Embed Size (px)
Citation preview

Journal of Dermatological Science 55 (2009) 99–107
Immunohistochemical survey of the distribution of epidermal melanoblastsand melanocytes during the development of UVB-induced pigmented spots
Rikako Furuya a,*, Yuzo Yoshida a, Osamu Moro a, Makoto Tsunenaga a, Hirofumi Aoki a,Jiro Kishimoto a, Ohji Ifuku a, Tomohisa Hirobe b,c
a Shiseido Research Center, Fukuura, Kanazawa-ku, Yokohama, Japanb Radiation Effect Mechanism Research Group, National Institute of Radiological Sciences, Anagawa, Inage-ku, Chiba, Japanc Graduate School of Science, Chiba University, Yayoi-cho, Inage-ku, Chiba, Japan
A R T I C L E I N F O
Article history:
Received 10 November 2008
Received in revised form 21 February 2009
Accepted 25 March 2009
Keywords:
UVB
Melanocyte
Solar lentigo
Pigmentation
TRP-1
TRP-2
c-kit
A B S T R A C T
Background: Repeated exposures to ultraviolet B radiation (UVB) induce pigmented spots on dorsal skin
of (HR-1 � HR/De) F1 hairless mouse. We showed previously that this mouse is suitable for studies of
melanocyte function.
Objective: To clarify the mechanism of development of pigmented spots induced by chronic UVB
exposure.
Methods: We used light and fluorescence microscopy to quantify changes in the numbers of
differentiated melanocytes containing melanin pigments (MM) and melanoblasts/melanocytes
immunohistochemically positive for tyrosinase-related protein (TRP)-1, TRP-2 (dopachrome tautomer-
ase), and c-kit in epidermis during the development of pigmented spots in hairless mice chronically
exposed to UVB (99 mJ/cm2, 3 times/week, 8 weeks).
Results: The change in the number of TRP-1-positive cells during chronic UVB exposure was similar to
that of MM: both increased dramatically during the stage of acute pigmentation, then decreased sharply
after cessation of UVB, concomitantly with depigmentation; subsequently they increased gradually with
the development of pigmented spots. In contrast, after two UVB exposures, no c-kit-positive cells were
detected, then the number gradually increased during UVB irradiation, and continued to increase after
cessation of irradiation; TRP-2-positive cells showed a rather similar pattern, except that they did not
disappear initially.
Conclusion: Our results indicate that chronic UVB irradiation induces differentiation and proliferation of
melanoblasts, followed by an increase of differentiated melanocytes, leading to the development of
pigmented spots. The sequence of expression of markers appeared to be c-kit, TRP-2, TRP-1, and finally
melanin, as it is during normal melanocyte differentiation.
� 2009 Japanese Society for Investigative Dermatology. Published by Elsevier Ireland Ltd. All rights reserved.
Contents lists available at ScienceDirect
Journal of Dermatological Science
journa l homepage: www. int l .e lsev ierhea l th .com/ journa ls / jods
1. Introduction
There have been numerous studies on the hyperpigmentationinduced by ultraviolet (UV) irradiation [1–8]. Various growthfactors, cytokines, or chemical mediators, such as basic fibroblastgrowth factor (bFGF; [9]), endothelins [10], granulocyte-macro-phage colony stimulating factor (GM-CSF; [11]), stem cell factor[12], nitric oxide [13] and arachidonic acid [14], are secreted fromUV-irradiated keratinocytes and stimulate proliferation and/ormelanogenesis of melanocytes. However, almost all of these
* Corresponding author at: Shiseido Research Center (KANAZAWA-HAKKEI), 2-
12-1 Fukuura, Kanazawa-ku, Yokohama 236-8643, Japan. Tel.: +81 45 590 6000;
fax: +81 45 590 6087.
E-mail address: [email protected] (R. Furuya).
0923-1811/$36.00 � 2009 Japanese Society for Investigative Dermatology. Published b
doi:10.1016/j.jdermsci.2009.03.012
reports dealt mainly with the acute effects of UV irradiation; thechronic effects of repeated UV exposures have not been wellstudied. In addition, many studies have focused on melanocytestates in pigmented spots caused by UV-irradiation, e.g., solarlentigo [15–19] and in aging skin [20–22], but there is littleinformation about long-term changes in melanocytes after UVirradiation. Naganuma et al. [23] reported that repeated UVB-exposures induce pigmented spots in Hos:(HR-1 � HR/De)F1 micelong after the cessation of irradiation. Hos:(HR-1 � HR/De)F1 micehave many melanocytes in the epidermis [24] and are suitable forthe study of melanocyte function. Pigmented spots induced by UVirradiation in mice may be a good animal model for humanlentigines and freckles.
We have shown that the proliferative activity of epidermalmelanoblasts and melanocytes in serum-free primary culture[25,26] changes during the development of pigmented spots
y Elsevier Ireland Ltd. All rights reserved.

R. Furuya et al. / Journal of Dermatological Science 55 (2009) 99–107100
induced by UVB irradiation [27], and keratinocytes are involvedin regulating this process [28–30]. However, little is knownabout the stages of differentiation of melanoblasts or melano-cytes present in the pigmented spots. In this study, we selectedthree differentiation markers, i.e., tyrosinase-related protein(TRP)-1, TRP-2 (dopachrome tautomerase, DCT), and c-kit. TRP-2and c-kit are mainly markers of early melanoblasts. In addition,tyrosinase, TRP-1, and TRP-2 are markers of differentiatedmelanocytes and key enzymes for melanin formation inmelanocytes. TRP-1 possesses 5,6-dihydroxyindole-2-carboxylicacid (DHICA)-oxidase activity [31,32]. TRP-2 converts dopa-chrome to DHICA [33], and is a marker of both melanoblasts andmelanocytes [34–36]. C-kit is the tyrosine kinase receptor, andits ligand is stem cell factor (steel factor, KIT-ligand, or mast cellgrowth factor). C-kit plays an important role in melanocytedevelopment, especially in the proliferation of early melano-blasts, and c-kit antibody is used for detecting melanoblasts[37,38]. At present, there is no marker able to strictly distinguishmelanocytes and melanoblasts. However, studies of skindevelopment and hair growth cycles suggest that these threeproteins are useful for detecting undifferentiated melanoblastsin mammalian skin.
In this study, we used light and fluorescence microscopy toquantify changes in the numbers of differentiated melanocytescontaining melanin pigments (MM) and melanoblasts/melano-cytes immunohistochemically positive for tyrosinase-relatedprotein (TRP)-1, TRP-2 (dopachrome tautomerase), and c-kit inepidermis during the development of pigmented spots in hairlessmice chronically exposed to UVB.
2. Materials and methods
2.1. Mice
Hos:(HR-1 � HR/De)F1 hairless mice were used in this study.They were given water and a commercial diet (CRF-1, OrientalYeast Co., Tokyo, Japan) ad libitum, and maintained at 24 � 1 8Cwith 40–60% relative humidity under 12 h on/12 h off fluorescentlighting. The present study was conducted in accordance with the
Fig. 1. The development of pigmented spots in the dorsal skin of (HR-1 � HR/De)F1 mice a
(UV irradiated; A–D, non-irradiated; J and K) and skin stained with DOPA (UV irradiated;
hyperpigmentation (A and E) and of age-matched untreated control of 11-week-old mice
the irradiated dorsal skin and dendritic DOPA-reactive melanocytes were seen except at
stage of depigmentation). The skin color was restored to the control level and there were
pigmented spots). (H) 20 weeks (the stage of development of small pigmented spots). (D
the cessation of UVB exposure. Pigmented spots were observed on the dorsal skin of irrad
pigmented spots gradually increased. (K) Non-irradiated 52-week-old mice (control of
guidelines of the National Institutes of Health, and was approved bythe Ethics Committee of Shiseido Research Center.
2.2. UV-irradiation and sampling time
Seven-week-old mice were exposed to 99 mJ/cm2 UVB from FL-20 SR fluorescent lamps 3 times a week for 8 weeks. Forimmunohistochemical experiments, epidermal sheets were pre-pared from dorsal skin of mice after two UVB exposures (the earlystage of UV irradiation), after UVB exposure 3 times per week for 4weeks (the acute hyperpigmentation stage), and at 3 (depigmen-tation stage), 8 (appearance of pigmented spots), 20 (developmentof small pigmented spots), and 37 (development of medium-sizedpigmented spots) weeks after the cessation of the 8-week UVBexposure schedule (Fig. 1). Untreated control samples were alsoobtained from non-irradiated mice at the corresponding times.
2.3. DOPA reaction of whole-mount specimens
Dorsal skin samples of Hos:(HR-1 � HR/De)F1 at various stagesafter UVB irradiation were cleaned of subcutaneous tissues. Theywere cut into small pieces (5 mm� 5 mm) and put into 70% ethanolsolution for 10 min, then absolute ethanol for 1 h, and finally intoxylene. The samples became transparent in xylene after more than30 min and were mounted on slide glasses with Canada balsam(Wako Pure Chemical Industries, Osaka, Japan). In the case of whole-mount DOPA (3,4-dihydroxyphenylalanine, Wako, Osaka, Japan)reaction, small skin pieces were fixed with buffered formalinsolution (pH 7.0) for 16–20 h at 4 8C. They were then rinsed indistilled water and preincubated with buffered 0.1% L-DOPA solution(pH 7.4) for 30 min at 37 8C. Then, they were incubated with fresh0.1% L-DOPA solution at 37 8C. After 18–24 h, the specimens werewashed with distilled water and put into 70% ethanol solution.Dehydration and mounting were performed as described above.
2.4. Preparation of epidermal sheets
Dorsal skin samples of Hos:(HR-1 � HR/De)F1 at various stagesafter UVB irradiation were cleaned of subcutaneous tissues, and
fter UVB irradiation. The sampling time (blue arrows) is also shown. Skin appearance
E–I) are shown. Skin 4 weeks after the initiation of UV irradiation (the stage of acute
(J) are shown. An even distribution of increased pigmentation was observed all over
hair follicles. (B and F) 3 weeks after the cessation of UVB exposure for 8 weeks (the
few melanocytes in the epidermis (F). (C and G) 8 weeks (the stage of appearance of
and I) 37 weeks (the stage of development of medium-sized pigmented spots) after
iated mice 8 weeks after the cessation of UVB exposure and the number and size of
D).

Fig. 2. Image processing of epidermal sheets. (A) Areas of sebaceous gland were
outlined (red), and excluded from effective skin area. (B and C) Areas of
keratinocytes containing pigments were outlined (red) and designated as
pigmented areas under the light field. Scale bar, 100 mm.
R. Furuya et al. / Journal of Dermatological Science 55 (2009) 99–107 101
the stratum corneum side of the samples was covered withcellophane adhesive tape. The samples were cut into small pieces(4 mm � 4 mm) and incubated in 2 N sodium bromide (NaBr) at37 8C. After 0.5 h, the epidermal sheets were mechanicallyseparated from the dermis with fine forceps.
2.5. Immunohistochemical staining
Epidermal sheets were fixed with acetone at 4 8C for 30 min,and rinsed in calcium-, magnesium-free phosphate-buffered saline(PBS(�), Wako, Osaka, Japan). The samples were blocked withnormal serum (HISTOFINER, Nichirei, Tokyo, Japan) for 2 h andincubated with primary antibodies overnight at 4 8C. The primaryantibodies used in this study were anti-TRP-1 (Pep1, gift from Dr.Tomita), anti-TRP-2 (Pep8, gift from Dr. Hearing), and anti-c-kit(ACK45, BD Pharmingen, NJ, USA). Normal rat IgG (IBL, Gunma,Japan) or rabbit IgG (Vector Laboratories, CA, USA) was used asnegative controls. Epidermal sheets were washed thoroughly withPBS(�) containing 0.05% Tween 20, then incubated with biotin-conjugated rat or mouse anti-IgG (Vector Laboratories) at roomtemperature for 1 h. They were re-washed thoroughly with PBS(�)containing 0.05% Tween 20, equilibrated with 0.1 M bicarbonate-buffered solution (pH 8.2) for 10 min, then incubated withFluorescein Avidin DCS (Vector Laboratories) at room temperaturefor 0.5 h in the dark.
2.6. Measurement of pigmented/non-pigmented areas and the
numbers of pigmented melanocytes and immunohistochemically
positive cells in the epidermis
We used a light and fluorescence microscope (BX50, Olympus,Tokyo, Japan) with a digital camera system (FUJIX Photograb-2500R SH-25/M, FUJIFILM, Tokyo, Japan) to acquire images, whichwere analyzed with WinroofR (Mitani Corporation, Fukui, Japan).Eight randomly selected areas of each specimen were analyzed.The statistical significance of differences was determined by usingStudent’s non-paired t-test for comparisons of groups of equal size.
2.6.1. Effective area
Since sebaceous glands do not contain melanocytes orimmunohistochemically positive cells, areas of sebaceous glandwere excluded. The total area excluding that of sebaceous glandswas taken as the effective area (Fig. 2A).
2.6.2. Pigmented/non-pigmented area
Pigmented areas contained keratinocytes were designated aspigmented areas (Fig. 2B and C), and the total effective area minuspigmented areas was taken as non-pigmented areas. The ratio ofpigmented/non-pigmented areas (percent) was calculated.
2.6.3. Number of pigmented melanocytes and
immunohistochemically positive cells in the epidermis
The number of differentiated melanocytes contained pigmentswas counted under a light microscope. The numbers of cellspositive to immunohistochemical staining for each of the markerswas counted under a fluorescence microscope. The number ofdifferentiated melanocytes containing pigments per unit area wascalculated based on the effective area.
3. Results
Whole mounts (Fig. 1) and epidermal sheets (Fig. 2) wereprepared from dorsal skin of mice at various stages during thedevelopment of pigmented spots after UVB irradiation (Fig. 1). Thepigmentation pattern could be visualized using epidermal sheets,and the number and distribution of melanocytes as well as
pigments produced could be observed through the skin surface(Fig. 2). Differentiated melanocytes containing melanin pigment(MM) and cells positive to TRP-1, TRP-2, and c-kit were countedand their numbers per 1 mm2 area were calculated (Figs. 3–7,Table 1).
In non-irradiated control skins, very few MM were present from7 to 35 weeks (Fig. 3). Although almost no pigments were observedduring this period, the numbers of cells positive for TRP-1, TRP-2,and c-kit were constant and 3–10-fold greater than that of MM
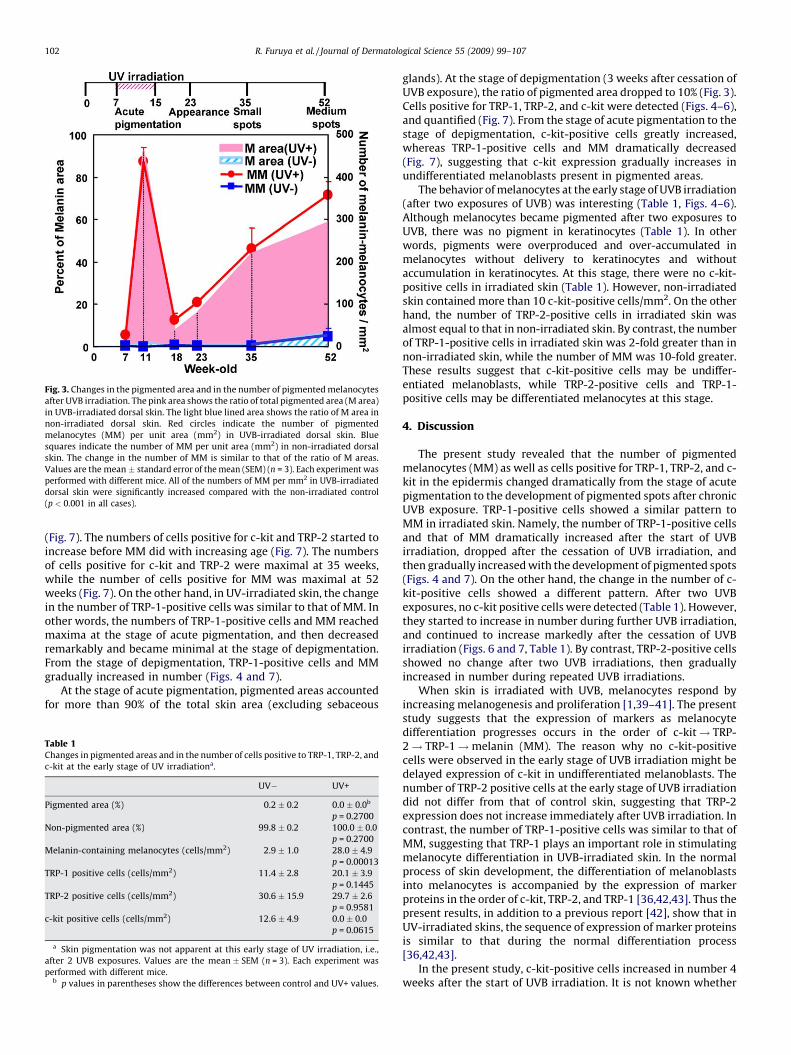
Fig. 3. Changes in the pigmented area and in the number of pigmented melanocytes
after UVB irradiation. The pink area shows the ratio of total pigmented area (M area)
in UVB-irradiated dorsal skin. The light blue lined area shows the ratio of M area in
non-irradiated dorsal skin. Red circles indicate the number of pigmented
melanocytes (MM) per unit area (mm2) in UVB-irradiated dorsal skin. Blue
squares indicate the number of MM per unit area (mm2) in non-irradiated dorsal
skin. The change in the number of MM is similar to that of the ratio of M areas.
Values are the mean � standard error of the mean (SEM) (n = 3). Each experiment was
performed with different mice. All of the numbers of MM per mm2 in UVB-irradiated
dorsal skin were significantly increased compared with the non-irradiated control
(p < 0.001 in all cases).
R. Furuya et al. / Journal of Dermatological Science 55 (2009) 99–107102
(Fig. 7). The numbers of cells positive for c-kit and TRP-2 started toincrease before MM did with increasing age (Fig. 7). The numbersof cells positive for c-kit and TRP-2 were maximal at 35 weeks,while the number of cells positive for MM was maximal at 52weeks (Fig. 7). On the other hand, in UV-irradiated skin, the changein the number of TRP-1-positive cells was similar to that of MM. Inother words, the numbers of TRP-1-positive cells and MM reachedmaxima at the stage of acute pigmentation, and then decreasedremarkably and became minimal at the stage of depigmentation.From the stage of depigmentation, TRP-1-positive cells and MMgradually increased in number (Figs. 4 and 7).
At the stage of acute pigmentation, pigmented areas accountedfor more than 90% of the total skin area (excluding sebaceous
Table 1Changes in pigmented areas and in the number of cells positive to TRP-1, TRP-2, and
c-kit at the early stage of UV irradiationa.
UV� UV+
Pigmented area (%) 0.2 � 0.2 0.0 � 0.0b
p = 0.2700
Non-pigmented area (%) 99.8 � 0.2 100.0 � 0.0
p = 0.2700
Melanin-containing melanocytes (cells/mm2) 2.9 � 1.0 28.0 � 4.9
p = 0.00013
TRP-1 positive cells (cells/mm2) 11.4 � 2.8 20.1 � 3.9
p = 0.1445
TRP-2 positive cells (cells/mm2) 30.6 � 15.9 29.7 � 2.6
p = 0.9581
c-kit positive cells (cells/mm2) 12.6 � 4.9 0.0 � 0.0
p = 0.0615
a Skin pigmentation was not apparent at this early stage of UV irradiation, i.e.,
after 2 UVB exposures. Values are the mean � SEM (n = 3). Each experiment was
performed with different mice.b p values in parentheses show the differences between control and UV+ values.
glands). At the stage of depigmentation (3 weeks after cessation ofUVB exposure), the ratio of pigmented area dropped to 10% (Fig. 3).Cells positive for TRP-1, TRP-2, and c-kit were detected (Figs. 4–6),and quantified (Fig. 7). From the stage of acute pigmentation to thestage of depigmentation, c-kit-positive cells greatly increased,whereas TRP-1-positive cells and MM dramatically decreased(Fig. 7), suggesting that c-kit expression gradually increases inundifferentiated melanoblasts present in pigmented areas.
The behavior of melanocytes at the early stage of UVB irradiation(after two exposures of UVB) was interesting (Table 1, Figs. 4–6).Although melanocytes became pigmented after two exposures toUVB, there was no pigment in keratinocytes (Table 1). In otherwords, pigments were overproduced and over-accumulated inmelanocytes without delivery to keratinocytes and withoutaccumulation in keratinocytes. At this stage, there were no c-kit-positive cells in irradiated skin (Table 1). However, non-irradiatedskin contained more than 10 c-kit-positive cells/mm2. On the otherhand, the number of TRP-2-positive cells in irradiated skin wasalmost equal to that in non-irradiated skin. By contrast, the numberof TRP-1-positive cells in irradiated skin was 2-fold greater than innon-irradiated skin, while the number of MM was 10-fold greater.These results suggest that c-kit-positive cells may be undiffer-entiated melanoblasts, while TRP-2-positive cells and TRP-1-positive cells may be differentiated melanocytes at this stage.
4. Discussion
The present study revealed that the number of pigmentedmelanocytes (MM) as well as cells positive for TRP-1, TRP-2, and c-kit in the epidermis changed dramatically from the stage of acutepigmentation to the development of pigmented spots after chronicUVB exposure. TRP-1-positive cells showed a similar pattern toMM in irradiated skin. Namely, the number of TRP-1-positive cellsand that of MM dramatically increased after the start of UVBirradiation, dropped after the cessation of UVB irradiation, andthen gradually increased with the development of pigmented spots(Figs. 4 and 7). On the other hand, the change in the number of c-kit-positive cells showed a different pattern. After two UVBexposures, no c-kit positive cells were detected (Table 1). However,they started to increase in number during further UVB irradiation,and continued to increase markedly after the cessation of UVBirradiation (Figs. 6 and 7, Table 1). By contrast, TRP-2-positive cellsshowed no change after two UVB irradiations, then graduallyincreased in number during repeated UVB irradiations.
When skin is irradiated with UVB, melanocytes respond byincreasing melanogenesis and proliferation [1,39–41]. The presentstudy suggests that the expression of markers as melanocytedifferentiation progresses occurs in the order of c-kit! TRP-2! TRP-1!melanin (MM). The reason why no c-kit-positivecells were observed in the early stage of UVB irradiation might bedelayed expression of c-kit in undifferentiated melanoblasts. Thenumber of TRP-2 positive cells at the early stage of UVB irradiationdid not differ from that of control skin, suggesting that TRP-2expression does not increase immediately after UVB irradiation. Incontrast, the number of TRP-1-positive cells was similar to that ofMM, suggesting that TRP-1 plays an important role in stimulatingmelanocyte differentiation in UVB-irradiated skin. In the normalprocess of skin development, the differentiation of melanoblastsinto melanocytes is accompanied by the expression of markerproteins in the order of c-kit, TRP-2, and TRP-1 [36,42,43]. Thus thepresent results, in addition to a previous report [42], show that inUV-irradiated skins, the sequence of expression of marker proteinsis similar to that during the normal differentiation process[36,42,43].
In the present study, c-kit-positive cells increased in number 4weeks after the start of UVB irradiation. It is not known whether
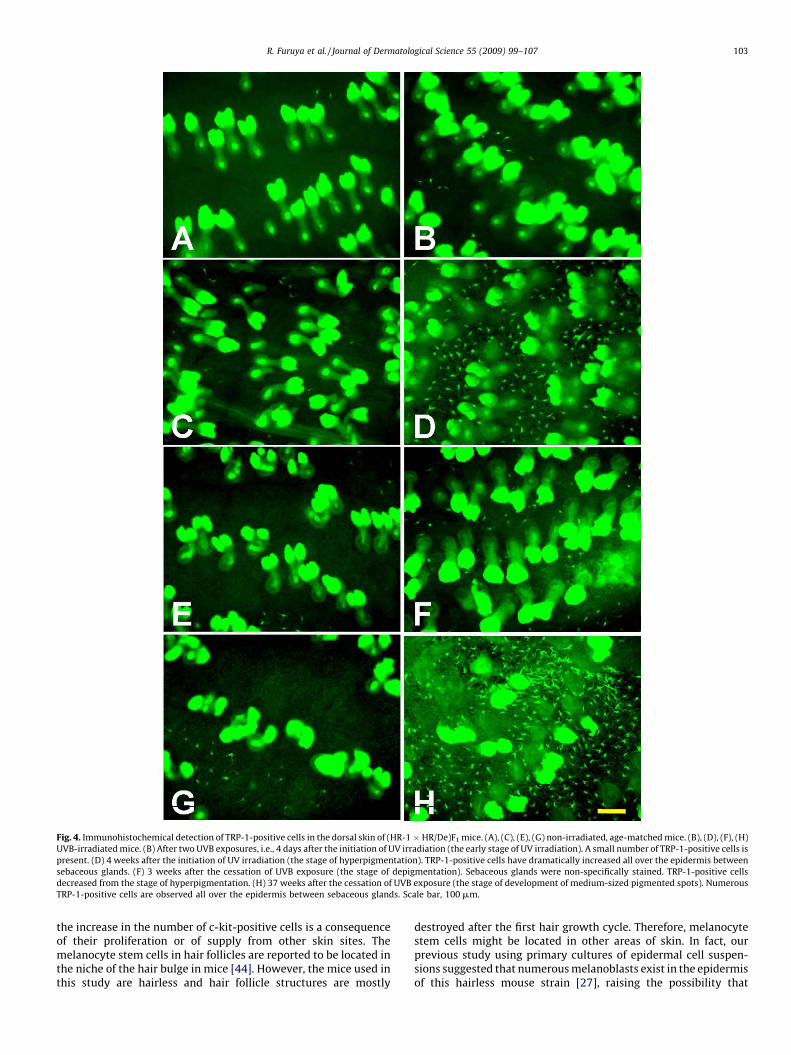
Fig. 4. Immunohistochemical detection of TRP-1-positive cells in the dorsal skin of (HR-1 � HR/De)F1 mice. (A), (C), (E), (G) non-irradiated, age-matched mice. (B), (D), (F), (H)
UVB-irradiated mice. (B) After two UVB exposures, i.e., 4 days after the initiation of UV irradiation (the early stage of UV irradiation). A small number of TRP-1-positive cells is
present. (D) 4 weeks after the initiation of UV irradiation (the stage of hyperpigmentation). TRP-1-positive cells have dramatically increased all over the epidermis between
sebaceous glands. (F) 3 weeks after the cessation of UVB exposure (the stage of depigmentation). Sebaceous glands were non-specifically stained. TRP-1-positive cells
decreased from the stage of hyperpigmentation. (H) 37 weeks after the cessation of UVB exposure (the stage of development of medium-sized pigmented spots). Numerous
TRP-1-positive cells are observed all over the epidermis between sebaceous glands. Scale bar, 100 mm.
R. Furuya et al. / Journal of Dermatological Science 55 (2009) 99–107 103
the increase in the number of c-kit-positive cells is a consequenceof their proliferation or of supply from other skin sites. Themelanocyte stem cells in hair follicles are reported to be located inthe niche of the hair bulge in mice [44]. However, the mice used inthis study are hairless and hair follicle structures are mostly
destroyed after the first hair growth cycle. Therefore, melanocytestem cells might be located in other areas of skin. In fact, ourprevious study using primary cultures of epidermal cell suspen-sions suggested that numerous melanoblasts exist in the epidermisof this hairless mouse strain [27], raising the possibility that
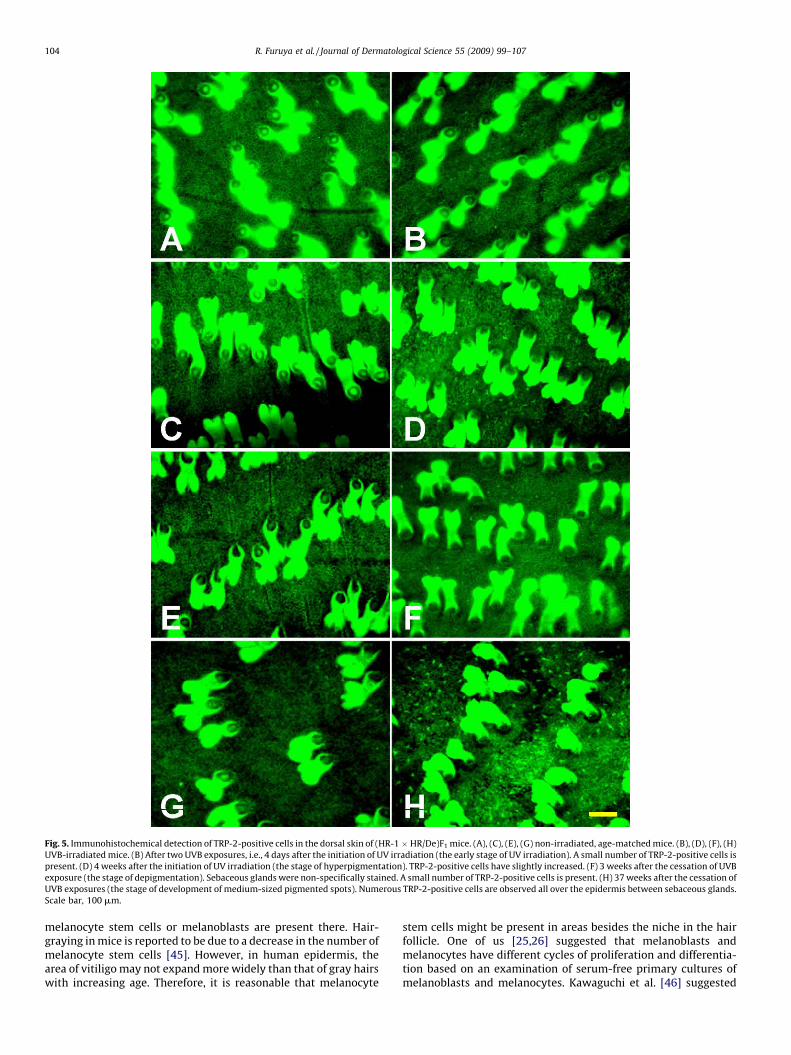
Fig. 5. Immunohistochemical detection of TRP-2-positive cells in the dorsal skin of (HR-1 � HR/De)F1 mice. (A), (C), (E), (G) non-irradiated, age-matched mice. (B), (D), (F), (H)
UVB-irradiated mice. (B) After two UVB exposures, i.e., 4 days after the initiation of UV irradiation (the early stage of UV irradiation). A small number of TRP-2-positive cells is
present. (D) 4 weeks after the initiation of UV irradiation (the stage of hyperpigmentation). TRP-2-positive cells have slightly increased. (F) 3 weeks after the cessation of UVB
exposure (the stage of depigmentation). Sebaceous glands were non-specifically stained. A small number of TRP-2-positive cells is present. (H) 37 weeks after the cessation of
UVB exposures (the stage of development of medium-sized pigmented spots). Numerous TRP-2-positive cells are observed all over the epidermis between sebaceous glands.
Scale bar, 100 mm.
R. Furuya et al. / Journal of Dermatological Science 55 (2009) 99–107104
melanocyte stem cells or melanoblasts are present there. Hair-graying in mice is reported to be due to a decrease in the number ofmelanocyte stem cells [45]. However, in human epidermis, thearea of vitiligo may not expand more widely than that of gray hairswith increasing age. Therefore, it is reasonable that melanocyte
stem cells might be present in areas besides the niche in the hairfollicle. One of us [25,26] suggested that melanoblasts andmelanocytes have different cycles of proliferation and differentia-tion based on an examination of serum-free primary cultures ofmelanoblasts and melanocytes. Kawaguchi et al. [46] suggested
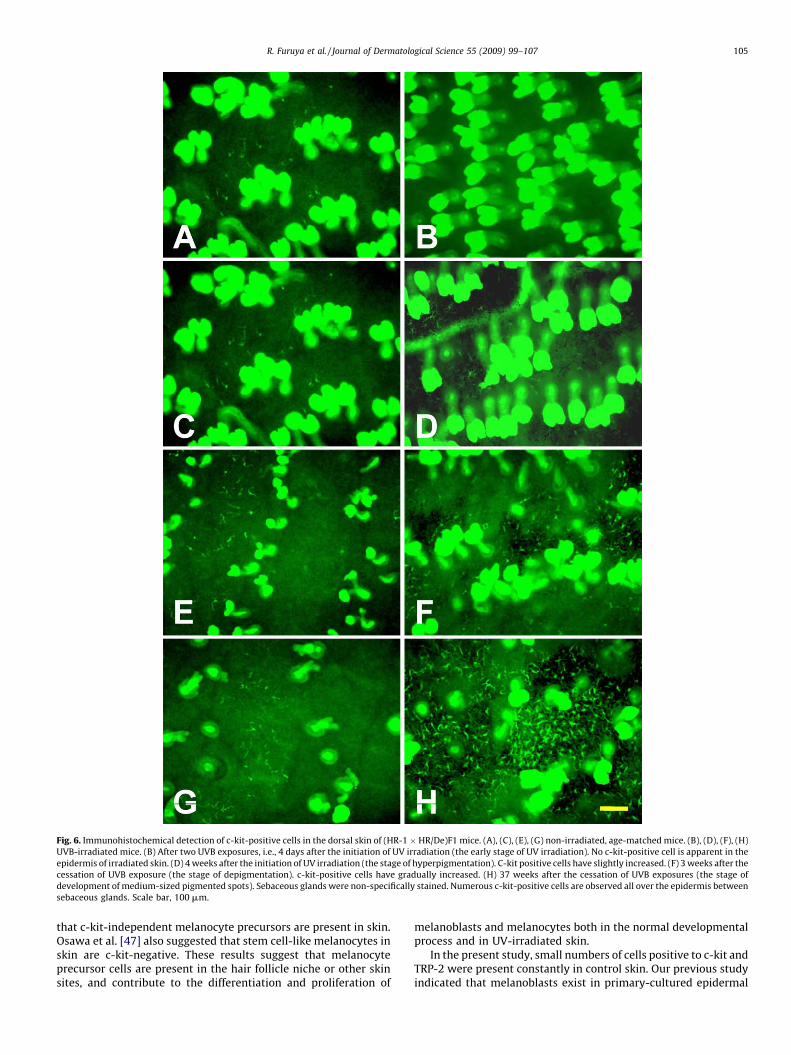
Fig. 6. Immunohistochemical detection of c-kit-positive cells in the dorsal skin of (HR-1 � HR/De)F1 mice. (A), (C), (E), (G) non-irradiated, age-matched mice. (B), (D), (F), (H)
UVB-irradiated mice. (B) After two UVB exposures, i.e., 4 days after the initiation of UV irradiation (the early stage of UV irradiation). No c-kit-positive cell is apparent in the
epidermis of irradiated skin. (D) 4 weeks after the initiation of UV irradiation (the stage of hyperpigmentation). C-kit positive cells have slightly increased. (F) 3 weeks after the
cessation of UVB exposure (the stage of depigmentation). c-kit-positive cells have gradually increased. (H) 37 weeks after the cessation of UVB exposures (the stage of
development of medium-sized pigmented spots). Sebaceous glands were non-specifically stained. Numerous c-kit-positive cells are observed all over the epidermis between
sebaceous glands. Scale bar, 100 mm.
R. Furuya et al. / Journal of Dermatological Science 55 (2009) 99–107 105
that c-kit-independent melanocyte precursors are present in skin.Osawa et al. [47] also suggested that stem cell-like melanocytes inskin are c-kit-negative. These results suggest that melanocyteprecursor cells are present in the hair follicle niche or other skinsites, and contribute to the differentiation and proliferation of
melanoblasts and melanocytes both in the normal developmentalprocess and in UV-irradiated skin.
In the present study, small numbers of cells positive to c-kit andTRP-2 were present constantly in control skin. Our previous studyindicated that melanoblasts exist in primary-cultured epidermal
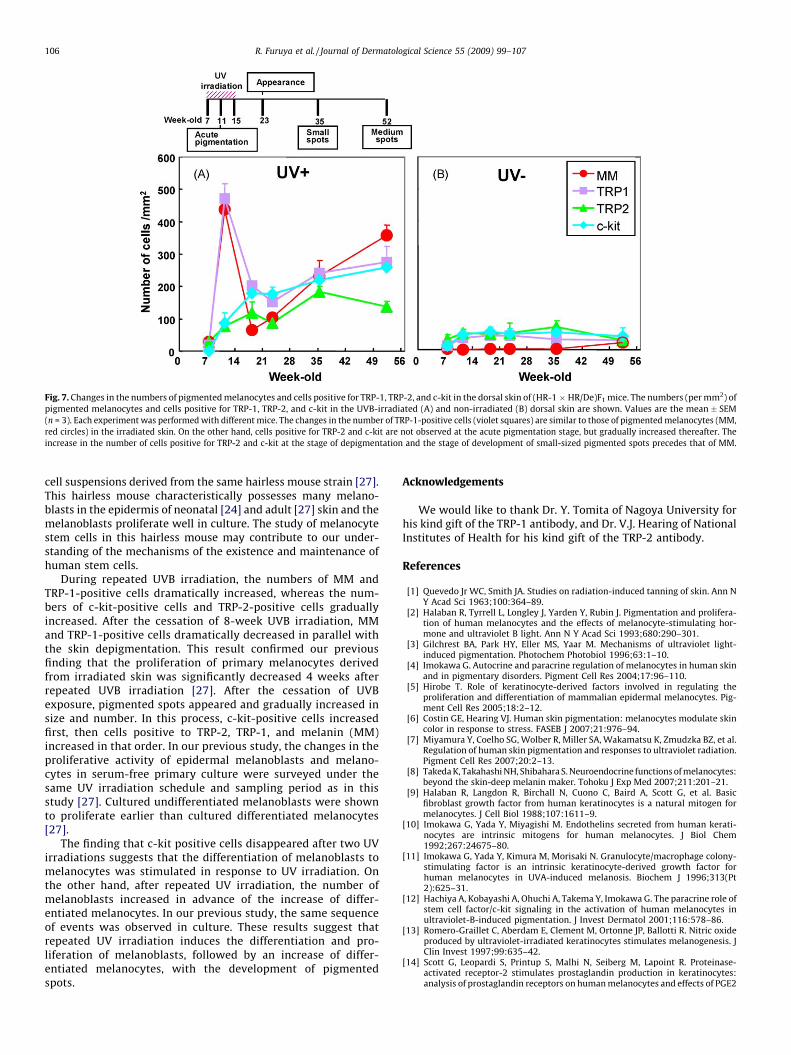
Fig. 7. Changes in the numbers of pigmented melanocytes and cells positive for TRP-1, TRP-2, and c-kit in the dorsal skin of (HR-1 � HR/De)F1 mice. The numbers (per mm2) of
pigmented melanocytes and cells positive for TRP-1, TRP-2, and c-kit in the UVB-irradiated (A) and non-irradiated (B) dorsal skin are shown. Values are the mean � SEM
(n = 3). Each experiment was performed with different mice. The changes in the number of TRP-1-positive cells (violet squares) are similar to those of pigmented melanocytes (MM,
red circles) in the irradiated skin. On the other hand, cells positive for TRP-2 and c-kit are not observed at the acute pigmentation stage, but gradually increased thereafter. The
increase in the number of cells positive for TRP-2 and c-kit at the stage of depigmentation and the stage of development of small-sized pigmented spots precedes that of MM.
R. Furuya et al. / Journal of Dermatological Science 55 (2009) 99–107106
cell suspensions derived from the same hairless mouse strain [27].This hairless mouse characteristically possesses many melano-blasts in the epidermis of neonatal [24] and adult [27] skin and themelanoblasts proliferate well in culture. The study of melanocytestem cells in this hairless mouse may contribute to our under-standing of the mechanisms of the existence and maintenance ofhuman stem cells.
During repeated UVB irradiation, the numbers of MM andTRP-1-positive cells dramatically increased, whereas the num-bers of c-kit-positive cells and TRP-2-positive cells graduallyincreased. After the cessation of 8-week UVB irradiation, MMand TRP-1-positive cells dramatically decreased in parallel withthe skin depigmentation. This result confirmed our previousfinding that the proliferation of primary melanocytes derivedfrom irradiated skin was significantly decreased 4 weeks afterrepeated UVB irradiation [27]. After the cessation of UVBexposure, pigmented spots appeared and gradually increased insize and number. In this process, c-kit-positive cells increasedfirst, then cells positive to TRP-2, TRP-1, and melanin (MM)increased in that order. In our previous study, the changes in theproliferative activity of epidermal melanoblasts and melano-cytes in serum-free primary culture were surveyed under thesame UV irradiation schedule and sampling period as in thisstudy [27]. Cultured undifferentiated melanoblasts were shownto proliferate earlier than cultured differentiated melanocytes[27].
The finding that c-kit positive cells disappeared after two UVirradiations suggests that the differentiation of melanoblasts tomelanocytes was stimulated in response to UV irradiation. Onthe other hand, after repeated UV irradiation, the number ofmelanoblasts increased in advance of the increase of differ-entiated melanocytes. In our previous study, the same sequenceof events was observed in culture. These results suggest thatrepeated UV irradiation induces the differentiation and pro-liferation of melanoblasts, followed by an increase of differ-entiated melanocytes, with the development of pigmentedspots.
Acknowledgements
We would like to thank Dr. Y. Tomita of Nagoya University forhis kind gift of the TRP-1 antibody, and Dr. V.J. Hearing of NationalInstitutes of Health for his kind gift of the TRP-2 antibody.
References
[1] Quevedo Jr WC, Smith JA. Studies on radiation-induced tanning of skin. Ann NY Acad Sci 1963;100:364–89.
[2] Halaban R, Tyrrell L, Longley J, Yarden Y, Rubin J. Pigmentation and prolifera-tion of human melanocytes and the effects of melanocyte-stimulating hor-mone and ultraviolet B light. Ann N Y Acad Sci 1993;680:290–301.
[3] Gilchrest BA, Park HY, Eller MS, Yaar M. Mechanisms of ultraviolet light-induced pigmentation. Photochem Photobiol 1996;63:1–10.
[4] Imokawa G. Autocrine and paracrine regulation of melanocytes in human skinand in pigmentary disorders. Pigment Cell Res 2004;17:96–110.
[5] Hirobe T. Role of keratinocyte-derived factors involved in regulating theproliferation and differentiation of mammalian epidermal melanocytes. Pig-ment Cell Res 2005;18:2–12.
[6] Costin GE, Hearing VJ. Human skin pigmentation: melanocytes modulate skincolor in response to stress. FASEB J 2007;21:976–94.
[7] Miyamura Y, Coelho SG, Wolber R, Miller SA, Wakamatsu K, Zmudzka BZ, et al.Regulation of human skin pigmentation and responses to ultraviolet radiation.Pigment Cell Res 2007;20:2–13.
[8] Takeda K, Takahashi NH, Shibahara S. Neuroendocrine functions of melanocytes:beyond the skin-deep melanin maker. Tohoku J Exp Med 2007;211:201–21.
[9] Halaban R, Langdon R, Birchall N, Cuono C, Baird A, Scott G, et al. Basicfibroblast growth factor from human keratinocytes is a natural mitogen formelanocytes. J Cell Biol 1988;107:1611–9.
[10] Imokawa G, Yada Y, Miyagishi M. Endothelins secreted from human kerati-nocytes are intrinsic mitogens for human melanocytes. J Biol Chem1992;267:24675–80.
[11] Imokawa G, Yada Y, Kimura M, Morisaki N. Granulocyte/macrophage colony-stimulating factor is an intrinsic keratinocyte-derived growth factor forhuman melanocytes in UVA-induced melanosis. Biochem J 1996;313(Pt2):625–31.
[12] Hachiya A, Kobayashi A, Ohuchi A, Takema Y, Imokawa G. The paracrine role ofstem cell factor/c-kit signaling in the activation of human melanocytes inultraviolet-B-induced pigmentation. J Invest Dermatol 2001;116:578–86.
[13] Romero-Graillet C, Aberdam E, Clement M, Ortonne JP, Ballotti R. Nitric oxideproduced by ultraviolet-irradiated keratinocytes stimulates melanogenesis. JClin Invest 1997;99:635–42.
[14] Scott G, Leopardi S, Printup S, Malhi N, Seiberg M, Lapoint R. Proteinase-activated receptor-2 stimulates prostaglandin production in keratinocytes:analysis of prostaglandin receptors on human melanocytes and effects of PGE2

R. Furuya et al. / Journal of Dermatological Science 55 (2009) 99–107 107
and PGF2alpha on melanocyte dendricity. J Invest Dermatol 2004;122:1214–24.
[15] Hodgson C. Senile lentigo. Arch Dermatol 1963;87:197–207.[16] Andersen WK, Labadie RR, Bhawan J. Histopathology of solar lentigines of the
face: a quantitative study. J Am Acad Dermatol 1997;36:444–7.[17] Cario-Andre M, Lepreux S, Pain C, Nizard C, Noblesse E, Taieb A. Perilesional vs.
lesional skin changes in senile lentigo. J Cutan Pathol 2004;31:441–7.[18] Noblesse E, Nizard C, Cario-Andre M, Lepreux S, Pain C, Schnebert S, et al. Skin
ultrastructure in senile lentigo. Skin Pharmacol Physiol 2006;19:95–100.[19] Aoki H, Moro O, Tagami H, Kishimoto J. Gene expression profiling analysis of
solar lentigo in relation to immunohistochemical characteristics. Br J Dermatol2007;156:1214–23.
[20] Gilchrest BA. Relationship between actinic damage and chronologic aging inkeratinocyte cultures of human skin. J Invest Dermatol 1979;72:219–23.
[21] Ortonne JP. Pigmentary changes of the ageing skin. Br J Dermatol1990;122(Suppl. 35):21–8.
[22] Yaar M, Gilchrest BA. Ageing and photoageing of keratinocytes and melano-cytes. Clin Exp Dermatol 2001;26:583–91.
[23] Naganumaa M, Yagi E, Fukuda M. Delayed induction of pigmented spots onUVB-irradiated hairless mice. J Dermatol Sci 2001;25:29–35.
[24] Furuya R, Akiu S, Naganuma M, Fukuda M, Hirobe T. The proliferation anddifferentiation of neonatal epidermal melanocytes in F1 hairless mice of HR-1 � HR/De in serum-free culture. J Dermatol 1998;25:211–21.
[25] Hirobe T. Melanocyte stimulating hormone induces the differentiation ofmouse epidermal melanocytes in serum-free culture. J Cell Physiol1992;152:337–45.
[26] Hirobe T. Basic fibroblast growth factor stimulates the sustained proliferationof mouse epidermal melanoblasts in a serum-free medium in the presence ofdibutyryl cyclic AMP and keratinocytes. Development 1992;114:435–45.
[27] Furuya R, Akiu S, Ideta R, Naganuma M, Fukuda M, Hirobe T. Changes in theproliferative activity of epidermal melanocytes in serum-free primary cultureduring the development of ultraviolet radiation B-induced pigmented spots inhairless mice. Pigment Cell Res 2002;15:348–56.
[28] Hirobe T, Furuya R, Akiu S, Ifuku O, Fukuda M. Keratinocytes control theproliferation and differentiation of cultured epidermal melanocytes fromultraviolet radiation B-induced pigmented spots in the dorsal skin of hairlessmice. Pigment Cell Res 2002;15:391–9.
[29] Hirobe T, Furuya R, Hara E, Horii I, Tsunenaga M, Ifuku O. Granulocyte-macrophage colony-stimulating factor (GM-CSF) controls the proliferationand differentiation of mouse epidermal melanocytes from pigmented spotsinduced by ultraviolet radiation B. Pigment Cell Res 2004;17:230–40.
[30] Hirobe T, Furuya R, Ifuku O, Osawa M, Nishikawa S. Granulocyte-macrophagecolony-stimulating factor is a keratinocyte-derived factor involved in regulat-ing the proliferation and differentiation of neonatal mouse epidermal mela-nocytes in culture. Exp Cell Res 2004;297:593–606.
[31] Jimenez-Cervantes C, Solano F, Kobayashi T, Urabe K, Hearing VJ, Lozano JA,et al. A new enzymatic function in the melanogenic pathway. The 5,6-dihy-
droxyindole-2-carboxylic acid oxidase activity of tyrosinase-related protein-1(TRP1). J Biol Chem 1994;269:17993–8000.
[32] Kobayashi T, Urabe K, Winder A, Jimenez-Cervantes C, Imokawa G, BrewingtonT, et al. Tyrosinase related protein 1 (TRP1) functions as a DHICA oxidase inmelanin biosynthesis. EMBO J 1994;13:5818–25.
[33] Tsukamoto K, Jackson IJ, Urabe K, Montague PM, Hearing VJ. A secondtyrosinase-related protein, TRP-2, is a melanogenic enzyme termed DOPA-chrome tautomerase. EMBO J 1992;11:519–26.
[34] Tai T, Eisinger M, Ogata S, Lloyd KO. Glycoproteins as differentiation markersin human malignant melanoma and melanocytes. Cancer Res 1983;43:2773–9.
[35] Vijayasaradhi S, Houghton AN. Purification of an autoantigenic 75-kDa humanmelanosomal glycoprotein. Int J Cancer 1991;47:298–303.
[36] Steel KP, Davidson DR, Jackson IJ. TRP-2/DT, a new early melanoblast marker,shows that steel growth factor (c-kit ligand) is a survival factor. Development1992;115:1111–9.
[37] Grichnik JM, Ali WN, Burch JA, Byers JD, Garcia CA, Clark RE, et al. KITexpression reveals a population of precursor melanocytes in human skin. JInvest Dermatol 1996;106:967–71.
[38] Nishikawa S, Kusakabe M, Yoshinaga K, Ogawa M, Hayashi S, Kunisada T, et al.In utero manipulation of coat color formation by a monoclonal anti-c-kitantibody: two distinct waves of c-kit-dependency during melanocyte devel-opment. EMBO J 1991;10:2111–8.
[39] Gilchrest BA, Blog FB, Szabo G. Effects of aging and chronic sun exposure onmelanocytes in human skin. J Invest Dermatol 1979;73:141–3.
[40] Quevedo WC, Szabo G, Virks J. Influence of age and UV on the populations ofdopa-positive melanocytes in human skin. J Invest Dermatol 1969;52:287–90.
[41] Quevedo Jr WC, Szabo G, Virks J, Sinesi SJ. Melanocyte populations in UV-irradiated human skin. J Invest Dermatol 1965;45:295–8.
[42] Kawaguchi Y, Mori N, Nakayama A. Kit(+) melanocytes seem to contribute tomelanocyte proliferation after UV exposure as precursor cells. J Invest Der-matol 2001;116:920–5.
[43] Wehrle-Haller B, Weston JA. Soluble and cell-bound forms of steel factoractivity play distinct roles in melanocyte precursor dispersal and survival onthe lateral neural crest migration pathway. Development 1995;121:731–42.
[44] Nishimura EK, Jordan SA, Oshima H, Yoshida H, Osawa M, Moriyama M, et al.Dominant role of the niche in melanocyte stem-cell fate determination. Nature2002;416:854–60.
[45] Nishimura EK, Granter SR, Fisher DE. Mechanisms of hair graying: incompletemelanocyte stem cell maintenance in the niche. Science 2005;307:720–4.
[46] Kawaguchi A, Chiba K, Tanimura Y, Motohashi T, Aoki H, Takeda T, et al.Isolation and characterization of Kit-independent melanocyte precursorsinduced in the skin of Steel factor transgenic mice. Dev Growth Differ2008;50:63–9.
[47] Osawa M, Egawa G, Mak SS, Moriyama M, Freter R, Yonetani S, et al. Molecularcharacterization of melanocyte stem cells in their niche. Development2005;132:5589–99.














![Toronto SCC epigenetics and aginginteresting skin lighteners on melanocytes looking atinteresting skin lighteners on melanocytes looking at Tyrosinase [TYR] and Ferritin [FTH1] gene](https://img.pdfslide.us/doc/110x75/602d4f8f53f48f1d883bdfdb/toronto-scc-epigenetics-and-aging-interesting-skin-lighteners-on-melanocytes-looking.jpg)














