Embed Size (px)
Citation preview

THE BIOSYNTHESIS OF STARCH IN SUGAR CANE
Alex G. Alexander
Agric$~ltuval Experiment Station, Ufiivevsity of Puevto Rico, R io Piedras, P.R.
INTRODUCTION
Althougll starch poses a problem in the factory by retarding filtration and crystalliza- tion rates of cane juices (Balch, 1953; Feulherade, ~gzg) , little attention has been given the mechanisms of starch formation in sugarcane. Of further importance is the knowledge that both glucose-I-phosphate* and uridine diphospl~oglucose, themselves major precursors of sucrose, are also involved in starch biosynthesis. The process of starch formation might .thus compete with mechanisms of sucrose synthesis.
In 1937 Cori and coworkers demonstrated that G-I-P was produced during the breakdown of glycogen by muscle extracts, and in so doing greatly advanced the un- derstanding of polysaccharide transformations. Prior to that time, scientists believed that glycogen breakdown was mediated by amylases, hydrolytic enzymes which offered no reasonable explanation for polysaccharide formation. In 1940, Hanes discovered a phosphorylase which promotes the synthesis and degradation of starch in the pea seed and potato tuber. Catalyzing the reaction
H,PO, + Starch G-I-P
starch phosphorylase is now known to be widely distributed in plant tissues. Other enzymes related to polysaccharide transformation in plants include tbe Q enzyme or branching enzyme (Gilbert et al., 1949, 1952; Alexander, 1 9 6 5 ~ ) ~ and the R enzyme (Hanes, 1940; Nelson and Rines, 1962).
More recently, Leloir and coworkers have demonstrated the formation of polysaccharide, by a bean starch granule preparation, according to the following I
I il UDPG $. Acceptor UDP + Alpha (I -+ 4) glucosyl-acceptor I Starch or a di or oligosaccharide of the maltose series was found to serve as the glucose acceptor. Akazawa et al. (1964) have studied a similar reaction with rice-grain prepara- tions.
The present report deals with starch formation in sugarcane, with special emphasis given to the properties of a cane-leaf starch pl~ospl~orylase.
EXPERIMENTAL PROCEDURE ,
I Enzyme preparation I I I
i
I
Plant materials from the varieties M. 336 and Uba Marot were prepared and extract-
* Abbreviations: Glucose-I-phosphate (G-I-P), urldine diphosphoglucose (UDPG), uridine triphosphate (UTP), uridine diphosphate (UDP), flavin adenine dinucleotide (FAD), adenosine triphosphate (ATP), and mlcromoles per rnillillter (pmoles/ml).

626 AGRICULTURE 1 1 ed in accordance with methods outlined previously (Alexander, 1g64a). This involved quick-freezing of leaves +2 and +3* in a mixture of dry ice and acetone, freeze drying and grinding at z°C, and extraction with buffer or distilled water. Protein was precipitated from clarified extracts with ammonium sulfate (pH 7.0, 22°C). Starch phosphorylase was not stable at room temperature, and, consequently, both undialyz- ed samples and those undergoing dialysis were maintained at 2°C.
Enzyme assay
Starch formation was determined colorimetrically by measuring the increased blue value of digests followin~treatment by potassium iodide-iodine solution. The proce- dure was particularly effective for measuring the sugarcane enzyme and has been used to follow the phosphorylase of potato (Arreguin-Lozano and Bonner, 1949) although a more common method involves the measurement of increased inorganic phosphate (Cori, 1940; Hanes, 1940; Green and Stumpf, 1942) .
The standard digest for starch phosphorylase was composed as follows: 0.5 ml , of o.rM succinate buffer (pH 6.25) ; G-I-P, sodium molybdate, and potassium iodide solutions sufficient to make 10, o.ooo~ and 0.002 ,umoles/ml of digest, respectively; 0.1 1111 of 2 percent soluble starch solution; and 0.5 ml of enzyme preparation con- taining 0.5 to 0.7 mg of protein. Final volume of the digest was 3.0 ml. All tests were run in duplicate or triplicate, with control vessels receiving all components of the digest except G-I-P. A second set of controls was maintained with G-I-P, starell, and . distilled water, in the absence of enzyme, to correct for an apparent slight interference by G-I-P with the starch-iodine reaction. Molybdate was added to inhibit phosphatase (Burma and Mortimer, 1956), and iodide to suppress the action of amylase (Alexandef-, 1965f).
The standard reaction was run for 25 minutes at 30°C, and thereupon was ter- minated with 2 ml of 10 percent trichloroacetic acid (TCA). The digest-TCA mixture was centrifuged at 3000 rpm for 10 minutes. To 0.5 ml of the supernatant liquid was added 0.5 ml of potassium iodide-iodine solution**, followed by 2 ml of distilled water. After allowing five minutes for color development, optical density of the solu- tion was measured at 580 mp with a Beclrman Model B Spectrophotometer. The amount of polysaccharide formed was determined by reference to a standard curve representing 0.01 to 0.25 mg of soluble potato starch.
The starch phosphorylase activity unit was arbitrarily defined as the amount of enzyme catalyzing the formation of 0.10 mg of starch under the prescribed condi- tions of the assay. Protein was determined colorimetrically by the technique of Sutherland et al. (1959). All phosphorylase activity measuren~ents were recorded in terms of specific activity (units per mg of protein).
* Other tissues sampled included ineristem, leaves I and o, leaves +6 and 4-7, sheaths $ 2 and +3, 8, to 10 nodes and internodes, and I to 3 nodes and internodes. All tissues were frozen, ground, and extracted in a manner identical to that described for leaves (Alexander, 19644.
** A stock iodine solution was prepared by dissolving o 20 g of iodine in IOO ml of potassium iod~de solution (8.3 g of KI dissolved In distilled water and brought to IOO ml). This peparation was stable. Dilutions (I : 10) were prepared daily for immediate use.
A. G. ALEXANDER 627
RESULTS AND DISCUSSION
I. Salt fractionation of cane-leaf extracts
Although starch phosphorylase is not a difficult system to measure, achievement of substantial purity has posed a definite problem. Green and Stumpf (1942) settled upon four successive series of fractionation with ammonium sulfate, after unsatis- factory efforts to obtain potato phosphorylase by alumina absorption. They also found that inactivation accompanied prolonged dialysis of the potato enzyme against distilled water. Hanes (1940) succeeded in removing considerable inert material from potato phosphorylase extracts by absorption upon alumina and kaolin. This prepara- tion still contained amylase, but more satisfactory results were gained by ammonium sulfate precipitation between 30 and 50 percent saturation. Arreguin-Lozano and Bonner (1949) also employed ammonium sulfate for obtaining potato phosphorylase.
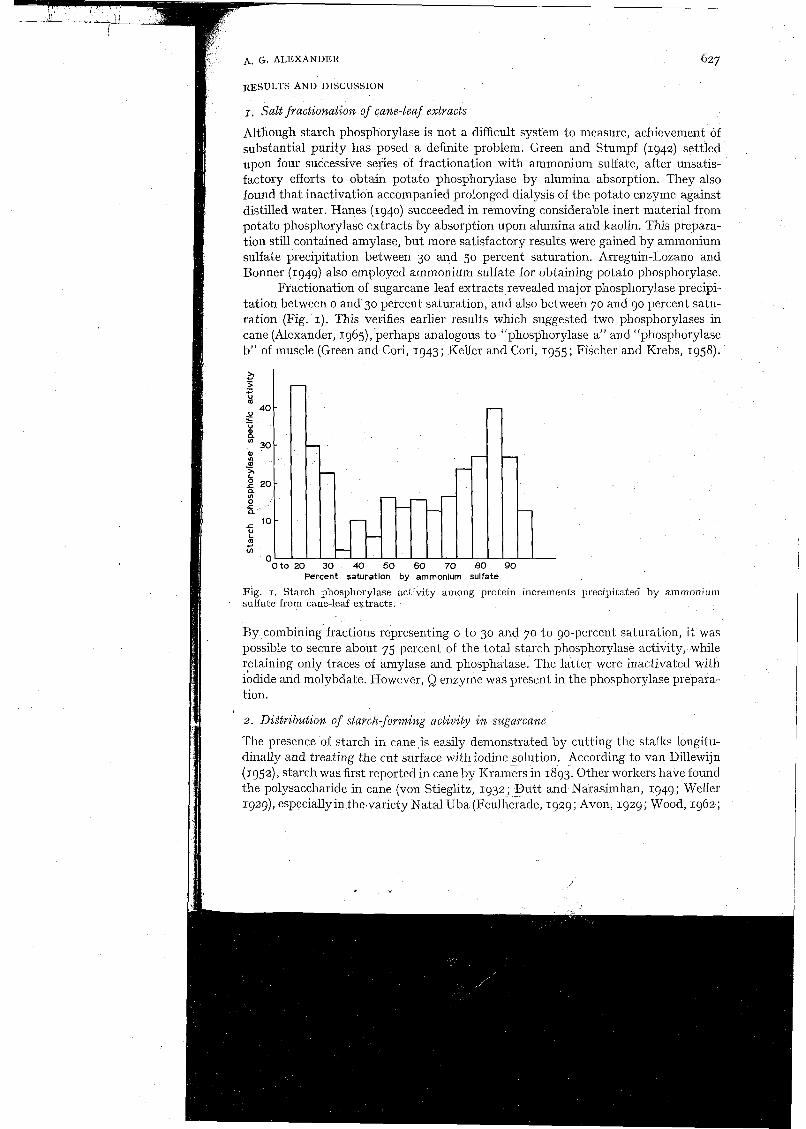
Fractionation of sugarcane leaf extracts revealed major phosphorylase precipi- tation between o and 30 percent saturation, and also between 70 and 90 percent satu- ration (Fig. I). This verifies earlier results which suggested two phosphorylases in cane (Alexander, 1965), perhaps analogous to "phosphorylase a" and "phosphorylase b" of muscle (Green and Cori, 1943; Keller and Cori, 1955; Fischer and Krebs, 1958).
Percent saturation by ammonium sulfate
Fig. I . Starch phosphorylase actlvity among protein increments precipitated by ammonium sulfate from cane-leaf extracts.
By combining fractions representing o to 30 and 70 to 90-percent saturation, it was possible to secure about 75 percent of the total starch phosphorylase activity, while retaining only traces of amylase and phosphatase. The latter were inactivated with iodide and molybdate. However, Q enzyme was present in the phosphorylase prepara- tion.
2. DiStribution of starch-forming activity in sugavcane
The presence of starch in cane is easily demonstrated by cutting the stalks longitu- dinally and treating the cut surface with iodine solution. According to van Dillewijn (1952), starch was first reported in cane by ~ r a m e r s in 1893. Other workers have found the polysaccbaride in cane (von Stieglitz, 1932; Dutt and Narasimhan, 1949; Weller 1929), especially in the variety Natal Uba (~eulherade, 1929; Avon, 1929 ; Wood, 1962 ;
AGRICULTURE
Haddon, 1927). Weller (1929) describes two types of starch accun~ulation in sugar cane. Traces of starch have been detected in the varieties Uba Marot and M. 336 a t this laboratory.
Starch is almost totally insoluble, and, therefore, its discovery in any given tissue should indicate the presence of a starch-forming mechanism. Actually, the appearance of starch might simply reflect an absence of amylase, and does not neces- sarily tell us where the richest source of starch phosphorylase lies. Table I summarizes the distribution of starch phosphorylase in mature plants of the variety Uba Marot, both in terms of specific activity and total starch-forming capacity per gram of tissue.
TABLE I
ACTIVITY O F STARCH PHOSPHORYLASE AMONG DIFFERENT TISSUES O F TWELVE MONTHS OLD SUGAR CANE
Tissue source Data Meristem Leaf classification (-1
and o)
Leaf Leaf Sheath Node Inter- Node Inter- ( + 2 (+6 ( + 6 (8-10) node (1-3) node
and +3) and f 7) and +7) (8-10) (1-3)
Specific activity 13.9 16.7
Calculated potential (tng of starchlg of tissue/zo min) 55.6 12.5
Greatest specific activity was recorded among internodes I to 3 and 8 to 10. However, so little protein was present in these tissues that their overall potential for starch synthesis was very low. Meristem revealed moderate specific activity, but high polysaccharide-forming potential, in view of its extremely high content of catalytic protein. But again, there is yery little meristem tissue in sugar cane. Leaves -I to +3 were ultimately selected as the source of starch phosphorylase for the present studies.
3. P~o$erties of calze starch phos$lzovylase
( a ) Stability and effects of dialysis When dealing with any enzyme preparation the investigator must know how long it can be reliably used, and which storage conditions will best preserve the active cata- lyst. Starch phosphorylase measured during the present investigations had already survived processes of quick freezing, freeze drying, grinding by metallic blades, stor- age within dried powder, water extraction and salt fractionation. Additional tests were conducted to learn the enzyme's stability when maintained under toluene and stored at room and refrigerator temperatures (Table 2).
TABLE 2
SPEClFIC ACTIVITY DECLINE O F CANE STARCH PHOSPHORYLASE STORED AT ROOM AND REFRIGERATOR
TEMPERATURES ' Storage Storage time (days) temperature ("C) o I 3 9
* Enzyme protein was dissolved in distilled water and stored dnder toluene.

A. G. ALEXANDER 629
Cane phosphorylase was slightly more stable at 2°C than 2z°C, yet activity declined marlsedly within one day of preparation. Consequently, the enzyme was prepared fresh daily througl~out these studies.
Dialysis of the enzyme preparation against distilled water caused about 87 per- cent of the activity to be lost within 8 hours (Table 3). Undialyzed preparations were used for the majority of subsequent tests.
TABLE 3
DECLINE O F STARCH PHOSPHORYLASE ACTIVITY OF CANE FOLLOWING DIALYSIS AGAINST DISTILLED
WATER
Dlalysis tune (hours) Data classification o 112 I 2 4 6 8 Specific actlvity 15.2 10.8 5.2 3.6 2.8 2.0 1.0
Investigators have not gained consistent results with phosphorylase, even when working with a single plant species. Green and Stumpf (1942) reported that potato phosphorylase was stable for several days at 0°C in dilute ammonium sulfate solu- tion (pH 6 to 7), wllereas deterioration of activity required a period of weeks. Dialysis of the enzyme caused serious inactivation only after prolonged periods (greater than 12
hours). Arreguin-Lozano and Bonner (1949) claimed that long contact with ammo- nium sulfate causes inactivation of the potato enzyme. Hanes (1940) also working with a potato source, found that dialysis against distilled water completely inactivated phosphorylase. On the other hand, Hanes found that potato phosphorylase in solutions containing 1.5 to 3 percent ammonium sulfate was quite stable, losing only around 10 percent of activity in 14 to 20 days, at 0°C.
Differences in behavior of potato phosphorylase is understandable when by necessity the plant materials are obtained froin local markets. Here such information as the potato variety, age, cultural practices during its growth, manner of harvest, conditions of storage, etc., are irretrievably lost, although such factors bear heavily on the properties of enzymes. Regarding sugar cane phosphorylase, it is felt at least one contaminant enzyme 'is present in the preparation which directly affects the polysaccl~aride-forming catalyst. This enzyme might be another phosphorylase, or a system such as that described by Iceller and Cori (1955) capable of cleaving "phospho- rylase a" into halves (the PR enzyme).
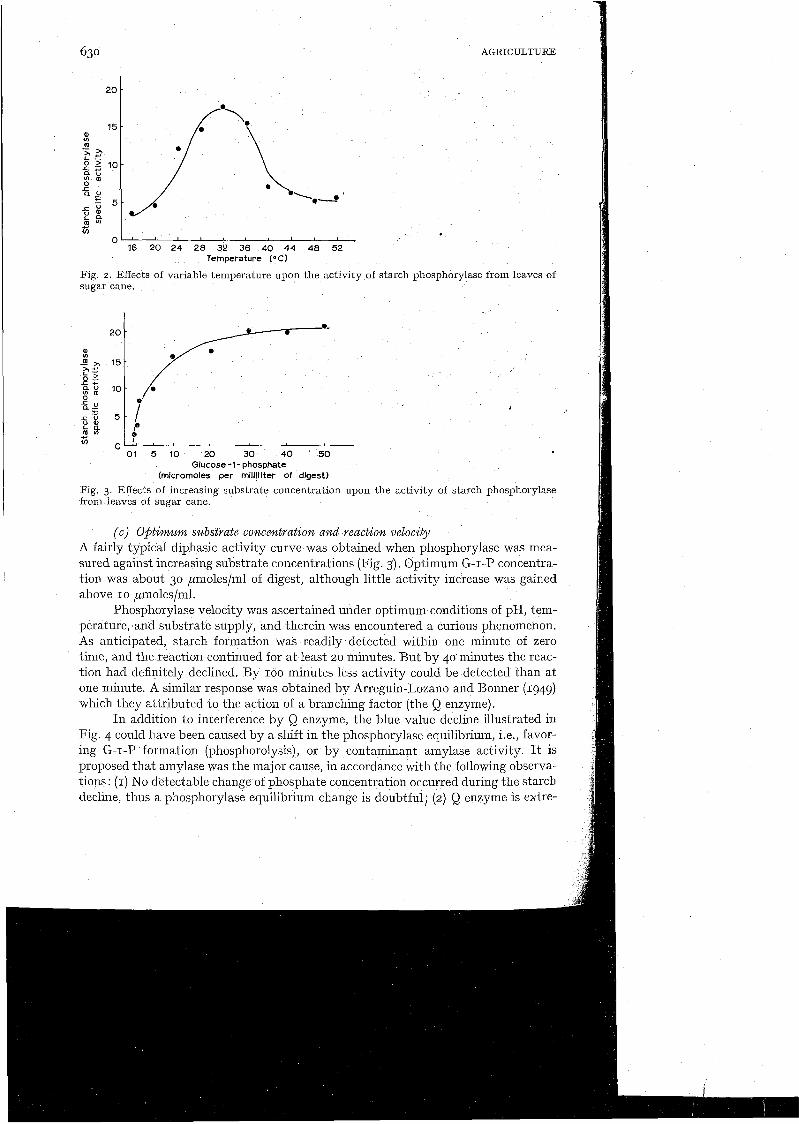
( b ) Effects of variable $H and temperature Cane-leaf preparations revealed no phosphorylase activity below pH 4.5, whereas an apex was achieved at pH 6.25 (Table 4). Activity declined somewllat above pH 6.25, but a moderately strong reaction could still be measured at pH 9.0. Optimum tkmperature for the reaction was about 32°C (Fig. 2).
TABLE 4
EFFECTS'OF VARIABLE pH UPON STARCH PHOSPHORYLASE FROM LEAVES O F IMMATURE SUGAR
CANE
pH* Data classification 3.5 4.04.5 5.05.5 6.0 6.25 6.50 6.75 7.0 7.5 8.0 Specific activity o o 3.8 8.0 12.4 20.8 30.4 20.4 1812 16.8 16.2 13.2 13.0 14.0
* The following buffers were employed: Acetate, p H 3.5-5.5; Succinate, p H 6.0-7.0; and tris (trishydroxymethylalnino methane) pH 7.5-9.0.
Temperature ('C)
Fig. 2. Effects of variable temperature upon the activity sugar cane.
starch phosphorylase from leaves of
Glucose -1 - phosphate (mlcromoles per mllllliter of dlgest)
Fig. 3 . Effects of increasing substrate concentration upon the activity of starch phosphorylase from leaves of sugar cane.
1 (c ) O$timum substrate co.ncentration and reaction velocity A fairly typical diphasic activity curve was obtained when phosphorylase was mea- sured against increasing substrate concentrations (Fig. 3). Optimum G-I-P concentra- tion was about 30 ,umoles/ml of digest, althougll little activity increase was gained above 10 ,umoles/ml.
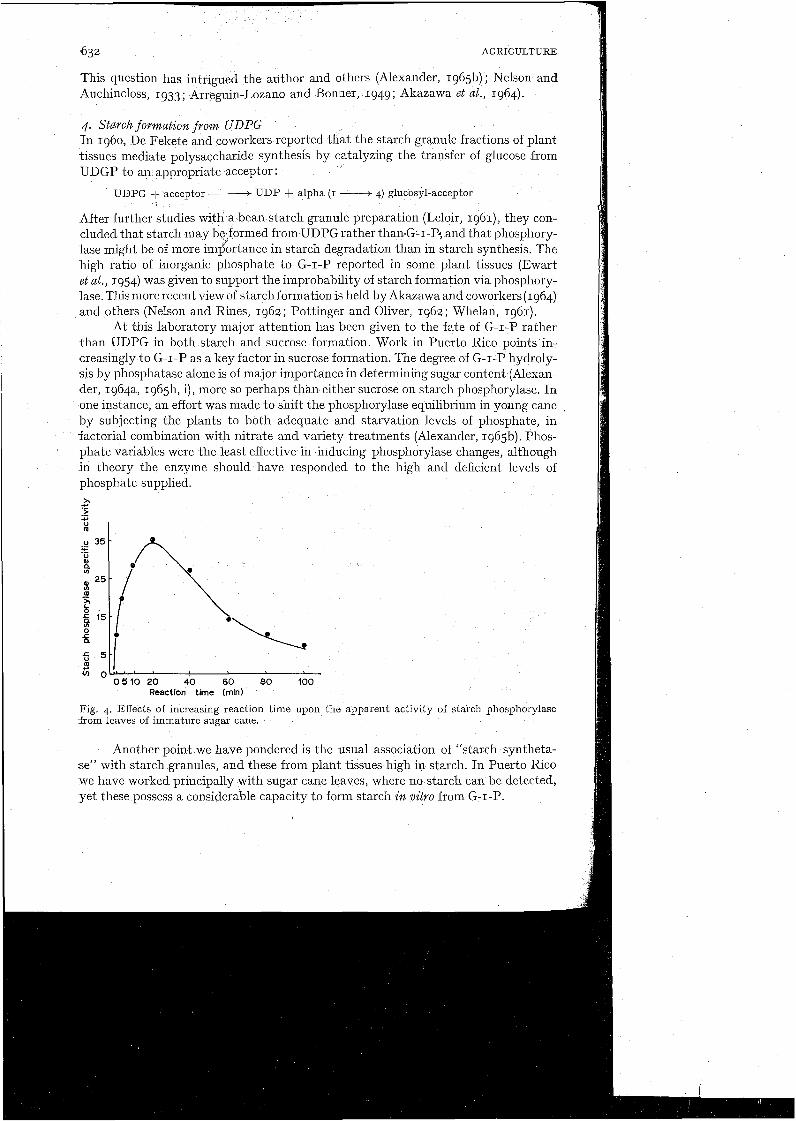
Phosphorylase velocity was ascertained under optimum conditions of pH, tem- perature, and substrate supply, and therein was encountered a curious phenomenon. As anticipated, starch formation was readily detected within one minute of zero time, and the reaction continued for at least 20 minutes. But by 40 minutes the reac- tion had definitely declined. By IOO minutes less activity could be detected than at one minute. A similar response was obtained by Arreguin-Lozano and Bonner (1949) which they attributed to the action of a branching factor (the Q enzyme).
In addition to interference by Q enzyme, the blue value decline illustrated in Fig. 4 could have been caused by a shift in the phosphorylase equilibrium, i.e., favor- ing G-I-P formation (phosphorolysis), or by contaminant amylase activity. I t is proposed that amylase was the major cause, in accordance with the following observa- tions : (I) No detectable change of phosphate concentration occurred during the starch decline, thus a pl~ospl~orylase equilibrium change is doubtful; (2) Q enzyme is extre-

A. G. ALEXANDER 63r
mely scarce in leaves -0 to +3 (Alexander, 1g65c), and could hardly have caused interference of the magnitude actually encountered; (3) starch decline was accompa- nied by an increase of reducing sugar in the digest.
The amylase suspected here could not be that which abundantly occupies cane- leaf tissues (Gilbert and Patrick, 1952). More than sufficient iodide was provided to totally inhibit that catalyst. Rather, a less prominent enzyme, an amylase closely associated with the Q enzyme, is more logically suspected. While studying the Q enzyme (Alexander, 1965~) we encountered an acid amylase which is apparently dormant in the absence of branched polysaccharide, since it became active only after the Q enzyme reaction had progressed ro to 20 minutes. This enzyme is logically classed as one of an alpha type (capable of acting upon the branched structure of amylopectin). I t is possible that during the present study enough amylopectin was formed by contaminant Q enzyme to initiate an alpha-amylase, which in turn catalyz- ed the destruction of starch (amylose plus amylopectin) observed during the phos- phorylase velocity measurements.
( d ) E f e c t s of additives A series of additives were tested for possible activator or inhibitor effects upon cane phosphorylase. These included magnesium (Mg), calcium (Ca), boron (B), fluoride (F) , bromide (Br) , iodide (I), arsenate (As), copper (Cu) , manganese (Mn) , cyanide (CN), zinc (Zn), and beta-glycerophosphate (P-GP). As indicated in Table 5, none of the factors caused enzyme inhibition at the tested rate of 1.0 ,umole/ml of digest.
TABLE 5
EFFECTS O F ADDITIVES UPON THE SPECIFIC ACTIVITY O F STARCH PHOSPHORYLASE FROM LEAVES O F
SUGAR CANE
Data Additive* classification Control*= Mg Ca B F Br I 4 s Cu &!In Mo CN Zn P-GP Specific activity 15.4 r4;9 21.3 16.5 24.5 21.8 18.6 18.6 17.0 13.3 18.6 21.8 15.4 21.8
* Each addltlve was supplied a t the rate of 1.0 ,~lmole/ml of dlgest. ** Control vessels received distilled water in place of addltive solution.
Fluoride caused an apparent activity increase. The F effect lends support to our con- tention that a Q enzyme-alpha amylase complex interferes with cane phosphorylase, since F was the most effective inhibitor discovered for the cane alpha-amylase (Alexander, 1965~).
Some increased activity was recorded in the presence of Ca, Br, CN, and P-GP. Green and Cori found that reducing agents increased solubility of muscle phosphory- lase but that KCN was less effective than glutathione and cysteine. According to Qruton and Simmonds (1963) P-GP is anoncolnpetitive inhibitor of muscle phosphory- lase a, although Green and Cori (1943) used P-GP to neutralize solutions and to create buffers for dialysis procedures during the preparation of crystalline phosphorylase. To the kuthor's knowledge, the most effective compound for altering the activity of cane phosphorylase is the substituted urea Diuron [3-(3,4-dicl11orophenyl)-I,I dimethylurea], which, at a concentration of 0.00256 ,umole/ml, caused nearly a three-fold activity increase (see p. 584). Curiously, Diuron was found to greatly inhibit the sucrose-forming mechanism of cane, thereby raising once again the question of direct relationships between sucrose and starch-forming pathways in higher plants.
632 AGRICULTURE
This question has intrigued the author and others (Alexander, 1965b); Nelson and Auchincloss, 1933; Arreguin-Lozano and Bonner, 1949; Ahazawa et al., 1964).
4. Starch f o r m a t i o ~ from UDPG In I ~ G o , De Felcete and coworkers reported that the starch granule fractions of plant tissues mediate polysaccharide synthesis by catalyzing the transfer of glucose from UDGP to an appropriate acceptor:
UDPG + acceptor UDP + alpha (I -4 4) glucosyl-acceptor
After further studies with a bean starch granule preparation (Leloir, 1961), they con- cluded that starch may be formed from UDPG rather than,G-I-P: and that phosphory- lase might be of more inldortance in starch degradation than in starch synthesis. The high ratio of inorganic phosphate to G-I-P reported in some plant tissues (Ewart et al., 1954) was given to support the improbability of starch formation via phosphory- lase. This more recent view of starch formation is held by Akazawa and coworkers (1964) and others (Nelson and Rines, 1962; Pottinger and Oliver, 1962; Whelan, 1961).
At this laboratory major attention has been given to the fate of G-I-P rather than UDPG in both starch and sucrose formation. Worlc in Puerto Rico points in- creasingly to G-I-P as a key factor in sucrose formation. The degree of G-I-P hydroly- sis by phosphatase alone is of major importance in determining sugar content (Alexan- der, 1964a, 196511, i), more so perhaps than either sucrose on starch phosphorylase. In one instance, an effort was made to shift the phosphorylase equilibrium in young cane ,
by subjecting the plants to both adequate and starvation levels of phosphate, in factorial combination with nitrate and variety treatments (Alexander, 196513). Phos- phate variables were the least effective in inducing phosphorylase changes, although in theory the enzyme should have responded to the high and deficient levels of phosphate supplied.
Reaction time (mln)
Pig. 4. Effects of increasing reaction time upon the apparent activity of starch phosphorylase from leaves of immature sugar cane.
Another point we have pondered is the usual association of "starch syntheta- se" with starch granules, and these from plant tissues high in starch. In Puerto Rico we have worked principally with sugar cane leaves, where no starch can be detected, yet these possess a considerable capacity to form starch ifi vitvo from G-I-P.

A. G. ALEXANDER 633
For the sake of biochemical perspective, some distinction needs to be drawn between transitionary $olysaccharide and the storage polysaccharide which may more rightfully be termed starch. In such tissues as the bean seed, rice grain, and potato tuber, starch-forming mechanisms may be relatively free from voracious amylases such as those in cane. The synthesis and accumulation of starch here is both logical and readily recognized. With cane leaf preparations, however, even under ideal i n vitro conditions for starch synthesis, the newly formed polysaccharide cannot be maintained much longer than 20-40 minutes (Fig. 4). I n vivo this product is probably
I hydrolyzed instantaneously. The subsequent failure of starch to accumulate can be incorrectly interpreted to mean that starch formation is of little consequence in cane. Yet a major concern with cane is the fate of the sucrose precursor G-I-P. From this standpoint, it must be emphasized that a transfer of the glucose radicle to polysaccha- ride, albeit only for a split second, still suffices to deplete G-I-P reserves as certainly as if glucose had entered permanent storage.
TABLE 6
STARCH FORMATION BY UNDIALYZED AND DIALYZED CANE ENZYME PREPARATIONS I N THE PRESENCE O F VARIABLE ADDITIVES
Exp. No. Item No. Digest composition Starch formation (unitslmg of protein)
I. Undialyzed enzyme
Enzyme, activator, G-I-P, plus H,O I
Enzyme, activator, plus 1.0 pmole/ml of UDPG Enzyme, activator, plus 10.0 pmoles/ml of UDGP Enzyme, activator, G-I-P, plus 1.0 pmole/n~l of UTP Enzyine, activator, G-I-P, plus 10.0 /~moles/ml of UTP Enzyme, activator, G-I-P, 1.0 pmole/inl of UDP,
plus 0.5 pinolelml of ATP Enzyme, actlvator, G-I-P, plus 10.0 /~inoles/ml of UDP,
plus 5.0 pmoles/ml of ATP Enzyme, activator, G-I-P, plus 0.1 pmole/ml of PAD
2. Dialyzed g Enzyme, activator, G-I-P, plus H,O 6.9 enzyme* ro Enzyme, activator, plus 1.0 pmole/ml of UDPG 26.7
I I Enzyme, activator, plus 10.0 pmoles/inl of UDPG 2.7 12 Enzyme, activator, G-I-P, plus 1.0 ,umole/inl of UTP 10.7 I3 Enzyme, activator, G-I-P, plus 10.0 ,umoles/ml of UTP 3.6 I4 Enzyme, activator, G-I-P, plus 1.0 pmole/inl of UDP,
plus 0.5 pmolelml of ATP 19.8 15 Enzyme activator, G-I-P, plus 10.0 pinoles/ml of UDP,
plus 5.0 ,urnoles/ml of ATP 6.9 I 6 Enzyme, activator, G-I-P, plus 0.1 ,~cinole/inl of FAD 18.6
* Enzyme preparations for experiment 2 were dialyzed for five hours against distilled water.
I t was of interest to learn whether or not cane-leaf preparations would catalyze . starch formation from UDPG as well as G-I-P (i.e., via "starch synthetase"). To this
end, a series of additives consisting of UDPG, UTP, UDP, ATP, and FAD were tested in the presence of dialyzed and undialyzed enzyme protein (Table 6). These
I enzyme preparations were identical to those used in cane phosphorylase experiments. Undialyzed enzyme catalyzed moderately strong starch phosphorylase in the
presence of G-I-P activator, and water (Table 6, treatment no. I). Additives served only to inhibit this activity, with inhibition being complete in the presence of 10.0

,umoles/ml of UDPG, 10.0 pmoles/ml of UDP plus 5.0 pmoleslml of ATP, and 0.1
,umole/ml of FAD. Dialyzed enzyme, on the other hand, caused an anticipated decline of phosphorylase, while permitting strong starch-forming activity in the presence of 1.0 ,umole/nil of UDPG. Of further significance was the stimulated activity when UTP, or UDP plus ATP, were added to the standard digest (Table 6, items 12 and 14). Thus, some provision for the transfer of glucose from UDPG to polysaccharide appears t o be present in cane. Very likely this is an enzymatic mechanism similar to the "starch synthetase" described by Leloir and coworkers (De Fekete, 1960; Leloir, 1961) .The evidence also points to a mechanism for the synthesis of UDPG, possibly via the reaction
UTP + G-I-P + UDPG + UDP
Thirdly, the formation of starch from a combination G-I-P, UDP, ATP, and activator (Table 6, treatment 14)~ suggests that the following sequence of reactions is promoted by the dialyzed cane preparation:
UDP + ATP , UTP + ADP UTP + G-I-P L- UDPG + UDP UDPG + activator ---- 4 polysaccharide + UDP
Reactions (I) and (2) are recognized in higher plants (Cardini et al., 1955; Burma and Mortimer, 1956; Turner, 1957; Dugger and Humphreys, 1960; Paladini and Leloir, 1952), including sugarcane (Alexander, 1964b; Hatch et.al., 1963), while reaction (3) is equivalent to that described by the Leloir group (1960, 196~) and Akazawa et al.' '(1964).
Table 6 emphasizes the importance of dialysis in distinguishing between two types of starch formation. These mechanisms would not seem to proceed happily together, since the dialysis step which made the UDPG system effective simultaneous- ly eliminated active starch phosphorylase. I t is suspected that an endogenous inhibi- tor is present in cane which prevents some step of the UDPG mechanism, and that phosphorylase requires an activator. Both are presumably lost during dialysis.
An interesting sidelight is suggested by the work of Leloir et al. (1961). They concluded that since the starch-synthesizing system takes place in whole grains, the in vitvo structural relationship between enzyme and polysaccharide is the same as in the intact plant tissue. Taking this a step further, it is obvious that our own cane phosphorylase preparations represent an artificial combination of cell constituents from disrupted plant tissues, including all potential inhibitors, irregardless of their in vivo role or previos localization. Confinement of an enzyme complex within whole grains is precisely the type of isolation needed to avoid such inhibitors, thereby making possible the formation of starch from UDPG.
Another point of interest is the fact that FAD completely inhibited undialysed phosphorylase (Table 6, item 8), yet caused marked stimulation of phosphorylase after it had been retarded by dialysis (Table 6, item 16). I t is tempting to speculate the role of FAD, but more work is needed before a plausible hypothesis can be put forward. We have previously suggested that FAD might affect glucose supply in cane as an essential cofactor of glucose oxidase (Alexander and Gonzalez-Ibafiez, 1964). I t now appears that FAD may be more closely associated with the fate of G-I-P. This possibility is strengthened by the fact that when activity of the cane sucrose-forming

A. G. ALEXANDER 635
mechanism was suppressed by dialysis, FAD was more effective than any other additive tested in returning the lost activity (Alexander, 1g64b).
5. Significance of starch-forming reactions in cane Starch can be a nuisance in the factory when present in large quantities (Balch, 1953; Feulherade, 1929). The cane variety Natal Uba was especially unfortunate in this respect (Haddon, 1927; Weller, 1929; Anon, 1929; Wood, 1962), although starch has not been considered a serious problem with varieties grown in Puerto Rico.
Perhaps more important is the capacity of cane to form a transitionary poly- saccharide, presumably amylose, rather than to accumulate starch as it is bnown in the potato tuber and starchy seeds. Thus an already limiting G-I-P reserve might be depleted by starch-forming and starch-hydrolyzing reactions in sugar cane, leaving no telltale signs of interference with sucrose production, when in fact such interference could be quite severe.
Results at this laboratory point increasingly toward overall relationships in cane between starch formation, sucrose formation, pathways of hexose metabolism, and the hydrolysis of hexose phosphates. In addition to starch-forming reactions discussed herein, we are also aware of a polysaccharide branching reaction (Alexander, 1g65c) and of enzymes capable of degrading both branched and unbranched polysac- charide. Maltase has also been measured in cane preparations (Alexander and Gon- zalez-Ibafiez, 1964), as was hexokinase, ATP-ase, phosphol~exose isomerase, inver- tases (Alexander, 1965e), and a series of phosphatases attacking hexose phosphates (Alexander, 1964a, 1g65d). A suggested overall relationship of these reactions to one another is illustrated by the following diagram:
4
Glucose UDP + UDPG UDPG + UDP
"Starch" y( )? I
I I 2 +Fruc. b 1b Amylose + Amylopectin +- Amylose + Glucose-I-P ~i Sucrose -+ Fructose + L L4 L lr I5 b 7
Glucose
~ a l t o s e ~li~osac'charides ~ a l t ' o s e 11 Glucose I I + 5 5 Fructose
6 Glucose
-> il I= - Glucose-6-P % Glucose +- I-I,PO, IP
ATP ADP
47 I
Fructose-6-P -% Fructose + H,PO, ADP
4 I Glycolysis and Krebs Cycle
I. Starch phosphorylase 7. ATP-ase 13. Glucose-I-P phosphatase 2. Q enzyme 8. Phosphohexose isomerase 14. UDPG-Fructose transglycosylase 3 . Beta-amylase g. Fructose-6-P phosphatase 15. Sucrose phosphorylase 4. Alpha-amylase 10. Glucose-6-P phosphatase 16. Beta-fructosiclase 5. Maltasc I I. Phosphogluco~nutase 17. Alpha-glucosidase 6. Hexolcinase 12. Starch synthetase

636 AGRICULTURE
Evidence for each of the enzymes indicated above has been encountered at this laboratory, with the exception of phosphogluco mutase. There is evidence also of UDPG formation from UTP and G-I-P, although no specific enzyme measurements have been attempted.
The critical position of G-I-P is at once evident in the above diagram. Numerous reactions appear capable of affecting G-I-P supply, and, without doubt, each of the essential pathways of sucrose formation, starch formation, and sugar metabolism stand to suffer should pools of G-I-P be depleted. In this capacity the inhibition of G-I-P phosphatase alone has been a consistent feature of sucrose promoting treat- ments. Enzymes hydrolyzing glucose-6-P, ATP, and starch have likewise been sup- pressed by treatments favoring high sucrose.
The exact need for powerful amylase in cane is very elusive. I t was proposed that one function of amylase is to insure that glucose is not drawn from active path- ways into storage form (Alexander, 1965a). The degradation of amylose chains back to individual glucose units, with subsequent phosphorylation to glucose-6-P, makes it theoretically possible for some of the glucose to reenter G-I-P reserves via phosphoglu- comutase. On the other hand, amylase suppression has been found associated with high sucrose (Alexander, 1964a, 1965b) which suggests that polysaccharide accumula- tion may tend to shift the starch phosphorylase equilibrium in favor of phosphoroly- sis, subsequently permitting more G-I-P to take part in sucrose-forming reactions.
I SUMMARY
Polysaccharide formation has been studied in sugarcane with particular emphasis given to starch phosphorylase, an enzyme similar to that encountered by earlier workers in extracts of muscle and the potato tuber. Cane phosphorylase catalyzes the production of a polysaccharide, yielding a typical starch-iodine reaction, in the presence of glucose-I-phosphate and a trace of starch activator.
Starch phosphorylase was extracted from freeze-dried tissues with distilled . water and precipitated with ammonium sulfate, primarily between o and 30 percent saturation, and again between 70 and go percent saturation. At least two phosporyla- ses are thus suspected in sugar cane. Variable enzyme activity was present in all tissues tested. Leaves and meristem revealed the greatest starch-forming potential.
The cane starch phosphorylase was unstable in aqueous solution and was rapidly inactivated by dialysis against distilledwater. Optimum pH was 6.25. No activity was detected below pH 4.5, but the enzyme was moderately active at pH values as high as 9.0. Optimum temperature was about 32°C. Approximately 30 micromoles of glu- cose-I-phosphate were needed per milliliter of digest to achieve a maximum reaction.
An unusual velocity curve was obtained, under optimum conditions of pH, tem- perature and substrate, in that an initially rapid reaction continued for only 20 to 40 minutes and then sharply declined. The simultaneous production of reducing sugars pointed to an interference by amylase, although adequate inhibitor was present to eliminate any known active amylase. I t was suggested that the cane preparations bear the potential to produce a polysaccharide-destroying catalyst whenever the polysac- charide accumulates above some minimum level.
Cane phosphorylase is readily inactivated by trichloroacetic acid, and ismoderat- ely stimulated by fluoride, calcium, and bromide. During another study it was learned

A. G . ALEXANDER 637
that the substituted urea Diuron greatly stimulates the starch-forming reaction. A second mechanism appears to be present in cane whereby polysaccharide is
formed from uridine diphosphoglucose in the presence of an appropriate acceptor. The reaction proceeds only after the enzyme preparation has been dialyzed, when starch phosphorylase is almost inactivated. Dialyzed enzyme plus uridine diphosphoglucose yielded p~l~saccharide, as did a combination of glucose-I-phosphate and uridine triphosphate with dialyzed enzyme. Furthermore, a combination of glucose-r-phos- phate, uridine diphosphate, and adenosine triphosphate yielded polysaccharide with dialyzed enzyme, indicating that additional mechanisms might be present for the production of uridine triphosphate and uridine diph~spho~lucose.
The significance of starch-forming reactions in cane is discussed with emphasis given to depletion of phosphorylated intermediates needed for other reactions. Glucose-I-phosphate in particular seems hard pressed. I t is possible that too little attention is given to starch in sugar cane since i t seldom accumulates in most cane varieties. In all probability, the accumulation of starch is prevented by amylases, whereas interference with sucrose formation might nevertheless proceed unnoticed.
REFERENCES
AIZAZAWA, T., MINAMIIZAWA, T AND MURATA, T. (1964) Enzymatic mechanism of starch synthesis in ripening rice grains. Plant Physiol., 39 : 371-8,
ALEXANDER, A. G. (1964~) ~ucrose-enzyme relationships in immature sugarcane as affected by varying levels of nitrate and potassium supplied in sand culture. J. Agr. Univ . P. R., 48: 165-231.
-- (1964b) The biosynthesis of sucrose in sugarcane. J . Agr. Uniu. P . R., 48 ( 4 ) : 265-283. ---- (1965a) ~ l ~ ~ s i o l o g i c a l studies of enzyines catalyzing the synthesis and hydrolysis of sucrose,
starch, and phosphorylated hexose in sugarcane. J . Agr. Univ . P . R., 49 (I) : 60-75 --- (1965b), Behavior of enzymes governing starch and sucrose-forming pathways in two sugar-
cane varieties supplied with variable .nitrate and phosphate in sand culture. J . Agr. Univ. P . R., 49 ( 2 ) : 153-175.
---- (1965~) The hydrolytic proteins of sugarcane. I. The Q enzyme. J . Agr. Uvziv. P . R., 49 ( 2 ) : 176-203.
---- (1b65d) The hydrolytic proteins of sugarcane. 11. The acid phosphatases. J . Agr. Unjv. P . R. , 49 (2 ) : 204-228.
---- (1g65e) The hydrolytic proteins of sugarcane. 111. The acid invertases. J . Agr. Univ. P . R. , 49 ( 3 ) : 287-307.
---- (1965f) The hydrolytic proteins of sugarcane. IV. Amylase. J . Agr. Uniu. P. R . , 49 ( 3 ) : 208- 324.
- (19659) Induction of varylng sugar levels In imlnature sugarcane by use of acid phosphatase ~nhibitors J Agt' U n z ~ P R., 49 (1) 35-59
- (1965h) Effects of tungsten and molybdenum upon sucrose content and hydlolytic enzymes of imlnatuie sugarcane J Agt' Unzv P . R , 49 (4) 429-442
- (1967) I n vztvo effects of Dluron [3-(3, 4-dichloropheny1)-I, I d~methylurea] upon enzymes catalyz~ng the synthesis and hydrolysis of sucrose, starch, and phosphorylated hexose in sugarcane Proc I S S C T . (1965). Thls volume, p 582.
ALEXANDER, A. G AND GONZALEZ-IBAREZ, J. (1964) Residual effects of Diu~ on [z-(3, 4-dichloro- phenyl) -I, I d~inethylurea] on the sugar content and sugal-metabol~zing enzymes of
" ratoon sugarcane 25 months following vailable soil tretment J Agr Unzv P R 48 ( 4 ) 284-303.
ANON (192~) Refiort of /hezo?flmzttee in Charge of the Experiment Statlon of the Hawanan Sugar Planters Associat~on, for the yea1 cnding Sept 30, 1929
ARREGUIN-I,OZANO, B AND BONNER, J (1949) Experiments on sucrose forinahon by potato tube~s as influenced by temperature Plant P h y ~ z o l , 2 4 . 720-38.
BALCH, R T. (1953) Further notes on starch in Louls~ana canes, juices and raw sugars. Sugar J , 15 . 11-15
BURMA, D P AND MORTIMER, D. C. (1956) The blospnthesis of uridlne diphosphate glucose In sugar beet leaf Arch Bzochem. and Bzofihys., 62 16-28.
CARDINI, E. E., LELOIR, L. E. AND CI-IIRIBOGA, J. (1955) The biosynthesis of sucrose. J . Bzol Chem. 214 ' 149-55.

638 AGRICULTURE
CORI. C. F., COLOWICIC, S. P AND CORI, G. T. (1937) The isolation and synthesis of glucose-I- phosphoric acid. J, Biol. Chem., 121: 465-77.
CORI, G. T. AKD CORI, C. F. (1940) The lrinetics of the enzymatic synthesis of glycogen from glu- cose-I-phosphate. J. Bzol. Chem., 135.
DE FEICETE, M. A. R., LELOIR, Id. F. AND CARDINI, C. E. (1960) Mechanism of starch biosynthesis. Nature, 187: 918-19.
DUGGER. W. M. AND HUMPHREYS. T. E. (1960) Influence of boron on enzymatic reactions associa- ~- ,
ted w ~ t h biosynthesis of sucrose. Plnnt Physiol., 35: 523-30. DUTT, N. L. AND NARASIMHAN, R. (1949) The occurrence of starch and its importance a5 a specific
character in Saccharum., Curr. Scz., 18: 346-7. EWART, IT. H., SIMINOVITCH, D. AND BRIGGS, I). R. (1954) Studies on the chemistry of the living
barlr of the blaclr locust in relation to its frost hardiness. VIII. Possible enzymatic processes involved in starch-sucrose interconvenslons. Plant Physzol , 29: 407-13.
FEULHERADE, L. (1929) Starch in Uba juices. S . A . S . J . , 13: 305-9. FISCHER, E. H. AND KREBS, E. G. (1958) The isolation and crystallization of rabbit slreletal
muscle phosphorylase. J . Biol. Chem., 231: 65-71. FRUTON, T. S. AND SIMMONDS, S. (1963) General Bzochemistry. John W11e)r and Sons, Inc., New
YO;^, p, 441. GILBERT, G. A. AND SWALLOW, A. J . (1949) The influence of ions on the activity of Q enzyme.
J . Chem. Soc., 4 : 2849-52. GILBERT. G. A. AND PATRICK, A. D. (1952) Enzvmes of the potato concerned in the synthesis of
\ - . , A
starch. T. The separation and crystallizatin of Q enzyme. Bzochem. J: 181-6. GREEN, A. A. AND CORI, G. T. (1943) Crystalline muscle phosphorylase. I . Pi-epaiation, properties,
and molecular weight. J . Bzol. Chem., 151: 21-30. GREEN, D. E. AND STUMPF, P . I<. (19~2) Starch phosphorylase of potato. 1. B z o ~ . ( ; h e f f ~ . , 124:
355-66. I-IADDON, E. (1927) Starch in sugar cane. Rev. Agrzc. Maurice, 4: 61-2. I-IANES, C. S. (194oa) The brealrdown and synthesis of starch by an enzyme system fro111 pea seeds.
Proc. Roy. Soc. (London), B 128: 421-50. -- (1940b) Reversible formation of starch from glucose-I-phosphate catalyzed by potato
phosphorylase. Proc. Roy. Soc. (London) , B 129 : 174-208. HATCH, M D., SACHER, J . A. AND GLASZIOU, I<. T. (1963) Sugar accumulation cycle In sugar cane.
I. Studies on enzymes of the cycle. Plant Physiol. 3 8 : 338-43. - HOBSON, P. N., WHELAN, W. J. AND PEAT, S. (1951) The enzymat~c synthesis and degradation of
starch. Part XIV. R enzyme. J. Chem. Soc., 2 : 1451-59. I<ELLER, P. J . AND CORI, G. T. (1955) Purification and properties of the phosphorylase-rupturing
enzyme. J . Bzol. Chem., 214: 127-134. LELOIR, L. F., DE FEICETE, M. A. R. AND CARDINI, C. E. (1961) Starch and oligosaccharide syn-
thesis from UDP-glucose. J . Bzol. Chem., 236: 636-41. NELSON, J . M. AND AUCHINCLOSS, R. (1933) The effects of glucose and fructose on the sucrose con-
tent in potato slices. J . Am. Chem. Soc. 55: 3769. NELSON, 0. E. AND RINES, H. W. (1962) The enzymatic deficiency in the waxy mutant of maize.
Bzochem. Bzophys. Res. Commun., 9 : 297-3. PALADINI, A. C. AND LELOIR, L. F. (1952) Studies on uridine diphosphate glucose. Bzochem. J., 59:
426-30. PEAT, S., WIIELAN, W. J., HOBSON, P. N. AND THOMAS, G. J . (1954) The enzymat~c synthesis and
degradat~on of starch. Part XIX. The act1011 of R enzyine of glycogen. J . Chem. Soc., 4 : 4440-5.
POTTINGER, P. K. AND OLIVER, I. T. (1962) The intracellular distribution of UDP-glucose-starch synthetase in potato tubers. Bzochzm. Bzophys. Acta, 58: 303-6.
SUTHERLAND, E. W., CORI, C. F., HAYNES, R. AND OLSEN, N. S. (1959) Purification of the hyper- glyceinic-glycogenolytic factor from insnlin and froin gastric mucosa. J . Bzol. Chem., 180: 825-17.
TURNER, j.-~. (1957) The biosynthesis of sucrose. Biochenz. J., 67: 450-6 VAN DILLEWIJN, C. (1952) Botany of Sugarcane. The Chronica Botanica Co., Waltham, Mass., p.
315. vON STIEGLITZ, C. R. (1932) The starch contents of some sugar canes m Queensland. Proc. Queensl.
Soc. Sug. Cane Technol., 3 : 63-8. WELLER, D. M. (1929) Accu~nulations of starch in the stallrs of Natal Uba cane grown on acid
soil. Haw. Plant. Rec., 34: 294-9. WHELAN, W. J. (1961) Recent advallces in the biochemistry of glycogen and starch. Nature. 190:
954-7. WOOD, G. H. (1962) Some factors influencing starch in sugar cane. Proc. S . African Sug. Technol.
Ass . : 123.

1 A. d. ALEXANDER 639 ! I Discussion
J. L. DU TOIT (S. Africa) : We have found in South Africa that there are big varietal differences in starch content. The Officinarum and Robustum varieties are all very low in starch, but the Barberi and Synensis varieties are very high with the Spontaneum varieties variable. The variety Uba has been mentioned as high in starch which is probably correct. However, Uba Marot, as mentioned in your paper, is a Spontaneum seedling and in no way related to UBA. MARETSKI (Puerto Rico) : Are the amylase levels in Uba Marot comparable to those in M. 336 ? A. ALEXANDER (Puerto Riqo) : The Uba Marot has a higher amylase content. G. E. BEAUCHAMP (Puerto Rico) : I thinlr we should congratulate Dr. Alexander for a very imnpor- tant piece of research well done. There appears to be an indication so far that his worlc applied to young seedlings may help to discover high sucrose yielding varieties and thus save lots of time and money in breeding work. A. J. VLITOS (Trinidad) : Is it not possible that some varieties might contain "lazy-amylase" and thereby store more starch than others ? Varietal differences in enzyme content may be important. For example, sucrose phosphorylase does not appear to occur in Pindar but does occur in Puerto Rico, (R. R.) varieties. DR. ALEXANDER: I do not know about "lazy-enzymes", but it does appear that where something goes wrong with the amylase system starch accumulates.





























