties of field-conditioned
soils were all significantly different to each other according to soil age, except by the
1.5-year-old as compared with the 7-year-old soils, and the 7-year-old as compared with
the 22-year-old soils (Table 2).
Figure 2. Scaling plots from principal co-ordinate analysis (PCO) of a) physico-chemical: density (g cm-3), gravel (%), available P and K (mg kg-1), EC (dS m-1), pH, total N and C (%), NO3
- and NH4+, (mg kg-1), and C:N ratio, and b) biological properties: microbial biomass-C (μg-
C g-1 soil), bacterial-feeding, fungal-feeding, omnivorous, predatory, plant-parasitic nematodes (per 100 g-1 soil), total CO2 rates produced from CLPPs (µg CO2 g-1soil h-1), and nematode channel ratio of field-conditioned soils from the chronosequence of restoration ages (1.5-, 7-, and 22-year-old, unmined soil). PCOs showed a significant effect of soil age, but not of plant species on the physico-chemical and biological properties of soils as revealed by two-way PERMANOVAs.
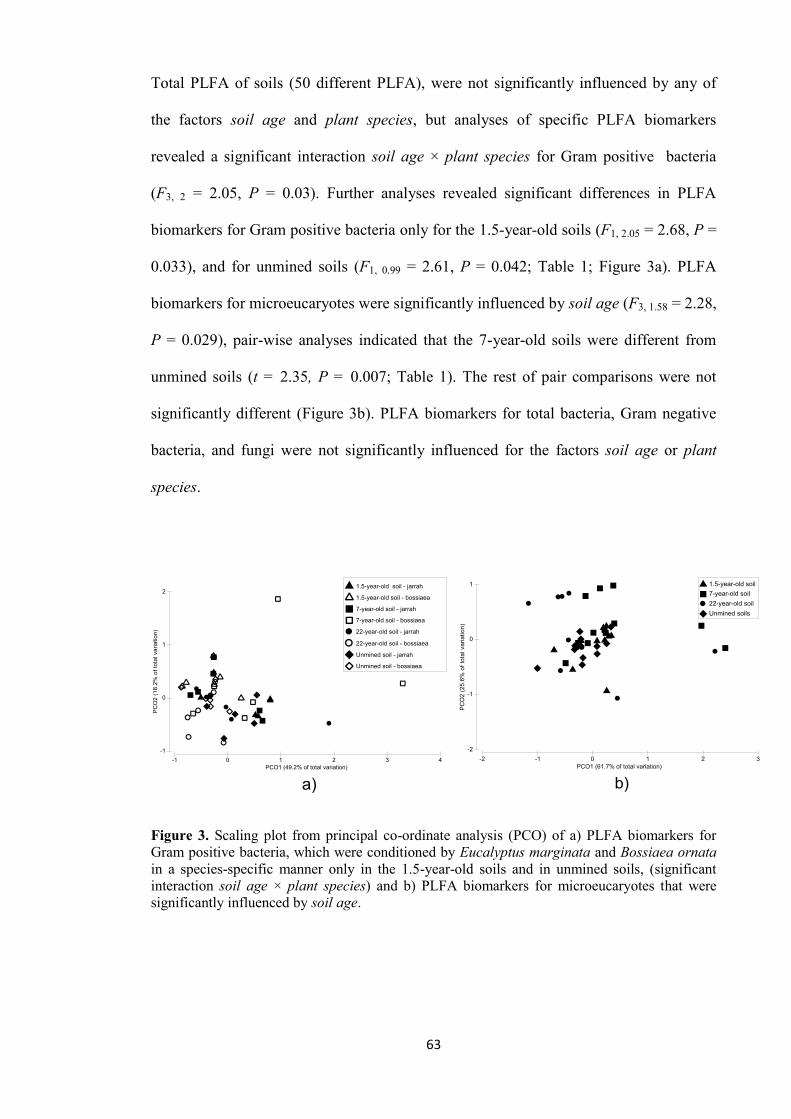
63
Total PLFA of soils (50 different PLFA), were not significantly influenced by any of
the factors soil age and plant species, but analyses of specific PLFA biomarkers
revealed a significant interaction soil age × plant species for Gram positive bacteria
(F3, 2 = 2.05, P = 0.03). Further analyses revealed significant differences in PLFA
biomarkers for Gram positive bacteria only for the 1.5-year-old soils (F1, 2.05 = 2.68, P =
0.033), and for unmined soils (F1, 0.99 = 2.61, P = 0.042; Table 1; Figure 3a). PLFA
biomarkers for microeucaryotes were significantly influenced by soil age (F3, 1.58 = 2.28,
P = 0.029), pair-wise analyses indicated that the 7-year-old soils were different from
unmined soils (t = 2.35, P = 0.007; Table 1). The rest of pair comparisons were not
significantly different (Figure 3b). PLFA biomarkers for total bacteria, Gram negative
bacteria, and fungi were not significantly influenced for the factors soil age or plant
species.
Figure 3. Scaling plot from principal co-ordinate analysis (PCO) of a) PLFA biomarkers for Gram positive bacteria, which were conditioned by Eucalyptus marginata and Bossiaea ornata in a species-specific manner only in the 1.5-year-old soils and in unmined soils, (significant interaction soil age × plant species) and b) PLFA biomarkers for microeucaryotes that were significantly influenced by soil age.
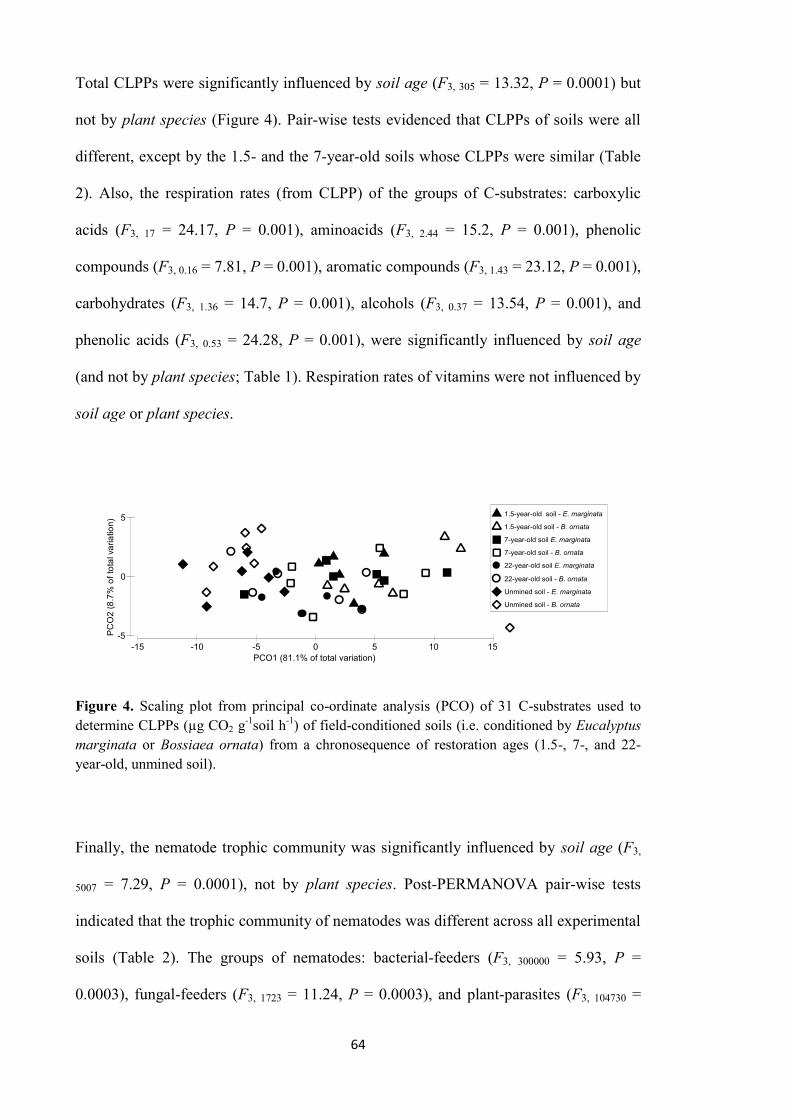
64
Total CLPPs were significantly influenced by soil age (F3, 305 = 13.32, P = 0.0001) but
not by plant species (Figure 4). Pair-wise tests evidenced that CLPPs of soils were all
different, except by the 1.5- and the 7-year-old soils whose CLPPs were similar (Table
2). Also, the respiration rates (from CLPP) of the groups of C-substrates: carboxylic
acids (F3, 17 = 24.17, P = 0.001), aminoacids (F3, 2.44 = 15.2, P = 0.001), phenolic
compounds (F3, 0.16 = 7.81, P = 0.001), aromatic compounds (F3, 1.43 = 23.12, P = 0.001),
carbohydrates (F3, 1.36 = 14.7, P = 0.001), alcohols (F3, 0.37 = 13.54, P = 0.001), and
phenolic acids (F3, 0.53 = 24.28, P = 0.001), were significantly influenced by soil age
(and not by plant species; Table 1). Respiration rates of vitamins were not influenced by
soil age or plant species.
Figure 4. Scaling plot from principal co-ordinate analysis (PCO) of 31 C-substrates used to determine CLPPs (µg CO2 g-1soil h-1) of field-conditioned soils (i.e. conditioned by Eucalyptus marginata or Bossiaea ornata) from a chronosequence of restoration ages (1.5-, 7-, and 22-year-old, unmined soil).
Finally, the nematode trophic community was significantly influenced by soil age (F3,
5007 = 7.29, P = 0.0001), not by plant species. Post-PERMANOVA pair-wise tests
indicated that the trophic community of nematodes was different across all experimental
soils (Table 2). The groups of nematodes: bacterial-feeders (F3, 300000 = 5.93, P =
0.0003), fungal-feeders (F3, 1723 = 11.24, P = 0.0003), and plant-parasites (F3, 104730 =
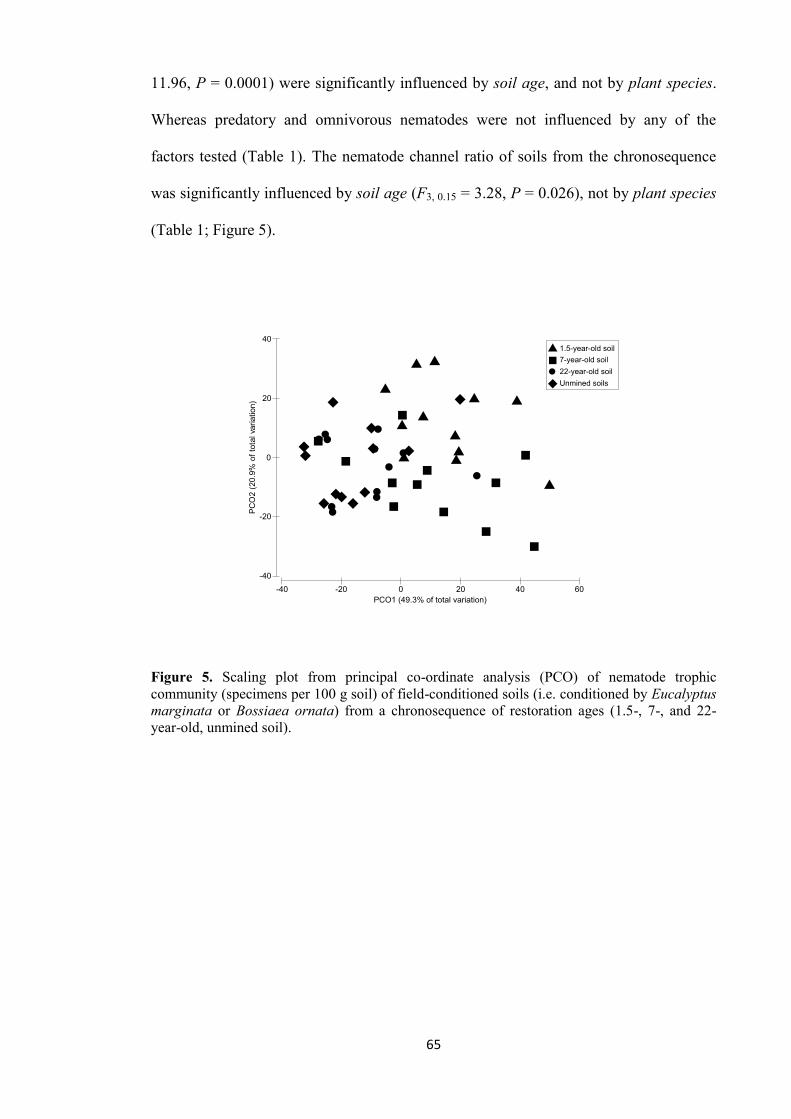
65
11.96, P = 0.0001) were significantly influenced by soil age, and not by plant species.
Whereas predatory and omnivorous nematodes were not influenced by any of the
factors tested (Table 1). The nematode channel ratio of soils from the chronosequence
was significantly influenced by soil age (F3, 0.15 = 3.28, P = 0.026), not by plant species
(Table 1; Figure 5).
Figure 5. Scaling plot from principal co-ordinate analysis (PCO) of nematode trophic community (specimens per 100 g soil) of field-conditioned soils (i.e. conditioned by Eucalyptus marginata or Bossiaea ornata) from a chronosequence of restoration ages (1.5-, 7-, and 22-year-old, unmined soil).
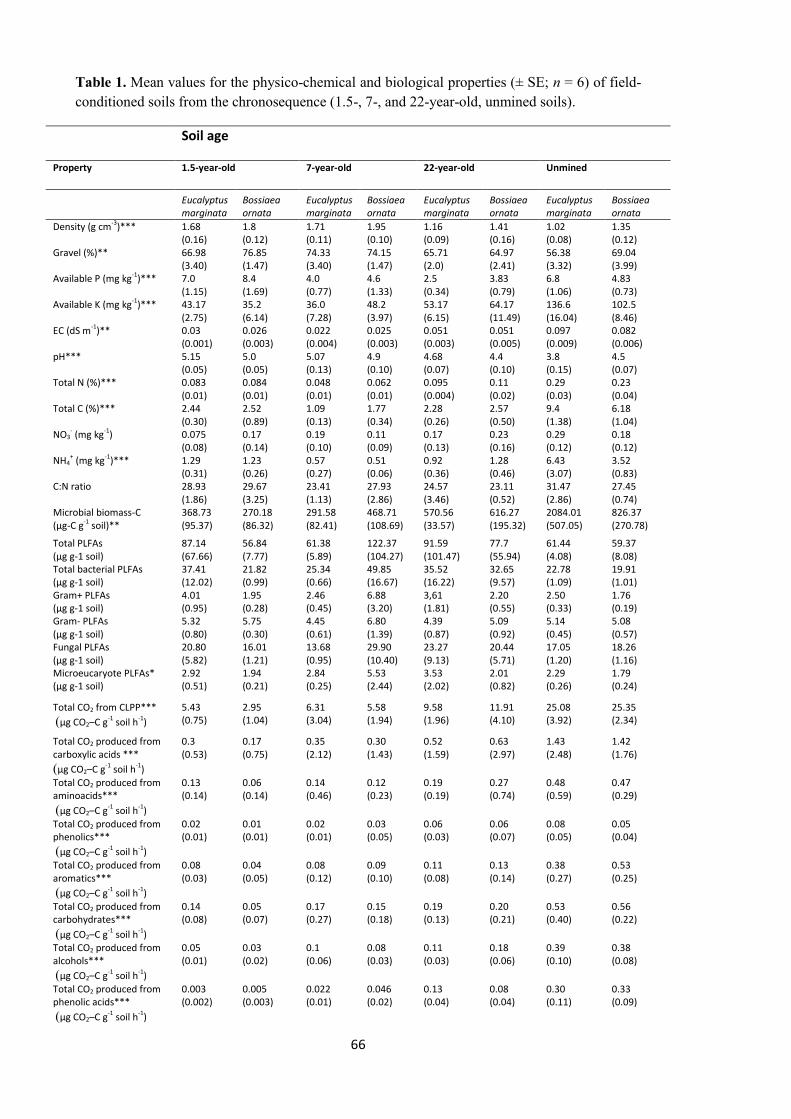
66
Table 1. Mean values for the physico-chemical and biological properties (± SE; n = 6) of field-conditioned soils from the chronosequence (1.5-, 7-, and 22-year-old, unmined soils).
Soil age
Property 1.5-year-old 7-year-old 22-year-old Unmined
Eucalyptus marginata
Bossiaea ornata
Eucalyptus marginata
Bossiaea ornata
Eucalyptus marginata
Bossiaea ornata
Eucalyptus marginata
Bossiaea ornata
Density (g cm-3)*** 1.68 (0.16)
1.8 (0.12)
1.71 (0.11)
1.95 (0.10)
1.16 (0.09)
1.41 (0.16)
1.02 (0.08)
1.35 (0.12)
Gravel (%)** 66.98 (3.40)
76.85 (1.47)
74.33 (3.40)
74.15 (1.47)
65.71 (2.0)
64.97 (2.41)
56.38 (3.32)
69.04 (3.99)
Available P (mg kg-1)*** 7.0 (1.15)
8.4 (1.69)
4.0 (0.77)
4.6 (1.33)
2.5 (0.34)
3.83 (0.79)
6.8 (1.06)
4.83 (0.73)
Available K (mg kg-1)*** 43.17 (2.75)
35.2 (6.14)
36.0 (7.28)
48.2 (3.97)
53.17 (6.15)
64.17 (11.49)
136.6 (16.04)
102.5 (8.46)
EC (dS m-1)** 0.03 (0.001)
0.026 (0.003)
0.022 (0.004)
0.025 (0.003)
0.051 (0.003)
0.051 (0.005)
0.097 (0.009)
0.082 (0.006)
pH*** 5.15 (0.05)
5.0 (0.05)
5.07 (0.13)
4.9 (0.10)
4.68 (0.07)
4.4 (0.10)
3.8 (0.15)
4.5 (0.07)
Total N (%)*** 0.083 (0.01)
0.084 (0.01)
0.048 (0.01)
0.062 (0.01)
0.095 (0.004)
0.11 (0.02)
0.29 (0.03)
0.23 (0.04)
Total C (%)*** 2.44 (0.30)
2.52 (0.89)
1.09 (0.13)
1.77 (0.34)
2.28 (0.26)
2.57 (0.50)
9.4 (1.38)
6.18 (1.04)
NO3- (mg kg-1) 0.075
(0.08) 0.17 (0.14)
0.19 (0.10)
0.11 (0.09)
0.17 (0.13)
0.23 (0.16)
0.29 (0.12)
0.18 (0.12)
NH4+ (mg kg-1)*** 1.29
(0.31) 1.23 (0.26)
0.57 (0.27)
0.51 (0.06)
0.92 (0.36)
1.28 (0.46)
6.43 (3.07)
3.52 (0.83)
C:N ratio 28.93 (1.86)
29.67 (3.25)
23.41 (1.13)
27.93 (2.86)
24.57 (3.46)
23.11 (0.52)
31.47 (2.86)
27.45 (0.74)
Microbial biomass-C (μg-C g-1 soil)**
368.73 (95.37)
270.18 (86.32)
291.58 (82.41)
468.71 (108.69)
570.56 (33.57)
616.27 (195.32)
2084.01 (507.05)
826.37 (270.78)
Total PLFAs (μg g-1 soil)
87.14 (67.66)
56.84 (7.77)
61.38 (5.89)
122.37 (104.27)
91.59 (101.47)
77.7 (55.94)
61.44 (4.08)
59.37 (8.08)
Total bacterial PLFAs (μg g-1 soil)
37.41 (12.02)
21.82 (0.99)
25.34 (0.66)
49.85 (16.67)
35.52 (16.22)
32.65 (9.57)
22.78 (1.09)
19.91 (1.01)
Gram+ PLFAs (μg g-1 soil)
4.01 (0.95)
1.95 (0.28)
2.46 (0.45)
6.88 (3.20)
3,61 (1.81)
2.20 (0.55)
2.50 (0.33)
1.76 (0.19)
Gram- PLFAs (μg g-1 soil)
5.32 (0.80)
5.75 (0.30)
4.45 (0.61)
6.80 (1.39)
4.39 (0.87)
5.09 (0.92)
5.14 (0.45)
5.08 (0.57)
Fungal PLFAs (μg g-1 soil)
20.80 (5.82)
16.01 (1.21)
13.68 (0.95)
29.90 (10.40)
23.27 (9.13)
20.44 (5.71)
17.05 (1.20)
18.26 (1.16)
Microeucaryote PLFAs* (μg g-1 soil)
2.92 (0.51)
1.94 (0.21)
2.84 (0.25)
5.53 (2.44)
3.53 (2.02)
2.01 (0.82)
2.29 (0.26)
1.79 (0.24)
Total CO2 from CLPP***
(μg CO2–C g-1 soil h-1)
5.43 (0.75)
2.95 (1.04)
6.31 (3.04)
5.58 (1.94)
9.58 (1.96)
11.91 (4.10)
25.08 (3.92)
25.35 (2.34)
Total CO2 produced from carboxylic acids ***
(μg CO2–C g-1 soil h-1)
0.3 (0.53)
0.17 (0.75)
0.35 (2.12)
0.30 (1.43)
0.52 (1.59)
0.63 (2.97)
1.43 (2.48)
1.42 (1.76)
Total CO2 produced from aminoacids***
(μg CO2–C g-1 soil h-1)
0.13 (0.14)
0.06 (0.14)
0.14 (0.46)
0.12 (0.23)
0.19 (0.19)
0.27 (0.74)
0.48 (0.59)
0.47 (0.29)
Total CO2 produced from phenolics***
(μg CO2–C g-1 soil h-1)
0.02 (0.01)
0.01 (0.01)
0.02 (0.01)
0.03 (0.05)
0.06 (0.03)
0.06 (0.07)
0.08 (0.05)
0.05 (0.04)
Total CO2 produced from aromatics***
(μg CO2–C g-1 soil h-1)
0.08 (0.03)
0.04 (0.05)
0.08 (0.12)
0.09 (0.10)
0.11 (0.08)
0.13 (0.14)
0.38 (0.27)
0.53 (0.25)
Total CO2 produced from carbohydrates***
(μg CO2–C g-1 soil h-1)
0.14 (0.08)
0.05 (0.07)
0.17 (0.27)
0.15 (0.18)
0.19 (0.13)
0.20 (0.21)
0.53 (0.40)
0.56 (0.22)
Total CO2 produced from alcohols***
(μg CO2–C g-1 soil h-1)
0.05 (0.01)
0.03 (0.02)
0.1 (0.06)
0.08 (0.03)
0.11 (0.03)
0.18 (0.06)
0.39 (0.10)
0.38 (0.08)
Total CO2 produced from phenolic acids***
(μg CO2–C g-1 soil h-1)
0.003 (0.002)
0.005 (0.003)
0.022 (0.01)
0.046 (0.02)
0.13 (0.04)
0.08 (0.04)
0.30 (0.11)
0.33 (0.09)
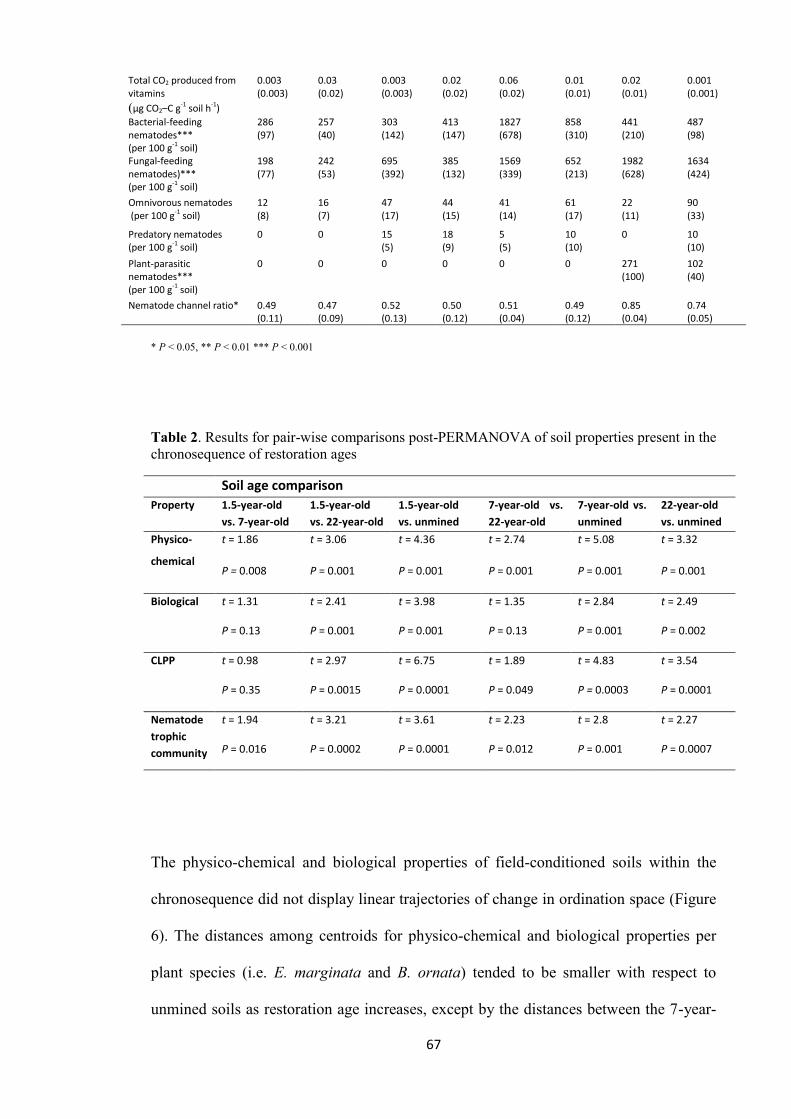
67
Total CO2 produced from vitamins
(μg CO2–C g-1 soil h-1)
0.003 (0.003)
0.03 (0.02)
0.003 (0.003)
0.02 (0.02)
0.06 (0.02)
0.01 (0.01)
0.02 (0.01)
0.001 (0.001)
Bacterial-feeding nematodes*** (per 100 g-1 soil)
286 (97)
257 (40)
303 (142)
413 (147)
1827 (678)
858 (310)
441 (210)
487 (98)
Fungal-feeding nematodes)***
(per 100 g-1 soil)
198 (77)
242 (53)
695 (392)
385 (132)
1569 (339)
652 (213)
1982 (628)
1634 (424)
Omnivorous nematodes (per 100 g-1 soil)
12 (8)
16 (7)
47 (17)
44 (15)
41 (14)
61 (17)
22 (11)
90 (33)
Predatory nematodes (per 100 g-1 soil)
0 0 15 (5)
18 (9)
5 (5)
10 (10)
0 10 (10)
Plant-parasitic nematodes*** (per 100 g-1 soil)
0 0 0 0 0 0 271 (100)
102 (40)
Nematode channel ratio* 0.49 (0.11)
0.47 (0.09)
0.52 (0.13)
0.50 (0.12)
0.51 (0.04)
0.49 (0.12)
0.85 (0.04)
0.74 (0.05)
* P < 0.05, ** P < 0.01 *** P < 0.001
Table 2. Results for pair-wise comparisons post-PERMANOVA of soil properties present in the chronosequence of restoration ages
Soil age comparison
Property 1.5-year-old
vs. 7-year-old
1.5-year-old
vs. 22-year-old
1.5-year-old
vs. unmined
7-year-old vs.
22-year-old
7-year-old vs.
unmined
22-year-old
vs. unmined
Physico-
chemical
t = 1.86 t = 3.06 t = 4.36 t = 2.74 t = 5.08 t = 3.32
P = 0.008 P = 0.001 P = 0.001 P = 0.001 P = 0.001 P = 0.001
Biological t = 1.31 t = 2.41 t = 3.98 t = 1.35 t = 2.84 t = 2.49
P = 0.13 P = 0.001 P = 0.001 P = 0.13 P = 0.001 P = 0.002
CLPP t = 0.98 t = 2.97 t = 6.75 t = 1.89 t = 4.83 t = 3.54
P = 0.35 P = 0.0015 P = 0.0001 P = 0.049 P = 0.0003 P = 0.0001
Nematode
trophic
community
t = 1.94 t = 3.21 t = 3.61 t = 2.23 t = 2.8 t = 2.27
P = 0.016 P = 0.0002 P = 0.0001 P = 0.012 P = 0.001 P = 0.0007
The physico-chemical and biological properties of field-conditioned soils within the
chronosequence did not display linear trajectories of change in ordination space (Figure
6). The distances among centroids for physico-chemical and biological properties per
plant species (i.e. E. marginata and B. ornata) tended to be smaller with respect to
unmined soils as restoration age increases, except by the distances between the 7-year-

68
old soils and unmined soils for the physico-chemical and biological properties
conditioned in the field by E. marginata, and the physico-chemical properties
conditioned in the field by B. ornata. The distances among centroids for the physico-
chemical properties conditioned in the field by E. marginata are: 1.5-year-old to
unmined = 5.01; 7-year-old to unmined = 6.13; 22-year-old to unmined = 4.35. The
distances among centroids for the biological properties conditioned in the field by E.
marginata are: 1.5-year-old soil to unmined = 3.65; 7-year-old to unmined = 3.75; 22-
year-old to unmined = 3.2. The distances among centroids for the physico-chemical
properties conditioned by B. ornata in the field are: 1.5-year-old soil to unmined = 4.29;
7-year-old to unmined = 4.52; 22-year-old to unmined = 3.03. The distances among
centroids for the biological properties conditioned in the field by B. ornata are: 1.5-
year-old soil to unmined = 3.7; 7-year-old to unmined = 3.4; 22-year-old to unmined =
2.6.
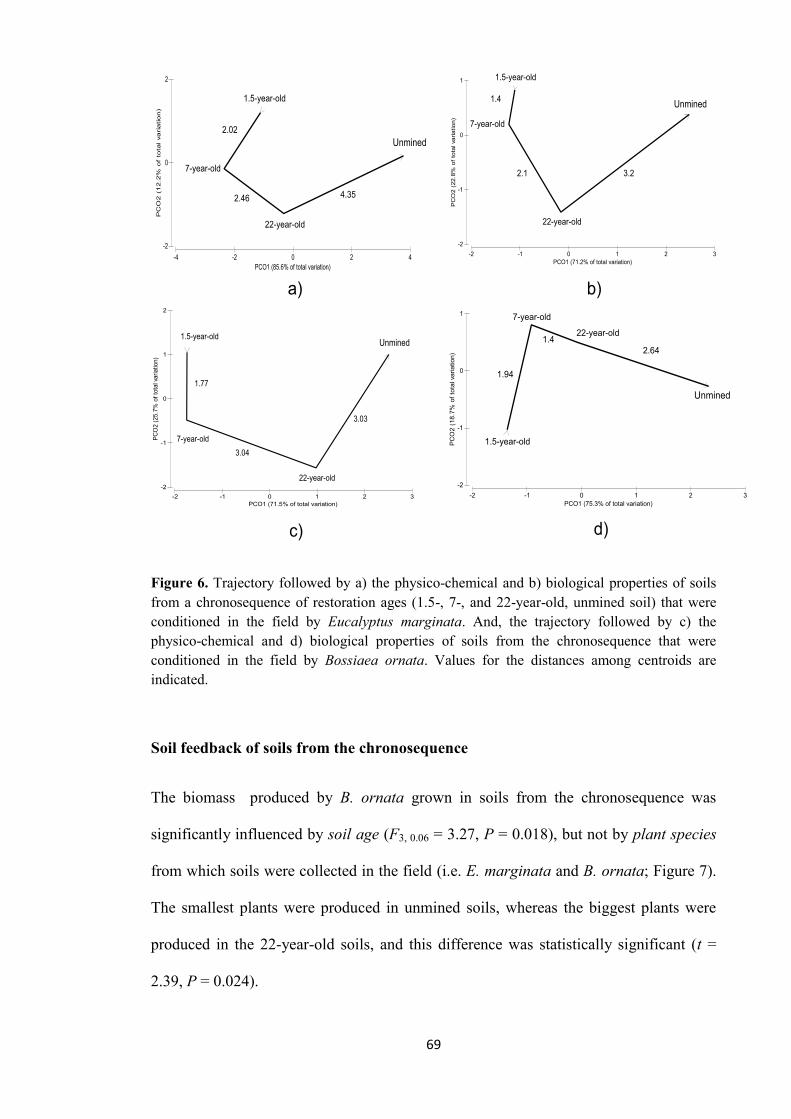
69
Figure 6. Trajectory followed by a) the physico-chemical and b) biological properties of soils from a chronosequence of restoration ages (1.5-, 7-, and 22-year-old, unmined soil) that were conditioned in the field by Eucalyptus marginata. And, the trajectory followed by c) the physico-chemical and d) biological properties of soils from the chronosequence that were conditioned in the field by Bossiaea ornata. Values for the distances among centroids are indicated.
Soil feedback of soils from the chronosequence
The biomass produced by B. ornata grown in soils from the chronosequence was
significantly influenced by soil age (F3, 0.06 = 3.27, P = 0.018), but not by plant species
from which soils were collected in the field (i.e. E. marginata and B. ornata; Figure 7).
The smallest plants were produced in unmined soils, whereas the biggest plants were
produced in the 22-year-old soils, and this difference was statistically significant (t =
2.39, P = 0.024).
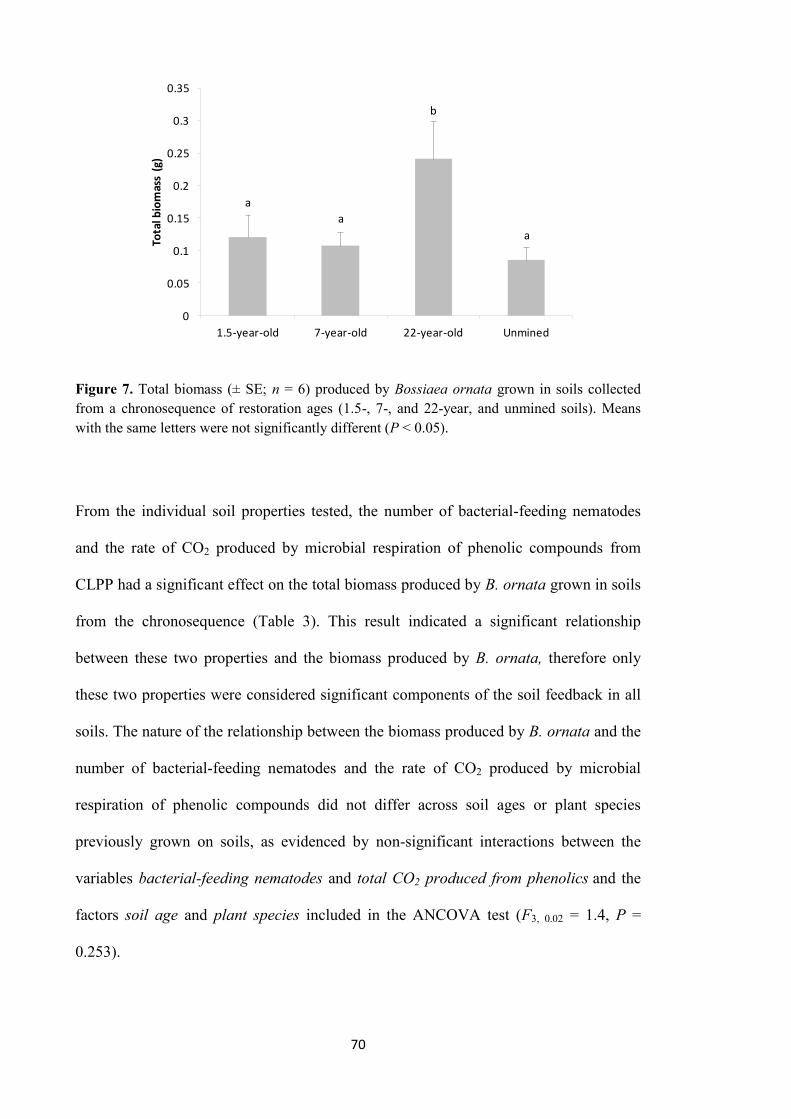
70
Figure 7. Total biomass (± SE; n = 6) produced by Bossiaea ornata grown in soils collected from a chronosequence of restoration ages (1.5-, 7-, and 22-year, and unmined soils). Means with the same letters were not significantly different (P < 0.05).
From the individual soil properties tested, the number of bacterial-feeding nematodes
and the rate of CO2 produced by microbial respiration of phenolic compounds from
CLPP had a significant effect on the total biomass produced by B. ornata grown in soils
from the chronosequence (Table 3). This result indicated a significant relationship
between these two properties and the biomass produced by B. ornata, therefore only
these two properties were considered significant components of the soil feedback in all
soils. The nature of the relationship between the biomass produced by B. ornata and the
number of bacterial-feeding nematodes and the rate of CO2 produced by microbial
respiration of phenolic compounds did not differ across soil ages or plant species
previously grown on soils, as evidenced by non-significant interactions between the
variables bacterial-feeding nematodes and total CO2 produced from phenolics and the
factors soil age and plant species included in the ANCOVA test (F3, 0.02 = 1.4, P =
0.253).
a
a
b
a
0
0.05
0.1
0.15
0.2
0.25
0.3
0.35
1.5-year-old 7-year-old 22-year-old Unmined
Tota
l bio
mas
s (g
)
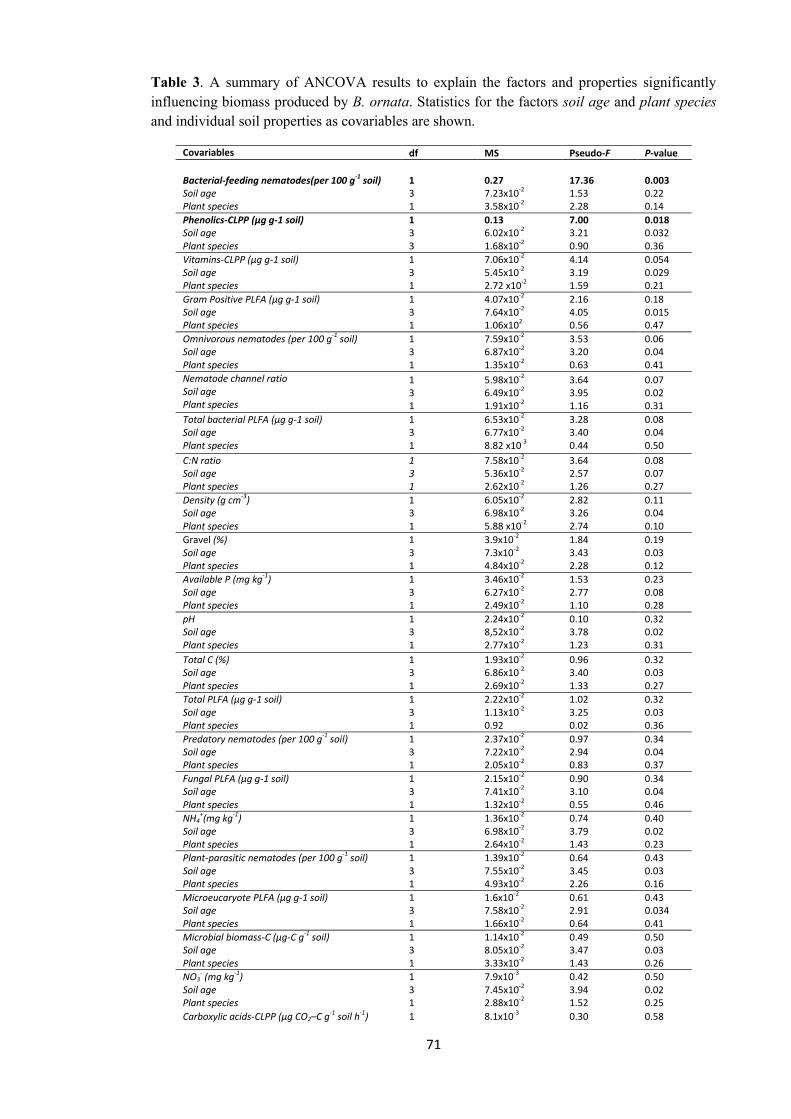
71
Table 3. A summary of ANCOVA results to explain the factors and properties significantly influencing biomass produced by B. ornata. Statistics for the factors soil age and plant species and individual soil properties as covariables are shown.
Covariables df MS Pseudo-F P-value
Bacterial-feeding nematodes(per 100 g-1 soil) 1 0.27 17.36 0.003 Soil age 3 7.23x10-2 1.53 0.22 Plant species 1 3.58x10-2 2.28 0.14
Phenolics-CLPP (μg g-1 soil) Soil age Plant species
1 3 3
0.13 6.02x10-2
1.68x10-2
7.00 3.21 0.90
0.018 0.032 0.36
Vitamins-CLPP (μg g-1 soil) Soil age Plant species
1 3 1
7.06x10-2
5.45x10-2
2.72 x10-2
4.14 3.19 1.59
0.054 0.029 0.21
Gram Positive PLFA (μg g-1 soil) Soil age Plant species
1 3 1
4.07x10-2
7.64x10-2
1.06x102
2.16 4.05 0.56
0.18 0.015 0.47
Omnivorous nematodes (per 100 g-1 soil) 1 7.59x10-2 3.53 0.06 Soil age 3 6.87x10-2 3.20 0.04 Plant species 1 1.35x10-2 0.63 0.41
Nematode channel ratio Soil age Plant species
1 3 1
5.98x10-2 6.49x10-2 1.91x10-2
3.64 3.95 1.16
0.07 0.02 0.31
Total bacterial PLFA (μg g-1 soil) Soil age Plant species
1 3 1
6.53x10-2
6.77x10-2
8.82 x10-3
3.28 3.40 0.44
0.08 0.04 0.50
C:N ratio Soil age Plant species
1 3 1
7.58x10-2 5.36x10-2 2.62x10-2
3.64 2.57 1.26
0.08 0.07 0.27
Density (g cm-3) 1 6.05x10-2 2.82 0.11 Soil age 3 6.98x10-2 3.26 0.04 Plant species 1 5.88 x10-2 2.74 0.10
Gravel (%) 1 3.9x10-2 1.84 0.19 Soil age 3 7.3x10-2 3.43 0.03 Plant species 1 4.84x10-2 2.28 0.12
Available P (mg kg-1) 1 3.46x10-2 1.53 0.23 Soil age 3 6.27x10-2 2.77 0.08 Plant species 1 2.49x10-2 1.10 0.28
pH Soil age Plant species
1 3 1
2.24x10-2 8,52x10-2 2.77x10-2
0.10 3.78 1.23
0.32 0.02 0.31
Total C (%) 1 1.93x10-2 0.96 0.32 Soil age Plant species
3 1
6.86x10-2 2.69x10-2
3.40 1.33
0.03 0.27
Total PLFA (μg g-1 soil) Soil age Plant species
1 3 1
2.22x10-2
1.13x10-2
0.92
1.02 3.25 0.02
0.32 0.03 0.36
Predatory nematodes (per 100 g-1 soil) 1 2.37x10-2 0.97 0.34 Soil age 3 7.22x10-2 2.94 0.04 Plant species 1 2.05x10-2 0.83 0.37
Fungal PLFA (μg g-1 soil) Soil age Plant species
1 3 1
2.15x10-2
7.41x10-2
1.32x10-2
0.90 3.10 0.55
0.34 0.04 0.46
NH4+(mg kg-1) 1 1.36x10-2 0.74 0.40
Soil age 3 6.98x10-2 3.79 0.02 Plant species 1 2.64x10-2 1.43 0.23
Plant-parasitic nematodes (per 100 g-1 soil) 1 1.39x10-2 0.64 0.43 Soil age 3 7.55x10-2 3.45 0.03 Plant species 1 4.93x10-2 2.26 0.16
Microeucaryote PLFA (μg g-1 soil) Soil age Plant species
1 3 1
1.6x10-2
7.58x10-2
1.66x10-2
0.61 2.91 0.64
0.43 0.034 0.41
Microbial biomass-C (μg-C g-1 soil) 1 1.14x10-2 0.49 0.50 Soil age 3 8.05x10-2 3.47 0.03 Plant species 1 3.33x10-2 1.43 0.26
NO3- (mg kg-1) 1 7.9x10-3 0.42 0.50
Soil age 3 7.45x10-2 3.94 0.02 Plant species 1 2.88x10-2 1.52 0.25
Carboxylic acids-CLPP (μg CO2–C g-1 soil h-1) 1 8.1x10-3 0.30 0.58

72
Soil age 3 7.42x10-2 2.73 0.051 Plant species 1 2.1x10-2 0.78 0.42
Total N (%) 1 4.94x10-3 0.20 0.65 Soil age 3 7.72x10-2 3.21 0.04 Plant species 1 2.68x10-2 1.11 0.30
EC (dS m-1) 1 4.1x10-3 0.19 0.66 Soil age 3 9.68x10-2 4.56 0.01 Plant species 1 3.63x10-2 1.71 0.19
Total C-CLPP (μg CO2–C g-1 soil h-1) 1 3.28x10-3 0.13 0.71 Soil age 3 7.33x10-2 2.87 0.055 Plant species 1 2.56x10-2 1.00 0.33
Aromatics-CLPP (μg g-1 soil) Soil age Plant species
1 3 1
2.1 x10-3
7.81x10-2 1.81x10-2
0.10 3.72 0.86
0.74 0.018 0.36
Gram Negative PLFA (μg g-1 soil) Soil age Plant species
1 3 1
2.9 x10-3
7.37x10-2
2.40x10-2
0.11 2.81 0.91
0.74 0.06 0.31
Phenolic acids-CLPP (μg CO2–C g-1 soil h-1) Soil age Plant species
1 3 1
2.15x10-3
7.74x10-2
1.77x10-2
8.23x10-2
2.96 0.680
0.78 0.04 0.44
Available K (mg kg-1) 1 1.2x10-3 6.06x10-2 0.79 Soil age 3 7.98x10-2 4.02 0.02 Plant species 1 2.95x10-2 1.44 0.25
Alcohols-CLPP (μg CO2–C g-1 soil h-1) Soil age Plant species
1 3 1
1.62x10-3
8.6x10-2
1.9x10-2
7.32x10-2
3.89 0.86
0.79 0.02 0.36
Aminoacids-CLPP (μg CO2–C g-1 soil h-1) Soil age Plant species
1 3 1
1.33x10-3
8.05x10-2
2.21x10-2
5.07x10-2
3.07 0.84
0.82 0.024 0.37
Carbohydrates-CLPP (μg CO2–C g-1 soil h-1) Soil age Plant species
1 3 1
1.40x10-3
9.52x10-2
1.32x10-2
5.92x10-2
4.04 0.56
0.83 0.019 0.45
Fungal-feeding nematodes (per 100 g-1 soil) 1 4.11x10-4 1.95x10-2 0.90 Soil age 3 7.39x10-2 3.51 0.03 Plant species 1 2.85x10-2 1.35 0.27
Individual PSF with respect to soil age
No statistical differences were found in the total biomass produced by B. ornata in
home (i.e. soils conditioned in the field by B. ornata) with respect to away soils (i.e.
soils conditioned in the field by E. marginata), therefore a neutral PSF was observed in
all soils from the chronosequence (Figure 8).
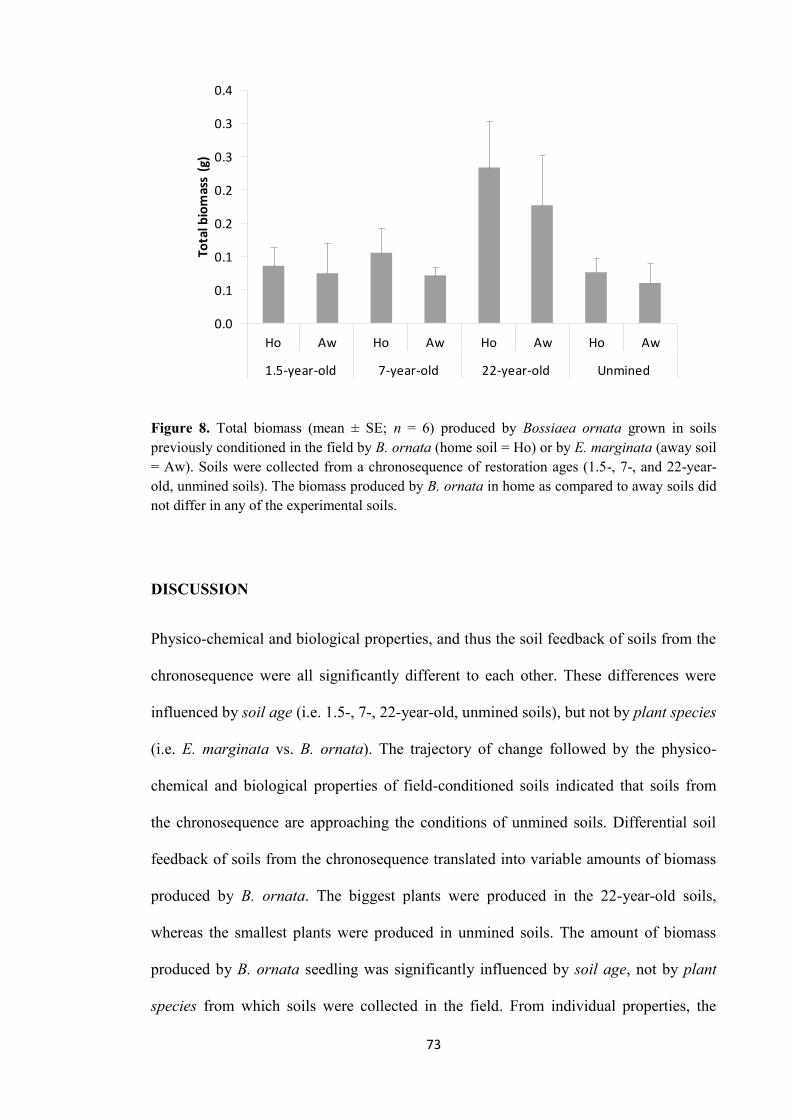
73
Figure 8. Total biomass (mean ± SE; n = 6) produced by Bossiaea ornata grown in soils previously conditioned in the field by B. ornata (home soil = Ho) or by E. marginata (away soil = Aw). Soils were collected from a chronosequence of restoration ages (1.5-, 7-, and 22-year-old, unmined soils). The biomass produced by B. ornata in home as compared to away soils did not differ in any of the experimental soils.
DISCUSSION
Physico-chemical and biological properties, and thus the soil feedback of soils from the
chronosequence were all significantly different to each other. These differences were
influenced by soil age (i.e. 1.5-, 7-, 22-year-old, unmined soils), but not by plant species
(i.e. E. marginata vs. B. ornata). The trajectory of change followed by the physico-
chemical and biological properties of field-conditioned soils indicated that soils from
the chronosequence are approaching the conditions of unmined soils. Differential soil
feedback of soils from the chronosequence translated into variable amounts of biomass
produced by B. ornata. The biggest plants were produced in the 22-year-old soils,
whereas the smallest plants were produced in unmined soils. The amount of biomass
produced by B. ornata seedling was significantly influenced by soil age, not by plant
species from which soils were collected in the field. From individual properties, the
0.0
0.1
0.1
0.2
0.2
0.3
0.3
0.4
Ho Aw Ho Aw Ho Aw Ho Aw
1.5-year-old 7-year-old 22-year-old Unmined
Tota
l bio
mas
s (g
)

74
number of bacterial-feeding nematodes and the rate of CO2 produced by microbial
respiration of phenolic compounds from CLPP were evidenced as significant
components of the soil feedback across soils from the chronosequence. These properties
had a strong and significant effect on the biomass produced by B. ornata in all soils.
Differential soil feedback of soils from the chronosequence did not translate into
variable individual PSF, and neutral PSF was associated with B. ornata across all soils.
The physico-chemical and biological properties of field-conditioned soils from the
chronosequence were all different to each other, and significantly influenced by soil age
(i.e. 1.5-, 7-, 22-year-old, unmined soils) as predicted. The time-dependent changing
nature of soils responds to biogeochemical and biological processes (Hobbie 1992;
Eviner 2004; Viketoft et al. 2005; Bezemer et al. 2006; De Bellis et al. 2007; Berg &
Smalla 2009). However, the results agreed (partially) with the initial prediction, since a
significant effect of plant species on soil properties was also expected. Particularly the
physico-chemical properties of the 1.5-year-old soils were not significantly influenced
by plant species as observed in the experiment described in Chapter 3. The discrepancy
in the results observed for this and the previous experiment for the 1.5-year-old soil can
be the result of both E. marginata and B. ornata causing similar changes to soils. This
behaviour was observed previously but only in unmined soils, where similar
conditioning processes of the chemical properties were found by E. marginata and B.
ornata (Chapter 3). Another possibility is that in the field, species-specific conditioning
of soil properties was constrained by strong soil age effects, since soil is a dynamic
evolving entity due to biogeochemical processes, climate influence, and changes
associated to biotic communities (Foth 1990; Lavelle & Spain 2003; Buscot & Varma
2005).

75
Experimentally, the discrepancy observed in case of the previous and this research
might be attributed to differences in soil conditioning processes present in each case.
Soil conditioning is influenced by the experimental procedure used, and for this
research, plant-conditioned soils were collected from single plants in the field, which is
known as the ‘natural experiment approach’ (Kulmatiski & Kardol 2008; Pernilla et al.
2010; van de Voorde et al. 2012), whereas for the previous work soils were collected
from root area of E. marginata, B. ornata, and Acacia pulchella, and from other
legumes, all soils were mixed, distributed into pots and conditioned under controlled
conditions in the glasshouse, this procedure is referred to as the ‘cultivation experiment
approach’ (Kulmatiski & Kardol 2008; Pernilla et al. 2010; van de Voorde et al. 2012).
These findings suggest that conditioning process is context-dependent and shows some
level of plasticity.
Properties such as density, gravel content, and pH decreased as restoration age
increased, probably in response to development of soil structure, stabilization of
aggregates, and biogeochemical processes (Jastrow et al. 1998; Malik & Scullion 1998;
Ward 2000; Banning et al. 2008). The concentrations of available K and EC increased
with increasing soil age, while available P, total N and total C decreased in the 7-year-
old soils as compared to the 1.5-year-old soils, but increased again in the 22-year-old
soils. Restoration of the jarrah forest includes N-P-K fertilisation early in the process
(Koch 2007b), and for this reason these nutrients are in high concentrations at early
stages of restoration as observed in the 1.5-year-old soils. With time, N-P-K
concentrations decrease due to plant uptake and leaching. As plant biomass production
increases and the process of organic matter recycling recovers, the concentrations of
nutrients and C in soil increase (Ward 2000; Banning et al. 2008; George et al. 2010;
Lin et al. 2011) as observed in the 22-year-old soils whose physico-chemical and
biological properties are close to those of unmined soils.

76
Microbial biomass-C increased with restoration age, whereby the oldest soils in the
chronosequence had values that were similar to that of unmined soils, a result which is
consistent with other studies (Banning et al. 2008). From PLFA biomarkers, only those
for Gram positive bacteria and microeucaryotes statistically differed in experimental
soils. Gram positive bacteria were conditioned by E. marginata and B. ornata in a
species-specific manner but only in the 1.5-year-old soils and in unmined soils. In these
two soil ages a higher concentration of PLFA biomarkers for Gram positive bacteria
were found under the influence of E. marginata as compared with B. ornata. Gram
positive bacteria usually form endospores, and their population is large in soils,
consequently, this bacterial group can survive under adverse conditions (Lavelle &
Spain 2003; Büdel 2005; Barriuso et al. 2008), such as those present in soil restored
post-bauxite mining. In degraded soils, Gram positive bacteria recover quickly because
these bacteria are very responsive to C inputs released to soil via root exudates (Jasper
2007; Caesar-TonThat et al. 2008). Bacterial populations developed under early-
successional stages of ecosystems are more sensitive to C inputs from plant species
(Jasper 2007; Papatheodorou 2008), as was observed for the 1.5-year-old soils, and not
for the other soils within the chronosequence. However, since root exudates are
produced in a species-specific (Marschner 1995; Uren 2001), I expected that
populations of Gram positive bacteria would be conditioned according to plant species
as was found during the experiment. Previous works have evidenced the capacity of E.
marginata to condition biotic communities of soil likely via root exudates (Malajczuk &
McComb 1977; Malajczuk 1979; Malajczuk & McComb 1979). In unmined soils, plant
conditioning of soil microbial populations (i.e. Gram positive bacteria) was expected
due to species-specific rhizodeposition released to soil for long time spans, a
phenomenon that has been evidenced by other works (Malajczuk & McComb 1979; De
Bellis et al. 2007; Zechmeister-Boltenstern et al. 2011; Prescott & Graystone 2013).

77
PLFA biomarkers for microeucaryotes were in higher concentration in the 7-year-old
soils but only when this soil age was compared with unmined soils. Since the rest of
comparisons for microeucaryote PLFAs among soils within the chronosequence were
not significantly different, I suggest this is a particular soil property of the 7-year-old
soils (i.e. an outlier).
Functional aspects of the microbial community (CLPP) differentiated only at late stages
of restoration, a result which is consistent with what has been previously observed
(Lalor et al. 2007). Populations of nematodes from diverse trophic groups augmented
with increasing restoration age, maximum numbers were observed in unmined soils
(except for bacterial-feeding nematodes). Plant-parasitic nematodes were exclusively
observed in the latter soils. According to the nematode channel ratio values, both the
bacterial-based as well as the fungal-based energy channel of decomposition were
simultaneously operating in soils across the chronosequence (i.e. nematode channel
ratio close or equal to 0.5). In contrast, in unmined soils a fungal-based energy channel
of organic matter decomposition operated (i.e. nematode channel ratio > 0.5; Moore &
Hunt 1988; Yeates 2003). Individual biological properties of soils from the
chronosequence tended to increase or ‘develop’ with increasing restoration age,
however, even after 22 years post-restoration, there are functional aspects of soil (i.e.
the nematode channel ratio) that are dissimilar from unmined soils.
Individual properties of field-conditioned soils of increasing restoration age tended to
reach the values of the properties of unmined soils as initially predicted. However, the
overlaid trajectory of multivariate change based on the physico-chemical and biological
properties of soils conditioned in the field by E. marginata and B. ornata was not linear.
Hypothetically, a maximum distinction between 1.5-year-old and unmined soils with
regard to physico-chemical and biological properties should be observed, and this

78
distinction should lessen as restoration age increases. Nonetheless distances between
centroids of the 7-year-old soils and the unmined soils were even longer than the
distances between centroids of 1.5-year-old soils and the unmined soils (except by the
biological properties conditioned by B. ornata). These results suggest that the properties
and thus the soil feedback of restored soils post-bauxite are recovering with increasing
time, however soils from the 7-year-old site selected for this experiment might be
following an alternative trajectory of change, and statistically this site can be considered
as an outlier (Suding et al. 2004; Grant 2006).
There was a differential soil feedback effect on the biomass produced by B. ornata
grown in soils from the chronosequence. Plant biomass was significantly influenced by
soil age as predicted, however any significant effect of plant species was observed.
These findings differed from the results observed in the experiment described in the
previous chapter, where plant species previously grown in soils had a significant effect
on the biomass produced by B. ornata in the 1.5-year-old soils (see Chapter 3).
However, as explained above, the experimental approach used in each one of the
experiments might have influenced the results (i.e. cultivation experiment approach
used in experiment described in Chapter 3 vs. natural experiment approach used in this
experiment; Kulmatiski & Kardol 2008; Pernilla et al. 2010; van de Voorde et al. 2012).
Differences in biomass produced by B. ornata in soils from the chronosequence suggest
that biomass production was influenced by changing contexts of soils (Manning et al.
2008; te Beest et al. 2009; Meijer et al. 2011). The 22-year-old soils exerted a positive
feedback on B. ornata that produced up to two times more biomass with respect to the
other soils. The values for the properties of the 22-year-old soils were increased, and
approached those of unmined soils, therefore, similar amounts of biomass were
expected to be produced by B. ornata in the 22-year-old and in unmined soils. However,

79
plants grown in the 22-year-old soils were significantly bigger that those grown in
unmined soils. The more marked differences between the 22-year-old soils and unmined
soils were the presence of plant-parasitic nematodes in the latter, and increased numbers
of bacterial-feeding and predatory nematodes in the 22-year-old soils. Therefore, the
increased biomass produced by B. ornata in the 22-year-old soils might respond to the
absence of plant-parasitic nematodes, and faster organic matter decomposition and
turnover due to the presence of bacterial-feeding and predatory nematodes. Plant-
parasitic nematodes can cause severe damage to plants due to root damage (Perry &
Tovar-Soto 2012), and are drivers of negative PSF in ecosystems (De Rooij-van der
Goes 1995; van der Putten & Peters 1997; van der Stoel et al. 2002). Bacterial-feeding
nematodes can mineralize a substantial amount of nitrogen that is available for plant
uptake (Freckman 1988), therefore can be considered drivers of positive PSF.
The number of bacterial-feeding nematodes and the rate of CO2 produced by microbial
respiration of phenolic compounds from CLPP were identified as significant
components of the soil feedback of soils from the chronosequence. These properties
showed a strong and significant influence on the biomass produced by B. ornata across
soils from the chronosequence. This finding supports the results of the increased
biomass produced by B. ornata in the 22-year-old soils, which might be a response of
increased bacterial-feeding nematodes in soil. Bacterial-feeding nematodes display
characteristics that make them an important group of soil organisms. They directly
influence plant productivity by releasing ammonia (i.e. the major excretory product of
nematodes) that is available for plant uptake (Freckman 1988; Yeates 2003; Postma-
Blaauw et al. 2005). Bacterial-feeding nematodes are diverse in ecosystems and
characteristics such as a short generation time and high fecundity allow these organisms
to quickly recover after soil disturbances (Freckman 1988; Bongers & Bongers 1998;
Yeates 2003; Holtkamp et al. 2008), including restoration post-bauxite mining.

80
Consequently their influence on plant productivity might start at early stages of plant
development and might persist at late stages of restoration. In the mid- and long-term,
bacterial feeding nematodes indirectly affect plant growth by dispersing microbes,
feeding on pathogenic bacteria, and being source of nutrients for other organisms within
the soil food web (Freckman 1988; Wardle 1999).
Three fenolic compounds were tested as substrates for CLPP, these were catechin,
coumarin and quercetin. These phenolic compounds are exuded by roots, and exhibit
antimicrobial and allelopathic properties (Lavelle & Spain 2003; Singh & Mukerji
2006; Bais et al. 2008; Hadacek 2008; Badri & Vivanco 2009). Chemicals with
antimicrobial and allelopathic action have been detected in exudates and litter of
Australian plants (Whitfield 1981; Deans 2002; D’Souza et al. 2004; Hussain et al.
2011; Lorenzo et al. 2013). Phenolic compounds released to soil via exudates and litter
are degraded (respired) by several microorganisms, for example pseudomonad (Inderjit
2005; Reuben et al. 2008; Zhang et al. 2009). Microbial degradation of C compounds is
substrate-dependent, my results indicated that phenolic compounds have been present in
soils and consequently degraded by soil microbes as evidenced by CLPP results. This
finding is expected because in jarrah forest, production of allelochemicals is critical for
plant persistence, since competition with other plants and soil organisms for moisture
and nutrients is intense, thus plants equipped with efficient inhibitory chemicals raise
their chances of survival (Willis 1999; Manoharachary & Mukerji 2006). I suggest that
respiration of phenolic compounds was evidenced as a key component of the soil
feedback because this is an indirect way to observe allelopathic properties of jarrah
forest plants, which is a key phenomenon for plant establishment and development (i.e.
biomass production) in jarrah forest.

81
The fact that biological properties were found to be components of the soil feedback of
soils from the chronosequence disagree with my initial prediction, I expected to observe
mainly physico-chemical properties significantly influencing the biomass produced by
B. ornata via soil feedback in recently-restored soils. Then, as restoration age increased,
I expected to observe mainly biological properties significantly influencing the biomass
produced by B. ornata via soil feedback at late-restored soils. This prediction was
formulated given that recently-restored soils display chemical properties such as
increased concentrations of P, N, and K that are consequence of fertiliser addition as
part of the restoration process (Koch 2007b). Biological properties of soil are likely to
influence plant growth via soil feedback in the mid-, long-term, and not in the short-
term due to the dependence of native plants on biologically-supplied nutrients such as N
and P (Lawrie 1981; Brundrett & Abbott 1991; Lambers et al. 2007), and decreasing
concentrations of nutrients from fertilisers.
Differential soil feedback of soils from the chronosequence did not translate into
differential individual PSF (i.e. magnitude and direction), that is, a neutral PSF was
associated with B. ornata. The results for the 22-year-old and unmined soils are
consistent with the prediction of neutral PSF associated with B. ornata. However, PSF
associated with B. ornata grown in the 1.5-year-old soils disagreed with the prediction
of negative PSF based on results from a previous experiment (see Chapter 3). In both,
this and the previous experiment recently-restored soils were the same age (i.e. 1.5-
year-old soils), however the results suggested that conditioning process is influenced by
soil contexts (Kulmatiski & Kardol 2008; Pernilla et al. 2010; van de Voorde et al.
2012). In the previous experiment soils were collected from different plants species,
mixed, and conditioned in the glasshouse, whereas for this research soils were collected
from single plants in the field, thus soils were conditioned under field conditions.

82
In conclusion, the results from the characterization of soils from the chronosequence
indicated that soil feedback of restored soils post-bauxite mining is recovering, but
alternative trajectories can occur as observed for the 7-year-old soil. However, these
results did not translate into altered patterns of biomass produced by B. ornata in the 7-
year-old soil. Instead, an increased biomass production was observed in the 22-year-old
soils, which suggested this soil age exerted a differential and positive soil feedback.
Despite this, individual PSF associated with B. ornata was not affected, and neutral PSF
was observed in all soils from the chronosequence.

83
CHAPTER 5
Long-term conditioning of soil by eucalypts and pines does not alter
the plant-soil feedback associated with native jarrah
INTRODUCTION
Plant-soil feedbacks can influence the restoration of native forests where soil has been
conditioned by non-native species for long periods of time (i.e. agricultural, forestry
activities, invader species; Kulmatiski et al. 2006; Jordan et al. 2008; Kulmatiski &
Beard 2008). Specifically, non-native species can condition soil properties in ways that
lead to reduced subsequent growth of native species (van Grunsven et al. 2007; Perkins
& Nowak 2012; Suding et al. 2013), due to alterations of nutrient cycling processes
(Bezemer et al. 2006; Kulmatiski et al. 2006; Kardol & Wardle 2010; Weidenhamer &
Callaway 2010), imbalance of microbial populations (i.e. free-living, plant-pathogens
and root symbionts; Indejit & van der Putten 2010), and negative effects of toxic
allelochemicals (Bertin et al. 2003; Badri & Vivanco 2009; Indejit & van der Putten
2010; Weidenhamer & Callaway 2010). Plant-conditioned properties of soil can persist
even after the removal of the non-native species (Kardol & Wardle 2010).
Soil conditioning operates through two mechanisms: 1) leaf litter deposition, and 2) root
deposition. In particular, litter quality is a species-specific plant trait (i.e. C:N ratio,
lignin and phenolic compounds content) that directly influences the chemical and
biological properties of soil. Elements released from litter during decomposition alter
the concentrations of soil macro and micronutrients in a species-specific manner
(Hobbie 1992; Guo & Sims 1999; Berg & McClaugherty 2008; Witt & Setälä 2010;
Uselman et al. 2012). As well, litter deposition drives changes in the microbial and
microfauna (i.e. nematode) community, including its size, composition, and
physiological properties (Berg & McClaugherty 2008). Generally, plant species that

84
produce labile litter (i.e. low C:N ratio) such as legumes, promote a microbial
community dominated by bacteria and their consumers (i.e. protozoa, bacterial-feeding
nematodes). In contrast, plant species that produce recalcitrant litter (i.e. high C:N ratio)
such as pines and eucalypts promote a microbial community dominated by fungi and
their consumers (i.e. fungal-feeding nematodes, collembolans, oribatid mites; Moore &
Hunt 1988; Scheu et al. 2005; Moore et al. 2007; Holtkamp et al. 2008; Witt & Setälä
2010).
In the jarrah forest of south-west Western Australia, the first restoration attempts post-
bauxite mining consisted of commercial plantations of fast-growing, exotic softwoods,
mainly pines (Pinus radiata) and cypresses (Cupressus sp.), as well as eastern
Australian hardwood eucalypts such as Sydney blue gum (Eucalyptus saligna; Koch
2007a; Koch 2007b). To date there are 3,600 ha of plantations of non-native species
within the jarrah forest ecoregion that will be harvested for timber in the near future and
then restored to native jarrah forest (Conservation Commission of Western Australia
2012). At these sites non-native trees have been conditioning the soil properties for
decades. The general aim of my research was to determine what differences (if any)
occurred in the soils after 35 years of conditioning by non-native species, and to
determine the effect of conditioning on the subsequent growth of jarrah (E. marginata)
seedlings (i.e. individual PSF).
Specifically, the research aimed to determine changes in properties of mined soils that
have been conditioned in the long-term by pines (P. radiata), Sydney blue gums (E.
saligna; both non-native, plantation trees), and jarrah. It was expected that conditioning
process of soil by the three plant species would be primarily driven by litter deposition,
thus influenced by litter characteristics of each species in particular. Pines and eucalypts
both produce recalcitrant litter, nonetheless, differences in conditioning processes occur

85
between the two genera associated with changes in the soil chemistry and biology
owing to differences in litter quality. Conditioning of soil by pines via litter deposition
includes acidification of soil, alteration of concentrations such as N, K, Mg, and Ca, and
low microbial activity (Feller 1983; Lavelle & Spain 2003; Ehrenfeld et al. 2005). In
contrast, allelopathy has been described as a key mechanism driving soil conditioning
by eucalypts. Litter extracts from eucalypts inhibit the germination of a diversity of
plant species as well as the growth of microorganisms such as nitrifying bacteria
(Bezuidenhout & Laing 2006; Kohli et al. 2006). For this investigation it was predicted
that the plant species (pine, jarrah and blue gum) would condition the soil in a species-
specific manner (Aponte et al. 2013), however, differences in soil conditioning would
be more marked after pine than eucalypt growth since the former presents litter with
composition that is different from eucalypts. In contrast, differences in conditioning
between blue gums and jarrah would be less distinct due to similar litter quality given
that both belong to the same family.
There is the potential for pines and eucalypts to condition soils differently but few tests
of the influence of the conditioned soils on plant growth, and in particular how
conditioning of forest soil by non-native species will influence the growth of native
species such as jarrah via soil feedbacks. This issue was addressed in the current study.
Jarrah displays a high degree of growth plasticity in response to resource availability
and the soil environment (Bleby et al. 2009). This plasticity suggests that E. marginata
is capable of producing constant biomass even under variable soil conditions. Therefore,
the second specific aim was to test PSF associated with jarrah plants grown in soils
conditioned by non-native plants by comparing their growth in soils conditioned by the
same species. I predicted that species-specific conditioning of soil by non-native plants
would little affect the biomass produced by jarrah seedlings (i.e. a neutral PSF would be
observed).

86
MATERIALS AND METHODS
Sampling site and experimental design
Soils for the experiment were collected from a site that had been cleared of jarrah forest,
then mined for bauxite, and then planted with diverse native and non-native eucalypt
species, pines and cypresses to create an arboretum in 1976 (i.e. 35 years prior to
sampling). Soil samples were collected in June 2011. At the site, six pines (P. radiata)
(Pinaceae), six jarrah (E. marginata) and six blue gums (E. saligna; both Myrtaceae)
were selected at random and four intact soil cores (0.785 L; diameter = 10 cm, depth =
10 cm) were taken from beneath the canopy area of each tree where litter is deposited.
Two cores were used to measure the physico-chemical and biological properties of soils
to determine species-specific conditioning (Figure 1A). These soils were sieved (2-mm
mesh) and stored at 4 °C pending analyses, and a subsample of soil was frozen (−20 °C)
and then subsequently freeze-dried pending phospholipid-fatty acid analysis (PLFA).
The two remaining cores were placed in bottom-sealed pots (1.2 L; diameter = 13.3 cm,
height = 12.4 cm) ready to use in the pot experiment.
Pots containing the intact soil cores were placed into a root cooling tank that was
housed in a glasshouse with temperatures ranging between 4.5 and 27.8 °C for the
duration of the experiment. To produce seedlings, seeds were surface-disinfected with
ethanol (70%) and bleach (50%) to prevent growth of pathogens and germinated at 14
°C in darkness. Several jarrah seedlings were planted in each pot, and once a plant
established, the superfluous seedlings were removed leaving only one plant per pot. As
a result there were jarrah seedlings growing in soil cores collected in the field from
jarrah trees (i.e. home soils) and in soil cores collected from pines or blue gums (i.e.
away soils). Pots were rotated within the tank every two weeks to negate a block effect
and watered once a week to 60% water-holding capacity. After four months growth

87
(from June to October 2011), plants were harvested; shoots and roots were removed,
oven-dried for one week at 60 °C and weighed. Plant biomass produced in home and
away soils was compared to determine individual PSF (magnitude and direction; Figure
1B).
Figure 1. Experimental design where intact soil cores were collected from beneath canopy of pines, jarrah and blue gums. The sampling site was mined for bauxite and planted with the pines, jarrah and blue gums 35 years earlier. One half of the cores were used to determine the properties conditioned in the field by the three tree species for 35 years by measuring their physico-chemical and biological soil properties (A). The second half of the soil cores were planted with jarrah seedlings in a home (i.e. conditioned by jarrah) and away (i.e. conditioned by pines or blue gums) arrangement. After four months, plants were harvested; the total biomass produced was measured and compared in home with respect to away soils to determine individual PSF (B).
Response variables
Refer to chapter two for methodologies used to determine response variables measured.
Statistical analyses
To determine the evidence for species-specific conditioning of field soils (Figure 1A)
the physico-chemical and biological properties (including PLFAs, CLPPs and nematode
trophic groups) of soils were compared using multivariate and univariate permutational
analyses of variance (PERMANOVA) followed by pair-wise tests. Although
PERMANOVA is a multivariate method, it can be used for univariate analyses. To
B
Intact soil cores conditioned by plant species in the field
Pines Plant-soil feedback
Biomass production home vs.
away soils
Jarrah seedlings
Jarrah Blue gums
Properties of field-conditioned
soils
A
Pine soil cores
Jarrah soilcores
Blue gum soil cores

88
perform PERMANOVA using one response variable and Euclidean distance yields
Fisher’s traditional univariate F-statistic (Anderson 2010). Scaling plots from principal
co-ordinate analysis (PCO; Anderson 2001) were built to visualise the results of
PERMANOVA analyses.
To determine individual PSF (i.e. magnitude and direction; Figure 1B), I used univariate
PERMANOVAs with total, shoot and root biomass produced in home vs. away soil as
response variables. A significant increase of the plant biomass production in home with
respect to away soil was considered a positive PSF. A significant decrease of the plant
biomass production in home soil with respect to away soil was considered a negative
PSF. No difference in biomass production in home with respect to away soil was
considered a neutral PSF.
For all the statistical analyses PRIMER v6 +PERMANOVA (PRIMER-E Ltd) was
used. For the multivariate analyses, permutation of residuals under a reduced model
was selected as permutation method as well as type III (partial) sum of squares. The
unrestricted permutation of raw data as permutation method and type III (partial) sum
of squares were used for the univariate analyses. In all cases data were transformed
when necessary and Euclidean distance was used to build resemblance matrices, except
for the analyses of CLPPs and the trophic groups of the nematode community when
Bray-Curtis coefficient and Manhattan distance were respectively utilized to build the
similarity matrices.
RESULTS
Properties of soils conditioned in the field by pines, jarrah, and blue gums
The physico-chemical properties of soils conditioned in the field were significantly
different according to plant species (F2, 26 = 2.83, P = 0.003; Figure 2a). Pair-wise
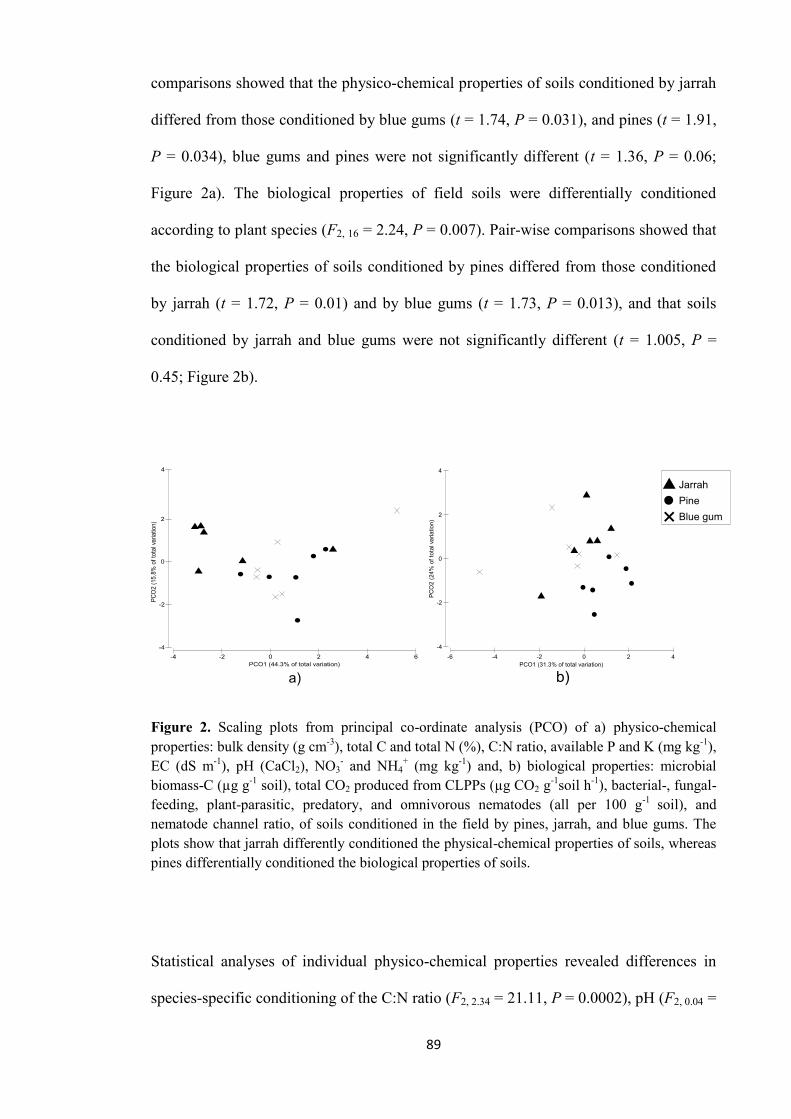
89
comparisons showed that the physico-chemical properties of soils conditioned by jarrah
differed from those conditioned by blue gums (t = 1.74, P = 0.031), and pines (t = 1.91,
P = 0.034), blue gums and pines were not significantly different (t = 1.36, P = 0.06;
Figure 2a). The biological properties of field soils were differentially conditioned
according to plant species (F2, 16 = 2.24, P = 0.007). Pair-wise comparisons showed that
the biological properties of soils conditioned by pines differed from those conditioned
by jarrah (t = 1.72, P = 0.01) and by blue gums (t = 1.73, P = 0.013), and that soils
conditioned by jarrah and blue gums were not significantly different (t = 1.005, P =
0.45; Figure 2b).
Figure 2. Scaling plots from principal co-ordinate analysis (PCO) of a) physico-chemical properties: bulk density (g cm-3), total C and total N (%), C:N ratio, available P and K (mg kg-1), EC (dS m-1), pH (CaCl2), NO3
- and NH4+ (mg kg-1) and, b) biological properties: microbial
biomass-C (µg g-1 soil), total CO2 produced from CLPPs (µg CO2 g-1soil h-1), bacterial-, fungal-feeding, plant-parasitic, predatory, and omnivorous nematodes (all per 100 g-1 soil), and nematode channel ratio, of soils conditioned in the field by pines, jarrah, and blue gums. The plots show that jarrah differently conditioned the physical-chemical properties of soils, whereas pines differentially conditioned the biological properties of soils.
Statistical analyses of individual physico-chemical properties revealed differences in
species-specific conditioning of the C:N ratio (F2, 2.34 = 21.11, P = 0.0002), pH (F2, 0.04 =

90
3.88, P = 0.042) and K concentration (F2, 37.7 = 18.2, P = 0.001). The C:N ratio of soils
conditioned by the three species were all significantly different (jarrah versus pine t =
6.02, P = 0.0016; jarrah versus blue gum t = 3.52, P = 0.0081; pine versus blue gum t =
3.24, P = 0.0113; Figure 3a). Soil pH conditioned by blue gums statistically differed
from pH of soils conditioned by pines (t = 2.78, P = 0.024; Figure 3b), and K
concentration of soils conditioned by the three species were statistically different from
each other (jarrah versus pine t = 2.63, P = 0.042; jarrah versus blue gum t = 5.14, P =
0.004; pine versus blue gum t = 3.93, P = 0 004; Figure 3c).
Statistical analyses of individual biological properties revealed species-specific
conditioning of the numbers of fungal-feeding nematodes (F2, 1904 = 3.77, P = 0.03),
omnivorous nematodes (F2, 5219 = 4.11, P = 0.039), and the nematode-channel ratio (F2,
0.06 = 4.6, P = 0.02). The numbers of fungal-feeding nematodes significantly differed in
soils conditioned by pines as compared with soils conditioned by jarrah (t = 2.41, P =
0.029; Figure 3d) and by blue gums (t = 2.21, P = 0.036). The numbers of fungal-
feeding nematodes present in soils conditioned by the two eucalypt species were not
significantly different from one another (t = 0.49, P = 0.76). The number of omnivorous
nematodes significantly differed in soils conditioned by pines with respect to soils
conditioned by jarrah (t = 3.05, P = 0.015; Figure 3e), but not with respect to soils
conditioned by blue gums (t = 1.91, P = 0.07). The numbers of omnivorous nematodes
present in soils conditioned by the two eucalypt species were not significantly different
(t = 0.69, P = 0.47). The nematode channel ratio conditioned by pines was different to
that conditioned by jarrah (t = 2.78, P = 0.033; Figure 3f) and by blue gums (t = 2.54, P
= 0.0188 respectively). Soil conditioned by pines presented the highest nematode
channel ratio.
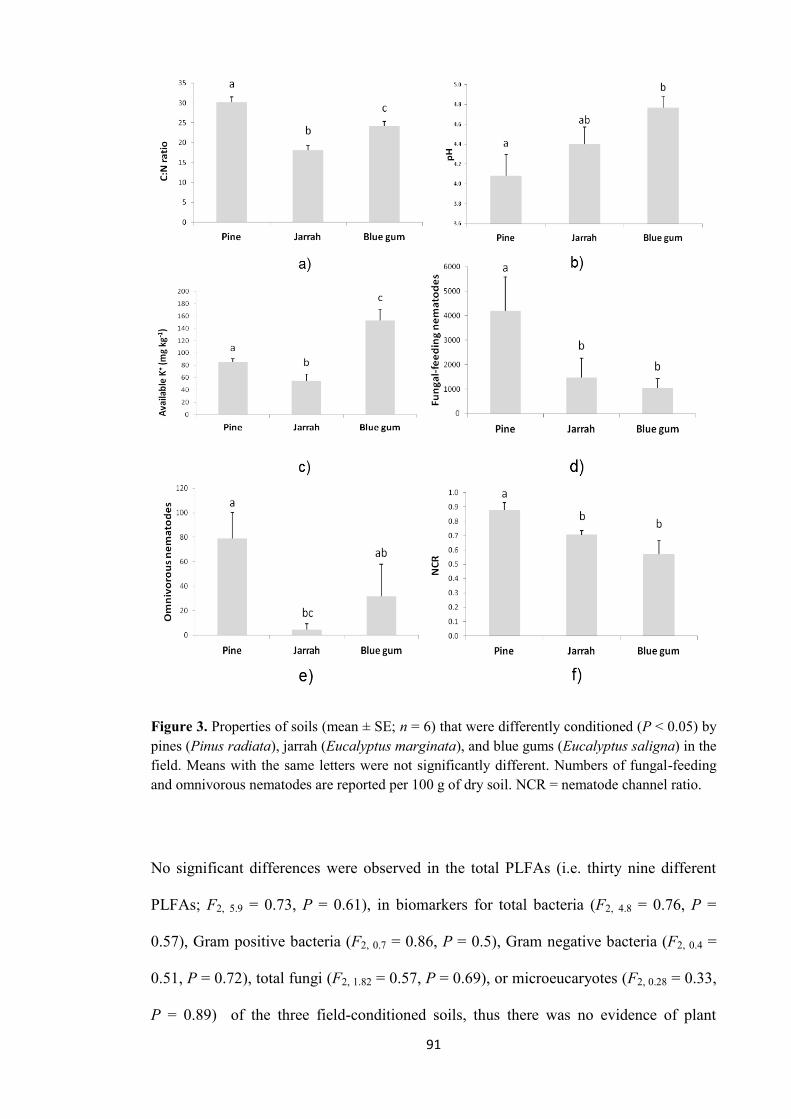
91
Figure 3. Properties of soils (mean ± SE; n = 6) that were differently conditioned (P < 0.05) by pines (Pinus radiata), jarrah (Eucalyptus marginata), and blue gums (Eucalyptus saligna) in the field. Means with the same letters were not significantly different. Numbers of fungal-feeding and omnivorous nematodes are reported per 100 g of dry soil. NCR = nematode channel ratio.
No significant differences were observed in the total PLFAs (i.e. thirty nine different
PLFAs; F2, 5.9 = 0.73, P = 0.61), in biomarkers for total bacteria (F2, 4.8 = 0.76, P =
0.57), Gram positive bacteria (F2, 0.7 = 0.86, P = 0.5), Gram negative bacteria (F2, 0.4 =
0.51, P = 0.72), total fungi (F2, 1.82 = 0.57, P = 0.69), or microeucaryotes (F2, 0.28 = 0.33,
P = 0.89) of the three field-conditioned soils, thus there was no evidence of plant
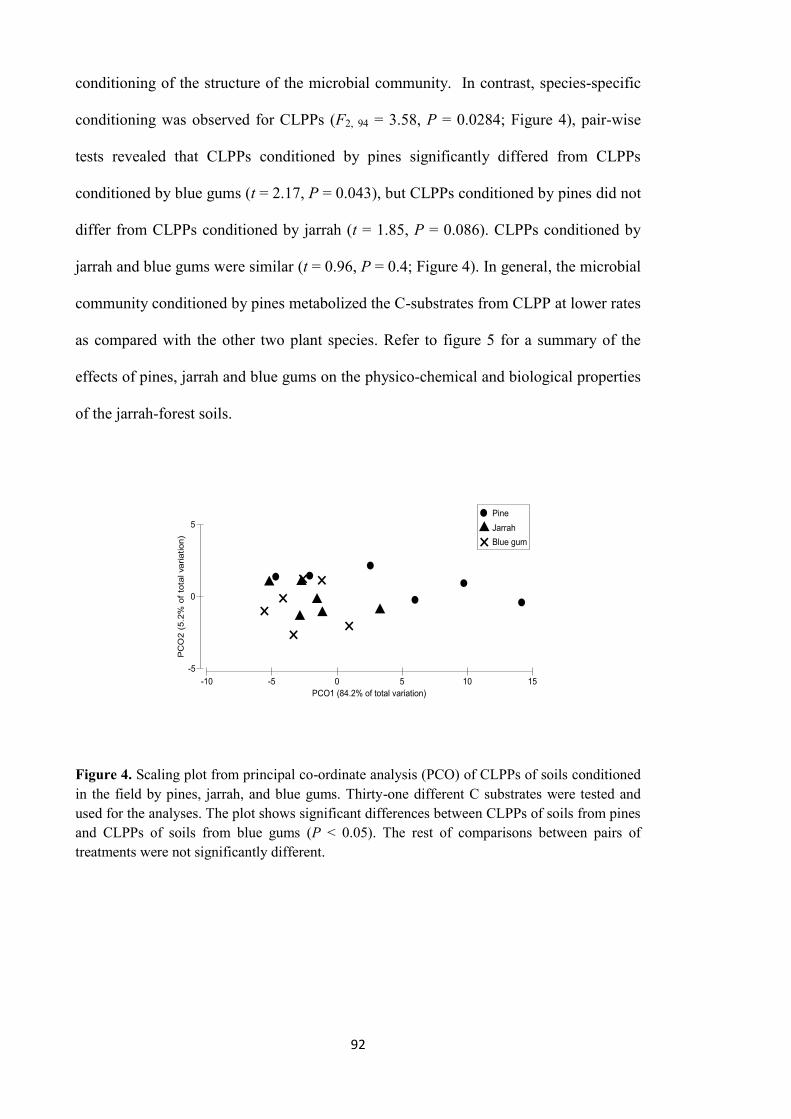
92
conditioning of the structure of the microbial community. In contrast, species-specific
conditioning was observed for CLPPs (F2, 94 = 3.58, P = 0.0284; Figure 4), pair-wise
tests revealed that CLPPs conditioned by pines significantly differed from CLPPs
conditioned by blue gums (t = 2.17, P = 0.043), but CLPPs conditioned by pines did not
differ from CLPPs conditioned by jarrah (t = 1.85, P = 0.086). CLPPs conditioned by
jarrah and blue gums were similar (t = 0.96, P = 0.4; Figure 4). In general, the microbial
community conditioned by pines metabolized the C-substrates from CLPP at lower rates
as compared with the other two plant species. Refer to figure 5 for a summary of the
effects of pines, jarrah and blue gums on the physico-chemical and biological properties
of the jarrah-forest soils.
Figure 4. Scaling plot from principal co-ordinate analysis (PCO) of CLPPs of soils conditioned in the field by pines, jarrah, and blue gums. Thirty-one different C substrates were tested and used for the analyses. The plot shows significant differences between CLPPs of soils from pines and CLPPs of soils from blue gums (P < 0.05). The rest of comparisons between pairs of treatments were not significantly different.
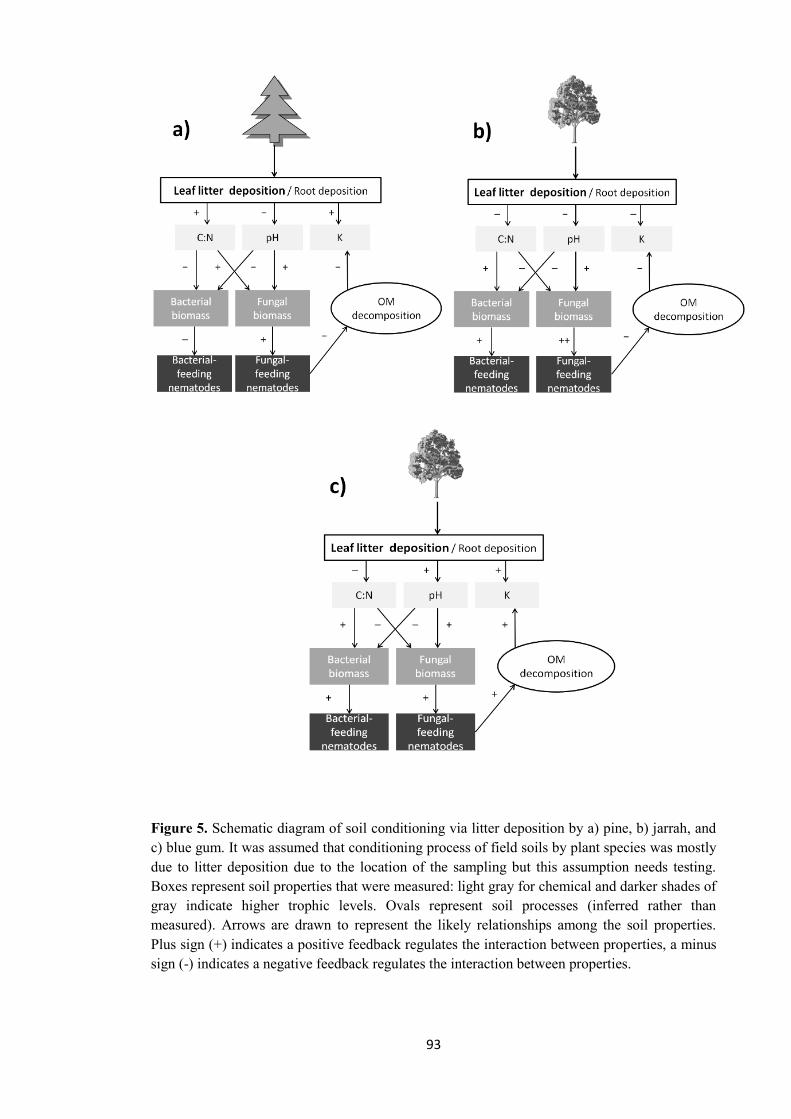
93
Figure 5. Schematic diagram of soil conditioning via litter deposition by a) pine, b) jarrah, and c) blue gum. It was assumed that conditioning process of field soils by plant species was mostly due to litter deposition due to the location of the sampling but this assumption needs testing. Boxes represent soil properties that were measured: light gray for chemical and darker shades of gray indicate higher trophic levels. Ovals represent soil processes (inferred rather than measured). Arrows are drawn to represent the likely relationships among the soil properties. Plus sign (+) indicates a positive feedback regulates the interaction between properties, a minus sign (-) indicates a negative feedback regulates the interaction between properties.
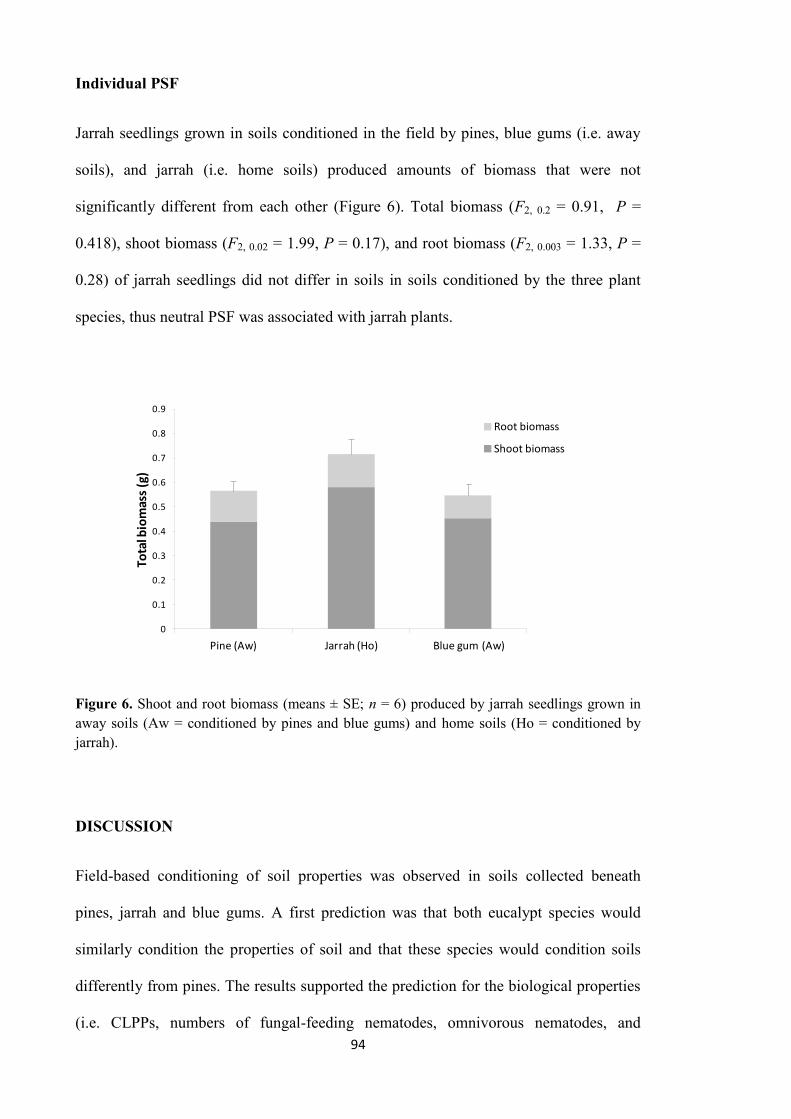
94
Individual PSF
Jarrah seedlings grown in soils conditioned in the field by pines, blue gums (i.e. away
soils), and jarrah (i.e. home soils) produced amounts of biomass that were not
significantly different from each other (Figure 6). Total biomass (F2, 0.2 = 0.91, P =
0.418), shoot biomass (F2, 0.02 = 1.99, P = 0.17), and root biomass (F2, 0.003 = 1.33, P =
0.28) of jarrah seedlings did not differ in soils in soils conditioned by the three plant
species, thus neutral PSF was associated with jarrah plants.
Figure 6. Shoot and root biomass (means ± SE; n = 6) produced by jarrah seedlings grown in away soils (Aw = conditioned by pines and blue gums) and home soils (Ho = conditioned by jarrah).
DISCUSSION
Field-based conditioning of soil properties was observed in soils collected beneath
pines, jarrah and blue gums. A first prediction was that both eucalypt species would
similarly condition the properties of soil and that these species would condition soils
differently from pines. The results supported the prediction for the biological properties
(i.e. CLPPs, numbers of fungal-feeding nematodes, omnivorous nematodes, and
0
0.1
0.2
0.3
0.4
0.5
0.6
0.7
0.8
0.9
Pine (Aw) Jarrah (Ho) Blue gum (Aw)
Tota
l bio
mas
s (g
)
Root biomass
Shoot biomass

95
nematode channel ratio), these soils were similarly conditioned by both eucalypt species
and conditioning by these trees differed from conditioning of soil by pines.
Nevertheless, the initial prediction was not supported by results of the physico-chemical
properties because these were differentially conditioned by the two eucalypt species. In
contrast, blue gums and pines similarly conditioned the physico-chemical properties of
soils. Especially relevant were the differences in conditioning of the properties C:N
ratio, pH, and available K. Despite the differences, species-specific conditioning of soil
did not translate into differences in the amounts of biomass produced by jarrah
seedlings and neutral PSF was observed as predicted. This result suggests that restoring
native jarrah trees to soils that have been conditioned by non-native species for decades
may not be as problematic as envisaged.
It was assumed that differences in properties of field-conditioned soils grown with the
three plant species were mainly the result of conditioning via leaf litter deposition rather
than root deposition. Jarrah trees produce a tap root system, from which lateral roots
and sinker roots emerge. Soil samples were taken from old tress (35-years-old), whose
roots are well developed (Szota et al. 2007). Because root exudation occurs mainly in
root hairs from younger roots (Bertin et al. 2003), it was assumed that in the first 10 cm
deep of soil, from which samples were collected, the influence of root deposition was
minimal. Moreover, soil samples were collected from beneath tree canopy where litter
deposition has occurred for 35 years. consequently, the main mechanism of
conditioning observed for these trees in the first 10 cm of soil was via leaf litter.
Conditioning of soil via litter deposition is directly related to the chemical composition
of litter (Hobbie 1992; Guo & Sims 1999; Berg & McClaugherty 2008; Witt & Setälä
2010; Uselman et al. 2012). For example, litter with high C:N ratio causes an increase
in the C:N ratio of soil and vice versa. Soils collected from beneath pines can have
higher C:N ratios (i.e. 28 – 36; O’Brien et al. 2003; Hopmans & Elms 2009) than soils

96
collected from beneath eucalypts (i.e. 14–23; Adams & Attiwill 1986; O’Brien et al.
2003) in response to differences in litter composition. These results are in keeping with
reports for soils previously collected from pines (i.e. C:N of 30), whereas soils collected
from jarrah and blue gums sites presented a C:N ratio within the range of 20–25 that
was slightly higher than the value reported in the literature.
Generally, litter with C:N ratios between 20 and 30 supply the necessary nutritional N-
requirements for soil microbes avoiding immobilization by microbial biomass, which in
turn, can limit plant growth (Foth 1990; Lavelle & Spain 2003; Johnson 2009; Hopkins
et al. 2010). Since soils collected from pines had the highest mean C:N ratio (i.e. 30) of
all soils, it was expected for the microorganisms inhabiting these soils to be more N-
limited as compared with those inhabiting soils conditioned by eucalypts. The results
supported this assertion; soils conditioned by pines presented the lowest microbial
respiration rates (from CLPPs) of the three field-conditioned soils, which indicated a
low microbial activity. Chemical analyses of soil revealed that the amounts of total N,
NO3–, and NH4
+ did not significantly differ in soils conditioned by pines from soils
conditioned by jarrah or blue gums. Therefore, N limitations in pine soils may be a
result of inaccessibility to N rather than low concentration of the nutrient. Pines can
release polyphenols that bind to organic N, then pines can utilize this N that is
inaccessible to other plants and microbes (Ehrenfeld et al. 2005). In soils conditioned by
eucalypt species, lower soil C:N ratios translated into higher rates of respired-C, which
indicated that soil microbes were less limited by N resources and were metabolically
more active than microbes in soils conditioned by pines.
The addition of leaf and woody litter to soil can cause soil pH to change due to release
of base cations, organic anions and N during litter decomposition (Bhupinderpal-Singh
& Rengel 2007; Berg & McClaugherty 2008). Due to species-specific litter chemistry,

97
changes in soil pH driven by litter deposition are also species-specific. The three plant
species used in the experiment produced changes in soil pH of different magnitude.
Soils collected from beneath pines presented the lowest pH of all soils (i.e. 4.0), which
is consistent with information reporting acidification of soil by this species (Feller 1983;
Laskowski et al. 1995; Lavelle & Spain 2003). The results also indicated that pH of
soils conditioned by jarrah and blue gums were less acidic than soils conditioned by
pines. Both eucalypt species conditioned soil pHs that were significantly different to
each other (i.e. pH = 4.5 for jarrah, and pH = 5 for blue gums) but that fall within the
range reported for eucalypt species (i.e. pH range of 3.8 – 6; Feller 1983; Adams &
Attiwill 1986; O’Brien et al. 2003; Turner & Lambert 2008; Spain et al. 2014). Species-
specific conditioning of soil pH by plants can, in turn, drive differential chemical and
biological properties of soil (Jones 1998; Uren 2001; Lavelle & Spain 2003; Bais et al.
2008; Richardson et al. 2009; Madigan et al. 2012). Overall, differences in litter quality
may ultimately impact structural and functional aspects of soil grown with pines, jarrah
and blue gums.
The structure of the microbial communities did not differ among the field-conditioned
soils. However, these findings could be a result of differential populations of microbial
predators, as the numbers of fungal-feeding and omnivorous nematodes were
significantly higher in soils conditioned by pines than in soils conditioned by the
eucalypt species. Increased populations of fungal-feeding and omnivorous nematodes
indirectly indicated an increased fungal biomass produced in soils conditioned by pines.
Soils conditioned by the two eucalypt species presented lower numbers of fungal-
feeding nematodes that were associated with higher pHs and lower C:N ratios as
compared with soil conditioned by pines. Understanding the interactions among the soil
food web is critical to being able to interpret these data.

98
The results for the nematode channel ratio indicated that an energy channel of organic
matter decomposition driven by the fungal compartment was operating in soils
conditioned by pines (i.e. nematode channel ratio = 0.88), which probably reflects the
recalcitrance of pine litter (Moore & Hunt 1988; Moore et al. 2003; Condron et al.
2010; Witt & Setälä 2010). Autochthonous fungi possess the unique ability to
decompose recalcitrant organic matter in a variety of ecosystems at slow rates (Lavelle
& Spain 2003; de Boer et al. 2005; Berg & McClaugherty 2008; Condron et al. 2010).
Both the bacterial- and fungal-based energy channels were operating in soils
conditioned by blue gums (i.e. nematode channel ratio = 0.57). Since the bacterial-based
energy channel is faster than the fungal one, quicker rates of organic matter
decomposition were occurring in soils conditioned by blue gums, probably in response
to the labile litter (Moore & Hunt 1988; Moore et al. 2003; Yeates 2003; Condron et al.
2010; Witt & Setälä 2010). Finally, the fungal-based energy channel was favoured over
the bacterial-based channel in soils conditioned by jarrah (i.e. nematode channel ratio =
0.7), which indicated a rate of organic matter decomposition that is intermediate
between pines and blue gums. Differences in organic matter decomposition rates
occurring in soils conditioned by the three plant species may have implications on soil
processes and ultimately on plant community structure and evolution (Hobbie 1992;
Eviner 2004; Weidenhamer & Callaway 2010).
Potassium is a cation that plays key roles in plant metabolism such as enzyme
activation, protein synthesis, photosynthesis, osmoregulation, cell extension, stomatal
movement, and phloem transport (Marschner 1995). Potassium is deposited in soil by
leaching and/or catabolic activity of microorganisms during decomposition of litter
(Ganjegunte et al. 2005; Berg &McClaugherty 2008). Amounts of K in soils are
expected to vary for two main reasons: the K concentrations present in fresh litter and
the rate of litter decomposition. Soils conditioned by blue gums, pines and jarrah,

99
presented the highest, intermediate, and the lowest concentrations of K respectively.
Litter from blue gums is reported to contain medium to high concentrations of K (i.e.
4.39–7.35 g kg-1; Lugo et al. 1990; Binkley & Ryan 1998), which might be quickly
released to soil due to an increased rate of litter decomposition (i.e. as suggested by the
lowest nematode channel ratio value of the three plant species). Litter from jarrah is
reported to contain lower K concentrations (i.e. 2.6 – 5.7 g kg-1; O’Connell et al. 1978;
Hingston et al. 1981), and according to the nematode channel ratio, litter from jarrah
might decompose at lower rates. These two aspects translated into low concentrations of
K in soils conditioned by jarrah plants. Litter from pines is reported to contain high K
concentrations (i.e. 7.31 g kg-1; Hopmans & Elms 2009), but the decomposition rate of
this litter type is slow (i.e. as suggested by the highest nematode channel ratio). As a
result, K concentrations in soil grown with pines were not as high as those conditioned
by blue gums.
The differences in the properties of soils conditioned in the field did not affect
individual PSF (i.e. magnitude or direction) and neutral PSF was observed for jarrah
seedlings during the pot experiment. This finding agreed with the initial prediction of
neutral PSF associated with jarrah. Previous work demonstrated that jarrah shows a high
aboveground architectural plasticity in response to varying environmental conditions
(Bleby et al. 2009). The results of this research suggest this plasticity is not exclusive to
the aboveground compartment, but to the belowground compartment as well, and is not
exclusive to morphological aspects but also to functional aspects. For that reason, jarrah
plants are able to maintain a stable biomass production despite the changing soil
properties, even those conditioned by different plant species. This ability is evident in
the field where jarrah establishes and grows well in non-disturbed but also in soil post-
bauxite mining (Koch & Samsa 2007), which presents modified properties due to
mining and restoration processes (i.e. reduced biological activity, pH, and C, and

100
increased P, N, K and gravel percentage; Ward 2000; Jasper 2007; George et al. 2010;
Tibbett 2010). Plant species displaying plastic growth responses including neutral PSFs
such as jarrah are ideal to use during restoration strategies. This trait allows restored
plants to establish, grow, and persist in restored sites.
In conclusion, non-native plant species can condition soils in ways that differ to native
species, which has implications for ecosystem functioning. This is critical for the
restoration of human-modified ecosystems, since the ultimate goal of most restoration
activities is a complete functional ecosystem. In restored jarrah forest post-bauxite
mining, despite alterations to soil properties are caused by non-native species, jarrah
seedlings were able to establish in these conditioned soils, without it affecting their
biomass production. This suggests that the jarrah–soil system displays a plastic PSF, a
trait of jarrah that might contribute to its persistence for long time spans, and to adapt to
modified soil conditions present in restored soil where jarrah establishes and grows
well.

101
CHAPTER 6
Concluding discussion
This thesis contributed to understand the influence of plant species (Chapter 3 & 5) and
soil conditions (Chapters 4 & 5) on PSF operating in jarrah forest. Plant-soil feedback
was examined in the contexts of unmined soil and restored soil post-bauxite mining.
Plant species of the jarrah forest contribute to PSF by conditioning the properties of soil
in species-specific manner (Chapter 3). The three plant species tested - Eucalyptus
marginata, Acacia pulchella, and Bossiaea ornata, conditioned the soil via changes to
chemical properties (Chapter 3; Figures 2 & 3, Chapter 4; Figure 2). Species-specific
conditioning of biological properties was evidenced only in soils from the field (Chapter
4; Figures 3 & 4), not in soils conditioned in the glasshouse. This finding indicated that
when studying PSF phenomena, the experimental approach used (‘natural experiment
approach’ vs. ‘cultivation experiment approach’; Kulmatiski & Kardol 2008; Pernilla et
al. 2010; van de Voorde et al. 2012) is a critical aspect of the research.
How jarrah forest plants condition the soil is likely to influence their establishment and
persistence in the field. For example, plant species such as E. marginata and B. ornata
conditioned the concentration of P efficiently when grown in unmined soil (Chapter 3;
Table 1). The conditioning capacity of soil P might be related to the long persistence of
both plant species (Abbott 1984; Bell 2001) given the low concentration of this nutrient
in jarrah forest soil (Hingston et al. 1989). Conditioning capacity of P was not altered
when the plants grew in restored soil (Chapter 3; Table 2), which suggests that
conditioning of P is a preserved plant trait that might help maintaining the flux of
nutrients in the ecosystem (Eviner & Hawkes 2008). Therefore after mining, restoration
of long-lived species such as E. marginata and B. ornata might be critical for the re-
establishment of the whole ecosystem functioning.

102
Acacia pulchella showed an inefficient capacity to condition P of soil, either unmined
or restored (Chapter 3; Tables 1 & 2). This trait might contribute to this species being
short-lived, A. pulchella populations decline four years after a disturbance such as fire
(Monk et al. 1981). Acacia pulchella conditioned the total N of unmined soil, through
nitrogen-fixation which is a trait generally recognised for its improvement of the soil
fertility. However, this conditioning capacity was affected by modified soil conditions
present in restored soil (Chapter 3; Tables 1 & 2). Hence it is likely soil functions
related to the N cycle could be affected, as previously suggested by studies that have
determined a conservative N cycle is developing in restored jarrah forest (i.e. 26 years
post-mining; Banning et al. 2008). A conservative N cycle developing in the ecosystem
implies that critical functions are recovering slowly.
Species-specific conditioning of soil by jarrah forest plants is likely to have major
influence on soil functioning. In unmined soil, E. marginata and B. ornata conditioned
higher C:N ratios (as compared with unplanted controls), whereas A. pulchella
conditioned lower C:N ratios (Chapter 3; Table 1). The results indicated that organic
matter decomposition is occurring at slower rates in soils influenced by E. marginata
and B. ornata, whereas faster decomposition rates occur in soils influenced by A.
pulchella. These patterns were not observed in modified soil (Chapter 3; Table 2), the
C:N ratios of restored soils grown with the three plant species did not differ from those
of unplanted controls, which suggests that altered patterns of conditioning can have
additional impacts on soil functioning.
Individual PSF associated with the plants used in this study was soil-context dependent
(De Deyn et al. 2004; Chapter 3; Figure 4). The results suggest that PSF is a
phenomenon that exhibits some level of plasticity. Field observations support my
findings, in that a neutral PSF associated with B. ornata might contribute to its

103
abundance and long-persistence in unmined soil. In contrast, this plant species shows
variable abundance in restored soil (Pang et al. 2010), which could be due, in part, to
the expression of negative PSF. Acacia pulchella was very responsive to changing soil
conditions (via soil feedback), and specifically to fertilisation that is part of restoration
practice. Despite the negative PSF observed in the experiment, A. pulchella produced up
to three times more biomass in restored soil as compared with unmined soil, which
agrees with the dominance of this plant species observed in early-restored sites. These
results have implications for the restoration of the jarrah forest, since altered PSF might
produce altered patterns of plant abundance and dominance.
Results from Chapter 4, supported the finding of soil conditions (the soil feedback)
shaping individual PSF associated to B. ornata seedlings. The biomass produced by this
plant species grown in soils from the chronosequence of restoration ages was
significantly influenced by soil age (time since restoration), but not by plant species
from which soils were collected in the field. Properties of soils from the
chronosequence approached those of unmined soil as restoration age increased, which
suggested a recovery of the soil feedback (Chapter 4; Figure 4). However, a differential
soil feedback effect was observed as variable amounts of biomass produced by B.
ornata. Particularly important was the positive soil feedback associated with the 22-
year-old soil (Chapter 4; Figure 6), where the biggest plants were produced as compared
with unmined soil. The increased biomass produced in the 22-year old soil might
respond to the absence of plant-parasitic nematodes (as compared with unmined soils),
and the increased numbers of bacterial-feeding nematodes (Chapter 4; Table 1). These
results suggest that decreased or increased numbers of key soil organisms contribute to
alter the patterns of plant dominance and abundance in restored jarrah forest. The
differential soil feedback observed in soils from the chronosequence of restoration ages
did not affect individual PSF associated with B. ornata, and neutral PSF was observed

104
in all soils (Chapter 4; Figure 8). The fact that B. ornata produced similar amounts of
biomass in home soils as compared with away soils reinforce the hypothesis of a PSF
influenced by soil conditions, and not by plant species conditioning of the soil.
Furthermore, the results described in Chapters 3 & 4, suggest that individual PSF
associated with representative jarrah forest plant species is driven by soil conditions, not
by plant species. However, PSF associated with jarrah (E. marginata) was the
exception. Different soil properties conditioned in the field by non-native species
(different contexts of soil; Chapter 5; Figures 2 & 3) did not affect individual PSF, and
neutral PSF was observed for jarrah seedlings in the pot experiment (Chapter 5; Figure
6). This finding suggested that PSF associated with jarrah is mainly driven by plant
species, and exhibits some level of plasticity (Bleby et al. 2009), which allows the plant
to maintain a stable biomass production despite changing contexts of soil. Field
observations support this assertion, jarrah establishes and grows well in unmined but
also in soil post-bauxite mining (Koch & Samsa 2007). A plastic PSF (neutral PSF) and
efficient conditioning capacity might contribute to the long persistence of jarrah in the
forest, and moreover, these traits might contribute to its dominance in the ecosystem. In
the context of restoration, plant species displaying plastic growth responses including
neutral PSF such as jarrah are ideal to use during restoration strategies because this trait
allows restored plants to establish, grow, and persist in restored sites.
Neutral PSF associated to the experimental plants used (i.e. long-lived and short-lived
plant species) during the three experiments (Chapters 3, 4 & 5) suggests that biomass
production of jarrah forest plants might be little affected by changing properties of soils.
This finding contradincts the accepted PSF theory, which indicates that negative PSF is
associated to plants grown in early-successional soils, whereas positive PSF is
associated to late-successional soils (Kardol et al. 2006). It is important to highlight that

105
most of the ecosystems studied from a PSF perspective follow a relay floristics model
of succession, (i.e. an orderly and predictable series of plant species replacements that
occur after a disturbance; Cattelino et al. 1979). The traditional PSF theory fits very
well within this model of succession, since PSF phenomena help explaining the orderly
series of plant replacement.
In the jarrah forest, plant succession follows an initial floristic model of succession that
describes how the initial plant community evolves only as changes in the abundance of
the species present in the landscape (Hobbs & Legg 1983; Norman et al. 2006) in
response to disturbances such as fire (Bell & Koch 1980). It is likely that PSF operating
in jarrah forest occurs differently as compared with other ecosystems so as to secure
plant persistence in the community. This could happen because the species that establish
early in succession are those that are still present later. The findings of my three
experiments fit very well within the initial floristic model of succession, since in order
to persist in the ecosystem, jarrah forest plants must condition properties of soil
efficiently, a plant trait that was observed during the experiments. Moreover, soil is an
evolving entity, soil properties change through time and these influence the patterns of
abundance and dominance of plant species. Therefore, a neutral PSF associated to jarrah
forest plants help explaining their persistence in the field, because jarrah forest plants
are able to maintain a stable biomass production despite changing soil properties.
Individual PSF associated to plant species was altered in restored soil, so it is likely the
patterns of dominance and abundance of plant species are modified in restored forest as
compared with unmined forest, with implications on the structure of the plant
community composition of restored forest.

106
REFERENCES
Abbott, L.K., Robson, A.D., 1977. The distribution and abundance of vesicular arbuscular endophytes in some western Australian soils. Aust J Bot 25:515-522.
Abbott, I. 1984, Comparisons of spatial pattern, structure, and tree composition between virgin and cut-over jarrah forest in Western Australia. For Ecol Manage 9:101-126.
Adams, M.A., Attiwill, P.M., 1986. Nutrient cycling and nitrogen mineralization in eucalypt forests of south eastern Australia II. Indices of nitrogen mineralization. Plant Soil 92:341-362.
Adl, S.M., 2003. The ecology of soil decomposition. CABI Publishing, UK.
Ahmad, I., Pichtel, J., Hayat, S. 2008. Plant-bacteria interactions. Wiley-VCH Verlag GmbH & Co. KGaA, Germany.
Alexander, R., Croft, K.D., Kagi, R.I., Shea, S., 1978. Saponins of Acacia pulchella. Aust J Chem 31: 2741-2744.
Altieri, M.A., 1999. The ecological role of biodiversity in agroecosystems. Agr Ecosyst Environ 74:19-31.
Amalesh, S., Das, G., Das, S.K., 2011. Roles of flavonoids in plants. Int J Pharm Sci Tech 6(1): 12-35.
Anderson, M.J., 2001. A new method for non-parametric multivariate analysis of variance. Austral Ecol 26:32–46.
Anderson, M.J., Gorley, R.N., Clarke, K.R., 2008. PERMANOVA+ for PRIMER: Guide to software and statistical methods. Institute of Information and Mathematical Sciences (IIMS) Massey University, Auckland.
Anderson, M.J., 2010. Detecting multivariate changes in biological assemblages: experimental design and data analysis. Multivariate analysis of complex experimental designs using PERMANOVA+ for PRIMER. Institute of Information and Mathematical Sciences (IIMS) Massey University, Auckland.
Aponte, C., García, L.V., Marañón, T., 2013. Tree species effects on nutrient cycling and soil biota: a feedback mechanism favouring species coexistence. For Ecol Manage http://dx.doi.org/10.1016/j.foreco.2013.05.035
Atkinson, D., 2006. Arbuscular mycorrhizal fungi and the form and functioning of the root system. In: Mukerji, K.G., Manoharachary, C., Singh, J. (eds) Microbial activity in the rhizosphere. Springer, Germany.
Badri, D.V., Vivanco, J.M., 2009. Regulation and function of root exudates. Plant Cell Environ 32:666-681.
Baeten, L., Vanhellemont, M., De Frenne, P., De Schrijver, A., Hermy, M., Verheyen, K., 2010. Plasticity in response to phosphorus and light availability in four forest herbs. Oecologia 163:1021-1032.
Bais, H.P., Broeckling, C.D., Vivanco, J.M., 2008. Root exudates modulate plant–microbe interactions in the rhizosphere. In: Karlovsky, P. (ed) Secondary metabolites in soil ecology. Springer, Berlin.

107
Banning, N.C., Grant, C.D., Jones, D.L., Murphy, D.V., 2008. Recovery of soil organic matter, organic matter turnover and nitrogen cycling in a post-mining forest rehabilitation chronosequence. Soil Biol Biochem 40: 2021–2031.
Banning, N.C., Lalor, B.M., Cookson, W.R., Grigg, A.H., Murphy, D.V. 2012. Analysis of soil microbial community level physicological profiles in native and post-mining rehabilitation forest: Which subtrates discriminate? Appl Soil Ecol 56:27-34.
Bardgett, D.R., Bowman, D.W., Kaufmann, R., Schmidt, K.S., 2005. A temporal approach to linking abovegroung and belowground ecology. Trends Ecol Evol 20(11):634-641.
Barriuso, J., Ramos, S.B., Lucas, J.A., Probanza, L.A., Garcia-Villaraco, A., Gutierrez, M.F.J 2008. Ecology, genetic diversity and screening strategies of plant growth promoting rhizobacteria. In: Ahmad, I., Pichtel, J.,Hayat, Shamsul (eds). Plant-bacteria interactions. Wiley-VCH Verlag GmbH & Co. KGaA, Germany.
Bell, D.T., Koch, J.M., 1980. Post-fire succession in the northern jarrah forest of Western Australia. Aust J Ecol 5:9-14.
Bell, D.T., Heddle, E.M., 1989. Floristic, morphologic and vegetational diversity. In: Dell, B., Havel, J.J., Malajczuk, N. (eds) The jarrah forest: a complex Mediterranean ecosystem. Kluwer Academic Publishers, The Netherlands.
Bell, D.T., 2001. Ecological response syndromes in the flora of southwestern Western Australia: fire resprouters versus reseeders. Bot Rev 67(4):417-440.
Bell, T.D., Hobbs, J.R., 2007. Jarrah forest ecosystem restoration: a foreword. Restor Ecol 15(4):S1-S2.
Berg, B., McClaugherty, C., 2008. Plant Litter: decomposition, humus formation, carbon sequestration. 2nd edition. Springer, Germany.
Berg, G., Smalla, K., 2009. Plant species and soil type cooperatively shape the structure and function of microbial communities in the rhizosphere. FEMS Microbiol Ecol 68:1-13.
Berg, M.P., 2010. Spatio-temporal structure in soil communities and ecosystem processes. In: Verhoef, H.A., Morin, P.J. (eds) Community Ecology: processes, models and applications. Oxford University Press, Great Britain.
Bertin, C., Yang, X., Weston, L.A., 2003. The role of root exudates and allelochemicals in the rhizosphere. Plant Soil 256:67-83.
Bever, D.J., 1994. Feedback between plants and their soil communities in an old field community. Ecology 75(7):1965-1977.
Bever, J.D., Westover, K.M., Antonovics, J., 1997. Incorporating the soil community into plant population dynamics: the utility of the feedback approach. J Ecol 85:561-573.
Bever, J.D., 2002. Host-specificity of AM fungal population growth rates can generate feedback on plant growth. Plant Soil 244:281-290.
Bever, J.D., 2003. Soil community feedback and the coexistence of competitors: conceptual frameworks and empirical tests. New Phytol 257:465-473.
Bever, J.D., Dickie, I.A., Facelli, E., Facelli, J.M., Klironomos, J., Moora, M., Rillig, M.C., Stock, W.D., Tibbett, M., Zobel, M., 2010. Rooting theories of plant community ecology in microbial interactions. Trends Ecol Evol 25:468-478.
Bever, J.D., Platt, T.G., Morton, E.R., 2012. Microbial population and community dynamics on plant roots and their feedbacks on plant communities. Annu Rev Microbiol 66:265-283.

108
Bezemer, T.M., Lawson, C.S., Hedlund, K., Edwards, A.R., Brook, A.J., Igual, J.M., Mortimer, S.M., van der Putten, W.H., 2006. Plant species and functional group effects on abiotic and microbial soil properties and plant–soil feedback responses in two grasslands. J Ecol 94:893-904.
Bezuidenhout, S.R., Laing, M., 2006. Allelopathy and its influence in soil systems. In: Uphoff, N., Ball, A.S., Fernandes, E., Herren, H., Husson, O., Laing, M., Palm, C., Pretty, J., Sanchez, P., Sanginga, N., Thies, J. (eds.) Biological approaches to sustainable soil systems. CRC Taylor & Francis Group, USA.
Bhupinderpal-Singh, Rengel, Z., 2007. The role of crop residues in improving soil fertility. In: Marschner, P., Rengel, Z. (eds.) Nutrient cycling in terrestrial ecosystems. Springer-Verlag, Berlin.
Binkley, D., Ryan, M.G., 1998. Net primary production and nutrient cycling in replicated stands of Eucalyptus saligna and Albizia facaltaria. For Ecol Manage 112:79-85.
Bleby, T.M., Colquhoun, I.J., Adams, M.A., 2009. Architectural plasticity in young Eucalyptus marginata on restored bauxite mines and adjacent natural forest in south-western Australia. Tree Physiol doi:10.1093/treephys/tpp044.
Bohn, H.L., McNeal, B.L., O’Connor, G.A. 2001. Soil Chemistry 3rd edition. John Wiley & Sons, Inc., USA.
Bonanomi, G., Mazzoleni, S., 2005. Soil history affects plant growth and competitive ability in herbaceous species. Community Ecol 6(1):23-28.
Bonanomi, G., Rietkerk, M., Dekker, S.C., Mazzoleni, S., 2005a. Negative plant–soil feedback and positive species interaction in an herbaceous plant community. Plant Ecol 181:269–278.
Bonanomi, G., Giannino, F., Mazzoleni, S., 2005b. Negative plant-soil feedback and species coexistence. Oikos 111:311-321.
Bongers, T., Bongers, M., 1998. Functional diversity of nematodes. Appl Soil Ecol 10:239-251.
Brandt, A.J., de Kroon, H., Reynolds, H.L., Burns, J.H., 2013. Soil heterogeneity generated by plant-soil feedbacks has implications for species recruitment and coexistence. J Ecol 101:277-286.
Bret-Harte, M.S., Shaver, G.R., Zoerner, J.P., Johnstone, J.F., Wagner, J.L., Chavez, A.S., Gunkelman, R.F., Lippert, S.C., Laundre, J.A., 2001. Developmental plasticity allows Betula nana to dominate tundra subjected to an altered environment. Ecology 82(1):18-32.
Brimecombe, M.J., De Leij, F.A.A.M., Lynch, J.M., 2001. The effect of root exudates on rhizosphere microbial populations. In: Pinton, R., Varanini, Z., Nannipieri, P. (eds) The rhizosphere: biochemistry and organic substances at the plant-soil interface. Marcel Dekker Editorial, USA.
Brown, M.T., 2005. Landscape restoration following phosphate mining: 30 years of co-evolution of science, industry and regulation. Ecol Eng 24:309-329.
Brundrett, M.C., Abbott, L.K., 1991. Roots of jarrah forest plants. I. Mycorrhizal associations. Aust J Bot 39:445–457.
Brundrett, M.C., Abbott, L.K., 1995. Mycorrhizal fungus propagules in the jarrah forest II. Spatial variability in inoculum levels. New Phytol 131:461-469.

109
Büdel, B., 2005. Microorganisms of biological crusts on soil surfaces. In: Buscot, F., Varma, A. (eds), 2005. Microorganisms in soil: roles in genesis and functions. Springer-Verlag, Berlin.
Burrows, N.D., 2008. Linking fire ecology and fire management in south-west Australian forest landscapes. Forest Ecol Mange 255(7):2394-2406.
Buscot, F., Varma, A. (eds), 2005. Microorganisms in soil: roles in genesis and functions. Springer-Verlag, Berlin.
Buscot, F., 2005. What are soils? In: Buscot, F., Varma, A., (eds.) Microorganisms in soil: roles in genesis and functions. Springer-Verlag, Berlin.
Caesar-TonThat, T.C., Caesar, A.J., Gaskin, J.F., Sainju, U.M., Stevens, W.B. 2008. Ecology and function of culturable microbes in soil aggregation. In: Liu, T-X (ed) Soil ecology research developments. Nova Science Publishers, New York.
Campbell, C.D., Chapman, S.J., Cameron, C.M., Davidson, M.S., Potts, J.M., 2003. A rapid microtiter plate method to measure carbon dioxide evolved from carbon substrate amendments so as to determine the physiological profiles of soil microbial communities by using whole soil. Appl Environ Microb 69:3593–3599.
Carvalhais, L.C., Dennis, P.G., Fedoseyenko, D., Hajirezaei, M.R., Borriss, R., von Wirén, N., 2011. Root exudation of sugars, amino acids, and organic acids by maize as affected by nitrogen, phosphorus, potassium, and iron deficiency. J Plant Nutr Soil Sc 174:3–11.
Castillo, P. & Vovlas, N. (2007). Pratylenchus (Nematoda: Pratylenchidae): diagnosis, biology, pathogenicityand management. Nematology Monographs & Perspectives, 6 (Series Editors: Hunt, D.J. & Perry,R.N.). Leiden, The Netherlands, Brill Academic Publishers, 555 pp.
Cattelino, P.J., Noble, I.R., Slatyer, R.O., Kessell, S.R., 1979. Predicting the multiple pathways of plant succession. Environ Manage 3(1):41-50.
Cavigelli, M.A., Robertson, P., Klug, M.J., 1995. Fatty acid methyl ester (FAME) profiles as measures of soil microbial community structure. Plant Soil 179:90-113
Cesco, S., Mimmo, T., Tonon, G., Tomasi, N., Pinton, R., Terzano, R., Neumann, G., Weisskopf, L., Renella, G., Landi, L., Nannipieri, P., 2012. Plant-borne flavonoids released into the rhizosphere: impact on soil bio-activities related to plant nutrition. A review. Biol Fertil Soils 48:123-149.
Chalkos, V.D., Karamanolo, K., 2006. Microorganisms and allelopathy: a one-sided approach. In: Reigosa, M.J., Pedrol, N., González, L. (eds) Allelopathy: A physiological process with ecological implications. Springer, The Netherlands.
Chapman, S.J., Campbell, C.D., Artz, R.R.E., 2007. Assessing CLPPs using MicroResp™, a comparison with Biolog and multi-SIR. J Soils Sediments 7(6):406–410.
Chodak, M., Pietrzykowski, M., Niklinska, M., 2009. Development of microbial properties in a chronosequence of sandy mine soils. Appl Soil Ecol 41:259-268.
Churchward, H.M., Dimmock, G.M., 1989. The soils and landforms of the northern jarrah forest. In: Dell, B., Havel, J.J., Malajczuk, N. (eds) The jarrah forest: a complex Mediterranean ecosystem. Kluwer Academic Publishers, The Netherlands.
Coleman, D.A., Crossley, D.A., Hendrix, P.F., 2004. Fundamentals of soil ecology 2nd edition. Elsevier Academic Press, Amsterdam.

110
Compan, S., Duffy, B., Nowak, J.,Clément, C., Ait Barka, E., 2005. Use of plant growth-promoting bacteria for biocontrol of plant diseases: principles, mechanisms of action, and future prospects. Appl Environ Microbiol 71(9):4951-4959.
Condron, L., Stark, C., O’Callaghan, M., Clinton, P., Huang, Z., 2010. The role of microbial communities in the formation and decomposition of soil organic matter. In: Dixon, G.R., Tilston, E.L. (eds) Soil microbiology and sustainable crop production. Springer, New York.
Conservation Commission of Western Australia, 2012. Draft forest management plan 2014 – 2023. Conservation Commission of Western Australia, Perth.
D’Souza, N.K., Colquhoun, I.J., Shearer, B.L., Hardy, G.E.S.J., 2004. The potential of five Western Australian native species for biological control of Phytophtora cinnamomi. Aust J Bot 52:267-279.
Dakora, F.D., Phillips, D.A., 2002. Root exudates as mediators of mineral acquisition in low-nutrient environments. Plant Soil 245:35-47.
De Bellis, T., Kernaghan, G., Widden, P., 2007. Plant community influences on soil microfungal assemblages in boreal mixed-wood forests. Mycologia 99(3):356-367.
de Boer, W., Folman, L.B., Summerbell, R.C., Boddy, L., 2005. Living in a fungal world: impact of fungi on soil bacterial niche development. FEMS Microbiol Rev 29:795–811.
De Deyn, B.G., Raaijmakers, E.C., van der Putten, H.W., 2004. Plant community development is affected by nutrients and soil biota. J Ecol 92:824-834.
De Rooij-van der Goes, P.C.E.M., 1995. The role of plant-parasitic nematodes and soil-borne fungi in the decline of Ammophila arenaria (L.) Link. New Phytol 129:661-669.
Deans, S.G., 2002. Antimicrobial activity of eucalyptus oil. In: Coppen, J.J.W. (ed) Eucalyptus: the genus Eucalyptus.Taylor & Francis, London.
Dell, B., Havel, J.J., Malajczuk N., 1989. The jarrah forest: a complex Mediterranean ecosystem. Kluwer Academic Publishers, The Netherlands.
Ehrenfeld, J.G., Ravit, B., Elgersma, K., 2005. Feedback in the plant-soil system. Annu Rev Environ Resour 30:75–115.
Eisenhauer, N., Reich, P.B., 2012. Above- and below-ground plant inputs both fuel soil food webs. Soil Biol Biochem 45:156-160.
Eviner, V.T., 2004. Plant traits that influence ecosystem processes vary independently among species. Ecology 85(8):2215-2229.
Eviner, V.T., Hawkes, C.V., 2008. Embracing variability in the application of plant–soil interactions to the restoration of communities and ecosystems. Restor Ecol 16(4):713-729.
Farrar, J., Hawes, M., Jones, D., Lindow, S., 2003. How roots control the flux of C to the rhizosphere. Ecology 84(4): 827-837.
Feller, M.C., 1983. Effects of an exotic conifer (Pinus radiata) plantation on forest nutrient cycling in Southeastern Australia. For Ecol Manage 7:77-102.
Ferris, H., Bongers, T., Goede, R.G.M., 2001. A framework for soil food web diagnostics: extension of the nematode faunal analysis concept. Appl Soil Ecol 18:13-29.

111
Ferris, H., Matute, M.M., 2003. Structural and functional succession in the nematode fauna of a soil food web. Appl Soil Ecol 23:93-110.
Foth, H.D., 1990. Fundamentals of soil science 8th edition. John Wiley & Sons USA.
Freckman, D.W., 1988. Bacterivorous nematodes and organic-matter decomposition. Agr Ecosyst Environ 24:195-217.
Frostegård, A., Bååth, E., 1996. The use of phospholipid fatty acid analysis to estimate bacterial and fungal biomass in soil. Biol Fertil Soils 22(1):59-65.
Fukami, T., Nakajima, M., 2013. Complex plant-soil interactions enhance plant species diversity by delaying community convergence. J Ecol 101:316-324.
Ganjegunte, G.K., Condron, L.M., Clinton, P.W., Davis, M.R., 2005. Effects of mixing radiata pine needles and understory litters on decomposition and nutrients release. Biol Fertil Soils 41:310-319.
Gardner, J.H., Bell, D.T., 2007. Bauxite mining restoration by Alcoa World Alumina Australia in Western Australia: social, political, historical, and environmental contexts. Restor Ecol 15(4):S3-S10.
Gentili, J., 1989. Climate of the jarrah forest. In: Dell, B., Havel, J.J., Malajczuk, N. (eds) The jarrah forest: a complex Mediterranean ecosystem. Kluwer Academic Publishers, the Netherlands.
George, S.J., Kelly, R.N., Greenwood, P.F., Tibbett, M., 2010. Soil carbon and litter development along a reconstructed biodiverse forest chronosequence of south-western Australia. Biogeochemistry 101:197–209.
George, S.J., Tibbett, M., Braimbridge, M.F., Davis, S.G., Vlahos, S., Ryan, M., 2006. Phosphorus fertiliser placement and seedling success in Australian jarrah forest. In: Fourie, A., Tibbett, M. (eds) Mine Closure. Australian Centre for Geomechanics, Perth.
Gong, X., Liu, Y., Li, Q., Wei, X., Guo, X., Niu, D., Zhang, W., Zhang, J., Zhang. L., 2013. Sub-tropic degraded red soil restoration: Is soil organic carbon build-up limited by nutrients supply. For Ecol Manage 300:77-87.
Gonzalez-Quiñones, V., Banning, N.C., Jimenez, B.R., Murphy, D.V. 2009. Influence of cold storage on soil microbial community level physicological profiles and implications for soil quality monitoring. Soil Biol Biochem 41:1574-1576.
Gorbushina, A.A., Krumbein, W.E., 2005. Role of microorganisms in wear down of rocks and minerals. In: Buscot, F., Varma, A. (eds) Microorganisms in soil: roles in genesis and functions. Springer-Verlag, Berlin.
Grant, C.D., 2006. State-and-transition successional model for bauxite mining rehabilitation in the jarrah forest of Western Australia. Restor Ecol 14(1):28-37.
Grierson, P.F., Adams, M.A., 2000. Plant species affect acid phosphatase, ergosterol and microbial P in a Jarrah (Eucalyptus marginata Donn ex Sm.) forest in south-western Australia. Soil Biol Biochem 32:1817-1827.
Griffiths, B.S., Christensen, S., Bonkowski, M., 2007. Microfaunal interactions in the rhizosphere, how nematodes and protozoa link above- and belowground processes. In: Cardon, Z.G., Whitbeck, J.L. (eds) The rizosphere: an ecological perspective. Elsevier, USA.

112
Guo, L.B., Sims, R.E.H., 1999. Litter decomposition and nutrient release via litter decomposition in New Zealand eucalypt short rotation forests. Agr Ecosyst Environ 75:133–140.
Gupta, V.V.S.R., Yeates, G.W., 1997. Soil microfauna as bioindicators of soil health. In: Pankhurst, C., Doube, B.M., Gupta, V.V.S.R. (eds) Biological indicators of soil health. CAB International, UK.
Hadacek, F., 2008. Constitutive secondary metabolites and soil fungi: defence against or facilitation of diversity. In: Karlovsky, P. (ed) Secondary metabolites in soil ecology. Springer-Verlag, Berlin.
Harris, J., 2009. Soil microbial communities and restoration ecology: facilitators or followers. Science 325:572-574.
Harrison, K.A., Bardgett, R.D., 2010. Influence of plant species and soil conditions on plant-soil feedback in mixed grassland communities. J Ecol 98:384-395.
Hingston, F.J., Dimmock, G.M., Turton, A.G., 1981. Nutrient distribution in a jarrah (Eucalyptus marginata Donn Ex Sm.) ecosystem in south-west Western Australia. For Ecol Manage 3:183-207.
Hingston, F.J., O’Connell, A.M., Grove, T.S., 1989. Nutrient cycling in jarrah forest. In: Dell, B., Havel, J.J., Malajczuk, N. (eds) The jarrah forest: a complex Mediterranean ecosystem. Kluwer Academic Publishers, The Netherlands.
Hirsch, A.M., Bauer, W.D., Bird, D.M., Cullimore, J., Tyler, B., Yoder, J.I., 2003. Molecular signals and receptors: controlling rhizosphere interactions between plants and other organisms. Ecology 84(4):858-868.
Hobbie, S.E., 1992. Effects of plant species on nutrient cycling. Trends Ecol Evol 7(10):336-339.
Hobbs, R.J., Legg, C.J., 1983. Markov models and initial floristic composition in heathland vegetation dynamics. Vegetatio 56:31–43.
Holtkamp, R., Kardol, P., van der Wal, A., Dekker, S.C., van der Putten, W.H., de Ruiter, P.C., 2008. Soil food web structure during ecosystem development after land abandonment. Appl Soil Ecol 39:23–34.
Hooper, D.J., Hallmann, J., Subbotin, S.A., 2005. Methods for extraction, processing and detection of plant and soil nematodes. In: Luc, M., Sikora, R.A., Bridge, J. (eds) Plant parasitic nematodes in subtropical and tropical agriculture 2nd edition. CAB Publishing, UK.
Hooper, D.U., Bignell, D.E., Brown, K.V., Brussard, L., Dangerfield, J.M., Wall, D.H., Wardle, D.A., Coleman, D.C., Giller, K.E., Lavelle, P., van der Putten, W.H., de Ruiter, P.C., Rusek, J., Whendee, L.S., Tiedje, J.M., Wolters, V., 2000. Interactions between aboveground and belowground biodiversity in terrestrial ecosystems: patterns, mechanisms, and feedbacks. BioScience 50(12):1049-1061.
Hopkins, D.W., Dungait, J.A.J., 2010. Soil microbiology and nutrient cycling. In: Dixon, G.R., Tilston, E.L. (eds) Soil microbiology and sustainable crop production. Springer, New York.
Hopmans, P., Elms, S.R., (2009) Changes in total carbon and nutrients in soil profiles and accumulation in biomass after a 30-year rotation of Pinus radiata on podzolized sands: impacts of intensive harvesting on soil resources. For Ecol Manage 258:2183-2193.

113
Hopper, S.D., 2009. OCBIL theory: towards an integrated understanding of the evolution, ecology and conservation of biodiversity on old, climatically buffered, infertile landscapes. Plant Soil 322:49-86.
Hussain, M.I., González, L., Souto, C., Reigosa, M.J., 2011. Ecophysicological responses of three native herbs to phytotoxic potential of invasive Acacia melanoxylon R. Br. Agroforest Syst 83:149-166.
Inderjit, Mallik, A.U., 1997. Effect of phenolic compounds on selected soil properties. Forest Ecol Manag 92:11-18.
Inderjit, 2005. Soil microorganisms: An important determinant of allelopathic activity. Plant Soil 274:227-236.
Inderjit, Weiner, J., 2001. Plant allelochemical interference or soil chemical ecology? Perspect Plant Ecol 4(1):3-12.
Inderjit, van der Putten, W.H., 2010. Impacts of soil microbial communities on exotic plant invasions. Trends Ecol Evol 25:512-519.
Insam, H., Domsch, H.K., 1988. Relationship between soil organic carbon and microbial biomass on chronosequences of reclamation sites. Microbial Ecol 15(2): 177-188.
Jasper, A.D., 2007. Beneficial soil microorganisms of the and their recovery in bauxite mine restoration in South Western Australia. Restor Ecol 15(4):S74-S84.
Jastrow, J.D., Miller, R.M., Lussenhop, J., 1998. Contributions of interacting biological mechanisms to soil aggregate stabilization in restored prairie. Soil Biol Biochem 30(7):905-916.
Johnson, C., 2009. Biology of soil science. Oxford Book Company, Delhi.
Jones, D.L., 1998. Organic acids in the rhizosphere – a critical review. Plant Soil 205:25-44.
Jordan, N.R., Larson, D.L., Huerd, S.C., 2008. Soil modification by invasive plants: effects on native and invasive species of mixed-grass prairies. Biol Invasions 10:177-190.
Kanesaki, Y., Los, D.A., Suzuki, I., Murata, N., 2010. Sensors and signal transducers of environmental stress in Cyanobacteria. In: Pareek, A., Sopory, S.K., Bohnert, H.J., Govindjee (eds) Abiotic stress adaptation in plants: physiological, molecular and genomic foundation. Springer, The Netherlands.
Kapoor, R., Mukerji, K.G., 2006. Rhizosphere microbial community dynamics. In: Mukerji, K.G., Manoharachary, C., Singh, J. (eds) Microbial activity in the rhizosphere. Springer, Germany.
Kardol, P., Bezemer, T.M., van der Putten, W.H., 2006. Temporal variation in plant–soil feedback controls succession. Ecol Lett 9: 1080–1088.
Kardol, P., Cornips, N.J., van Kempen, M.M.L., Bakx-Schotman, J.M.T., van der Putten, W.H., 2007. Microbe-mediated plant-soil feedback causes historical contingency effects in plant community assembly. Ecol Monogr 77:147-162.
Kardol, P., Wardle, D.A., 2010. How understanding aboveground-belowground linkages can assist restoration ecology. Trends Ecol Evol 25(11):670-679.
Kardol, P., De Deyn, G.B., Laliberté, E., Mariotte, P., Hawkes, C.V., 2013. Biotic plant-soil feedbacks across temporal scales. J Ecol 101:309-315.

114
Keeley, J.E., Bond, W.J., Bradstock, R.A., Pausas, J.G.,Rundel, P.W., 2012. Fire in Mediterranean ecosystems: Ecology, evolution and management. Cambridge University Press, USA.
Koch, J.M., 2007a. Bauxite mining restoration by Alcoa World Alumina Australia in Western Australia: social, political, historical, and environmental contexts. Restor Ecol 15(4):S3-S10.
Koch, J.M., 2007b. Alcoa’s mining and restoration process in south Western Australia. Restor Ecol 15(4):S11-S16.
Koch, J.M., Samsa, G., 2007. Restoring jarrah forest trees after bauxite mining in Western Australia. Restor Ecol 15(4):S17-S25.
Kohli, R.K., Batish, D.R., Singh, H.P., 2006. Allelopathic interactions in agroecosystems. In: Reigosa, M.J., Pedrol, N., González, L. (eds) Allelopathy: a physiological process with ecological implications. Springer, The Netherlands.
Kulmatiski A, Beard KH, Stark JM (2006) Soil history as a primary control on plant invasion in abandoned agricultural fields. J Appl Ecol 43:868-876.
Kulmatiski, A., Beard, H.K., Stevens, R.J., Cobbold, M.S., 2008. Plant-soil feedbacks: a meta-analytical review. Ecol Lett 11:980-992.
Kulmatiski, A., Beard, H.K., 2008. Decoupling plant-growth from land-use legacies in soil microbial communities. Soil Biol Biochem 40:1059-1068.
Kulmatiski, A., Kardol, P., 2008. Getting plant-soil feedbacks out of the greenhouse: experimental and conceptual approaches. Prog Bot 69:449-472.
Kulmatiski, A., Beard, H.K., 2011. Long-term plant growth legacies overwhelm short-term plant growth effects on soil microbial community structure. Soil Biol Biochem 43:823-830.
Kylafis, G., Loreau, M., 2008. Ecological and evolutionary consequences of niche construction for its agent. Ecol Lett 11:1072-1081.
Kyle, G.P., Beard, K.H., Kulmatiski, A., 2007. Reduced soil compaction enhances growth of non-native species. Plant Ecol 193(2):223-232.
Ladygina, N., Hedlund, K., 2010. Plant species influence microbial diversity and carbon allocation in the rhizosphere. Soil Biol Biochem 42:162-168.
Lalor, E.L., Cookson, W.R., Murphy, D.V., 2007. Comparison of two methods that assess soil community level physiological profiles in a forest ecosystem. Soil Biol Biochem 39:454–462.
Lambers, H., Raven, J.A., Shaver, G.R., Smith, S.E., 2007. Plant nutrient-acquisition strategies change with soil age. Trends Ecol Evol 23(2):95-103.
Lambers, H., Bishop, J.G., Hopper, S.D., Laliberté, E., Zuñiga-Feest, A., 2012. Phosphorus-mobilization ecosystem engineering: the roles of cluster roots and carboxylate exudation in young P-limited ecosystems. Ann Bot-London 110:329-348.
Laskowski, R., Niklinska, M., Maryanski, M., 1995. The dynamics of chemical elements in forest litter. Ecology 76(5):1393-1406.
Lavelle, P., Spain, A.V., 2003. Soil Ecology. Kluwer Academic Publishers, USA
Lawrie, A.C., 1981. Nitrogen fixation by native Australian legumes. Aust J Bot 29:143-157.

115
Lin, D.S., Greenwood, P.F., George, S.J., Somerfield P.J., Tibbett, M., 2011. The development of soil organic matter in restored biodiverse jarrah forests of South-Western Australia as determined by ASE and GCMS. Environ Sci Pollut R 18:1070–1078.
Liu, Y., Yu, S., Xie, Z., Staehelin, C., 2012. Analysis of a negative plant–soil feedback in a subtropical monsoon forest. J Ecol 100:1019-1028.
Lorenzo, P., Pereira, C.S., Rodríguez-Echeverría, S., 2013. Differential impact on soil microbes of allelopathic compounds released by the invasive Acacia dealbata Link. Soil Biol Biochem 57:156-163.
Lotina-Hennsen, B., King-Díaz, B., Aguilar, M.I., Hernandez-Terrones, M.G., 2006. Plant secondary metabolites. Targets and mechanisms of allelopathy. In: Reigosa, M.J., Pedrol, N., González, L. (eds) Allelopathy: A physiological process with ecological implications. Springer, The Netherlands.
Lugo, A.E., Cuevas, E., Sánchez, M.J., 1990. Nutrients and mass in litter and top soil of ten tropical tree plantations. Plant Soil 125:263-280.
Madigan, M.T., Martinko, J.M., Stahl, D.A., Clark, D.P., 2012. Brock Biology of Microorganisms 13th edition. Benjamin Cummings Editorial, USA.
Malajczuk, N., McComb, A.J., 1977. Root exudates from Eucalyptus calophylla R.Br. and Eucalyptus marginata Donn. ex Sm. seedlings and their effect on Phytophthora cinnamomi Rands. Aust J Bot 25:501-514.
Malajczuk, N., McComb, A.J., 1979. The microflora of unsuberized roots of Eucalyptus calophylla R.Br. and Eucalyptus marginata Donn ex Sm. seedlings grown in soil suppressive and conducive to Phytophthora cinnamomi Rands. I Rhizosphere bacteria, actinomycetes and fungi. Aust J Bot 27:235-254.
Malajczuk, N., 1979. The microflora of unsuberized roots of Eucalyptus calophylla R. Br. and Eucalyptus marginata Donn ex Sm. seedlings grown in soils suppressive and conducive to Phytophthora cinnamomi Rands. II Mycorrhizal roots and associated microflora. Aust J Bot 27:255-272.
Malik, A., Scullion, J., 1998. Soil development on restored opencast coal sites with particular reference to organic matter and aggregate stability. Soil Use Manage 14:234-239.
Maluf, de S.F., Ferreira, B.J.L., 2004. Restoration of seasonal semideciduous forests in Brazil: influence of age and restoration design on forest structure. For Ecol Manage 191:185-200.
Mangan, S.A., Schnitzer, S.A., Herre, E.A., Mack, K.M.L., Valencia, M.C., Sanchez, E.I., Bever, J.D., 2010. Negative plant–soil feedback predicts tree-species relative abundance in a tropical forest. Nature 466 doi: 10.1038/nature09273.
Manning, P., Morrison, S.A., Bonkowski, M., Bardgett, R.D., 2008. Nitrogen enrichment modifies plant community structure via changes to plant-soil feedback. Oecologia, 157(4): 661-673.
Manoharachary, C., Mukerji, K.G., 2006. Rhizosphere biology – an overview. In: Mukerji, K.G., Manoharachary, C., Singh, J. (eds) Microbial activity in the rhizosphere. Springer, Germany.
Marschner, H., 1995. Mineral nutrition of higher plants 2nd edition. Academic Press, Great Britain.

116
Martin, B.C., George, S.J., Price, C.A., Ryan, M.H., Tibbett, M., 2014. The role of root exuded low molecular weight organic anions in facilitating petroleum hydrocarbon degradation: current knowledge and future directions. Sci Total Environ 472:642-653.
Meijer, S.S., Holmgren, M., van der Putten, W.H., 2011. Effects of plant–soil feedback on tree seedling growth under arid conditions. J Plant Ecol doi: 10.1093/jpe/rtr011
Menta, C., Conti, F.D., Pinto, S., Leoni, A., Lozano-Fondón, C., 2013. Monitoring soil restoration in an open-pit mine in northern Italy. Appl Soil Ecol doi.org/10.1016/j.apsoil.2013.07.013
Miki, T., Ushio, M., Fukui, S., Kondoh, M., 2010. Functional diversity of microbial decomposers facilitates plant coexistence in a plant–microbe–soil feedback model. PNAS 107(32):14251-14256.
Miki, T., 2012. Microbe-mediated plant-soil feedback and its roles in a changing world. Ecol Res 27:509-520.
Mills, E.K., Bever, D.J., 1998. Maintenance of diversity within plant communities: soil pathogens as agents of negative feedback. Ecology 79(5):1595-1601.
Monk, D., Pate, J.S., Loneragan, W.A., 1981. Biology of Acacia pulchella R.Br. with special reference to symbiotic nitrogen fixation. Aust J Bot 29: 579–592.
Moore, J.C., Hunt, H.W., 1988. Resource compartmentation and the stability of real ecosystems. Nature 33:261-263.
Moore, J.C., McCann, K., Setäla, H., de Ruiter, P.C., 2003. Top-down is bottom-up: does predation in the rhizosphere regulate aboveground dynamics? Ecology 84:846-857.
Moore, J.C., McCann, K., de Ruiter, P.C., 2007. Soil rhizosphere food webs, their stability, and implications for soil processes in ecosystems. In: Cardon, Z.G., Whitbeck, J.L. (eds) The rhizosphere: an ecological perspective. Elsevier, USA.
Mummey, D.L., Stahl, P.D., Buyer, J.S., 2002. Soil microbiological properties 20 years after surface mine reclamation: spatial analysis of reclaimed and undisturbed sites. Soil Biol Biochem 34:1717-1725.
Myers, N., Mittermeier, R.A., Mittermeier, C.G., da Fonseca, G.A.B., Kent, J., 2000. Biodiversity hotspots for conservation priorities. Nature 403:853-858.
Nautiyal, C.S., Srivastava, S., Chauhan, P.S., 2008. Rhizosphere colonization: molecular determinants from plant-microbe coexistence perspective. In: Nautiyal, C.S., Dion, P. (eds) Molecular mechanisms of plant and microbe coexistence. Springer, Germany.
Neher, D.A., 1999. Soil community composition and ecosystem processes: comparing agricultural ecosystems with natural ecosystems. In: Lefroy, E.C., Hobbs, R.J., O’Connor, M.H., Pate, J.S. (eds) Agriculture as a mimic of natural ecosystems. Kluwer Academic Publishers, The Netherlands.
Nelson, E.B., 2006. Rhizosphere regulation of preinfection behaviour of oomycete plant pathogens. In: Mukerji, K.G., Manoharachary, C., Singh, J. (eds) Microbial activity in the rhizosphere. Springer, Germany.
Neumann, G., Römheld, V., 1999. Root excretion of carboxylic acids and protons in phosphorus-deficient plants. Plant Soil 211:121-130.
Neumann, G., Römheld, V., 2001. The release of root exudates as affected by the plant’s physicological status. In: Pinton, R., Varanini, Z., Nannipieri, P. (eds) The rhizosphere:

117
biochemistry and organic substances at the plant-soil interface. Marcel Dekker Editorial, USA.
Neumann, G., 2007. Root exudates and nutrient cycling. In: Marschner, P., Rengel, Z. (eds) Nutrient cycling in terrestrial ecosystems. Springer-Verlag, Berlin.
Norman, M.A., Koch, J.M., Grant, C.D., Morald, T.K., Ward, S.C., 2006. Vegetation succession after bauxite mining in Western Australia. Restor Ecol 14(2):278-288.
O’Brien, N.D., Attiwill, P.M., Weston, C.J., 2003. Stability of soil organic matter in Eucalyptus regnans forests and Pinus radiata plantations in south eastern Australia. For Ecol Manage 185:249-261.
O’Connell, A.M., Grove, T.S., Dimmock, G.M., 1978. Nutrients in the litter of jarrah forest soils. Aust J Ecol 3:253-260.
Pang, J., Ryan, M.H., Tibbett, M., Cawthray, G.R., Siddique, K.H.M., Bolland, M.D.A., Denton, M.D., Lambers, H., 2010. Variation in morphological and physiological parameters in herbaceous perennial legumes in response to phosphorus supply. Plant Soil 331:241-255.
Papatheodorou, E.M., 2008. Responses of soil microbial communities to climatic and human impacts in Mediterranean regions. In: Liu, T-X (ed) Soil ecology research developments. Nova Science Publishers, New York.
Paul, M., Catterall, C.P., Pollard, P.C., Kanowski, J., 2010. Recovery of soil properties and functions in different rainforest restoration pathways. For Ecol Manage 259:2083-2092.
Pekin, B.K., Wittkuhn, R.S., Boer, M.M., MacFarlane, C., Grierson, P., 2012. Response of plant species and life form diversity to variable fire histories and biomass in the jarrah forest of south-west Australia. Aust Ecol 37:330-338.
Perkins, L.B., Nowak, R.S., 2012. Soil conditioning and plant-soil feedbacks affect competitive relationships between native and invasive grasses. Plant Ecol 213:1337-1344.
Pernilla, B.E., van der Putten, W.H., Bakker, E.J., Verhoeven, K.J.F., 2010. Plant–soil feedback: experimental approaches, statistical analyses and ecological interpretations. J Ecol 98:1063-1073.
Perry, R.N., Tovar-Soto, A., 2012. Biology and host-parasite interactions. In: Manzanilla-López, R.H., Marbán-Mendoza, N. (eds) Practical plant Nematology. Biblioteca Básica de Agricultura, Mexico.
Philippot, L., Germon, J.C., 2005. Contribution of bacteria to initial input and cycling of nitrogen in soil. In: Buscot, F., Varma, A. (eds) Microorganisms in soil: roles in genesis and functions. Springer-Verlag, Berlin.
Ponge, J.F., 2013. Plant-soil feedbacks mediated by humus forms: A review. Soil Biol Biochem 57:1048-1060.
Postma-Blaauw, M.B., de Vries, F.T., de Goede, R.G.M., Bloem, J., Faber, J.H., Brussaard, L., 2005. Within-trophic group interactions of bacterivorous nematode species and their effects on the bacterial community and nitrogen mineralization. Oecologia 142:428-439.
Pregitzer, K.S., Azk, D.R., Loya, W.M., Karberg, N.J., King, J.S., Burton, A.J., 2007. The contribution of root – rhizosphere interactions to biogeochemical cycles in a changing world. In: Cardon, Z.G., Whitbeck, J.L. (eds) The rhizosphere: an ecological perspective. Elsevier, USA.

118
Pregitzer, C.C., Bailey, J.K., Hart, S.C., Schweitzer, J.A., 2010. Soils as agents of selection: feedbacks between plants and soils alter seedling survival and performance. Evol Ecol 24:1045–1059.
Prescott, C.E., Graystone, S.J. 2013. Tree species influence on microbial communities in litter and soil: current knowledge and research needs. Forest Ecol Manage 309:19-27.
Prober, S.M., Lunt, I.D., Morgan, J.W. 2009. Rapid internal plant-soil feedbacks lead to alternative stable states in temperate Australian grassy woodlands. In: Hoobs, R.J., Suding, K.N. (eds) New models for ecosystem dynamics and restoration. Island Press, USA.
Ranganayaki, N., Tilak, K.V.B.R., Manoharachary, C., Mukerji, K.G., 2006. Methods and techniques for isolation, enumeration and characterization of rhizosphere microorganisms In: Mukerji, K.G., Manoharachary, C., Singh, J. (eds) Microbial activity in the rhizosphere. Springer, Germany.
Rayment, G.E., Higginson, F.R., 1992. Australian laboratory handbook of soil and water chemical methods: Australian soil and land survey handbook Vol 3. Inkata Press, Australia.
Reuben, S., Bhinu, V.S., Swarup, S. 2008. Rhizosphere metabolomics:methods and applications. In: Karlovsky, P. (ed) Secondary metabolites in soil ecology. Springer, Berlin.
Reynolds, H.L., Packer, A., Bever, J.D., Clay, K., 2003. Grassroots ecology: plant–microbe–soil interactions as drivers of plant community structure and dynamics. Ecology 84(9):2281-2291.
Ribeiro, R.R., Venancio, M.S., de Barros, L.C., 2004. Tropical Rain Forest regeneration in an area degraded by mining in Mato Grosso State, Brazil. For Ecol Manage 190:323-333.
Richardson, A.E., Barea, J.M., McNeill, A.M., Prigent-Combaret, C., 2009. Acquisition of phosphorus and nitrogen in the rhizosphere and plant growth promotion by microorganisms. Plant Soil 321:305–339.
Rock, C.D., Sakata, Y., Quatrano, R.S., 2010. Stress signalling I: The role of abscisic acid (ABA) In: Pareek, A., Sopory, S.K., Bohnert, H.J., Govindjee (eds) Abiotic stress adaptation in plants: physiological, molecular and genomic foundation. Springer, The Netherlands.
Sardans, J., Peñuelas, J., 2013. Plant-soil interactions in Mediterranean forest and shrublands: impacts of climatic change. Plant Soil 365:1-33.
Sbrana, C., 2006. Fungal recognition responses to host derived signals by arbuscular mycorrhizal fungi. In: Mukerji, K.G., Manoharachary, C., Singh, J. (eds) Microbial activity in the rhizosphere. Springer, Germany.
Scheu, S., Ruess, L., Bonkowski, M., 2005. Interactions between microorganisms and soil micro- and mesofauna. In: Buscot, F., Varma, A. (eds) Microorganisms in soils: roles in genesis and functions. Springer-Verlag, Berlin.
Schimann, H., Petit-Jean, C., Guitet, S., Reis, T., Domenach, A.M., Roggy, J.-C. 2012. Microbial bioindicators of soil functioning after disturbance: The case of gold mining in tropical rainforests of French Guiana. Ecol Indic 20:34-41.
Schipper, L.A., Degens, B.P., Sparling, G.P., Duncan, L.C., 2001. Changes in microbial heterotrophic diversity along five plant successional sequences. Soil Biol Biochem 33:2093-2103.

119
Scholefield, D., Jewkes, E., Bol, Roland., 2007. Nutrient cycling budgets in managed pastures. In: Marschner, P., Rengel, Z. (eds) Nutrient cycling in terrestrial ecosystems. Springer-Verlag, Berlin.
Shannon, S., Flory, S.L., Reynolds, H., 2012. Competitive context alters plant-soil feedback in an experimental woodland community. Oecologia 169:235-243.
Shono, K., Davies, S.J., Kheng, C.Y., 2006. Regeneration of native plant species in restored forests on degraded lands in Singapore. For Ecol Mange 237:574-582.
Singh, G., Mukerji, K.G., 2006. Root exudates as determinant of rhizospheric microbial diversity. In: Mukerji, K.G., Manoharachary, C., Singh, J. (eds) Microbial activity in the rhizosphere. Springer, Germany.
Solaiman, M.Z., Abbott, L.K., 2003. Phosphorus uptake by a community of arbuscular mycorrhizal fungi in jarrah forest. Plant Soil 248:313-320.
Spain, A.V., Tibbett, M., Hinz, D.A., Ludwig, J.A., Tongway, D.J., 2014. The mining-restoration system and ecosystem development following bauxite mining in a biodiverse environment of the seasonally dry tropics, Northern Territory, Australia. In: Tibbett, M. (ed) Mining in Ecologically Sensitive Landscapes. CSIRO Press, In Press
Spring, S., Schulze, R., Overmann, J., Schleifer, K-H., 2000. Identification and characterization of ecologically significant prokaryotes in the sediment of freshwater lakes: molecular and cultivation studies. FEMS Microbiol Rev 24(5):573-590.
Standish, R.J., Tibbett, M., Vlahos, S., Stokes, B.A., Hobbs, R.J., 2010. Effect of fertiliser on early-successional jarrah forest restored after bauxite mining in south-western Australia. In: Fourie, A., Tibbett, M., Wiertz, J. (eds) Mine Closure. Australian Centre for Geomechanics, Perth.
Suding, K.N., Gross, K.L., Houseman, G.R. (2004) Alternative states and positive feedbacks in restoration ecology. Trends Ecol Evol 19(1):46-53.
Suding, K.N., Hobbs, R.J., 2008. Threshold models in restoration and conservation: a developing framework. Trends Ecol Evol 24:271-279.
Suding, K.N., Harpole, W.S., Fukami, T., Kulmatiski, A., MacDougall, A.S., Stein, C., van der Putten, W.H., 2013. Consequences of plant-soil feedbacks in invasion. J Ecol 101:298-308.
te Beest, M., Stevens, N., Olff, N., van der Putten, W.H., 2009. Plant–soil feedback induces shifts in biomass allocation in the invasive plant Chromolaena odorata. J Ecol 97:1281–1290.
Tibbett, M., 2010. Large-scale mine site restoration of Australian eucalypt forests after bauxite mining: soil management and ecosystem development. In: Batty, L.C., Hallberg, K. (eds) Ecology of industrial pollution. Cambridge University Press, UK.
Topp, G.C., Parkin, G.W., Ferré, T.P.A. Soil water content In: Carter, M.R., Gregorich, E.G. (eds) 2008. Soil sampling and methods of analysis 2nd edition. CRC Press, USA.
Treonis, A.M, Ostle, N.J., Stott, A.W., Primrose, R., Grayston, S.J., Ineson, P., 2004. Identification of groups of metabolically-active rhizosphere microorganisms by stable isotope probing of PLFAs. Soil Biol Biochem 36:533-537.
Turner, J., Lambert, M.J., 2008. Nutrient cycling in age sequences of two Eucalyptus plantation species. For Ecol Manage 255:1701-1712.

120
Uren, N.C., 2001. Types, amounts, and possible functions of compounds released into the rhizosphere by soil-grown plants. In: Pinton, R., Varanini, Z., Nannipieri, P. (eds) The Rhizosphere: Biochemistry and organic substances at the soil-plant interface. Marcel Dekker Editorial, USA.
Uselman, S.M., Qualls, R.G., Lilienfein, J., 2012. Quality of soluble organic C, N, and P produced by different types and species of litter: root litter versus leaf litter. Soil Biol Biochem 54:57-67.
van de Voorde, T.F.J., van der Putten, W.H., Bezemer, T.M., 2011. Intra- and interspecific plant–soil interactions, soil legacies and priority effects during old-field succession J Ecol doi: 10.1111/j.1365-2745.2011.01815.
van der Heijden, M.G.A., Klironomos, J.N., Ursic, M., Moutoglis, P., Streitwolf-Engel, R., Boller, T., Wiemken, A., Sanders, I.R., 1998. Mycorrhizal fungal diversity determines plant biodiversity, ecosystem variability and productivity. Nature 396: 69–72.
van der Heijden, M.G.A., Bardgett, R.D., van Straalen, N.M., 2008. The unseen majority: soil microbes as drivers of plant diversity and productivity in terrestrial ecosystems. Ecol Lett 11:296-310.
van der Putten, W.H., Peters, B.A.M., 1997. How soil-borne pathogens may affect plant competition. Ecology 78(6):1785-1795.
van der Putten, W.H., 2000. Pathogen-driven forest diversity. Nature 404:232-233.
van der Putten, W.H., Bardgett, R.D., de Ruiter, P.C., Hol, W.H.G., Meyer, K.M., Bezemer, T.M., Bradford, M.A., Christensen, S., Eppinga, M.B., Fukami, T., Hemerik, L., Molofsky, J., Schädler, M., Scherber, C., Strauss, S.Y., Vos, M., Wardle, D.A., 2009. Empirical and theoretical challenges in aboveground–belowground ecology. Oecologia 161:1–14.
van der Putten, W.H., Bardgett, R.D., Bever, J.D., Bezemer, T.M., Casper, B.B., Fukami, T., Kardol, P., Klironomos, J.N., Kulmatiski, A., Schweitzer, J.A., Suding, K.N., van de Voorde, T.F.J., Wardle, D.A., 2013. Plant-soil feedbacks: the past, present and future challenges. J Ecol 101(2):265-276.
van der Stoel, C.D., van der Putten, W.H., Duyts, H., 2002. Development of a negative plant-soil feedback in the extension zone of the clonal grass Ammophila arenaria following root formation and nematode colonization. J Ecol 90:978-988.
van Grunsven, R.H.A., van der Putten, W.H., Bezemer, T.M., Tamis, W.L.M., Berendse, F., Veenendaal, E.M., 2007. Reduced plant-soil feedback of plant species expanding their range as compared to natives. J Ecol 95:1050-1057.
Viketoft, M., Palmborg, C., Sohlenius, B., Huss-Danell, K., Bengtsson, J., 2005. Plant species effects on soil nematode communities in experimental grasslands. Appl Soil Ecol 30:90-103.
Ward, S.C., 2000. Soil development on restored bauxite mines in south-west Australia. Aust J Soil Res 38(2):453.
Wardle, D.A., 1999. How soil food webs make plants grow. Trends Ecol Evol 14 (11):418-420.
Wardle, D.A., Bardgett, D.R., Klironomos, N.J., Setäla, H., van der Putten, H.W., Wall, H.D. 2004. Ecological linkages between aboveground and belowground biota. Science 304:1629-1633.
Wardle, D.A., 2006. The influence of biotic interactions on soil biodiversity. Ecol Lett 9:870-886.

121
Wardle, D.A., Bardgett, R.D., Klironomos, J.N., Setälä, H., van der Putten, W.H., Wall, D.H., 2004. Ecological linkages between aboveground and belowground biota. Science 304(5677):1629-1633.
Weidenhamer, J.D., Callaway, R.M., 2010. Direct and indirect effects of invasive plants on soil chemistry and ecosystem function. J Chem Ecol 36:59-69.
Weir, T.L., Park, S-W., Vivanco, J.M., 2004. Biochemical and physiological mechanisms mediated by allelochemicals. Curr Opin Plant Biol 7:472-479.
Werner, D., 2001. Organic signals between plants and microorganisms. In: Pinton, R., Varanini, Z., Nannipieri, P. (eds) The Rhizosphere: Biochemistry and organic substances at the soil-plant interface. Marcel Dekker Editorial, USA.
Whisenant, S.G. 1999, Repairing damaged wildlands: A process-oriented, landscape-scale Approach. Cambridge University Press.
White, D.C., Davies, W.M., Nickels, J.S., Kings, J.D., Bobbie, R.J., 1979. Determination of the sedimentary microbial biomass by extractible lipid phosphate. Oecologia 40: 51-62.
Whitfield, F.B., Shea, S.T., Gillen, K.J., Shaw, K.J., 1981. Volatile components from the roots of Acacia pulchella R.Br. and their effect on Phytophtora cinnamomi Rands. Aust J Bot 29:195-208.
Willis, R., 1999. Australian studies on allelopathy in Eucalyptus: A review. In: Inderjit, Dakshini, K.M.M., Foy, L.F. (eds) Principles and practices in plant ecology: allelochemical interactions. CRC Press, USA.
Witt, C., Setälä, H., 2010. Do plant species of different resource qualities form dissimilar energy channels below-ground? Appl Soil Ecol 44:270-278.
Xiaoyun, C., Manqiang, L., Feng, H., Xiaofang, M., Huixin, L., 2007. Contributions of soil micro-fauna (protozoa and nematodes) to rhizosphere ecological functions. Acta Ecol Sin 27(8):3132-3143.
Yeates, G.W., Bongers, T., M de Goede, R.G., Freckman, D.W., Georgieva, S.S., 1993. Feeding habits in soil nematode families and genera – An outline for soil ecologists. J Nematol 25(3):315-331.
Yeates, G.W., 2003. Nematodes as soil indicators: functional and biodiversity aspects. Biol Fertil Soils 37:199–210.
Yeates, G.W., Ferris, H., Moens, T., van der Putten, W.H., 2009. The role of nematodes in ecosystems. In: Wilson MJ, Kakouli-Duarte T (eds) Nematodes as environmental indicators. CABI, UK.
Zavaleta-Mejía, E., Kaloshian, I., 2012. Plant-nematode interactions. In: Manzanilla-López, R.H., Marbán-Mendoza, N. (eds) Practical plant Nematology. Biblioteca básica de Agricultura, México.
Zechmeister-Boltenstern, S., Michel, K., Pfeffer, M. 2011. Soil microbial community structure in European forests in relation to forest type and atmospheric nitrogen deposition. Plant Soil 343(1):37-50.
Zelles, L., Bai, Q.Y., 1993. Fractionation of fatty acids derived from soil lipids by soil phase extraction and their quantitative analysis by GC–MS. Soil Biol Biochem 25:495-507.
Zelles, L., 1999. Fatty acid patterns of phospholipids and lipopolysaccharides in the characterisation of microbial communities in soil: a review. Biol Fetil Soils 29:111-129.

122
Zhang, Z-Y., Pan, L-P., Li, H-H., 2009. Isolation, identification and characterization of soil microbes which degrade phenolic allelochemicals. J Appl Microbiol 108:1839-1849.

123
APPENDIX 1: Carbon substrates (31 in total) used for community-level physiological profiles of soils (CLPPs) Carbohydrates Carboxylic
acids Amino acids
Phenolic acids
Aromatics Various phenolics
Vitamins Alcohols
L-rhamnose Formic L-alanine Cinnamic acid
Urocanic acid
Catechin Thiamine D-mannitol
D-galactose L-ascorbic L-asparagine
Imidazole Coumarin
a-ketobutyric L-methionine
Syringic acid
Quercetin
a-ketoglutaric L-valine Fumaric L-
threonine
L-tartaric L-glutamic acid
Glycolic Succinic Malic Citric D-galacturonic Quinic Lactic

124
APPENDIX 2: PUBLICATIONS
NOTE: Journal article removed due to copyright restrictions. Orozco-Aceves, M., Standish, R. J., & Tibbett, M. (2015). Soil conditioning and plant-soil feedbacks in a modified forest ecosystem are soil-context dependent. Plant and Soil, 390(1-2), 183-194. 10.1007/s11104-015-2390-z

Forest Ecology and Management 338 (2015) 92–99
Contents lists available at ScienceDirect
Forest Ecology and Management
journal homepage: www.elsevier .com/ locate/ foreco
Long-term conditioning of soil by plantation eucalypts and pines doesnot affect growth of the native jarrah tree
http://dx.doi.org/10.1016/j.foreco.2014.11.0070378-1127/� 2014 Elsevier B.V. All rights reserved.
Abbreviations: PSF, plant–soil feedback; PLFA, phospholipid-fatty acid profile;CLPP, community level physiological profile; PCO, principal co-ordinate analysis.⇑ Corresponding author at: Cranfield Soil and Agrifood Institute, Cranfield
University, Cranfield, Bedfordshire MK43 0AL, UK. Tel./fax: +44 (0) 1234750111x2991.
E-mail address: [email protected] (M. Tibbett).
Martha Orozco-Aceves a, Rachel J. Standish b, Mark Tibbett a,c,⇑a School of Earth and Environment, University of Western Australia, M087, 35 Stirling Hwy, Crawley, WA 6009, Australiab School of Plant Biology, University of Western Australia, M090, 35 Stirling Hwy, Crawley, WA 6009, Australiac Cranfield Soil and Agrifood Institute, Cranfield University, Cranfield, Bedfordshire MK43 0AL, UK
a r t i c l e i n f o
Article history:Received 3 September 2014Received in revised form 3 November 2014Accepted 11 November 2014
Keywords:Bauxite miningPlant–soil feedbackSoil biologySoil chemistrySoil feedbackRestoration
a b s t r a c t
Plant species can condition the physico-chemical and biological properties of soil in ways that modifyplant growth via plant–soil feedback (PSF). Plant growth can be positively affected, negatively affectedor neutrally affected by soil conditioning by the same or other plant species. Soil conditioning by otherplant species has particular relevance to ecological restoration of historic ecosystems because sites setaside for restoration are often conditioned by other, potentially non-native, plant species. We investigatedchanges in properties of jarrah forest soils after long-term (35 years) conditioning by pines (Pinus radiata),Sydney blue gums (Eucalyptus saligna), both non-native, plantation trees, and jarrah (Eucalyptus margin-ata; dominant native tree). Then, we tested the influence of the conditioned soils on the growth of jarrahseedlings. Blue gums and pines similarly conditioned the physico-chemical properties of soils, which dif-fered from soil conditioning caused by jarrah. Especially important were the differences in conditioning ofthe properties C:N ratio, pH, and available K. The two eucalypt species similarly conditioned the biologicalproperties of soil (i.e. community level physiological profile, numbers of fungal-feeding nematodes,omnivorous nematodes, and nematode channel ratio), and these differed from conditioning caused bypines. Species-specific conditioning of soil did not translate into differences in the amounts of biomassproduced by jarrah seedlings and a neutral PSF was observed. In summary, we found that decades of soilconditioning by non-native plantation trees did not influence the growth of jarrah seedlings and willtherefore not limit restoration of jarrah following the removal of the plantation trees.
� 2014 Elsevier B.V. All rights reserved.
1. Introduction
Plant–soil feedbacks (PSF) can influence the restoration ofnative forests where soil has been conditioned by non-native spe-cies for long periods of time (i.e. agricultural, forestry activities,invader species; Kulmatiski et al., 2006; Jordan et al., 2008;Kulmatiski and Beard, 2008). Specifically, non-native species cancondition soil properties in ways that lead to reduced growth ofnative species (van Grunsven et al., 2007; Perkins and Nowak,2012; Suding et al., 2013), due to alterations of nutrient cyclingprocesses (Bezemer et al., 2006; Kulmatiski et al., 2006; Kardoland Wardle, 2010; Weidenhamer and Callaway, 2010), imbalance
of microbial communities (i.e. free-living organisms, plant-patho-gens and root symbionts; Inderjit and van der Putten (2010)),and negative effects of toxic allelochemicals (Bertin et al., 2003;Badri and Vivanco, 2009; Inderjit and van der Putten, 2010;Weidenhamer and Callaway, 2010). Plant-conditioned propertiesof soil can persist for long periods of time even after the removalof the non-native species (Kardol and Wardle, 2010).
Soil conditioning operates through two mechanisms: (1) leaflitter deposition, and (2) root deposition. In particular, litter qual-ity is a species-specific plant trait (i.e. C:N ratio, lignin and pheno-lic content) that directly influence the chemical and biologicalproperties of soil. Elements released from litter during decompo-sition alter the concentrations of soil macro and micronutrients ina species-specific manner (Hobbie, 1992; Guo and Sims, 1999;Berg and McClaugherty, 2008; Witt and Setälä, 2010; Uselmanet al., 2012). As well, litter deposition drives changes in themicrobial and microfauna (i.e. nematode) community, includingits biomass, composition, and physiological properties (Berg and

M. Orozco-Aceves et al. / Forest Ecology and Management 338 (2015) 92–99 93
McClaugherty, 2008). Generally, plant species that produce labilelitter (i.e. low C:N ratio) such as legumes, promote a microbialcommunity dominated by bacteria and their consumers (i.e. pro-tozoa, bacterial-feeding nematodes). In contrast, plant speciesthat produce recalcitrant litter (i.e. high C:N ratio) such as pinesand eucalypts promote a microbial community dominated byfungi and their consumers (i.e. fungal-feeding nematodes, collem-bolans, oribatid mites; Moore and Hunt, 1988; Scheu et al., 2005;Moore et al., 2007; Holtkamp et al., 2008; Witt and Setälä, 2010).Thus, in this manner soil conditioning occurs via differences inleaf litter quality.
Bauxite has been mined in the jarrah forest of south-west Wes-tern Australia since 1964. Initial attempts to restore mine sitespost-bauxite mining consisted of commercial plantations of fast-growing, exotic softwoods, mainly pines (Pinus radiata) andcypresses (Cupressus sp.), as well as eastern Australian hardwoodeucalypts such as Sydney blue gum (Eucalyptus saligna; Koch,2007a,b). To date there are 3600 ha of plantations of non-nativespecies within the jarrah forest ecoregion that will be harvestedfor timber in the near future and then restored to native jarrah for-est (Conservation Commission of Western Australia, 2012). Atthese sites non-native trees have been conditioning the soil for35 years, causing species-specific changes to soil properties. Here,we tested species-specific conditioning of soil by pines (P. radiata),Sydney blue gums (E. saligna; both non-native, plantation trees),and jarrah (Eucalyptus marginata; dominant native tree). Wasexpected that the conditioning process of soil by the three plantspecies would be primarily driven by litter deposition, thus influ-enced by litter characteristics of each species in particular. Pinesand eucalypts both produce recalcitrant litter; nonetheless, differ-ences in conditioning processes occur between the two generaassociated with changes in the soil chemistry and biology owingto differences in litter quality.
Conditioning of soil by pines via litter deposition includes acid-ification of soil, alteration of mineral concentrations such as N, K,Mg, and Ca, and low microbial activity (Feller, 1983; Lavelle andSpain, 2003; Ehrenfeld et al., 2005). In contrast, allelopathy hasbeen described as a key mechanism driving soil conditioning byeucalypts. Litter extracts from eucalypts inhibit the germinationof a diversity of plant species as well as the growth of microorgan-isms such as nitrifying bacteria (Bezuidenhout and Laing, 2006;Kohli et al., 2006). There is the potential for pines and eucalyptsto condition soils differently but no tests of the influence of theconditioned soils on plant growth, and in particular how condition-ing of forest soil by non-native species will influence the growth ofnative species such as jarrah via PSF are available. Jarrah displays ahigh degree of growth plasticity in response to resource availabilityand the soil environment (Bleby et al., 2009). This plasticity sug-gests that jarrah is capable of producing constant biomass evenunder variable soil conditions.
The aims of this research were to determine what differences (ifany) occurred in the soils after 35 years of conditioning by non-native species, and to determine the effect of conditioning on thesubsequent growth of jarrah (E. marginata) seedlings (i.e. the neteffect of the PSF). We predicted that the plant species (pine, jarrahand blue gum) would condition the soil in a species-specific man-ner (Aponte et al., 2013), however, differences in soil conditioningwould be more marked after pine than eucalypt growth since theformer presents litter with composition that is different from thatof the eucalypts. In contrast, differences in conditioning betweenblue gums and jarrah would be less distinct due to similar litterquality given that both belong to the same genus. The second spe-cific aim was to test the net PSF associated with jarrah plantsgrown in soils conditioned by non-native plants by comparingtheir growth in soils conditioned by jarrah. We predicted that spe-cies-specific conditioning of soil by non-native plants would not
affect the biomass produced by jarrah seedlings (i.e. a neutralPSF would be observed).
2. Materials and methods
2.1. Sampling site and experimental design
Soils for the experiment were collected from a site that hadbeen cleared of jarrah forest, mined for bauxite, and then plantedwith diverse native and non-native eucalypt species, pines andcypresses to create an arboretum in 1976 (i.e. 35 years prior tosampling). Soil samples were collected in June 2011. At the site,six pines (P. radiata: Pinaceae), six jarrah (E. marginata: Myrtaceae)and six blue gums (E. saligna: Myrtaceae) were selected at randomand four intact soil cores (0.785 L; diameter = 10 cm,depth = 10 cm) were taken from beneath the canopy area of eachtree where litter is deposited. Two cores were used to measurethe physico-chemical and biological properties of soils to deter-mine species-specific conditioning. These soils were sieved (2-mm mesh) and stored at 4 �C pending analyses, and a subsampleof soil was frozen (�20 �C) and then subsequently freeze-driedpending phospholipid-fatty acid analysis (PLFA). The two remain-ing cores were placed in bottom-sealed pots (1.2 L; diame-ter = 13.3 cm, height = 12.4 cm) ready to use in the pot experiment.
Pots containing the intact soil cores were placed into a rootcooling tank that was housed in a glasshouse with temperaturesranging between 4.5 and 27.8 �C for the duration of the experi-ment. To produce seedlings, seeds were surface-disinfected withethanol (70%) and bleach (50%) to prevent growth of pathogensand germinated at 14 �C in darkness. Several jarrah seedlings wereplanted in each pot, and once a plant established, the superfluousseedlings were removed leaving only one plant per pot. As a resultthere were jarrah seedlings growing in soil cores collected in thefield from jarrah trees (i.e. home soils) and in soil cores collectedfrom pines or blue gums (i.e. away soils). Pots were rotated withinthe tank every two weeks to negate a block effect and wateredonce a week to 60% water-holding capacity. After four monthsgrowth (from June to October 2011), plants were harvested; shootsand roots were removed, oven-dried for one week at 60 �C andweighed.
2.2. Response variables
2.2.1. Soil chemical propertiesTotal N (%), total C (%), available (bicarbonate extractable) P
(mg kg�1) and K (mg kg�1), pH (CaCl2) and EC (dS m�1) were deter-mined by the commercial laboratory Cummings Smith BritishPetroleum (CSBP, Bibra Lake Western Australia). Total N and totalC were measured in a combustion analyser (LECO� USA), availableP, available K, pH, EC, were determined using standard methodsdescribed in Rayment and Higginson (1992). Ammonium (NH4
+
mg kg�1) and nitrate (NO3� mg kg�1) analyses were done simulta-
neously by the Skalar Autoanalyser (Skalar Analytical, The Nether-lands) in laboratories at the University of Western Australia.
2.2.2. Soil biology and associated properties2.2.2.1. Microbial biomass-carbon (C). Microbial biomass-C wasextracted by chloroform-fumigation technique described inColeman et al. (2004), and then measured with a Total Organic-CAnalyser (Shimadzu 5000A, Japan). For calculations a kEC factor of0.45 was used and data were reported on a dry soil basis (lg C g�1
dry soil).
2.2.2.2. Phospholipid-fatty acid (PLFA) profiles. Phospholipid-fattyacid profiles of soils were determined following the procedure

94 M. Orozco-Aceves et al. / Forest Ecology and Management 338 (2015) 92–99
described in White et al. (1979). Solid phase extraction tubes (sil-ica-bonded polar, 1 mL volume, Supelclean™) were used to sepa-rate phospholipids from neutral lipids and glycolipids.Phospholipids were subjected to a mild alkaline hydrolysis to formfatty acid methyl esters (FAMEs) as described by Zelles and Bai(1993). Individual PLFAs were detected using gas chromatographwith flame ionization detector (Agilent 7890A Dean Switch GCFID).Fifty lL (75 lg mL�1) of methyl-nonadecanoate (Me-C19 Sigma)were added to FAMEs extracts as internal standard. PLFAs wereidentified by comparing their retention times with those ofqualitative standards of methyl esters (Supelco�). To calculateconcentrations of FAMEs, the following formula was used:
lgindividual fatty acid g�1 soil ¼ PFAME � lgStd
PISTD �W
where PFAME and PISTD are the peak areas of fatty acid and the inter-nal standard respectively; lgStd is the concentration of the internalstandard (lg lL�1 solvent), and W is the dry weight of soil used (g).Finally, biomarkers for specific groups of soil organisms such as bac-teria (15:0i, 15:0a, 16:0i, 17:0i, 17:0cy), fungi (18:2x6,9, 18:1x9),and microeucaryotes (22:0, 24:0, 25:0; Zelles, 1999; Treonis et al.,2004; Bezemer et al., 2006) were determined.
2.2.2.3. Community level physiological profile (CLPP) of soils. Todetermine CLPPs of the soils the MicroResp™ methodology wasused (Campbell et al., 2003; Lalor et al., 2007). Types of C sub-strates, as well as concentrations of substrates and reagents in cre-sol red detector plates were the same used by Lalor et al. (2007).After a 6 h incubation period at 25 �C, absorbance of detector plates(at 590 nm) was determined using a microplate plate reader (ASYSHitech, EXPERT 96) and headspace %CO2 was calculated with thecalibration curve equation experimentally obtained utilizing theplate reader mentioned above: 0.6(exp(�9.95(x � 0.42)) + 0.06.Finally the CO2 rate (lg CO2–C g�1 soil h�1) was calculated asfollows:
%CO2100 � vol� 44
22� 1244� 273
ð273þTÞsoilfwt�soil%dwt
100h
where %CO2 is the absorbance value interpolated in the calibrationcurve; 44, 22, 12 and 273 are constants; T is the incubation temper-ature (�C), vol is the headspace volume in microplate wells (ll), soil-fwt is the fresh weight of soil per well (g), h is the incubation time inhours and soil%dwt is soil sample %dry weight.
2.2.2.4. Nematode trophic community. Nematodes were extractedfrom 100 g of fresh soil (per sample) using a mistifier as describedin Hooper et al. (2005). Aqueous extracts were collected and nem-atodes within them were sorted per1 mL extract using a countingcell (Sedgewick Rafter cell, Graticules England) observed at 100�magnification (eyepiece = 10�; lens = 10�) in a compound micro-scope (Olympus Vanox). Nematodes of the following trophicgroups were identified and counted according to morphologicalstructures described in Yeates et al. (1993): bacterial feeders (BF;Rhabditida, Enoplida), fungal feeders (FF; Tylenchida), omnivorous(Dorylaimida), predators (Mononchida) and plant parasites(Tylenchida, Dorylaimida). Data were expressed as the number ofnematodes of each group per 100 g of dry soil. Additionally, thenematode channel ratio (FF/FF + BF) was calculated as an indicatorof the decomposition channel (i.e. bacterial or fungal) working insoils (Yeates, 2003).
2.3. Statistical analyses
To determine the evidence for species-specific conditioning offield soils the physico-chemical and biological properties (includ-
ing total PLFAs and specific biomarkers for groups of organisms,CLPPs and nematode trophic groups) of soils were compared usingmultivariate and univariate permutational analyses of variance(PERMANOVA) followed by pair-wise tests. Although PERMANOVAis a multivariate method, it can be used for univariate analyses.Performing PERMANOVA using one response variable and Euclid-ean distance yields Fisher’s traditional univariate F-statistic(Anderson, 2010). Scaling plots from principal co-ordinate analysis(PCO; Anderson, 2001) were built to visualize the results of PER-MANOVA analyses. Vectors showing the properties causing themajor effects on the separation of clusters along the main PCOs(i.e. Pearson correlation coefficient with PCO1 and PCO2 > 0.7)were included in some of the plots.
To determine the net effects of the PSF (i.e. magnitude anddirection), we used univariate PERMANOVAs with total, shootand root biomass produced in home vs. away soil as response vari-ables. A significant increase of the plant biomass production inhome with respect to away soil was considered a net positivePSF. A significant decrease of the plant biomass production inhome soil with respect to away soil was considered a net negativePSF. No difference in biomass production in home with respect toaway soil was considered a neutral PSF.
For all the statistical analyses PRIMER v6 + PERMANOVA (PRI-MER-E Ltd) was used. For the multivariate analyses, permutationof residuals under a reduced model was selected as permutationmethod as well as type III (partial) sum of squares. The unrestrictedpermutation of raw data as permutation method and type III (par-tial) sum of squares were used for the univariate analyses. In allcases data were transformed when necessary and Euclidean dis-tance was used to build resemblance matrices, except for the anal-yses of CLPPs and the trophic groups of the nematode communitywhen Bray–Curtis coefficient and Manhattan distance were respec-tively utilized to build the similarity matrices.
3. Results
3.1. Properties of soils conditioned in the field by pines, jarrah, andblue gums
The physico-chemical properties of soils conditioned in the fieldwere significantly different according to plant species (F2,26 = 2.83,P = 0.003; Fig. 1a). Pair-wise comparisons showed that the physico-chemical properties of soils conditioned by jarrah differed fromthose conditioned by blue gums (t = 1.74, P = 0.031), and pines(t = 1.91, P = 0.034), and that soils conditioned by blue gums andpines were not significantly different from each other (t = 1.36,P = 0.06). The biological properties of field soils were differentiallyconditioned according to plant species (F2,16 = 2.24, P = 0.007;Fig. 1b). Pair-wise comparisons showed that the biological proper-ties of soils conditioned by pines differed from those conditionedby jarrah (t = 1.72, P = 0.01) and soils conditioned by blue gums(t = 1.73, P = 0.013), and that soils conditioned by jarrah and bluegums were not significantly different from each other (t = 1.005,P = 0.45).
Statistical analyses of individual physico-chemical properties(Table 1) revealed differences in species-specific conditioning ofthe C:N ratio (F2,2.34 = 21.11, P = 0.0002), pH (F2,0.04 = 3.88,P = 0.042) and K concentration (F2,37.7 = 18.2, P = 0.001). The C:Nratio of soils conditioned by the three species were all significantlydifferent (jarrah versus pine t = 6.02, P = 0.0016; jarrah versus bluegum t = 3.52, P = 0.0081; pine versus blue gum t = 3.24, P = 0.0113).Soil pH conditioned by blue gums statistically differed from pH ofsoils conditioned by pines (t = 2.78, P = 0.024), and K concentrationof soils conditioned by the three species were statistically differentfrom each other (jarrah versus pine t = 2.63, P = 0.042; jarrah
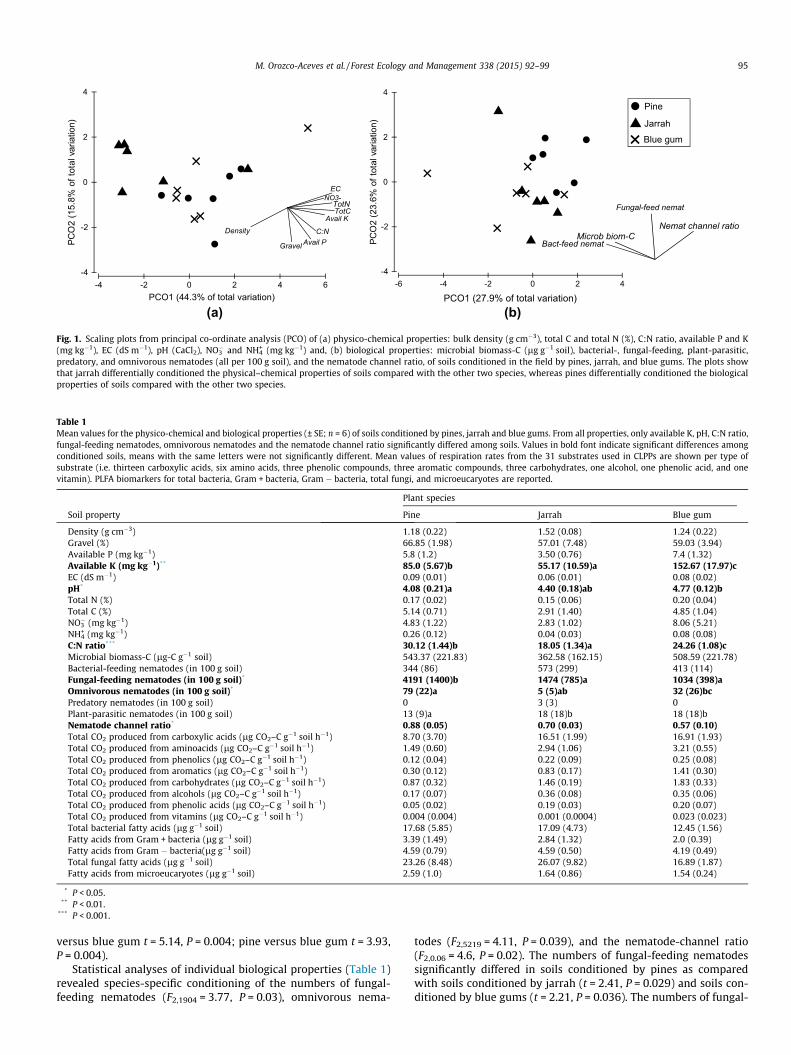
(a) (b)
Fig. 1. Scaling plots from principal co-ordinate analysis (PCO) of (a) physico-chemical properties: bulk density (g cm�3), total C and total N (%), C:N ratio, available P and K(mg kg�1), EC (dS m�1), pH (CaCl2), NO3
� and NH4+ (mg kg�1) and, (b) biological properties: microbial biomass-C (lg g�1 soil), bacterial-, fungal-feeding, plant-parasitic,
predatory, and omnivorous nematodes (all per 100 g soil), and the nematode channel ratio, of soils conditioned in the field by pines, jarrah, and blue gums. The plots showthat jarrah differentially conditioned the physical–chemical properties of soils compared with the other two species, whereas pines differentially conditioned the biologicalproperties of soils compared with the other two species.
Table 1Mean values for the physico-chemical and biological properties (± SE; n = 6) of soils conditioned by pines, jarrah and blue gums. From all properties, only available K, pH, C:N ratio,fungal-feeding nematodes, omnivorous nematodes and the nematode channel ratio significantly differed among soils. Values in bold font indicate significant differences amongconditioned soils, means with the same letters were not significantly different. Mean values of respiration rates from the 31 substrates used in CLPPs are shown per type ofsubstrate (i.e. thirteen carboxylic acids, six amino acids, three phenolic compounds, three aromatic compounds, three carbohydrates, one alcohol, one phenolic acid, and onevitamin). PLFA biomarkers for total bacteria, Gram + bacteria, Gram � bacteria, total fungi, and microeucaryotes are reported.
Plant species
Soil property Pine Jarrah Blue gum
Density (g cm�3) 1.18 (0.22) 1.52 (0.08) 1.24 (0.22)Gravel (%) 66.85 (1.98) 57.01 (7.48) 59.03 (3.94)Available P (mg kg�1) 5.8 (1.2) 3.50 (0.76) 7.4 (1.32)Available K (mg kg�1)** 85.0 (5.67)b 55.17 (10.59)a 152.67 (17.97)cEC (dS m�1) 0.09 (0.01) 0.06 (0.01) 0.08 (0.02)pH* 4.08 (0.21)a 4.40 (0.18)ab 4.77 (0.12)bTotal N (%) 0.17 (0.02) 0.15 (0.06) 0.20 (0.04)Total C (%) 5.14 (0.71) 2.91 (1.40) 4.85 (1.04)NO3� (mg kg�1) 4.83 (1.22) 2.83 (1.02) 8.06 (5.21)
NH4+ (mg kg�1) 0.26 (0.12) 0.04 (0.03) 0.08 (0.08)
C:N ratio*** 30.12 (1.44)b 18.05 (1.34)a 24.26 (1.08)cMicrobial biomass-C (lg-C g�1 soil) 543.37 (221.83) 362.58 (162.15) 508.59 (221.78)Bacterial-feeding nematodes (in 100 g soil) 344 (86) 573 (299) 413 (114)Fungal-feeding nematodes (in 100 g soil)* 4191 (1400)b 1474 (785)a 1034 (398)aOmnivorous nematodes (in 100 g soil)* 79 (22)a 5 (5)ab 32 (26)bcPredatory nematodes (in 100 g soil) 0 3 (3) 0Plant-parasitic nematodes (in 100 g soil) 13 (9)a 18 (18)b 18 (18)bNematode channel ratio* 0.88 (0.05) 0.70 (0.03) 0.57 (0.10)Total CO2 produced from carboxylic acids (lg CO2–C g�1 soil h�1) 8.70 (3.70) 16.51 (1.99) 16.91 (1.93)Total CO2 produced from aminoacids (lg CO2–C g�1 soil h�1) 1.49 (0.60) 2.94 (1.06) 3.21 (0.55)Total CO2 produced from phenolics (lg CO2–C g�1 soil h�1) 0.12 (0.04) 0.22 (0.09) 0.25 (0.08)Total CO2 produced from aromatics (lg CO2–C g�1 soil h�1) 0.30 (0.12) 0.83 (0.17) 1.41 (0.30)Total CO2 produced from carbohydrates (lg CO2–C g�1 soil h�1) 0.87 (0.32) 1.46 (0.19) 1.83 (0.33)Total CO2 produced from alcohols (lg CO2–C g�1 soil h�1) 0.17 (0.07) 0.36 (0.08) 0.35 (0.06)Total CO2 produced from phenolic acids (lg CO2–C g�1 soil h�1) 0.05 (0.02) 0.19 (0.03) 0.20 (0.07)Total CO2 produced from vitamins (lg CO2–C g�1 soil h�1) 0.004 (0.004) 0.001 (0.0004) 0.023 (0.023)Total bacterial fatty acids (lg g�1 soil) 17.68 (5.85) 17.09 (4.73) 12.45 (1.56)Fatty acids from Gram + bacteria (lg g�1 soil) 3.39 (1.49) 2.84 (1.32) 2.0 (0.39)Fatty acids from Gram � bacteria(lg g�1 soil) 4.59 (0.79) 4.59 (0.50) 4.19 (0.49)Total fungal fatty acids (lg g�1 soil) 23.26 (8.48) 26.07 (9.82) 16.89 (1.87)Fatty acids from microeucaryotes (lg g�1 soil) 2.59 (1.0) 1.64 (0.86) 1.54 (0.24)
* P < 0.05.** P < 0.01.
*** P < 0.001.
M. Orozco-Aceves et al. / Forest Ecology and Management 338 (2015) 92–99 95
versus blue gum t = 5.14, P = 0.004; pine versus blue gum t = 3.93,P = 0.004).
Statistical analyses of individual biological properties (Table 1)revealed species-specific conditioning of the numbers of fungal-feeding nematodes (F2,1904 = 3.77, P = 0.03), omnivorous nema-
todes (F2,5219 = 4.11, P = 0.039), and the nematode-channel ratio(F2,0.06 = 4.6, P = 0.02). The numbers of fungal-feeding nematodessignificantly differed in soils conditioned by pines as comparedwith soils conditioned by jarrah (t = 2.41, P = 0.029) and soils con-ditioned by blue gums (t = 2.21, P = 0.036). The numbers of fungal-
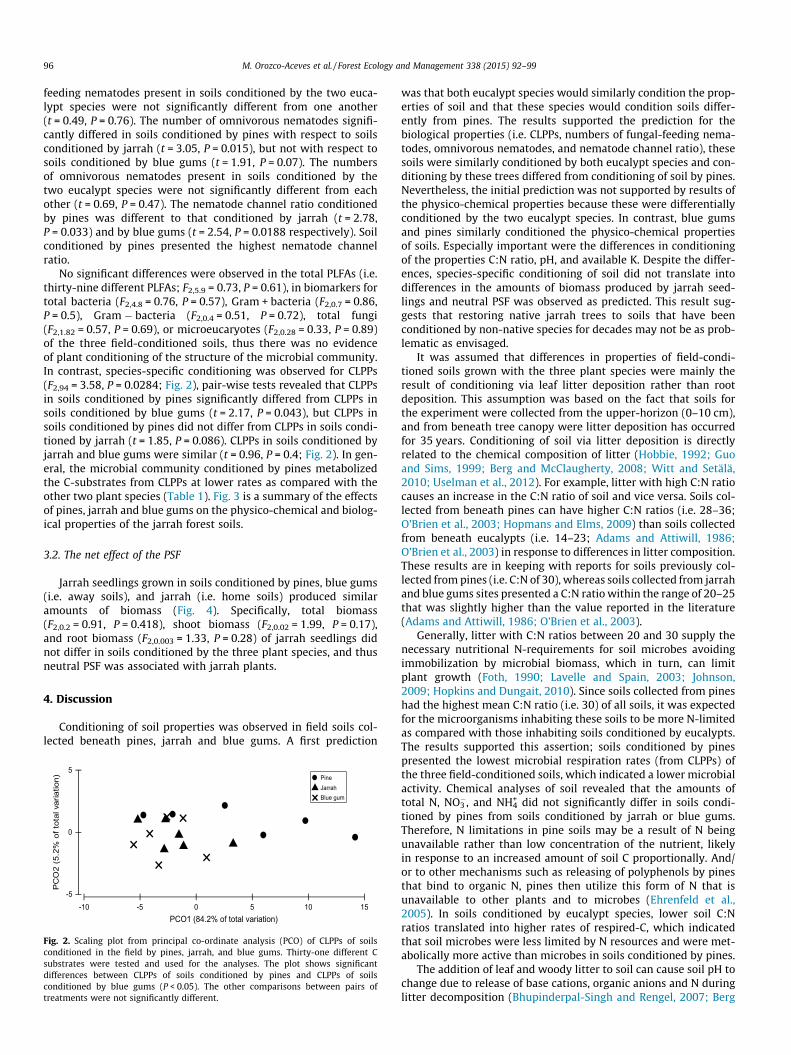
96 M. Orozco-Aceves et al. / Forest Ecology and Management 338 (2015) 92–99
feeding nematodes present in soils conditioned by the two euca-lypt species were not significantly different from one another(t = 0.49, P = 0.76). The number of omnivorous nematodes signifi-cantly differed in soils conditioned by pines with respect to soilsconditioned by jarrah (t = 3.05, P = 0.015), but not with respect tosoils conditioned by blue gums (t = 1.91, P = 0.07). The numbersof omnivorous nematodes present in soils conditioned by thetwo eucalypt species were not significantly different from eachother (t = 0.69, P = 0.47). The nematode channel ratio conditionedby pines was different to that conditioned by jarrah (t = 2.78,P = 0.033) and by blue gums (t = 2.54, P = 0.0188 respectively). Soilconditioned by pines presented the highest nematode channelratio.
No significant differences were observed in the total PLFAs (i.e.thirty-nine different PLFAs; F2,5.9 = 0.73, P = 0.61), in biomarkers fortotal bacteria (F2,4.8 = 0.76, P = 0.57), Gram + bacteria (F2,0.7 = 0.86,P = 0.5), Gram � bacteria (F2,0.4 = 0.51, P = 0.72), total fungi(F2,1.82 = 0.57, P = 0.69), or microeucaryotes (F2,0.28 = 0.33, P = 0.89)of the three field-conditioned soils, thus there was no evidenceof plant conditioning of the structure of the microbial community.In contrast, species-specific conditioning was observed for CLPPs(F2,94 = 3.58, P = 0.0284; Fig. 2), pair-wise tests revealed that CLPPsin soils conditioned by pines significantly differed from CLPPs insoils conditioned by blue gums (t = 2.17, P = 0.043), but CLPPs insoils conditioned by pines did not differ from CLPPs in soils condi-tioned by jarrah (t = 1.85, P = 0.086). CLPPs in soils conditioned byjarrah and blue gums were similar (t = 0.96, P = 0.4; Fig. 2). In gen-eral, the microbial community conditioned by pines metabolizedthe C-substrates from CLPPs at lower rates as compared with theother two plant species (Table 1). Fig. 3 is a summary of the effectsof pines, jarrah and blue gums on the physico-chemical and biolog-ical properties of the jarrah forest soils.
3.2. The net effect of the PSF
Jarrah seedlings grown in soils conditioned by pines, blue gums(i.e. away soils), and jarrah (i.e. home soils) produced similaramounts of biomass (Fig. 4). Specifically, total biomass(F2,0.2 = 0.91, P = 0.418), shoot biomass (F2,0.02 = 1.99, P = 0.17),and root biomass (F2,0.003 = 1.33, P = 0.28) of jarrah seedlings didnot differ in soils conditioned by the three plant species, and thusneutral PSF was associated with jarrah plants.
4. Discussion
Conditioning of soil properties was observed in field soils col-lected beneath pines, jarrah and blue gums. A first prediction
Fig. 2. Scaling plot from principal co-ordinate analysis (PCO) of CLPPs of soilsconditioned in the field by pines, jarrah, and blue gums. Thirty-one different Csubstrates were tested and used for the analyses. The plot shows significantdifferences between CLPPs of soils conditioned by pines and CLPPs of soilsconditioned by blue gums (P < 0.05). The other comparisons between pairs oftreatments were not significantly different.
was that both eucalypt species would similarly condition the prop-erties of soil and that these species would condition soils differ-ently from pines. The results supported the prediction for thebiological properties (i.e. CLPPs, numbers of fungal-feeding nema-todes, omnivorous nematodes, and nematode channel ratio), thesesoils were similarly conditioned by both eucalypt species and con-ditioning by these trees differed from conditioning of soil by pines.Nevertheless, the initial prediction was not supported by results ofthe physico-chemical properties because these were differentiallyconditioned by the two eucalypt species. In contrast, blue gumsand pines similarly conditioned the physico-chemical propertiesof soils. Especially important were the differences in conditioningof the properties C:N ratio, pH, and available K. Despite the differ-ences, species-specific conditioning of soil did not translate intodifferences in the amounts of biomass produced by jarrah seed-lings and neutral PSF was observed as predicted. This result sug-gests that restoring native jarrah trees to soils that have beenconditioned by non-native species for decades may not be as prob-lematic as envisaged.
It was assumed that differences in properties of field-condi-tioned soils grown with the three plant species were mainly theresult of conditioning via leaf litter deposition rather than rootdeposition. This assumption was based on the fact that soils forthe experiment were collected from the upper-horizon (0–10 cm),and from beneath tree canopy were litter deposition has occurredfor 35 years. Conditioning of soil via litter deposition is directlyrelated to the chemical composition of litter (Hobbie, 1992; Guoand Sims, 1999; Berg and McClaugherty, 2008; Witt and Setälä,2010; Uselman et al., 2012). For example, litter with high C:N ratiocauses an increase in the C:N ratio of soil and vice versa. Soils col-lected from beneath pines can have higher C:N ratios (i.e. 28–36;O’Brien et al., 2003; Hopmans and Elms, 2009) than soils collectedfrom beneath eucalypts (i.e. 14–23; Adams and Attiwill, 1986;O’Brien et al., 2003) in response to differences in litter composition.These results are in keeping with reports for soils previously col-lected from pines (i.e. C:N of 30), whereas soils collected from jarrahand blue gums sites presented a C:N ratio within the range of 20–25that was slightly higher than the value reported in the literature(Adams and Attiwill, 1986; O’Brien et al., 2003).
Generally, litter with C:N ratios between 20 and 30 supply thenecessary nutritional N-requirements for soil microbes avoidingimmobilization by microbial biomass, which in turn, can limitplant growth (Foth, 1990; Lavelle and Spain, 2003; Johnson,2009; Hopkins and Dungait, 2010). Since soils collected from pineshad the highest mean C:N ratio (i.e. 30) of all soils, it was expectedfor the microorganisms inhabiting these soils to be more N-limitedas compared with those inhabiting soils conditioned by eucalypts.The results supported this assertion; soils conditioned by pinespresented the lowest microbial respiration rates (from CLPPs) ofthe three field-conditioned soils, which indicated a lower microbialactivity. Chemical analyses of soil revealed that the amounts oftotal N, NO3
�, and NH4+ did not significantly differ in soils condi-
tioned by pines from soils conditioned by jarrah or blue gums.Therefore, N limitations in pine soils may be a result of N beingunavailable rather than low concentration of the nutrient, likelyin response to an increased amount of soil C proportionally. And/or to other mechanisms such as releasing of polyphenols by pinesthat bind to organic N, pines then utilize this form of N that isunavailable to other plants and to microbes (Ehrenfeld et al.,2005). In soils conditioned by eucalypt species, lower soil C:Nratios translated into higher rates of respired-C, which indicatedthat soil microbes were less limited by N resources and were met-abolically more active than microbes in soils conditioned by pines.
The addition of leaf and woody litter to soil can cause soil pH tochange due to release of base cations, organic anions and N duringlitter decomposition (Bhupinderpal-Singh and Rengel, 2007; Berg
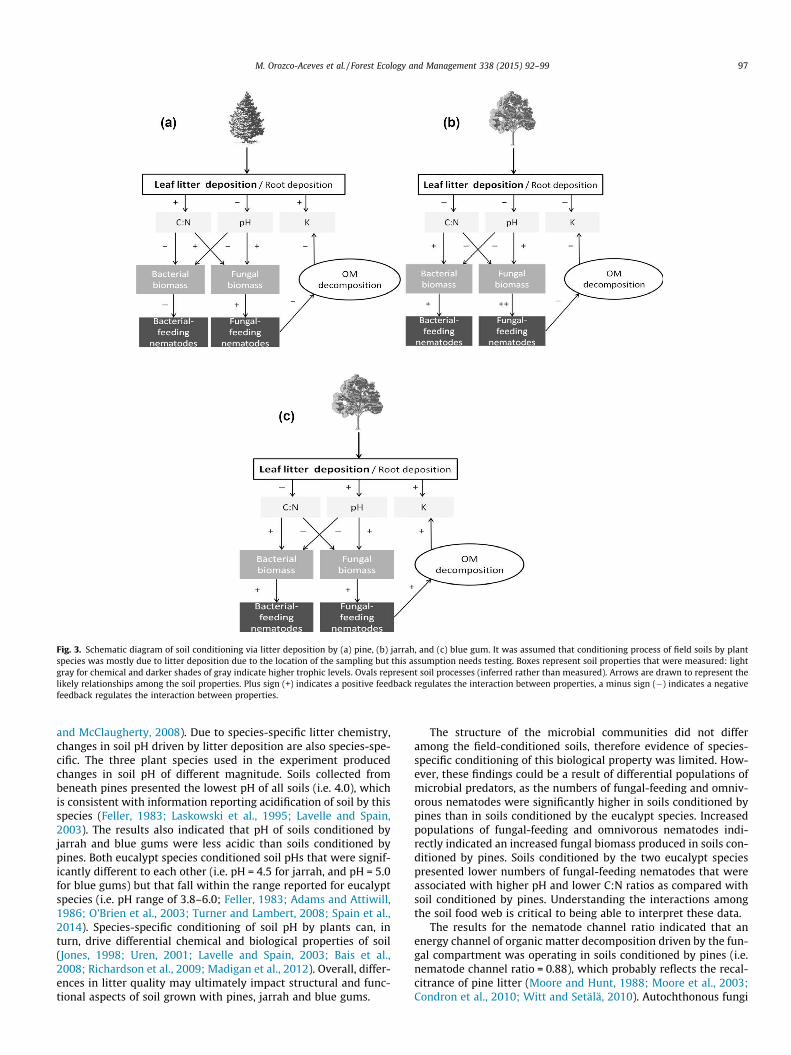
Fig. 3. Schematic diagram of soil conditioning via litter deposition by (a) pine, (b) jarrah, and (c) blue gum. It was assumed that conditioning process of field soils by plantspecies was mostly due to litter deposition due to the location of the sampling but this assumption needs testing. Boxes represent soil properties that were measured: lightgray for chemical and darker shades of gray indicate higher trophic levels. Ovals represent soil processes (inferred rather than measured). Arrows are drawn to represent thelikely relationships among the soil properties. Plus sign (+) indicates a positive feedback regulates the interaction between properties, a minus sign (�) indicates a negativefeedback regulates the interaction between properties.
M. Orozco-Aceves et al. / Forest Ecology and Management 338 (2015) 92–99 97
and McClaugherty, 2008). Due to species-specific litter chemistry,changes in soil pH driven by litter deposition are also species-spe-cific. The three plant species used in the experiment producedchanges in soil pH of different magnitude. Soils collected frombeneath pines presented the lowest pH of all soils (i.e. 4.0), whichis consistent with information reporting acidification of soil by thisspecies (Feller, 1983; Laskowski et al., 1995; Lavelle and Spain,2003). The results also indicated that pH of soils conditioned byjarrah and blue gums were less acidic than soils conditioned bypines. Both eucalypt species conditioned soil pHs that were signif-icantly different to each other (i.e. pH = 4.5 for jarrah, and pH = 5.0for blue gums) but that fall within the range reported for eucalyptspecies (i.e. pH range of 3.8–6.0; Feller, 1983; Adams and Attiwill,1986; O’Brien et al., 2003; Turner and Lambert, 2008; Spain et al.,2014). Species-specific conditioning of soil pH by plants can, inturn, drive differential chemical and biological properties of soil(Jones, 1998; Uren, 2001; Lavelle and Spain, 2003; Bais et al.,2008; Richardson et al., 2009; Madigan et al., 2012). Overall, differ-ences in litter quality may ultimately impact structural and func-tional aspects of soil grown with pines, jarrah and blue gums.
The structure of the microbial communities did not differamong the field-conditioned soils, therefore evidence of species-specific conditioning of this biological property was limited. How-ever, these findings could be a result of differential populations ofmicrobial predators, as the numbers of fungal-feeding and omniv-orous nematodes were significantly higher in soils conditioned bypines than in soils conditioned by the eucalypt species. Increasedpopulations of fungal-feeding and omnivorous nematodes indi-rectly indicated an increased fungal biomass produced in soils con-ditioned by pines. Soils conditioned by the two eucalypt speciespresented lower numbers of fungal-feeding nematodes that wereassociated with higher pH and lower C:N ratios as compared withsoil conditioned by pines. Understanding the interactions amongthe soil food web is critical to being able to interpret these data.
The results for the nematode channel ratio indicated that anenergy channel of organic matter decomposition driven by the fun-gal compartment was operating in soils conditioned by pines (i.e.nematode channel ratio = 0.88), which probably reflects the recal-citrance of pine litter (Moore and Hunt, 1988; Moore et al., 2003;Condron et al., 2010; Witt and Setälä, 2010). Autochthonous fungi
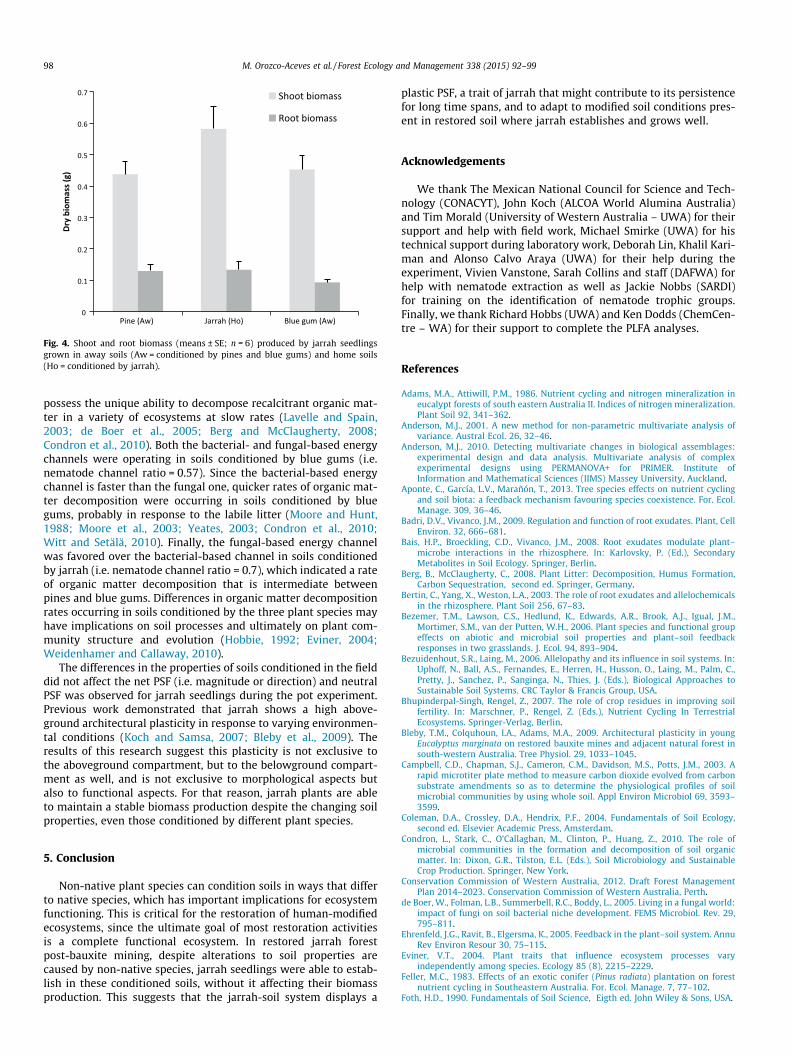
0
0.1
0.2
0.3
0.4
0.5
0.6
0.7
Pine (Aw) Jarrah (Ho) Blue gum (Aw)
Dry
biom
ass (
g)
Shoot biomass
Root biomass
Fig. 4. Shoot and root biomass (means ± SE; n = 6) produced by jarrah seedlingsgrown in away soils (Aw = conditioned by pines and blue gums) and home soils(Ho = conditioned by jarrah).
98 M. Orozco-Aceves et al. / Forest Ecology and Management 338 (2015) 92–99
possess the unique ability to decompose recalcitrant organic mat-ter in a variety of ecosystems at slow rates (Lavelle and Spain,2003; de Boer et al., 2005; Berg and McClaugherty, 2008;Condron et al., 2010). Both the bacterial- and fungal-based energychannels were operating in soils conditioned by blue gums (i.e.nematode channel ratio = 0.57). Since the bacterial-based energychannel is faster than the fungal one, quicker rates of organic mat-ter decomposition were occurring in soils conditioned by bluegums, probably in response to the labile litter (Moore and Hunt,1988; Moore et al., 2003; Yeates, 2003; Condron et al., 2010;Witt and Setälä, 2010). Finally, the fungal-based energy channelwas favored over the bacterial-based channel in soils conditionedby jarrah (i.e. nematode channel ratio = 0.7), which indicated a rateof organic matter decomposition that is intermediate betweenpines and blue gums. Differences in organic matter decompositionrates occurring in soils conditioned by the three plant species mayhave implications on soil processes and ultimately on plant com-munity structure and evolution (Hobbie, 1992; Eviner, 2004;Weidenhamer and Callaway, 2010).
The differences in the properties of soils conditioned in the fielddid not affect the net PSF (i.e. magnitude or direction) and neutralPSF was observed for jarrah seedlings during the pot experiment.Previous work demonstrated that jarrah shows a high above-ground architectural plasticity in response to varying environmen-tal conditions (Koch and Samsa, 2007; Bleby et al., 2009). Theresults of this research suggest this plasticity is not exclusive tothe aboveground compartment, but to the belowground compart-ment as well, and is not exclusive to morphological aspects butalso to functional aspects. For that reason, jarrah plants are ableto maintain a stable biomass production despite the changing soilproperties, even those conditioned by different plant species.
5. Conclusion
Non-native plant species can condition soils in ways that differto native species, which has important implications for ecosystemfunctioning. This is critical for the restoration of human-modifiedecosystems, since the ultimate goal of most restoration activitiesis a complete functional ecosystem. In restored jarrah forestpost-bauxite mining, despite alterations to soil properties arecaused by non-native species, jarrah seedlings were able to estab-lish in these conditioned soils, without it affecting their biomassproduction. This suggests that the jarrah-soil system displays a
plastic PSF, a trait of jarrah that might contribute to its persistencefor long time spans, and to adapt to modified soil conditions pres-ent in restored soil where jarrah establishes and grows well.
Acknowledgements
We thank The Mexican National Council for Science and Tech-nology (CONACYT), John Koch (ALCOA World Alumina Australia)and Tim Morald (University of Western Australia – UWA) for theirsupport and help with field work, Michael Smirke (UWA) for histechnical support during laboratory work, Deborah Lin, Khalil Kari-man and Alonso Calvo Araya (UWA) for their help during theexperiment, Vivien Vanstone, Sarah Collins and staff (DAFWA) forhelp with nematode extraction as well as Jackie Nobbs (SARDI)for training on the identification of nematode trophic groups.Finally, we thank Richard Hobbs (UWA) and Ken Dodds (ChemCen-tre – WA) for their support to complete the PLFA analyses.
References
Adams, M.A., Attiwill, P.M., 1986. Nutrient cycling and nitrogen mineralization ineucalypt forests of south eastern Australia II. Indices of nitrogen mineralization.Plant Soil 92, 341–362.
Anderson, M.J., 2001. A new method for non-parametric multivariate analysis ofvariance. Austral Ecol. 26, 32–46.
Anderson, M.J., 2010. Detecting multivariate changes in biological assemblages:experimental design and data analysis. Multivariate analysis of complexexperimental designs using PERMANOVA+ for PRIMER. Institute ofInformation and Mathematical Sciences (IIMS) Massey University, Auckland.
Aponte, C., García, L.V., Marañón, T., 2013. Tree species effects on nutrient cyclingand soil biota: a feedback mechanism favouring species coexistence. For. Ecol.Manage. 309, 36–46.
Badri, D.V., Vivanco, J.M., 2009. Regulation and function of root exudates. Plant, CellEnviron. 32, 666–681.
Bais, H.P., Broeckling, C.D., Vivanco, J.M., 2008. Root exudates modulate plant–microbe interactions in the rhizosphere. In: Karlovsky, P. (Ed.), SecondaryMetabolites in Soil Ecology. Springer, Berlin.
Berg, B., McClaugherty, C., 2008. Plant Litter: Decomposition, Humus Formation,Carbon Sequestration, second ed. Springer, Germany.
Bertin, C., Yang, X., Weston, L.A., 2003. The role of root exudates and allelochemicalsin the rhizosphere. Plant Soil 256, 67–83.
Bezemer, T.M., Lawson, C.S., Hedlund, K., Edwards, A.R., Brook, A.J., Igual, J.M.,Mortimer, S.M., van der Putten, W.H., 2006. Plant species and functional groupeffects on abiotic and microbial soil properties and plant–soil feedbackresponses in two grasslands. J. Ecol. 94, 893–904.
Bezuidenhout, S.R., Laing, M., 2006. Allelopathy and its influence in soil systems. In:Uphoff, N., Ball, A.S., Fernandes, E., Herren, H., Husson, O., Laing, M., Palm, C.,Pretty, J., Sanchez, P., Sanginga, N., Thies, J. (Eds.), Biological Approaches toSustainable Soil Systems. CRC Taylor & Francis Group, USA.
Bhupinderpal-Singh, Rengel, Z., 2007. The role of crop residues in improving soilfertility. In: Marschner, P., Rengel, Z. (Eds.), Nutrient Cycling In TerrestrialEcosystems. Springer-Verlag, Berlin.
Bleby, T.M., Colquhoun, I.A., Adams, M.A., 2009. Architectural plasticity in youngEucalyptus marginata on restored bauxite mines and adjacent natural forest insouth-western Australia. Tree Physiol. 29, 1033–1045.
Campbell, C.D., Chapman, S.J., Cameron, C.M., Davidson, M.S., Potts, J.M., 2003. Arapid microtiter plate method to measure carbon dioxide evolved from carbonsubstrate amendments so as to determine the physiological profiles of soilmicrobial communities by using whole soil. Appl Environ Microbiol 69, 3593–3599.
Coleman, D.A., Crossley, D.A., Hendrix, P.F., 2004. Fundamentals of Soil Ecology,second ed. Elsevier Academic Press, Amsterdam.
Condron, L., Stark, C., O’Callaghan, M., Clinton, P., Huang, Z., 2010. The role ofmicrobial communities in the formation and decomposition of soil organicmatter. In: Dixon, G.R., Tilston, E.L. (Eds.), Soil Microbiology and SustainableCrop Production. Springer, New York.
Conservation Commission of Western Australia, 2012. Draft Forest ManagementPlan 2014–2023. Conservation Commission of Western Australia, Perth.
de Boer, W., Folman, L.B., Summerbell, R.C., Boddy, L., 2005. Living in a fungal world:impact of fungi on soil bacterial niche development. FEMS Microbiol. Rev. 29,795–811.
Ehrenfeld, J.G., Ravit, B., Elgersma, K., 2005. Feedback in the plant–soil system. AnnuRev Environ Resour 30, 75–115.
Eviner, V.T., 2004. Plant traits that influence ecosystem processes varyindependently among species. Ecology 85 (8), 2215–2229.
Feller, M.C., 1983. Effects of an exotic conifer (Pinus radiata) plantation on forestnutrient cycling in Southeastern Australia. For. Ecol. Manage. 7, 77–102.
Foth, H.D., 1990. Fundamentals of Soil Science, Eigth ed. John Wiley & Sons, USA.

M. Orozco-Aceves et al. / Forest Ecology and Management 338 (2015) 92–99 99
Guo, L.B., Sims, R.E.H., 1999. Litter decomposition and nutrient release via litterdecomposition in New Zealand eucalypt short rotation forests. Agric EcosystEnviron 75, 133–140.
Hobbie, S.E., 1992. Effects of plant species on nutrient cycling. Trends Ecol. Evol. 7(10), 336–339.
Holtkamp, R., Kardol, P., van der Wal, A., Dekker, S.C., van der Putten, W.H., deRuiter, P.C., 2008. Soil food web structure during ecosystem development afterland abandonment. Appl Soil Ecol 39, 23–34.
Hooper, D.J., Hallmann, J., Subbotin, S.A., 2005. Methods for extraction, processingand detection of plant and soil nematodes. In: Luc, M., Sikora, R.A., Bridge, J.(Eds.), Plant Parasitic Nematodes in Subtropical and Tropical Agriculture,second ed. CAB Publishing, UK.
Hopkins, D.W., Dungait, J.A.J., 2010. Soil microbiology and nutrient cycling. In:Dixon, G.R., Tilston, E.L. (Eds.), Soil Microbiology and Sustainable CropProduction. Springer, New York.
Hopmans, P., Elms, S.R., 2009. Changes in total carbon and nutrients in soil profilesand accumulation in biomass after a 30-year rotation of Pinus radiata onpodzolized sands: impacts of intensive harvesting on soil resources. For. Ecol.Manage. 258, 2183–2193.
Inderjit, van der Putten, W.H., 2010. Impacts of soil microbial communities onexotic plant invasions. Trends Ecol. Evol. 25, 512–519.
Johnson, C., 2009. Biology of Soil Science. Oxford Book Company, Delhi.Jones, D.L., 1998. Organic acids in the rhizosphere – a critical review. Plant Soil 205,
25–44.Jordan, N.R., Larson, D.L., Huerd, S.C., 2008. Soil modification by invasive plants:
effects on native and invasive species of mixed-grass prairies. Biol. Invasions 10,177–190.
Kardol, P., Wardle, D.A., 2010. How understanding aboveground–belowgroundlinkages can assist restoration ecology. Trends Ecol. Evol. 25 (11), 670–679.
Koch, J.M., 2007a. Bauxite mining restoration by Alcoa World Alumina Australia inWestern Australia: social, political, historical, and environmental contexts.Restor. Ecol. 15 (4), S3–S10.
Koch, J.M., 2007b. Alcoa’s mining and restoration process in south WesternAustralia. Restor. Ecol. 15 (4), S11–S16.
Koch, J.M., Samsa, G., 2007. Restoring jarrah forest trees after bauxite mining inWestern Australia. Restor. Ecol. 15 (4), S17–S25.
Kohli, R.K., Batish, D.R., Singh, H.P., 2006. Allelopathic interactions inagroecosystems. In: Reigosa, M.J., Pedrol, N., González, L. (Eds.), Allelopathy: APhysiological Process With Ecological Implications. Springer, The Netherlands.
Kulmatiski, A., Beard, K.H., Stark, J.M., 2006. Soil history as a primary control onplant invasion in abandoned agricultural fields. J. Appl. Ecol. 43, 868–876.
Kulmatiski, A., Beard, H.K., 2008. Decoupling plant-growth from land-use legacies insoil microbial communities. Soil Biol. Biochem. 40, 1059–1068.
Lalor, E.L., Cookson, W.R., Murphy, D.V., 2007. Comparison of two methods thatassess soil community level physiological profiles in a forest ecosystem. SoilBiol. Biochem. 39, 454–462.
Laskowski, R., Niklinska, M., Maryanski, M., 1995. The dynamics of chemicalelements in forest litter. Ecology 76 (5), 1393–1406.
Lavelle, P., Spain, A.V., 2003. Soil Ecology. Kluwer Academic Publishers, USA.Madigan, M.T., Martinko, J.M., Stahl, D.A., Clark, D.P., 2012. Brock Biology of
Microorganisms, Thirteenth ed. Benjamin Cummings Editorial, USA.Moore, J.C., Hunt, H.W., 1988. Resource compartmentation and the stability of real
ecosystems. Nature 33, 261–263.Moore, J.C., McCann, K., Setäla, H., de Ruiter, P.C., 2003. Top-down is bottom-up:
does predation in the rhizosphere regulate aboveground dynamics? Ecology 84,846–857.
Moore, J.C., McCann, K., de Ruiter, P.C., 2007. Soil rhizosphere food webs, theirstability, and implications for soil processes in ecosystems. In: Cardon, Z.G.,Whitbeck, J.L. (Eds.), The Rhizosphere: An Ecological Perspective. Elsevier, USA.
O’Brien, N.D., Attiwill, P.M., Weston, C.J., 2003. Stability of soil organic matter inEucalyptus regnans forests and Pinus radiata plantations in south easternAustralia. For. Ecol. Manage. 185, 249–261.
Perkins, L.B., Nowak, R.S., 2012. Soil conditioning and plant–soil feedbacks affectcompetitive relationships between native and invasive grasses. Plant Ecol. 213,1337–1344.
Rayment, G.E., Higginson, F.R., 1992. Australian Laboratory Handbook of Soil andWater Chemical Methods: Australian Soil and Land Survey Handbook, vol. 3.Inkata Press, Australia.
Richardson, A.E., Barea, J.M., McNeill, A.M., Prigent-Combaret, C., 2009. Acquisitionof phosphorus and nitrogen in the rhizosphere and plant growth promotion bymicroorganisms. Plant Soil 321, 305–339.
Scheu, S., Ruess, L., Bonkowski, M., 2005. Interactions between microorganisms andsoil micro- and mesofauna. In: Buscot, F., Varma, A. (Eds.), Microorganisms inSoils: Roles in Genesis and Functions. Springer-Verlag, Berlin.
Spain, A.V., Tibbett, M., Hinz, D.A., Ludwig, J.A., Tongway, D.J., 2014. The mining-restoration system and ecosystem development following bauxite mining in abiodiverse environment of the seasonally dry tropics, Northern Territory,Australia. In: Tibbett, M. (Ed.), Mining in Ecologically Sensitive Landscapes.CSIRO Press (in press).
Suding, K.N., Harpole, W.S., Fukami, T., Kulmatiski, A., MacDougall, A.S., Stein, C., vander Putten, W.H., 2013. Consequences of plant–soil feedbacks in invasion. J.Ecol. 101, 298–308.
Treonis, A.M., Ostle, N.J., Stott, A.W., Primrose, R., Grayston, S.J., Ineson, P., 2004.Identification of groups of metabolically-active rhizosphere microorganisms bystable isotope probing of PLFAs. Soil Biol. Biochem. 36, 533–537.
Turner, J., Lambert, M.J., 2008. Nutrient cycling in age sequences of two Eucalyptusplantation species. For. Ecol. Manage. 255, 1701–1712.
Uren, N.C., 2001. Types, amounts, and possible functions of compounds releasedinto the rhizosphere by soil-grown plants. In: Pinton, R., Varanini, Z., Nannipieri,P. (Eds.), The Rhizosphere: Biochemistry and Organic Substances at the Soil–Plant Interface. Marcel Dekker Editorial, USA.
Uselman, S.M., Qualls, R.G., Lilienfein, J., 2012. Quality of soluble organic C, N, and Pproduced by different types and species of litter: root litter versus leaf litter. SoilBiol. Biochem. 54, 57–67.
van Grunsven, R.H.A., van der Putten, W.H., Bezemer, T.M., Tamis, W.L.M., Berendse,F., Veenendaal, E.M., 2007. Reduced plant–soil feedback of plant speciesexpanding their range as compared to natives. J. Ecol. 95, 1050–1057.
Weidenhamer, J.D., Callaway, R.M., 2010. Direct and indirect effects of invasiveplants on soil chemistry and ecosystem function. J. Chem. Ecol. 36, 59–69.
White, D.C., Davies, W.M., Nickels, J.S., Kings, J.D., Bobbie, R.J., 1979. Determinationof the sedimentary microbial biomass by extractible lipid phosphate. Oecologia40, 51–62.
Witt, C., Setälä, H., 2010. Do plant species of different resource qualities formdissimilar energy channels below-ground? Appl Soil Ecol 44, 270–278.
Yeates, G.W., Bongers, T., de Goede, R.G.M., Freckman, D.W., Georgieva, S.S., 1993.Feeding habits in soil nematode families and genera – an outline for soilecologists. J Nematol 25 (3), 315–331.
Yeates, G.W., 2003. Nematodes as soil indicators: functional and biodiversityaspects. Biol. Fertil. Soils 37, 199–210.
Zelles, L., Bai, Q.Y., 1993. Fractionation of fatty acids derived from soil lipids by soilphase extraction and their quantitative analysis by GC–MS. Soil Biol. Biochem.25, 495–507.
Zelles, L., 1999. Fatty acid patterns of phospholipids and lipopolysaccharides in thecharacterisation of microbial communities in soil: a review. Biol. Fertil. Soils 29,111–129.


















![[d], [o] If one doctor doctors another doctor does the doctor who doctors the doctor doctor the doctor the way the doctor he is doctoring doctors? Or](https://img.pdfslide.us/doc/110x75/56649e995503460f94b9c732/d-o-if-one-doctor-doctors-another-doctor-does-the-doctor-who-doctors-the.jpg)










