Embed Size (px)
DESCRIPTION
mjk
Citation preview

OR IG INAL
ART ICLE
Participation of central imidazoline binding
sites in antinociceptive effect of ethanol and
nicotine in rats
Manish Manohar Aglawe, Brijesh Gulabrao Taksande,
Sharvari Shambabu Kuldhariya, Chandrabhan Tukaram Chopde,
Milind Janrao Umekar, Nandkishor Ramdas Kotagale*2Division of Neuroscience, Department of Pharmacology, Smt. Kishoritai Bhoyar College of Pharmacy, New Kamptee,
Nagpur 441002, Maharashtra, India
Keywords
antinociception,
ethanol,
imidazoline Receptors,
nicotine4
Received 6 July 2012;
revised 6 February 2013;
accepted 28 March 2013
*Correspondence and reprints:
ABSTRACT
Despite synergistic morbidity and mortality, concomitant consumption of alcohol
and tobacco is increasing, and their antinociceptive effect has been linked with
co-abuse. Present study was designed to investigate the role of imidazoline binding
sites in the antinociceptive effect of nicotine, ethanol, and their combination. Sepa-
rate group of male Sprague–Dawley rats (200–250 g) were treated with different
doses of alcohol (0.50–2 g/kg, i.p.) or nicotine (0.25–1 mg/kg, i.p.), and their
combination evaluated in tail flick test. Influence of endogenous imidazoline bind-
ing site ligands, agonist, and antagonists were determined by their prior treatment
with effective or subeffective doses of either ethanol or nicotine. Ethanol, nicotine,
or their subeffective dose combination exhibited significant antinociceptive effects
in dose-dependent manner. Antinociceptive3 effect of ethanol and nicotine was sig-
nificantly augmented by intracerebroventricular (i.c.v.) administration of endoge-
nous imidazoline receptor ligands, harmane (25 lg/rat, i.c.v.) and agmatine
(10 lg/rat, i.c.v.), as well as imidazoline I1/a2 adrenergic receptor agonist, cloni-
dine (2 lg/rat, i.c.v.), I1 agonist moxonidine (25 lg/rat, i.c.v.), and imidazoline I2agonist, 2-BFI (10 lg/rat, i.c.v.). Conversely, antinociception elicited by ethanol or
nicotine or their subeffective dose combination was antagonized by pretreatment
with imidazoline I1 antagonist, efaroxan (10 lg/rat, i.c.v.), and I2 antagonist,
idazoxan (4 lg/rat, i.c.v.), at their per se ineffective doses. These findings project
imidazoline binding ligands as important therapeutic molecules for central antino-
ciceptive activity as well as may reduce the co-abuse potential of alcohol and
nicotine.
INTRODUCT ION
Concomitant consumption of alcohol and tobacco
remains high despite the observed synergistic morbidity
in society. Repeatedly, scientific studies have cautioned
about the synergistic risks for various cancers with
concurrent drinking and smoking [1–3]. Several factors
including common genetic mechanisms, pharmacoki-
netic and pharmacodynamic interactions predisposing
to increased rewarding effects, and/or counteracting
toxic effects of alcohol by nicotine have been proposed
for co-abuse [4–16]. In addition, a number of clinical
[17,18] and animal studies [19] have provided evi-
dences for common genetic predisposition for their
co-abuse.
Pharmacokinetic data revealed that nicotine dose
dependently reduces blood alcohol levels in rats [14].
Interestingly, alcohol-induced ataxia as well as cognitive
impairments was also protected by nicotine [20–23].
Moreover, combination of alcohol and nicotine found to
1
2
3
4
5
6
7
8
9
10
11
12
13
14
15
16
17
18
19
20
21
22
23
24
25
26
27
28
29
30
31
32
33
34
35
36
37
38
39
40
41
42
43
44
45
46
47
48
49
50
ª 2013 The Authors Fundamental and Clinical Pharmacology © 2013 Soci�et�e Franc�aise de Pharmacologie et de Th�erapeutique
Fundamental & Clinical Pharmacology 1
doi: 10.1111/fcp.12034
FC
P1
20
34B
Dispa
tch:
9.4.13
Jour
nal:
FCP
CE:L
iyag
atAli
JournalName
ManuscriptNo.
Autho
rRec
eive
d:No.
ofpa
ges:
10PE
:Kav
itha

exhibit increased rewarding effects associated with exag-
gerated dopamine release in the nucleus accumbens
shell of rats [24,25].
Alcohol consumption in humans [26,27] and admin-
istration to animals [28] result in analgesia or antino-
ciception. Similarly, nicotine or nicotinic agonists
produce analgesic effect in humans [29,30] and in
animals [31–35]. Available neurological and
pharmacological evidences suggest the existence of
close relationship between reinforcement and analgesia.
Moreover, it is proposed that additive or synergistic
antinociceptive effects induced by alcohol and nicotine
may contribute to their co-abuse [36,37].
Imidazoline binding sites have currently attracted
attention in nociception as well as drug addiction [38–
40]. Moreover, the brain structures that are involved
in the drug abuse and pain perception including hypo-
thalamus, hippocampus, amygdala, etc., are rich in
imidazoline binding sites and its endogenous ligands
[41].
Imidazoline binding sites are a family of unique non-
adrenergic high-affinity binding sites that exist in three
major subclasses (I1, I2, and I3) based upon their ligand
selectivity, subcellular distribution, and physiological
functions [42–45]. In human brain, I1 receptors are
distributed in regional manner with highest density in
striatum, pallidum, and gyrus dentatus of hippocam-
pus, amygdala, and substantia nigra [46]. The I2 bind-
ing sites (I2A and I2B) are allosteric and are located on
monoamine oxidases [45,47,48]. The interaction of
imidazoline binding sites and ligands on ethanol and
nicotine intake, their dependence, and withdrawal state
is well documented [38,49,50]. Furthermore, the
involvement of imidazoline I1/I2 endogenous ligands
like agmatine, harmane, and b-carboline in nociception
as well as addiction is now fairly well established. How-
ever the influence of imidazoline binding site modula-
tion on antinociceptive effect of ethanol and nicotine
remains unexplored.
In view of this background, this study was under-
taken to investigate the effect of imidazoline binding
sites ligands on ethanol- and nicotine-induced antinoci-
ceptive effect in rats using tail flick assay method
reflecting spinal nociception in rats.
MATER IALS AND METHODS
Animals
Adult healthy Sprague–Dawley rats (200–250 g) were
used. The rats were kept four per cage (640 9 410 9
250 mm height) or individually after intracerebroven-
tricular (i.c.v.) cannulation, in a room with controlled
temperature (25 � 2 °C) and maintained on a 12:12 h
light/dark cycle (on/off at 07:00 am/07:00 pm) with
free access to food and water. All experimental proce-
dures were approved by the Institutional Animal Ethi-
cal Committee and executed in strict accordance with
the guidelines for the care and use of laboratory
animals (CPCSEA, India 5).
Drugs
Nicotine hydrogen tartrate, agmatine sulfate, clonidine
hydrochloride, moxonidine hydrochloride, harmane,
efaroxan hydrochloride, and idazoxan hydrochloride
were purchased from Sigma-Aldrich Co., USA 6, while
2-(2-benzofuranyl)-2-imidazoline hydrochloride (2-BFI)
was purchased from Tocris Biosciences, UK 7.
Ethanol (99%) was purchased from Merck chemicals,
Mumbai, India. Ethanol was diluted or nicotine was
dissolved in physiologic saline (0.9%) and administered
by intraperitoneal (i.p.) route in a volume of 1 mL/kg
of body weight. Agmatine, moxonidine, 2-BFI, efaro-
xan, and idazoxan were injected by intracerebroven-
tricular (i.c.v., 2 lL/rat) route. For i.c.v. administration
of drugs, dilutions were made with artificial cerebrospi-
nal fluid (aCSF) of following composition: 0.2 M NaCl,
0.02 M NaH2CO3, 2 mM KCl, 0.5 mM KH2PO4, 1.2 mM
CaCl2, 1.8 mM MgCl2, 0.5 mM Na2SO4, and 5.8 mM
D-glucose.
Intracerebroventricular cannula implantation and
drug administration
For the study involving i.c.v. administration of drugs, rats
were anesthetized with thiopental sodium (60 mg/kg,
i.p.; Abbott Pharmaceuticals Ltd., Mumbai, India) and
stereotaxically (David Kopf Instruments, CA, USA 8)
implanted [49,50] 24-gauge stainless steel guide cannula
(C313G/Spc, plastic UK 9). Stereotaxic coordinates used
were �0.8 mm posterior, +1.3 mm lateral to midline,
and �3.5 mm ventral to bregma according to Paxinos
and Watson [51]. The guide cannulae were then fixed to
skull with dental cement (DPI-RR cold cure, acrylic pow-
der; Dental Product of India, Mumbai, India) and secured
in two stainless steel screws. A 28-gauge stainless steel
dummy cannula was used to occlude the guide cannula
when not in use. Following surgery, the animals were
placed individually in cages and each subject was allowed
to recover at least for 7 days before being tested for tail
flick latency. Rats were then randomly assigned to
different groups (n = 6 per group) and habituated to the
ª 2013 The Authors Fundamental and Clinical Pharmacology ª 2013 Soci�et�e Franc�aise de Pharmacologie et de Th�erapeutique
Fundamental & Clinical Pharmacology
2 M.M. Aglawe et al.
1
2
3
4
5
6
7
8
9
10
11
12
13
14
15
16
17
18
19
20
21
22
23
24
25
26
27
28
29
30
31
32
33
34
35
36
37
38
39
40
41
42
43
44
45
46
47
48
49
50

testing environment by transferring to experimental
room and twice daily handling for 1 week. Drugs were
injected (2 lL/rat) bilaterally over a 1-min period with a
microliter syringe (Hamilton, Reno, NV, USA) connected
by PE-10 polyethylene tubing to a 33-gauge internal
cannula (C313 I/Spc, plastic one, internal diameter
0.18 mm, outer diameter 0.20 mm) that extended
0.5 mm beyond the guide cannula. The internal cannula
was held in position for another 1 min before being
slowly withdrawn to prevent backflow and promote diffu-
sion of drugs.
Confirmation of cannula placement
At the end of all experiments, dilute India ink
(5 lL/rat, i.c.v.) was injected and animals were killed
by an overdose of thiopental sodium. The brain of each
animal was dissected out and sliced in coronal plane to
verify the placement of the guide cannula and distribu-
tion of ink in the ventricles. In 13% of animals, guide
cannulae were found incorrectly placed. Data of only
those animals that showed uniform distribution of ink
in the ventricles were considered for statistical analysis.
Assessment of nociception
Nociception was assessed with tail flick apparatus
(INCO, India10 ). It consisted of an electrically heated
nichrome wire (1/8 mm) as a radiant heat source.
A desired intensity was adjusted by supplying constant
current (6 �A) to wire so that a sudden flick of the tail
occurred in about 5–6 s. The basal reaction time (pre-
drug reaction time) to radiant heat was determined by
placing the tip (dorsal 2–3 cm) of the tail on hot wire.
A cutoff period of 15 s was observed to avoid damage
to the tail [52,53]. Animals failing to withdraw tail in
5–6 s were rejected from study. At least three basal
reaction times for each rat with the interval of 1 min
were recorded. Animals were then randomly divided
into different groups each containing six animals for
further studies as described below. All the measure-
ments were taken by the skilled observer blind to the
treatment given.
Effect of ethanol and nicotine on nociception
Animals were randomly divided into different groups
(n = 6), and each group was assigned to one of the fol-
lowing treatment. Firstly, we determined the acute
antinociceptive effect of various doses of ethanol (8%
w/v in saline, 0.50–2 g/kg, i.p.) or nicotine (0.25–
1 mg/kg, i.p.) or their combination or saline (1 mL/kg,
i.p.) in tail flick assay. For combination studies,
submaximal or ineffective doses of both the drugs were
used. Ethanol was always administered 30 min before
and nicotine 20 min before the tests. Individual rat
was subjected to tail flick test, and reaction latency
was determined.
Effect of endogenous ligands of imidazoline
binding sites on ethanol- or nicotine-induced
antinociception
In these experiments, rats were treated with endogenous
ligands of imidazoline binding sites viz. agmatine
(10 lg/rat, i.c.v.) or harmane (25 lg/rat, i.c.v.) or aCSF
(2 lL/rat, i.c.v.) 10 min before i.p. injection of vehicle or
subeffective dose of ethanol (8% w/v; 0.5 g/kg, i.p.) or
nicotine (0.25 mg/kg, i.p.) or saline (1 mL/kg, i.p.).
Effect of imidazoline receptor agonists on
ethanol- and nicotine-induced antinociception
Separate group of rats (n = 4–16) were administered
either with aCSF (2 lL/rat, i.c.v.) or agmatine (10–
40 lg/rat, i.c.v.) or harmane (25–100 lg/rat, i.c.v.) or
clonidine (2–8 lg/rat, i.c.v.) or moxonidine (25–
100 lg/rat, i.c.v.) or 2-BFI (10–40 lg/rat, i.c.v.) or
efaroxan (10–40 lg/rat, i.c.v.) or idazoxan (4–
16 lg/rat, i.c.v.) 10 min before being subjected to tail
flick test to determine the reaction latency of individual
animal.
Separate groups of rats were treated with imidazoline
I1/a2 receptor agonist moxonidine (25 lg/rat, i.c.v.) or
clonidine (2 lg/rat, i.c.v.) or imidazoline I2 receptor
agonist 2-BFI (10 lg/rat, i.c.v.) or aCSF (2 lL/rat,
i.c.v.) 10 min before i.p. injection of vehicle or subeffec-
tive dose of ethanol (8% w/v; 0.5 g/kg, i.p.) or nicotine
(0.25 mg/kg, i.p.) or saline (1 mL/kg, i.p.). Individual
rat was subjected to tail flick test, and reaction latency
was determined as mentioned earlier.
Effect of imidazoline receptor antagonists on
ethanol- and nicotine-induced antinociception
For the antagonism study, rats were treated with imi-
dazoline I1 receptor antagonist, efaroxan (10 lg/rat,
i.c.v.), or imidazoline I2 receptor antagonist, idazoxan
(4 lg/rat, i.c.v.), or aCSF (2 lL/rat, i.c.v.) 10 min prior
to antinociceptive dose of ethanol (8% w/v; 1 g/kg,
i.p.) or nicotine (0.5 mg/kg, i.p.) or combination of
subeffective dose of ethanol (0.5 g/kg, i.p.) and nicotine
(0.25 mg/kg, i.p.) or saline (1 mL/kg, i.p.).
In combination studies, the same chronological
sequence as described above was applied, that is,
imidazoline receptor antagonist administration was
ª 2013 The Authors Fundamental and Clinical Pharmacology ª 2013 Soci�et�e Franc�aise de Pharmacologie et de Th�erapeutique
Fundamental & Clinical Pharmacology
Ethanol/nicotine antinociception modulated by imidazoline receptor 13
1
2
3
4
5
6
7
8
9
10
11
12
13
14
15
16
17
18
19
20
21
22
23
24
25
26
27
28
29
30
31
32
33
34
35
36
37
38
39
40
41
42
43
44
45
46
47
48
49
50

followed by alcohol and 10 min later by nicotine, and
the tests were conducted 20 min after nicotine admin-
istration. Individual rat was subjected to tail flick test,
and reaction latency was determined.
The doses of endogenous imidazoline binding site
ligands, imidazoline receptor agonists, and antagonists
used here are selected on the basis of preliminary
experiments conducted at our laboratory and available
literature.
Data analysis
Antinociception was calculated as the percentage of
maximum possible effect by the (% MPE) formula: %
MPE = 100 9 (drug time control time/cutoff time con-
trol time). Percentage of MPE values were calculated
for each animal and were statistically analyzed with
one-way analysis of variance (ANOVA). Means were com-
pared with post hoc Dunnett’s or Newman–Keuls test.
P-values < 0.05 were considered significant.
RESULTS
Antinociceptive effect of ethanol and nicotine
As shown in Figure 1a, ethanol (1–2 g/kg, i.p.) dose
dependently increased tail flick latency as compared to
saline-treated group [F(5, 35) = 18.56, P < 0.001]
demonstrating its analgesic potential. Post hoc Dunnett’s
test revealed that ethanol 1, 1.5, and 2 g/kg, i.p.
increased baseline latency by 114% (P < 0.001), 150%
(P < 0.001), and 243% (P < 0.001), respectively. How-
ever, lower doses of ethanol (0.5 g/kg, i.p.) failed to
produce any analgesic effect.
Similarly, administration of nicotine (0.5, 0.75, and
1 mg/kg, i.p.) dose dependently increased baseline tail
flick latency by 141% (P < 0.05), 198% (P < 0.001),
and 222% (P < 0.001), respectively [F(4, 29) = 15.87,
P < 0.001] (Figure 1b). Nicotine at the dose of
0.25 mg/kg, i.p. could not produce significant antinoci-
ception in rats.
Figure 1c depicts the effects executed by combination
of submaximal doses of alcohol (0.5 g/kg, i.p.) and nico-
tine (0.25 mg/kg, i.p.) in the tail flick assay. Subeffective
dose combination of alcohol and nicotine produced sig-
nificant increases in latency time as compared to saline-
treated group [F(3, 21) = 15.76, P < 0.001]. Post hoc
Dunnett’s comparisons indicated the significant potenti-
ation of the antinociceptive effect of per se ineffective
doses of nicotine and ethanol. The doses of nicotine or
ethanol used here did not influenced latency time in tail
flick assay as compared to saline-treated rats.
(a)
(b)
(c)
Figure 1 Antinociceptive effect of ethanol (a), nicotine (b), and
ineffective dose combination of ethanol and nicotine (c) in tail
flick test. Separate group of rats (n = 6) were treated either with
saline (1 ml/kg, i.p.) or ethanol (0.1–2 g/kg), 30 min before or
nicotine (0.1–2 mg/kg, i.p.) 20 min before or ineffective dose
combination of ethanol (0.5 g/kg, i.p.) and nicotine (0.25 mg/kg,
i.p.) at appropriate time before the test and reaction latency was
measured. *P < 0.05, **P < 0.01, ***P < 0.001 vs. respective
control group (one-way ANOVA post hoc Dunnett’s test).
$P < 0.001 vs. ethanol treatment, #P < 0.001 vs. nicotine
treatment (one-way ANOVA post hoc Newman–Keuls test).
ª 2013 The Authors Fundamental and Clinical Pharmacology ª 2013 Soci�et�e Franc�aise de Pharmacologie et de Th�erapeutique
Fundamental & Clinical Pharmacology
4 M.M. Aglawe et al.
1
2
3
4
5
6
7
8
9
10
11
12
13
14
15
16
17
18
19
20
21
22
23
24
25
26
27
28
29
30
31
32
33
34
35
36
37
38
39
40
41
42
43
44
45
46
47
48
49
50
Endogenous imidazoline ligands potentiated
antinociceptive effect of ethanol and nicotine
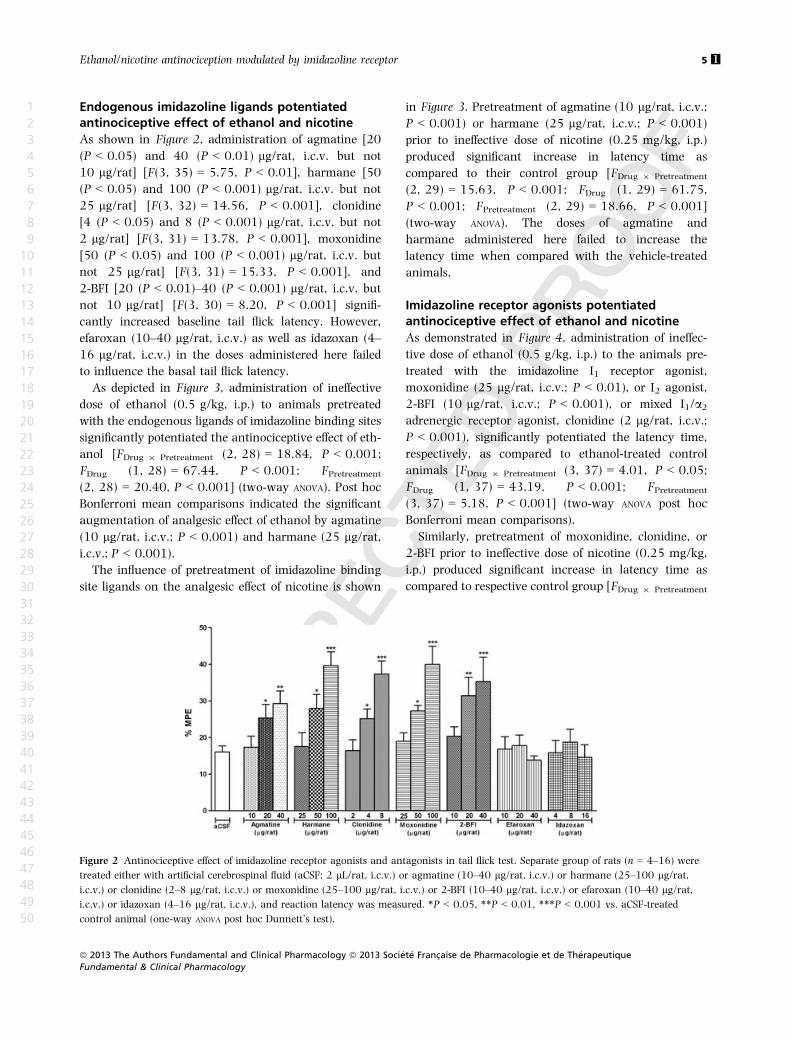
As shown in Figure 2, administration of agmatine [20
(P < 0.05) and 40 (P < 0.01) lg/rat, i.c.v. but not
10 lg/rat] [F(3, 35) = 5.75, P < 0.01], harmane [50
(P < 0.05) and 100 (P < 0.001) lg/rat, i.c.v. but not
25 lg/rat] [F(3, 32) = 14.56, P < 0.001], clonidine
[4 (P < 0.05) and 8 (P < 0.001) lg/rat, i.c.v. but not
2 lg/rat] [F(3, 31) = 13.78, P < 0.001], moxonidine
[50 (P < 0.05) and 100 (P < 0.001) lg/rat, i.c.v. but
not 25 lg/rat] [F(3, 31) = 15.33, P < 0.001], and
2-BFI [20 (P < 0.01)–40 (P < 0.001) lg/rat, i.c.v. but
not 10 lg/rat] [F(3, 30) = 8.20, P < 0.001] signifi-
cantly increased baseline tail flick latency. However,
efaroxan (10–40 lg/rat, i.c.v.) as well as idazoxan (4–
16 lg/rat, i.c.v.) in the doses administered here failed
to influence the basal tail flick latency.
As depicted in Figure 3, administration of ineffective
dose of ethanol (0.5 g/kg, i.p.) to animals pretreated
with the endogenous ligands of imidazoline binding sites
significantly potentiated the antinociceptive effect of eth-
anol [FDrug 9 Pretreatment (2, 28) = 18.84, P < 0.001;
FDrug (1, 28) = 67.44, P < 0.001; FPretreatment
(2, 28) = 20.40, P < 0.001] (two-way ANOVA). Post hoc
Bonferroni mean comparisons indicated the significant
augmentation of analgesic effect of ethanol by agmatine
(10 lg/rat, i.c.v.; P < 0.001) and harmane (25 lg/rat,
i.c.v.; P < 0.001).
The influence of pretreatment of imidazoline binding
site ligands on the analgesic effect of nicotine is shown
in Figure 3. Pretreatment of agmatine (10 lg/rat, i.c.v.;
P < 0.001) or harmane (25 lg/rat, i.c.v.; P < 0.001)
prior to ineffective dose of nicotine (0.25 mg/kg, i.p.)
produced significant increase in latency time as
compared to their control group [FDrug 9 Pretreatment
(2, 29) = 15.63, P < 0.001; FDrug (1, 29) = 61.75,
P < 0.001; FPretreatment (2, 29) = 18.66, P < 0.001]
(two-way ANOVA). The doses of agmatine and
harmane administered here failed to increase the
latency time when compared with the vehicle-treated
animals.
Imidazoline receptor agonists potentiated
antinociceptive effect of ethanol and nicotine
As demonstrated in Figure 4, administration of ineffec-
tive dose of ethanol (0.5 g/kg, i.p.) to the animals pre-
treated with the imidazoline I1 receptor agonist,
moxonidine (25 lg/rat, i.c.v.; P < 0.01), or I2 agonist,
2-BFI (10 lg/rat, i.c.v.; P < 0.001), or mixed I1/a2adrenergic receptor agonist, clonidine (2 lg/rat, i.c.v.;
P < 0.001), significantly potentiated the latency time,
respectively, as compared to ethanol-treated control
animals [FDrug 9 Pretreatment (3, 37) = 4.01, P < 0.05;
FDrug (1, 37) = 43.19, P < 0.001; FPretreatment
(3, 37) = 5.18, P < 0.001] (two-way ANOVA post hoc
Bonferroni mean comparisons).
Similarly, pretreatment of moxonidine, clonidine, or
2-BFI prior to ineffective dose of nicotine (0.25 mg/kg,
i.p.) produced significant increase in latency time as
compared to respective control group [FDrug 9 Pretreatment
Figure 2 Antinociceptive effect of imidazoline receptor agonists and antagonists in tail flick test. Separate group of rats (n = 4–16) were
treated either with artificial cerebrospinal fluid (aCSF; 2 lL/rat, i.c.v.) or agmatine (10–40 lg/rat, i.c.v.) or harmane (25–100 lg/rat,
i.c.v.) or clonidine (2–8 lg/rat, i.c.v.) or moxonidine (25–100 lg/rat, i.c.v.) or 2-BFI (10–40 lg/rat, i.c.v.) or efaroxan (10–40 lg/rat,
i.c.v.) or idazoxan (4–16 lg/rat, i.c.v.), and reaction latency was measured. *P < 0.05, **P < 0.01, ***P < 0.001 vs. aCSF-treated
control animal (one-way ANOVA post hoc Dunnett’s test).
ª 2013 The Authors Fundamental and Clinical Pharmacology ª 2013 Soci�et�e Franc�aise de Pharmacologie et de Th�erapeutique
Fundamental & Clinical Pharmacology
Ethanol/nicotine antinociception modulated by imidazoline receptor 15
1
2
3
4
5
6
7
8
9
10
11
12
13
14
15
16
17
18
19
20
21
22
23
24
25
26
27
28
29
30
31
32
33
34
35
36
37
38
39
40
41
42
43
44
45
46
47
48
49
50

(3, 39) = 3.40, P < 0.05; FDrug (1, 39) = 47.82,
P < 0.001; FPretreatment (3, 39) = 5.78, P < 0.01] (two-
way ANOVA). Post hoc Bonferroni multiple mean compari-
sons with aCSF + nicotine-treated control group
exhibited significant potentiation in % MPE by moxoni-
dine (25 lg/rat, i.c.v.; P < 0.001) or 2-BFI (10 lg/rat,
i.c.v.; P < 0.01) or clonidine (2 lg/rat, i.c.v.;
P < 0.001). Moxonidine, clonidine, and 2-BFI in the
dose used here did not influence the basal latency time
in tail flick assay (Figure 4).
Imidazoline receptor antagonists attenuated
antinociceptive effect of ethanol and nicotine
Two-way ANOVA indicated that pretreatment of rats with
imidazoline I1 antagonist, efaroxan (10 lg/rat, i.c.v.), or
I2 antagonist, idazoxan (4 lg/rat, i.c.v.), significantly
attenuated the ethanol (1 g/kg, i.p.)-induced increased
latency time in tail flick assay [FDrug 9 Pretreatment
(2, 28) = 4.06, P < 0.05; FDrug (1, 28) = 31.11,
P < 0.001; FPretreatment (2, 28) = 3.98, P < 0.05] (Fig-
ure 5). Post hoc Bonferroni multiple comparisons
between the means demonstrated the decreased latency
time in efaroxan (10 lg/rat, i.c.v.) and idazoxan (4 lg/
rat, i.c.v.) pretreated animals indicating the antagonistic
potential of these agents in the antinociceptive effect of
ethanol.
Similar treatment of imidazoline receptor antago-
nists, efaroxan (10 lg/rat, i.c.v.) and idazoxan
(4 lg/rat, i.c.v.), before nicotine (0.5 mg/kg, i.p.) also
antagonized the nicotine-induced increases in latency
[FDrug 9 Pretreatment (2, 28) = 4.77, P < 0.05; FDrug
(1, 28) = 23.99, P < 0.001; FPretreatment (2, 28) =
3.96, P < 0.05] (two-way ANOVA post hoc Bonferroni
Test; Figure 5). Efaroxan or idazoxan administered
alone or with saline did not influence the latency time
in tail flick assay.
Imidazoline receptor antagonists attenuated
synergistic antinociceptive effect of ethanol and
nicotine
As depicted in Figure 6, pretreatment with efaroxan
(10 lg/rat, i.c.v.) or idazoxan (4 lg/rat, i.c.v.) blocked
the potentiated antinociceptive effect elicited by
ineffective dose combination of nicotine (0.25 mg/kg,
i.p.) and ethanol (0.5 g/kg, i.p.) [FDrug 9 Pretreatment
(2, 27) = 8.28, P < 0.01; FDrug (1, 27) = 21.24,
P < 0.001; FPretreatment (2, 27) = 11.36, P < 0.001]
(two-way ANOVA post hoc Bonferroni test).
Figure 3 Potentiation of antinociceptive effect of ethanol and
nicotine by endogenous imidazoline binding site ligands in tail
flick test. Separate group of rats (n = 5–6) were treated either
with artificial cerebrospinal fluid (aCSF; 2 lL/rat, i.c.v.) or
harmane (25 lg/rat, i.c.v.) or agmatine (10 lg/rat, i.c.v.) 10 min
before saline (1 mL/kg, i.p.) or ethanol (0.5 g/kg, i.p.) or nicotine
(0.25 mg/kg, i.p.), and reaction latency was measured 30 min
after alcohol or 20 min after nicotine administration. *P < 0.001
vs. aCSF + saline control group, $P < 0.001 vs. aCSF + ethanol,
#P < 0.001 vs. aCSF + nicotine (two-way ANOVA post hoc
Bonferroni multiple comparison test).
Figure 4 Potentiation of antinociceptive effect of ethanol and
nicotine by imidazoline receptor agonist in tail flick test. Separate
group of rats (n = 4–6) were treated either with artificial
cerebrospinal fluid (aCSF; 2 lL/rat, i.c.v.) or moxonidine (25 lg/
rat, i.c.v.) or 2-BFI (10 lg/rat, i.c.v.) or clonidine (2 lg/rat, i.c.v.)
10 min before saline (1 mL/kg, i.p.), ethanol (0.5 g/kg, i.p.), or
nicotine (0.25 mg/kg, i.p.), and reaction latency was measured
20 min after alcohol or 10 min after nicotine administration.
*, $P < 0.01; **, $$P < 0.01 vs. ethanol/nicotine-treated
respective control group (two-way ANOVA post hoc Bonferroni
multiple comparison test).
ª 2013 The Authors Fundamental and Clinical Pharmacology ª 2013 Soci�et�e Franc�aise de Pharmacologie et de Th�erapeutique
Fundamental & Clinical Pharmacology
6 M.M. Aglawe et al.
1
2
3
4
5
6
7
8
9
10
11
12
13
14
15
16
17
18
19
20
21
22
23
24
25
26
27
28
29
30
31
32
33
34
35
36
37
38
39
40
41
42
43
44
45
46
47
48
49
50

DISCUSS ION
Over the years, it has been a great scientific challenge to
produce centrally acting analgesic without abuse poten-
tial. Currently, opioidergic drugs are the only clinically
available central analgesics. Present study provides
functional evidences for the involvement of imidazoline
binding sites in antinociceptive effects of alcohol and
nicotine. It is well known that alcohol and nicotine
traditionally being used as strong pain-relieving sub-
stances. In present study, we found that ethanol and
nicotine as well as their subeffective combination pro-
duce dose-dependent increase in latency time in tail flick
assay of spinal nociception. These results are in well
agreement with earlier findings of Campbel et al.
[54,55]. Indeed, these drugs are known for their
co-abuse potential and share common pathways for
their reward as well as analgesic property [36,37]. Con-
siderable efforts are being directed toward developing
more effective therapy for treatment of alcoholism and
nicotine addiction. The challenge is compounded by
conditions of comorbid dependence on alcohol and nico-
tine [56]. The 12imidazoline receptor binding sites are of
particular relevance to the comorbid occurrence of alco-
holism and heavy smoking. It is intimately involved in
perception and modulation of pain, while it plays major
role in reward [57]. It is not surprising therefore that
the rewarding and antinociceptive effects of alcohol and
nicotine, either individually or combined, may be closely
intertwined with the endogenous imidazoline system.
Several clinical and preclinical studies have demon-
strated therapeutic potential of imidazoline binding site
ligands [58,59]. Interestingly, several behavioral effects
of ethanol and nicotine including conditioned hyperlo-
comotion, anhedonia, anxiety, withdrawal, locomotor
sensitization, intake, and development of dependence or
withdrawal syndrome are found to be associated with
imidazoline binding sites [38,40,49,50,60–66]. Immu-
nocytochemical studies suggest that imidazoline bind-
ing sites are expressed in brain regions involved in
pain perception and response to painful stimuli [41].
Further I2 sites can be detected in spinal cord and
implicated in modulation of nociceptive processing
[67]. However, it remains unknown as to whether they
enhanced or inhibit nociception. In view of this, pres-
ent study investigated the effect of endogenous imidaz-
oline ligands on antinociceptive effect of ethanol and
nicotine in tail flick assay representing spinal
nociception. Agmatine and harmane are the potent
endogenous ligands that activate imidazoline I1 and I2
Figure 5 Influence of imidazoline receptor antagonists on the
antinociceptive effect executed by ethanol and nicotine in tail flick
test. Separate group of rats (n = 6) were treated either with
artificial cerebrospinal fluid (aCSF; 2 lL/rat, i.c.v.) or idazoxan
(4 lg/rat, i.c.v.) or efaroxan (10 lg/rat, i.c.v.) 10 min before saline
(1 mL/kg, i.p.), ethanol (1 g/kg, i.p.), or nicotine (0.5 mg/kg, i.p.),
and reaction latency was measured 20 min after alcohol or 10 min
after nicotine administration. *P < 0.001 vs. vehicle control group,
$P < 0.01 vs. alcohol-treated group, # 11vs. nicotine-treated
group (two-way ANOVA post hoc Bonferroni multiple comparison
test).
Figure 6 Influence of imidazoline receptor antagonists on the
antinociceptive effect executed by noneffective dose combination
of ethanol and nicotine in tail flick test. Separate group of rats
(n = 6) were treated either with artificial cerebrospinal fluid
(aCSF; 2 lL/rat, i.c.v.) or idazoxan (4 lg/rat, i.c.v.) or efaroxan
(10 lg/rat, i.c.v.) 10 min before saline (1 mL/kg, i.p.) or ethanol
(0.5 g/kg, i.p.) and nicotine (0.25 mg/kg, i.p.), and reaction
latency was measured 20 min after alcohol or 10 min after
nicotine administration. #P < 0.001 vs. vehicle control group,
*P < 0.001 vs. alcohol- and nicotine-treated group (two-way
ANOVA post hoc Bonferroni multiple comparison test).
ª 2013 The Authors Fundamental and Clinical Pharmacology ª 2013 Soci�et�e Franc�aise de Pharmacologie et de Th�erapeutique
Fundamental & Clinical Pharmacology
Ethanol/nicotine antinociception modulated by imidazoline receptor 17
1
2
3
4
5
6
7
8
9
10
11
12
13
14
15
16
17
18
19
20
21
22
23
24
25
26
27
28
29
30
31
32
33
34
35
36
37
38
39
40
41
42
43
44
45
46
47
48
49
50

receptor. It is important to note that these agents
themselves exhibit antinociceptive profile in neuropath-
ic pain [68,69]. Interestingly, we found that pretreat-
ment of these agents potentiated the antinociceptive
effect of ethanol as well as nicotine. This suggests that
imidazoline binding sites act as common neural sub-
strate responsible for analgesic effect of alcohol and
nicotine. Moreover, evidences also suggest the involve-
ment of imidazoline binding sites in addiction liability
[38–40]. In our earlier study, we also demonstrated
the involvement of imidazoline binding sites in ethanol
withdrawal anxiety [50]. Moreover, agmatine, an
endogenous receptor ligand attenuated the develop-
ment as well as expression of nicotine, induced locomo-
tor sensitization [49]. This is interesting in view of the
therapeutic potential of imidazoline agents to counter-
act the comorbidity of alcohol and nicotine.
It is also reported that several imidazoline receptor
agonists including moxonidine, clonidine, 2-BFI, and
BU-224 possess analgesic property [67,70]. Hence to
confirm above finding, we observed the effect of imidaz-
oline receptor agonists on antinociceptive potential of
ethanol and nicotine. We found that clonidine, mixed
agonist of imidazoline I1/adrenergic a2 receptor, moxo-
nidine, imidazoline I1 receptor agonist, and 2-BFI,
imidazoline I2 receptor agonist, significantly increased
the antinociceptive effect of nicotine and ethanol. Con-
versely, antinociceptive effect of ethanol and nicotine
or their subeffective dose combination was significantly
inhibited by pretreatment with imidazoline I1 antago-
nist, efaroxan, as well as imidazoline I2/adrenergic a2receptor antagonist, idazoxan.
Thus, our result clearly suggests the involvement of
imidazoline I1 and I2 receptors in analgesic effects of
ethanol and nicotine. However, most of the agents
used in this study like clonidine, agmatine, and idazo-
xan possess moderate affinity toward a2 adrenergic
receptors. Further analgesic property of clonidine and
agmatine has been linked to a2 adrenergic receptors
modulation [68]. Thus, possible involvement of a2adrenergic receptors in antinociceptive effect of ethanol
and nicotine needs further investigation. Moreover,
agmatine and harmane also act on NMDA [71,72] and
GABAA receptors [73], respectively, and hence the con-
tribution of these targets could not be ruled out and
needs further investigation.
The antinociceptive activity of nicotine or alcohol
has been linked to their co-abuse [36,37]. Therefore, it
can be inferred from the present study that imidazoline
agents may act as important therapeutic molecules for
the treatment of alcohol and nicotine addiction as well
as their co-abuse potential. However, extensive bio-
chemical and pharmacological studies are required to
prove this hypothesis.
CONFL ICT OF INTEREST
All the authors report no conflict of interest.
REFERENCES
1 Olsen J., Sabreo S., Fasting U. Interaction of alcohol and
tobacco as risk factors in cancer of the laryngeal region.
J. Epidemiol. Community Health (1985) 39 165–168.
2 Johnson K.A., Jennison K.M. The drinking–smoking syndrome
and social context. Int. J. Addict. (1992) 27 749–792.
3 Castellsague X., Munoz N., Stefani D.E., Victoria C.G.,
Quintana M.J. Independent and joint effects of tobacco
smoking and alcohol drinking on the risk of oesophageal
cancer in men and women. Int. J. Cancer (1999) 82
657–664.
4 Yoshida K., Engel J., Liljequist S. The effect of chronic ethanol
administration of high affinity 3H-nicotinic binding in rat
brain. Naunyn Schmiedebergs Arch. Pharmacol. (1982) 321
74–76.
5 Signs S.A., Schechter M.D. Nicotine-induced potentiation of
ethanol discrimination. Pharmacol. Biochem. Behav. (1986)
24 769–771.
6 Booker T.K., Collins A.C. Long-term ethanol treatment elicits
changes in nicotinic receptor binding in only a few brain
regions. Alcohol (1997) 14 131–140.
7 Penland S., Hoplight B., Obernier J., Crews F.T. Effects of
nicotine on ethanol dependence and brain damage. Alcohol
(2001) 24 45–54.
8 Howard L.A., Ahluwalia J.S., Lin S.K., Sellers E.M., Tyndale
R.F. CYP2E1_1D regulatory polymorphism: association with
alcohol and nicotine dependence. Pharmacogenetics (2003)
13 321–328.
9 Howard L.A., Miksys S., Hoffmann E., Mash D., Tyndale R.F.
Brain CYP2E1 is induced by nicotine and ethanol in rat and
is higher in smokers and alcoholics. Br. J. Pharmacol. (2003)
138 1376–1386.
10 Rose J.E., Brauer L.H., Behm F.M., Cramblett M., Calkins
K., Lawhon D. Psychopharmacological interactions between
nicotine and ethanol. Nicotine Tob. Res. (2004) 6 133–
144.
11 Korkosz A., Zatorski P., Taracha E., Plaznik A., Kostowski W.,
Bienkowski P. Ethanol blocks nicotine-induced seizures in
mice: comparison with midazolam and baclofen. Alcohol
(2006) 40 151–157.
12 Korkosz A., Zatorski P., Taracha E., Plaznik A., Kostowski W.,
Bienkowski P. Effects of ethanol on nicotine-induced
conditioned place preference in C57BL/6J mice. Prog.
Neuropsychopharmacol. Biol. Psychiatry (2006) 30 1283–
1290.
ª 2013 The Authors Fundamental and Clinical Pharmacology ª 2013 Soci�et�e Franc�aise de Pharmacologie et de Th�erapeutique
Fundamental & Clinical Pharmacology
8 M.M. Aglawe et al.
1
2
3
4
5
6
7
8
9
10
11
12
13
14
15
16
17
18
19
20
21
22
23
24
25
26
27
28
29
30
31
32
33
34
35
36
37
38
39
40
41
42
43
44
45
46
47
48
49
50

13 Korkosz A., Scinska A., Taracha E. et al. Nicotine-induced
conditioned taste aversion in the rat: effects of ethanol.
Eur. J. Pharmacol. (2006) 537 99–105.
14 Parnell S.E., West J.R., Chen W.J. Nicotine decreases blood
alcohol concentrations in adult rats: a phenomenon
potentially related to gastric function. Alcohol. Clin. Exp. Res.
(2006) 30 1408–1413.
15 King A., McNamara P., Conrad M., Cao D. Alcohol-induced
increases in smoking behavior for nicotinized and
denicotinized cigarettes in men and women.
Psychopharmacology (2009) 207 107–117.
16 Ribeiro-Carvalho A., Lima C.S., Medeiros A.H. et al.
Combined exposure to nicotine and ethanol in adolescent
mice: effects on the central cholinergic systems during short
and long term withdrawal. Neuroscience (2009) 162 1174–
1186.
17 Connor J.P., Young R.M., Lawford B.R., Saunders J.B., Ritchie
T.L., Noble E.P. Heavy nicotine and alcohol use in alcohol
dependence is associated with D2 dopamine receptor (DRD2)
polymorphism. Addict. Behav. (2006) 32 310–319.
18 Volk H.E., Scherrer J.F., Bucholz K.K. et al. Evidence for
specificity of transmission of alcohol and nicotine dependence
in an offspring of twins design. Drug Alcohol Depend. (2006)
87 225–232.
19 Le D., Li Z., Funk D., Shram M., Li T.K., Shaham Y. Increased
vulnerability to nicotine self-administration and relapse in
alcohol-naive offspring of rats selectively bred for high alcohol
intake. J. Neurosci. (2006) 26 1872–1879.
20 Rezvani A.H., Levin E.D. Nicotine-alcohol interactions and
cognitive function in rats. Pharmacol. Biochem. Behav.
(2002) 72 865–872.
21 Al-Rajaie S., Dar M.S. Behavioral interaction between
nicotinic and ethanol: possible modulation by mouse
cerebellar glutamate. Alcohol. Clin. Exp. Res. (2006)
30 1223–1233.
22 Al-Rajaie S., Dar M.S. Antagonism of ethanol ataxia by
intracerebellar nicotine: possible modulation by mouse
cerebellar nitric oxide and cGMP. Brain Res. Bull. (2006)
69 187–196.
23 Ceballos N.A., Tivis R., Lawton-Craddock A., Nixond S.J.
Nicotine and cognitive efficiency in alcoholics and elicit
stimulant abusers: implication of smoking cessation for
substance users in treatment. Subst. Use Misuse (2006)
41 265–268.
24 Tizabi Y., Copeland R.L. Jr, Louis V.A., Taylor R.E. Effects of
combined systemic alcohol and central nicotine
administration into VTA on dopamine release in the nucleus
accumbens. Alcohol. Clin. Exp. Res. (2002) 26 394–399.
25 Tizabi Y., Manaye K.F., Smoot D.T., Taylor R.E. Nicotine
inhibits ethanol-induced toxicity in cultured cerebral cortical
cells. Neurotox. Res. (2004) 6 311–316.
26 Cutter H.S., O’Farell T.J. Experience with alcohol and the
endogenous opioid system in ethanol analgesia. Addict.
Behav. (1987) 12 331–343.
27 Woodrow K.M., Eltherington L.G. Feeling no pain: alcohol as
an analgesic. Pain (1988) 32 159–163.
28 Boada J., Feria M., Sanz E. Inhibitory effect of naloxone on
ethanol induced antinociception in mice. Pharmacol. Res.
Commun. (1981) 13 673–678.
29 Fertig J.B., Pomerleau O.F., Sanders B. Nicotine-produced
antinociception in minimally deprived smokers and ex-
smokers. Addict. Behav. (1986) 11 239–248.
30 Pomerleau O.F., Turk D.C., Fertig J.B. The effects of cigarette
smoking on pain and anxiety. Addict. Behav. (1984) 9 265–
271.
31 Phan D.V., Doda M., Bite A., Gyorgy L. Antinociceptive activity
of nicotine. Acta Physiol. Acad. Sci. (1973) 44 85–93.
32 Damaj M.I., Welech S.P., Martin B.R. Involvement of calcium
and L-type channels in nicotine-induced antinociception.
J. Pharmacol. Exp. Ther. (1993) 266 1330–1338.
33 Khan I.M., Buerkle H., Taylor P., Yaksh T.L. Nociceptive and
antinociceptive responses to intrathecally administered
nicotinic agonists. Neuropharmacology (1998) 37 1515–
1525.
34 Simons C.T., Cuellar J.M., Moore J.A. et al. Nicotinic receptor
involvement in antinociception induced by exposure to
cigarette smoke. J. Neurosci. (2005) 26 1872–1879.
35 Vincler M. Neuronal nicotinic receptors as targets for novel
analgesics. Expert Opin. Investig. Drugs (2005) 14 1191–
1198.
36 Franklin K.B.J. Analgesia and the neural substrate of reward.
Neurosci. Biobehav. Rev. (1989) 13 149–154.
37 Franklin K.B.J. Analgesia and abuse potential: an accidental
association or a common substrate? Pharmacol. Biochem.
Behav. (1998) 59 993–1002.
38 Uzbay I.T., Yesilyurt O., Celik T., Ergun H., Isimer A. Effects
of agmatine on ethanol withdrawal syndrome in rats. Behav.
Brain Res. (2000) 107 153–159.
39 Gentili F., Cardinaletti C., Carrieri A. et al. Involvement of
I2-imidazoline binding sites in positive and negative morphine
analgesia modulatory effects. Eur. J. Pharmacol. (2006) 553
73–81.
40 Lewis B., Wellmann K., Barron S. Agmatine reduces balance
deficits in a rat model of third trimester binge-like alcohol
exposure. Pharmacol. Biochem. Behav. (2007) 88 114–121.
41 Ruggiero D.A., Regunathan S., Wang H., Milner T.A., Reis
D.J. Immunocytochemical localization of an imidazoline
receptor protein in the central nervous system. Brain Res.
(1998) 780 270–293.
42 Michel M.C., Insel P.A. Are there multiple imidazoline binding
sites? Trends Pharmacol. Sci. (1989) 10 342–344.
43 Michel M.C., Ernsberger P. Keeping an eye on the I site:
imidazoline-preferring receptors. Trends Pharmacol. Sci.
(1992) 13 369–370.
44 Parini A., Moudanos C.G., Pizzinat N., Lanier S.M. The elusive
family of imidazoline binding sites. Trends Pharmacol. Sci.
(1996) 17 13–16.
45 Eglen R.M., Hudson A.L., Kendall D.A. et al. ‘Seeing through
a glass darkly’: casting light on imidazoline ‘I’ sites. Trends
Pharmacol. Sci. (1998) 19 381–390.
46 De Vos H., Bricca G., De Keyser J., De Backer J.P., Bousquet
P., Vauquelin G. Imidazoline receptors, non-adrenergic
ª 2013 The Authors Fundamental and Clinical Pharmacology ª 2013 Soci�et�e Franc�aise de Pharmacologie et de Th�erapeutique
Fundamental & Clinical Pharmacology
Ethanol/nicotine antinociception modulated by imidazoline receptor 19
1
2
3
4
5
6
7
8
9
10
11
12
13
14
15
16
17
18
19
20
21
22
23
24
25
26
27
28
29
30
31
32
33
34
35
36
37
38
39
40
41
42
43
44
45
46
47
48
49
50

idazoxan binding sites and alpha 2-adrenoceptors in the
human central nervous system. Neuroscience (1994) 59
589–598.
47 Tesson F., Prip-Buus C., Lemoine A., Pegorier J.P., Parini A.
Subcellular distribution of imidazoline-guanidinium-receptive
sites in human and rabbit liver. Major localization to the
mitochondrial outer membrane. J. Biol. Chem. (1991) 266
155–160.
48 Raddatz R., Savic S.L., Bakthavachalam V. et al. Imidazoline-
binding domains on monoamine oxidase B and
subpopulations of enzyme. J. Pharmacol. Exp. Ther. (2000)
292 1135–1145.
49 Kotagale N.R., Taksande B.G., Gahane A.Y., Ugale R.R.,
Chopde C.T. Repeated agmatine treatment attenuates nicotine
sensitization in mice: modulation by alpha2-adrenoceptors.
Behav. Brain Res. (2010) 213 161–174.
50 Taksande B.G., Kotagale N.R., Patel M.R., Shelkar G.P., Ugale
R.R., Chopde C.T. Agmatine, an endogenous imidazoline
receptor ligand modulates ethanol anxiolysis and withdrawal
anxiety in rats. Eur. J. Pharmacol. (2010) 637 89–101.
51 Paxinos G., Watson C. The rat brain in stereotaxic
co-ordinates, Elsevier Academic Press, Amsterdam, 2005.
52 Kamei J., Sodeyama M., Tsuda M., Suzuki T., Nagase H.
Antinociceptive effect of buprenorphine in mu1-opioid
receptor deficient CXBK mice. Life Sci. (1997) 60 333–337.
53 Kamei J., Zushida K. Effect of mexiletine on thermal allodynia
and hyperalgesia in diabetic mice. Jpn. J. Pharmacol. (2000)
84 89–92.
54 Campbel V.C., Taylor R.E., Tizabi Y. Effects of selective opioid
receptor antagonists on alcohol-induced and nicotine-induced
antinociception. Alcohol. Clin. Exp. Res. (2007) 31 1435–
1440.
55 Campbel V.C., Taylor R.E., Tizabi Y. Antinociceptive effect of
alcohol and nicotine: involvement of the opioid system. Brain
Res. (2006) 1097 71–77.
56 Meyerhoff D.J., Tizabi Y., Staley J.K., Durazzo T.C., Glass J.M.,
Nixon S.J. Smoking comorbidity in alcoholism:
neurobiological and neurocognitive consequences. Alcohol.
Clin. Exp. Res. (2006) 30 253–264.
57 Ruiz-Durantez E., Torrecilla M., Pineda J., Ugedo L.
Attenuation of acute and chronic effects of morphine by the
imidazoline receptor ligand 2-(2-benzofuranyl)-2-imidazoline
in rat locus coeruleus neurons. Br. J. Pharmacol. (2003) 138
494–500.
58 Head G.A., Mayorov D.N. Imidazoline receptors, novel agents
and therapeutic potential. Cardiovasc. Hematol. Agents Med.
Chem. (2006) 4 17–32.
59 Li J.X., Zhang Y. Imidazoline I2 receptors: target for new
analgesics? Eur. J. Pharmacol. (2011) 658 49–56.
60 Rommelspacher H., May T., Susilo R. Beta-carbolines and
tetrahydroisoquinolines: detection and function in mammals.
Planta Med. (1991) 57 S85–S92.
61 Rommelspacher H., Dufeu P., Schmidt L.G. Harman and
norharman in alcoholism: correlations with psychopathology
and long-term changes. Alcohol. Clin. Exp. Res. (1996) 20
3–8.
62 Mao L., Abdel-Rahman A.A. Synergistic behavioral
interaction between ethanol and clonidine in rats: role of
alpha-2 adrenoceptors. J. Pharmacol. Exp. Ther. (1996) 279
443–449.
63 Spies C.D., Dubisz N., Neumann T. Therapy of alcohol
withdrawal syndrome in intensive care unit patients following
trauma. Results of a prospective randomised trial. Crit. Care
Med. (1996) 24 414–422.
64 Dobrydnjov I., Axelsson K., Berggren L., Samarutel J.,
Holmstrom B. Intrathecal and oral clonidine as prophylaxis
for postoperative alcohol withdrawal syndrome: a randomized
double-blinded study. Anesth. Analg. (2004) 98 738–744.
65 Zaniewska M., McCreary A.C., Sezer G., Przegalinski E., Filip
M. Effects of agmatine on nicotine-evoked behavioral
responses in rats. Pharmacol. Rep. (2008) 60 645–654.
66 Semenova S., Markou A. The alpha2 adrenergic receptor
antagonist idazoxan, but not the serotonin-2A receptor
antagonist M100907, partially attenuated reward deficits
associated with nicotine, but not amphetamine, withdrawal
in rats. Eur. Neuropsychopharmacol. (2010) 20 731–746.
67 Diaz A., Mayet S., Dickenson A.H. BU-224 produces spinal
antinociception as an agonist at imidazoline I2 receptors. Eur.
J. Pharmacol. (1997) 333 9–15.
68 Yesilyurt O., Uzbay I.T. Agmatine potentiates the analgesic
effect of morphine by an alpha(2)-adrenoceptor-mediated
mechanism in mice. Neuropsychopharmacol. J. (2001) 25
98–103.
69 Aricioglu F., Korcegez E., Ozyalcin S. Effect of harmane on
mononeuropathic pain in rats. Ann. N. Y. Acad. Sci. (2003)
1009 180–184.
70 Fairbanks C.A., Posthumus I.J., Kitto K.F., Stone L.S., Wilcox
G.L. Monoxidine, a selective imidazoline/alpha2 adrenergic
receptor agonist, synergizes with morphine and deltorphin 2
to inhibit substance P- induced behaviour in mice. Pain
(2000) 84 13–20.
71 Yang X.C., Reis D.J. Agmatine selectively blocks the NMDA
subclass of glutamate receptor channels in cultured mouse
hippocampal neurons. J. Pharmacol. Exp. Ther. (1999) 288
544–549.
72 Olmos G., DeGregorio-Rocasolano N., Paz Regalado M.,
Gasull T., Assumpcio Boronat M., Trullas R. Protection by
imidazol(ine) drugs and agmatine of glutamate-induced
neurotoxicity in cultured cerebellar granule cells through
blockade of NMDA receptor. Br. J. Pharmacol. (1999) 127
1317–1326.
73 Komissarov I.V., Abramets I.I. Characteristics of GABA-
potentiating effect of harmane. Biull. Eksp. Biol. Med. (1982)
94 61–63.
ª 2013 The Authors Fundamental and Clinical Pharmacology ª 2013 Soci�et�e Franc�aise de Pharmacologie et de Th�erapeutique
Fundamental & Clinical Pharmacology
10 M.M. Aglawe et al.
1
2
3
4
5
6
7
8
9
10
11
12
13
14
15
16
17
18
19
20
21
22
23
24
25
26
27
28
29
30
31
32
33
34
35
36
37
38
39
40
41
42
43
44
45
46
47
48
49
50

Author Query Form
Journal: FCP
Article: 12034
Dear Author,
During the copy-editing of your paper, the following queries arose. Please respond to these by marking up
your proofs with the necessary changes/additions. Please write your answers on the query sheet if there is
insufficient space on the page proofs. Please write clearly and follow the conventions shown on the attached
corrections sheet. If returning the proof by fax do not write too close to the paper’s edge. Please remember
that illegible mark-ups may delay publication.
Many thanks for your assistance.
Query reference Query Remarks
1 AUTHOR: Please check the edit made in the running title.
2 AUTHOR: Please identify and encircle the forename and surname of all
authors.
3 AUTHOR: Please check the edit made in the sentence ‘Antinociceptive effect of
ethanol and…’.
4 AUTHOR: Please check keywords.
5 AUTHOR: Please give address information for ‘CPCSEA’: town.
6 AUTHOR: Please give address information for ‘Sigma-Aldrich Co.’: town, state.
7 AUTHOR: Please give address information for ‘Tocris Biosciences’: town.
8 AUTHOR: Please give address information for ‘David Kopf Instruments’: town.
9 AUTHOR: Please give address information for ‘plastic UK’: town and country.
10 AUTHOR: Please give address information for ‘INCO’: town.
11 AUTHOR: Please define ‘#’ in Figure 5.
12 AUTHOR: Please check the edit made in the sentence ‘The imidazoline receptor
binding sites are of…’.

O n c e y o u h a v e A c r o b a t R e a d e r o p e n o n y o u r c o m p u t e r , c l i c k o n t h e C o m m e n t t a b a t t h e r i g h t o f t h e t o o l b a r :
S t r i k e s a l i n e t h r o u g h t e x t a n d o p e n s u p a t e x tb o x w h e r e r e p l a c e m e n t t e x t c a n b e e n t e r e d .‚ H i g h l i g h t a w o r d o r s e n t e n c e .‚ C l i c k o n t h e R e p l a c e ( I n s ) i c o n i n t h e A n n o t a t i o n ss e c t i o n .‚ T y p e t h e r e p l a c e m e n t t e x t i n t o t h e b l u e b o x t h a ta p p e a r s .
T h i s w i l l o p e n u p a p a n e l d o w n t h e r i g h t s i d e o f t h e d o c u m e n t . T h e m a j o r i t y o ft o o l s y o u w i l l u s e f o r a n n o t a t i n g y o u r p r o o f w i l l b e i n t h e A n n o t a t i o n s s e c t i o n ,p i c t u r e d o p p o s i t e . W e ’ v e p i c k e d o u t s o m e o f t h e s e t o o l s b e l o w :S t r i k e s a r e d l i n e t h r o u g h t e x t t h a t i s t o b ed e l e t e d .
‚ H i g h l i g h t a w o r d o r s e n t e n c e .‚ C l i c k o n t h e S t r i k e t h r o u g h ( D e l ) i c o n i n t h eA n n o t a t i o n s s e c t i o n .
H i g h l i g h t s t e x t i n y e l l o w a n d o p e n s u p a t e x tb o x w h e r e c o m m e n t s c a n b e e n t e r e d .‚ H i g h l i g h t t h e r e l e v a n t s e c t i o n o f t e x t .‚ C l i c k o n t h e A d d n o t e t o t e x t i c o n i n t h eA n n o t a t i o n s s e c t i o n .‚ T y p e i n s t r u c t i o n o n w h a t s h o u l d b e c h a n g e dr e g a r d i n g t h e t e x t i n t o t h e y e l l o w b o x t h a ta p p e a r s .
M a r k s a p o i n t i n t h e p r o o f w h e r e a c o m m e n tn e e d s t o b e h i g h l i g h t e d .‚ C l i c k o n t h e A d d s t i c k y n o t e i c o n i n t h eA n n o t a t i o n s s e c t i o n .‚ C l i c k a t t h e p o i n t i n t h e p r o o f w h e r e t h e c o m m e n ts h o u l d b e i n s e r t e d .‚ T y p e t h e c o m m e n t i n t o t h e y e l l o w b o x t h a ta p p e a r s .

I n s e r t s a n i c o n l i n k i n g t o t h e a t t a c h e d f i l e i n t h ea p p r o p r i a t e p a c e i n t h e t e x t .‚ C l i c k o n t h e A t t a c h F i l e i c o n i n t h e A n n o t a t i o n ss e c t i o n .‚ C l i c k o n t h e p r o o f t o w h e r e y o u ’ d l i k e t h e a t t a c h e df i l e t o b e l i n k e d .‚ S e l e c t t h e f i l e t o b e a t t a c h e d f r o m y o u r c o m p u t e ro r n e t w o r k .‚ S e l e c t t h e c o l o u r a n d t y p e o f i c o n t h a t w i l l a p p e a ri n t h e p r o o f . C l i c k O K .
I n s e r t s a s e l e c t e d s t a m p o n t o a n a p p r o p r i a t ep l a c e i n t h e p r o o f .‚ C l i c k o n t h e A d d s t a m p i c o n i n t h e A n n o t a t i o n ss e c t i o n .‚ S e l e c t t h e s t a m p y o u w a n t t o u s e . ( T h e A p p r o v e ds t a m p i s u s u a l l y a v a i l a b l e d i r e c t l y i n t h e m e n u t h a ta p p e a r s ) .‚ C l i c k o n t h e p r o o f w h e r e y o u ’ d l i k e t h e s t a m p t oa p p e a r . ( W h e r e a p r o o f i s t o b e a p p r o v e d a s i t i s ,t h i s w o u l d n o r m a l l y b e o n t h e f i r s t p a g e ) .
A l l o w s s h a p e s , l i n e s a n d f r e e f o r m a n n o t a t i o n s t o b e d r a w n o n p r o o f s a n d f o rc o m m e n t t o b e m a d e o n t h e s e m a r k s . .‚ C l i c k o n o n e o f t h e s h a p e s i n t h e D r a w i n gM a r k u p s s e c t i o n .‚ C l i c k o n t h e p r o o f a t t h e r e l e v a n t p o i n t a n dd r a w t h e s e l e c t e d s h a p e w i t h t h e c u r s o r .‚
T o a d d a c o m m e n t t o t h e d r a w n s h a p e ,m o v e t h e c u r s o r o v e r t h e s h a p e u n t i l a na r r o w h e a d a p p e a r s .‚
D o u b l e c l i c k o n t h e s h a p e a n d t y p e a n yt e x t i n t h e r e d b o x t h a t a p p e a r s .