Embed Size (px)
Citation preview

Synergistic Anti-Myeloma Activity of the Proteasome Inhibitor Marizomib and the IMiD® Immunomodulatory Drug Pomalidomide
Deepika Sharma Das1, Arghya Ray1, Yan Song1, Paul Richardson1, Mohit Trikha2, Dharminder Chauhan1,¶,*, and Kenneth C. Anderson1,¶,*
1LeBow Institute for Myeloma Therapeutics and Jerome Lipper Myeloma Center, Department of Medical Oncology, Dana-Farber Cancer Institute, Harvard Medical School, Boston, MA
2Triphase Accelerator, San Diego, CA
Abstract
The proteasome inhibitor bortezomib is an effective therapy for the treatment of relapsed and
refractory multiple myeloma (RRMM); however, prolonged treatment can be associated with
toxicity, peripheral neuropathy and drug resistance. Our earlier studies showed that the novel
proteasome inhibitor marizomib is distinct from bortezomib in its chemical structure, mechanisms
of action and effects on proteasomal activities, and that it can overcome bortezomib resistance.
Pomalidomide, like lenalidomide, has potent immunomodulatory activity and has been approved
by the US Food and Drug Administration for the treatment of RRMM. Here, we demonstrate that
combining low concentrations of marizomib with pomalidomide induces synergistic anti-MM
activity. Marizomib plus pomalidomide-induced apoptosis is associated with: 1) activation of
caspase-8, caspase-9, caspase-3 and PARP cleavage; 2) downregulation of cereblon (CRBN),
IRF4, MYC and MCL1; and 3) suppression of chymotrypsin-like, caspase-like, and trypsin-like
proteasome activities. CRBN-siRNA attenuates marizomib plus pomalidomide-induced MM cells
death. Furthermore, marizomib plus pomalidomide inhibits the migration of MM cells and
tumour-associated angiogenesis, as well as overcomes cytoprotective effects of bone marrow
microenvironment. In human MM xenograft model studies, the combination of marizomib and
pomalidomide is well tolerated, inhibits tumour growth and prolongs survival. These preclinical
studies provide the rationale for on-going clinical trials of combined marizomib and
pomalidomide to improve outcome in patients with RRMM.
¶Address correspondence: Kenneth C. Anderson and Dharminder Chauhan, Dana-Farber Cancer Institute, Dept. of Medicine, Harvard Medical School, 44 Binney Street, Mayer 557, Boston, MA 02115, USA, Tel: +1 617-632-2144, Fax: +1 617-632-6140, [email protected]; [email protected].*Joint Senior authorship
Authors’ contributions: DSD performed the experiments, interpreted data and wrote the manuscript; YS and AR helped in animal studies; PR provided clinical samples; MT reviewed the manuscript; DC designed research, analysed data and wrote the manuscript; KCA analysed data and wrote the manuscript.
Conflict-of-interest disclosure MT is an employee of Triphase Accelerator; KCA is on Advisory board of Celgene, Onyx, Gilead, Sonofi Aventis and is a Scientific founder of Acetylon and Oncopep; DC is consultant to Triphase Accelerator. The remaining authors declare no competing financial interests.
Supplementary information is available online.
HHS Public AccessAuthor manuscriptBr J Haematol. Author manuscript; available in PMC 2016 December 01.
Published in final edited form as:Br J Haematol. 2015 December ; 171(5): 798–812. doi:10.1111/bjh.13780.
Author M
anuscriptA
uthor Manuscript
Author M
anuscriptA
uthor Manuscript

Keywords
Marizomib; Pomalidomide; Multiple myeloma; cancer and drug therapy
Introduction
The ubiquitin proteasome pathway (UPP) is a validated therapeutic target in multiple
myeloma (MM), evidenced by the US Food and Drug Administration (FDA) approval of
bortezomib (Velcade™) and carfilzomib (Kyprolis™).(Kane, et al 2003, Richardson, et al
2003, Richardson, et al 2005, Siegel, et al 2012, Vij, et al 2012a, Vij, et al 2012b) Even
though bortezomib and carfilzomib therapies are major advances, they are associated with
possible off-target toxicities and the development of drug-resistance.(Atrash, et al 2015, Cai,
et al 2014, Harvey 2014, Huber, et al 2015, Lonial, et al 2005, Richardson, et al 2006,
Wanchoo, et al 2014) Our previous studies showed that the novel proteasome inhibitor
marizomib(Feling, et al 2003) is distinct from bortezomib and triggers apoptosis even in
MM cells resistant to bortezomib therapies.(Chauhan, et al 2005a) These preclinical data
provided the basis for the on-going phase-1 clinical trials of marizomib in patients with
relapsed/refractory MM (RRMM).(Potts, et al 2011, Richardson, et al 2011) In addition, we
showed that the combination of marizomib with the immunomodulatory agent lenalidomide
induces synergistic anti-MM activity.(Chauhan, et al 2010)
Pomalidomide, like lenalidomide, is a thalidomide analogue with potent immunomodulatory
activity. Based on increased progression-free survival(Gras 2013, Richardson, et al 2013,
Richardson, et al 2014), pomalidomide has been approved by the FDA for the treatment of
patients with RRMM who have received at least two prior therapies, including lenalidomide
and bortezomib, and who showed disease progression on or within 60 days of completion of
the most recent therapy.(Gras 2013, Richardson, et al 2013, Richardson, et al 2014)
In the present study, we characterize the effects of the combination of marizomib and
pomalidomide treatment against MM cell lines and primary patient cells resistant to
conventional and novel therapies. Both in vitro models and in vivo MM xenograft models
demonstrate that marizomib plus pomalidomide trigger synergistic anti-MM activity and
overcome drug resistance. Our preclinical studies support the continuation of clinical trials
of combined marizomib and pomalidomide to improve outcome in patients with RRMM.
Materials and methods
Cell culture and reagents
Human MM cell lines MM.1S, MM.1R, INA-6, ARP-1, RPMI-8226, DOX40, LR5,
ANBL-6.WT (wild type), and ANBL-6-bortezomib-resistant (ANBL-6.BR), as well as
peripheral blood mononuclear cells (PBMCs) from normal healthy donors, were cultured in
RPMI-1640 medium supplemented with complete medium (10% fetal bovine serum, 100
units/ml penicillin, 100 μg/ml streptomycin, and 2 mM L-glutamine) at 37°C and 5% CO2.
ANBL-6.WT and ANBL-6.BR cell lines were kindly provided by Dr. Robert Orlowski (MD
Anderson Cancer Center, Texas). Bone marrow stromal cells (BMSCs) were cultured in
Das et al. Page 2
Br J Haematol. Author manuscript; available in PMC 2016 December 01.
Author M
anuscriptA
uthor Manuscript
Author M
anuscriptA
uthor Manuscript

Dulbecco’s modified Eagle medium supplemented with complete medium. Patient CD138+
MM cells, BMSCs and plasmacytoid dendritic cells (pDC) were isolated and cultured as
described previously.(Chauhan, et al 2009) Informed consent was obtained from all patients,
in accordance with an Institutional Review Board approved clinical protocol. Marizomib
was obtained from Triphase Accelerator Corporation (San Diego, CA, USA), and
pomalidomide was purchased from Selleck chemicals (Houston, TX, USA).
Cytotoxicity and apoptosis assays
Cell viability in MM cell lines, patient MM cells and normal PBMCs were assessed by 3-
(4,5-dimethylthiazol-2-yl)-2,5-diphenyltetrazolium bromide (MTT)/water-soluble
tetrazolium salt 1 (WST-1) assay. MM cell growth assessment in co-culture studies with
BMSCs or pDCs were performed using bromodeoxyuridine (BrdU) cell proliferation kits as
previously described.(Chauhan, et al 2008a) Apoptosis was quantified using FACSCanto
(BD Biosciences, San Jose, CA, USA). Caspase-8 and -9 fluorometric assay kits
(ALX-850-222-K101 and ALX-850-224-K101, Enzo Life Sciences, Farmingdale, NY) were
utilized to measure caspase-8 and caspase-9 enzymatic activity.
In vitro migration and capillary-like tube structure formation assays
The migration assay was performed using 24-well Transwell plates (Millipore, Billerica,
MA, USA) in the presence of 10% fetal bovine serum, and migrating cells were quantified
by measuring the fluorescence intensity, as previously described.(Chauhan, et al 2010)
Angiogenesis was determined in vitro by matrigel capillary-like tube structure formation
assay, as previously described.(Chauhan, et al 2008a) Human vascular endothelial cells
(HUVECs)(American Type Culture Collection [ATCC], Manassas, VA, USA) were
maintained in endothelial cell growth medium-2 supplemented with 5% FBS. After 3
passages, HUVEC viability was measured using Trypan blue exclusion assay; less than 5%
of cell death was observed with single or combined agents.
Western blotting and proteasome activity assays
Immunoblot analysis was performed using antibodies (Abs) against poly ADP ribose
polymerase (PARP) (BD Bioscience Pharmingen, San Diego, CA), cereblon (CRBN)
(Novus Biologicals), caspase-8, caspase-9, caspase-3, IRF4, MYC (c-Myc), MCL1,
HSP90AA1 (Hsp-90), HSPA1A (Hsp-70), HSPB1 (Hsp-27) or GAPDH (Cell Signaling,
Beverly, MA). Blots were then developed by enhanced chemiluminescence (Amersham,
Arlington Heights, IL). In vitro proteasome activity was performed using fluorogenic
peptide substrates, as previously described.(Chauhan, et al 2008a)
Transfection assays
CRBN knockdown experiment was performed using CRBN-siRNA (Origene, Rockville,
MD, USA). MM.1S cells were transfected with CRBN-siRNA or scrambled (scr)-siRNA
using the cell line Nucleofactor Kit V solution (Amaxa Biosystems/Lonza, Allendale, NJ,
USA), as per the manufacturer’s instructions. After cells were pre-treated with
pomalidomide for 24 h, marizomib was added for an additional 24h, followed by analysis of
cell viability by WST-1 assay.
Das et al. Page 3
Br J Haematol. Author manuscript; available in PMC 2016 December 01.
Author M
anuscriptA
uthor Manuscript
Author M
anuscriptA
uthor Manuscript

Human plasmacytoma xenograft model
The Animal Care and Use Committee of Dana Farber Cancer Institute approved all
experiments involving animals. The subcutaneous MM.1S mouse xenograft model was
generated as previously described(Chauhan, et al 2005a, Chauhan, et al 2008a, Chauhan, et
al 2010) CB-17 SCID-mice (n=30; Taconic, Petersburgh, NY, USA) were subcutaneously
inoculated with 5.0 × 106 MM.1S cells in 100 μl of serum-free RPMI 1640 medium.
Tumour size was measured in two dimensions using calipers; tumour volume was calculated
using the formula: V = 0.5 a x b2, where a and b are the long and short diameter of the
tumour, respectively. Animals were sacrificed when their tumours reached 2 cm3.
When tumours were measurable (~150 mm3), approximately 3 weeks after MM-cell
injection, mice (5 mice/group) were divided into random groups blindly and treated orally
with vehicle alone, marizomib (0.15 mg/kg), pomalidomide (0.5 mg/kg or 2.5 mg/kg) or
marizomib (0.15 mg/kg) plus pomalidomide (0.5 mg/kg or 2.5 mg/kg) for 24 days on a
twice-weekly schedule for marizomib and on 4 consecutive days weekly for pomalidomide.
Immunostaining
Sections of tumours harvested from mice were stained with anti-Ki67, anti-Caspase-3,
Factor VIII, or vascular endothelial growth factor receptor 1 (VEGFR1) Abs, as previously
described.(Chauhan, et al 2010) Immunostained tissues were imaged with a Zeiss
AxioImager M1 Microscope (Zeiss, Oberkochen, Germany) (Chauhan, et al 2010).
Statistical analysis
Statistical significance of differences observed in drug-treated vs. control culture was
determined using the Student’s t test. Statistical significance of differences observed in
tumour volumes in marizomib-, pomalidomide- or marizomib plus pomalidomide-treated
mice was determined using a Student’s t test. P values <0.05 were considered statistically
significant. Survival of mice was determined by GraphPad Prism software (GraphPad
Software, La Jolla, CA). Isobologram analysis(Chou and Talalay 1984) was performed by
using the CalcuSyn software program (Biosoft, Ferguson, MO, and Cambridge, United
Kingdom). A combination index value < 1.0 indicates synergism, and values > 1.0 indicate
antagonism.
Results
Anti-MM activity of combined low concentrations of marizomib and pomalidomide in vitro
Human MM-cell lines (MM.1S, INA-6, RPMI-8226, MM.1R, Dox-40, LR5, ANBL6.WT
and ANBL6.BR) were pretreated with pomalidomide for 24h; marizomib was then added for
an additional 24h, followed by assessment for cell viability using MTT assays. For the
combination studies, we utilized marizomib and pomalidomide across a range of
concentrations. An analysis of synergistic anti-MM activity using the method of Chou and
Talalay (1984) demonstrated a significant decrease in viability of all cell lines in response to
treatment with combined low concentrations of marizomib and pomalidomide compared
with either agent alone (Fig 1A–1D, and Supplementary Fig 1A–1E). Isobologram
analysis(Chou and Talalay 1984) confirmed synergistic anti-MM activity, with a
Das et al. Page 4
Br J Haematol. Author manuscript; available in PMC 2016 December 01.
Author M
anuscriptA
uthor Manuscript
Author M
anuscriptA
uthor Manuscript

combination index of < 1.0 in all MM cell lines tested. Importantly, the combination of low
concentrations of marizomib and pomalidomide overcame resistance to both conventional
and novel anti-MM agents. For example, marizomib plus pomalidomide decreased viability
of cell lines resistant to bortezomib (ANBL6.BR), dexamethasone (MM.1R), doxorubicin
(Dox-40) and melphalan (LR5) (Fig 1, and Supplementary Fig 1). Additionally, synergistic
anti-MM activity of combined marizomib and pomalidomide was also observed against
TP53-null ARP-1 MM cells (Fig 1C).
To determine whether the combination of low concentrations of marizomib and
pomalidomide similarly affected purified patient MM cells, we examined purified (CD138+)
MM cells from 2 newly diagnosed (Patients 6 and 7) and 5 patients with MM refractory to
multiple therapies (Fig 1E), including dexamethasone (Patient 1), bortezomib/
dexamethasone (Patients 2, 4 and 5), and lenalidomide/bortezomib/dexamethasone (Patient
3). MM was considered refractory to a specific therapy when disease progressed on that
therapy or relapsed within two months of discontinuing therapy. At the concentrations used,
either agent was moderately active in cells isolated from Patients 1 and 2 and was weakly
active or inactive in cells isolated from Patients 3–7. Importantly, a significant decrease in
cell viability of all patient MM cells was noted after combination therapy as compared to
either compound alone (p < 0.05 for all patients; Fig 1E). In contrast, marizomib (1.25 nM)
plus pomalidomide (2.5 μM) did not significantly decrease the viability of normal PBMCs
(Fig 1F), suggesting a favourable therapeutic index for this combination regimen.
Combined low concentrations of marizomib and pomalidomide block migration of MM cells and angiogenesis
Migration of MM cells and associated angiogenesis contributes to the progression of MM.
(Giuliani, et al 2003, Podar, et al 2001, Vacca, et al 2003) To determine whether marizomib
plus pomalidomide affects these processes, we utilized Transwell insert systems and in vitro
tubule formation assays. Serum alone increased MM.1S cell migration; importantly,
marizomib (1.25 nM) plus pomalidomide (2.5 μM) inhibited serum-dependent MM.1S cell
migration, as evidenced by a marked decrease in the number of migrating crystal violet-
stained cells (Fig 2A). Cells were >90% viable before and after performing the migration
assay, excluding the possibility that inhibition of migration was due to cell death. Our
findings show that marizomib plus pomalidomide blocked MM cell migration, which may
prevent homing of MM cells to the BM.
To determine whether marizomib plus pomalidomide exhibits anti-angiogenic activity, we
utilized in vitro capillary-like tube structure formation assays. Specifically, we performed
matrigel capillary-like tube structure formation assays: HUVECs plated onto matrigel
differentiate and form capillary-like tube structures reflecting in vivo neovascularization, a
process dependent on cell-matrix interaction, cellular communication and cellular motility.
Treatment of HUVECs with combined low concentrations of marizomib and pomalidomide,
but not either agent alone, significantly decreased tubule formation (Fig 2B). HUVEC
viability was assessed using the trypan blue exclusion assay and < 10% cell death was
observed with either single agents or the combined treatment. These findings suggest that
the combination of marizomib and pomalidomide block angiogenesis.
Das et al. Page 5
Br J Haematol. Author manuscript; available in PMC 2016 December 01.
Author M
anuscriptA
uthor Manuscript
Author M
anuscriptA
uthor Manuscript

Combination of low concentrations of marizomib plus pomalidomide inhibits bone marrow stromal cells (BMSCs)- and plasmacytoid dendritic cells (pDCs)-induced growth of MM cells
The MM-host BM microenvironment confers growth, survival and drug resistance in MM
cells.(Anderson 2007, Chauhan, et al 1996) Therefore, we next examined whether the
combination of marizomib plus pomalidomide retains its anti-MM activity in the presence of
the MM BM milieu. MM.1S cells were cultured with or without patient BMSCs in the
presence or absence of marizomib, pomalidomide, or marizomib plus pomalidomide. Low
concentrations of marizomib plus pomalidomide significantly inhibited BMSC-induced
growth of MM.1S cells, as assessed by WST-1 assays (Fig 2C).
Our recent studies have identified an integral role of pDCs in MM pathogenesis.(Chauhan,
et al 2009, Ray, et al 2015, Ray, et al 2014) The functional significance of increased
numbers of pDCs in the MM BM is evidenced by our observations that pDCs are relatively
resistant to novel and conventional therapies, protect tumour cells from therapy-induced
cytotoxicity, promote tumour growth and survival as well as suppress immune responses.
(Chauhan, et al 2009, Ray, et al 2015, Ray, et al 2014) We therefore examined the effect of
marizomib plus pomalidomide on pDC-induced MM cell growth. MM.1S cells (5 x 104
cells/200 μl) and patient pDCs (1 x 104 cells/200 μl) were cultured either alone or together.
Cells were pre-treated with pomalidomide for 24 h; marizomib was then added for an
additional 24 h; and growth was measured using a WST-1 assay. A significant inhibition of
pDC-induced MM.1S cell growth was observed in response to treatment with marizomib
plus pomalidomide (Fig 2D). Together, our data suggests that the combination of marizomib
and pomalidomide has the ability to overcome BMSC- or pDC-mediated MM cell growth
and drug resistance.
Mechanisms mediating the anti-MM activity of marizomib plus pomalidomide
As shown in Fig 1, the combination of low concentrations of marizomib plus pomalidomide
decreased the viability of MM cell lines and patient cells. This decrease in viability was due
to induction of apoptosis, as determined by Annexin V/propidium iodide staining and flow
cytometry (Supplementary Fig 2). Moreover, marizomib plus pomalidomide-induced
apoptosis in MM cell lines (MM.1S, RPMI-8226, Dox-40) was associated with PARP
cleavage (Fig 3A) and the activation of caspase-3, caspase-8 and caspase-9 (Fig 3B).
Biochemical inhibitors of caspase-3, caspase-8, caspase-9 or pan-caspases abrogated
marizomib plus pomaldiomide-induced cytotoxicity (Fig 3C). Of note, blockade of either
caspase-8 or caspase-9 led to a similar degree of inhibition of marizomib plus
pomalidomide-induced apoptosis.
Effects of marizomib plus pomalidomide on chymotrypsin-like (CT-L), caspase-like (C-L) and trypsin-like (T-L) proteasome activities
The primary target of proteasome inhibitors is the proteasome, which mediates degradation
of proteins via three (CT-L, T-L and C-L) proteolytically active sites.(Arendt and
Hochstrasser 1997, Heinemeyer, et al 1997) Therefore, we next examined the effect of
combined marizomib and pomalidomide on CT-L, C-L, and T-L activity. The combination
of low concentrations of marizomib (1.25 nM) and pomalidomide (2.5 μM) induced a
Das et al. Page 6
Br J Haematol. Author manuscript; available in PMC 2016 December 01.
Author M
anuscriptA
uthor Manuscript
Author M
anuscriptA
uthor Manuscript

marked inhibition of all three proteasomal activities versus either agent alone at these
concentrations (Fig 4A), suggesting that pomalidomide enhances the ability of marizomib to
target all three proteasome activities.
Effect of marizomib plus pomalidomide on cereblon and associated downstream signaling
The primary target of pomalidomide is the E3 ubiquitin ligase, cereblon (CRBN).(Lu, et al
2014) Immunoblot analysis of CRBN expression in MM cells and normal healthy donor
PBMCs showed significantly higher CRBN expression in MM cell lines than in normal
PBMCs (Fig 4B). We therefore examined the functional role of CRBN during marizomib
plus pomalidomide-induced cytotoxicity using loss-of-function studies. The functional
specificity of CRBN-siRNA was confirmed by a marked decrease in CRBN levels (Fig 4C,
immunoblot). Importantly, transfection of CRBN-siRNA, but not negative-control
(scrambled) siRNA significantly inhibited marizomib plus pomalidomide-induced apoptosis
in MM.1S cells (Fig 4C; p < 0.005). We found no statistically significant difference upon
treatment with marizomib alone in either CRBN siRNA or scrambled siRNA transfected
cells. Non-transfected cells showed results similar to those noted in scrambled siRNA
transfected cells (data not shown). These findings suggest that marizomib plus
pomalidomide-triggered apoptosis in MM cells is mediated, at least in part, via CRBN.
CRBN is linked to the activation of the downstream IRF4/MYC signalling axis.(Thakurta, et
al 2014) IRF4 is a survival factor for MM cells, and MYC is a direct target of IRF4 in
activated B cells and MM cells. Conversely, IRF4 is also a direct target of MYC
transactivation, generating an autoregulatory circuit in MM cells.(Shaffer, et al 2008)
Importantly, MYC coordinates cellular growth, metabolism and proliferation.(Holien, et al
2012) In this study, treatment of MM.1S and RPMI-8226 cells with marizomib plus
pomalidomide led to reduction of CRBN, IRF4 and MYC protein expression (Fig 4D and
Supplementary Fig 3). NF-κB (NFKB1) is another key mediator of MM cell survival.
(Chauhan, et al 1996) Interestingly, lenalidomide inhibits IRF4, which in turn
downregulates NF-κB in a CRBN-dependent manner.(Zhang, et al 2013) In our study,
treatment of MM cells with low doses of marizomib and pomalidomide also blocks NF-κB
activity (Supplementary Fig 4). Together, these findings suggest that marizomib plus
pomalidomide-induced apoptosis in MM cells is associated with downregulation of the
CRBN/IRF4/MYC signalling axis.
We next examined whether the combination of marizomib and pomalidomide overcomes
mechanism(s) that confer survival and drug-resistance in MM cells. For example, the anti-
apoptotic protein MCL1 confers drug resistance in MM cells(Fan, et al 2014), and we found
a marked decrease in MCL1 levels in marizomib and pomalidomide-treated cells versus
cells treated with either agent alone (Fig 4D). In addition, prior studies link heat shock
proteins (HSPB1, HSPA1A, HSP90AA1) with drug resistance.(Chauhan, et al 2005b,
Chauhan, et al 2003, Fribley, et al 2004, Landowski, et al 2005, Obeng, et al 2006)
Importantly, we found that the combination of low concentrations of marizomib and
pomalidomide did not induce HSPB1, HSPA1A or HSP90AA1 protein levels (Fig 4E).
Given the role of MCL1 and heat shock proteins in the development of drug-resistance, our
Das et al. Page 7
Br J Haematol. Author manuscript; available in PMC 2016 December 01.
Author M
anuscriptA
uthor Manuscript
Author M
anuscriptA
uthor Manuscript

data suggest that combined low doses of marizomib and pomalidomide may result in less
frequent occurrence of drug resistance in MM.
Combined marizomib and pomalidomide inhibit human MM cell growth in vivo and prolongs survival in the MM.1S MM xenograft mouse model
Having defined the efficacy of combined marizomib and pomalidomide in targeting MM
cells in vitro, we next validated these findings in vivo using our murine xenograft model of
human MM.(Chauhan, et al 2008a, Chauhan, et al 2010) This model has been useful in
validating the novel anti-MM therapies bortezomib and lenalidomide, leading to their
translation to clinical trials and FDA approval for the treatment of MM. A marked reduction
in tumour growth, as well as an increased prolongation of survival, was observed in
marizomib plus pomalidomide-treated mice versus mice receiving either agent alone (Fig
5A and 5B). These data demonstrate the effectiveness of the anti-tumour activity of
marizomib plus pomalidomide in vivo. The combination of marizomib and pomalidomide
treatment was well tolerated, as evidenced by the lack of weight loss even after 3 weeks of
treatment (Fig 5C). Blood chemistry profiles of marizomib plus pomalidomide-treated mice
showed that haemoglobin, bilirubin and creatinine levels were not statistically significant
altered (Fig 5D). Together, these findings suggest that combining marizomib and
pomalidomide markedly reduced tumour growth and is well tolerated in vivo.
We next examined the apoptotic activity of this drug combination in vivo using
immunostaining for caspase-3 activation and terminal deoxynucleotidyl transferase dUTP
nick end labeling (TUNEL) staining of tumours harvested from mice treated with vehicle
alone, marizomib, pomalidomide, or marizomib plus pomalidomide. The combination of
marizomib plus pomalidomide increased the number of cleaved caspase-3- and TUNEL-
positive apoptotic tumour cells compared to treatment with vehicle or either agent alone (Fig
6A and 6B). Furthermore, a decrease in the proliferation marker Ki67 was also noted in
tumours excised from mice receiving marizomib plus pomalidomide (Fig 6C). In concert
with our in vitro data (Fig 4D), tumours from mice treated with marizomib plus
pomalidomide showed decreased MCL1 and IRF4 levels (Fig 6D).
MM cell growth is associated with angiogenesis, and vascular endothelial growth factor
(VEGF) plays a role in this process.(Anderson 2012) Our in vitro data showed that the
combination of marizomib and pomalidomide exerts anti-angiogenic activity (Fig 2A and
2B). To determine whether marizomib and pomalidomide trigger anti-angiogenic activity in
vivo, we evaluated tumours harvested from mice by immunostaining using two distinct
markers of angiogenesis, Factor VIII and VEGFR1. As seen in Fig 6E, low doses of
marizomib or pomalidomide alone triggered a very modest decrease in the number of Factor
VIII-positive cells compared to sections from control-treated tumours, whereas combination
treatment decreased the number of Factor VIII-positive cells. Similarly, marizomib plus
pomalidomide markedly decreased the number of VEGFR1-positive cells (Fig 6F). These
data suggest that marizomib plus pomalidomide inhibits tumour-associated angiogenic
activity.
Das et al. Page 8
Br J Haematol. Author manuscript; available in PMC 2016 December 01.
Author M
anuscriptA
uthor Manuscript
Author M
anuscriptA
uthor Manuscript

Discussion
We show that the combination of low concentrations of marizomib and pomalidomide
decreased viability of MM cell lines and primary patient tumour cells, without affecting
normal PBMC viability. Our data show the anti-MM activity of marizomib plus
pomalidomide against MM cell lines, including those sensitive and resistant to conventional
and novel therapies, as well as representing distinct cytogenetic profiles.(Bergsagel, et al
1996, Bergsagel and Kuehl 2005) For example, we studied isogenic cell lines
dexamethasone-sensitive MM.1S(Greenstein, et al 2003) and dexamethasone-resistant MM.
1R with t(14;16) translocation and MAF (c-maf) overexpression; INA-6, an interleukin 6–
dependent cell line with NRAS activating mutation; and RPMI-8266 with TP53, KRAS and
EGFR mutations.(Avet-Loiseau, et al 2007, Bergsagel, et al 1996, Bergsagel and Kuehl
2005, Burger, et al 2001, Davies, et al 2003, Greenstein, et al 2003) Although combined
treatment with marizomib and pomalidomide decreased viability in all MM cell lines, the
50% inhibitory concentration of this combination regimen was different for each cell line,
which may be due to their distinct genetic heterogeneity and/or drug-resistance
characteristics.
We observed maximal synergistic anti-MM activity of combined marizomib and
pomalidomide when MM cells were pretreated with pomalidomide for 24 h followed by
addition of marizomib for another 24 h versus the simultaneous addition of these agents
(data not shown). It is likely that pomalidomide induces distinct apoptotic signalling versus
marizomib and/or upregulates proteasome load in MM cells, which together primes the cells
to undergo robust cell death in response to subsequent treatment with even low
concentrations of marizomib.
To determine whether the combination of marizomib and pomalidomide overcomes
bortezomib resistance, we used previously characterized(Kuhn, et al 2009) bortezomib-
sensitive (ANBL-6.WT) and -resistant (ANBL-6.BR) MM cell lines. We found significant
anti-MM activity of marizomib plus pomalidomide against ANBL-6.BR, confirming the
ability of this regimen to overcome bortezomib-resistance. Importantly, we observed similar
responses to marizomib plus pomalidomide in patient MM cells resistant to therapies
including bortezomib, lenalidomide or Dex. Moreover, the combination of marizomib and
pomalidomide overcomes the MM cell growth advantage conferred by the MM-host BM
microenvironment.
Mechanistic analysis showed that anti-MM activity of marizomib plus pomalidomide is
associated with the activation of the caspase cascade, inhibition of proteasomal activities,
and downregulation of both CRBN/IRF4/MYC signalling axis and MCL1. Studies using
biochemical inhibitors showed that blockade of either caspase-8 or caspase-9 led to a similar
degree of inhibition of marizomib plus pomalidomide-induced apoptosis. Of note, our earlier
study showed that the combination of marizomib with the immunomodulatory drug
lenalidomide relies more on caspase-8 activity than caspase-9 for inducing cell
death(Chauhan, et al 2010). Nonetheless, our data suggest that marizomib plus
pomalidomide-induced MM cell apoptosis is dependent, at least in part, on both caspase-8-
and caspase-9-mediated signalling pathways.
Das et al. Page 9
Br J Haematol. Author manuscript; available in PMC 2016 December 01.
Author M
anuscriptA
uthor Manuscript
Author M
anuscriptA
uthor Manuscript

Previous studies showed that marizomib inhibits CT-L, C-L and T-L proteasome activities.
(Chauhan, et al 2008b, Chauhan, et al 2005a) Our prior study also showed that lenalidomide
enhances marizomib-induced inhibition of proteasome function(Chauhan, et al 2010). In an
analogous manner, we found that the combined low concentrations of marizomib and
pomalidomide block all three proteasomal activities versus either agent alone at these low
concentrations. The mechanism whereby pomolidamide enhances the ability of marizomib
to potently inhibit proteasome activity remains to be defined. Given the immunomodulatory
characteristics of pomalidomide, it is likely that pomalidomide affects immunoproteasome
activities (CT-Li, C-Li and T-Li), which in turn may alter function of constitutive
proteasome activities. This notion is supported by earlier findings that proteasome active
sites allosterically regulate each other: for example, occupancy of C-L sites triggers the T-L
activity of proteasomes in a compensatory manner.(Kisselev, et al 2003) Indeed, additional
loss-of-function studies are required to delineate the functional role of each proteasome
subunit.
Another possible mechanism mediating the marizomib plus pomalidomide-induced
blockade of proteasome activity is the involvement of E3 ligase CRBN within the ubiquitin
proteasome pathway. CRBN is a direct protein target of pomalidomide.(Ito, et al 2010, Lu,
et al 2014) A recent study showed that CRBN modulates proteasome activity by binding to
the 20S core proteasome subunit beta type 4.(Lee, et al 2012) It is therefore possible that
CRBN may similarly affect the β5, β2 or β1 proteasome subunits to modulate overall
proteasome activity. Our genetic studies using CRBN-siRNA suggest that marizomib plus
pomalidomide-induced cell death is mediated, at least in part, by CRBN. A further evidence
for the involvement of CRBN is noted by a marked decrease in CRBN associated signalling
via IRF4 and MYC. Together, these data suggest that marizomib plus pomalidomide-
induced apoptosis is associated with a marked inhibition of all three proteasome activities,
as well as CRBN-associated downstream signalling via IRF4/MYC.
In addition to activating pro-apoptotic signaling, the combination of marizomib and
pomalidomide also downregulates proteins associated with survival and drug resistance. For
example, MCL1 is a pro-survival member of the BCL2 family, which plays a crucial role in
MM pathogenesis and drug resistance.(Fan, et al 2014) We found that low concentrations of
marizomib plus pomalidomide reduced the levels of anti-apoptotic protein MCL1. Similarly,
prior studies have linked bortezomib-induced apoptosis with the upregulation of heat shock
proteins, which confer drug resistance in MM.(Chauhan, et al 2005b, Mitsiades, et al 2002,
Obeng, et al 2006) In this context, our data shows that marizomib plus pomalidomide
combination therapy does not induce HSP90AA1, HSPA1A or HSPB1. These data suggest
that drug resistance may be less frequent in patients receiving this combined low dose
regimen.
In vitro data showing anti-MM activity of marizomib plus pomalidomide was validated in
vivo using the human MM.1S xenograft mouse model. A marked reduction in tumour
progression and prolongation of survival was observed in marizomib plus pomalidomide-
treated mice versus mice receiving either agent alone. Moreover, marizomib plus
pomalidomide treatment was well tolerated, because differences in body weight and overall
appearance were not observed. The remarkable anti-MM and anti-angiogenic activity of
Das et al. Page 10
Br J Haematol. Author manuscript; available in PMC 2016 December 01.
Author M
anuscriptA
uthor Manuscript
Author M
anuscriptA
uthor Manuscript

marizomib plus pomalidomide in vivo was confirmed by IHC analysis of tumours harvested
from control- and combination-treated mice using molecular markers of apoptosis
(caspase-3 cleavage, TUNEL staining), proliferation (Ki67), and associated angiogenesis
(Factor VIII and VEGFR1). Tumour lysates showed decreased MCL1 and IRF4 levels after
marizomib plus pomalidomide treatment. Therefore, these findings indicate a dual effect of
combining marizomib plus pomalidomide: increased apoptosis and decreased MM-cell
proliferation.
Collectively, our preclinical studies demonstrate potent in vitro and in vivo anti-MM activity
of marizomib plus pomalidomide at low doses that are well tolerated in a human
plasmacytoma xenograft mouse model. These findings provide the rationale for the
continuation of clinical trials of low-dose combination marizomib plus pomalidomide
therapy to enhance cytotoxicity, overcome drug resistance, reduce side effects, and improve
patient outcome in MM.
Supplementary Material
Refer to Web version on PubMed Central for supplementary material.
Acknowledgments
Grant Support: This investigation was supported by National Institutes of Health Specialized Programs of Research Excellence (SPORE) grant P50100707, PO1-CA078378, and RO1 CA050947. K.C.A. is an American Cancer Society Clinical Research Professor.
References
Anderson KC. Targeted therapy of multiple myeloma based upon tumor-microenvironmental interactions. Exp Hematol. 2007; 35:155–162. [PubMed: 17379101]
Anderson KC. The 39th David A. Karnofsky Lecture: bench-to-bedside translation of targeted therapies in multiple myeloma. J Clin Oncol. 2012; 30:445–452. [PubMed: 22215754]
Arendt C, Hochstrasser M. Identification of the yeast 20S proteasome catalytic centers and subunit interactions required for active-site formation. Proc Natl Acad Sci U S A. 1997; 94:7156–7161. [PubMed: 9207060]
Atrash S, Tullos A, Panozzo S, Bhutani M, Van Rhee F, Barlogie B, Usmani SZ. Cardiac complications in relapsed and refractory multiple myeloma patients treated with carfilzomib. Blood Cancer J. 2015; 5:e272. [PubMed: 25594159]
Avet-Loiseau H, Attal M, Moreau P, Charbonnel C, Garban F, Hulin C, Leyvraz S, Michallet M, Yakoub-Agha I, Garderet L, Marit G, Michaux L, Voillat L, Renaud M, Grosbois B, Guillerm G, Benboubker L, Monconduit M, Thieblemont C, Casassus P, Caillot D, Stoppa AM, Sotto JJ, Wetterwald M, Dumontet C, Fuzibet JG, Azais I, Dorvaux V, Zandecki M, Bataille R, Minvielle S, Harousseau JL, Facon T, Mathiot C. Genetic abnormalities and survival in multiple myeloma: the experience of the Intergroupe Francophone du Myelome. Blood. 2007; 109:3489–3495. [PubMed: 17209057]
Bergsagel PL, Kuehl WM. Molecular pathogenesis and a consequent classification of multiple myeloma. J Clin Oncol. 2005; 23:6333–6338. [PubMed: 16155016]
Bergsagel PL, Chesi M, Nardini E, Brents LA, Kirby SL, Kuehl WM. Promiscuous translocations into immunoglobulin heavy chain switch regions in multiple myeloma. Proc Natl Acad Sci U S A. 1996; 93:13931–13936. [PubMed: 8943038]
Burger R, Guenther A, Bakker F, Schmalzing M, Bernand S, Baum W, Duerr B, Hocke GM, Steininger H, Gebhart E, Gramatzki M. Gp130 and ras mediated signaling in human plasma cell line
Das et al. Page 11
Br J Haematol. Author manuscript; available in PMC 2016 December 01.
Author M
anuscriptA
uthor Manuscript
Author M
anuscriptA
uthor Manuscript

INA-6: a cytokine-regulated tumor model for plasmacytoma. Hematol J. 2001; 2:42–53. [PubMed: 11920233]
Cai X, Bhattacharyya S, Plitt A, Raibagkar P, Labuzetta JN, Schleicher SM, Munshi NC, Klein JP. Management of Posterior Reversible Encephalopathy Syndrome Induced by Carfilzomib in a Patient With Multiple Myeloma. J Clin Oncol. 2014 Epub ahead of print 5 May 2014. 10.1200/JCO.2013.49.6166
Chauhan D, Uchiyama H, Akbarali Y, Urashima M, Yamamoto K, Libermann TA, Anderson KC. Multiple myeloma cell adhesion-induced interleukin-6 expression in bone marrow stromal cells involves activation of NF-kappa B. Blood. 1996; 87:1104–1112. [PubMed: 8562936]
Chauhan D, Li G, Shringarpure R, Podar K, Ohtake Y, Hideshima T, Anderson KC. Blockade of Hsp27 overcomes Bortezomib/proteasome inhibitor PS-341 resistance in lymphoma cells. Cancer Res. 2003; 63:6174–6177. [PubMed: 14559800]
Chauhan D, Catley L, Li G, Podar K, Hideshima T, Velankar M, Mitsiades C, Mitsiades N, Yasui H, Letai A, Ovaa H, Berkers C, Nicholson B, Chao TH, Neuteboom ST, Richardson P, Palladino MA, Anderson KC. A novel orally active proteasome inhibitor induces apoptosis in multiple myeloma cells with mechanisms distinct from Bortezomib. Cancer Cell. 2005a; 8:407–419. [PubMed: 16286248]
Chauhan D, Hideshima T, Anderson KC. Proteasome inhibition in multiple myeloma: therapeutic implication. Annu Rev Pharmacol Toxicol. 2005b; 45:465–476. [PubMed: 15822185]
Chauhan D, Singh A, Brahmandam M, Podar K, Hideshima T, Richardson P, Munshi N, Palladino MA, Anderson KC. Combination of proteasome inhibitors bortezomib and NPI-0052 trigger in vivo synergistic cytotoxicity in multiple myeloma. Blood. 2008a; 111:1654–1664. [PubMed: 18006697]
Chauhan D, Bianchi G, Anderson KC. Targeting the UPS as therapy in multiple myeloma. BMC Biochem. 2008b; 9(Suppl 1):S1. [PubMed: 19007431]
Chauhan D, Singh AV, Brahmandam M, Carrasco R, Bandi M, Hideshima T, Bianchi G, Podar K, Tai YT, Mitsiades C, Raje N, Jaye DL, Kumar SK, Richardson P, Munshi N, Anderson KC. Functional interaction of plasmacytoid dendritic cells with multiple myeloma cells: a therapeutic target. Cancer Cell. 2009; 16:309–323. [PubMed: 19800576]
Chauhan D, Singh AV, Ciccarelli B, Richardson PG, Palladino MA, Anderson KC. Combination of novel proteasome inhibitor NPI-0052 and lenalidomide trigger in vitro and in vivo synergistic cytotoxicity in multiple myeloma. Blood. 2010; 115:834–845. [PubMed: 19965674]
Chou TC, Talalay P. Quantitative analysis of dose-effect relationships: the combined effects of multiple drugs or enzyme inhibitors. Adv Enzyme Regul. 1984; 22:27–55. [PubMed: 6382953]
Davies FE, Dring AM, Li C, Rawstron AC, Shammas MA, O’Connor SM, Fenton JA, Hideshima T, Chauhan D, Tai IT, Robinson E, Auclair D, Rees K, Gonzalez D, Ashcroft AJ, Dasgupta R, Mitsiades C, Mitsiades N, Chen LB, Wong WH, Munshi NC, Morgan GJ, Anderson KC. Insights into the multistep transformation of MGUS to myeloma using microarray expression analysis. Blood. 2003; 102:4504–4511. [PubMed: 12947006]
Fan F, Tonon G, Bashari MH, Vallet S, Antonini E, Goldschmidt H, Schulze-Bergkamen H, Opferman JT, Sattler M, Anderson KC, Jager D, Podar K. Targeting Mcl-1 for multiple myeloma (MM) therapy: drug-induced generation of Mcl-1 fragment Mcl-1(128–350) triggers MM cell death via c-Jun upregulation. Cancer Lett. 2014; 343:286–294. [PubMed: 24120758]
Feling RH, Buchanan GO, Mincer TJ, Kauffman CA, Jensen PR, Fenical W. Salinosporamide A: a highly cytotoxic proteasome inhibitor from a novel microbial source, a marine bacterium of the new genus salinospora. Angew Chem Int Ed Engl. 2003; 42:355–357. [PubMed: 12548698]
Fribley A, Zeng Q, Wang CY. Proteasome inhibitor PS-341 induces apoptosis through induction of endoplasmic reticulum stress-reactive oxygen species in head and neck squamous cell carcinoma cells. Mol Cell Biol. 2004; 24:9695–9704. [PubMed: 15509775]
Giuliani N, Colla S, Lazzaretti M, Sala R, Roti G, Mancini C, Bonomini S, Lunghi P, Hojden M, Genestreti G, Svaldi M, Coser P, Fattori PP, Sammarelli G, Gazzola GC, Bataille R, Almici C, Caramatti C, Mangoni L, Rizzoli V. Pro-angiogenetic properties of human myeloma cells: production of angiopoietin-1 and its potential relationship with myeloma-induced angiogenesis. Blood. 2003; 102:638–45. [PubMed: 12649156]
Das et al. Page 12
Br J Haematol. Author manuscript; available in PMC 2016 December 01.
Author M
anuscriptA
uthor Manuscript
Author M
anuscriptA
uthor Manuscript

Gras J. Pomalidomide for patients with multiple myeloma. Drugs Today (Barc). 2013; 49:555–562. [PubMed: 24086951]
Greenstein S, Krett NL, Kurosawa Y, Ma C, Chauhan D, Hideshima T, Anderson KC, Rosen ST. Characterization of the MM.1 human multiple myeloma (MM) cell lines. A model system to elucidate the characteristics, behavior, and signaling of steroid-sensitive and -resistant MM cells. Exp Hematol. 2003; 31:271–282. [PubMed: 12691914]
Harvey RD. Incidence and management of adverse events in patients with relapsed and/or refractory multiple myeloma receiving single-agent carfilzomib. Clin Pharmacol. 2014; 6:87–96. [PubMed: 24855395]
Heinemeyer W, Fischer M, Krimmer T, Stachon U, Wolf D. The active sites of the eukaryotic 20 S proteasome and their involvement in subunit precursor processing. J Biol Chem. 1997; 272:25200–25209. [PubMed: 9312134]
Holien T, Vatsveen TK, Hella H, Waage A, Sundan A. Addiction to c-MYC in multiple myeloma. Blood. 2012; 120:2450–2453. [PubMed: 22806891]
Huber EM, Heinemeyer W, Groll M. Bortezomib-Resistant Mutant Proteasomes: Structural and Biochemical Evaluation with Carfilzomib and ONX 0914. Structure. 2015; 23:407–417. [PubMed: 25599643]
Ito T, Ando H, Suzuki T, Ogura T, Hotta K, Imamura Y, Yamaguchi Y, Handa H. Identification of a primary target of thalidomide teratogenicity. Science. 2010; 327:1345–1350. [PubMed: 20223979]
Kane RC, Bross PF, Farrell AT, Pazdur R. Velcade: U.S. FDA approval for the treatment of multiple myeloma progressing on prior therapy. Oncologist. 2003; 8:508–513. [PubMed: 14657528]
Kisselev AF, Garcia-Calvo M, Overkleeft HS, Peterson E, Pennington MW, Ploegh HL, Thornberry NA, Goldberg AL. The caspase-like sites of proteasomes, their substrate specificity, new inhibitors and substrates, and allosteric interactions with the trypsin-like sites. J Biol Chem. 2003; 278:35869–35877. [PubMed: 12815064]
Kuhn D, Bjorklund C, Magarotto V, Mathews J, Wang M, Baladandayuthapani V, Orlowski R. Bortezomib resistance is mediated by increased signaling through the insulin-like growth factor-1/Akt axis. Blood (ASH Annual Meeting Abstracts). 2009; 114:2739.
Landowski TH, Megli CJ, Nullmeyer KD, Lynch RM, Dorr RT. Mitochondrial-mediated disregulation of Ca2+ is a critical determinant of Velcade (PS-341/bortezomib) cytotoxicity in myeloma cell lines. Cancer Res. 2005; 65:3828–3836. [PubMed: 15867381]
Lee KM, Lee J, Park CS. Cereblon inhibits proteasome activity by binding to the 20S core proteasome subunit beta type 4. Biochem Biophys Res Commun. 2012; 427:618–622. [PubMed: 23026050]
Lonial S, Waller EK, Richardson PG, Jagannath S, Orlowski RZ, Giver CR, Jaye DL, Francis D, Giusti S, Torre C, Barlogie B, Berenson JR, Singhal S, Schenkein DP, Esseltine DL, Anderson J, Xiao H, Heffner LT, Anderson KC, Investigators SC. Risk factors and kinetics of thrombocytopenia associated with bortezomib for relapsed, refractory multiple myeloma. Blood. 2005; 106:3777–3784. [PubMed: 16099887]
Lu G, Middleton RE, Sun H, Naniong M, Ott CJ, Mitsiades CS, Wong KK, Bradner JE, Kaelin WG Jr. The myeloma drug lenalidomide promotes the cereblon-dependent destruction of Ikaros proteins. Science. 2014; 343:305–309. [PubMed: 24292623]
Mitsiades N, Mitsiades CS, Poulaki V, Chauhan D, Fanourakis G, Gu X, Bailey C, Joseph M, Libermann TA, Treon SP, Munshi NC, Richardson PG, Hideshima T, Anderson KC. Molecular sequelae of proteasome inhibition in human multiple myeloma cells. Proc Natl Acad Sci U S A. 2002; 99:14374–14379. [PubMed: 12391322]
Obeng EA, Carlson LM, Gutman DM, Harrington WJ Jr, Lee KP, Boise LH. Proteasome inhibitors induce a terminal unfolded protein response in multiple myeloma cells. Blood. 2006; 107:4907–4916. [PubMed: 16507771]
Podar K, Tai YT, Davies FE, Lentzsch S, Sattler M, Hideshima T, Lin BK, Gupta D, Shima Y, Chauhan D, Mitsiades C, Raje N, Richardson P, Anderson KC. Vascular endothelial growth factor triggers signaling cascades mediating multiple myeloma cell growth and migration. Blood. 2001; 98:428–435. [PubMed: 11435313]
Potts BC, Albitar MX, Anderson KC, Baritaki S, Berkers C, Bonavida B, Chandra J, Chauhan D, Cusack JC Jr, Fenical W, Ghobrial IM, Groll M, Jensen PR, Lam KS, Lloyd GK, McBride W,
Das et al. Page 13
Br J Haematol. Author manuscript; available in PMC 2016 December 01.
Author M
anuscriptA
uthor Manuscript
Author M
anuscriptA
uthor Manuscript

McConkey DJ, Miller CP, Neuteboom ST, Oki Y, Ovaa H, Pajonk F, Richardson PG, Roccaro AM, Sloss CM, Spear MA, Valashi E, Younes A, Palladino MA. Marizomib, a proteasome inhibitor for all seasons: preclinical profile and a framework for clinical trials. Curr Cancer Drug Targets. 2011; 11:254–284. [PubMed: 21247382]
Ray A, Tian Z, Das DS, Coffman RL, Richardson P, Chauhan D, Anderson KC. A novel TLR-9 agonist C792 inhibits plasmacytoid dendritic cell-induced myeloma cell growth and enhance cytotoxicity of bortezomib. Leukemia. 2014; 28:1716–1724. [PubMed: 24476765]
Ray A, Das DS, Song Y, PGR, Chauhan DKCA. Targeting PD1-PDL1 immune checkpoint in plasmacytoid dendritic cells interactions with T cells, natural killer cells, and multiple myeloma cells. Leukemia. 2015 (In press). 10.1038/leu.2015.11
Richardson P, Barlogie B, Berenson J, Singhal S, Jagannath S, Irwin D, Rajkumar SV, Srkalovic G, Alsina M, Alexanian R, Siegel D, Orlowski RZ, Kuter D, Limentani SA, Lee S, Hideshima T, Esseltine DL, Kauffman M, Adams J, Schenkein DP, Anderson KC. A multicenter phase II multicenter study of Bortezomib in patients with relapsed and refractory multiple myeloma. N Engl J Med. 2003; 348:2609–2617. [PubMed: 12826635]
Richardson PG, Sonneveld P, Schuster MW, Irwin D, Stadtmauer EA, Facon T, Harousseau JL, Ben-Yehuda D, Lonial S, Goldschmidt H, Reece D, San-Miguel JF, Blade J, Boccadoro M, Cavenagh J, Dalton WS, Boral AL, Esseltine DL, Porter JB, Schenkein D, Anderson KC. Assessment of Proteasome Inhibition for Extending Remissions I. Bortezomib or high-dose dexamethasone for relapsed multiple myeloma. N Engl J Med. 2005; 352:2487–2498. [PubMed: 15958804]
Richardson PG, Briemberg H, Jagannath S, Wen PY, Barlogie B, Berenson J, Singhal S, Siegel DS, Irwin D, Schuster M, Srkalovic G, Alexanian R, Rajkumar SV, Limentani S, Alsina M, Orlowski RZ, Najarian K, Esseltine D, Anderson KC, Amato AA. Frequency, characteristics, and reversibility of peripheral neuropathy during treatment of advanced multiple myeloma with bortezomib. J Clin Oncol. 2006; 24:3113–3120. [PubMed: 16754936]
Richardson P, Spencer A, Cannell P, Harrison SJ, Catley L, Underhill C, Zimmerman T, Hofmeister C, Jakubowiak A, Laubach J, Palladini M, Longenecker A, Wear S, Lloyd KG, Hannah AL, Reich S, Spear M, Anderson KC. Phase 1 Clinical Evaluation of Twice-Weekly Marizomib (NPI-0052), a Novel Proteasome Inhibitor, in Patients with Relapsed/Refractory Multiple Myeloma (MM). Blood (ASH Annual meeting abstracts). 2011; 118:302.
Richardson PG, Siegel D, Baz R, Kelley SL, Munshi NC, Laubach J, Sullivan D, Alsina M, Schlossman R, Ghobrial IM, Doss D, Loughney N, McBride L, Bilotti E, Anand P, Nardelli L, Wear S, Larkins G, Chen M, Zaki MH, Jacques C, Anderson KC. Phase 1 study of pomalidomide MTD, safety, and efficacy in patients with refractory multiple myeloma who have received lenalidomide and bortezomib. Blood. 2013; 121:1961–1967. [PubMed: 23243282]
Richardson PG, Siegel DS, Vij R, Hofmeister CC, Baz R, Jagannath S, Chen C, Lonial S, Jakubowiak A, Bahlis N, Song K, Belch A, Raje N, Shustik C, Lentzsch S, Lacy M, Mikhael J, Matous J, Vesole D, Chen M, Zaki MH, Jacques C, Yu Z, Anderson KC. Pomalidomide alone or in combination with low-dose dexamethasone in relapsed and refractory multiple myeloma: a randomized phase 2 study. Blood. 2014; 123:1826–1832. [PubMed: 24421329]
Shaffer AL, Emre NC, Lamy L, Ngo VN, Wright G, Xiao W, Powell J, Dave S, Yu X, Zhao H, Zeng Y, Chen B, Epstein J, Staudt LM. IRF4 addiction in multiple myeloma. Nature. 2008; 454:226–231. [PubMed: 18568025]
Siegel DS, Martin T, Wang M, Vij R, Jakubowiak AJ, Lonial S, Trudel S, Kukreti V, Bahlis N, Alsina M, Chanan-Khan A, Buadi F, Reu FJ, Somlo G, Zonder J, Song K, Stewart AK, Stadtmauer E, Kunkel L, Wear S, Wong AF, Orlowski RZ, Jagannath S. A phase 2 study of single-agent carfilzomib (PX-171-003-A1) in patients with relapsed and refractory multiple myeloma. Blood. 2012; 120:2817–2825. [PubMed: 22833546]
Thakurta A, Gandhi AK, Waldman MF, Bjorklund C, Ning Y, Mendy D, Schafer P, Lopez-Girona A, Lentzsch S, Schey SA, Calle Y, Chelliah R, Orlowski RZ, Madan A, Avet-Loiseau H, Chopra R. Absence of mutations in cereblon (CRBN) and DNA damage-binding protein 1 (DDB1) genes and significance for IMiD therapy. Leukemia. 2014; 28:1129–1131. [PubMed: 24166296]
Vacca A, Ria R, Ribatti D, Semeraro F, Djonov V, Di Raimondo F, Dammacco F. A paracrine loop in the vascular endothelial growth factor pathway triggers tumor angiogenesis and growth in multiple myeloma. Haematologica. 2003; 88:176–185. [PubMed: 12604407]
Das et al. Page 14
Br J Haematol. Author manuscript; available in PMC 2016 December 01.
Author M
anuscriptA
uthor Manuscript
Author M
anuscriptA
uthor Manuscript

Vij R, Siegel DS, Jagannath S, Jakubowiak AJ, Stewart AK, McDonagh K, Bahlis N, Belch A, Kunkel LA, Wear S, Wong AF, Wang M. An open-label, single-arm, phase 2 study of single-agent carfilzomib in patients with relapsed and/or refractory multiple myeloma who have been previously treated with bortezomib. Br J Haematol. 2012a; 158:739–748. [PubMed: 22845873]
Vij R, Wang M, Kaufman JL, Lonial S, Jakubowiak AJ, Stewart AK, Kukreti V, Jagannath S, McDonagh KT, Alsina M, Bahlis NJ, Reu FJ, Gabrail NY, Belch A, Matous JV, Lee P, Rosen P, Sebag M, Vesole DH, Kunkel LA, Wear SM, Wong AF, Orlowski RZ, Siegel DS. An open-label, single-arm, phase 2 (PX-171-004) study of single-agent carfilzomib in bortezomib-naive patients with relapsed and/or refractory multiple myeloma. Blood. 2012b; 119:5661–5670. [PubMed: 22555973]
Wanchoo R, Khan S, Kolitz JE, Jhaveri KD. Carfilzomib-related acute kidney injury may be prevented by N-acetyl-L-cysteine. J Oncol Pharm Pract. 2014; 21:313–6. [PubMed: 24748581]
Zhang LH, Kosek J, Wang M, Heise C, Schafer PH, Chopra R. Lenalidomide efficacy in activated B-cell-like subtype diffuse large B-cell lymphoma is dependent upon IRF4 and cereblon expression. Br J Haematol. 2013; 160:487–502. [PubMed: 23252516]
Das et al. Page 15
Br J Haematol. Author manuscript; available in PMC 2016 December 01.
Author M
anuscriptA
uthor Manuscript
Author M
anuscriptA
uthor Manuscript

Key points
1. Combining low concentrations of marizomib and pomalidomide triggers
synergistic cytotoxicity in MM cells and overcomes bortezomib-resistance.
2. Combination of marizomib with pomalidomide represents a promising novel
therapy in patients with relapsed and refractory MM.
Das et al. Page 16
Br J Haematol. Author manuscript; available in PMC 2016 December 01.
Author M
anuscriptA
uthor Manuscript
Author M
anuscriptA
uthor Manuscript
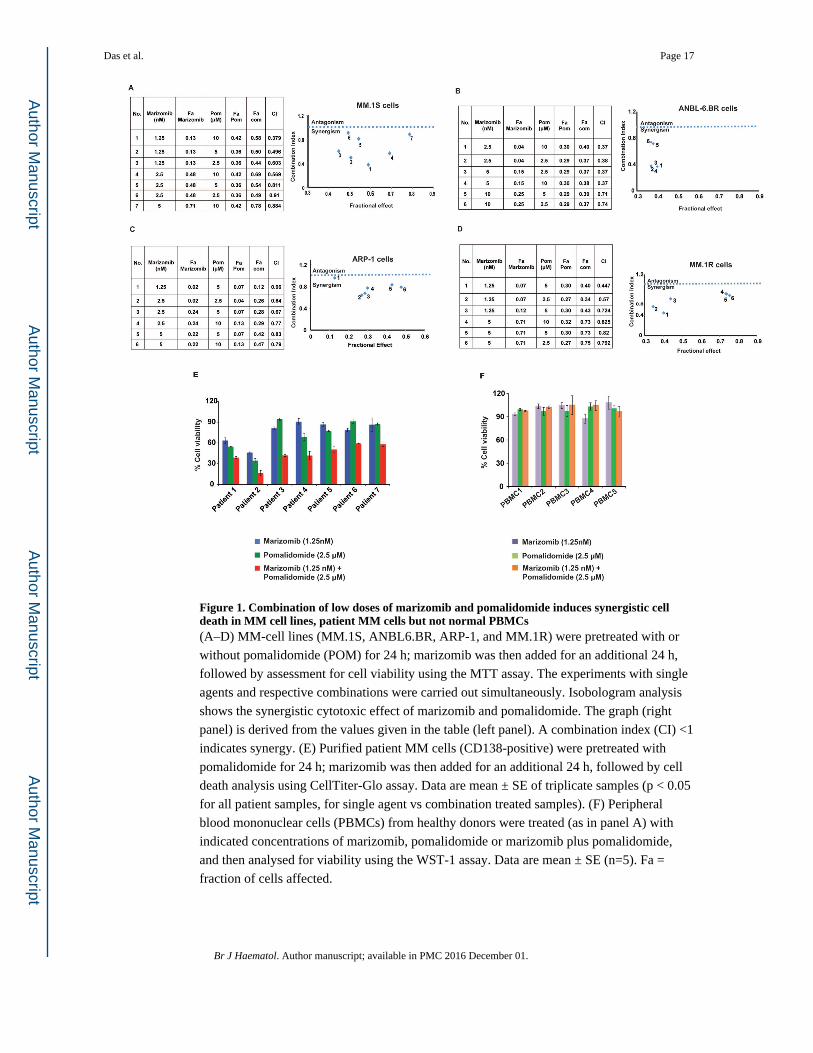
Figure 1. Combination of low doses of marizomib and pomalidomide induces synergistic cell death in MM cell lines, patient MM cells but not normal PBMCs(A–D) MM-cell lines (MM.1S, ANBL6.BR, ARP-1, and MM.1R) were pretreated with or
without pomalidomide (POM) for 24 h; marizomib was then added for an additional 24 h,
followed by assessment for cell viability using the MTT assay. The experiments with single
agents and respective combinations were carried out simultaneously. Isobologram analysis
shows the synergistic cytotoxic effect of marizomib and pomalidomide. The graph (right
panel) is derived from the values given in the table (left panel). A combination index (CI) <1
indicates synergy. (E) Purified patient MM cells (CD138-positive) were pretreated with
pomalidomide for 24 h; marizomib was then added for an additional 24 h, followed by cell
death analysis using CellTiter-Glo assay. Data are mean ± SE of triplicate samples (p < 0.05
for all patient samples, for single agent vs combination treated samples). (F) Peripheral
blood mononuclear cells (PBMCs) from healthy donors were treated (as in panel A) with
indicated concentrations of marizomib, pomalidomide or marizomib plus pomalidomide,
and then analysed for viability using the WST-1 assay. Data are mean ± SE (n=5). Fa =
fraction of cells affected.
Das et al. Page 17
Br J Haematol. Author manuscript; available in PMC 2016 December 01.
Author M
anuscriptA
uthor Manuscript
Author M
anuscriptA
uthor Manuscript

Figure 2. Combined low doses of marizomib and pomalidomide block migration, tubule formation, and cytoprotective effects of BMSCs and pDCs(A) Migration assay: MM.1S cells were pretreated with pomalidomide for 12h, and then
marizomib was added for an additional 6h; cells were more than 90% viable at this time
point. The cells were washed and cultured in serum-free medium. After 2h incubation, cells
were plated on a fibronectin-coated polycarbonate membrane in the upper chamber of
transwell inserts and exposed for 4h to serum-containing medium in the lower chamber.
Cells migrating to the bottom face of the membrane were fixed with 90% ethanol and
stained with crystal violet. A total of 3 randomly selected fields were examined for cells that
Das et al. Page 18
Br J Haematol. Author manuscript; available in PMC 2016 December 01.
Author M
anuscriptA
uthor Manuscript
Author M
anuscriptA
uthor Manuscript

had migrated from the top to the bottom chambers. (Left panel) Image is representative of 2
experiments with similar results. (Right panel) The bar graph represents quantification of
migrated cells. Data are mean ± SE (p < 0.05). (B) Human vascular endothelial cells were
cultured in the presence or absence of combined low doses of marizomib plus pomalidomide
for 48 h, and then assessed for in vitro angiogenesis using matrigel capillary-like tube
structure formation assays (Left panel). Image is representative from 3 experiments with
similar results. In vitro angiogenesis is reflected by capillary tube branch formation (dark
brown). (Right panel) The bar graph represents quantification of capillary-like tube structure
formation in response to indicated agents: Branch points in several random view fields/well
were counted, values were averaged, and statistically significant differences were measured
using Student’s t test. (C–D) MM.1S cells were cultured in bone marrow stromal cells
(BMSC)- or plasmacytoid dendritic cells (pDC)-coated or uncoated wells with control
medium, marizomib, pomalidomide or marizomib plus pomalidomide. Cell proliferation
was assessed by Brdu colorimetric assay. Data are mean ± SE (n=3; p < 0.05, for control vs
combination treated samples).
Das et al. Page 19
Br J Haematol. Author manuscript; available in PMC 2016 December 01.
Author M
anuscriptA
uthor Manuscript
Author M
anuscriptA
uthor Manuscript
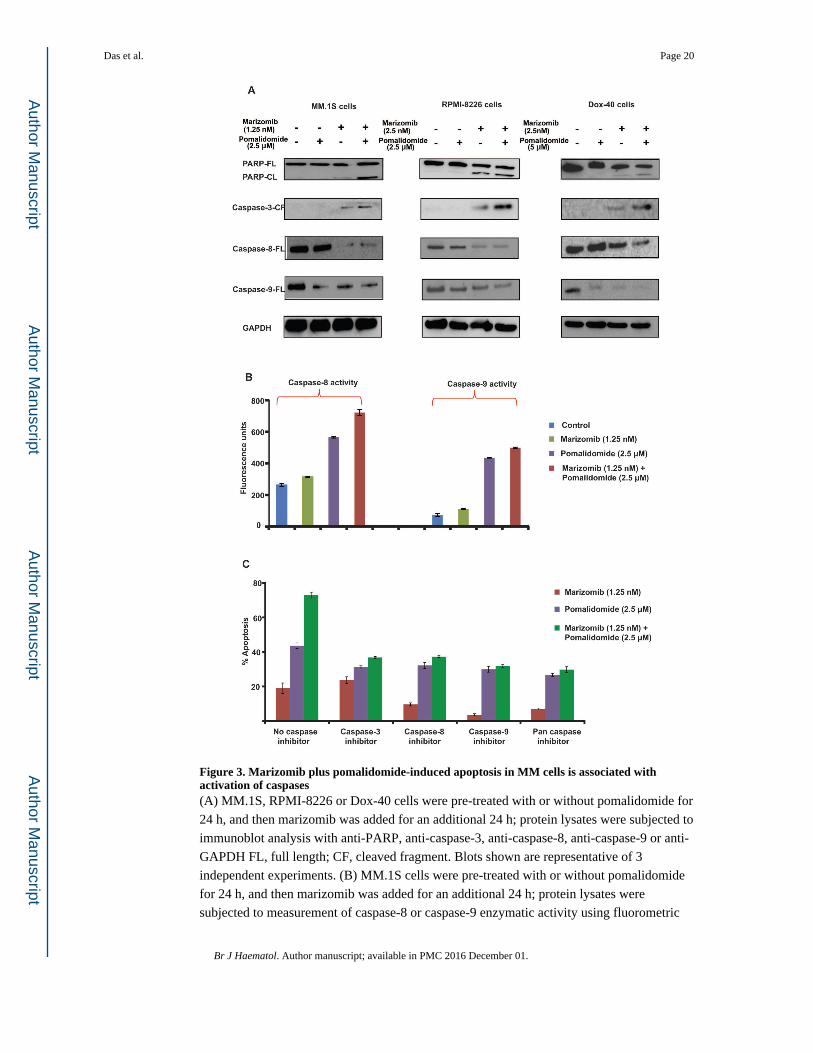
Figure 3. Marizomib plus pomalidomide-induced apoptosis in MM cells is associated with activation of caspases(A) MM.1S, RPMI-8226 or Dox-40 cells were pre-treated with or without pomalidomide for
24 h, and then marizomib was added for an additional 24 h; protein lysates were subjected to
immunoblot analysis with anti-PARP, anti-caspase-3, anti-caspase-8, anti-caspase-9 or anti-
GAPDH FL, full length; CF, cleaved fragment. Blots shown are representative of 3
independent experiments. (B) MM.1S cells were pre-treated with or without pomalidomide
for 24 h, and then marizomib was added for an additional 24 h; protein lysates were
subjected to measurement of caspase-8 or caspase-9 enzymatic activity using fluorometric
Das et al. Page 20
Br J Haematol. Author manuscript; available in PMC 2016 December 01.
Author M
anuscriptA
uthor Manuscript
Author M
anuscriptA
uthor Manuscript

kit. Data are mean ± SE (n=3; p < 0.05, for single agent vs combination treated samples) (C)
MM.1S cells were treated with indicated agents (as in panel ‘A’) in the presence or absence
of biochemical inhibitors of caspase-3, caspase-8, caspase-9, or pan-caspase and then
analyzed for apoptosis using WST-1 assay. Data are mean ± SE (n=3; p < 0.05, for control
vs combination treated samples).
Das et al. Page 21
Br J Haematol. Author manuscript; available in PMC 2016 December 01.
Author M
anuscriptA
uthor Manuscript
Author M
anuscriptA
uthor Manuscript

Figure 4. Mechanisms mediating anti-MM activity of marizomib plus pomalidomide(A) MM.1S cells were pretreated with or without pomalidomide for 12 h, and then
marizomib was added for an additional 12 h; cells were harvested and cytosolic extracts
were then analysed for chymotrypsin- like (CT-L), caspase-like (C-L) and trypsin-like (T-L)
proteasome activities. Results are represented as percentage inhibition in proteasome
activities in drug-treated versus vehicle control. Data are mean ± SE (n=3; p < 0.05, for
single agent vs combination treated samples) (B) Total protein lysates from the indicated
MM cell lines and normal healthy donor PBMCs were subjected to immunoblot analysis
with anti-CRBN or anti-GAPDH Abs. (C) MM.1S cells were transfected with siRNA-
Das et al. Page 22
Br J Haematol. Author manuscript; available in PMC 2016 December 01.
Author M
anuscriptA
uthor Manuscript
Author M
anuscriptA
uthor Manuscript

CRBN or scr-siRNA for 24h. Transfected cells were pretreated with or without
pomalidomide for 24h; marizomib was then added for an additional 24h, followed by
analysis for apoptosis using WST-1 assay. Data are mean ± SE (n=3; p < 0.05, for single
agent vs combination treated samples) Immunoblot shows CRBN expression in cells
transfected with scr-siRNA or CRBN-siRNA. (D and E) MM.1S cells were treated with
indicated agents (as treated in panel ‘A’); protein lysates were subjected to immunoblot
analysis with anti-CRBN, anti-IRF4, anti-MYC, anti-MCL1, anti-HSP90AA1, anti-
HSPA1A, anti-HSPB1 or anti-GAPDH Abs.
Das et al. Page 23
Br J Haematol. Author manuscript; available in PMC 2016 December 01.
Author M
anuscriptA
uthor Manuscript
Author M
anuscriptA
uthor Manuscript
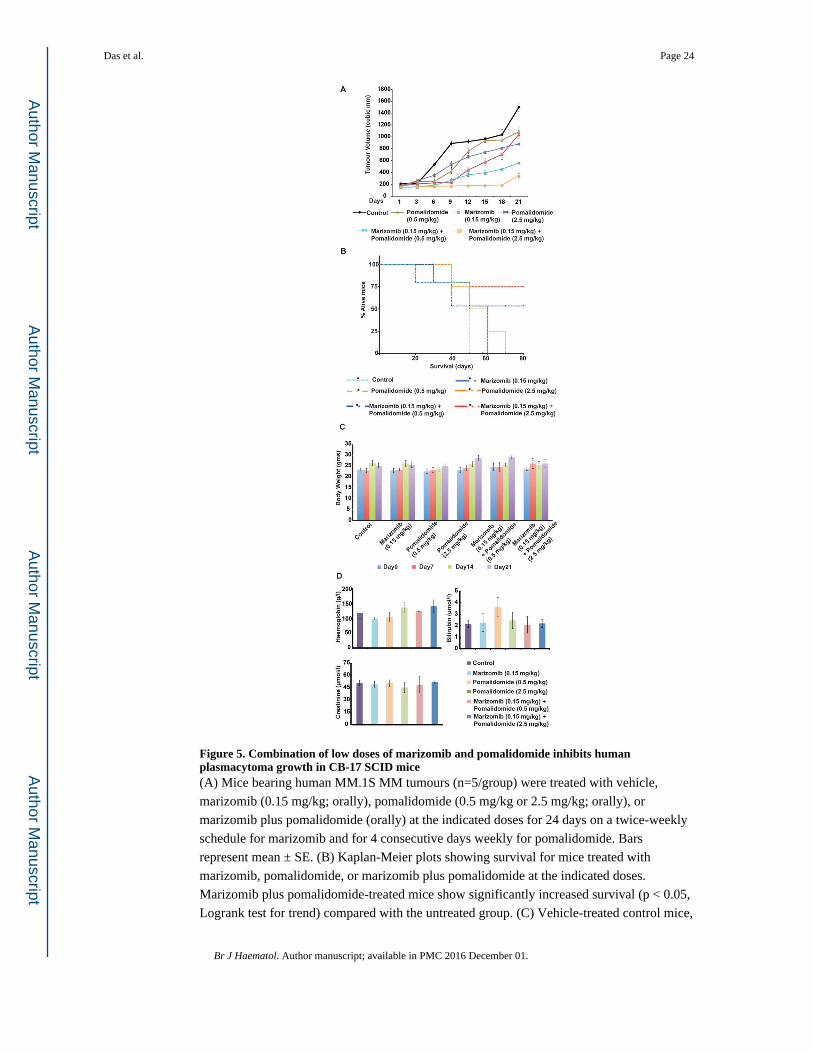
Figure 5. Combination of low doses of marizomib and pomalidomide inhibits human plasmacytoma growth in CB-17 SCID mice(A) Mice bearing human MM.1S MM tumours (n=5/group) were treated with vehicle,
marizomib (0.15 mg/kg; orally), pomalidomide (0.5 mg/kg or 2.5 mg/kg; orally), or
marizomib plus pomalidomide (orally) at the indicated doses for 24 days on a twice-weekly
schedule for marizomib and for 4 consecutive days weekly for pomalidomide. Bars
represent mean ± SE. (B) Kaplan-Meier plots showing survival for mice treated with
marizomib, pomalidomide, or marizomib plus pomalidomide at the indicated doses.
Marizomib plus pomalidomide-treated mice show significantly increased survival (p < 0.05,
Logrank test for trend) compared with the untreated group. (C) Vehicle-treated control mice,
Das et al. Page 24
Br J Haematol. Author manuscript; available in PMC 2016 December 01.
Author M
anuscriptA
uthor Manuscript
Author M
anuscriptA
uthor Manuscript

as well as mice in the marizomib, pomalidomide, or marizomib plus pomalidomide-treated
cohorts, were weighed every week. The average changes in body weight are shown. (D)
Mice were treated with vehicle, marizomib, pomalidomide, or marizomib plus
pomalidomide (as in panel ‘A’) for 24 days; blood samples were then obtained and
subjected to analysis for serum bilirubin, hemoglobin, and creatinine levels using
Quantichrom Creatinine, Bilirubin, and Haemoglobin Assay kit (BioAssay Systems,
Hayward, CA, USA).
Das et al. Page 25
Br J Haematol. Author manuscript; available in PMC 2016 December 01.
Author M
anuscriptA
uthor Manuscript
Author M
anuscriptA
uthor Manuscript

Figure 6. Effect of marizomib plus pomalidomide on apoptosis, MCL1, IRF4 and neovascularization in vivo in xenografted MM tumours(A and B) Apoptotic cells in sections of tumours harvested from vehicle control-,
marizomib-, pomalidomide-, or marizomib (0.15 mg/kg) plus pomalidomide (2.5 mg/kg)-
treated mice were identified by immunostaining for activated caspase-3 (green cells) or
TUNEL. Tumour sections were obtained on day 24. Images were obtained with a Leica
SP5X laser scanning confocal microscope (40x magnification). (C) Tumours harvested from
mice (as in panel ‘A’) were immunostained with Ki67 Abs (40x magnification). (D) Tumour
lysates from control and drug-treated mice were subjected to immunoblot analysis using
anti-MCL1, anti-IRF4, or anti-GAPDH Abs. Lanes 1 to 6 represent lysates of tumours
Das et al. Page 26
Br J Haematol. Author manuscript; available in PMC 2016 December 01.
Author M
anuscriptA
uthor Manuscript
Author M
anuscriptA
uthor Manuscript

harvested from mice receiving the following treatments: lane 1, vehicle alone (control); lane
2, marizomib (0.15 mg/kg); lane 3, pomalidomide (0.5 mg/kg); lane 4, pomalidomide (2.5
mg/kg); lane 5, marizomib (0.15 mg/kg) plus pomalidomide (2.5 mg/kg); and lane 6,
marizomib (0.15 mg/kg) plus pomalidomide (0.5mg/kg). (E and F) Tumours harvested from mice (as in panel ‘A’) were immunostained with Factor VIII or VEGFR1 Abs. Images
were obtained with a Leica SP5X laser scanning confocal microscope (40x magnification).
Photographs (A, B, C, E, and F) shown are representative of similar observations in 2 mice
receiving the same treatment.
Das et al. Page 27
Br J Haematol. Author manuscript; available in PMC 2016 December 01.
Author M
anuscriptA
uthor Manuscript
Author M
anuscriptA
uthor Manuscript





























