Embed Size (px)
Citation preview

J. Cell Sci. 3 3 ) 371-383 (1978) 371Printed in Great Britain © Company of Biologists Limited
STIMULATION OF DENDRITOGENESIS IN THE
EPIDERMAL MELANOCYTES OF NEWBORN
MICE BY MELANOCYTE-STIMULATING
HORMONE
TOMOHISA HIROBE
Department of Biology, Faculty of Education,luate University, Ueda, Morioka, 020 Japan
SUMMARY
The number of dendrites and the total length of dendrites in the epidermal melanocytespositive for the dopa reaction were shown to increase when newborn mice were injected witha-MSH or DBc-AMP. Moreover, both indices of the degree of dendritogenesis increasedwhen skin explants of newborn mice were cultured in medium containing a-MSH or DBc-AMP. Electron-microscopic observation showed that the number of melanosomes was alsoincreased by a-MSH treatment. Many mature melanosomes were observed in the dendritesof the epidermal melanocytes of a-MSH-injected mice. Highly dendritic melanocytes seemto be the cells stimulated by MSH to form melanosomes and translocate them to dendrites.Dendritogenesis stimulated by the hormone was suppressed by actinomycin D or cycloheximide,suggesting that the dendritogenesis in the epidermal melanocytes requires de novo transcriptionand translation.
INTRODUCTION
It is well established that MSH (melanocyte-stimulating hormone) causes reversibledispersion of melanosomes in the melanophores of fishes (Chavin, 1956) and ofamphibians (Shizume, Lerner & Fitzpatrick, 1954; Wright & Lerner, i960). Inmammals, however, the effects of MSH on melanocytes have not been fully investi-gated. Krogh (1926) reported the existence of MSH in the circulating blood ofmammals. Zondek & Krohn (1932) measured the amount of MSH in the pituitarygland of several species. However, the stimulative effect of MSH on mammalianmelanocytes was observed only when Lerner, Shizume & Bunding (1954) broughtabout the darkening of human skin by intramuscular injection of an alkali-treatedextract from hog pituitary gland. Later, Lerner & McGuire (1961) reported thatsynthetic a-MSH and purified /?-MSH from hog pituitary glands also brought aboutdarkening of human skin. They repeated the experiments and confirmed the stimula-tive effect of MSH on the human pigmentary system (Lerner, Snell, Chanco-Turner& McGuire, 1966).
In mice, a few reports concerning the effects of MSH on hair colour have beenpresented. Namely, elevated endogenous (Geschwind & Huseby, 1966) and exo-genous (Geschwind, 1966) a-MSH caused a darkening of the hair of mice. Thecoats of deer mice {Peromyscus maniculatus) became lighter 7 weeks after hypo-

372 T. Hirobe
physectomy (Bronson, Eleftheriou & Dezell, 1969). Moreover, Pomerantz & Chuang(1970) reported that in newborn mice tyrosinase activity in skin homogenates waselevated after /?-MSH treatment. A question remains as to whether MSH stimulatesthe dendritogenesis of melanocytes or not - namely, whether MSH acts in the mouseby causing the slow dispersion of melanosomes within the melanocytes and bringsabout an increased production of melanin or not.
The present experiments were designed to evaluate the effect of a-MSH onepidermal melanocytes. The number of dendrites of melanocytes was counted andthe total length of dendrites was measured in the light microscope. The numberof melanosomes within melanocytes was also counted in the electron microscope, asan index of melanin content.
MATERIALS AND METHODS
The materials used in this study were newborn infants of the house mouse, Mus musatlus,of strain C57BL/10J. They were injected subcutaneously at the dorsal side with a-MSH(a gift from Ciba-Geigy) or DBc-AMP (iV6, O2'-dibutyryl-adenosine-3',5'-cyclic monophos-phate, Sigma). In some cases, they were injected with actinomycin D (Merck) or cycloheximide(Wako). These chemicals were dissolved in Hanks' BSS (balanced salt solution). After thetreatment, pieces of skin were excised from the dorsal side of mice and fixed with formalinin phosphate buffer (pH 70) for 24 h at 4 CC. They were then washed with distilled waterand incubated with o ' i%L-dopa solution in phosphate buffer (pH 7-4) for 24 h at 37 °C(Hirobe & Takeuchi, 1977a).
Alternatively, the dorsal skin was subjected to the organ culture system described previously(Hirobe & Takeuchi, 19776). Pieces of the dorsal skin of 2-day-old mice of the same strainwere excised and rinsed 3 times with culture medium, and were cultured on a glass filterplaced in a Petri dish with 5 ml of TC 199 (Ciba Kessei) in an atmosphere of 95 % O2 and5 % CO2 at 37 °C, pH 7-2, for 12 h. a-MSH, DBc-AMP, 5'AMP (s'-adenosine monophosphate,Kyowa) or theophylline (Wako) were added to the culture medium. In some cases, actinomycinD or cycloheximide was added to the culture medium. These chemicals were dissolved inHanks' BSS. After the treatment, pieces of the cultured skin were fixed and then subjected tothe dopa reaction in the same manner.
The specimens treated in vivo and in vitro were embedded in paraffin and were sectionedat 10 fim and counterstained with eosin. 240 dopa-positive cells (melanocytes) per specimenwere being scored for the number of dendrites and the total length of dendrites.
The dorsal skins of a-MSH-injected mice and Hanks' BSS-injected mice as controls weresubjected to the electron microscopy. The method for the electron-microscopic observationwas reported previously (Hirobe & Takeuchi, 1978). The dorsal skin was fixed with 4 %glutaraldehyde (Wako) in 0 1 M phosphate buffer (pH 74) and postfixed with 1% osmiumtetroxide (Merck) in 0 1 M phosphate buffer (pH 74) for 2 h at 2-4 CC. After the fixation, itwas dehydrated with ethanol and embedded in epon8i2. The ultrathin sections were cut inan LKB Ultrotome, stained with uranyl acetate and lead citrate, and examined with HitachiHS-9 electron microscope. Both experiments were triplicated. The number of melanosomeswas counted for 240 microphotograms of nucleate cells containing identifiable melanosomes.
RESULTS
Changes in the number of dendrites and the total length of dendrites in the epidermalmelanocytes of newborn mice
The dorsal skins of C57BL/10J strain mice were fixed at 1, 2, and 3 days of ageafter birth and were subjected to the dopa reaction. Fig. 1 shows the changes in the
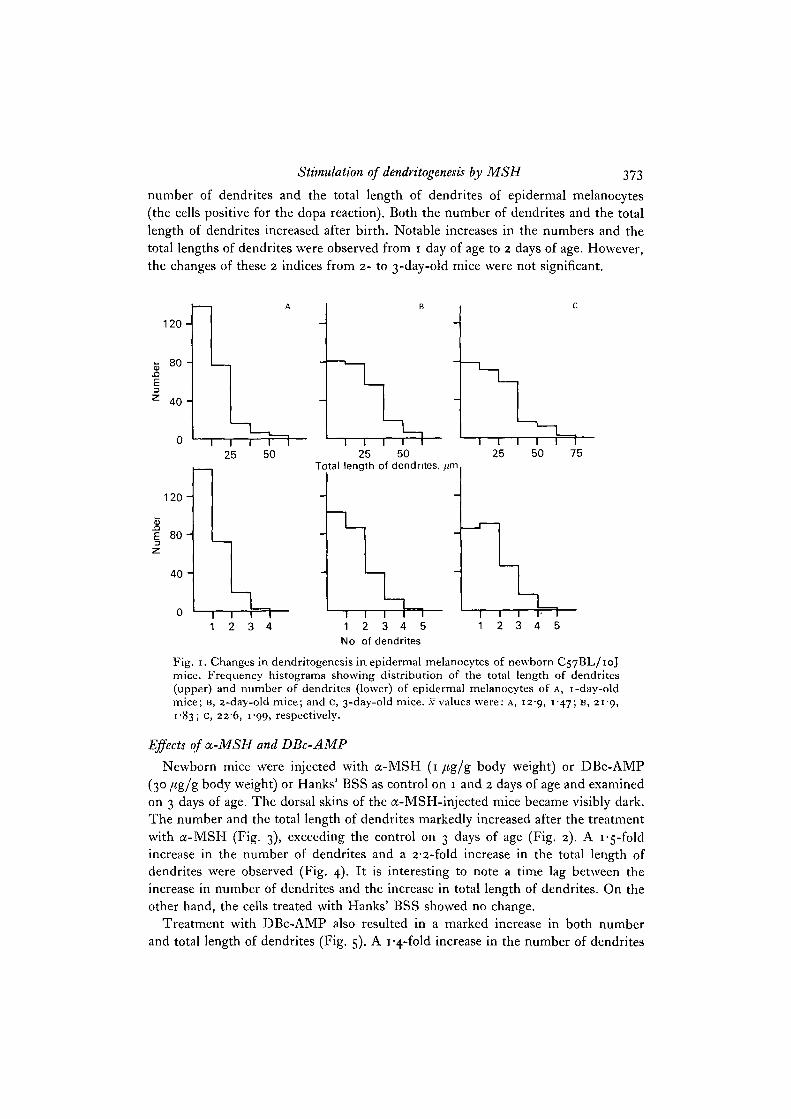
Stimulation of dendritogenesis by MSH 373
number of dendrites and the total length of dendrites of epidermal melanocytes(the cells positive for the dopa reaction). Both the number of dendrites and the totallength of dendrites increased after birth. Notable increases in the numbers and thetotal lengths of dendrites were observed from 1 day of age to 2 days of age. However,the changes of these 2 indices from 2- to 3-day-old mice were not significant.
1 2 0 -
80 -
40 -
1 2 0 -
J5E 8 0 -
40 -
25 50Total length of dendrites, //m
1 2 3 4i i r1 2 3 4 5
No of dendrites
1 I l~~12 3 4 5
Fig. 1. Changes in dendritogenesis in epidermal melanocytes of newborn C57BL/10Jmice. Frequency histograms showing distribution of the total length of dendrites(upper) and number of dendrites (lower) of epidermal melanocytes of A, 1-day-oldmice; B, 2-day-old mice; and c, 3-day-old mice, .v values were: A, 129, 147; B, 219,1 83; C, 226, 1 99, respectively.
Effects of a-MSH and DBc-AMP
Newborn mice were injected with a-MSH (1 /tg/g body weight) or DBc-AMP(30 /tg/g body weight) or Hanks' BSS as control on 1 and 2 days of age and examinedon 3 days of age. The dorsal skins of the a-MSH-injected mice became visibly dark.The number and the total length of dendrites markedly increased after the treatmentwith a-MSH (Fig. 3), exceeding the control on 3 days of age (Fig. 2). A i-5-foldincrease in the number of dendrites and a 2-2-fold increase in the total length ofdendrites were observed (Fig. 4). It is interesting to note a time lag between theincrease in number of dendrites and the increase in total length of dendrites. On theother hand, the cells treated with Hanks' BSS showed no change.
Treatment with DBc-AMP also resulted in a marked increase in both numberand total length of dendrites (Fig. 5). A 1 "4-fold increase in the number of dendrites
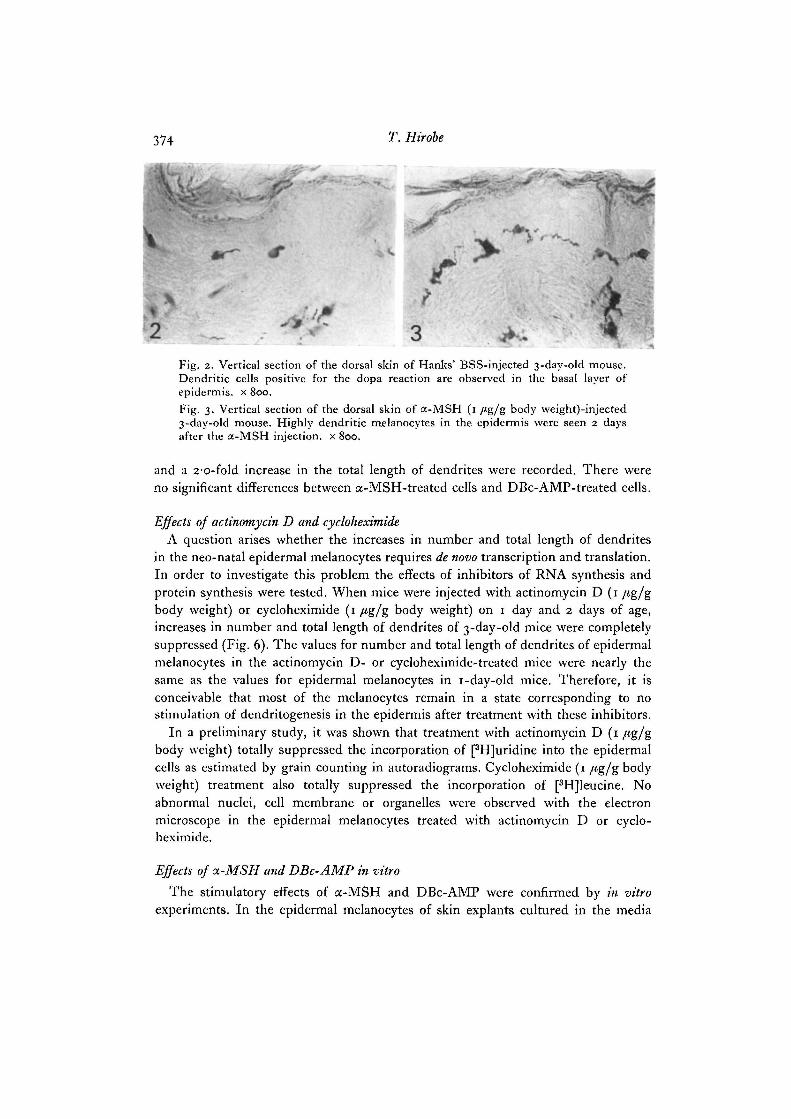
374 T. Hirobe
•*y. „
•4Fig. 2. Vertical section of the dorsal skin of Hanks' BSS-injected 3-day-old mouse.Dendritic cells positive for the dopa reaction are observed in the basal layer ofepidermis, x 800.Fig. 3. Vertical section of the dorsal skin of a-MSH (1 /<g/g body \veight)-injected3-day-old mouse. Highly dendritic melanocytes in the epidermis were seen 2 daysafter the a-MSH injection, x 800.
and a 2-o-fold increase in the total length of dendrites were recorded. There wereno significant differences between a-MSH-treated cells and DBc-AMP-treated cells.
Effects of actinomycin D and cycloheximideA question arises whether the increases in number and total length of dendrites
in the neo-natal epidermal melanocytes requires de novo transcription and translation.In order to investigate this problem the effects of inhibitors of RNA synthesis andprotein synthesis were tested. When mice were injected with actinomycin D (1 /tg/gbody weight) or cycloheximide (1 /<g/g body Weight) on 1 day and 2 days of age,increases in number and total length of dendrites of 3-day-old mice were completelysuppressed (Fig. 6). The values for number and total length of dendrites of epidermalmelanocytes in the actinomycin D- or cycloheximide-treated mice were nearly thesame as the values for epidermal melanocytes in 1-day-old mice. Therefore, it isconceivable that most of the melanocytes remain in a state corresponding to nostimulation of dendritogenesis in the epidermis after treatment with these inhibitors.
In a preliminary study, it was shown that treatment with actinomycin D (1 /tg/gbody weight) totally suppressed the incorporation of [3H]uridine into the epidermalcells as estimated by grain counting in autoradiograms. Cycloheximide (1 /tg/g bodyweight) treatment also totally suppressed the incorporation of [3H]leucine. Noabnormal nuclei, cell membrane or organelles were observed with the electronmicroscope in the epidermal melanocytes treated with actinomycin D or cyclo-heximide.
Effects of a-MSH and DBc-AMP in vitro
The stimulatory effects of a-MSH and DBc-AMP were confirmed by in vitroexperiments. In the epidermal melanocytes of skin explants cultured in the media
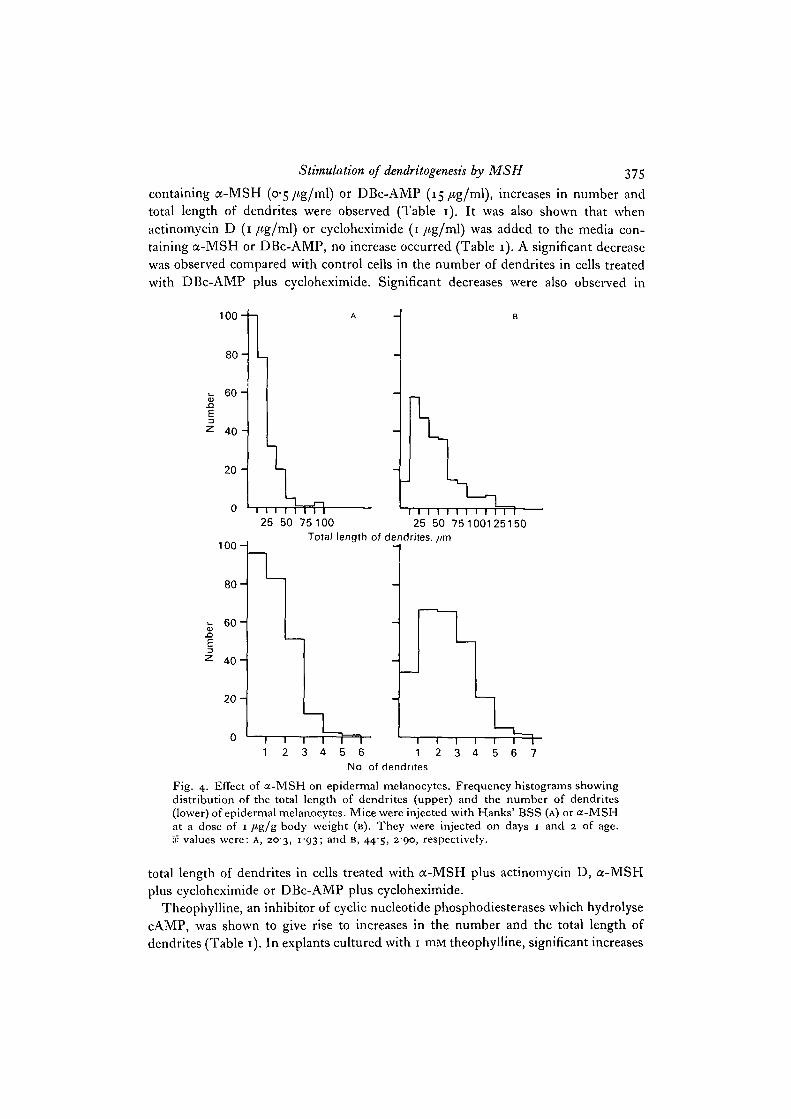
Stimulation of dendritogenesis by MSH 375
containing a-MSH (o'5/ig/ml) or DBc-AMP (15/tg/ml), increases in number andtotal length of dendrites were observed (Table 1). It was also shown that whenactinomycin D (1 /tg/ml) or cycloheximide (1 /tg/ml) was added to the media con-taining a-MSH or DBc-AMP, no increase occurred (Table 1). A significant decreasewas observed compared with control cells in the number of dendrites in cells treatedwith DBc-AMP plus cycloheximide. Significant decreases were also observed in
.aE
100-
8 0 -
6 0 -
4 0 "
20 -
0
A
'—1
1 1 1 1 i 1
10CH
8 0 -
15.aE
40 H
2 0 -
25 50 75 100 25 50 75100125150Total length of dendrites, //m
1 2 3 41 r ~5 6 1 2
No of dendrites3 4 5 6 7
Fig. 4. Effect of a-MSH on epidermal melanocytes. Frequency histograms showingdistribution of the total length of dendrites (upper) and the number of dendrites(lower) of epidermal melanocytes. Mice were injected with Hanks' BSS (A) or a-MSHat a dose of 1 /tg/g body weight (B). They were injected on days 1 and 2 of age.x values were: A, 203, i"93; and B, 445, 290, respectively.
total length of dendrites in cells treated with a-MSH plus actinomycin D, a-MSHplus cycloheximide or DBc-AMP plus cycloheximide.
Theophylline, an inhibitor of cyclic nucleotide phosphodiesterases which hydrolysecAMP, was shown to give rise to increases in the number and the total length ofdendrites (Table 1). In explants cultured with 1 mni theophylline, significant increases
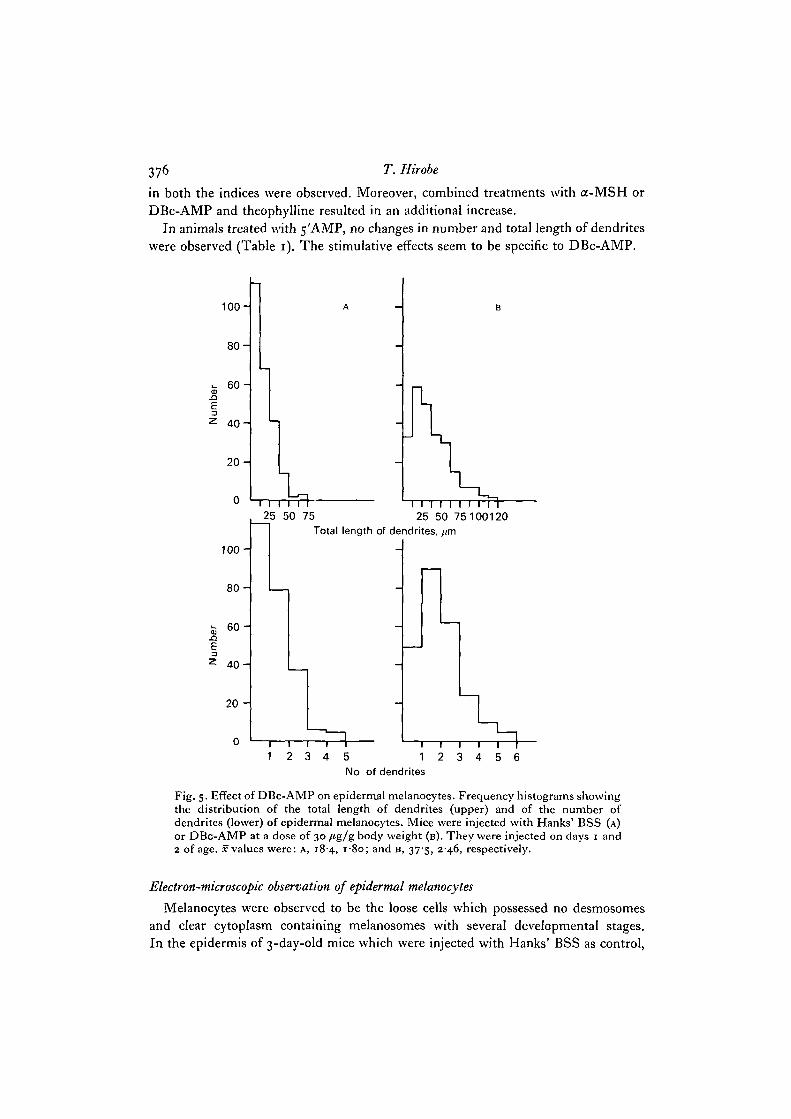
376 T. Hirobe
in both the indices were observed. Moreover, combined treatments with a-MSH orDBc-AMP and theophylline resulted in an additional increase.
In animals treated with 5'AMP, no changes in number and total length of dendriteswere observed (Table 1). The stimulative effects seem to be specific to DBc-AMP.
1 0 0 -
8 0 -
E3
6 0 -
4 0 -
2 0 -
100-
8 0 -
6 0 -
4 0 -
20 -
I I T I 725 50 75
1 1 I 1 I I I 1 I 125 50 75 100120
Total length of dendrites, /im
1 2 3 4 5 1No of dendrites
1 I p2 3 4 5
Fig. 5- Effect of DBc-AMP on epidermal melanocytes. Frequency histograms showingthe distribution of the total length of dendrites (upper) and of the number ofdendrites (lower) of epidermal melanocytes. Mice were injected with Hanks' BSS (A)or DBc-AMP at a dose of 30 /tg/g body weight (B). They were injected on days 1 and2 of age. x values were: A, 184, 180; and B, 37'5, 2-46, respectively.
Electron-microscopic observation of epidermal melanocytes
Melanocytes were observed to be the loose cells which possessed no desmosomesand clear cytoplasm containing melanosomes with several developmental stages.In the epidermis of 3-day-old mice which were injected with Hanks' BSS as control,

Stimulation of dendritogenesis by MSH 377
160-
120-
8 0 -
40 -
i—i—i—i—m r25 50 75 25 50
Total length of dendrites, //m
25 50
160-
120-
CD
•g 8 0 -
4 0 -
1 2 3 4 5—i—i—r=r—
1 2 3 4
No of dendrites
1 2 3 4
Fig. 6. Effect of actinomycin D or cycloheximide on epidermal melanocytes. Fre-quency histograms showing distribution of the total length of dendrites (upper) andthe number of dendrites (lower) of epidermal melanocytes. Mice were injected withHanks' BSS (A) or actinomycin D at a dose of i /*g/g body weight (B) or cyclohexi-mide at a dose of i /tg/g body weight (c). They were injected on days i and 2 ofage. x values were: A, 217, 1-79; B, 12-5, i-43; and C, 13-1, 1-43, respectively.
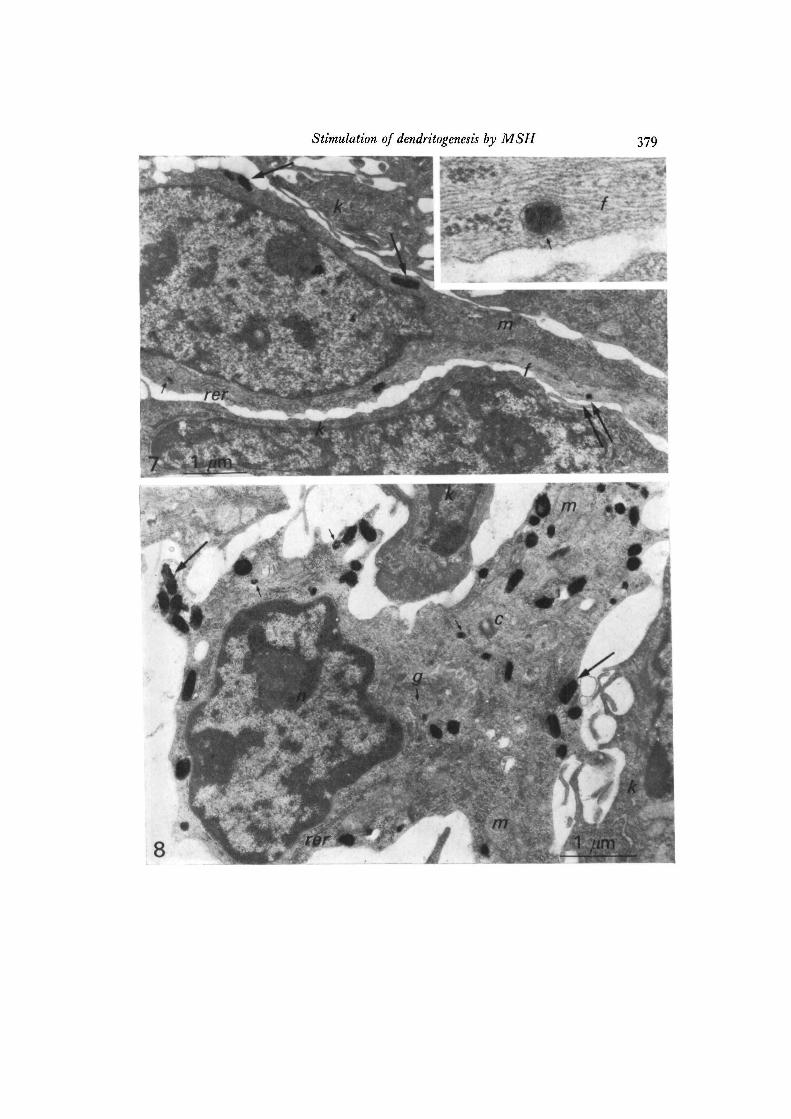
melanocytes with several melanosomes were found (Fig. 7). On the other hand, theepidermis of a-MSH-treated mice contained melanocytes with numerous fullymelanized melanosomes (Fig. 8). In the a-MSH-treated cells, many melanosomeswere observed in the area of the dendrites. The numbers of melanosomes of differentstages were recorded for 240 microphotograms of nucleate cells containing identifiablemelanosomes for the Hanks' BSS-treated control skin and for the a-MSH-treatedskin, respectively. The stage of melanosome maturation was categorized accordingto Fitzpatrick, Hori, Toda & Seiji (1969). Stages I and II include immature pre-melanosomes, while melanized melanosomes are classified as stages III and IV. Inthe a-MSH-treated cells, an increase in the number of melanosomes per cell wasalso observed (Table 2). The a-MSH-treated cells contained many more stagesIII + IV melanosomes than the control cells. On the other hand, a-MSH-treated

378 T. Hirobe
cells contained fewer stages I+ 11 melanosomes than control cells (Fig. 9). Therefore,
it is conceivable that melanin content within the melanocyte is markedly increased
by MSH treatment.
Table 1. Effect of OL-MSH and DBc-AMP on the epidermalmelanocytes in vitro
Number of Total lengthGroup dendrites of dendrites
Cultured control 1000 ±3-1" 100015-36
a-MSH (0-5 /tg/ml) 1299 ±44C 1323 i S ^ "a-MSH (05 /tg/ml)+ theophylline (1 IJM) i54o±4-6e 200-5 ± 7 7 /
a-MSH (0-5 /ig/ml) + actinomycin D (1 fig/ml) 97-3±3'2" j%(>±^-<)h
a-MSH (05/tg/ml) + cycloheximide (1/tg/ml) 101-7 + 3-3' 73'8±53J
DBc-AMP (15 /tg/ml) 1438 ± 4-2* 168-5 ± 6-8'DBc-AMP (15/tg/ml)+ theophylline (1 m.M) i56-2±47'" 185-817-6"DBc-AMP (15/tg/ml) + actinomycin D (1 /ig/ml) 94-615-2° 103-61 3-8''DBc-AMP (15/tg/ml)+ cycloheximide (1/tg/ml) 8o-4 + 2-8" 73'7i3-i r
Theophylline (1 mM) 119-2! 3-9* 122-018-2'5'AMP (0-03 mM) 104-1137" 96-018-o1'
Pieces of skin were cultured in TC 199 with or without a-MSH or DBc-AMP for 12I1 at
37 °C.Each value is the mean is.E. (standard error) from 240 epidermal melanocytes in 8 different
skin explants and expressed as percent of the cultured control. Control of 2 days of age(IOI-3 1 3-5, 1007 + 7-3) was not significantly different from the cultured control.
(-test, significant differences: a-c (P < 0001); a-e (P < o-ooi); a-k (P < 0001); a-111(P < o-ooi); a-q (P < o-ooi); as (oooi < P < 0005); b-d (P < 0001); b-f (P < 0001);b-h(oooi < P < 0005); 6-y (P < o-ooi);b-l(P < o-ooi);b-n(P < o-ooi);b-r (P < o-ooi);b-t (002 < P < 005); c-e (P < 0001); d-f (P < 0001).
N o t significant differences: a—g, a—i, a—o, a—u, b—p, b—v, e—nt,f—n, k—m, l—n.
DISCUSSION
Lerner & McGuire (1961), injecting human skin with a-MSH or /?-MSH, haveshown that these hormones are effective agents for darkening the skin. The dispersionof melanosomes seemed to occur within the melanocytes in a manner similar to thatseen in the melanophores of frogs and fishes following exposure to MSH, since
Fig. 7. Electron micrograph of an epidermal melanocyte (Hanks' BSS-treatedcontrol). One melanosome is seen in the dendrite. Inset shows the connexion ofa stage III melanosome with intermediate filaments (a higher magnification of theportion indicated by double arrow). Small arrow, stage III melanosome; largearrow, stage IV melanosome; /, intermediate filament (10-nm filament); k, basalkeratinocyte; m, mitochondrion; n, nucleus; rer, rough endoplasmic reticulum.x 17300. Inset, x 100000.Fig. 8. Electron micrograph of an epidermal melanocyte (a-MSH-treated). Numerousmature melanosomes are seen, particularly in the dendrite areas. Small arrow,stage III melanosome; large arrow, stage IV melanosome; c, centriole; g, Golgiapparatus; k, basal keratinocyte; m, mitochondrion; n, nucleus; rer, rough endo-plasmic reticulum. x 21000.
Stimulation of dendritogenesis by MSH 379
38o T. Hirobe
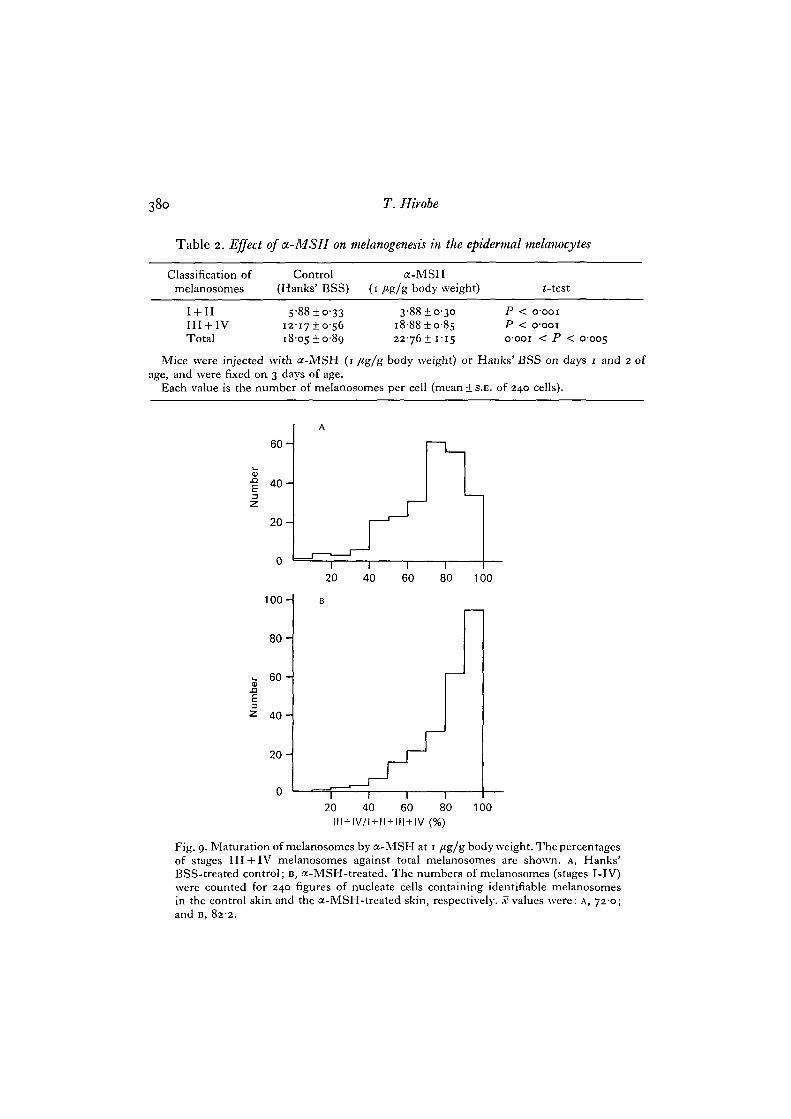
Table 2. Effect of a-MSH on melanogenesis in the epidermal melanocytes
Classification of Control a-MSHmelanosomes (Hanks' BSS) (1 /tg/g body weight) <-test
I + IIIII + IVTotal
1 2 -
i 8 -
88 ±o-331710-5605 ±0-89
3-88 + 0-30i8-88±o-8s22-76! 1-15
PPo-
< 0001
< 0001
001 < P 0-005
Mice were injected with a-MSH (1 /*g/g body weight) or Hanks' BSS on days 1 and 2 ofage, and were fixed on 3 days of age.
Each value is the number of melanosomes per cell (mean + S.E. of 240 cells).
6 0 -
4 0 -
2 0 -
a>. aE3
6 0 -
4 0 -
2 0 -
20 40 60 80 100
100H B
8 0 -
1 r2 0 4 0 6 0 8 0
III + IV/I + II + III + IV (%)
100
Fig. 9. Maturation of melanosomes by a-MSH at 1 /tg/gbody weight. The percentagesof stages III + IV melanosomes against total melanosomes are shown, A, Hanks'BSS-treated control; B, a-MSH-treated. The numbers of melanosomes (stages I-IV)were counted for 240 figures of nucleate cells containing identifiable melanosomesin the control skin and the a-MSH-treated skin, respectively, x values were: A, 720;and B, 822.

Stimulation of dendritogenesis by MSH 381
darkening begins almost within 24 h. They assumed that newly synthesized melaninmight account for the prolonged hyperpigmentation. On the other hand, Snell(1962) reported that /?-MSH produced no obvious macroscopic changes in colourof guinea pigs. However, histological examination showed that the dendrites ofepidermal melanocytes became larger and more complex. Therefore, in guinea pigs,stimulation of dendritogenesis in the epidermal melanocytes seems to result in nodarkening of coat colour. The experiments with a-MSH on guinea pigs showed thesame tendency (Snell, 1964). In the present study, it was shown that the colour ofnewborn mouse skin darkened and the number and total length of dendrites markedlyincreased 48 h after the injection with a-MSH (1 /*g/g body weight). Therefore, itis conceivable that MSH brings about the darkening of mouse body skin by stimu-lating the dendritogenesis of epidermal melanocytes.
There have been several reports concerning the effect of MSH on dissociatedmammalian epidermal melanocytes (Klaus & Snell, 1967; Kitano, 1976). However,only a few studies to investigate the effects of MSH on epidermal melanocytes inorgan culture have been reported. In the present study, stimulation of dendritogenesisof epidermal melanocytes of newborn mouse skin in organ culture was recognized12 h after the addition of a-MSH (0-5 /tg/ml) into the culture medium.
Electron-microscopic observations showed that the numbers of melanosomes withinthe epidermal melanocytes and the percentages of mature melanosomes against totalmelanosomes within the melanocytes increased significantly in the a-MSH-treatedgroups. Many mature melanosomes were observed in the dendrites of a-MSH-injected mice. Therefore, it is possible that MSH stimulates new melanosomeformation, and the maturity of melanosomes as well as the translocation of maturemelanosomes to tKe dendrite areas. The above-mentioned effects of a-MSH atelectron-microscopic levels seem to be comparable to the increases in number andtotal length of dendrites revealed in histological preparations for light microscope.
In the present study, the effects of DBc-AMP mimicked those of a-MSH. Theincreases in number and total length of dendrites were observed after injection ofDBc-AMP at a dose of 30 /tg/g body weight. The effect of DBc-AMP was revealedin the 2 indices showing the degree of dendritogenesis that were observed 12 h afteraddition of DBc-AMP (15 /tg/ml) into the culture medium. Therefore, it is probablethat the stimulation of dendritogenesis in the epidermal melanocytes by MSH ismediated through cAMP. This assumption is supported by the results with theo-phylline, which induced in organ culture increases in both number and total lengthof dendrites in epidermal melanocytes, similar to that induced by a-MSH or DBc-AMP.
The experiments in which actinomycin D and cycloheximide are involved showedthat in most melanocytes dendritogenesis remained unstimulated. The resultsindicate the possibility that dendritogenesis in epidermal melanocytes requires denovo RNA synthesis and protein synthesis. The stimulation of dendritogenesis inepidermal melanocytes in organ culture by a-MSH or DBc-AMP was also almostcompletely suppressed by addition of actinomycin D or cycloheximide to the culturemedia. The results suggest that the enhancement of dendritogenesis in response to
25 CEL 33

382 T. Hirobe
MSH or cAMP requires de novo transcription and translation. A mechanism, of thesame kind seems to be involved in both normal and MSH-stimulated dendritogenesisin melanocytes, since the dendritogenesis in both cases requires de novo transcriptionand translation.
We have presented evidence that MSH induces melanogenesis in the epidermalmelanoblasts of newborn mouse skin in vivo (Hirobe & Takeuchi, 1977a, 1978) andin vitro (Hirobe & Takeuchi, 19776). A problem which remains unresolved is thediscrimination between the mechanism involved in the inducing effect of MSH onthe differentiation of melanoblasts and that involved in the stimulating effect ofMSH on the activities of differentiated melanocytes.
The author expresses his thanks to Dr T. Takeuchi of Tohoku University for his criticalreading of the manuscript.
Thanks are also due to Dr W. Rittel for providing synthetic a-MSH.
REFERENCES
BRONSON, F. H., ELEFTHERIOU, B. E. & DEZELL, H. E. (1969). Melanocyte-stimulating activityfollowing adrenalectomy in deermice. Proc. Soc. exp. Biol. Med. 130, 527-529.
CHAVIN, W. (1956). Pituitary-adrenal control of melanization in xanthic goldfish, Carassiusauratus L. jf. exp. Zool. 133, 1-45.
FITZPATRICK, T. B., HORI, Y., TODA, K. & SEIJI, M. (1969). Melanin 1969: some definitionsand problems. Jap. J. Derm. ser. B 79, 278-282.
GESCHWIND, I. I. (1966). Change in hair color in mice induced by injection of a-MSH.Endocrinology 79, 1165-1167.
GESCHWIND, I. I. & HUSEBY, R. A. (1966). Melanocyte-stimulating activity in a transplantablemouse pituitary tumor. Endocrinology 79, 97-105.
HIROBE, T. & TAKEUCHI, T. (1977a). Induction of melanogenesis in the epidermal melano-blasts of newborn mouse skin by MSH. jf. Embryol. exp. Morph. 37, 79-90.
HIROBE, T . & TAKEUCHI, T. (19776). Induction of melanogenesis in vitro in the epidermalmelanoblasts of newborn mouse skin by MSH. In Vitro 13, 311-315.
HIROBE, T. & TAKEUCHI, T. (1978). Changes of organelles associated with the differentiationof epidermal melanocytes in the mouse, jf. Embryol. exp. Morph. 43, 107-121.
KITANO, Y. (1976). Effects of melanocyte stimulating hormone and theophylline on humanmelanocytes in vitro. Archs Derm. Res. 255, 163-168.
KLAUS, S. N. & SNELL, R. S. (1967). The response of mammalian epidermal melanocytes inculture to hormones, jf. invest. Derm. 48, 352—358.
KROGH, A. (1926). The pituitary (posterior lobe) principle in circulating blood, jf. Pharmac.exp. Ther. 29, 177-189.
LERNER, A. B. & MCGUIRE, J. S. (1961). Effect of alpha- and beta-melanocyte stimulatinghormones on the skin colour of man. Nature, Lond. 189, 176-179.
LERNER, A. B., SHIZUME, K. & BUNDING, I. (1954). The mechanism of endocrine control ofmelanin pigmentation, jf. din. Endocr. Metab. 14, 1463-1490.
LERNER, A. B., SNELL, R. S., CHANCO-TURNER, M. L. & MCGUIRE, J. S. (1966). Vitiligo and
sympathectomy. The effect of sympathectomy and a-melanocyte stimulating hormone.Archs Derm. Syph. 94, 269-278.
POMERANTZ, S. H. & CHUANG, L. (1970). Effects of /?-MSH, cortisol and ACTH on tyrosinasein the skin of newborn hamsters and mice. Endocrinology 87, 302-310.
SHIZUME, K., LERNER, A. B. & FITZPATRICK, T. B. (1954). In vitro bioassay for the melanocytestimulating hormone. Endocrinology 54, 553-560.
SNELL, R. S. (1962). Effect of the melanocyte stimulating hormone of the pituitary on melano-cytes and melanin in the skin of guinea-pigs. J. Endocr. 25, 249-258.

Stimulation of dendntogenesis by MSH 383
SNELL, R. S. (1964). Effect of the alpha melanocyte stimulating hormone of the pituitary onmammalian epidermal melanocyte. J. invest. Derm. 42, 337-347-
WRIGHT, M. R. & LERNER, A. B. (i960). On the movement of pigment granules in frogmelanocyte. Endocrinology 66, 599-609.
ZONDEK, B. & KROHN, H. (1932). Hormon des Zwischenlappens der Hypophyse (lntermedin).II. lntermedin im Organismus (Hypophyse, Gehirn). Klin. Wchnschr. n , 849-853.
{Received 7 March 1978)
25-2








![Journal of Pigmentary Disorders - Novoxel the number of vessels and epidermal pigmentation in melasma [18]. Electron microscopy studies have revealed that the melanocytes are filled](https://img.pdfslide.us/doc/110x75/5af01d5e7f8b9a572b8ef73b/journal-of-pigmentary-disorders-the-number-of-vessels-and-epidermal-pigmentation.jpg)






![D a l J u Journal of Pigmentary Disorders DOI · between the number of vessels and epidermal pigmentation in melasma [18]. Electron microscopy studies have revealed that the melanocytes](https://img.pdfslide.us/doc/110x75/5bef8e0209d3f274038bc962/d-a-l-j-u-journal-of-pigmentary-disorders-doi-between-the-number-of-vessels.jpg)