Embed Size (px)
Citation preview

Indian Journal of Experimental Biology Vol. 37, May 1999, pp. 491-495
In vitro pollen(s) - novel source of genetic variability in safflower (Carthamus tinctorius, L)
P Seeta, K Talat & SY Anwar· Department of Genetics, Osmania University, Hyderabad 500 007, India
Received 6 August /998; revised 13 January /999
For the first time, use of in vitro pollen(s) as a source for generating new genetic variability in safflower is reported. In vitro flower formation from cotyledonary explants of 2-3 days old seedlings was (52%) in cv: "Manjira" and (47%) in cv: "A-I" on MS medium supplemented with 0.5mg/L BAP and 0.1 mg/L NAA Fertility of the pollen(s) harvested from in vitro induced flowers was 80-88%. In vitro flowers of cv: "Manjira" were used as pollinators in crosses with the female parents (Manjira and KAS-I) grown in thr. field . F I hybrids obtained using in vitro pollen(s) and KAS-\ as female parent, exhibited significant improvement for characters, viz. plant height, number of primary and secondary branches and number of capitula per plant, besides, number of seeds per capitulum and seed yield per plant over the F I hybrids recovered from the cross where both the parents (Manjira and KAS-I) were grown in the field . F I hybrids with dominant phenotypes obtained from the cross where cv: "Manjira" was grown as female parent in the field and was pollinated with in vitro induced flowers of Manjira, segregated into 3 dominant : I small / striped recessive phenotypes in F2-generation. The results clearly suggest that in vitro pollen(s) can be used as an alternate source in creating genetic variability which can be utilized in conventional breeding programmes aimed at the improvement of certain agronomic traits, viz. seed yield and oil content.
Safflower is an important oil seed crop and the oil is preferred over other seed oils because of its high degree of polyunsaturation and elevated levels of cctocopherol. Oil seeds with their oil and high protein meal products are the most valuable commodity in the world trade. The process of oil seed crop improvement is complex, where simultaneous improvement in seed oil content with desired fatty acids profile without altering the seed yield is required. Therefore, rational use of different methods and tools is required to restructure the genetic architecture of oil yielding crop plants. Protocols for plantlet regeneration I , in vitro induction of capitulum2.3, induction of androgenic haploids4
.5 in safflower have been de
veloped. Besides, conventional mutagenesis6•7 and
somaclonal variationS have been used as tools in creating genetic variability. In vitro flowering is useful in generating somaclonal variations and in studies aimed at understanding the flowering process9
• However, the present report describes the results, first of its kind, on the use of in vitro pollen(s) as a source for ge.nerating new genetic variability in safflower.
Materials and Methods Three genotypes (Manjira, A-I and KAS-I) being
·Corr~spondent author
maintained at Plant Genetics experimental farm, Department of Genetics, Osmania University were used.
Efficiency oj growth regulators on the induction oj in vitro fiowers--Coty1edons devoid of embryos were excised from 2-3 days old seedlings and inoculated on MS basal medium supplemented with various concentrations of BAP (0.1, 0.5 and I mg/L) and NAA (0.05 , 0.1 and 0.5 mg/L) The cultures were maintained at 2000-2500 lux and 25°±l °C for 4 weeks. To test the efficiency of different concentrations of growth regulator in the induction of in vitro flowers, least significance difference (LSD) test was employed.
Pollen Jertility-Induced in vitro flower was put on the slide and the florets were gently removed by teasing with the needles. A drop of 1% acetocarmine was added. The pollen fertility was determined as the ratio of number of stained pollen (viable) to the number of pollen counted.
Hybridization with in vitro pollen(s}--Basic procedure of hybridization was essentially the same as described eariier lO with minor modifications. 'KAS-I' was grown in vivo in the field and was used as female parent and was poUinated with pollen from induced in vitro flowers of 'Manjira'. A parallel set of hybridisation was also conducted with the same cultivars
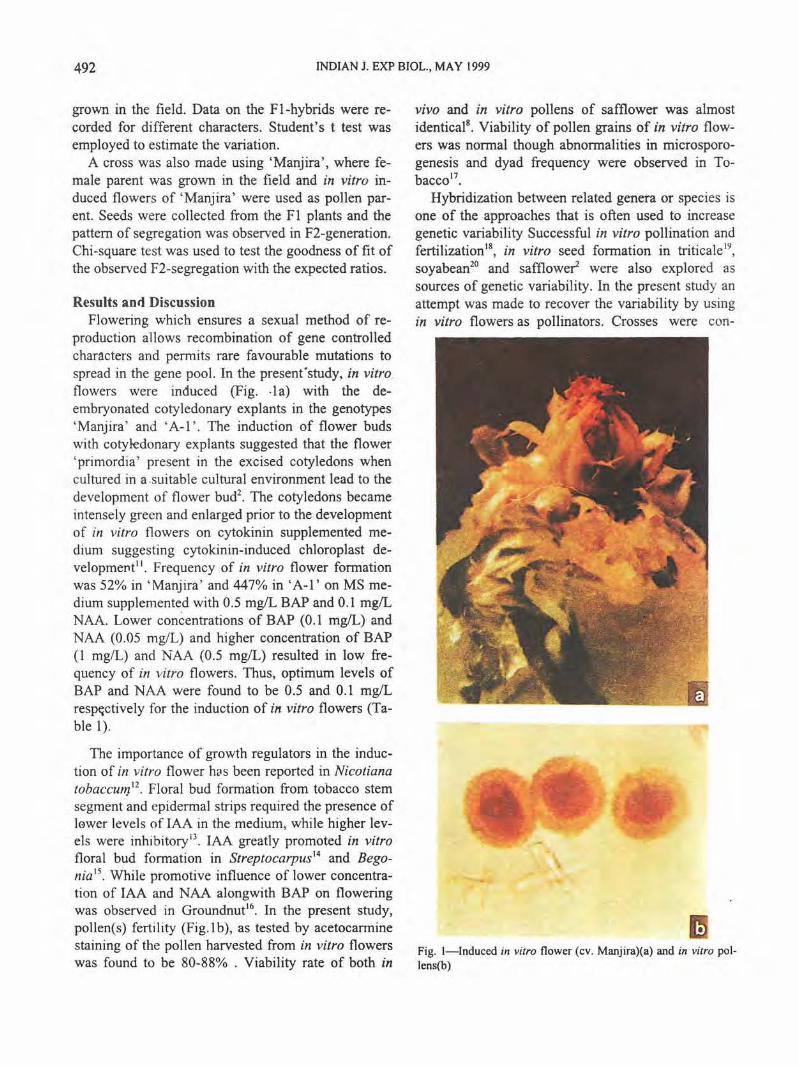
492 INDIAN J. EXP BIOL., MAY 1999
grown in the field. Data on the F1-hybrids were recorded for different characters. Student's t test was employed to estimate the variation.
A cross was also made using 'Manjira', where female parent was grown in the field and in vitro induced flowers of 'Manjira' were used as pollen parent. Seeds were collected from the F1 plants and the pattern of segregation was observed in F2-generation. Chi-square test was used to test the goodness of fit of the observed F2-segregation with the expected ratios.
Results ancl Discussion Flowering which ensures a sexual method of re
production allows recombination of gene controlled characters and permits rare favourable mutations to spread in the gene pool. In the presenfstudy. in vitro flowers were induced (Fig. ·1a) with the deembryonated cotyledonary explants in the genotypes 'Manjira' and 'A-1 ' . The induction of flower buds with cotyledonary explants suggested that the flower 'primordia' present in the excised cotyledons when cultured in a .suitable cultural environment lead to the development of flower bud2
. The cotyledons became intensely green and enlarged prior to the development of in vitro flowers on cytokinin supplemented medium suggesting cytokinin-induced chloroplast development''. Frequency of in vitro flower formation was 52% in 'Manjira' and 447% in 'A-1' on MS medium supplemented with 0.5 mg!L BAP and 0.1 mg/L NAA. Lower concentrations of BAP (0.1 mg!L) and NAA (0.05 mg/L) and higher concentration of BAP (1 mg/L) and NAA (0.5 mg/L) resulted in low frequency of in vitro flowers. Thus, optimum levels of BAP and NAA were found to be 0.5 and 0.1 mg!L resp~ctively for the induction of in vitro flowers (Table 1).
The importance of growth regulators in the induction of in vitro flower h<Js been reported in Nicotiana tobaccun] 12
• Floral bud formation from tobacco stem segment and epidermal strips required the presence of l0wer levels of IAA in the medium, while higher levels were inhibitoryJ3. IAA greatly promoted in vitro floral bud formation in Streptocarpus 14 and Begonia15. While promotive influence of lower concentration of IAA and NAA alongwith BAP on flowering was observed in Groundnut 16
• In the present study, pollen(s) fertility (Fig.lb), as tested by acetocarmine staining of the pollen harvested from in vitro flowers was found to be 80-88% . Viability rate of both in
vivo and in vitro pollens of safflower was almost identical8
• Viability of pollen grains of in vitro flowers was normal though abnormalities in microsporogenesis and dyad frequency were observed in Tobacco17.
Hybridization between related genera or species is one of the approaches that is often used to increase genetic variability Successful in vitro pollination and fertilization 18
, in vitro seed formation in triticale 19,
soyabean20 and safflower were also explored as sources of genetic variability. In the present study an attempt was made to recover the variability by using in vitro flowers as pollinators. Crosses were con-
• Fig. !-Induced in vitro flower (cv. Manjira)(a) and in vitro pollens(b)
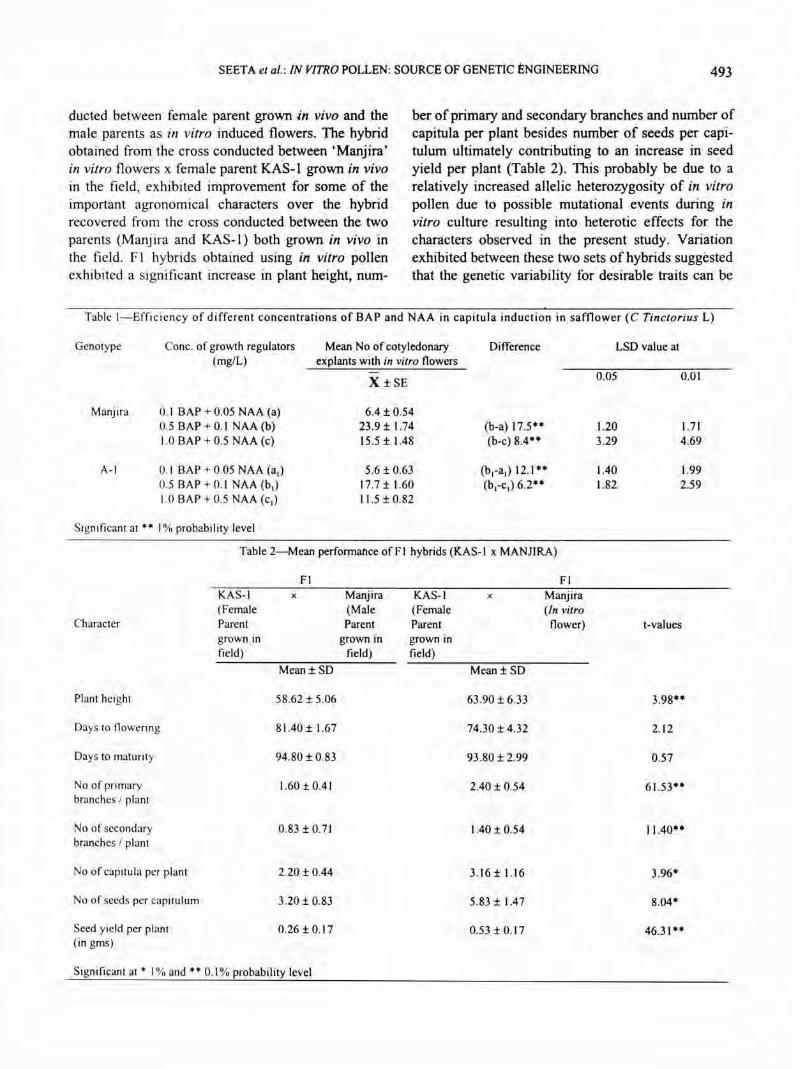
SEETA et al.: IN VITRO POLLEN: SOURCE OF GENETIC ENGINEERING 493
ducted between female parent grown in vivo and the male parents as in vitro induced flowers. The hybrid obtained from the cross conducted between 'Manjira' in vitro flowers x female parent KAS-I grown in vivo in the field , exhibited improvement for some of the important agronomical characters over the hybrid recovered from the cross conducted between the two parents (Manj ira and KAS-l) both grown in vivo in the field. Fl hybrids obtained using in vitro pollen exhibited a significant increase in plant height, num-
ber of primary and secondary branches and number of capitula per plant besides number of seeds per capitulum ultimately contributing to an increase in seed yield per plant (Table 2). This probably be due to a relatively increased allelic heterozygosity of in vitro pollen due to possible mutational events during in vitro culture resulting into heterotic effects for the characters observed in the present study. Variation exhibited between these two sets of hybrids suggested that the genetic variability for desirable traits can be
Tab le I-Efficiency of different concentrations of BAP and NAA in capitula induction in safflower (C Tinctorius L)
Genotype Conc. of growth regulators Mean No of cotyledonary Difference (mg/L) explants with in vitro flowers
X ±SE
Manjira 0.1 BAP + 0.05 NAA (a) 6.4 ± 0.54 0.5 BAP + 0.1 NAA (b) 23 .9 ± 1.74 (h-a) 17.5** 1.0 BAP + 0.5 NAA (c) 15.5±1.48 (b-c) 8.4**
A-I 0.1 BAP + 0.05 NAA (al) 5.6 ± 0.63 (hl-al) 12.1 ** 0.5 BAP + 0. 1 NAA (bl) 17.7± 1.60 (bl-cl) 6.2** 1 0 BAP + 0.5 NAA (c l) 11.5 ± 0.82
Signifi cant at ** I % probability level
Table 2-Mean performance of FI hybrids (KAS-I x MANJlRA)
Character
Plant height
Days to flowerin g
Days to maturity
No of primary branches / pl ant
No of secondary branches / pl ant
No of capi tula per pl ant
No of seeds per capi tulum
Seed yie ld per plant (i n gms)
KAS-I (Female Parent grown in field)
FI x
Mean ± SD
58.62 ± 5.06
81.40 ± 1.67
94.80 ± 0.83
1.60 ± 0.41
0.83 ± 0.71
2.20 ± 0.44
3.20 ± 0.83
0.26 ± 0.17
Significant at * I % and * * 0.1 % probability level
FI Manjira KAS-I x Manjira (Male (Female (In vitro Parent Parent flower)
grown in grown in field) field)
Mean ± SD
63.90 ± 6.33
74.30 ± 4.32
93.80 ± 2.99
2.40 ± 0.54
1.40 ± 0.54
3.16 ± \.16
5.83 ± 1.47
0.53 ± 0.17
LSD value at
0.05 0.01
1.20 1.71 3.29 4.69
1.40 1.99 1.82 2.59
t-values
3.98**
2.12
0.57
61.53**
11.40**
3.96*
8.04*
46.31**
494 INDIAN J. EXP BIOL., MAY 1999
Table :>--Segregation pattern of "seed size" and "seed hull structure" in F2 generation of cross (Manjira ~ x Manjira in vitro flower
d')
Seed Character
Seed Size (Medium vs Small)
Seed hull structure (Unstriped vs Striped)
P~0.05
Total No. of plants
79
\09
recovered by using in vitro pollen.
Dominant
58 (Medium)
80 (Un striped)
Inheritance pattern of the trait "seed size" was observed in F2-generation obtained from the cross where 'Manjira' grown in vivo under field condition was used as female and was crossed with in vitro induced flower of 'Manjira' as pollinator. Female par~ ent (Manjira) had the dominant phenotype i.e. "Medium size" seeds. Cotyledonary explant used for in vitro induction of capitulum (pollinator) was also from the "medium size" seeds of 'Manjira' . The F 1-hybrid obtained exhibited the dominant phenotype ('medium size' seed). However, the most interesting observation was the segregation of this character, i.e. seed size in F2-generation into 3 "medium seed" and 1 "small seed" with a goodness of fit for the expected chi-square value 0.105, P ~ 0.05 indicating monogenic pattern of inheritance (Table 3). Seed size (medium vs small) is a monogenic trait where 'medium' is dominant over ' small'21.
Inheritance pattern of an another important trait "seed hull structure" was also studied in F2-generation; Hybrids were recovered from the cross where in vivo grown 'Manjira ' ("Unstriped") was crossed with in vitro flowers of 'Manjira' (pollinator) .induced from the cotyledonary explant of "Unstriped" seed. The hybrid exhibited "unstriped" (dominant phenotype) seed in F I-generation similar to that of parent's seed phenotype. However in F2, segregation into 3· "unstriped" : 1 "striped:' was observed which showed goodness of fit with the expected chi-square value 0.145, P ~ 0.05 (Table 3). Seed hull stIucture (un striped vs striped) is a ~onogenic trait where ' unstriped! (STP) is dominant over 'striped ' (Stp)21 .
Segregation into recessive phenotypes observed in F2-generation can be explained only when it is as-
Phenotype F2 ratio x2 value
Recessive
21 3 : I 0.105 (Small)
29 3 : I 0.145 (Striped)
sumed that the Fl-hybrids in both the crosses must have been heterozygous with the dominant alleles contributed by the female parent (in vivo) grown in the field and recessive alleles by the in vitro pollen(s) as a result of possible mutational events during in vitro cultural passage for the induction of in vitro flowers.
Data recorded on yield components and other traits of Fl-hybrids obtained by using in vitro pollen(s), segregation of characters, viz. "seed size" and "hull structure" in F2-generation clearly suggests that the in vitro pollen(s) serve as a potential, novel and alternate source in generating variability for subsequent utilization in conventional breeding programmes aimed at the improvement of safflower crop for characters like 'seed yield' and 'oil content' as seeds with "stripped hull" (reduced hull proportion) yield more oil compared to "un striped" hull with increased hull proportion22.
References I Tejovathi G & Anwar S Y, Indian Natf Sci Acad (B), 59
( 1993) 633 . 2 Tejovathi G & Anwar S Y, PI Sci Lett 36 (1984) 165. 3 Yashodhara V, Tejovathi G & Anwar S Y, J Cyfol Genet 28
(1993) 137. 4 Rajendar Prasad B, Khadeer M A, Seeta P & Anwar S Y,
Indian J up BioI, 28 (1990) 924. 5 Rajendar Prasad B, Khadeer M A, Seeta P & Anwar S Y, PI
Cell Rep, 10(1991) 48. 6 Khadeer M A & Anwar S Y, Cytologia, 66 ( 1991) 517. 7 Talat K & Anwar S Y, Adv PI Sci, II (2) (1998) (in press). 8 Seeta P, : Induction and isolation of somaclonal variants in
safflower (Carthamus tinctorius L) PhD Thesis - Osmania University, Hyderabad, 1992.
9 Kulakami V M, Chowdhari A N, Shah B S & Harinarayana G, Indian J up BioI, 33, (1995) 142.
\0 Classen C E, Agron J, 42 (8) (1950) 381.

SEETA et al. : IN VITRO POLLEN: SOURCE OF GENETIC ENGINEERING 495
11 Sugiura M, Bot Mag, 76 (1963) 309_
12 Tran Than van M, Thisien M & Chy1ah A, Planta, 119 (1974) 149_
13 Wardell W L & Skoog F, Plant Physiol, 44 (1966) 1402_
14 Rossino L M F & Nitsch, JPCR Acad Sci, 263 (1996) 1379_
15 Ringe F & Nitsch J P, Plant Cell Physiol, 9 (1968) 639_
16 Narasimhulu S B & Reddy G M, Theor : Appl Genet, 69 (1984) 87_
17 Bridgen M P & Veilleux R E, Plant Cell Tiss and Org Cult, 13 (1988)_
18 Kanta K, Rangaswamy N S & Maheswari P, Nature, 194 (1962) 1214_
19 Reddy V D & Reddy G M, Genet Agr, 42 (1988) 151 _ 20 Dickens & Staden J V J, Plant Physiol, 20 (1985) 83_
21 Knowles P F, Adv In Agron, \0 (1958) 289_
22 Urie A L, Proc Of First Int SajJ10wer Conf Davis CA USA , (1981) 264_




![ElevatedEvolutionaryRatesamongFunctionallyDiverged ...downloads.hindawi.com/journals/ijeb/2011/274975.pdf · vertebrate genitalia [3]. Both Darwin and Eberhard explain this higher](https://img.pdfslide.us/doc/110x75/5fb951a42cfc9c0d1c1a37bc/elevatedevolutionaryratesamongfunctionallydiverged-vertebrate-genitalia-3.jpg)
























![Improving KVM x86 Nested Virtualization · qemu-system-x86-19066 [030] kvm_inj_virq: irq 210 ⇐ re-inject interrupt to L2 qemu-system-x86-19066 [030] kvm_entry: vcpu 15 ⇐ Resume](https://img.pdfslide.us/doc/110x75/5f067d127e708231d4183e65/improving-kvm-x86-nested-virtualization-qemu-system-x86-19066-030-kvminjvirq.jpg)