Embed Size (px)
Citation preview

BIOLOGY OF REPRODUCTION 44, 1177-1182 (1991)
1177
In Vitro Fertilization of Goat Oocyte&
A. I. YOUNIS, K. A. ZUELKE, K. M. HARPER, M. A. L. OLIVEIRA,3 and B. G. BRACKETI’2
Department of Physiology and Pharmacology, College of Veterinaiy Medicine
The University of Georgia, Athens, Georgia 30602
ABSTRACT
Experiments were carried out to achieve fertilization (lyE) and initial embryonic development of goat oocytes in vitro. Oo-cyte/cumulus complexes were recovered from large follicles (>7 mm) of hormonally treated does and from 1-6-mm follicles
of ovaries from hormonally superstimulated and nontreated goats. Three different sperm treatment/IVF media were used: definedmedium (Brackets and Oliphant, Biol Reprod 1975; 12:260-274 111) with modifications (mDM); TALP (Bavister and Yanagimachi,
Biol Reprod 1977; 16:228-237 [2]), as modified by Parrish et al. (Theriogenology 1986; 25:591-600 [3]), i.e. modified TALP
(mTALP); and HEPES-buffered M199 with modifications (mH-M199). Immature oocytes (from 1-6 mm, small antral follicles)
were cultured for in vitro maturation (WM) in M199 buffered with bicarbonate and with modifications including supplementation
with 20% (v/v) goat serum (mB-M199) with either (a) 100 �g L}l/ml, (b) 5 �g FSH/ml, or (c) no added gonadotropin control.
Insemination of (in vivo or in vitro) matured oocytes was performed with swim-up separated and heparin-treated freshly ejac-
ulated sperm; additionally, caffeine was included in the mDM treatment. Use of mDM yielded better results than mTALP or mH-
M199 (p < .05). Results with oocytes after IVM were significantly better than those obtained with oocytes matured in vivo
(68.4% vs. 45.5%, p <0.05). Presence of LH or FSH during oocyte maturation improved both the WM and WE results over
those of the control (p < 0.05). The highest proportion of fertilized oocytes (fertilization rate) was achieved by combining the
use of mDM for sperm and WE with IVM in the presence of LH. LH provided the highest proportion of inseminated oocytes that
cleaved, 39.5% vs. 23.3 when WM was with FSH (p < 0.05). For fertilization, mDM afforded the best results (p < 0.05) whether
oocytes were matured in vivo or in vitro. Three pregnancies were initiated after oviductal transfer of 2. and 4-cell stage embryos
resulting from WM in the presence of LH, WE, and culture.
INTRODUCTION
In vitro fertilization (IVF) with development to live young
has been reported in several large domestic species in-
cluding cattle [4, 5) sheep [6,7], pigs [6], and goats [8]. Mor-
phological description of sperm-egg union [9] and ultra-
structural studies of goat zygotes obtained in vivo have
revealed many morphological features common to other
mammalian species [10].
Requirements for in vitro capacitation of mammalian
spermatozoa have been investigated extensively [11, 12]. The
defined medium (DM) for rabbit IVF [1] was modified and
used for treatment of goat spermatozoa [13-15]. Kusunoki
et al. [15] have shown that the acrosome reaction can be
induced by incubating washed ejaculated goat spermatozoa
in DM prepared without BSA. The motility of goat sper-
matozoa can be reversibly inhibited by elevation of CO2
tension [14]. Bou and Hanada [13], using zona-free hamster
eggs in vitro, demonstrated the capacitation of ejaculated
goat sperm following treatment with calcium ionophore
Accepted February 11, 1991.
Received October 15, 1990.
‘Supported by Veterinary Medical Experiment Station (91-222), and Program in
Biological Resources and Biotechnology, The University of Georgia.
2Correspondence: FAX: 404-542-5743.
3Current address: MAL. Oliveira, Universidade Federal Rural de Pernambuco,
Departamento de Medicina veterinaria, Rua D. Manoel de Medeiros S/N, Dois Ir-
maos, 50000 Recife PR, Brasilia
A23187 and caffeine. A lamb and a kid resulting from IVF
of in vivo-matured oocytes with A23187-capacitated sperm
were reported in Japan by Hanada [8].
Oocytes recovered surgically from follicles of eCG-treated
pregnant goats have been shown to be competent for fer-
tilization and development [16]. Xenogenous fertilization of
goat follicular or ovulated oocytes in the rabbit [17] or sheep
[18] oviduct has resulted in embryos (2- to 8-cell stages)
that developed into blastocysts and live offspring.
Objectives of the present studies are to develop LYF
technology for utility in goat breeding and to provide zy-
gotes for utility in production of transgenic animals capable
of expressing valuable molecular species. Progress in de-
velopment of a repeatable IVF procedure for goat oocytes
matured in vivo and in vitro, and the developmental po-
tential of resulting IVF embryos, reported earlier in prelim-
inary form [19], is reported here.
Animal Treatments
MATERIALS AND METHODS
Experiments were carried out during the fall of 1989.
Twenty-three doe goats (Angora, Alpine, Nubian, and Tog-
genburg) maintained on pasture isolated from males were
used as oocyte donors. Nine additional goats were used as
recipients for embryos resulting from IVF. Some of the oo-
cyte donors were hormonally superstimulated by admin-
Dow
nloaded from https://academ
ic.oup.com/biolreprod/article-abstract/44/6/1177/2763115 by guest on 05 M
ay 2019

1178 YOUNIS ET AL.
istration of a total of 20 mg of FSH (F.S.H.-P, Schering Corp.,
Kenilworth, NJ) in six divided (evening and morning) i.m.
doses over 3 days: 5.0, 5.0; 2.5, 2.5; 2.5, 2.5 mg, respectively.
Concurrent with the fourth FSH injection, the donors re-
ceived a single i.m. injection of 125 �tg prostaglandin F2�
(PGF2a, Lutalyse, Upjohn Co., Kalamazoo, MI). Each oocyte
donor was anesthetized with i.m. injections of xylazine (0.2
mg/kg body weight, Rompun, Bay Vet Division, Miles Lab-
oratories, Inc., Shawnee, KS) and ketamine hydrochloride
(2-3 mg/kg, Vetalar, Parke-Davis Division, Warner-Lambert
Co., Morris Plain, NJ) and prepared for aseptic surgery.
Ovariectomy was performed 24 h after the last FSH injec-
tion, 48 h after PGFZa. Laparotomy and post-operative care
were as described below for embryo transfer.
Oocyte Treatment
In a controlled environmental room at 38#{176}C,the ovaries
were placed in a sterile petri dish containing 10 ml HEPES-
buffered medium 199, with Earle’s salts and L-glutamine (H-M199; Sigma Chemical Co., cat. no. 2520, St. Louis, MO)
modified by addition of sodium pyruvate (112 p.g/ml) and
gentamicin sulfate (50 �g/ml), mH-M199 without BSA or
serum. Under a stereomicroscope (16x), all large follicles
greater than 7 mm in diameter, were aspirated using a 20-
gauge needle attached to a 1-cc syringe. Oocytes with sur-
rounding cumulus cells were washed (4 times) in drops of
mH-M 199 without BSA or serum and evaluated for matu-
ration. Matured oocytes, i.e. with expanded cumulus cells
and/or visible first polar bodies, were randomly transfered
to droplets of medium for in vitro insemination, which was
initiated within 3 h of ovariectomy. Oocytes with signs of
degeneration were not used. Immature oocytes were sim-
ilarly obtained following dissection of small antral follicles
(1-6 mm in diameter) in mH-M199 without BSA or serum.
The oocyte/cumulus complexes selected as previously de-
scribed in the cow [22] were randomly assigned to treat-
ment groups and cultured for 24-25 h in bicarbonate-buff-
ered M199 (b-M199) with Earle’s salts and L-glutamine (Sigma,
cat. no. M5017) to which additions of sodium pyruvate (50
�g/ml), glucose (5.5 mg/mI), gentamicin sulfate (50 pg/
ml), and goat serum (heat-inactivated and mycoplasma-free;
GIBCO, Grand Island, NY) to 20% (v/v) were made. This
medium (mB-M199) was further supplemented with 100 �g
LH (NIADDK-0LH-26)/ml [21] or 5 p�g FSH (NIADDK-0FSH-
17)/mI [20] or nothing (no added hormone control). Con-
tamination of the NIADDK-oLH-26 with other anterior
pituitary hormones was negligible; determined by RIA,
contamination as percent by weight was <0.1% GH, <0.5%
TSH, <0.5% FSH, <0.1% PRL, and <0.5% vasopressin on
a molar basis (Parlow, A.F., Technical report no. 157, Pi-
tuitary Hormones and Antisera Center, 1000 W. Carson St.,
Torrance, CA 90509). The NM and all subsequent incuba-
tions of ova were maintained in droplets of medium under
silicon oil (Aldrich Chemical Co., Inc., Milwaukee, WI) and
a humidified 5% C02, 5% 02, and 90% N2 atmosphere at
38#{176}C.
Sperm Treatment and IVF
Fresh semen was obtained by electroejaculation or via
an artificial vagina from a purebred Nubian buck of proven
fertility. Three approaches identified by media tested for
sperm preparation were mDM, mTALP, and mH-M199. More
specifically, the three media were as follows: (1) defined
medium DM [1), prepared with 6.0 mg BSA/ml (rather than
3.0 mg/mI) and with gentamicin sulfate (50 �.tg/ml) and no
penicillin added, designated here as mDM; (2) Tyrode’s
medium modified to include albumin, lactate, and pyru-
vate, TAIP [2) prepared with HEPES (2.3 mg/ml) and with
other additions described by Parrish et al. [H-TALP, 3], re-
ferred to as mTALP here; (3) H-M199 supplemented as
described above and with 6.0 mg BSA/ml, designated mH-
M199. Each of these media contained BSA that was essen-
tially fatty acid-free (Sigma, cat. no. A-751 1). To each of the
above sperm-containing media, heparin (sodium salt, GradeII from porcine intestinal mucosa; Sigma, cat. no. H7005)
was added for 5 mm (10 p.g/ml) prior to transfer of sperm
to oocytes (insemination). Caffeine (388 �tg/ml final con-
centration) was also included in the mDM sperm treatment.
Routinely, a combination of two fresh ejaculates, from the
same buck, were diluted with 1.0 ml sperm medium to be
tested (mDM, mTALP, or mH-M199) just after collection and
initial gross evaluation of the semen (volume, color, gen-
eral appearance) and prior to examination for motility, con-
centration, and morphology. Sperm and IVF procedures were
similar to those used for cattle [21, 22]. Briefly, in each ex-
periment 0.2 ml of sperm suspension was overlaid with 1
ml sperm medium in each of six small test tubes (12 X 55-
mm Falcon, Becton-Dickinson Labware, Lincoln Park, NJ)
held at a 45#{176}angle at 38#{176}Cfor 1 h. Then, the top 0.8 ml
from each tube (swim-up fraction) was removed and pooled
into a sterile centrifuge tube (15 ml) and centrifuged at 350x g for 10 mm. After the supernatant was discarded, the
sperm pellet was resuspended with 4 ml fresh medium and
centrifuged as before. The resulting sperm pellet was trans-
ferred into 100 �tl of heparin (10 p�g/ml)-containing sperm
medium in a microcentrifuge tube. At this point, 388 p.g/
ml caffeine was also included in the mDM treatments.
Motility, concentration and morphology were assessed. Si-
multaneously with sperm incubation, oocytes were evalu-
ated (40 x) and those with properly expanded cumuli weretransferred into droplets. Aliquots of sperm suspensions (6-
10 �.d) to give a final concentration of 106 sperm/mi were
added to the droplets, each 50-pA droplet containing 5 oo-
cytes. Oocytes were incubated with sperm for 24 h.
In Vitro Culture and Embryo Transfer
Ova were removed from sperm-containing media for co-
culture on a monolayer of cumulus cells in mH-M199 pre-
Dow
nloaded from https://academ
ic.oup.com/biolreprod/article-abstract/44/6/1177/2763115 by guest on 05 M
ay 2019

IN VITRO FERTILIZATION OF GOAT OOCYTES 1179
pared with 10% goat serum (v/v) and no BSA. The pro-
cedure for preparation of the cumulus cell monolayer was
similar to that used for cattle [22]. This involved culture of
cumulus cells isolated during oocyte retrieval and inde-
pendently cultured until their use as monolayers at the end
of the insemination interval. Ova were examined for pro-
gressive development in culture at successive 24-h inter-
vals.
Animals were taken off feed and water 24 h prior to
administration of drugs for anesthesia. For embryo transfer
the reproductive tract was exposed via a lateral flank inci-
sion in does previously treated with acepromazine (0.2 mg/
kg, Bay Vet Division, Miles Laboratories), xylazmne (0.13 mg/
kg), and ketamine (2-3 mg/kg). Embryos in 2- to 4-cell
stages were surgically transferred via a tomcat catheter (4.5
cm, 3.5 French, open-ended catheter; Sherwood Medicals,
St. Louis, MO) into recipients’ oviducts ipsilateral to cor-
pora lutea. Five embryos in less than 50 p.l medium (mH-M199 with 10% goat serum) were deposited approximately
2 cm into the oviductal ampulla of each of nine recipient
does given PGF2a 120 h earlier. Closure was routine and
recovery from anesthesia was complete within 3 h. Goats
were maintained in the surgical area until standing, then
moved to a stall where food and water were provided dur-
ing a 24-h interval before return to pasture. Antibiotics were
used as indicated and additional postoperative care includ-
ing suture removal was routine.
Statistical Analyses
Data included the numbers of large follicles (>7 mm in
diameter) per doe (both ovaries) and the numbers of ma-
tured and immature oocytes recovered per doe; data are
reported as mean ± SEM per treated donor and per un-
treated donor; proportions of oocytes completing matura-
tion (maturation rate); each mature ovum exhibiting an ex-
panded cumulus oophorous, Metaphase II chromosomal
plate, and/or a first polar body; proportions of inseminated
oocytes (ova) that were fertilized (fertilization rate), as-
sessed at 48 h postinsemination; fertilized ova that had two
pronuclei + sperm tail or that were cleaved; and cleavagerate, indicating proportion of inseminated ova that cleaved
to at least the 2-cell stage within 48 h after insemination.
Representative ova (5-10) from each group were mounted
on slides, fixed in acetic alcohol for 24 h, and stained with
1% aceto-orcein stain. Nomarski optics facilitated identifi-
cation of Metaphase II chromosomal configurations, first
polar bodies, pronuclei and sperm tail remnants. Degen-
erated oocytes were found with heterogeneous-appearingcytoplasm, fragmentation, absence of a normal surrounding
complement of cumulus cells, etc.Proportions were compared by Chi-square analysis and
differences of p <0.05 were considered significant.
RESULTS
Qocyte Recovery and Hormonal Responses
The numbers of visible large follicles and the mean
numbers of oocytes isolated from treated and untreated do-
nors are shown in Table 1. The mean number of large fol-licles (at least 7 mm in diameter) seen (both ovaries) after
hormonal superstimulation was 33.7 ± 14.9 (range of 9-
65), which was 15-fold higher than for untreated donors
(2.2 ± 1.0). Total oocytes aspirated from ovaries of treated
does averaged 29.5 ± 10.6; of these, a mean of 3.9 (13.2%)
were degenerated and, hence, eliminated from further study.
Total oocytes aspirated from ovaries of untreated does av-
eraged 17.2 ± 4.4; on average, only 0.8 (4.7%) were dis-
carded due to their degenerated appearance at time of re-
covery. Numbers of oocytes aspirated from large follicles
judged to be in vivo-matured by their expanded cumulus
cells and/or first polar bodies are reported (Table 1). The
discrepancy between numbers of matured oocytes (2.8) and
large follicles (2.2) aspirated in the untreated group might
be explained by recovery of mature oocytes from medium-sized (5-6 mm diameter) follicles or aspiration of adjacent
follicles. Fixation and staining of representative ova en-
abled confirmation of nuclear maturation as evidenced by
Metaphase II chromosome plates and/or first polar bodies.
The mean number of in vivo-matured oocytes was 13.8 ±
6.0 per treated donor, 5-fold higher than for nontreated do-
nors (2.8 ± 1.8). Immature oocytes recovered from follicles<7 mm diameter of hormonally treated and untreated doeswere characterized by the presence of compact and unex-
panded cumuli and homogeneously dark ooplasm. Thesewere cultured and matured in vitro in experiments re-ported below. The mean numbers of immature oocytes se-
TABLE 1. Comparison of large ovarian follicles and oocytes recovered from hormonally
superstimulated and untreated doe goats.
Hormonallytreated
No. ofdoes
Num bers per doe (mean ± SEMI
Large follicles±7 mm diameter
In vivo-maturedoocytes
Immature oocytes
from follicles<7 mm diameter
Yes
No1211
33.7 ± 14.922b ± 1.0
13.8� ± 6.0
28d ± 1.811.8 ± 3.413.6 ± 3.2
“Hormonal treatment (3-day protocol described in text) consisted of 20 mg FSH and 125 �g PGF2�#{149}�da > b, c > d (p < 0.05).
Dow
nloaded from https://academ
ic.oup.com/biolreprod/article-abstract/44/6/1177/2763115 by guest on 05 M
ay 2019
1180 YOUNIS El AL.
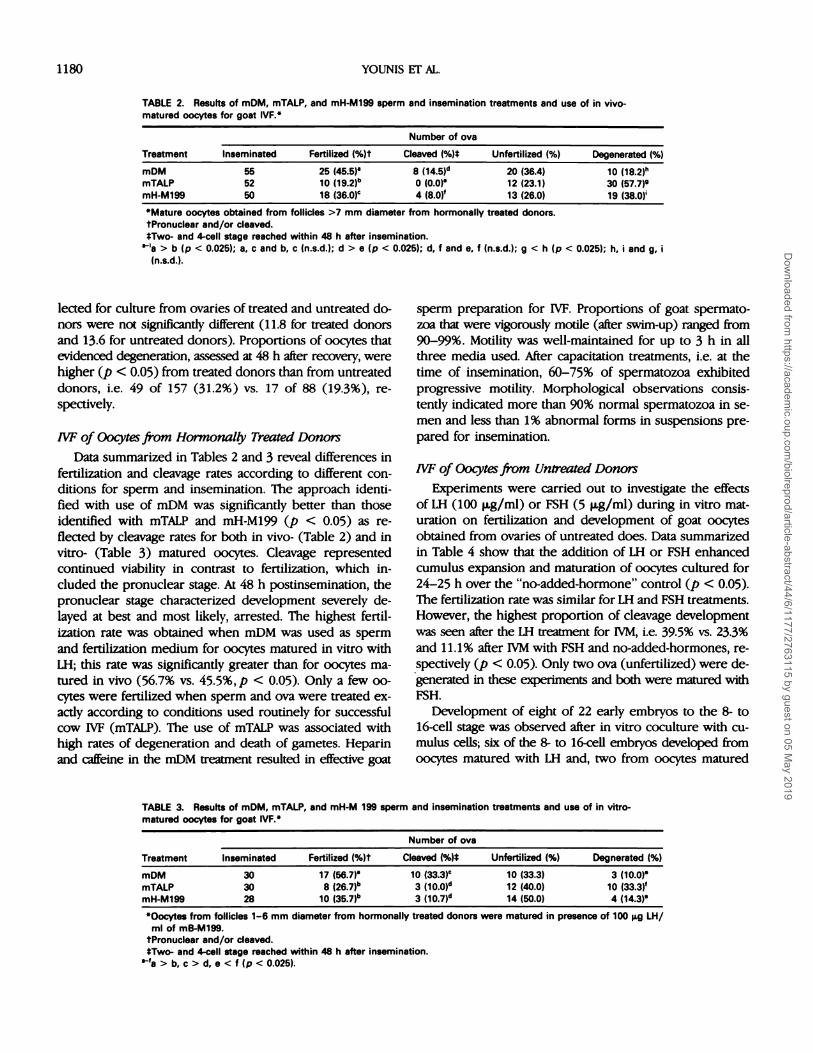
TABLE 2. Results of mDM, mTALP, and mH-M199 sperm and insemination treatments and use of in vivo-matured oocytes for goat IVF.”
Treatment
Number of ova
Inseminated Fertilized (%)t Cleaved (%)� Unfertilized (%) Degenerated (%)
mDM 55 25 (45.5)” 8 (1451d 20 (36.4) 10 (18.2)”mTALP 52 10 (19�2)b 0 (0.0)” 12 (23.1) 30 (57.7)9mH-M199 50 18 (36.0)c 4 (8.0)’ 13 (26.0) 19 (38.0)’
“Mature oocytes obtained from follicles >7 mm diameter from hormonally treated donors.
tPronuclear and/or cleaved.tTwo- and 4-cell stage reached within 48 h after insemination.
> b (p < 0.025); a, c and b, c (n.s.d.); d > e (p < 0.025); d, f and e, f (n.s.d.); g < h (p < 0.025); h, i and g, i
(n.s.d.).
lected for culture from ovaries of treated and untreated do-
nors were not significantly different (11.8 for treated donors
and 13.6 for untreated donors). Proportions of oocytes that
evidenced degeneration, assessed at 48 h after recovery, were
higher (p < 0.05) from treated donors than from untreated
donors, i.e. 49 of 157 (31.2%) vs. 17 of 88 (19.3%), re-
spectively.
IVF of Oocytes from Hormonally Treated Donors
Data summarized in Tables 2 and 3 reveal differences in
fertilization and cleavage rates according to different con-
ditions for sperm and insemination. The approach identi-
fied with use of mDM was significantly better than those
identified with mTALP and mH-M199 (p < 0.05) as re-
flected by cleavage rates for both in vivo- (Table 2) and in
vitro- (Table 3) matured oocytes. Cleavage represented
continued viability in contrast to fertilization, which in-
cluded the pronuclear stage. At 48 h postinsemination, the
pronuclear stage characterized development severely de-
layed at best and most likely, arrested. The highest fertil-
ization rate was obtained when mDM was used as sperm
and fertilization medium for oocytes matured in vitro withLH; this rate was significantly greater than for oocytes ma-
tured in vivo (56.7% vs. 45.5%,p < 0.05). Only a few oo-
cytes were fertilized when sperm and ova were treated ex-
actly according to conditions used routinely for successful
cow IVF (mTALP). The use of mTALP was associated withhigh rates of degeneration and death of gametes. Heparin
and caffeine in the mOM treatment resulted in effective goat
sperm preparation for IVF. Proportions of goat spermato-
zoa that were vigorously motile (after swim-up) ranged from
90-99%. Motility was well-maintained for up to 3 h in all
three media used. After capacitation treatments, i.e. at the
time of insemination, 60-75% of spermatozoa exhibited
progressive motility. Morphological observations consis-
tently indicated more than 90% normal spermatozoa in se-
men and less than 1% abnormal forms in suspensions pre-
pared for insemination.
IVF of Qocytes from Untreated Donors
Experiments were carried out to investigate the effects
of LH (100 p.g/ml) or FSH (5 p.g/ml) during in vitro mat-
uration on fertilization and development of goat oocytes
obtained from ovaries of untreated does. Data summarized
in Table 4 show that the addition of LH or FSH enhanced
cumulus expansion and maturation of oocytes cultured for
24-25 h over the “no-added-hormone” control (p < 0.05).
The fertilization rate was similar for LH and FSH treatments.
However, the highest proportion of cleavage development
was seen after the LH treatment for NM, i.e. 39.5% vs. 23.3%
and 11.1% after NM with FSH and no-added-hormones, re-
spectively (� < 0.05). Only two ova (unfertilized) were de-
generated in these experiments and both were matured with
FSH.
Development of eight of 22 early embryos to the 8- to
16-cell stage was observed after in vitro coculture with cu-mulus cells; six of the 8- to 16-cell embryos developed from
oocytes matured with LH and, two from oocytes matured
TABLE 3. Results of mDM, mTALP, and mH-M 199 sperm and insemination treatments and use of in vitro-matured oocytes for goat IVF.
Treatment
Number of ova
Inseminated Fertilized (%)t Cleaved (%)� Unfertilized (%) Degnerated (%)
mDM 30 17 (56.7)” 10 (33#{149}3)C 10 (33.3) 3 (10.0)”
mTALP 30 8 (267)b3 (100)d 12 (40.0) 10 (33.3)’
mH-M199 28 10 (35.7)” 3 (,07)d 14 (50.0) 4 (14.3)”
“Oocytes from follicles 1-6 mm diameter from hormonally treated donors were matured in presence of 100 �g LH/ml of mB-M199.
tPronuclear and/or cleaved.*Two- and 4-cell stage reached within 48 h after insemination.
> b, c > d, e < f (p < 0.025).
Dow
nloaded from https://academ
ic.oup.com/biolreprod/article-abstract/44/6/1177/2763115 by guest on 05 M
ay 2019

TABLE 4. Effects of in vitro maturation of goat oocytes with LH (100
�g/ml) or FSH (5 �g/ml) on IVF and cleavage.”
Number of ova
“Sperm preparation and IVF were in mDM.
tPronuclear and/or cleaved.
lTwo- to 4-cell stage reached within 48 h after insemination.
“9a>b,c>d,e>f>g(p<0.05).
IN VITRO FERTILIZATION OF GOAT OOCY�ES 1181
Maturation
treatment
Oocytes
cultured
Matured
(%)Fertilizedt
(%)Cleaved�
(%)
LH 38 38 (100.0)” 26 (68.4)’ 15 (39.5)”
FSH 33 30 (90.0)” 18 (60.7)’ 7 (23.3)’
mB-M199 15 9 (506)b 4 (444)d 1(11.1)”
with FSH (Table 4). Tubal transfer of five 2- to 4-cell stage
embryos developing after maturation in vivo with LH and
IVF in mDM to each of nine recipients resulted in three
pregnancies confirmed by ultrasonography. Although three
of nine recipients sustained embryonic development initi-
ated by IVF, resorption occurred in two recipients at ap-
proximately 2 mo, and the other pregnancy resulted in
abortion of a 3.5-mo fetus.
DISCUSSION
This report is the first to document pregnancy initiation
by embryo transfer after in vitro maturation and fertiliza-
tion of goat oocytes. Abortion of one and resorption of the
other two pregnancies suggested transfer of five embryos
to each recipient doe to be excessive and inappropriate to
allow complete gestational development. The initial report
of term development in this species involved use of in vivo-
matured oocytes [8]. Our goal was to establish an IVF sys-
tem whereby goat oocytes matured in vivo or in vitro could
be fertilized and cultured to stages suitable for gene injec-
tion and embryo transfer. 1W technology requires meth-
odologies for sperm preparation, oocyte maturation, fertil-
ization, and for fulfilling developmental potential in
conjunction with embryo transfer to appropriate recipients.
Heparmn added in all sperm treatments appeared to be ef-
fective for in vitro capacitation of goat spermatozoa. The
medium seemed to be more determinant than the pres-
ence of heparin in relation to resulting proportions of 00-
cytes undergoing fertilization and cleavage. Best results were
obtained when mDM was used as sperm and fertilization
medium. Heparin apparently binds to spermatozoa and plays
a role in sperm uptake of calcium [12,23]. Heparin has been
shown to capacitate fresh or frozen/thawed bull sperm in
a dose-dependent manner that leads to fertilization rates
above 70% [11]. Niwa and Ohgada [24] reported that caf-
feine acts synergistically with heparmn to enhance efficacy
of sperm for bovine IVF. Addition of caffeine (2 mM) to
heparin in mTALP treatment of goat sperm for IVF did not
enhance results (Younis and Brackett, unpublished data).
For oocyte maturation, two approaches were taken to
rescue ovarian oocytes from their normal fate of degen-
eration and to induce development of others. The first was
superstimulation of does with exogenous hormones to en-
hance folliculogenesis and preovulatory maturation; this was
followed by isolation and insemination of in vivo-matured
oocytes. Second, oocytes were treated to accomplish in vivo
maturation. The numbers of antral follicles developing in
ovaries of hormonally treated goats were greater than for
controls and also greater than reported for the sheep [25]
or for the cow [26, 27]. Effects of exogenous hormones in
stimulation of goat follicles have been investigated [27, 28].
The two most commonly used gonadotropin preparations
for superovulation are eCG and FSH (of porcine origin).
Comparison of effectiveness of these hormones in goats re-
vealed FSH to be superior [29,30,31]. Ovulation and IVF
rates were improved by FSH, although abnormal endocrine
patterns of estradiol, progesterone, and LH were observed
in the treated goats [30].
In the present work, an initial high dose [5.0 mg, BID.)
followed by a constant dose of 2.5 mg FSH twice daily for
2 additional days (20 mg total) led to an average of 33.7
large follicles per animal. This is higher than the average
number of corpora lutea (17.7 per cycle) reported by Nuti
et al. [31] to result from progesterone pretreatment fol-
lowed by a constant dose of 2.5 mg FSH twice daily for 3
days (15 mg total). Including those retrieved from small
follicles requiring in vitro maturation, many more oocyteswere recovered for IVF than were accounted for by grossly
visible (surface) follicles. Oocytes from hormonally treated
donors resulted in lower fertilization and cleavage devel-
opment than those obtained from untreated donors. This
probably reflected improper follicular and oocyte matura-
tion induced by exogenous FSH treatments. This conclu-
sion is further supported by the higher proportion of de-
generation seen among oocytes from hormonally treated
donors. Several experiments have shown that most prep-
arations used for superovulating sheep and cattle adversely
affect a high proportion of the oocytes and lead to abnor-
mal maturation including abnormal protein synthesis [32],
a high proportion of degenerated oocytes [33, 34], prema-
turely activated oocytes [34], and oocytes unable to mature
beyond the diakinetic stage [34]. Callensen et al. [33] have
reported that certain superovulatory treatments have been
associated with disturbed normal oocyte and follicular de-
velopment leading to embryos of inferior quality in the cow.
The ability of a mature oocyte or cumulus-oocyte complex
to fertilize and develop normally into live offspring re-
quires appropriate dynamics within its microenvironment
during the preovulatory, maturational interval.
Current efforts to rescue ovarian oocytes by isolation as
immature oocytes have demonstrated that 90-100% of the
cultured oocytes could be matured to Metaphase II in vitro
in the presence of gonadotropmns and serum. Not unex-
pectedly in light of our recent work in cattle [19,20,25],
best results of fertilization were obtained with oocytes that
were matured in medium containing a high concentration
Dow
nloaded from https://academ
ic.oup.com/biolreprod/article-abstract/44/6/1177/2763115 by guest on 05 M
ay 2019

1182 YOUNIS ET AL.
of LH. The 68.4% fertilization rate afforded by present con-
ditions was higher than that (57.7%) observed by Song and
Iritani [10] who cultured oocytes with compact cumulus cells
for 25 h and inseminated them with epididymal sperma-
tozoa preincubated for 6 h in a modified KRB medium. Also,
the present fertilization rate was higher than that (49.4%)
described by Hanada [8] and apparently without the high
incidence of polyspermy, 33.3% of fertilized oocytes. Mor-
phological aspects of goat fertilization have been published
[10,36]. Recent progress has been made in culture of IVF
goat embryos up to the 16-cell stage in the presence of
oviductal cells [37]. Attempts to achieve xenogenous fertil-
ization of goat follicular or ovulated oocytes in the rabbit
[16] or sheep [17] oviduct have had limited success; a low
proportion of embryos (2- to 8-cell stage) have been shown
to develop into blastocysts or live offspring.
IVF in many species has evolved into a useful research
tool. Animal biotechnology programs hold promise for
achieving, in a single generation, improvements in com-
mercial livestock. Application of IVF technology in the goat
promises more rapid success in gene transfer experimen-
tation than is possible in cattle and other livestock with longer
generational intervals. An aim of experiments reported here
was to obtain a large number of synchronized zygotes suit-
able for microinjection of novel genes. Other applications
of IVF technology in goats will include improvements in
production by extending the use of valuable semen, eval-
uation of sperm and/or oocyte quality, and in many facets
of experimental embryology.
ACKNOWLEDGMENTS
The authors thank Amanda Latimer for her support and encouragement; Dr. Roy
Silcox, Levent Kestintepe, and Katharine Mackie for assistance in various aspects of
this project; and Joanne Foster for secretarial assistance. The gift of purified pituitary
hormone from NIDDK of Maryland School of Medicine, is gratefully acknowledged.
REFERENCES
1. Bracken BG, Oliphant G. Capacitation of rabbit spermatozoa in vitro. Biol Reprod
1975; 12:260-274.
2. Bavister BD, Yanagimachi R The effects of sperm extracts and energy sources
on the motility and acrosome reaction of hamster spermatozoa in vitro. Biol
Reprod 1977; 16:228-237.
3. Parrish B� Susko-Parish JL, Leibfriend-Rutledge ML, Critser ES, Eyestone WH, First
NL. Bovine in vitro fertilization with frozen.thawed semen. Theriogenology 1986;
25:591-600.
4. Bracken BG, Bousquet D, Boice ML, Donawick WJ, Evans JF, Dressel MA. Normal
development following in vitro fertilization in the cow. Biol Reprod 1982; 27:147-
158.
5. Bracken BG, Keefer Cl, Troop CG, Donawick WJ, Bennett KA. Bovine twins re-
sulting from in vitro fertilization. Theriogenology 1984; 21:224.
6. Cheng WTK, Moor RM, Polge C. In vitro fertilization of pig and sheep oocvtes
matured in vivo and in vitro. Theriogenology 1986; 25:146 (abstract).
7. Crozet N, Huneau D, Desmedt V, Theron MC, Szollosi D, Torres S, Sevellec C.
In vitro fertilization with normal development in the sheep. Gamete Res 1987;
16:159-170.
8. Hanada A. In vitro fertilization in goat. Jpn J Anim Reprod 1985; 31:21-26.
9. song FIB, Iritani A. In vitro fertilization of goat follicular oocytes with epididymal
spermatozoa capacitated in a chemically defined medium. Proc 3rd MAP Anim
Sd Cong. Seoul, Korea; 1985; 1:463.
10. Crozet N, Theron MC, Chemineua P. Ukrastructure of in vivo fertilization in the
goat. Gamete Ret 1987; 18:191-199.
11. First NL, Parrish JJ. Sperm maturation and in vitro fertilization. Proc 11th tnt
Congr Anim Reprod and Al. 1988; 5:160-168.
12. Toyoda Y, Naito K P/F in domestic animals. In: Bavister BD, CumminsJ, Roldan
E (ed), Fertilization in Mammals. Norwell, MA. Serono Symposia; 1990: 335-347.
13. Bou 5, Hanada A. Penetration of zona-free hamster eggs in vitro by ejaculated
goat spermatozoa after treatment with ionophore A23187. Jpn J Anim Reprod
1985; 31(3):115-116.
14. Kato 5, Iritani A, Nishikawa Y. Effects of CO2 on motility of spermatozoa of farm
animals. Jnp J Zootech Sci 1977; 48:573-575.
15. Kusunoki H, Sakaue M, Kato S, Kanda S. Induction of acrosome reaction in ejac-
ulated goat spermatozoa by preincubation in chemically defined medium. J Exp
Zool 1989; 249:322-328.
16. Lacalandra GM, Minoia P, Dell’Aquila ME, Barile VL, Costantini V. Surgical re-
covery of oocytes from follicles of pregnant goats. Proc 11th lot Congr Anim
Reprod and Al. 1988; 2:169-171.
17. Duan EK, Wang GY, Ma BH, Wang JC. Xenogenous fertilization of goat ovulated
oocytes in the rabbit oviduct. Theriogenology 1990; 33:219 (abstract).
18. Walters DL, Kiehm DJ, Daniel 5J, Armstrong DT. Fertilization and embryo de-
velopment in sheep and goat oviducts following intra- and inter-specific oocyte
transfer. Theriogenology 1986; 25:219 (abstract).
19. Younis Al, Zuelke KA, Harper KM, Oliveira MA.L, Bracken BG. Pregnancies after
embryo transfer of in vitro fertilized goat oocytes. Biol Reprod 1990; 42 (suppl
1):128 (abstract).
20. Younis Al, Bracken BG, Fayrer-Hosken RA. Influence of serum and hormones on
maturation and fertilization of bovine oocytes in vitro. Gamete Res 1989; 23:189-
201.
21. Bracken BG, Younis Al, Fayrer-Hosken RA. Enhanced viability after in vitro fer-
tilization of bovine oocytes matured in vivo with high concentrations of lutein-
izing hormone. Fertil Steril 1989; 52:319-324.
22. Younis Al, Bracken BG. In vitro development of bovine oocytes into morula and
blastocysts. Theriogenology 1990; 33:361 (abstract).
23. Parrish JJ,5usko-Parish JL, Winer MA, First NL. Capacitation of bovine sperm by
heparin. Biol Reprod 1988; 38:1171-1180.
24. Niwa K, ohgada 0. Synergistic effect of caffeine and heparin on in fertilization
of cattle oocyte matured in culture. Theriogenology 1988; 30:733-741.
25. Bindon DM, Piper LR, Cahill LP, Driancourt MA. O’Shea T. Genetic and hormonal
factors affecting superovulation. Theriogenology 1986; 25:53-70.
26. Sirard MA, Lambert RI), Beland R, Bernard C. The effects of repeated laparos-
copic surgery used for ovarian examination and follicular aspiration in cows.
Anim Reprod Sci 1985; 9:25-30.
27. Lambert RU, Sirard MA, Bernard C, Beland R, Rioux JE, Leclere P, Menard Dl’,
Bedoya M. In vitro fertilization of bovine oocytes matured in vivo and collected
at laparoscopy. Theriogenology 1986; 25:117-133,
28. Arnoah RA, Gelaye S. Superovulation, synchronization and breeding of does. Small
Rum Res 1990; 3:63-72.
29. Armstrong UT, Pfitzner Al’, Warnes GM, Ralph MM, Seamark ES. Endocrine re-
sponses of goats after induction of superovulation with PMSG and FSH. J Reprod
Fertil 1983; 395-401.
30. Armstrong UT, Pfltzner AP, Warnes GM, Seamark ES. Superovulation treatments
and embryo transfer in Angora goats. J Reprod Fertil 1983; 403-410.
31. Nuti LC, Minhas BS, Baker WC, Capehart j5, Marrak P. Superovulation and re-
covery of zygotes from Nubian and Alpine goats. Theriogenology 1987; 28:481-
488.32. Moor RM, Osbom JC, Crosby JM. Gonadotropin-induced abnormalities in sheep
oocytes after superovulation. J Reprod Fertil 985; 74:167-172.
33. Callesen H, Greve T, Hyttel P. Pre-ovulatory endocrinology and oocyte matura-
tion in superovulated cattle. Theriogenology 1986; 25:71-86.
34. Dieleman SJ, Bevers MM, Kruip Th.AM. Maturational stage of oocytes at ovulation
in PMSG-superovulated cows. Proc 11th mt Congr Anim Reprod and Al. 1988;
2:153 (abstract 35).
35. Zuelke KA, Brackett BG. Luteinizing hormone-enhanced in vitro maturation of
bovine oocytes with and without protein supplementation. Biol Reprod 1990;
43:784-787.
36. Chartrain I, Niar A, King WA, Picard L, St. Pierre H. Development of the nucleolus
in early goat embryos. Gamete Res 1987; 18:201-213.
37. Sakkas D, BaC PA, Cameron AWN. Development of preimplantation goat (Capri
bircu.s) embryos in vivo and in vitro. J Reprod Fertil 1989; 87:359-365.
Dow
nloaded from https://academ
ic.oup.com/biolreprod/article-abstract/44/6/1177/2763115 by guest on 05 M
ay 2019





























