Embed Size (px)
Citation preview

In Vivo and In Vitro Inhibition of Monocyte Adhesion toEndothelial Cells and Endothelial Adhesion Molecules by
Eicosapentaenoic AcidHideto Yamada, Masayuki Yoshida, Yasutaka Nakano, Takayoshi Suganami, Noriko Satoh,
Tomoya Mita, Kosuke Azuma, Michiko Itoh, Yukio Yamamoto, Yasutomi Kamei, Minoru Horie,Hirotaka Watada, Yoshihiro Ogawa
Objective—A large-scale, prospective, randomized clinical trial has recently revealed that the addition of highly purifiedeicosapentaenoic acid (EPA) to low-dose statin therapy significantly reduces the incidence of major coronary events. Here weinvestigated in vivo and in vitro effect of EPA on monocyte adhesion to endothelial cells and adhesion molecules.
Methods and Results—A new en face immunohistochemistry of endothelial surface in combination with confocal microscopyrevealed marked reduction of lipopolysaccharide (LPS)-induced monocyte adhesion to the aortic endothelium in parallel withthe suppression of vascular cell adhesion molecule 1 (VCAM-1) and nuclear translocation of nuclear factor-�B p65 inEPA-treated mice relative to vehicle-treated groups. In an in vitro adhesion assay system under physiological flow conditions,EPA inhibited LPS-induced monocyte adhesion and endothelial adhesion molecules. We found significant decrease in plasmaconcentrations of soluble intercellular adhesion molecule 1 (sICAM-1) and sVCAM-1 in patients with the metabolicsyndrome after a 3-month administration of highly purified EPA (1.8 g daily). Multivariate regression analysis revealed thatEPA administration is the only independent determinant of sICAM-1 and sVCAM-1.
Conclusions—This study provides evidence that EPA inhibits monocyte adhesion to endothelial cells in parallel with thesuppression of endothelial adhesion molecules in vivo and in vitro. (Arterioscler Thromb Vasc Biol. 2008;28:2173-2179.)
Key Words: adhesion molecules � endothelial cells � EPA � monocytes � TLR4
Obesity may be viewed as a state of chronic low-gradeinflammation and confers a higher risk of atheroscle-
rotic diseases.1 There is considerable evidence that obeseadipose tissue is markedly infiltrated by macrophages,2,3
suggesting that they may participate in the inflammatorypathways that are activated in obese adipose tissue. Using anin vitro coculture system composed of adipocytes and mac-rophages, we have demonstrated that saturated fatty acidsreleased from adipocytes via the macrophage-induced lipol-ysis serve as a naturally occurring ligand for Toll-likereceptor 4 (TLR4) to induce the inflammatory changes inmacrophages through nuclear factor-�B (NF-�B) activa-tion.4,5 Notably, n-3 polyunsaturated fatty acids (n-3 PUFAs)such as eicosapentaenoic acid (EPA) and docosahexaenoicacid are unable to activate macrophages and can evenantagonize the proinflammatory effect of saturated fatty acidsor LPS,6 a well-defined exogenous ligand for TLR4,7 therebyhighlighting the antiinflammatory effect of n-3 PUFAs.
Atherosclerosis is a complex pathological process that isassociated with vascular wall dysfunction and inflammation.8
The monocyte-endothelial cell interaction may play a crucialrole in atherosclerotic plaque formation.9 Indeed, the adhe-sion of circulating monocytes to the intimal endothelial cellmonolayer is thought to be one of the earliest events, whichis mediated through complex interactions among multipleadhesion molecules and their counterreceptors expressed byboth endothelial cells and monocytes, such as selectins,intercellular adhesion molecule-1 (ICAM-1), vascular celladhesion molecule 1 (VCAM-1), and �1- and �2-integrins.10
Several previous reports indicated that the TLR4/NF-�Bsignaling pathway in both endothelial cells and monocytes/macrophages is involved in the development of vasculardysfunction and atherosclerosis11–13 and expression of adhe-sion molecules is regulated through NF-�B activation.14
Therefore, modulation of monocyte adhesion to the vascularendothelium may represent an attractive therapeutic target foratherosclerosis.
In epidemiological and clinical trials, fish oil rich in n-3PUFAs or n-3 PUFAs have reduced the incidence of coronary
Original received June 6, 2008; final version accepted September 24, 2008.From the Department of Molecular Medicine and Metabolism (H.Y., T.S., M.I., Y.K., Y.O.), Medical Top Track (MTT) Program, Medical Research
Institute (Y.Y.), Global Center of Excellence Program, International Research Center for Molecular Science in Tooth and Bone Diseases (Y.O.); andBioethics Research Center (M.Y.), Tokyo Medical and Dental University; Cardiovascular and Respiratory Medicine (H.Y., Y.N., M.H.), Shiga Universityof Medical Science, Otsu; Clinical Research Institute for Endocrine Metabolic Disease (N.S.), National Hospital Organization, Kyoto Medical Center,Kyoto; and the Department of Medicine, Metabolism, and Endocrinology (T.M., K.A., H.W.), Juntendo University School of Medicine, Tokyo, Japan.
Correspondence to Yoshihiro Ogawa, Department of Molecular Medicine and Metabolism, Medical Research Institute, Tokyo Medical and DentalUniversity, 1-5-45 Yushima, Bunkyo-ku, Tokyo 113-8510. E-mail [email protected]
© 2008 American Heart Association, Inc.
Arterioscler Thromb Vasc Biol is available at http://atvb.ahajournals.org DOI: 10.1161/ATVBAHA.108.171736
2173
by guest on June 2, 2018http://atvb.ahajournals.org/
Dow
nloaded from
by guest on June 2, 2018http://atvb.ahajournals.org/
Dow
nloaded from
by guest on June 2, 2018http://atvb.ahajournals.org/
Dow
nloaded from
by guest on June 2, 2018http://atvb.ahajournals.org/
Dow
nloaded from
by guest on June 2, 2018http://atvb.ahajournals.org/
Dow
nloaded from
by guest on June 2, 2018http://atvb.ahajournals.org/
Dow
nloaded from
by guest on June 2, 2018http://atvb.ahajournals.org/
Dow
nloaded from
by guest on June 2, 2018http://atvb.ahajournals.org/
Dow
nloaded from
by guest on June 2, 2018http://atvb.ahajournals.org/
Dow
nloaded from
by guest on June 2, 2018http://atvb.ahajournals.org/
Dow
nloaded from
by guest on June 2, 2018http://atvb.ahajournals.org/
Dow
nloaded from
by guest on June 2, 2018http://atvb.ahajournals.org/
Dow
nloaded from
by guest on June 2, 2018http://atvb.ahajournals.org/
Dow
nloaded from
by guest on June 2, 2018http://atvb.ahajournals.org/
Dow
nloaded from
by guest on June 2, 2018http://atvb.ahajournals.org/
Dow
nloaded from
by guest on June 2, 2018http://atvb.ahajournals.org/
Dow
nloaded from
by guest on June 2, 2018http://atvb.ahajournals.org/
Dow
nloaded from
by guest on June 2, 2018http://atvb.ahajournals.org/
Dow
nloaded from
by guest on June 2, 2018http://atvb.ahajournals.org/
Dow
nloaded from

heart disease.15 A large-scale, prospective, randomized clin-ical trial, or the Japan EPA Lipid Intervention Study (JELIS),has demonstrated that the addition of highly purified EPA, theonly class of n-3 PUFAs used clinically to treat hyperlipid-emia, to low-dose statin therapy significantly reduces theincidence of major coronary events,16 suggesting the pleio-tropic effect of EPA in addition to its well-known lipidlowering effect.17 It is, therefore, tempting to speculate thebeneficial effect of n-3 PUFAs on monocyte-endothelial cellinteractions during the course of atherosclerosis. Indeed, acouple of reports showed that n-3 PUFAs inhibit monocyte–endothelial cell interactions in vitro18; however, there is nodirect evidence for the inhibitory effect of EPA on monocyte-endothelial cell adhesion and endothelial adhesion moleculesin vivo. Moreover, there has been no report showing theinhibitory effect of EPA on endothelial adhesion moleculeexpression in patients with the metabolic syndrome, a majorrisk of atherosclerotic diseases.
Using a new en face immunohistochemistry of endothelialsurface in combination with confocal microscopy, we dem-onstrate for the first time that EPA markedly inhibits LPS-induced monocyte adhesion to the mouse aortic endotheliumin parallel with the suppression of endothelial adhesionmolecules and nuclear translocation of NF-�B p65 in vivo.The inhibitory effect is also confirmed under physiologicalflow conditions in vitro. We also show that highly purifiedEPA significantly reduces soluble ICAM-1 (sICAM-1) andsoluble VCAM-1 (sVCAM-1) in patients with the metabolicsyndrome. This study provides in vivo and in vitro evidence thatEPA inhibits monocyte adhesion to endothelial cells in parallelwith the suppression of endothelial adhesion molecules.
Materials and MethodsMaterialsPreparation and characterization of EPA used in vivo and in vitrowere reported elsewhere.6 All other reagents were purchased fromSigma and Nacalai Tesque unless otherwise noted.
AnimalsAll animal experiments were conducted in accordance to the guide-lines of Tokyo Medical and Dental University Committee on AnimalResearch (No. 0060026). Details are described in supplementalMethods (available online at http://atvb.ahajournals.org.).
New En Face Method for Optimal Observationof Endothelial SurfaceDetails are described in supplemental Methods.
En Face Analysis of Adhesion Molecule andNF-�B Activation on Aortic Endothelial SurfaceAfter fixation, the aorta was dissected carefully from the aortic archto lower thoracic region and was immersed in 10% buffered formalinfor 1 hour at 4°C. The thoracic aorta was removed of fat and cut openlongitudinally along the ventral side with scissors and rinsed 3 timeswith phosphate-buffered saline (PBS) containing 0.05% Tween 20(PBS-T). The samples were incubated overnight at 4°C with PBS-Tcontaining antimouse VCAM-1 and NF-�B p65 (p65) polyclonalantibodies (Santa Cruz Biotechnology Inc; 1:100) after 30 minutesblocking with PBS-T containing 10% donkey serum, and then rinsed3 times with PBS-T. It was followed by incubation with PBS-Tcontaining the secondary donkey antibody labeled with Alexa Fluor488 and 594 fluorophore molecules (Molecular Probes Inc; 1:600)for 1 hour at room temperature. The samples were rinsed 3 times
with PBS-T, placed on a slide glass with the intimal side up, andcovered with VECTORSHIELD Mounting Medium with DAPI(Vector Laboratories Inc). Each sample was viewed immediatelywith a FV1000 confocal system (Olympus). Three to 6 pictures ofeach field were captured at various focal lengths and counted aspositive cells per 1000 endothelial cells.
Details regarding cell culture, monocyte adhesion assay, fluores-cent immunobinding assay, western blot analysis, human study,statistical analysis are described in supplemental Methods.
ResultsEffect of EPA on LPS-Induced MonocyteAdhesion to Aortic Endothelium and EndothelialExpression of Adhesion Molecules In VivoMonocyte accumulation and atherosclerotic lesion formationare known to occur reproducibly at specific sites in thearterial tree, such as arterial branches.19 Using a newlydeveloped new enface method for optimal observation ofendothelial surface (NEMOes), we examined the effect ofEPA on LPS-induced monocyte adhesion to the endothelialsurface surrounding the orifice of intercostal arteries ofthoracic aorta in C57BL/6J mice. The number of monocytesadhering to the aortic endothelium was significantly in-creased in LPS-treated mice relative to vehicle-treated mice(P�0.01; Figure 1 and supplemental Figure I), which wassignificantly inhibited in EPA-treated group relative to con-trol group 6 hour after LPS treatment (P�0.01; Figure 1).The inhibitory effect tended to be observed up to 24 hoursafter LPS treatment (supplemental Figure I). In this study,serum EPA concentrations in EPA-treated and control groupswere consistent with previous reports (supplemental FigureIIA).20,21 Serum TG concentrations tended to be decreased byEPA treatment, although not statistically significant (supple-mental Figure IIB and IIC).
Confocal microscopic analysis of the aortic endotheliumrevealed that endothelial surface expression of VCAM-1 issignificantly increased in LPS-treated mice relative tovehicle-treated mice (P�0.01; Figure 2), which is signifi-cantly inhibited by EPA treatment (P�0.01; Figure 2).
Effect of EPA on LPS-Induced MonocyteAdhesion to Endothelial Cells and EndothelialExpression of Adhesion Molecules In VitroUsing an in vitro adhesion assay system under physiologicalflow conditions, we examined the effect of EPA onmonocyte-endothelial interaction; monocyte rolling and ad-hesion on the HUVEC monolayers (Figure 3A and supple-mental Figure III). The monocyte rolling and adhesion weresignificantly increased in LPS-treated HUVECs relative tovehicle-treated HUVECs (P�0.01; Figure 3A), which weresignificantly inhibited in HUVECs treated with EPA(P�0.01; Figure 3A). Palmitate, a major saturated fatty acidwhich can activate the NF-�B pathway via TLR4,4 was alsoused to induce monocyte-endothelial interaction. Interest-ingly, monocyte adhesion was significantly increased inpalmitate-treated HUVECs relative to vehicle-treatedHUVECs 4 hours after palmitate treatment (P�0.01; supple-mental Figure IV), which was significantly inhibited by EPAtreatment (P�0.01; supplemental Figure IV). The tendencywas observed up to 24 hours after palmitate treatment
2174 Arterioscler Thromb Vasc Biol December 2008
by guest on June 2, 2018http://atvb.ahajournals.org/
Dow
nloaded from

(supplemental Figure IV). We also found that the monocyterolling and adhesion are significantly increased in tumornecrosis factor (TNF)�-treated HUVECs relative to vehicle-treated HUVECs (P�0.01; supplemental Figure VA), whichis significantly inhibited in HUVECs treated with EPA(P�0.01; supplemental Figure VA).
Fluorescent immunobinding assay revealed that expressionof ICAM-1 and VCAM-1 is significantly increased in LPS-treated HUVECs relative to vehicle-treated HUVECs(P�0.01; Figure 3B), which is significantly inhibited inHUVECs treated with EPA (P�0.01; Figure 3B). In thisstudy, there was no appreciable difference in E-selectinexpression between LPS- and vehicle-treated groups 24 hoursafter LPS treatment. We also found that expression ofICAM-1 and VCAM-1 is significantly increased in TNF�-treated HUVECs relative to vehicle-treated HUVECs
(P�0.01; supplemental Figure VB), which is significantlyinhibited in HUVECs treated with EPA (P�0.01; supplemen-tal Figure VB).
Effect of EPA on LPS-Induced Intracellular SignalTransduction In Vitro and In VivoWestern blot analysis showed that the amount of Inhibitor of�B� (I�B-�) protein is reduced in total cell lysates in parallelwith the increase in phospho-p65 (p-p65) and p65 in nuclearextracts from LPS-treated HUVECs relative to vehicle-treated HUVECs (Figure 4A and 4B). The LPS-induceddecrease in I�B-� and increase in nuclear p-p65 and p65 wasreversed in HUVECs treated with EPA (Figure 4A and 4B).We also found that phospho-p38 mitogen-activated proteinkinase (p-p38) is increased in total cell lysates from LPS-treated HUVECs relative to vehicle-treated HUVECs, whichis inhibited by EPA treatment (Figure 4C).
We also examined whether EPA reduces LPS-inducedincrease in nuclear translocation of p65 in vivo. Confocalmicroscopic analysis revealed that nuclear translocation ofp65 in endothelial cells is significantly increased in parallelwith the increase in VCAM-1 expression in LPS-treatedgroup relative to vehicle-treated group (P�0.05; Figure 5Aand 5B). The LPS-induced increase in nuclear translocationof p65 was significantly inhibited in parallel with VCAM-1expression in EPA-treated group relative to vehicle-treatedgroup (P�0.01; Figure 5A and 5B).
LPS (-)EP
A (+
)EP
A (-
)LPS (+)A
LPSVehicle
30
10
20
40
0
(Cel
ls /
mm
2 )
**B
**EPA (-) EPA (+)
Figure 1. Effect of EPA on monocytes adhering to the aortic en-dothelium of mice. A, Representative en face views of immuno-histochemical staining with Mac-2 antibody for monocytesadhering to the aortic endothelium of mice 6 hours after i.p.injection of LPS with EPA treatment. Arrowheads denote Mac-2–positive cells. Scale bar, 100 �m. B, Quantification of Mac-2–positive cells. **P�0.01. n�5.
LPS
(+)
(VC
AM
-1-p
ositi
ve c
ells
/ 10
00 c
ells
)B
EPA (-) (-)
Vehicle
100
200
300
400
0
500
600 **
A(+) / (-) )+( / )+( )-( / )-(
LPS / EPA
**
Figure 2. Effect of EPA on VCAM-1 expression on the aorticendothelium of mice. A, Representative en face views of immu-nohistochemical staining with VCAM-1 antibody for the aorticendothelium of mice 24 hours after i.p. injection of LPS withEPA treatment. Endothelial cell nuclei and VCAM-1 are in blueand green, respectively. Scale bar, 10 �m. B, Quantification ofVCAM-1–positive cells. **P�0.01. n�10 to 15 visual fields.
Yamada et al Monocyte Adhesion and EPA 2175
by guest on June 2, 2018http://atvb.ahajournals.org/
Dow
nloaded from

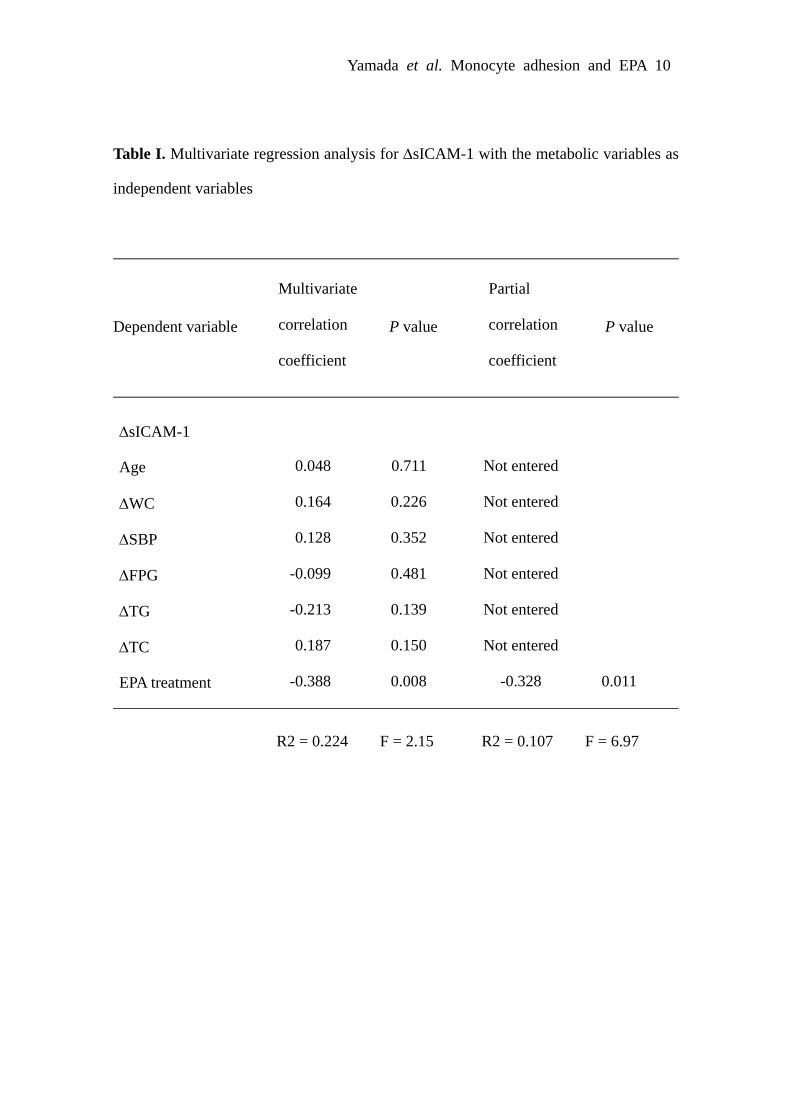
Effect of EPA on Soluble Adhesion Moleculesin HumansTo obtain insight into the effect of EPA on monocyteadhesion to endothelial cells in humans, we examinedwhether highly purified EPA decreases plasma concentra-tions of sICAM-1 and sVCAM-1 in patients with the meta-bolic syndrome. There was no significant difference in themeasured variables tested between EPA and control groupsbefore EPA administration (Table). After a 3-month EPAadministration, plasma EPA concentrations were significantlyincreased in EPA group relative to control group (P�0.01).In EPA group, plasma triglyceride (TG) concentrations weresignificantly decreased relative to control group (P�0.01),although there were no difference in body mass index (BMI),waist circumference (WC), systolic blood pressure (SBP),fasting plasma glucose (FPG), plasma concentrations of totalcholesterol (TC), high-density lipoprotein cholesterol (HDL-C), and low-density lipoprotein cholesterol (LDL-C). Bothplasma sICAM-1 and sVCAM-1 concentrations were signif-icantly reduced after EPA administration (P�0.01), butunchanged in control group (Table). To determine the factorsindependently influencing the changes in plasma sICAM-1and sVCAM-1 concentrations, multivariate regression analy-sis was performed (supplemental Tables I and II). PlasmasICAM-1 and sVCAM-1 concentrations were inversely cor-related only with EPA treatment.
DiscussionUsing NEMOes in combination with confocal microscopicanalysis of en face aortic endothelium, we demonstrated for thefirst time that EPA markedly inhibits LPS-induced monocyte
adhesion to the aortic endothelium, nuclear translocation ofNF-�B p65, and endothelial surface expression of VCAM-1 inmice. The new technique, originally described by Verna et al,22
will provide a unique opportunity to study monocyte adhesion toand intracellular signal transduction in endothelial cells in vivo.There is a report by Sethi et al that oxidized EPA is able toinhibit LPS-induced monocyte rolling and adhesion in themouse mesenteric venules in vivo.23 However, whether EPAinhibits monocyte adhesion to the aortic endothelium underhigher shear stress has never been addressed. To our knowledge,this study is the first in vivo observation that EPA inhibitsmonocyte adhesion to the aortic endothelial cells and reducesaortic endothelial expression of adhesion molecules. Theseobservations are consistent with a previous report by Matsumotoet al that a long-term EPA treatment reduces atheroscleroticplaque formation during the late phase of atherosclerosis inApoE-deficient mice.24 Furthermore, we found that LPS-induced monocyte adhesion tends to be reduced with coadmin-istration of EPA and statins relative to administration of statinsalone in vitro and in vivo (unpublished data Yamada H, YoshidaM, Watada H, Ogawa Y, 2008). These observations support theconcept that EPA is able to delay and prevent the progression ofatherosclerosis at both the early and late phases of atherosclero-sis beyond its lipid lowering effect. Using the in vitro adhesionassay system, we previously demonstrated that pharmacologicalinhibition of endothelial adhesion molecules results in thereduction of monocyte-endothelial cell adhesion.25,26 In thisstudy, we found that EPA results in marked inhibition ofLPS-induced increase in monocyte adhesion in parallel with the
IκκB-α
Actin
0 h 1 h 2 h
(+)EPA (-) (-) (+) (-) (+)(-) (-) (-)LPS LPS LPSVehicle
A
LPS LPSVehicle LPSVehicle
p65
EPA (-) (-) (+) (-) (+)(-) (-) (-) (+)
p-p65
B
Lamin
p-p38
p38
0 h 1 h 2 h
EPA (-) (-) (+) (-) (+)(-) (-) (-)
LPS
(+)
LPS LPSVehicle
C
0 h 2 h
Vehicle
Vehicle
Vehicle
Vehicle
Vehicle1 h
Figure 4. Effect of EPA on LPS-induced intracellular signalingpathways in HUVECs. Representative Western blot analysisusing total cell lysates (A and C) and nuclear extracts (B) fromHUVECs before and 1 hour and 2 hours after LPS treatmentwith EPA treatment. A, I�B-� expression. B, p-p65 and p65expression. C, p-p38 and p38 expression. Blots are representa-tive of 4 to 5 separate experiments.
B
(Rel
ativ
e flu
ores
cent
uni
ts)
0
20
40
60
80
100
Vehicle LPS Vehicle LPS
ICAM-1 VCAM-1 E-selectin
******
**
Vehicle LPS
2
4
0
** **
AR
ollin
gEPA (-) EPA (+)
EPA (-) EPA (+)
Vehicle LPS
6
2
4
8
0
(HL6
0 ce
lls /
HPF
) ****
Adh
esio
n
Vehicle LPS0
10
20
30(R
elat
ive
fluor
esce
nt u
nits
)
0
10
20
30
40
(Rel
ativ
e flu
ores
cent
uni
ts)
(HL6
0 ce
lls /
HPF
)
Figure 3. Effect of EPA on LPS-induced monocyte adhesionand expression of endothelial adhesion molecules in HUVECs.A, The number of rolling and adhesive monocytes on HUVECs24 hours after LPS treatment with EPA treatment under physio-logical flow conditions. **P�0.01. n�10 high power fields. Dataare representative of 3 separate experiments. B, Endothelial sur-face expression of ICAM-1, VCAM-1, and E-selectin in HUVECs24 hours after LPS treatment with EPA treatment. **P�0.01.n�3. Data are representative of 3 separate experiments.
2176 Arterioscler Thromb Vasc Biol December 2008
by guest on June 2, 2018http://atvb.ahajournals.org/
Dow
nloaded from

suppression of endothelial adhesion molecules and NF-�Bactivation in HUVECs. The LPS-induced increase in nucleartranslocation of NF-�B p65 is also inhibited in mice treated withEPA in vivo. Although EPA may also act on circulatingmonocytes and thus modulate their adhesion to endothelial cellsin vivo,27 it is likely that EPA inhibition of monocyte adhesionto endothelial cells is mediated at least in part through thesuppression of endothelial adhesion molecules.
The molecular mechanism by which EPA reduces expres-sion of endothelial adhesion molecules is unclear at present.In this study, we demonstrated that EPA inhibits LPS-inducednuclear translocation of p65 in endothelial cells both in vivoand in vitro, which is consistent with our previous observa-tion that EPA suppresses NF-�B activity in macrophagestreated with LPS or saturated fatty acids.6 We also found thatEPA inhibits LPS-induced increase in p-p38, which is knownto mediate NF-�B activation.14 It is, therefore, conceivablethat the upstream signaling of I�B-� and p38 is important forthe antiinflammatory effect of EPA. There is a report sug-gesting that peroxisome proliferator-activated receptor(PPAR) � interacts with p65 and interferes with NF-�Bactivation.28 Indeed, Sethi et al reported that an i.p. injectionof oxidized EPA suppresses monocyte rolling and adhesion tothe venous endothelium via a PPAR�-dependent pathway.23
In this regard, PPAR� has been shown to export NF-�B p65from the nucleus via direct protein–protein interaction, thusattenuating NF-�B activity.29 Moreover, a PPAR� antagonistGW9662 has reversed EPA-induced inhibition of ICAM-1mRNA expression in endothelial cells in vitro.21 However,we found that GW9662 fails to reverse the EPA-inducedinhibition of monocyte adhesion in the in vitro adhesion assaysystem (unpublished data Yamada H, Yoshida M, Ogawa Y,2008). On the other hand, Arita et al demonstrated thatresolvinE1, an endogenous lipid mediator derived from EPA,exerts an antiinflammatory effect through interaction withleukotriene B4 receptor BLT1 and chemerin receptorChemR23.30 Further studies are required to elucidate howEPA inhibits the monocyte-endothelial interaction.
Evidence has accumulated indicating that elevated levelsof circulating free fatty acids, especially saturated free fattyacids derived from dietary animal fat or via the macrophage-induced adipocyte lipolysis in obese adipose tissue, maypotentially contribute to the development of the inflammatorychanges and dysfunction in the vascular wall.31–33 For in-stance, Kim et al recently reported that a high-fat diet inducesvascular insulin resistance and inflammatory responses bysignaling through the TLR4/NF-�B pathway and suggestedthat such effect is mediated at least in part by saturated freefatty acids.34 In this study, we also found that palmitateinduces monocyte adhesion to endothelial cells in the in vitroadhesion assay system, which is inhibited by EPA treatment.These observations, taken together, suggest that the TLR4/NF-�B pathway plays a role in the deleterious effect ofsaturated free fatty acids on monocyte-endothelial cell inter-action in the vasculature. The above discussion supports theconcept that antagonism of the TLR4/NF-�B pathway invascular wall offers a novel therapeutic strategy to prevent ortreat obesity-related atherosclerotic diseases. Using the invitro coculture of adipocytes and macrophages, we havedemonstrated that EPA reverses the coculture-induced reduc-
Table. Clinical Characteristics and Metabolic ParametersBefore and After the Follow-Up Period
Control Group EPA Group
Before After Before After
Sex, male/female 11/19 12/18
Age 52.9�2.75 50.3�2.48
BMI, kg/m2 29.7�0.79 29.7�0.79 30.0�0.84 29.8�0.86
WC, cm 94.7�2.02 95.2�2.04 95.0�2.03 94.9�2.05
SBP, mm Hg 136�3.42 133�3.26 135�3.15 140�3.42
FPG, mg/dl 115�4.94 120�7.43 111�9.02 108�9.30
TC, mg/dl 203�5.52 206�5.66 205�6.85 199�5.39
TG, mg/dl 164�13.1 159�12.4 178�16.7 131�10.5**
HDL-C, mg/dl 57.2�2.57 54.2�2.97 53.3�2.74 52.5�2.40
LDL-C, mg/dl 125�5.31 128�5.31 130�4.54 123�4.59
EPA, �g/ml 73.0�10.1 69.3�7.76 76.2�9.06 143�14.6**‡
sICAM-1, ng/ml 210�9.66 210�10.2 224�13.9 204�13.0**
sVCAM-1, ng/ml 622�38.3 624�38.8 676�55.4 569�36.6**
Data are expressed as the mean�SEM. **P�0.01 vs before determined by2-tailed, paired t test. ‡P�0.01 vs control determined by Student t test.
(+) / (-)
LPS
(+)
B
(-) / (-)
EPA (-) (-)
Vehicle
(+) / (+)
**
A
100
200
300
400
0
(p65
-pos
itive
cel
ls /
1000
cel
ls)
LPS / EPA
*
Figure 5. Effect of EPA on nuclear translocation of p65 in aortic en-dothelial cells of mice. A, Representative images of nuclear transloca-tion of p65 in aortic endothelial cells of mice 24 hours after i.p. injec-tion of LPS with EPA treatment. Endothelial Cell nuclei, VCAM-1, andp65 are in blue, green, and red, respectively. The upper row showsdouble staining, and the lower, triple staining. Scale bar, 10 �m. B,Quantification of endothelial cells positive for nuclear translocation ofp65. *P�0.05. **P�0.01. n�10 to 15 visual fields.
Yamada et al Monocyte Adhesion and EPA 2177
by guest on June 2, 2018http://atvb.ahajournals.org/
Dow
nloaded from

tion of adiponectin secretion at least in part by suppressingthe saturated fatty acids/TLR4/NF-�B pathway in macro-phages and suggested that the beneficial effect of EPA isattributable at least partly to the improvement of obesity-induced adipose tissue inflammation.6 It is, therefore, con-ceivable that EPA reduces the incidence of obesity-inducedatherosclerotic diseases through the suppression of adipocyte-macrophage interaction in the adipose tissue and monocyte-endothelial interaction in the vascular tissue.
Soluble forms of endothelial adhesion molecules have beenclinically used as surrogate markers to reflect endothelialsurface expression of adhesion molecules. There are previousreports showing that treatment with n-3 PUFAs reducessignificantly sICAM-1 in patients with dyslipidemia.35,36
However, whether n-3 PUFAs have an inhibitory effect onendothelial expression of adhesion molecules in the metabol-ic syndrome has not been addressed so far. In this study, wedemonstrated for the first time that highly purified EPAdecreases both plasma sICAM-1 and sVCAM-1 concentra-tions in patients who met the criteria of the metabolicsyndrome or a higher risk group of atherosclerosis. Indeed,we also confirmed that cardio-ankle vascular index, a newindex of arterial stiffness independent of blood pressure,37
and other biomarkers such as adiponectin and high-sensitivityC-reactive protein are significantly improved relative tocontrol group as short as 3-month EPA administration (un-published data Satoh N, Ogawa Y, 2008). These observa-tions, taken together, suggest that the inhibitory effect of EPAon monocyte-endothelial interaction may also contribute tothe marked antiatherogenic effect of EPA in patients at higherrisk of atherosclerosis. In this study, multivariate regressionanalysis revealed that EPA reduces both sICAM-1 andsVCAM-1 independently of other metabolic parameters.These observations support the concept that EPA acts directlyon endothelial cells to reduce levels of endothelial adhesionmolecules and prevents the progression of atherosclerosis andmajor coronary events. However, this study does not excludethe possibility that the EPA-induced reduction in TG influ-ences the reduction in soluble adhesion molecules.
In conclusion, this study provides in vivo and in vitroevidence that EPA inhibits monocyte adhesion to endothelialcells in parallel with the suppression of endothelial adhesionmolecules, thereby highlighting the pleiotropic effect of EPA.Given the potentially important role of monocyte adhesion inthe pathogenesis of atherosclerosis, the data of this studycould be relevant to the protective effects of EPA in patientsat risk for cardiovascular disease.
AcknowledgmentsThe authors thank Hajime Yamakage for technical assistance.
Sources of FundingThis work was supported in part by a Grant-in-Aid for ScientificResearch from the Ministry of Education, Culture, Sports, Science,and Technology of Japan, and Ministry of Health, Labor, andWelfare of Japan, and research grants from Daiwa Securities HealthFoundation, Mitsubishi Pharma Research Foundation, Takeda Sci-ence Foundation, and Japan Research Foundation for Clinical Phar-macology. Michiko Itoh is Research Fellow of the Japan Society forthe Promotion of Science.
DisclosuresNone.
References1. Berg AH, Scherer PE. Adipose tissue, inflammation, and cardiovascular
disease. Circ Res. 2005;96:939–949.2. Weisberg SP, McCann D, Desai M, Rosenbaum M, Leibel RL, Ferrante
AW Jr. Obesity is associated with macrophage accumulation in adiposetissue. J Clin Invest. 2003;112:1796–1808.
3. Xu H, Barnes GT, Yang Q, Tan G, Yang D, Chou CJ, Sole J, Nichols A,Ross JS, Tartaglia LA, Chen H. Chronic inflammation in fat plays acrucial role in the development of obesity-related insulin resistance.J Clin Invest. 2003;112:1821–1830.
4. Suganami T, Tanimoto-Koyama K, Nishida J, Itoh M, Yuan X, MizuaraiS, Kotani H, Yamaoka S, Miyake K, Aoe S, Kamei Y, Ogawa Y. Role ofthe Toll-like receptor 4/NF-kappaB pathway in saturated fatty acid-induced inflammatory changes in the interaction between adipocytes andmacrophages. Arterioscler Thromb Vasc Biol. 2007;27:84–91.
5. Suganami T, Nishida J, Ogawa Y. A paracrine loop between adipocytesand macrophages aggravates inflammatory changes: role of free fattyacids and tumor necrosis factor alpha. Arterioscler Thromb Vasc Biol.2005;25:2062–2068.
6. Itoh M, Suganami T, Satoh N, Tanimoto-Koyama K, Yuan X, Tanaka M,Kawano H, Yano T, Aoe S, Takeya M, Shimatsu A, Kuzuya H, Kamei Y,Ogawa Y. Increased adiponectin secretion by highly purified eicosapen-taenoic acid in rodent models of obesity and human obese subjects.Arterioscler Thromb Vasc Biol. 2007;27:1918–1925.
7. Akira S, Takeda K, Kaisho T. Toll-like receptors: critical proteins linkinginnate and acquired immunity. Nat Immunol. 2001;2:675–680.
8. Ross R. Atherosclerosis–an inflammatory disease. N Engl J Med. 1999;340:115–126.
9. Cybulsky MI, Iiyama K, Li H, Zhu S, Chen M, Iiyama M, Davis V,Gutierrez-Ramos JC, Connelly PW, Milstone DS. A major role for VCAM-1,but not ICAM-1, in early atherosclerosis. J Clin Invest. 2001;107:1255–1262.
10. Luscinskas FW, Kansas GS, Ding H, Pizcueta P, Schleiffenbaum BE,Tedder TF, Gimbrone MA Jr. Monocyte rolling, arrest and spreading onIL-4-activated vascular endothelium under flow is mediated viasequential action of L-selectin, beta 1-integrins, and beta 2-integrins.J Cell Biol. 1994;125:1417–1427.
11. Michelsen KS, Wong MH, Shah PK, Zhang W, Yano J, Doherty TM,Akira S, Rajavashisth TB, Arditi M. Lack of Toll-like receptor 4 ormyeloid differentiation factor 88 reduces atherosclerosis and alters plaquephenotype in mice deficient in apolipoprotein E. Proc Natl Acad SciU S A. 2004;101:10679–10684.
12. Kim F, Tysseling KA, Rice J, Pham M, Haji L, Gallis BM, Baas AS,Paramsothy P, Giachelli CM, Corson MA, Raines EW. Free fatty acidimpairment of nitric oxide production in endothelial cells is mediated byIKKbeta. Arterioscler Thromb Vasc Biol. 2005;25:989–994.
13. Xu XH, Shah PK, Faure E, Equils O, Thomas L, Fishbein MC, LuthringerD, Xu XP, Rajavashisth TB, Yano J, Kaul S, Arditi M. Toll-likereceptor-4 is expressed by macrophages in murine and human lipid-richatherosclerotic plaques and upregulated by oxidized LDL. Circulation.2001;104:3103–3108.
14. Liu SF, Malik AB. NF-kappa B activation as a pathological mechanismof septic shock and inflammation. Am J Physiol Lung Cell Mol Physiol.2006;290:L622–L645.
15. Din JN, Newby DE, Flapan AD. Omega 3 fatty acids and cardiovasculardisease–fishing for a natural treatment. BMJ. 2004;328:30–35.
16. Yokoyama M, Origasa H, Matsuzaki M, Matsuzawa Y, Saito Y, IshikawaY, Oikawa S, Sasaki J, Hishida H, Itakura H, Kita T, Kitabatake A,Nakaya N, Sakata T, Shimada K, Shirato K. Effects of eicosapentaenoicacid on major coronary events in hypercholesterolaemic patients (JELIS):a randomised open-label, blinded endpoint analysis. Lancet. 2007;369:1090–1098.
17. Mizuguchi K, Yano T, Tanaka Y, Ishibashi M, Masada A, Mizota M, FukutakeK, Saito Y. Mechanism of the lipid-lowering effect of ethyl all-cis-5,8,11,14,17-icosapentaenoate. Eur J Pharmacol. 1993;231:121–127.
18. Chen H, Li D, Chen J, Roberts GJ, Saldeen T, Mehta JL. EPA and DHAattenuate ox-LDL-induced expression of adhesion molecules in humancoronary artery endothelial cells via protein kinase B pathway. J Mol CellCardiol. 2003;35:769–775.
19. Iiyama K, Hajra L, Iiyama M, Li H, DiChiara M, Medoff BD, Cybulsky MI.Patterns of vascular cell adhesion molecule-1 and intercellular adhesion
2178 Arterioscler Thromb Vasc Biol December 2008
by guest on June 2, 2018http://atvb.ahajournals.org/
Dow
nloaded from

molecule-1 expression in rabbit and mouse atherosclerotic lesions and at sitespredisposed to lesion formation. Circ Res. 1999;85:199–207.
20. Hagiwara S, Makita Y, Gu L, Tanimoto M, Zhang M, Nakamura S,Kaneko S, Itoh T, Gohda T, Horikoshi S, Tomino Y. Eicosapentaenoicacid ameliorates diabetic nephropathy of type 2 diabetic KKAy/Ta mice:involvement of MCP-1 suppression and decreased ERK1/2 and p38phosphorylation. Nephrol Dial Transplant. 2006;21:605–615.
21. Koto T, Nagai N, Mochimaru H, Kurihara T, Izumi-Nagai K, Satofuka S,Shinoda H, Noda K, Ozawa Y, Inoue M, Tsubota K, Oike Y, Ishida S.Eicosapentaenoic acid is anti-inflammatory in preventing choroidal neo-vascularization in mice. Invest Ophthalmol Vis Sci. 2007;48:4328–4334.
22. Verna L, Ganda C, Stemerman MB. In vivo low-density lipoproteinexposure induces intercellular adhesion molecule-1 and vascular celladhesion molecule-1 correlated with activator protein-1 expression. Arte-rioscler Thromb Vasc Biol. 2006;26:1344–1349.
23. Sethi S, Ziouzenkova O, Ni H, Wagner DD, Plutzky J, Mayadas TN. Oxidizedomega-3 fatty acids in fish oil inhibit leukocyte-endothelial interactions throughactivation of PPAR alpha. Blood. 2002;100:1340–1346.
24. Matsumoto M, Sata M, Fukuda D, Tanaka K, Soma M, Hirata Y, Nagai R.Orally administered eicosapentaenoic acid reduces and stabilizes atheroscleroticlesions in ApoE-deficient mice. Atherosclerosis. 2008;197:524–533.
25. Yoshida M, Takano Y, Sasaoka T, Izumi T, Kimura A. E-selectin poly-morphism associated with myocardial infarction causes enhancedleukocyte-endothelial interactions under flow conditions. ArteriosclerThromb Vasc Biol. 2003;23:783–788.
26. Nakamura N, Yoshida M, Umeda M, Huang Y, Kitajima S, Inoue Y,Ishikawa I, Iwai T. Extended exposure of lipopolysaccharide fractionfrom Porphyromonas gingivalis facilitates mononuclear cell adhesion tovascular endothelium via Toll-like receptor-2 dependent mechanism.Atherosclerosis. 2008;196:59–67.
27. Swirski FK, Libby P, Aikawa E, Alcaide P, Luscinskas FW, WeisslederR, Pittet MJ. Ly-6Chi monocytes dominate hypercholesterolemia-associated monocytosis and give rise to macrophages in atheromata.J Clin Invest. 2007;117:195–205.
28. Delerive P, De Bosscher K, Besnard S, Vanden Berghe W, Peters JM,Gonzalez FJ, Fruchart JC, Tedgui A, Haegeman G, Staels B. Peroxisome
proliferator-activated receptor alpha negatively regulates the vascularinflammatory gene response by negative cross-talk with transcriptionfactors NF-kappaB and AP-1. J Biol Chem. 1999;274:32048–32054.
29. Kelly D, Campbell JI, King TP, Grant G, Jansson EA, Coutts AG,Pettersson S, Conway S. Commensal anaerobic gut bacteria attenuateinflammation by regulating nuclear-cytoplasmic shuttling ofPPAR-gamma and RelA. Nat Immunol. 2004;5:104–112.
30. Arita M, Ohira T, Sun YP, Elangovan S, Chiang N, Serhan CN. ResolvinE1 selectively interacts with leukotriene B4 receptor BLT1 and ChemR23to regulate inflammation. J Immunol. 2007;178:3912–3917.
31. Staiger H, Staiger K, Stefan N, Wahl HG, Machicao F, Kellerer M,Haring HU. Palmitate-induced interleukin-6 expression in humancoronary artery endothelial cells. Diabetes. 2004;53:3209–3216.
32. Steinberg HO, Paradisi G, Hook G, Crowder K, Cronin J, Baron AD. Freefatty acid elevation impairs insulin-mediated vasodilation and nitric oxideproduction. Diabetes. 2000;49:1231–1238.
33. Watanabe S, Tagawa T, Yamakawa K, Shimabukuro M, Ueda S. Inhi-bition of the renin-angiotensin system prevents free fatty acid-inducedacute endothelial dysfunction in humans. Arterioscler Thromb Vasc Biol.2005;25:2376–2380.
34. Kim F, Pham M, Luttrell I, Bannerman DD, Tupper J, Thaler J, Hawn TR,Raines EW, Schwartz MW. Toll-like receptor-4 mediates vascularinflammation and insulin resistance in diet-induced obesity. Circ Res.2007;100:1589–1596.
35. Abe Y, El-Masri B, Kimball KT, Pownall H, Reilly CF, Osmundsen K,Smith CW, Ballantyne CM. Soluble cell adhesion molecules in hypertri-glyceridemia and potential significance on monocyte adhesion. Arte-rioscler Thromb Vasc Biol. 1998;18:723–731.
36. Hjerkinn EM, Seljeflot I, Ellingsen I, Berstad P, Hjermann I, Sandvik L,Arnesen H. Influence of long-term intervention with dietary counseling,long-chain n-3 fatty acid supplements, or both on circulating markers ofendothelial activation in men with long-standing hyperlipidemia. Am JClin Nutr. 2005;81:583–589.
37. Satoh N, Shimatsu A, Kato Y, Araki R, Koyama K, Okajima T, TanabeM, Ooishi M, Kotani K, Ogawa Y. Evaluation of the cardio-anklevascular index, a new indicator of arterial stiffness independent of bloodpressure, in obesity and metabolic syndrome. Hypertens Res. In press.
Yamada et al Monocyte Adhesion and EPA 2179
by guest on June 2, 2018http://atvb.ahajournals.org/
Dow
nloaded from

Horie, Hirotaka Watada and Yoshihiro OgawaTomoya Mita, Kosuke Azuma, Michiko Itoh, Yukio Yamamoto, Yasutomi Kamei, Minoru
Hideto Yamada, Masayuki Yoshida, Yasutaka Nakano, Takayoshi Suganami, Noriko Satoh,Endothelial Adhesion Molecules by Eicosapentaenoic Acid
In Vivo and In Vitro Inhibition of Monocyte Adhesion to Endothelial Cells and
Print ISSN: 1079-5642. Online ISSN: 1524-4636 Copyright © 2008 American Heart Association, Inc. All rights reserved.
Greenville Avenue, Dallas, TX 75231is published by the American Heart Association, 7272Arteriosclerosis, Thrombosis, and Vascular Biology
doi: 10.1161/ATVBAHA.108.1717362008;
2008;28:2173-2179; originally published online October 23,Arterioscler Thromb Vasc Biol.
http://atvb.ahajournals.org/content/28/12/2173World Wide Web at:
The online version of this article, along with updated information and services, is located on the
http://atvb.ahajournals.org/content/suppl/2008/10/24/ATVBAHA.108.171736.DC1Data Supplement (unedited) at:
http://atvb.ahajournals.org//subscriptions/
at: is onlineArteriosclerosis, Thrombosis, and Vascular Biology Information about subscribing to Subscriptions:
http://www.lww.com/reprints
Information about reprints can be found online at: Reprints:
document. Question and AnswerPermissions and Rightspage under Services. Further information about this process is available in the
which permission is being requested is located, click Request Permissions in the middle column of the WebCopyright Clearance Center, not the Editorial Office. Once the online version of the published article for
can be obtained via RightsLink, a service of theArteriosclerosis, Thrombosis, and Vascular Biologyin Requests for permissions to reproduce figures, tables, or portions of articles originally publishedPermissions:
by guest on June 2, 2018http://atvb.ahajournals.org/
Dow
nloaded from

LPS (-)EP
A (+
)EP
A (-
)LPS (+)(A)
Figure I
LPS
30
Vehicle
(Cel
ls /
mm
2 )
10
20
40
0
EPA (-) EPA (+)(B)**

Figure II
Vehicle LPS
(μg/
ml) 200
100
300
0
**
**
EPA(A)
(mg/
dl)
Vehicle LPS
(B) (C)TG
120
(mg/
dl)
0
40
80
TG
Vehicle LPS
*120
0
40
80
EPA (-) EPA (+)
EPA (-) EPA (+)

Figure III
Flow assay (Movie)
LPS (-)
EPA
(+)
EPA
(-)
LPS (+)Movie I
Movie II
Movie III
Movie IV

4
0
2
4
0
2
PalmitateVehicleEPA (-) (-) (+)
PalmitateVehicleEPA (-) (-) (+)
Figure IV
(HL6
0 ce
lls /
HPF
)
(HL6
0 ce
lls /
HPF
)
24 h****
4 h
Adh
esio
n
Adh
esio
n

(A) EPA (-) EPA (+)
****
Rol
ling
(HL6
0 ce
lls /
HPF
)A
dhes
ion
(HL6
0 ce
lls /
HPF
)
2
4
0
6
24
8
0
10****
Vehicle TNFα Vehicle TNFα
(Rel
ativ
e flu
ores
cent
uni
ts)
0
20
40
60
80
100
0
10
20
30
40
(Rel
ativ
e flu
ores
cent
uni
ts)
50
0
10
20
(Rel
ativ
e flu
ores
cent
uni
ts)
(B)
ICAM-1 VCAM-1 E-selectin
EPA (-) EPA (+)
Vehicle TNFα Vehicle TNFα Vehicle TNFα
**** ** **
Figure V

Yamada et al. Monocyte adhesion and EPA 1
Online Supplement
Methods
Animals
Male C57BL/6J mice were purchased from CLEA (Tokyo, Japan). Seven to 9-week-old
mice were housed in individual cages in a temperature-, humidity-, and light-controlled
room (12-h light and 12-h dark cycle) and allowed free access to water and a PUFA-free
diet (fish meal-free F1; 362 kcal/100 g, 4.4% energy as fat; Funabashi Farm, Chiba,
Japan) supplemented with or without 5% EPA (wt/wt) for 1 week (EPA-treated and
control groups, respectively).1 All diets were changed every day to avoid oxidation. At
the end of the experiment, mice were injected LPS (1 mg/kg) or saline (vehicle) after
overnight fast and thereafter sacrificed under anesthesia by intraperitoneal (i.p.)
injection of sodium pentobarbital (1 mg/kg; Nembutal, Abbott Laboratories, Abbott
Park, IL).
New En face Method for Optimal Observation of Endothelial Surface (NEMOes)
Fixation and preparation of the mouse aorta were performed as described.2, 3 Briefly, the
samples were incubated with anti mouse Mac-2 monoclonal antibody (Dako
Corporation, Carpinteria, CA) followed by incubation with biotinylated goat anti-mouse
IgG and reaction with horseradish peroxidase-conjugated streptavidin (Dako). Staining
was completed after incubation with a substrate-chromogen solution and
counterstaining with hematoxylin. Six to 10 pictures of each field were captured at
various focal lengths with an automatically regulated Z-stepper using an Image-Pro4.5J
(Planetron Co., Tokyo, Japan) to select the clearest images automatically and produce a
composite image of the entire thoracic aorta.
Measurement of Serum EPA and Triglyceride Concentrations

Yamada et al. Monocyte adhesion and EPA 2
Serum Triglyceride (TG) concentrations were measured as described.1 Serum EPA
concentrations were determined by gas chromatography.4
Cell Culture
Human umbilical vein endothelial cells (HUVECs) and HL60, a promyelocytic human
leukocyte cell line were obtained from Sanko Junyaku (Tokyo, Japan) and American
Type Culture Collection (Rockville, MD), respectively, and cultured as described.5
Monocyte Adhesion Assay
The protocols of monocyte adhesion assay under physiological flow conditions were
described.6 In brief, HUVEC monolayers on coverslips were treated with 1%
FBS-containing RPMI-1640 medium supplemented with or without EPA (50 μmol/L)
for 1 h followed by stimulation with LPS (1 μg/ml), palmitate (200 μmol/L), or tumor
necrosis factor-α (TNFα) (Research and Diagnostic Systems, Minneapolis, MN) (5
ng/ml) for the indicated period. The coverslips were positioned in a parallel-plate flow
chamber and mounted on an IX70 inverted microscope (Olympus, Tokyo, Japan). The
flow chamber was perfused for 2 to 3 min with perfusion medium (PBS containing
0.2% human serum albumin), over which HL60 cells were suspended to achieve a final
concentration of 1 × 105 cells/ml and drawn through the chamber. The flow rate was
controlled to generate a calculated shear stress of 1.0 dyne/cm2 using a PHD2000
syringe pump (Harvard Apparatus Inc., Holliston, MA). The entire period of perfusion
was recorded on videotape and captured images were analyzed to determine the number
of rolling and adherent HL60 cells on HUVEC monolayers in 10 randomly selected 20
× microscopic fields. Cells were considered to be “adherent” after about 10 sec of stable
contact with the monolayer.
Fluorescent Immunobinding Assay

Yamada et al. Monocyte adhesion and EPA 3
Fluorescent immunobinding assay was carried out as described.5, 7 In brief, HUVECs
were incubated with the following primary antibodies; mouse anti-human E-selectin
monoclonal antibody (mAb) (7A9), anti-human ICAM-1 mAb (Hu5/3), and anti-human
VCAM-1 mAb (E1/6), at a concentration of 10 μg/ml for 1 h on ice, followed by a
fluorescein isothiocyanate-conjugated goat anti-mouse IgG for 1 h on ice. Cell
surface-associated fluorescence was detected using a Cytofluor II fluorescent plate
reader (Applied Biosystems Inc., Foster City, CA).
Western Blot Analysis
Total cell lysates and nuclear extracts were prepared as described previously.7 In brief,
10 μg of protein obtained from 1× 107 HUVECs plated in a 10 cm dish was placed in
each lane in 7.5~12.5% sodium dodecyl sulfate-polyacrylamide gel electrophoresis
followed by Western blot analysis using antibodies against NF-κB p65 (p65) (Santa
Cruz Biotechnology), Inhibitor of κBα (IκB-α), phospho-p65 (Ser536) (93H1) (p-p65),
p38 mitogen-activated protein kinase (MAPK) (p38), phospho-p38 MAPK
(Thr180/Tyr182) (p-p38) (Cell Signaling Technology Inc., Danvers, MA), Lamin A/C
(Millipore, Billerica, MA), and actin. Immunoblots were developed with horseradish
peroxidase-conjugated secondary antibodies and detected using an ECL Advance
Western Blotting Detection Kit (GE Healthcare Bio-Sciences Corp., NJ) and observed
with a LAS-3000 mini system (Fuji Photo Film, Tokyo, Japan).
Human Study
A total of 60 Japanese obese patients with dyslipidemia (23 men and 37 women, mean
age 51.6 ± 1.8 years, mean BMI 29.8 ± 0.6 kg/m2, mean hemoglobin A1c (HbA1c) 6.0
± 0.2 %) were recruited in our clinics. They all met the metabolic syndrome (MS)
criteria by the modified National Cholesterol Education Program-Adult treatment Panel
III (NCEP-ATP III) definition.8 The study protocol was approved by the ethical

Yamada et al. Monocyte adhesion and EPA 4
committee on human research of Kyoto Medical Center and Medical Research Institute,
Tokyo Medical and Dental University, and all participants gave written informed
consent.
Patients were assigned to one of the following treatment groups (a single-blind
and run-in period randomization, which patients received); those treated for 3 months
with either diet alone (control group) or diet plus EPA (1.8 g daily) (EPA group). At the
beginning and the end of the study, we measured body mass index (BMI), waist
circumference (WC), systolic blood pressure (SBP), fasting plasma glucose (FPG),
plasma concentrations of total cholesterol (TC), triglyceride (TG), high density
lipoprotein cholesterol (HDL-C), low density lipoprotein cholesterol (LDL-C), and EPA
according to the standard procedures. Plasma concentrations of sICAM-1 and
sVCAM-1 were measured using commercially available immunoassays (Research and
Diagnostic Systems, Minneapolis, MN).9 Lipid lowering medications such as statins and
fibrates were excluded. Several patients in both groups had medication with oral
antidiabetic agents and antihypertensive agents and there were no changes in medication
during the study.
Statistical Analysis
Data are presented as mean ± SEM, and P < 0.05 was considered statistically significant.
In cell culture and animal studies, Bartlett test was used to verify the homogeneity of
variances followed by one-way ANOVA to determine the overall difference between
groups. When the variance was not homogenous, the data were analyzed with
Kruskal-Wallis test. If one-way ANOVA or Kruskal-Wallis test was significant,
differences between individual groups were estimated using Scheffe test.

Yamada et al. Monocyte adhesion and EPA 5
In human study, repeated measures ANOVA (control and EPA groups × before and
after the treatment) was used to access the comparative effect of EPA treatment on the
measured variables. A two-tailed, paired t-test was applied for the evaluation of changes
from baseline conditions to those at 3 months in the same group. Comparisons of the
means between the two groups at baseline or post-treatment (abbreviated as Δ) were
performed by Student’s t-test (Table 1). Changes from baseline conditions to those at 3
months were abbreviated as Δ. Multivariate regression analysis was performed to
elucidate factors related to ΔsICAM-1 and ΔsVCAM-1 in all subjects. The following
variables were assessed: age, ΔWC, ΔSBP, ΔFPG, ΔTG, ΔTC, and EPA treatment
(supplemental Table I and II). All statistical analyses were performed using the Stat
View program version 5.0 for Windows (SAS Institute, Cary, NC) and Excel 2003
(Microsoft, Redmond, WA) with the add-in software Statcel 2.10
References
1. Itoh M, Suganami T, Satoh N, Tanimoto-Koyama K, Yuan X, Tanaka M,
Kawano H, Yano T, Aoe S, Takeya M, Shimatsu A, Kuzuya H, Kamei Y, Ogawa
Y. Increased adiponectin secretion by highly purified eicosapentaenoic acid in
rodent models of obesity and human obese subjects. Arterioscler Thromb Vasc
Biol. 2007;27:1918-1925.
2. Azuma K, Watada H, Niihashi M, Otsuka A, Sato F, Kawasumi M, Shimada S,
Tanaka Y, Kawamori R, Mitsumata M. A new En face method is useful to
quantitate endothelial damage in vivo. Biochem Biophys Res Commun.
2003;309:384-390.
3. Mita T, Otsuka A, Azuma K, Uchida T, Ogihara T, Fujitani Y, Hirose T,
Mitsumata M, Kawamori R, Watada H. Swings in blood glucose levels

Yamada et al. Monocyte adhesion and EPA 6
accelerate atherogenesis in apolipoprotein E-deficient mice. Biochem Biophys
Res Commun. 2007;358:679-685.
4. Koto T, Nagai N, Mochimaru H, Kurihara T, Izumi-Nagai K, Satofuka S,
Shinoda H, Noda K, Ozawa Y, Inoue M, Tsubota K, Oike Y, Ishida S.
Eicosapentaenoic acid is anti-inflammatory in preventing choroidal
neovascularization in mice. Invest Ophthalmol Vis Sci. 2007;48:4328-4334.
5. Yoshida M, Takano Y, Sasaoka T, Izumi T, Kimura A. E-selectin polymorphism
associated with myocardial infarction causes enhanced leukocyte-endothelial
interactions under flow conditions. Arterioscler Thromb Vasc Biol.
2003;23:783-788.
6. Yoshida M, Sawada T, Ishii H, Gerszten RE, Rosenzweig A, Gimbrone MA, Jr.,
Yasukochi Y, Numano F. HMG-CoA reductase inhibitor modulates
monocyte-endothelial cell interaction under physiological flow conditions in
vitro: involvement of Rho GTPase-dependent mechanism. Arterioscler Thromb
Vasc Biol. 2001;21:1165-1171.
7. Nakamura N, Yoshida M, Umeda M, Huang Y, Kitajima S, Inoue Y, Ishikawa I,
Iwai T. Extended exposure of lipopolysaccharide fraction from Porphyromonas
gingivalis facilitates mononuclear cell adhesion to vascular endothelium via
Toll-like receptor-2 dependent mechanism. Atherosclerosis. 2008;196:59-67.
8. Executive Summary of The Third Report of The National Cholesterol Education
Program (NCEP) Expert Panel on Detection, Evaluation, And Treatment of High
Blood Cholesterol In Adults (Adult Treatment Panel III). JAMA.
2001;285:2486-2497.
9. Satoh N, Shimatsu A, Yamada K, Aizawa-Abe M, Suganami T, Kuzuya H,
Ogawa Y. An alpha-glucosidase inhibitor, voglibose, reduces oxidative stress

Yamada et al. Monocyte adhesion and EPA 7
markers and soluble intercellular adhesion molecule 1 in obese type 2 diabetic
patients. Metabolism. 2006;55:786-793.
10. Kashiwazaki N, Okuda Y, Seita Y, Hisamatsu S, Sonoki S, Shino M, Masaoka T,
Inomata T. Comparison of glycerol, lactamide, acetamide and dimethylsulfoxide
as cryoprotectants of Japanese white rabbit spermatozoa. J Reprod Dev.
2006;52:511-516.

Yamada et al. Monocyte adhesion and EPA 8
Legends to Supplemental Figures
Figure I. Effect of EPA on monocytes adhering to the aortic endothelium of mice 24
h after LPS treatment
(A) Representative en face views of immunohistochemical staining with Mac-2
antibody for monocytes adhering to aortic endothelium of mice 24 h after i.p. injection
of LPS with EPA treatment. Arrowheads denote Mac-2-positive cells. Scale bar, 100 μm.
(B) Quantification of Mac-2-positive cells. ** P < 0.01. n = 5 to 7.
Figure II. Serum concentrations of TG and EPA in mice 6 h and 24 h after LPS
treatment
(A) Serum EPA concentrations in mice 6 h and 24 h after i.p. injection of LPS with EPA
treatment. n = 4 to 5. (B) Serum TG concentrations in mice 6 h after i.p. injection of
LPS with EPA treatment. n = 6 to 7. (C) Serum TG concentrations in mice 24 h after i.p.
injection of LPS with EPA treatment. n = 4 to 9. * P < 0.05 and ** P < 0.01.
Figure III. Representative movies of LPS-induced monocyte rolling and adhesion
in vitro
Representative movies of rolling and adhesive monocytes on HUVECs 24 h after LPS
treatment under physiological flow conditions (available online at
http://atvb.ahajournals.org.). Movie I, Vehicle-treated HUVECs without EPA treatment.
Movie II, Vehicle-treated HUVECs with EPA treatment. Movie III, LPS-treated
HUVECs without EPA treatment. Movie IV, LPS-treated HUVECs with EPA treatment.
Figure IV. Effect of EPA on palmitate-induced monocyte adhesion in HUVECs
The number of adhesive monocytes on HUVECs 4 h and 24 h after palmitate treatment
with EPA treatment under physiological flow conditions. ** P < 0.01. n = 10 high
power fields. Data are representative of 3 separate experiments.
Figure V. Effect of EPA on TNFα-induced monocyte adhesion and expression of

Yamada et al. Monocyte adhesion and EPA 9
endothelial adhesion molecules in HUVECs
(A) The number of rolling and adhesive monocytes on HUVECs 24 h after TNFα
treatment with EPA treatment under physiological flow conditions. ** P < 0.01. n = 10
high power fields. Data are representative of 3 separate experiments. (B) Endothelial
surface expression of ICAM-1, VCAM-1, and E-selectin in HUVECs 24 h after TNFα
treatment with EPA treatment. ** P < 0.01. n = 3. Data are representative of 3 separate
experiments.
Yamada et al. Monocyte adhesion and EPA 10
Table I. Multivariate regression analysis for ΔsICAM-1 with the metabolic variables as
independent variables
Dependent variable P value P value
Multivariate
correlation
coefficient
Partial
correlation
coefficient
0.048
0.164
0.128
-0.099
-0.213
0.187
-0.388
0.711
0.226
0.352
0.481
0.139
0.150
0.008
0.011
ΔsICAM-1
Age
ΔWC
ΔSBP
ΔFPG
ΔTG
ΔTC
EPA treatment
Not entered
Not entered
Not entered
Not entered
Not entered
Not entered
-0.328
R2 = 0.224 F = 2.15 R2 = 0.107 F = 6.97
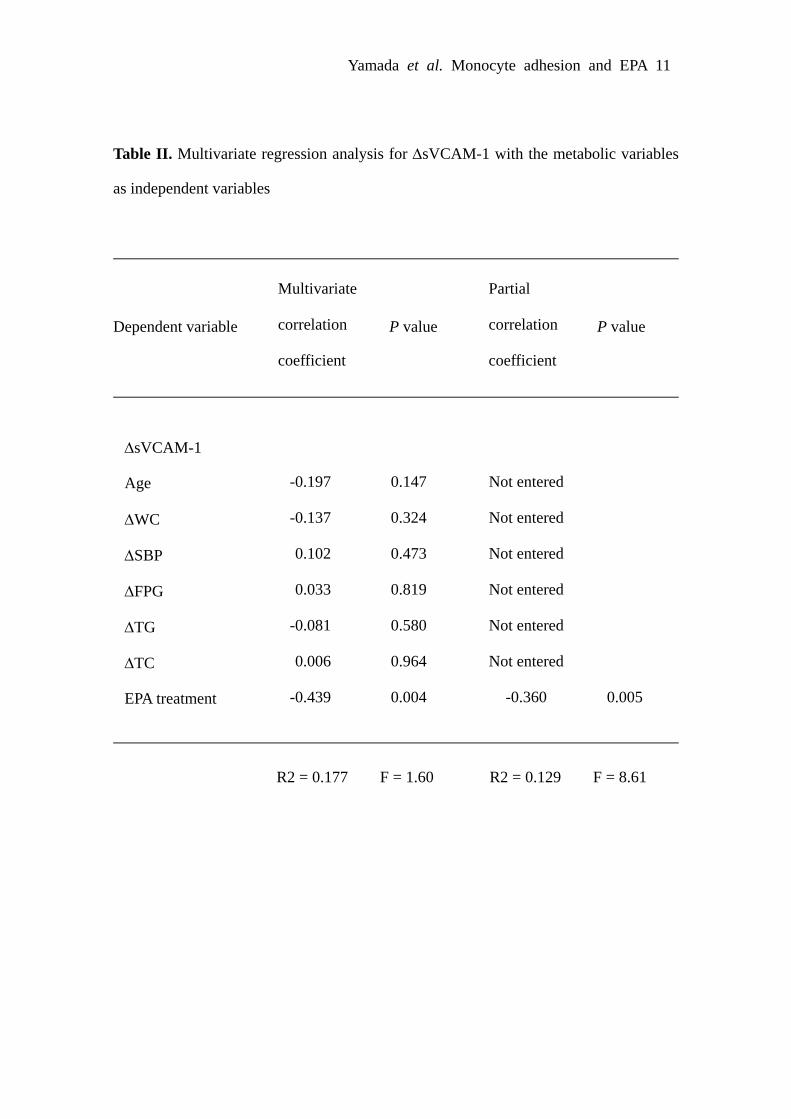
Yamada et al. Monocyte adhesion and EPA 11
Table II. Multivariate regression analysis for ΔsVCAM-1 with the metabolic variables
as independent variables
Dependent variable P value P value
Multivariate
correlation
coefficient
Partial
correlation
coefficient
-0.197
-0.137
0.102
0.033
-0.081
0.006
-0.439
0.147
0.324
0.473
0.819
0.580
0.964
0.004
0.005
ΔsVCAM-1
Age
ΔWC
ΔSBP
ΔFPG
ΔTG
ΔTC
EPA treatment
Not entered
Not entered
Not entered
Not entered
Not entered
Not entered
-0.360
R2 = 0.177 F = 1.60 R2 = 0.129 F = 8.61
![Biochimica et Biophysica Acta · 2020. 1. 4. · 2.7. Flow based endothelial cell assays Adhesion assays were performed as previously described [29]. Briefly, HDMEC cell suspension](https://img.pdfslide.us/doc/110x75/60e7bc4aef30b643b144251d/biochimica-et-biophysica-acta-2020-1-4-27-flow-based-endothelial-cell-assays.jpg)





![Cimetidine Inhibits Cancer Cell Adhesion to Endothelial ...[CANCER RESEARCH 60, 3978–3984, July 15, 2000] Cimetidine Inhibits Cancer Cell Adhesion to Endothelial Cells and Prevents](https://img.pdfslide.us/doc/110x75/608554adacb16d34563deda1/cimetidine-inhibits-cancer-cell-adhesion-to-endothelial-cancer-research-60.jpg)










![Cimetidine Inhibits Cancer Cell Adhesion to Endothelial Cells …cancerres.aacrjournals.org/content/canres/60/14/3978.full.pdf · [CANCER RESEARCH 60, 3978–3984, July 15, 2000]](https://img.pdfslide.us/doc/110x75/5d1f4ecb88c9934c378d6bc5/cimetidine-inhibits-cancer-cell-adhesion-to-endothelial-cells-cancer-research.jpg)


![Viability, proliferation and adhesion of smooth muscle ... filerecruitment of adjacent endothelial cells (EC) and endothelial progenitor cells (EPC) [13]. This requires a selective](https://img.pdfslide.us/doc/110x75/5e20b942a0730f09e1657e0b/viability-proliferation-and-adhesion-of-smooth-muscle-of-adjacent-endothelial.jpg)








