Embed Size (px)
Citation preview

Small Molecule Therapeutics
Immunotherapy for Hepatoma Using a Dual-Function Vectorwith Both Immunostimulatory and Pim-3–Silencing Effects
Qie Guo1,2, Peixiang Lan1, Xin Yu1, Qiuju Han1, Jian Zhang1, Zhigang Tian1,3, and Cai Zhang1
AbstractTumorigenesis is an immortalization process in which the growth of normal cells is uncontrolled and
programmed cell death is suppressed. Molecular biologic and immunologic studies have revealed that the
aberrant expression of some proto-oncogenes boosts proliferation and inhibits apoptosis, which is vital for
tumor development. The hypofunction of the host immune system also drives the development andmetastasis
ofmalignant tumors. Pim-3, amember of thePim family, is aberrantly expressed in several cancers.Data suggest
thatPim-3 inhibits apoptosis byphosphorylating theproapoptoticBH3-onlyproteinBad.Here,weconstructeda
dual-function small hairpinRNA(shRNA)vector containing an shRNAtargetingPim-3 andaTLR7-stimulating
ssRNA. Stimulation with this bi-functional vector in vitro promoted significant apoptosis of Hepa1-6 cells by
regulating the expression of apoptosis-related proteins and induced secretion of type I IFNs.Most importantly,
this bi-functional vector more effectively inhibited subcutaneous Hepa1-6 cell growth than did single shRNA
and ssRNA treatment in vivo. Natural killer (NK), CD4þT, andCD8þT cells andmacrophageswere required for
effective tumor suppression, and CD4þ T cells were shown to play a helper role in the activation of NK cells,
possibly by regulating the secretion of Th1 or Th2 cytokines. This ssRNA–shRNA bi-functional vector may
represent a promising approach for tumor therapy. Mol Cancer Ther; 13(6); 1503–13. �2014 AACR.
IntroductionAccumulated genetic and epigenetic changes that alter
the proliferation and survival pathways of normal cellshave resulted in cellular transformation and progressivetumor growth (1). Evasion of apoptosis and self-sufficien-cy in growth signals are essential for malignant growth.The proto-oncogene family Provirus integrating siteMoloney murine leukemia virus (Pim) is a highly con-served serine/threonine kinase family that has beenimplicated in cancer progression (2, 3). Three Pim kinases(Pim-1, -2, and -3) have been identified. Pim-1 and Pim-2induce cell-cycle progression in cooperation with c-Myc,acting as inhibitors of apoptosis in hematologic malig-nancies and some solid tumors (2, 3). The newest memberof the family, Pim-3, is aberrantly expressed in severalcancers, particularly those of endoderm-derived organs,
including liver, pancreas, colon, and stomach (4–7). Datasuggest that Pim-3 inhibits apoptosis by phosphorylatingand inactivating the proapoptotic BH3-only protein Bad(4–6). Pim-3 mRNA and protein were detected in humanhepatocellular carcinoma (HCC) tissues and cell lines,but not in normal hepatocytes and liver tissues. Silencingof Pim-3 by RNA interference inhibited growth andenhanced apoptosis in hepatoma cells (7). Thus, Pim-3kinase may be a candidate molecular target for cancertherapy.
Tumor pathogenesis also involves a process calledcancer immunoediting, a temporal transition fromimmune-mediated tumor elimination in early phases oftumor development to immune escape of establishedtumors (8). The ability to evade immune recognition andto suppress immune reactivity are the main methodswhereby cancers evade immune destruction (9, 10). Hostimmunosuppression, mediated by tumor cells, is charac-terized by incompetence of cytotoxic T lymphocytes(CTL), massive secretion of suppressing cytokines (suchas IL-10 and TGF-b), and activation of Treg cells, leadingto functional deficiencies in CTLs, CD4þ Th1 cells, ornatural killer (NK) cells (9–11). Thus, tumor therapymustrestimulate the immune response, in addition to suppres-sing oncogene expression.
Toll-like receptors (TLR) are pattern recognition recep-tors that trigger the innate immune response and primethe antigen-specific adaptive immune response by recog-nizing conserved structures in pathogens. TLRs areimportant in protective immunity against cancer andinfection (12). TLRs are expressed by immune and non-
Authors' Affiliations: 1Institute of Immunopharmacology & Immunother-apy, School of Pharmaceutical Sciences, Shandong University, Jinan;2Department of Pharmacy, The Affiliated Hospital of Medical College,Qingdao University, Qingdao, Shandong; and 3Department of Microbiol-ogy and Immunology, School of Life Sciences, University of Science andTechnology of China, Hefei, Anhui, China
Note: Supplementary data for this article are available at Molecular CancerTherapeutics Online (http://mct.aacrjournals.org/).
Corresponding Author: Cai Zhang, Institute of Immunopharmacology &Immunotherapy, School of Pharmaceutical Sciences, Shandong Univer-sity, 44 Wenhua West Road, Jinan 250012, China. Phone: 86-531-8838-3782; Fax: 86-531-8838-3782; E-mail: [email protected]
doi: 10.1158/1535-7163.MCT-13-0722
�2014 American Association for Cancer Research.
MolecularCancer
Therapeutics
www.aacrjournals.org 1503
on November 13, 2018. © 2014 American Association for Cancer Research. mct.aacrjournals.org Downloaded from
Published OnlineFirst April 10, 2014; DOI: 10.1158/1535-7163.MCT-13-0722

immune cells, and their ligands represent promisingimmune stimulators that could stimulate both innate andadaptive immune cells (12). Interferons (IFN) secretionfollowing TLR-mediated activation of IFN-regulatory fac-tors (IRF) is regardedas the central coordinator of immunerevival (13). TLR7 or TLR8, expressed in endosomes,recognizes natural nucleoside structures, such as viralsingle-stranded RNA (ssRNA) and synthetic compounds,for example imidazoquinolines (14, 15). U- or GU-richssRNAs, such as ssRNA40 derived fromHIV-1, are potentTLR7 activators (12, 14, 16). Binding of TLR7 with itsagonists triggers a signaling cascade, which comprisesrecruitment of MyD88, activation of the NF-kB and IRF7pathway, and production of type I IFN and inflammatorycytokines. TLR7 stimulation can prime activation of NKand T cells directly or with the help of activated antigen-presenting cells (APC) and exhibit antitumor immuneresponses (16, 17).
Here, we constructed a dual-function small hairpinRNA (shRNA) vector containing an shRNA targetingPim-3 and a TLR7-stimulating ssRNA. Stimulation withthis bi-functional vector in vitro promoted significantapoptosis of Hepa1-6 cells and induced secretion of typeI IFNs. Importantly, the vector more effectively inhibitedsubcutaneousHepa1-6 cell growth thandid single shRNAand ssRNAtreatment in vivo.NK,CD4þ, CD8þTcells, andmacrophages were required for effective tumor suppres-sion. CD4þ T cells were shown play a helper role inactivating NK cells. The bi-functional vector may repre-sent a promising approach for tumor therapy.
Materials and MethodsPlasmid construction and lentiviral packaging
Transcription of each shRNA oligonucleotide targetingPim-3 (sense-loop-antisense) was designed as a syntheticduplex with overhanging ends identical to those createdby restriction enzymedigestion (BamHI at the 50 andEcoRIat the 30), andwas cloned into vector pTZU6þ1 vector thatcontains a U6 polymerase-III (pol-III) promoter. TheshRNA template sequences are shown in SupplementaryTable S1.
Transcription of each ssRNA oligonucleotide syntheticduplex sequence (sense-terminator) was designed using asimilar overhanging ends procedure to the shRNA, andcloned into expression vector pSIREN, which contains aU6 pol-III promoter. ssRNA template sequences areshown in Supplementary Table S1. To create the dual-functional vector, the U6þshRNA in pTZU6þ1-shRNAwas digested by HindIII and EcoRI, and inserted to pSI-REN-ssRNA.
pSIREN-control, ssRNA, shRNA, or dual vectors werecloned into a lentiviral pGCSIL-GFP plasmid, and trans-fected into 293 T cells. Forty-eight hours later, culturesupernatant was collected and filtered through a 0.45-mmfilter. Viruses (LV-ctrl, LV-ssRNA, LV-shRNA, LV-dual)were harvested by centrifugation at 70,000� g at 4�C for 2hours. Harvested viruses were aliquoted and stored at�80�C.
Cell cultureMouse hepatoma cell linesHepa1-6 andH22 (Cell Bank
of the Chinese Academy of Sciences, Shanghai, China)and normal mouse hepatocyte cell line BNL.CL2 (Amer-ican Type Culture Collection, ATCC) were maintained inDMEM medium (GIBCO/BRL) supplemented with 10%heat-inactivated FBS. These cell lines were used within6 months of receipt. Cells were never used above 10passages and were cultured at 37�C in a humidifiedatmosphere with 5% CO2.
Animals, tumor challenge, and treatmentC57BL/6 mice (6–8 weeks old; Experimental Animal
Center of Beijing University, Beijing, China) were main-tained under specific pathogen-free (SPF) conditions. TheCommittee on the Ethics of Animal Experiments of theShandong University approved all the animal studies.
1� 106Hepa1-6 cells were injected subcutaneously intothe right flank of C57BL/6 mice. After 2 weeks, LV-ctrl,LV-ssRNA, LV-shRNA, LV-dual (MOI¼ 50)were admin-istered intratumorally once a week for 2 weeks. Afteranother 2 weeks, the mice were sacrificed and the tumorvolume was calculated by length � width2/2.
Human samplesHCC and nontumor liver tissue samples were obtained
from the Shandong Provincial Hospital (Jinan, China)under the National Regulation of Clinical Sampling inChina. Both were immediately fresh frozen and stored at�80�C for further use in real-time PCR and Westernblotting assays.
Semiquantitative reverse transcription-PCR andreal-time PCR analysis
Total RNAwas extracted by the TRizol regent (Invitro-gen) and cDNAs were synthesized using Superscript IIIReverse Transcriptase (Invitrogen), followed by real-timePCRand semiquantitative reverse transcription (RT)-PCRanalysis. For semiquantitative RT-PCR, cDNAwas ampli-fied using pairs of primers (RiboBio) that specificallyamplify Pim-3 or other genes, according to the manufac-turer’s protocol. For real-time PCR analysis, cDNA wasamplified with the assistance of SYBR green (Bio-Rad).Relative gene expression was determined in comparisonwith that of GAPDHor b-actin. PCRprimers are providedin Supplementary Table S2.
Western blottingCells were collected and lysed on ice using a total
protein extraction reagent (Beyotime). The protein sam-ples (30 mg/lane) were separated by sodium dodecylsulfate polyacrylamide gel electrophoresis (SDS-PAGE)and transferred to a nitrocellulose membrane (Millipore).The membrane was blocked in Tris-buffered saline with5% (w/v) nonfat dry milk, and then incubated withprimary antibodies over night at 4�C, followed by incu-bation with horseradish peroxidase–conjugated second-ary antibody for 50 minutes at room temperature.
Guo et al.
Mol Cancer Ther; 13(6) June 2014 Molecular Cancer Therapeutics1504
on November 13, 2018. © 2014 American Association for Cancer Research. mct.aacrjournals.org Downloaded from
Published OnlineFirst April 10, 2014; DOI: 10.1158/1535-7163.MCT-13-0722

Immunoreactive proteins were visualized using Molecu-lar Imager ChemiDoc XRS System BioradChemiDoc XRS(Bio-Rad). Rabbit anti-human pim-3, anti-mouse pim-3,anti-mouse Bad, anti-mouse p-Bad, anti-mouse NF-kB,and anti-mouse p-NFkBmAbs were purchased from CellSignalingTechnology (NewEnglandBioLabs Inc.). Rabbitanti-mouse p-PKR, anti-mouse IkB-a, anti-mouse bcl-2,anti-mousebcl-XL, anti-mouseBim, andanti-mouseTLR7mAbs were obtained from Santa Cruz Biotechnology.
NK cytotoxicity assaysThe ability of spleen lymphocytes to kill Hepa1-6 cells
was evaluated by CFSE/7-AAD flow cytometry assay, aspreviously described (18). Briefly, Hepa1-6 cells wereincubated with 1 mL of CFSE (2 mmol/L; MolecularProbes) for 10 minutes at 37�C and then washed. Spleenlymphocyteswere isolated and added to the target cells ateffector/target ratios of 50:1, 25:1, and 12.5:1, respectively,for 4 hours. Following a further wash, cells were labeledfor 15 minutes with 7-AAD (optimized at 0.25 mg/mL;Sigma-Aldrich) to identify dead cells. The cells were thenanalyzed via flow cytometer (FACScalibur). Cytotoxicitywas calculated as follows:% lysis¼ (CFSE/7-AADdoublepositive cells/CFSE positive cells) � 100%.
Measurement of apoptosisStaining for Annexin V-FITC/PI (BestBio) via flow
cytometry was used to detect apoptosis of tumor cells.The percentage of cells that were Annexin V-positiverepresented the proportion of apoptotic cells. Alternative-ly, apoptosiswas alsomeasuredbyTUNEL stainingusinga One Step TUNEL Apoptosis Assay Kit (Beyotime).Nuclear staining was evaluated under a light microscopevia DAPI staining (Beyotime). A commercial Enzyme-Linked Immunosorbent Assay (ELISA) Kit (KeyingMei)detected the level of caspase-8 in cell lysates, according tothe manufacturer’s instruction.
ELISA for cytokine detectionThe levels of cytokines (IFN-a, IFN-b, IFN-g , TNF-a, IL-
4, and IL-10) in culture supernatants from Hepa1-6 cellsand in the serum of tumor-bearing mice were detected byELISA kits (ExCellBiology).
Flow cytometry analysisSplenic lymphocytes were isolated to analyze the per-
centages and activation of NK and T cells after the treat-ment with shRNA, ssRNA, and dual vectors. The expres-sion of NKG2D, NKG2A, and PD-1 on lymphocytes andNKG2D ligands on Hepa1-6 cells was also detected. Cellswere harvested, blockedwith anti-FcgRmAb, and stainedwith labeledmAbs at 4�C for 45minutes. For intracellularIFN-g staining, splenic cells were cultured in RPMI 1640containing 10% FCS, and treated with monensin (Sigma)for 4 hours to inhibit extracellular secretion of cytokines.The antibodies used were: FITC-conjugated NK1.1,PerCP-Cy5.5-conjugated CD3e (BD Biosciences); FITC-conjugated CD4, PerCP-Cy5.5-conjugated CD8, PE-con-
jugated CD69, APC-conjugated NKG2D, and PE-conju-gated NKG2A (eBiosciences); and carboxyfluorescein-conjugated RAE-1, FITC-conjugated MULT-1, PE-conju-gated H-60, and PE-conjugated IFN-g mAb (R&D Sys-tems). All stained cells were analyzed using a flow cyt-ometer, and the data were processed with WinMDI2.9 software (Scripps Research Institute, La Jolla, CA).
Lymphocyte depletion and TLR7 inhibitionCell depletion mAbs were purified from PK136
(a-NK1.1), GK1.5 (a-CD4), and 2.43 (a-CD8a) hybridomacell lines. To deplete cells, tumor-bearing mice wereinjected intraperitoneally with 1 mg of mAb for 3 days(19). To deplete macrophages, 1 mg liposomes containingDMDP (dichloromethylene diphosphonate; Sigma) wasadministered intraperitoneally into C57BL/6 48 hoursbefore treatment (20). The LV-dual vector (MOI ¼ 50)was then administered intratumorally once a week for2 weeks. To ablate the function of TLR7, IRS661 (50-TGCTTGCAAGCTTGCAAGCA-30; TAKARA), a decoyanalog that interferes with the combination of TLR7 andssRNA (21, 22), was administered intravenously beforeLV-dual vector treatment.
Histochemical analysisTumor tissues were excised and fixed in 10% neutral
buffered formalin, embedded in paraffin, and sectionedand stained with hematoxylin and eosin to assess mor-phologic changes and lymphocyte infiltration.
Statistical analysesStatistical analysis was performed using a paired Stu-
dent t test andMann–WhitneyU test. P values < 0.05wereconsidered significant.
ResultsPim-3 is aberrantly expressed in mouse hepatomacell lines and HCC tissues
First, we detected the expression of Pim-3 in mousehepatoma cell lines Hepa1-6 and H22, and in the normalhepatocyte cell line BNL.CL2.Pim-3washighly expressedin Hepa1-6 and H22 cell lines, but was weakly expressedinnormal hepatocytes (Supplementary Fig. S1A).Westernblotting also showed that Pim-3 expression was higher inhepatoma Hapa1-6 and H22 cell lines, and low in BNL.CL2 cells (Supplementary Fig. S1B). Pim-3 expression inhuman primary hepatocellular carcinoma cells was sig-nificantly higher in tumor tissues than in nontumor tis-sues at both themRNAandprotein levels (SupplementaryFig. S1C andS1D). These results suggested a critical role ofPim-3 in tumorigenesis and pathogenesis of liver cancer.
Construction of a dual-function vector with bothimmunostimulatory ssRNA and Pim-3-silencingshRNA
To clarify the role of Pim-3 in cell growth and apoptosisof hepatomas and to stimulate an immune response andsilence Pim-3 expression simultaneously, we constructed
Immunotherapy for Hepatoma Using a Dual-Function Vector
www.aacrjournals.org Mol Cancer Ther; 13(6) June 2014 1505
on November 13, 2018. © 2014 American Association for Cancer Research. mct.aacrjournals.org Downloaded from
Published OnlineFirst April 10, 2014; DOI: 10.1158/1535-7163.MCT-13-0722
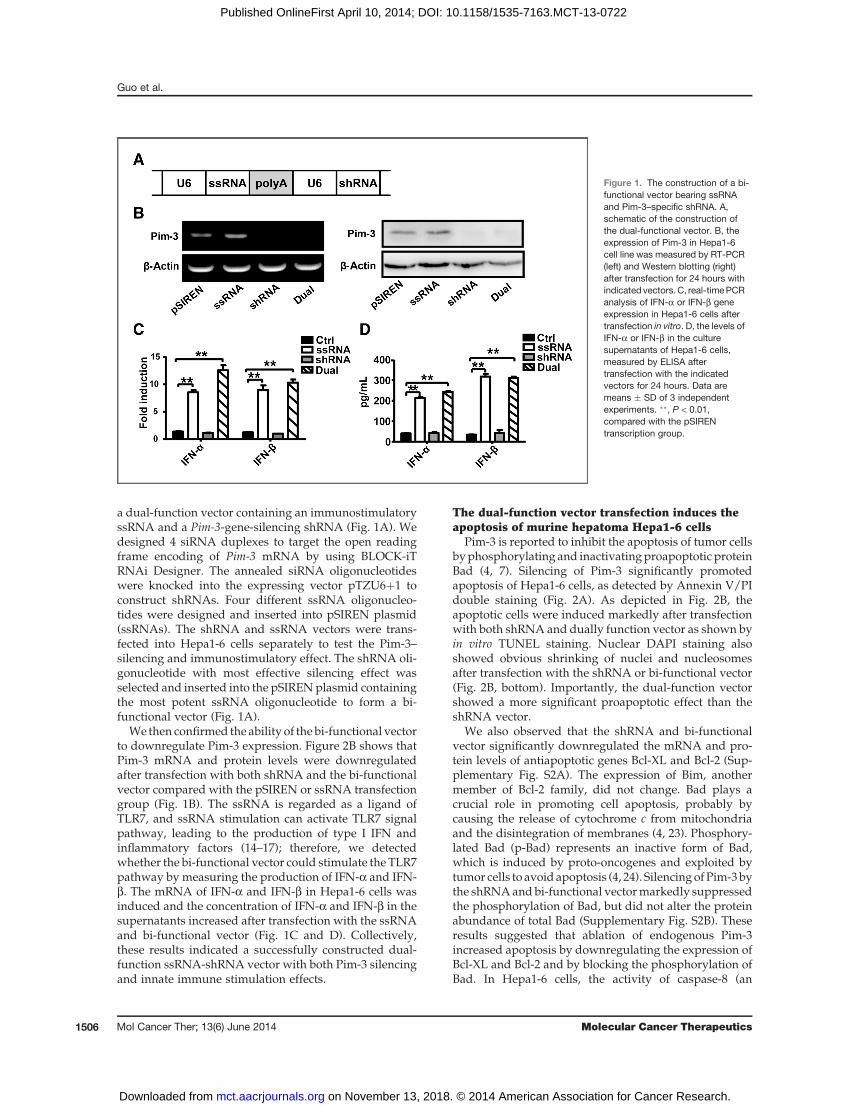
a dual-function vector containing an immunostimulatoryssRNA and a Pim-3-gene-silencing shRNA (Fig. 1A). Wedesigned 4 siRNA duplexes to target the open readingframe encoding of Pim-3 mRNA by using BLOCK-iTRNAi Designer. The annealed siRNA oligonucleotideswere knocked into the expressing vector pTZU6þ1 toconstruct shRNAs. Four different ssRNA oligonucleo-tides were designed and inserted into pSIREN plasmid(ssRNAs). The shRNA and ssRNA vectors were trans-fected into Hepa1-6 cells separately to test the Pim-3–silencing and immunostimulatory effect. The shRNA oli-gonucleotide with most effective silencing effect wasselected and inserted into the pSIRENplasmid containingthe most potent ssRNA oligonucleotide to form a bi-functional vector (Fig. 1A).
We then confirmed the ability of the bi-functional vectorto downregulate Pim-3 expression. Figure 2B shows thatPim-3 mRNA and protein levels were downregulatedafter transfection with both shRNA and the bi-functionalvector compared with the pSIREN or ssRNA transfectiongroup (Fig. 1B). The ssRNA is regarded as a ligand ofTLR7, and ssRNA stimulation can activate TLR7 signalpathway, leading to the production of type I IFN andinflammatory factors (14–17); therefore, we detectedwhether the bi-functional vector could stimulate the TLR7pathway by measuring the production of IFN-a and IFN-b. The mRNA of IFN-a and IFN-b in Hepa1-6 cells wasinduced and the concentration of IFN-a and IFN-b in thesupernatants increased after transfection with the ssRNAand bi-functional vector (Fig. 1C and D). Collectively,these results indicated a successfully constructed dual-function ssRNA-shRNA vector with both Pim-3 silencingand innate immune stimulation effects.
The dual-function vector transfection induces theapoptosis of murine hepatoma Hepa1-6 cells
Pim-3 is reported to inhibit the apoptosis of tumor cellsbyphosphorylating and inactivatingproapoptotic proteinBad (4, 7). Silencing of Pim-3 significantly promotedapoptosis of Hepa1-6 cells, as detected by Annexin V/PIdouble staining (Fig. 2A). As depicted in Fig. 2B, theapoptotic cells were induced markedly after transfectionwith both shRNA and dually function vector as shown byin vitro TUNEL staining. Nuclear DAPI staining alsoshowed obvious shrinking of nuclei and nucleosomesafter transfection with the shRNA or bi-functional vector(Fig. 2B, bottom). Importantly, the dual-function vectorshowed a more significant proapoptotic effect than theshRNA vector.
We also observed that the shRNA and bi-functionalvector significantly downregulated the mRNA and pro-tein levels of antiapoptotic genes Bcl-XL and Bcl-2 (Sup-plementary Fig. S2A). The expression of Bim, anothermember of Bcl-2 family, did not change. Bad plays acrucial role in promoting cell apoptosis, probably bycausing the release of cytochrome c from mitochondriaand the disintegration of membranes (4, 23). Phosphory-lated Bad (p-Bad) represents an inactive form of Bad,which is induced by proto-oncogenes and exploited bytumor cells to avoid apoptosis (4, 24). SilencingofPim-3bythe shRNAandbi-functional vectormarkedly suppressedthe phosphorylation of Bad, but did not alter the proteinabundance of total Bad (Supplementary Fig. S2B). Theseresults suggested that ablation of endogenous Pim-3increased apoptosis by downregulating the expression ofBcl-XL and Bcl-2 and by blocking the phosphorylation ofBad. In Hepa1-6 cells, the activity of caspase-8 (an
Figure 1. The construction of a bi-functional vector bearing ssRNAand Pim-3–specific shRNA. A,schematic of the construction ofthe dual-functional vector. B, theexpression of Pim-3 in Hepa1-6cell line was measured by RT-PCR(left) and Western blotting (right)after transfection for 24 hours withindicated vectors. C, real-timePCRanalysis of IFN-a or IFN-b geneexpression in Hepa1-6 cells aftertransfection in vitro. D, the levels ofIFN-a or IFN-b in the culturesupernatants of Hepa1-6 cells,measured by ELISA aftertransfection with the indicatedvectors for 24 hours. Data aremeans � SD of 3 independentexperiments. ��, P < 0.01,compared with the pSIRENtranscription group.
Guo et al.
Mol Cancer Ther; 13(6) June 2014 Molecular Cancer Therapeutics1506
on November 13, 2018. © 2014 American Association for Cancer Research. mct.aacrjournals.org Downloaded from
Published OnlineFirst April 10, 2014; DOI: 10.1158/1535-7163.MCT-13-0722
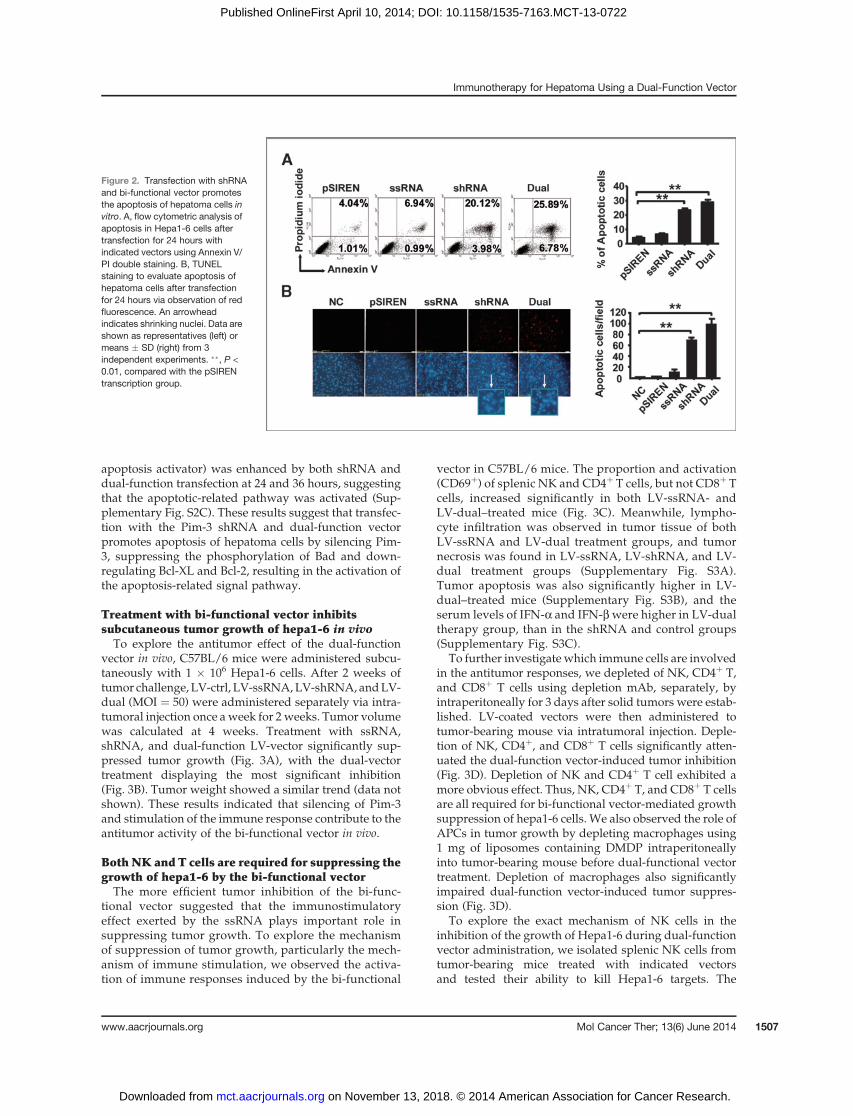
apoptosis activator) was enhanced by both shRNA anddual-function transfection at 24 and 36 hours, suggestingthat the apoptotic-related pathway was activated (Sup-plementary Fig. S2C). These results suggest that transfec-tion with the Pim-3 shRNA and dual-function vectorpromotes apoptosis of hepatoma cells by silencing Pim-3, suppressing the phosphorylation of Bad and down-regulating Bcl-XL and Bcl-2, resulting in the activation ofthe apoptosis-related signal pathway.
Treatment with bi-functional vector inhibitssubcutaneous tumor growth of hepa1-6 in vivoTo explore the antitumor effect of the dual-function
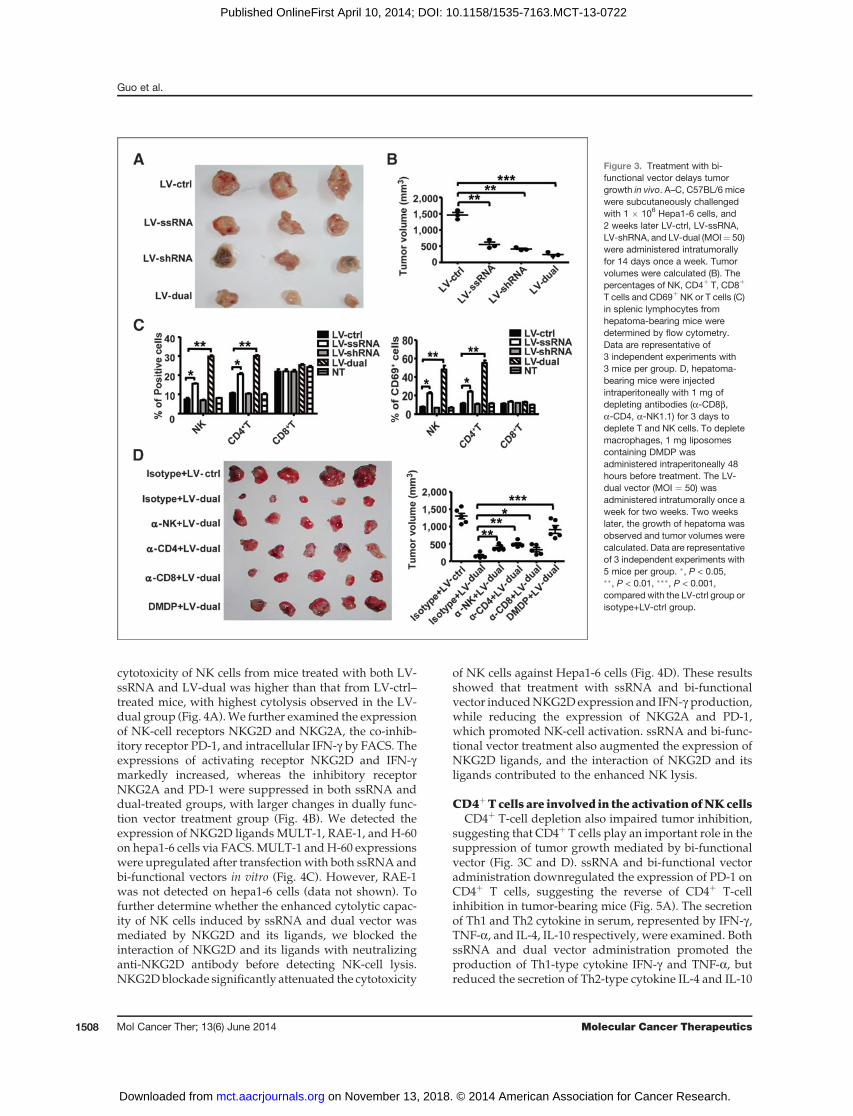
vector in vivo, C57BL/6 mice were administered subcu-taneously with 1 � 106 Hepa1-6 cells. After 2 weeks oftumor challenge, LV-ctrl, LV-ssRNA,LV-shRNA, andLV-dual (MOI ¼ 50) were administered separately via intra-tumoral injection once aweek for 2weeks. Tumor volumewas calculated at 4 weeks. Treatment with ssRNA,shRNA, and dual-function LV-vector significantly sup-pressed tumor growth (Fig. 3A), with the dual-vectortreatment displaying the most significant inhibition(Fig. 3B). Tumor weight showed a similar trend (data notshown). These results indicated that silencing of Pim-3and stimulation of the immune response contribute to theantitumor activity of the bi-functional vector in vivo.
Both NK and T cells are required for suppressing thegrowth of hepa1-6 by the bi-functional vectorThe more efficient tumor inhibition of the bi-func-
tional vector suggested that the immunostimulatoryeffect exerted by the ssRNA plays important role insuppressing tumor growth. To explore the mechanismof suppression of tumor growth, particularly the mech-anism of immune stimulation, we observed the activa-tion of immune responses induced by the bi-functional
vector in C57BL/6 mice. The proportion and activation(CD69þ) of splenic NK and CD4þ T cells, but not CD8þ Tcells, increased significantly in both LV-ssRNA- andLV-dual–treated mice (Fig. 3C). Meanwhile, lympho-cyte infiltration was observed in tumor tissue of bothLV-ssRNA and LV-dual treatment groups, and tumornecrosis was found in LV-ssRNA, LV-shRNA, and LV-dual treatment groups (Supplementary Fig. S3A).Tumor apoptosis was also significantly higher in LV-dual–treated mice (Supplementary Fig. S3B), and theserum levels of IFN-a and IFN-bwere higher in LV-dualtherapy group, than in the shRNA and control groups(Supplementary Fig. S3C).
To further investigate which immune cells are involvedin the antitumor responses, we depleted of NK, CD4þ T,and CD8þ T cells using depletion mAb, separately, byintraperitoneally for 3 days after solid tumors were estab-lished. LV-coated vectors were then administered totumor-bearing mouse via intratumoral injection. Deple-tion of NK, CD4þ, and CD8þ T cells significantly atten-uated the dual-function vector-induced tumor inhibition(Fig. 3D). Depletion of NK and CD4þ T cell exhibited amore obvious effect. Thus, NK, CD4þ T, and CD8þ T cellsare all required for bi-functional vector-mediated growthsuppression of hepa1-6 cells. We also observed the role ofAPCs in tumor growth by depleting macrophages using1 mg of liposomes containing DMDP intraperitoneallyinto tumor-bearing mouse before dual-functional vectortreatment. Depletion of macrophages also significantlyimpaired dual-function vector-induced tumor suppres-sion (Fig. 3D).
To explore the exact mechanism of NK cells in theinhibition of the growth of Hepa1-6 during dual-functionvector administration, we isolated splenic NK cells fromtumor-bearing mice treated with indicated vectorsand tested their ability to kill Hepa1-6 targets. The
Figure 2. Transfection with shRNAand bi-functional vector promotesthe apoptosis of hepatoma cells invitro. A, flow cytometric analysis ofapoptosis in Hepa1-6 cells aftertransfection for 24 hours withindicated vectors using Annexin V/PI double staining. B, TUNELstaining to evaluate apoptosis ofhepatoma cells after transfectionfor 24 hours via observation of redfluorescence. An arrowheadindicates shrinking nuclei. Data areshown as representatives (left) ormeans � SD (right) from 3independent experiments. ��, P <0.01, compared with the pSIRENtranscription group.
Immunotherapy for Hepatoma Using a Dual-Function Vector
www.aacrjournals.org Mol Cancer Ther; 13(6) June 2014 1507
on November 13, 2018. © 2014 American Association for Cancer Research. mct.aacrjournals.org Downloaded from
Published OnlineFirst April 10, 2014; DOI: 10.1158/1535-7163.MCT-13-0722
cytotoxicity of NK cells from mice treated with both LV-ssRNA and LV-dual was higher than that from LV-ctrl–treated mice, with highest cytolysis observed in the LV-dual group (Fig. 4A).We further examined the expressionof NK-cell receptors NKG2D and NKG2A, the co-inhib-itory receptor PD-1, and intracellular IFN-g by FACS. Theexpressions of activating receptor NKG2D and IFN-gmarkedly increased, whereas the inhibitory receptorNKG2A and PD-1 were suppressed in both ssRNA anddual-treated groups, with larger changes in dually func-tion vector treatment group (Fig. 4B). We detected theexpression of NKG2D ligands MULT-1, RAE-1, and H-60on hepa1-6 cells via FACS.MULT-1 andH-60 expressionswere upregulated after transfection with both ssRNA andbi-functional vectors in vitro (Fig. 4C). However, RAE-1was not detected on hepa1-6 cells (data not shown). Tofurther determine whether the enhanced cytolytic capac-ity of NK cells induced by ssRNA and dual vector wasmediated by NKG2D and its ligands, we blocked theinteraction of NKG2D and its ligands with neutralizinganti-NKG2D antibody before detecting NK-cell lysis.NKG2Dblockade significantly attenuated the cytotoxicity
of NK cells against Hepa1-6 cells (Fig. 4D). These resultsshowed that treatment with ssRNA and bi-functionalvector inducedNKG2Dexpression and IFN-g production,while reducing the expression of NKG2A and PD-1,which promoted NK-cell activation. ssRNA and bi-func-tional vector treatment also augmented the expression ofNKG2D ligands, and the interaction of NKG2D and itsligands contributed to the enhanced NK lysis.
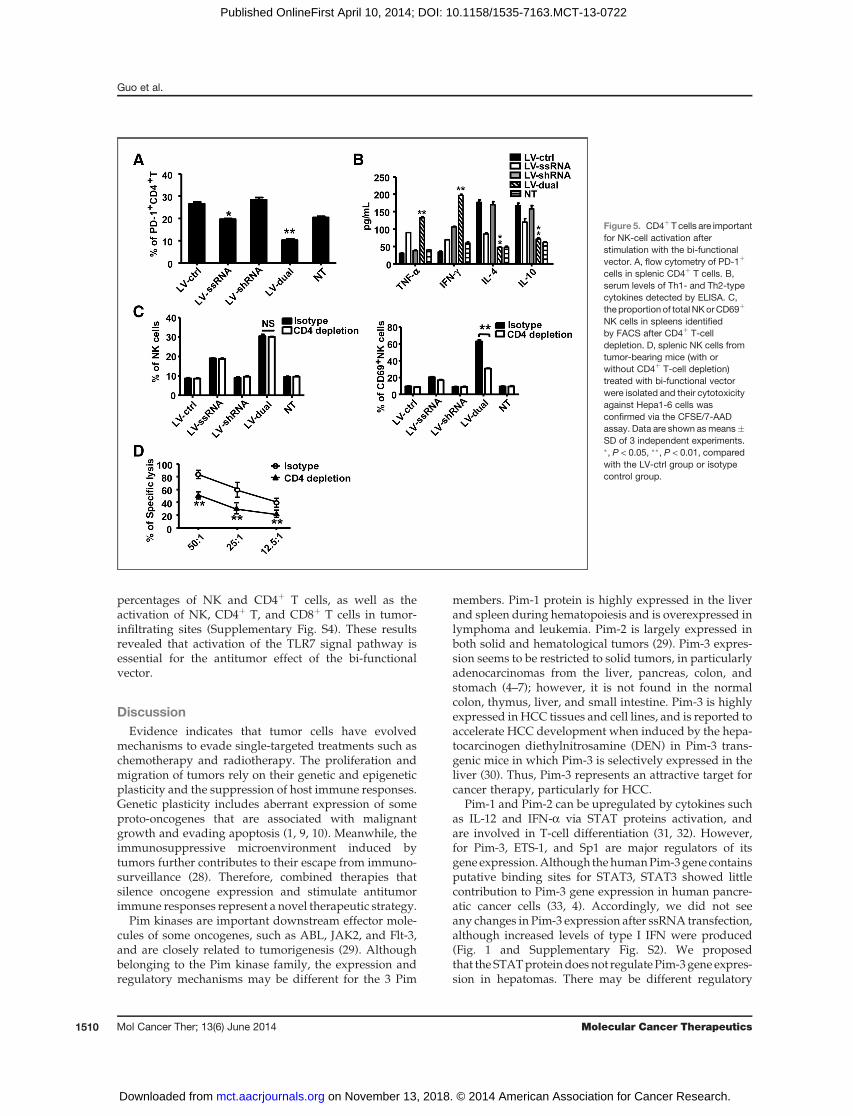
CD4þTcells are involved in the activationofNKcellsCD4þ T-cell depletion also impaired tumor inhibition,
suggesting that CD4þ T cells play an important role in thesuppression of tumor growth mediated by bi-functionalvector (Fig. 3C and D). ssRNA and bi-functional vectoradministration downregulated the expression of PD-1 onCD4þ T cells, suggesting the reverse of CD4þ T-cellinhibition in tumor-bearing mice (Fig. 5A). The secretionof Th1 and Th2 cytokine in serum, represented by IFN-g ,TNF-a, and IL-4, IL-10 respectively, were examined. BothssRNA and dual vector administration promoted theproduction of Th1-type cytokine IFN-g and TNF-a, butreduced the secretion of Th2-type cytokine IL-4 and IL-10
Figure 3. Treatment with bi-functional vector delays tumorgrowth in vivo. A–C, C57BL/6 micewere subcutaneously challengedwith 1 � 106 Hepa1-6 cells, and2 weeks later LV-ctrl, LV-ssRNA,LV-shRNA, and LV-dual (MOI¼ 50)were administered intratumorallyfor 14 days once a week. Tumorvolumes were calculated (B). Thepercentages of NK, CD4þ T, CD8þ
T cells and CD69þ NK or T cells (C)in splenic lymphocytes fromhepatoma-bearing mice weredetermined by flow cytometry.Data are representative of3 independent experiments with3 mice per group. D, hepatoma-bearing mice were injectedintraperitoneally with 1 mg ofdepleting antibodies (a-CD8b,a-CD4, a-NK1.1) for 3 days todeplete T and NK cells. To depletemacrophages, 1 mg liposomescontaining DMDP wasadministered intraperitoneally 48hours before treatment. The LV-dual vector (MOI ¼ 50) wasadministered intratumorally once aweek for two weeks. Two weekslater, the growth of hepatoma wasobserved and tumor volumes werecalculated. Data are representativeof 3 independent experiments with5 mice per group. �, P < 0.05,��, P < 0.01, ���, P < 0.001,compared with the LV-ctrl group orisotype+LV-ctrl group.
Guo et al.
Mol Cancer Ther; 13(6) June 2014 Molecular Cancer Therapeutics1508
on November 13, 2018. © 2014 American Association for Cancer Research. mct.aacrjournals.org Downloaded from
Published OnlineFirst April 10, 2014; DOI: 10.1158/1535-7163.MCT-13-0722

(Fig. 5B). We hypothesized that treatment with ssRNAand bi-functional vector promoted CD4þ T-cell prolifer-ation and activation (Fig. 3C), and shifted the balance ofTh1 and Th2 cytokine secretion by CD4þ T cells towardTh1, which contributed to the antitumor immuneresponse.CD4þTcells providehelp forNK-cell activation (25, 26);
therefore, we determined whether depletion of CD4þ Tcells impaired the activation and function of NK cells.CD4þ T-cell elimination did not influence the proportionof splenic NK cells; however, NK-cell activation wasinhibited significantly in the bi-functional vector treat-ment group (Fig. 5C).Moreover, the cytotoxicity of splenicNKcellswas significantly reduced afterCD4þT cellsweredepleted (Fig. 5D). Thus CD4þ T cells provide a necessaryhelp for NK-cell activation and play important role in theantitumor effect exerted by the bi-functional vector.
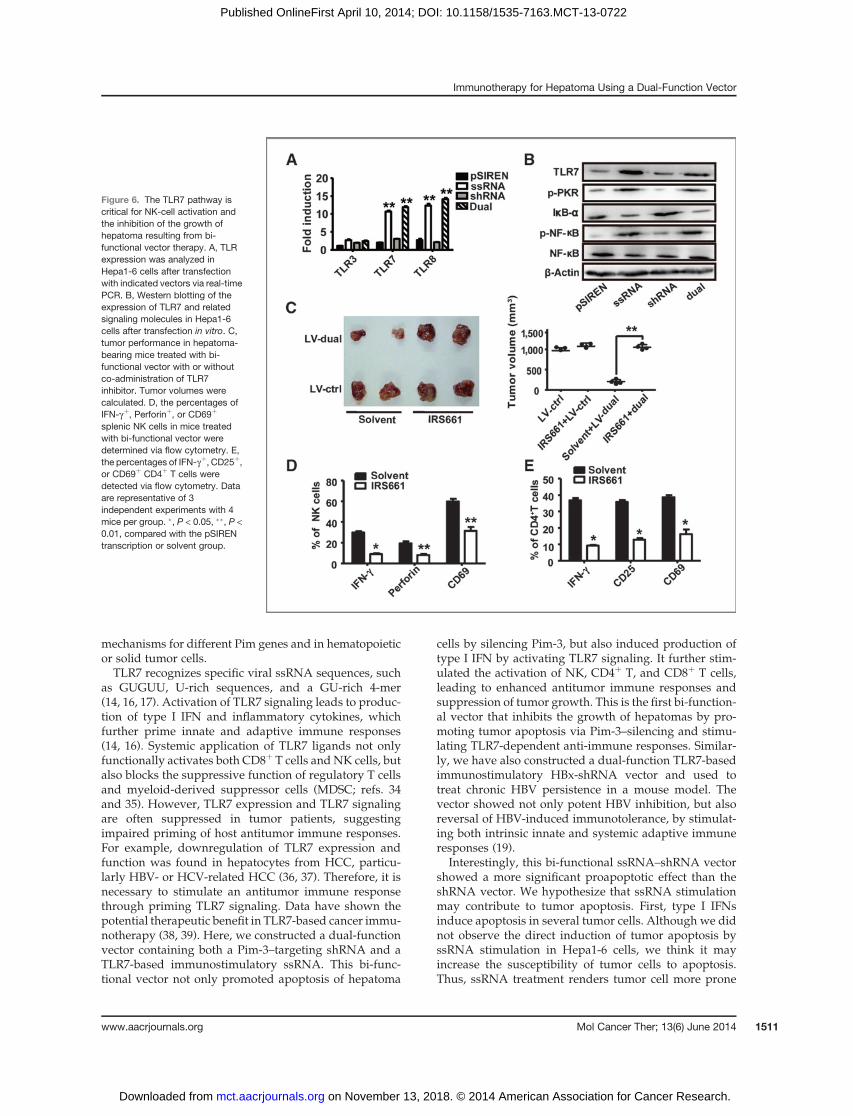
Activation of the TLR7 signal pathway is importantfor the antitumoreffectmediatedby thebi-functionalvectorTLR7 and TLR8, also known as "nucleic acid-sensing
TLRs," were originally identified as recognizing imidazo-quinoline derivatives such as imiquimod, resiquimod (R-848), and guanine analogs such as loxoribine (which haveantiviral and antitumor properties; ref. 17). TLR7 alsorecognizes ssRNA derived from RNA viruses, such asvesicular stomatitis virus, influenza A virus, and human
immunodeficiency virus (27). To further determine themechanism whereby ssRNA recognition leads toincreased NK and CD4þ T-cell activation during dualvector treatment, we evaluated TLR7 and TLR8 expres-sion in Hepa1-6 cells after dual vector transfection. Thegene expression of TLR7 and TLR8, but not TLR3, inHepa1-6 cells was significantly induced after transfectionwith ssRNA and dual-function vectors (Fig. 6A). Theprotein level of TLR7 also increased (Fig. 6B). Both vectorsalso promoted phosphorylation of NF-kB and degrada-tion of IkB-a, indicating the activation of the TLR7 signalpathway. Interestingly, the phosphorylation of anotherpattern-recognition receptor, PKR, was also upregulatedafter transfection with ssRNA and bi-functional vectors(Fig. 6B). To further confirm the role of TLR7 in the dualvector-induced immunostimulatory effect and tumorinhibition, we suppressed the TLR7 signal pathway byadministering a TLR7 inhibitor, IRS661, a decoy analogthat interferes with the combination of TLR7 and ssRNA(21), to tumor-bearingmice intravenously before LV-dualvector treatment. We found that IRS661 treatment nearlycompletely eliminated dual vector-induced tumor sup-pression (Fig. 6C). In addition, IRS661 treatment signifi-cantly attenuated the expression of IFN-g , perforin, andCD69 in/onNK cells (Fig. 6D), aswell as the expression ofIFN-g , CD25, and CD69 in/on CD4þ T cells (Fig. 6E),suggesting the functional impairment of NK and CD4þ Tcells. Similarly, IRS661 treatment markedly decreased the
Figure 4. NKcells are involved in thesuppression of hepatomasmediated by the bi-functionalvector in a NKG2D-dependentmanner. A, the cytotoxicity of NKcellswas determined bymeasuring(using CFSE/7-AAD assay) theability of splenic lymphocytes intreated mice to kill Hepa1-6 cells.B, the percentages of NKG2Dþ,NKG2Aþ, IFN-gþ, or PD-1þ NKcells were determined via FACS. C,the expression of NKG2D ligandsH-60 and MULT-1 on Hepa1-6cells was confirmed via FACS aftertransfection with indicated vectorsfor 24 hours. D, the cytotoxicity ofsplenic NK cells from hepatoma-bearing mice treated with bi-functional vectors againstHepa1-6cells, determined by the CFSE/7-AAD assay, after incubation with orwithout NKG2D blocking mAbs.Data are representative ormeans�SD of 3 independent experiments.�, P < 0.05, ��, P < 0.01, comparedwith the LV-ctrl or LV-ctrlþ isotypegroup.
Immunotherapy for Hepatoma Using a Dual-Function Vector
www.aacrjournals.org Mol Cancer Ther; 13(6) June 2014 1509
on November 13, 2018. © 2014 American Association for Cancer Research. mct.aacrjournals.org Downloaded from
Published OnlineFirst April 10, 2014; DOI: 10.1158/1535-7163.MCT-13-0722
percentages of NK and CD4þ T cells, as well as theactivation of NK, CD4þ T, and CD8þ T cells in tumor-infiltrating sites (Supplementary Fig. S4). These resultsrevealed that activation of the TLR7 signal pathway isessential for the antitumor effect of the bi-functionalvector.
DiscussionEvidence indicates that tumor cells have evolved
mechanisms to evade single-targeted treatments such aschemotherapy and radiotherapy. The proliferation andmigration of tumors rely on their genetic and epigeneticplasticity and the suppression of host immune responses.Genetic plasticity includes aberrant expression of someproto-oncogenes that are associated with malignantgrowth and evading apoptosis (1, 9, 10). Meanwhile, theimmunosuppressive microenvironment induced bytumors further contributes to their escape from immuno-surveillance (28). Therefore, combined therapies thatsilence oncogene expression and stimulate antitumorimmune responses represent a novel therapeutic strategy.
Pim kinases are important downstream effector mole-cules of some oncogenes, such as ABL, JAK2, and Flt-3,and are closely related to tumorigenesis (29). Althoughbelonging to the Pim kinase family, the expression andregulatory mechanisms may be different for the 3 Pim
members. Pim-1 protein is highly expressed in the liverand spleen during hematopoiesis and is overexpressed inlymphoma and leukemia. Pim-2 is largely expressed inboth solid and hematological tumors (29). Pim-3 expres-sion seems to be restricted to solid tumors, in particularlyadenocarcinomas from the liver, pancreas, colon, andstomach (4–7); however, it is not found in the normalcolon, thymus, liver, and small intestine. Pim-3 is highlyexpressed in HCC tissues and cell lines, and is reported toaccelerate HCC development when induced by the hepa-tocarcinogen diethylnitrosamine (DEN) in Pim-3 trans-genic mice in which Pim-3 is selectively expressed in theliver (30). Thus, Pim-3 represents an attractive target forcancer therapy, particularly for HCC.
Pim-1 and Pim-2 can be upregulated by cytokines suchas IL-12 and IFN-a via STAT proteins activation, andare involved in T-cell differentiation (31, 32). However,for Pim-3, ETS-1, and Sp1 are major regulators of itsgene expression.Although thehumanPim-3gene containsputative binding sites for STAT3, STAT3 showed littlecontribution to Pim-3 gene expression in human pancre-atic cancer cells (33, 4). Accordingly, we did not seeany changes in Pim-3 expression after ssRNA transfection,although increased levels of type I IFN were produced(Fig. 1 and Supplementary Fig. S2). We proposedthat the STATproteindoes not regulatePim-3gene expres-sion in hepatomas. There may be different regulatory
Figure 5. CD4þT cells are importantfor NK-cell activation afterstimulation with the bi-functionalvector. A, flow cytometry of PD-1þ
cells in splenic CD4þ T cells. B,serum levels of Th1- and Th2-typecytokines detected by ELISA. C,the proportion of total NKorCD69þ
NK cells in spleens identifiedby FACS after CD4þ T-celldepletion. D, splenic NK cells fromtumor-bearing mice (with orwithout CD4þ T-cell depletion)treated with bi-functional vectorwere isolated and their cytotoxicityagainst Hepa1-6 cells wasconfirmed via the CFSE/7-AADassay. Data are shown asmeans�SD of 3 independent experiments.�, P < 0.05, ��, P < 0.01, comparedwith the LV-ctrl group or isotypecontrol group.
Guo et al.
Mol Cancer Ther; 13(6) June 2014 Molecular Cancer Therapeutics1510
on November 13, 2018. © 2014 American Association for Cancer Research. mct.aacrjournals.org Downloaded from
Published OnlineFirst April 10, 2014; DOI: 10.1158/1535-7163.MCT-13-0722
mechanisms for different Pim genes and in hematopoieticor solid tumor cells.TLR7 recognizes specific viral ssRNA sequences, such
as GUGUU, U-rich sequences, and a GU-rich 4-mer(14, 16, 17). Activation of TLR7 signaling leads to produc-tion of type I IFN and inflammatory cytokines, whichfurther prime innate and adaptive immune responses(14, 16). Systemic application of TLR7 ligands not onlyfunctionally activates both CD8þ T cells andNK cells, butalso blocks the suppressive function of regulatory T cellsand myeloid-derived suppressor cells (MDSC; refs. 34and 35). However, TLR7 expression and TLR7 signalingare often suppressed in tumor patients, suggestingimpaired priming of host antitumor immune responses.For example, downregulation of TLR7 expression andfunction was found in hepatocytes from HCC, particu-larly HBV- or HCV-related HCC (36, 37). Therefore, it isnecessary to stimulate an antitumor immune responsethrough priming TLR7 signaling. Data have shown thepotential therapeutic benefit in TLR7-based cancer immu-notherapy (38, 39). Here, we constructed a dual-functionvector containing both a Pim-3–targeting shRNA and aTLR7-based immunostimulatory ssRNA. This bi-func-tional vector not only promoted apoptosis of hepatoma
cells by silencing Pim-3, but also induced production oftype I IFN by activating TLR7 signaling. It further stim-ulated the activation of NK, CD4þ T, and CD8þ T cells,leading to enhanced antitumor immune responses andsuppression of tumor growth. This is the first bi-function-al vector that inhibits the growth of hepatomas by pro-moting tumor apoptosis via Pim-3–silencing and stimu-lating TLR7-dependent anti-immune responses. Similar-ly, we have also constructed a dual-function TLR7-basedimmunostimulatory HBx-shRNA vector and used totreat chronic HBV persistence in a mouse model. Thevector showed not only potent HBV inhibition, but alsoreversal of HBV-induced immunotolerance, by stimulat-ing both intrinsic innate and systemic adaptive immuneresponses (19).
Interestingly, this bi-functional ssRNA–shRNA vectorshowed a more significant proapoptotic effect than theshRNA vector. We hypothesize that ssRNA stimulationmay contribute to tumor apoptosis. First, type I IFNsinduce apoptosis in several tumor cells. Although we didnot observe the direct induction of tumor apoptosis byssRNA stimulation in Hepa1-6 cells, we think it mayincrease the susceptibility of tumor cells to apoptosis.Thus, ssRNA treatment renders tumor cell more prone
Figure 6. The TLR7 pathway iscritical for NK-cell activation andthe inhibition of the growth ofhepatoma resulting from bi-functional vector therapy. A, TLRexpression was analyzed inHepa1-6 cells after transfectionwith indicated vectors via real-timePCR. B, Western blotting of theexpression of TLR7 and relatedsignaling molecules in Hepa1-6cells after transfection in vitro. C,tumor performance in hepatoma-bearing mice treated with bi-functional vector with or withoutco-administration of TLR7inhibitor. Tumor volumes werecalculated. D, the percentages ofIFN-gþ, Perforinþ, or CD69þ
splenic NK cells in mice treatedwith bi-functional vector weredetermined via flow cytometry. E,the percentages of IFN-gþ, CD25þ,or CD69þ CD4þ T cells weredetected via flow cytometry. Dataare representative of 3independent experiments with 4mice per group. �, P < 0.05, ��, P <0.01, compared with the pSIRENtranscription or solvent group.
Immunotherapy for Hepatoma Using a Dual-Function Vector
www.aacrjournals.org Mol Cancer Ther; 13(6) June 2014 1511
on November 13, 2018. © 2014 American Association for Cancer Research. mct.aacrjournals.org Downloaded from
Published OnlineFirst April 10, 2014; DOI: 10.1158/1535-7163.MCT-13-0722

to undergo apoptosis when Pim-3 is silenced. Second, thessRNA in the bi-functional vectormay enhance the silenc-ing effect of Pim-3-shRNA, and subsequently promote theproapoptotic effect. The exact mechanism of these effectsneeds to be further investigated.
To determine which immune cells are responsible forthe tumor regression process,we depleted ofNK,CD4þT,CD8þ T cells, and macrophages using depletion mAb orliposomes containing DMDP, respectively. We deter-mined that both NK and T cells are required for effectivetumor suppression (Fig. 3D), whereas NK cells showenhanced cytotoxicity against hepatoma via augmentedNKG2D–NKG2D ligands interaction (Fig. 4). The criticalrole of NK cells in TLR7/8 activation-mediated antitumorresponses has been reported (34, 40), and most studiesshowed that the activation of NK cells through TLR7/8recognition requires the help of APCs; however, TLR7/8signaling may exert a direct activating role on NK cells(34, 40, 41). We also demonstrated the critical role ofmacrophages in the tumor suppression mediated by thedual-function vector. We hypothesize that macrophagesand other APCs provide indispensable helper role forboth NK- and T-cell activation, possibly by activation ofTLR7 on APCs. Therefore, macrophages, CD4þ T, NK,and CD8þ T cells all contribute to the observed tumorregression. In addition, we found that CD4þ T cells pro-vide a helper role inNK-cell activation, predominantly bysecreting Th1-type cytokines. This is in agreement withobservations in other cancermodels (42, 43). Surprisingly,the inhibition of TLR7 with IRS661 completely abrogatedvector-induced tumor regression (Fig. 6). We assume thatTLR7 activation is the first issue for immune cells activa-tion-induced tumor suppression. First, type I IFNsinduced by TLR7 signaling may directly contribute to theactivation of immune cells and tumor suppression; sec-ond, type I IFNs-activated NK, CD4þ T, and CD8þ T cellsexert enhanced cyotoxicity to tumor cells; third, type IIFNs might contribute to tumor apoptosis, as described
above. However, why TLR7 inhibitor treatment can near-ly completely eliminate the dual vector-induced tumorsuppression requires further investigation.
Despite accumulating evidence showing the strongimmune activation induced by TLR7 stimulation and thesuccessful immunotherapy of skin tumors by TLR7 ago-nists when applied topically, their systemic use for thetreatment of cancer has been delayed because of TLR7tolerance by repeated administration (44, 45). Here, ther-apy with the dual-function vector provided a sustainedand long-lasting stimulation rather than short-livedimmune activation by TLR7 agonists, and thus will avoidthe TLR7 tolerance induced by repeated administration.This strategy might represent a promising therapeuticapproach in future therapy forHCCor other solid tumors,in which Pim-3 is aberrantly expressed.
Disclosure of Potential Conflicts of InterestNo potential conflicts of interest were disclosed.
Authors' ContributionsConception and design: Q. Guo, P. Lan, J. Zhang, Z. Tian, C. ZhangDevelopment of methodology: Q. GuoAcquisition of data (provided animals, acquired and managed patients,provided facilities, etc.): Q. Guo, P. LanAnalysis and interpretation of data (e.g., statistical analysis, biostatis-tics, computational analysis): Q. Guo, Q. Han, J. Zhang, C. ZhangWriting, review, andor revision of themanuscript:Q.Guo, X. Yu, Z. Tian,C. ZhangStudy supervision: Z. Tian
Grant SupportC.Zhangwas supported bygrants from theNational 973Basic Research
Program of China (#2013CB944901), the Natural Science Foundation ofChina (#81273220, #31200651), and the Young and Middle-aged ScientistAward of Shandong Province (#BS2010YY033).
The costs of publication of this article were defrayed in part by thepayment of page charges. This article must therefore be hereby markedadvertisement in accordance with 18 U.S.C. Section 1734 solely to indicatethis fact.
Received August 29, 2013; revised March 19, 2014; accepted April 3,2014; published OnlineFirst April 10, 2014.
References1. Hanahan D, Weinberg RA. The hallmarks of cancer. Cell 2000;100:
57–70.2. AdamM,PogacicV,BenditM,ChappuisR,NawijnMC,Duyster J, et al.
Targeting PIM kinases impairs survival of hematopoietic cells trans-formed by kinase inhibitor–sensitive and kinase inhibitor–resistantforms of Fms-Like tyrosine kinase 3 and BCR/ABL. Cancer Res2006;66:7–14.
3. Brault L, Gasser C, Bracher F, Huber K, Knapp S, Schwaller J. PIMserine/threonine kinases in the pathogenesis and therapy of hemato-logic malignancies and solid cancers. Haematologica 2010;95:1004–15.
4. Mukaida N, Wang YY, Li YY. Roles of Pim-3, a novel survival kinase, intumorigenesis. Cancer Sci 2011;102:1437–42.
5. Li YY, PopivanovaBK,Nagai Y, IshikuraH, Fujii C,MukaidaN. Pim-3, aproto-oncogene with serine/threonine kinase is aberrantly expressedin human pancreatic cancer and phosphorylates Bad to block Bad-mediate apoptosis in human pancreatic cancer cell lines activity.Cancer Res 2006;66:6741–7.
6. Popivanova BK, Li YY, Zheng H, Omura K, Fujii C, Tsuneyama K, et al.Proto-oncogene, Pim-3 with serine/threonine kinase activity, is aber-
rantly expressed in human colon cancer cells and can prevent Bad-mediated apoptosis. Cancer Sci 2007;98:321–8.
7. Fujii C, Nakamoto Y, Lu P. Aberrant expression of serine/threoninekinase Pim-3 in hepatocellular carcinoma development and its role inthe proliferation of human hepatoma cell lines. Int J Cancer 2005;114:209–18.
8. Bui JD, Schreiber RD. Cancer immunosurveillance, immunoeditingand inflammation: independent or interdependent processes? CurrOpin Immunol 2007;19:203–8.
9. Hanahan D, Weinberg RA. Hallmarks of cancer: the next generation.Cell 2011;144:646–74.
10. Cavallo F, De Giovanni C, Nanni P, Forni G, Lollini PL. 2011: theimmune hallmarks of cancer. Cancer Immunol Immunother 2011;60:319–26.
11. Fu YX. New immune therapy targets tumor-associated environment:from bone marrow to tumor site. Cell Mol Immunol 2012;9:1–2.
12. Kawai T, Akira S. The role of pattern-recognition receptors in innateimmunity: update on Toll-like receptors. Nat Immunol 2010, 11:373–84.
13. Noppert SJ, Fitzgerald KA, Hertzog PJ. The role of type I interferons inTLR responses. Immunol Cell Biol 2007;85:446–57.
Guo et al.
Mol Cancer Ther; 13(6) June 2014 Molecular Cancer Therapeutics1512
on November 13, 2018. © 2014 American Association for Cancer Research. mct.aacrjournals.org Downloaded from
Published OnlineFirst April 10, 2014; DOI: 10.1158/1535-7163.MCT-13-0722

14. Heil F, Hemmi H, Hochrein H, Ampenberger F, Kirschning C, Akira S,et al. Species-specific recognition of single-stranded RNA via toll-likereceptor 7 and 8. Science 2004;303:1526–9.
15. Cervantes JL,WeinermanB, BasoleC, Salazar JC. TLR8: the forgottenrelative revindicated. Cell Mol Immunol 2012;9:434–8.
16. Alter G, Suscovich TJ, Teigen N, Meier A, Streeck H, Brander C, et al.Single-stranded RNA derived from HIV-1 serves as a potent activatorof NK cells. J Immunol 2007;178:7658–66.
17. Smits EL, Ponsaerts P, Berneman ZN, Van Tendeloo VF. The use ofTLR7 and TLR8 ligands for the enhancement of cancer immunother-apy. Oncologist 2008;13:859–75.
18. Li Z, Zhang C, Zhou Z, Zhang J, Zhang J, Tian Z. Small intestinalintraepithelial lymphocytes expressing CD8 and T cell receptor gd areinvolved in bacterial clearance during Salmonella enterica serovarTyphimurium infection. Infect Immun 2012;80:565–74.
19. Lan P, Zhang C, Han Q, Zhang J, Tian Z. Therapeutic recovery of HBV-induced hepatocyte-intrinsic immune defect reverses systemic adap-tive immune tolerance. Hepatology 2013;58:73–85.
20. Van Rooijen N, Sanders A. Liposome mediated depletion of macro-phages: mechanism of action, preparation of liposomes and applica-tions. J Immunol Methods 1994;174:83–93.
21. Barrat FJ, Meeker T, Gregorio J, Chan JH, Uematsu S, Akira S, et al.Nucleic acids of mammalian origin can act as endogenous ligands forToll-like receptors and may promote systemic lupus erythematosus.J Exp Med 2005;202:1131–9.
22. Pawar RD, Ramanjaneyulu A, Kulkarni OP, Lech M, Segerer S, AndersHJ. Inhibition of Toll-like receptor-7 (TLR-7) or TLR-7 plus TLR-9attenuates glomerulonephritis and lung injury in experimental lupus.J Am Soc Nephrol 2007;18:1721–31.
23. Yan B, Zemskova M, Holder S, Chin V, Kraft A, Koskinen PJ, et al. ThePIM-2 kinase phosphorylates BAD on serine 112 and reverses BAD-induced cell death. J Biol Chem 2003;278:45358–67.
24. Aho TL, Sandholm J, Peltola KJ, Mankonen HP, Lilly M, Koskinen PJ.Pim-1 kinase promotes inactivation of the pro-apoptotic Bad proteinby phosphorylating it on the Ser112 gatekeeper site. FEBS Lett2004;571:43–9.
25. Romagnani C, Della Chiesa M, Kohler S, Moewes B, Radbruch A,Moretta L, et al. Activation of human NK cells by plasmacytoiddendritic cells and its modulation by CD4þ T helper cells and CD4þ
CD25hi T regulatory cells. Eur J Immunol 2005;35:2452–8.26. Bihl F, Pecheur J, Br�eart B, PouponG, Cazareth J, Julia V, et al. Primed
antigen-specific CD4þ T cells are required for NK cell activation in vivoupon Leishmania major infection. J Immunol 2010;185:2174–81.
27. Diebold SS, Kaisho T, Hemmi H, Akira S, Reis e Sousa C. Innate-antiviral responses by means of TLR7-mediated recognition of single-stranded RNA. Science 2004;303:1529–31.
28. Swartz MA, Iida N, Roberts EW, Sangaletti S, Wong MH, Yull FE, et al.Tumor microenvironment complexity: emerging roles in cancer ther-apy. Cancer Res 2012;72:2473–80.
29. Swords R, Kelly K, Carew J, Nawrocki S, Mahalingam D, Sarantopou-los J, et al. The Pim kinases: new targets for drug development. CurrDrug Targets 2011;12:2059–66.
30. Wu Y, Wang YY, Nakamoto Y, Li YY, Baba T, Kaneko S, et al.Accelerated hepatocellular carcinoma development in mice expres-
sing the Pim-3 transgene selectively in the liver. Oncogene 2010;29:2228–37.
31. AhoTL, LundRJ, Ylikoski EK,MatikainenS, LahesmaaR,KoskinenPJ.Expression of human pim family genes is selectively up-regulated bycytokines promoting T helper type 1, but not T helper type 2, celldifferentiation. Immunology 2005;116:82–8.
32. MatikainenS,SarenevaT,Ronni T, LehtonenA,KoskinenPJ, JulkunenI. Interferon-alpha activates multiple STAT proteins and upregulatesproliferation-associated IL-2Ra, c-myc, and pim-1 genes in human Tcells. Blood 1999;93:1980–91.
33. Li YY, Wu Y, Tsuneyama K, Baba T, Mukaida N. Essential contributionof Ets-1 to constitutive Pim-3 expression in human pancreatic cancercells. Cancer Sci 2009;100:396–404.
34. Bourquin C, Schmidt L, Lanz AL, Storch B, Wurzenberger C, Anz D,et al. Immunostimulatory RNA oligonucleotides induce an effectiveantitumoral NK cell response through the TLR7. J Immunol2009;183:6078–86.
35. Anz D, Koelzer VH, Moder S, Thaler R, Schwerd T, Lahl K, et al.Immunostimulatory RNA blocks suppression by regulatory T cells.J Immunol 2010;184:939–46.
36. Lin KJ, Lin TM, Wang CH, Liu HC, Lin YL, Eng HL. Down-regulation ofToll-like receptor 7 expression in hepatitis-virus-related human hepa-tocellular carcinoma. Hum Pathol 2013;44:534–41.
37. Chang S, Kodys K, Szabo G. Impaired expression and function of toll-like receptor 7 in hepatitis C virus infection in human hepatoma cells.Hepatology 2010;51:35–42.
38. Khairuddin N, Gantier MP, Blake SJ, Wu SY, Behlke MA, Williams BR,et al. siRNA-induced immunostimulation through TLR7 promotesantitumoral activity against HPV-driven tumors in vivo. Immunol CellBiol 2012;90:187–96.
39. Fotin-Mleczek M, Duchardt KM, Lorenz C, Pfeiffer R, Ojki�c-Zrna S,Probst J, et al. Messenger RNA-based vaccines with dual activityinduce balanced TLR-7 dependent adaptive immune responses andprovide antitumor activity. J Immunother 2011;34:1–15.
40. Berger M, Ablasser A, Kim S, Bekeredjian-Ding I, Giese T, Endres S,et al. TLR8-driven IL-12-dependent reciprocal and synergistic activa-tion of NK cells andmonocytes by immunostimulatory RNA. J Immun-other 2009;32:262–71.
41. Hart OM, Athie-Morales V, O'Connor GM, Gardiner CM. TLR7/8-mediated activation of human NK cells results in accessory cell-dependent IFN-g production. J Immunol 2005;175:1636–42.
42. Perez-Diez A, Joncker NT, Choi K, Chan WF, Anderson CC, Lantz O,et al. CD4 cells can bemore efficient at tumor rejection than CD8 cells.Blood 2007;109:5346–54.
43. Li Z, Pradera F, Kammertoens T, Li B, Liu S, Qin Z. Cross-talk betweenT cells and innate immune cells is crucial for IFN-g-dependent tumorrejection. J Immunol 2007;179:1568–76.
44. Tsukada K, Kitazawa T, Fukushima A, Okugawa S, Yanagimoto S,Tatsuno K, et al. Macrophage tolerance induced by stimulation withToll-like receptor 7/8 ligands. Immunol Lett 2007;111:51–6.
45. Bourquin C, Hotz C, Noerenberg D, Voelkl A, Heidegger S, Roetzer LC,et al. Systemic cancer therapy with a small molecule agonist of toll-likereceptor 7 can be improved by circumventing TLR tolerance. CancerRes 2011;71:5123–33.
www.aacrjournals.org Mol Cancer Ther; 13(6) June 2014 1513
Immunotherapy for Hepatoma Using a Dual-Function Vector
on November 13, 2018. © 2014 American Association for Cancer Research. mct.aacrjournals.org Downloaded from
Published OnlineFirst April 10, 2014; DOI: 10.1158/1535-7163.MCT-13-0722

2014;13:1503-1513. Published OnlineFirst April 10, 2014.Mol Cancer Ther Qie Guo, Peixiang Lan, Xin Yu, et al.
Silencing Effects−Both Immunostimulatory and Pim-3 Immunotherapy for Hepatoma Using a Dual-Function Vector with
Updated version
10.1158/1535-7163.MCT-13-0722doi:
Access the most recent version of this article at:
Material
Supplementary
http://mct.aacrjournals.org/content/suppl/2014/04/10/1535-7163.MCT-13-0722.DC1
Access the most recent supplemental material at:
Cited articles
http://mct.aacrjournals.org/content/13/6/1503.full#ref-list-1
This article cites 45 articles, 19 of which you can access for free at:
Citing articles
http://mct.aacrjournals.org/content/13/6/1503.full#related-urls
This article has been cited by 1 HighWire-hosted articles. Access the articles at:
E-mail alerts related to this article or journal.Sign up to receive free email-alerts
Subscriptions
Reprints and
To order reprints of this article or to subscribe to the journal, contact the AACR Publications Department at
Permissions
Rightslink site. Click on "Request Permissions" which will take you to the Copyright Clearance Center's (CCC)
.http://mct.aacrjournals.org/content/13/6/1503To request permission to re-use all or part of this article, use this link
on November 13, 2018. © 2014 American Association for Cancer Research. mct.aacrjournals.org Downloaded from
Published OnlineFirst April 10, 2014; DOI: 10.1158/1535-7163.MCT-13-0722





























