Embed Size (px)
Citation preview

of May 17, 2018.This information is current as
Transfer Model of ColitisIL-4 Exacerbates Disease in a Th1 Cell
M. RennickMenon, Frank Brombacher, Michael W. Leach and Donna Madeline M. Fort, Robin Lesley, Natalie J. Davidson, Satish
http://www.jimmunol.org/content/166/4/2793doi: 10.4049/jimmunol.166.4.2793
2001; 166:2793-2800; ;J Immunol
Referenceshttp://www.jimmunol.org/content/166/4/2793.full#ref-list-1
, 27 of which you can access for free at: cites 53 articlesThis article
average*
4 weeks from acceptance to publicationFast Publication! •
Every submission reviewed by practicing scientistsNo Triage! •
from submission to initial decisionRapid Reviews! 30 days* •
Submit online. ?The JIWhy
Subscriptionhttp://jimmunol.org/subscription
is online at: The Journal of ImmunologyInformation about subscribing to
Permissionshttp://www.aai.org/About/Publications/JI/copyright.htmlSubmit copyright permission requests at:
Email Alertshttp://jimmunol.org/alertsReceive free email-alerts when new articles cite this article. Sign up at:
Print ISSN: 0022-1767 Online ISSN: 1550-6606. Immunologists All rights reserved.Copyright © 2001 by The American Association of1451 Rockville Pike, Suite 650, Rockville, MD 20852The American Association of Immunologists, Inc.,
is published twice each month byThe Journal of Immunology
by guest on May 17, 2018
http://ww
w.jim
munol.org/
Dow
nloaded from
by guest on May 17, 2018
http://ww
w.jim
munol.org/
Dow
nloaded from

IL-4 Exacerbates Disease in a Th1 Cell TransferModel of Colitis1
Madeline M. Fort,2* Robin Lesley,* Natalie J. Davidson,* Satish Menon,* Frank Brombacher,†
Michael W. Leach,‡ and Donna M. Rennick*
IL-4 is associated with Th2-type immune responses and can either inhibit or, in some cases, promote Th1-type responses. We testedthe effect of IL-4 treatment on the development of inflammation in the CD41CD45RBhigh T cell transfer model of colitis, whichhas been characterized as a Th1-dependent disease. IL-4 treatment significantly accelerated the development of colitis in immu-nodeficient recipients (recombinase-activating gene-2 (Rag2)2/2) of CD41CD45RBhigh T cells. Quantitative analysis of mRNAexpression in the colons of IL-4-treated mice showed an up-regulation of both Th1- and Th2-associated molecules, includingIFN-g, IP-10, MIG, CXCR3, chemokine receptor-8, and IL-4. However, cotreatment with either IL-10 or anti-IL-12 mAb effec-tively blocked the development of colitis in the presence of exogenous IL-4. These data indicate that IL-4 treatment exacerbatesa Th1-mediated disease rather than induces Th2-mediated inflammation. As other cell types besides T cells express the receptorfor IL-4, the proinflammatory effects of IL-4 on host cells in Rag22/2 recipients were assessed. IL-4 treatment was able tomoderately exacerbate colitis in Rag22/2 mice that were reconstituted with IL-4Ra-deficient (IL-4Ra2/2) CD41CD45RBhigh Tcells, suggesting that the IL-4 has proinflammatory effects on both non-T and T cells in this model. IL-4 did not cause colitis inRag22/2 mice in the absence of T cells, but did induce an increase in MHC class II expression in the lamina propria of the colon,which was blocked by cotreatment with IL-10. Together these results indicate that IL-4 can indirectly promote Th1-type inflam-mation in the CD41CD45RBhigh T cell transfer model of colitis. The Journal of Immunology,2001, 166: 2793–2800.
T cell-dependent immune responses are generally dividedinto two types, Th1 and Th2. Th1-type responses occurwhen APCs release IL-12, which, in turn, induces the
differentiation of CD41 T cells to produce IL-2 and IFN-g (1, 2).When uncontrolled, Th1 responses can result in chronic inflam-matory diseases, such as diabetes, arthritis, and multiple sclerosis.In animal models, inhibition of IL-12 will prevent the developmentof these autoimmune diseases by blocking the development ofIFN-g-producing CD41 T cells (3, 4). Thus, the ability to controlthe development of Th1-type T cells may be critical for the pre-vention of some chronic inflammatory diseases.
Th2-type responses are characterized by the development ofCD41 T cells, which secrete IL-4, IL-5, IL-10, and IL-13 (1, 2).Some Th2-type cytokines, especially IL-10 and IL-4, are known tosuppress the development of Th1 T cells (3, 5, 6). IL-10 has beenshown to suppress many Th1-type inflammatory responses in vivo,including LPS-induced endotoxic shock, contact hypersensitivity,experimental autoimmune encephalomyelitis, and collagen-in-duced arthritis (7–11). Like IL-10, IL-4 treatment can suppress orameliorate the development of Th1-type inflammatory diseases.IL-4 treatment has been shown to suppress the development ofproteoglycan-induced arthritis and to prevent cartilage destructionin a collagen-induced model of arthritis (12, 13). In addition, IL-4
inhibits the development of diabetes in both NOD and transgenicmouse models as well as the development of experimental auto-immune encephalomyelitis (5, 14–18). Furthermore, the combina-tion of IL-4 and IL-10 is more effective than either cytokine alonein ameliorating glomerulonephritis and delayed-type hypersensi-tivity in mice primed withLeishmania major(19, 20). These datasuggest that Th2-associated cytokines are potent inhibitors of Th1-type immune responses.
The ability of Th2-associated cytokines to inhibit the develop-ment of deleterious Th1-mediated immune responses suggestedthat they may be useful therapeutics for ablating certain autoim-mune diseases (5, 6). However, there is increasing evidence thatIL-4, in particular, can aid in Th1-type inflammatory responses.IL-4 treatment of rats significantly worsened the development ofTh1-type experimental autoimmune uveoretinitis and resulted inincreased production of IFN-g, TNF-a, and NO in recall responsesin vitro to retinal Ag (21). Furthermore, IL-4-deficient (IL-42/2)mice have severely impaired anti-tumor responses that are char-acterized by an inability to develop CD81 anti-tumor CTLs and bydecreased production of splenic IFN-g (22). IL-42/2 mice are alsounable to produce enough IFN-g and IL-12 to develop immunity toCandida albicansinfection (23). Recently, IL-4 has been shown tobe critical for the development of alloreactive CD41 T cells, andblocking IL-4 prolongs the survival of allogeneic skin grafts (24).These data indicate that IL-4 may be important for the develop-ment of some Th1-type inflammatory responses.
Crohn’s disease is a chronic inflammatory condition of the in-testinal tract that has been characterized as a Th1-mediated dis-ease. T cells isolated from the colons of patients with Crohn’sdisease produce large amounts of IFN-g and TNF-a and little IL-4or IL-10 (25–27). Many mouse models of chronic intestinal in-flammation, or colitis, are also characterized by massive infiltra-tion of IFN-g-producing CD41 T cells in the colon. IL-102/2
mice, IL-22/2 mice, Gai22/2 mice, Stat32/2 mice, Tge26 mice
*Department of Molecular Biology, DNAX Research Institute for Molecular andCellular Biology, Palo Alto, CA 94304;†Department of Immunology, University ofCape Town, Cape Town, South Africa; and‡Schering-Plough Research Institute,Lafayette, NJ 07848
Received for publication July 14, 2000. Accepted for publication December 8, 2000.
The costs of publication of this article were defrayed in part by the payment of pagecharges. This article must therefore be hereby markedadvertisementin accordancewith 18 U.S.C. Section 1734 solely to indicate this fact.1 DNAX Research Institute is supported by Schering-Plough Corp.2 Address correspondence and reprint requests to Dr. Madeline M. Fort, DNAX Re-search Institute, 901 California Avenue, Palo Alto, CA 94304. E-mail address:[email protected]
Copyright © 2001 by The American Association of Immunologists 0022-1767/01/$02.00
by guest on May 17, 2018
http://ww
w.jim
munol.org/
Dow
nloaded from

reconstituted with wild-type (WT)3 bone marrow cells, immuno-deficient recipients of CD41CD45RBhigh T cells, and trinitroben-zenesulfonic acid (TNBS)-treated SJL/J mice all develop Th1-typecolitis (28–32). In many of these models disease is either pre-vented, or at least ameliorated, by early treatment with IL-10 orneutralizing Ab to IL-12 (26, 28, 30). The role of IL-4 in Th1-typeintestinal disease is less clear. In one study IL-4 treatment did notsuppress the development of colitis inscidmice transplanted withCD41CD45RBhigh T cells (33). In contrast, infection of mice withadenovirus containing the IL-4 gene reduced inflammation in ratswith TNBS-induced colitis (34).
We have evaluated the effect of IL-4 treatment in a modifiedversion of the original T cell transfer model of colitis (29, 35). Inthis modified model the transfer of CD41CD45RBhigh T cells fromeither WT mice or IL-102/2 mice with established Th1-type dis-ease into 129 SvEv Rag22/2 mice results in colitis characterizedby diffuse inflammatory cellular infiltrates, epithelial hyperplasia,and, when severe, ulcers and transmural inflammation. The result-ing colitis can be significantly ameliorated by treatment with anti-IL-12 mAb or rIL-10 in vivo (36, 37). We transferredCD41CD45RBhigh T cells from either WT or IL-102/2 mice intoRag22/2 immunodeficient mice and treated them daily with IL-4.Our data show that IL-4 treatment has the ability to exacerbateTh1-type colonic inflammation in this model.
Materials and MethodsMice
Recombinase-activating gene 2-deficient (Rag22/2) mice on either 129SvEv or BALB/c background and WT 129 SvEv mice were obtained fromTaconic Farms (Germantown, NY) or from a colony maintained at theDNAX Animal Care Facility. IL-102/2 129 SvEv mice and IL-4Ra-chain-deficient (IL-4Ra2/2) BALB/c mice were from colonies maintained underspecific pathogen-free conditions at the DNAX Animal Care Facility (38, 39).
In vivo treatments
Purified recombinant murine IL-4 and IL-10 were made as previously de-scribed and contained,0.1 E.U/mg protein of endotoxin (37, 40). Micewere given 10–20mg of IL-4 i.p. daily for 28 days. For IL-10 treatment,mice were given 20mg i.p. daily. Saline (13HBSS; BioWhittaker, Walk-ersville, MD) was given daily as a negative control. Purified anti-murineIL-12 mAb (C.17.8.20) or isotype control mAb (MP4–25D2, anti-humanIL-4; no cross-reactivity with murine IL-4) was given to mice i.p. weekly,at 2 mg/dose for the duration of the experiment. All cytokine and Abtreatments were started on the day of T cell transfer (day 0).
Cell isolations and transfers
CD41CD45RBhigh splenic T cells were obtained by cell sorting. Briefly,splenocytes were first enriched for CD41 T cells by red cell lysis andmagnetic bead depletion using lineage-specific rat mAbs supernatants(10%, v/v): B220 (B cells) and Ter119 (erythrocytes). mAb-stained cellswere removed in a magnetic field using goat anti-rat IgG (Fc) and goatanti-rat IgG (H1L)-coated magnetic beads (PerSeptive Diagnostics, Cam-bridge, MA). The remaining cells were then stained with anti-CD4-FITCand anti-CD45RB-PE (both from PharMingen, San Diego, CA). Two-colorcell sorting was performed using a FACStar Plus (Becton Dickinson,Mountain View, CA). The sorted CD41CD45RBhigh T cells were.98%pure upon reanalysis. The purified CD41CD45RBhigh cells (23 105) wereinjected i.p. into 129 SvEv or BALB/c Rag22/2 recipient mice, dependingon the donor cells used. Four weeks after T cell transfer, mice were sac-rificed and analyzed for the presence of enterocolitis.
Histologic analysis of colitis
Microscopic examination of mouse colons was performed in a blindedfashion by the same pathologist (M.W.L.) on formalin-fixed tissue sectionsstained with hematoxylin and eosin as previously described (36). Longi-
tudinal sections of the entire length of the colon were evaluated, taking intoaccount both the number of lesions and their severity. Five regions of thecolon (cecum, ascending, transverse and descending colon, and rectum)were graded semiquantitatively as 0 (no change) to 5 (most severe change).The grading represents the incidence and severity of inflammatory lesionsbased on infiltrates, goblet cell loss, crypt abscesses, ulcerations, and fi-brosis. The summation of the score for each of five segments of the colonprovides a total disease score per mouse (from 0–25), where 0–1 indicatesno change, 2–5 indicates mild disease, 6–10 indicates moderate disease,and 11–20 indicates severe disease. No mice in these studies have a score.20, because such severe disease results in death.
Quantitative mRNA (TaqMan) analysis in colon
Total RNA was isolated from whole colon samples using Qiagen RNeasycolumns (Qiagen, Valencia, CA), according to the manufacturer’s instruc-tions. Total RNA (5mg) was reverse transcribed into cDNA using randomhexamers (Promega, Madison, WI). The expression of IFN-g, IL-4, MIG,IP-10, eotaxin, TARC, CCR3, CCR4, CCR8, CXCR3, and TNF-a wasdetermined by a method for real-time quantitative PCR using the ABI 7700sequence detector system (Perkin-Elmer Applied Biosystems, Foster City,CA). Briefly, 50 ng of total cDNA was in a reaction volume of 25ml thatcontained final concentrations of 13PCR buffer; 200mM dATP, dCTP,and dGTP; 400mM dUTP; 4 mM MgCl2; 1.25 U of AmpliTaq DNApolymerase; 0.5 U of Amp-Erase uracil-N-glycosylase; 900 nM of eachprimer; and 250 nM probe. The thermal cycling conditions included 50°Cfor 2 min and 95°C for 10 min, followed by 40 cycles of amplification at95°C for 15 s and 55°C for 1 min for denaturing and annealing-extension,respectively. Sense and antisense primers as well as probes used for thedetection of the genes of interest were predeveloped TaqMan assay re-agents (Perkin-Elmer Applied Biosystems). Primers and probes were de-signed to ensure that no cross-reactivity with other genes would occur.Each primer-probe pair was tested on a panel of cDNA plasmids containinga variety of cytokines, chemokines, and chemokine receptors. The probesfor each message were labeled at the 59end with a reporter fluorescent dye,FAM, and at the 39end with a quench fluorescent dye, TAMRA. Fluores-cence detection of FAM was performed at the end of each cycle. Thequantity of cDNA of the gene of interest was directly related to the amountof FAM detected after 40 cycles. cDNA plasmids containing the gene ofinterest were used as a standard curve, ranging from 100 to 0.01 pg. Fromthis standard curve, the amount of cDNA of the gene of interest was cal-culated in femtograms per 50 ng of total cDNA. As an internal control, 18Sribosomal RNA (rRNA) expression was measured in each sample in amultiplex assay; the probe for rRNA was labeled at the 59 end with thereporter fluorescent dye VIC. The amount of 18S rRNA was correlated tothe cycle at which VIC fluorescence was first detected (cycle thresholdvalue). To correct for any variation in the amount of RNA between indi-vidual samples, the mean cycle threshold value for 18S rRNA was calcu-lated for all samples and subtracted from each individual cycle thresholdvalue, then this difference was raised to the second power and multiplied bythe FAM value (in femtograms per 50 ng of total cDNA) for each sample.Thus, the amount of cDNA of the gene of interest in each sample could bedirectly compared with the amounts detected in all other samples.
Immunohistochemical analysis
Briefly, colon pieces from saline- and IL-4-treated Rag22/2 mice werecleaned of feces and frozen in tissue-freezing medium (Triangle Biomed-ical Sciences, Durham, NC). Frozen blocks were cut on a cryostat, and8-mm-thick sections were cut onto gelatinized glass slides, dried, and fixedfor 10 min with acetone. Sections were preincubated with 13PBS (Gen-eMate, Kaysville, UT) and 10% normal mouse serum (NMS; Jackson Im-munoResearch Laboratories, West Grove, PA) for 15 min. Rat anti-mouseI-Ab (PharMingen) was diluted in PBS/10% NMS and incubated on sec-tions for 1 h at room temperature. Rat IgG2b (PharMingen) was used as anisotype control. Sections were rinsed in 13PBS and stained with HRP-conjugated sheep anti-rat IgG (The Binding Site, Birmingham, U.K.) di-luted in PBS/10% NMS for 30 min at room temperature. AEC (Vector,Burlingame, CA) was used as a substrate for HRP. Sections were coun-terstained with hematoxylin (Vector), and then sequentially rinsed in H20,1% ammonium hydroxide, and PBS. Sections were mounted with cover-slips and observed by light microscopy.
Statistics
All data were analyzed using a statistical program (InstatP, GraphPad, SanDiego, CA). Student’st test or nonparametric Mann-Whitney test was usedto determine statistical significance between groups, withp # 0.05 con-sidered significant.
3 Abbreviations used in this paper: WT, wild type; IL-102/2, IL-10-deficient, IL-4Ra2/2, IL-4R a-chain-deficient; Rag22/2, recombinase-activating gene-2-deficient;TNBS, trinitrobenzenesulfonic acid; IP-10; rRNA, ribosomal RNA; NMS, normalmouse serum; LP, lamina propria.
2794 IL-4 TREATMENT PROMOTES Th1-TYPE COLITIS
by guest on May 17, 2018
http://ww
w.jim
munol.org/
Dow
nloaded from

ResultsIL-4 exacerbates disease in the CD41CD45RBhigh T celltransfer model of colitis
To test the effect of IL-4 on a Th1-type murine model of colitis,WT CD41CD45RBhigh T cells were transferred into immunode-ficient Rag22/2 129 SvEv mice, and the recipient mice weretreated daily with either saline (vehicle control) or 20mg IL-4. Inthis model untreated recipient mice develop moderate to severecolitis 8–12 wk after T cell transfer (29, 36, 37). However, IL-4treatment exacerbated the disease such that the majority of micewere moribund 4 wk after T cell transfer. Therefore, the experi-ment was terminated, and the mice were analyzed histologically
for signs of colitis. Five of eight mice had moderate to severecolitis by 4 wk, while saline-treated mice had few histologicalsigns of disease at that time point (Fig. 1A). As WTCD41CD45RBhigh T cells are uncommitted to either a Th1 or aTh2 phenotype, we considered the possibility that IL-4 treatmentwas pushing naive T cells to an inflammatory Th2 phenotype,rather than having a proinflammatory effect on Th1-type T cells.Therefore, we repeated the experiment using CD41CD45RBhigh Tcells from 4- to 6-mo-old IL-102/2 129 SvEv mice that alreadyhad signs of chronic Th1-type colitis (29, 40). The IL-102/2
CD41CD45RBhigh T cells caused mild colitis in recipientRag22/2 mice within 4 wk (Fig. 1B; mean colitis score, 3.3; Fig.2A). However, IL-4 treatment was able to clearly exacerbate thecolitis seen in Rag22/2 recipients of IL-102/2 CD41CD45RBhigh
T cells (Figs. 1Band 2B; mean colitis score, 10.9). Because, in ourhands, IL-102/2 CD41CD45RBhigh T cells cause more consistentand aggressive disease in Rag22/2 recipients than T cells fromWT mice, we performed all subsequent transfer experiments withCD41CD45RBhigh T cells from IL-102/2 mice. (M. Fort and D.Rennick, unpublished observations).
IL-4 treatment does not switch the colitis to a Th2-typeinflammation
The ability of exogenous IL-4 to exacerbate colitis in this T celltransfer model of colitis lead us to investigate whether IL-4 in-duced a switch from a Th1-dependent to a Th2-dependent disease.Therefore, we looked for differential gene expression in the colonsof Rag22/2 mice 28 days after reconstitution with IL-102/2
CD41CD45RBhigh T cells and treatment with either saline or IL-4.Using a very sensitive, quantitative method of PCR analysis, co-lonic total RNA was tested for the presence of mRNA of variousTh1- and Th2-type-associated cytokines, chemokines, and chemo-kine receptors. IL-4 treatment induced a significant increase in the
FIGURE 1. In vivo treatment with IL-4 accelerates the colitis caused byCD41CD45RBhigh T cells. Purified 23 105 CD41CD45RBhigh T cellsfrom either WT (A) or IL-102/2 (B) mice were transferred into Rag22/2
mice. Mice were treated daily i.p. with saline or 20mg of IL-4 for 28 days.At the end of 28 days mice were sacrificed, and the colons of individualmice were scored for colitis by histological analysis (seeMaterials andMethods). Each symbol represents an individual animal. Bar indicatesmean colitis score for each group.A andB, IL-4-treated mice had signif-icantly higher colitis scores compared with saline-treated mice (p5 0.002andp 5 0.016, respectively, by Mann-WhitneyU test).
FIGURE 2. IL-4 exacerbates colitis in T cell-reconsti-tuted Rag22/2 mice. Representative photomicrographs ofthe descending colon of Rag22/2 mice. When present,colitis was usually diffuse.A, Example of moderate colitisin a mouse receiving IL-102/2 CD41CD45RBhigh T cellsand treated with saline. This was the most severe diseaseseen in this group at 4 wk post-transfer.B, Example ofsevere disease typically seen in mice receiving IL-102/2
CD41CD45RBhigh T cells and treated with IL-4. There isgreater epithelial hyperplasia and more inflammation thatseparates intestinal glands and extends into the submu-cosa than shown inA. A multinucleated giant cells is alsopresent (left sideat the base of the glands).C and D,Normal appearing colon from mice receiving IL-102/2
CD41CD45RBhigh T cells and treated with IL-4 andIL-10 (C) or with IL-4 and anti-IL-12 mAb (D).E andF,Normal-appearing colon from nonreconstituted Rag22/2
mice treated with saline (E) or IL-4 (F). Hematoxylin andeosin,3100.
2795The Journal of Immunology
by guest on May 17, 2018
http://ww
w.jim
munol.org/
Dow
nloaded from
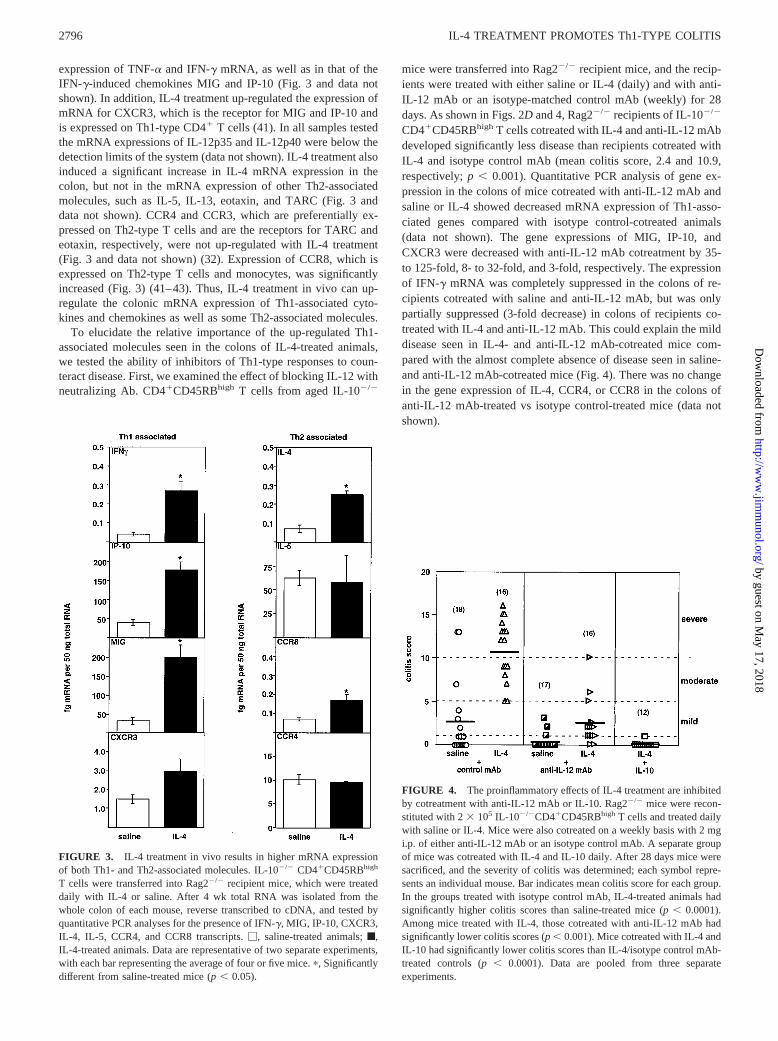
expression of TNF-a and IFN-g mRNA, as well as in that of theIFN-g-induced chemokines MIG and IP-10 (Fig. 3 and data notshown). In addition, IL-4 treatment up-regulated the expression ofmRNA for CXCR3, which is the receptor for MIG and IP-10 andis expressed on Th1-type CD41 T cells (41). In all samples testedthe mRNA expressions of IL-12p35 and IL-12p40 were below thedetection limits of the system (data not shown). IL-4 treatment alsoinduced a significant increase in IL-4 mRNA expression in thecolon, but not in the mRNA expression of other Th2-associatedmolecules, such as IL-5, IL-13, eotaxin, and TARC (Fig. 3 anddata not shown). CCR4 and CCR3, which are preferentially ex-pressed on Th2-type T cells and are the receptors for TARC andeotaxin, respectively, were not up-regulated with IL-4 treatment(Fig. 3 and data not shown) (32). Expression of CCR8, which isexpressed on Th2-type T cells and monocytes, was significantlyincreased (Fig. 3) (41–43). Thus, IL-4 treatment in vivo can up-regulate the colonic mRNA expression of Th1-associated cyto-kines and chemokines as well as some Th2-associated molecules.
To elucidate the relative importance of the up-regulated Th1-associated molecules seen in the colons of IL-4-treated animals,we tested the ability of inhibitors of Th1-type responses to coun-teract disease. First, we examined the effect of blocking IL-12 withneutralizing Ab. CD41CD45RBhigh T cells from aged IL-102/2
mice were transferred into Rag22/2 recipient mice, and the recip-ients were treated with either saline or IL-4 (daily) and with anti-IL-12 mAb or an isotype-matched control mAb (weekly) for 28days. As shown in Figs. 2Dand 4, Rag22/2 recipients of IL-102/2
CD41CD45RBhigh T cells cotreated with IL-4 and anti-IL-12 mAbdeveloped significantly less disease than recipients cotreated withIL-4 and isotype control mAb (mean colitis score, 2.4 and 10.9,respectively;p , 0.001). Quantitative PCR analysis of gene ex-pression in the colons of mice cotreated with anti-IL-12 mAb andsaline or IL-4 showed decreased mRNA expression of Th1-asso-ciated genes compared with isotype control-cotreated animals(data not shown). The gene expressions of MIG, IP-10, andCXCR3 were decreased with anti-IL-12 mAb cotreatment by 35-to 125-fold, 8- to 32-fold, and 3-fold, respectively. The expressionof IFN-g mRNA was completely suppressed in the colons of re-cipients cotreated with saline and anti-IL-12 mAb, but was onlypartially suppressed (3-fold decrease) in colons of recipients co-treated with IL-4 and anti-IL-12 mAb. This could explain the milddisease seen in IL-4- and anti-IL-12 mAb-cotreated mice com-pared with the almost complete absence of disease seen in saline-and anti-IL-12 mAb-cotreated mice (Fig. 4). There was no changein the gene expression of IL-4, CCR4, or CCR8 in the colons ofanti-IL-12 mAb-treated vs isotype control-treated mice (data notshown).
FIGURE 3. IL-4 treatment in vivo results in higher mRNA expressionof both Th1- and Th2-associated molecules. IL-102/2 CD41CD45RBhigh
T cells were transferred into Rag22/2 recipient mice, which were treateddaily with IL-4 or saline. After 4 wk total RNA was isolated from thewhole colon of each mouse, reverse transcribed to cDNA, and tested byquantitative PCR analyses for the presence of IFN-g, MIG, IP-10, CXCR3,IL-4, IL-5, CCR4, and CCR8 transcripts.M, saline-treated animals;f,IL-4-treated animals. Data are representative of two separate experiments,with each bar representing the average of four or five mice.p, Significantlydifferent from saline-treated mice (p, 0.05).
FIGURE 4. The proinflammatory effects of IL-4 treatment are inhibitedby cotreatment with anti-IL-12 mAb or IL-10. Rag22/2 mice were recon-stituted with 23 105 IL-102/2CD41CD45RBhigh T cells and treated dailywith saline or IL-4. Mice were also cotreated on a weekly basis with 2 mgi.p. of either anti-IL-12 mAb or an isotype control mAb. A separate groupof mice was cotreated with IL-4 and IL-10 daily. After 28 days mice weresacrificed, and the severity of colitis was determined; each symbol repre-sents an individual mouse. Bar indicates mean colitis score for each group.In the groups treated with isotype control mAb, IL-4-treated animals hadsignificantly higher colitis scores than saline-treated mice (p, 0.0001).Among mice treated with IL-4, those cotreated with anti-IL-12 mAb hadsignificantly lower colitis scores (p, 0.001). Mice cotreated with IL-4 andIL-10 had significantly lower colitis scores than IL-4/isotype control mAb-treated controls (p, 0.0001). Data are pooled from three separateexperiments.
2796 IL-4 TREATMENT PROMOTES Th1-TYPE COLITIS
by guest on May 17, 2018
http://ww
w.jim
munol.org/
Dow
nloaded from

To confirm that IL-4 was acting by exacerbating a Th1 response,the ability of IL-10 to ameliorate the effects of IL-4 was also tested.Daily IL-10 cotreatment completely suppressed the developmentof colitis in the presence of IL-4 (Figs 2Cand 4). These datasuggest that the colitis seen in IL-4-treated recipients was depen-dent on the production of IL-12 and was inhibited by the presenceof exogenous IL-10, and thus fits the paradigm of a Th1-type in-flammatory disease.
IL-4 need not act directly on T cells to augment a Th1-typeinflammatory response
The receptor for IL-4 consists of the IL-4Ra-chain and IL-2Rg-chain and is expressed on many cell types, including T cells, Bcells, monocytes, and nonhemopoietic cells, including intestinalepithelial cells (44–47). Therefore, IL-4 treatment in vivo may ac-celerate colitis by acting on T cells, microenvironmental cells, or both.To differentiate the effects of exogenous IL-4 on the Rag22/2 hostcells vs donor T cells, we made use of T cells from mice that aredeficient in the IL-4Ra-chain (IL-4Ra2/2). CD41CD45RBhigh Tcells from IL-4Ra2/2 BALB/c mice were transferred into Rag22/2
BALB/c mice, and the mice were treated for 28 days with eitherIL-4 or saline. As shown in Fig. 5, saline-treated recipients ofIL-4Ra2/2 T cells developed mild-moderate colitis (mean score,4.8) within 4 wk. Of the IL-4-treated recipients, 9 of 13 mice hadmoderate to severe disease, even though the transferred T cellscould not respond to IL-4 (Fig. 5). However, because of the largevariability in disease score within each group, the results were notstatistically significant. Nevertheless, the trend toward exacerbateddisease following IL-4 treatment suggests a proinflammatory effecton the microenvironmental cells that express functional IL-4R.
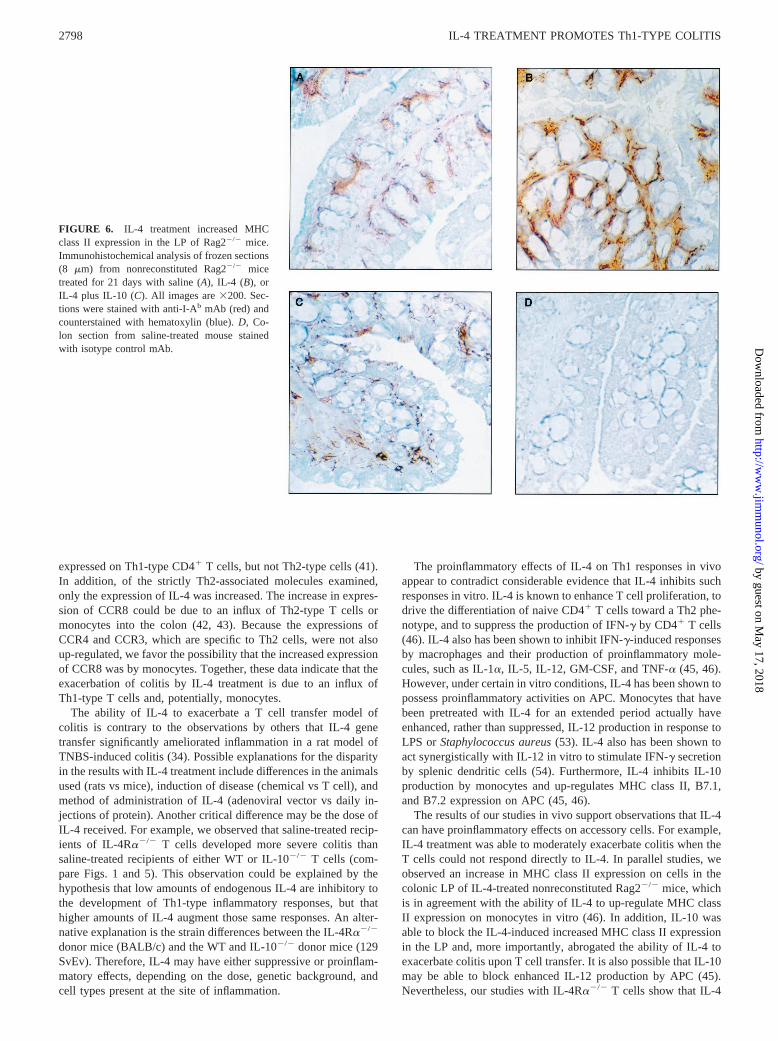
To test whether IL-4 treatment could induce an inflammatoryresponse in the absence of T cells, nonreconstituted Rag22/2 micewere treated daily with IL-4 or saline control for 21–28 days. Asshown in Table I, 9 of 10 nonreconstituted Rag22/2 mice treatedwith IL-4 remained disease free, and only one mouse had verymild colitis. Histological analysis failed to show obvious increasesin mononuclear or polynuclear infiltrates in the colons of IL-4-treated Rag22/2 mice (Fig. 2,E and F). To determine whetherthere were changes in macrophage or dendritic cell populations,immunohistochemical analyses of frozen sections from colon tis-sue were performed. No difference in the expression of F4/80,CD11b, CD11c, or CD4 was found in colons from IL-4-treatednonreconstituted Rag22/2 mice compared with saline-treated con-trols (data not shown). However, there was a consistent increase in
MHC class II expression in the lamina propria (LP) of IL-4-treatednonreconstituted Rag22/2 mice (Fig. 6). Furthermore, cotreatmentof Rag22/2 with IL-10 suppressed this increase in MHC class IIexpression in the LP by IL-4 (Fig. 6). Thus, IL-4 treatment maypromote colitis in the presence of T cells by increasing Ag pre-sentation by resident macrophages and/or dendritic cells. Treat-ment of nonreconstituted Rag22/2 mice with IL-4 did cause a3-fold increase in spleen size (Table I) due to extramedullary he-mopoiesis, as has been previously described by others (48). Thesplenomegaly was not significantly altered by cotreatment withIL-10. As IL-10 effectively blocked the exacerbation of colitis andthe increased MHC class II expression in the LP, the developmentof extramedullary hemopoiesis and splenomegaly appeared to be adistinct effect of IL-4 treatment. These data show that IL-4 canhave effects on hemopoietic cells in the absence of T or B cells, butthe development of colitis is strictly T cell dependent in this model.
DiscussionOur results clearly show that IL-4, which is typically associatedwith Th2-type responses, exacerbated an intestinal Th1-type in-flammatory disease. The inability of IL-4 to inhibit a T cell transfermodel of colitis has been previously described, but our data extendthese results to show that IL-4 treatment significantly acceleratesthe onset and the severity of the colitis seen in Rag22/2 recipientsof either WT or IL-102/2 CD41CD45RBhigh T cells (37). Co-treatment with either IL-10 or anti-IL-12 mAb greatly diminishedthe effects of IL-4 on the development of colitis. Both anti-IL-12mAb and IL-10 treatments have been shown to prevent colitis inseveral models of Th1-type colitis, including IL-102/2 mice andthe transfer of WT or IL-102/2 CD41CD45RBhigh T cells to im-munodeficient mice (29, 36, 37, 49). Our observation that IL-4 canaccelerate an IL-12-dependent model of colitis is distinct fromTh2-type models of colitis, which are strictly dependent on thepresence of IL-4. For example, BALB/c mice treated intrarectallywith TNBS and SJL/J mice treated intrarectally with oxazolonewill develop intestinal inflammation that is ameliorated by treat-ment with anti-IL-4 mAb, but not by anti-IL-12 mAb (50, 51).
The conclusion that IL-4 treatment promoted a Th1-type inflam-matory response is further supported by the greatly enhanced geneexpression of IFN-g, MIG, IP-10, and CXCR3 in the colons ofIL-4-treated mice. Furthermore, anti-IL-12 mAb cotreatment re-duced the expression of Th1-associated genes as well as the se-verity of disease. MIG and IP-10 are both produced by monocytesand lymphocytes in response to IFN-g. MIG has chemotactic ac-tivity for activated T cells, while IP-10 is a chemoattractant foractivated CD41 T cells, monocytes, and NK cells (52). CXCR3 isthe receptor for both MIG and IP-10 and has been shown to be
FIGURE 5. IL-4 enhances colitis caused by IL-4Ra-deficient T cells.CD41CD45RBhigh T cells from IL-4Ra2/2 mice were transferred intoRag22/2 recipients, which were treated daily with IL-4 or saline for 4 wk.The severity of colitis was determined as described in Fig. 1. Each symbolrepresents an individual animal; data were pooled from two separate ex-periments. Bar indicates mean colitis scores for saline- and IL-4-treatedmice (4.8 and 7.3, respectively).
Table I. IL-4 cannot induce colitis in the absence of T cells but cancause splenomegalya
TreatmentIncidenceof Colitis
Average SpleenWeight (g)
Saline 0/9 0.0656 0.022b
IL-4 1/10c 0.188p6 0.024IL-4 1 IL-10 0/8 0.148p6 0.016
a Nonreconstituted 129 SvEV Rag22/2 mice were treated daily with saline or 20mg IL-4 6 20 mg IL-10 for 21–28 days. Mice were then sacrificed and analyzed forpresence of splenomegaly (by spleen weight) and colitis (by histological analysis).
b Mean6 SEM.c One mouse with colitis score5 2.p, Significantly different from saline-treated mice,p , 0.05, Mann-Whitney non-
parametric test.
2797The Journal of Immunology
by guest on May 17, 2018
http://ww
w.jim
munol.org/
Dow
nloaded from
expressed on Th1-type CD41 T cells, but not Th2-type cells (41).In addition, of the strictly Th2-associated molecules examined,only the expression of IL-4 was increased. The increase in expres-sion of CCR8 could be due to an influx of Th2-type T cells ormonocytes into the colon (42, 43). Because the expressions ofCCR4 and CCR3, which are specific to Th2 cells, were not alsoup-regulated, we favor the possibility that the increased expressionof CCR8 was by monocytes. Together, these data indicate that theexacerbation of colitis by IL-4 treatment is due to an influx ofTh1-type T cells and, potentially, monocytes.
The ability of IL-4 to exacerbate a T cell transfer model ofcolitis is contrary to the observations by others that IL-4 genetransfer significantly ameliorated inflammation in a rat model ofTNBS-induced colitis (34). Possible explanations for the disparityin the results with IL-4 treatment include differences in the animalsused (rats vs mice), induction of disease (chemical vs T cell), andmethod of administration of IL-4 (adenoviral vector vs daily in-jections of protein). Another critical difference may be the dose ofIL-4 received. For example, we observed that saline-treated recip-ients of IL-4Ra2/2 T cells developed more severe colitis thansaline-treated recipients of either WT or IL-102/2 T cells (com-pare Figs. 1 and 5). This observation could be explained by thehypothesis that low amounts of endogenous IL-4 are inhibitory tothe development of Th1-type inflammatory responses, but thathigher amounts of IL-4 augment those same responses. An alter-native explanation is the strain differences between the IL-4Ra2/2
donor mice (BALB/c) and the WT and IL-102/2 donor mice (129SvEv). Therefore, IL-4 may have either suppressive or proinflam-matory effects, depending on the dose, genetic background, andcell types present at the site of inflammation.
The proinflammatory effects of IL-4 on Th1 responses in vivoappear to contradict considerable evidence that IL-4 inhibits suchresponses in vitro. IL-4 is known to enhance T cell proliferation, todrive the differentiation of naive CD41 T cells toward a Th2 phe-notype, and to suppress the production of IFN-g by CD41 T cells(46). IL-4 also has been shown to inhibit IFN-g-induced responsesby macrophages and their production of proinflammatory mole-cules, such as IL-1a, IL-5, IL-12, GM-CSF, and TNF-a (45, 46).However, under certain in vitro conditions, IL-4 has been shown topossess proinflammatory activities on APC. Monocytes that havebeen pretreated with IL-4 for an extended period actually haveenhanced, rather than suppressed, IL-12 production in response toLPS orStaphylococcus aureus(53). IL-4 also has been shown toact synergistically with IL-12 in vitro to stimulate IFN-g secretionby splenic dendritic cells (54). Furthermore, IL-4 inhibits IL-10production by monocytes and up-regulates MHC class II, B7.1,and B7.2 expression on APC (45, 46).
The results of our studies in vivo support observations that IL-4can have proinflammatory effects on accessory cells. For example,IL-4 treatment was able to moderately exacerbate colitis when theT cells could not respond directly to IL-4. In parallel studies, weobserved an increase in MHC class II expression on cells in thecolonic LP of IL-4-treated nonreconstituted Rag22/2 mice, whichis in agreement with the ability of IL-4 to up-regulate MHC classII expression on monocytes in vitro (46). In addition, IL-10 wasable to block the IL-4-induced increased MHC class II expressionin the LP and, more importantly, abrogated the ability of IL-4 toexacerbate colitis upon T cell transfer. It is also possible that IL-10may be able to block enhanced IL-12 production by APC (45).Nevertheless, our studies with IL-4Ra2/2 T cells show that IL-4
FIGURE 6. IL-4 treatment increased MHCclass II expression in the LP of Rag22/2 mice.Immunohistochemical analysis of frozen sections(8 mm) from nonreconstituted Rag22/2 micetreated for 21 days with saline (A), IL-4 (B), orIL-4 plus IL-10 (C). All images are3200. Sec-tions were stained with anti-I-Ab mAb (red) andcounterstained with hematoxylin (blue).D, Co-lon section from saline-treated mouse stainedwith isotype control mAb.
2798 IL-4 TREATMENT PROMOTES Th1-TYPE COLITIS
by guest on May 17, 2018
http://ww
w.jim
munol.org/
Dow
nloaded from

may have some direct proinflammatory effect on Th1 cells. It isalso likely that IL-4 has proinflammatory effects on nonhemopoi-etic cells. Colonic epithelial cells express IL-4R and can increaseICAM-1 expression in response to IL-4, and thus may contributeto the exacerbation of disease in this model (47, 55).
While there is much evidence to support the role of IL-4 inallergic Th2-type responses, the role of IL-4 in the development ofan IL-12-dependent, IFN-g-producing immune response in vivoneeds to be more fully elucidated. In particular, which microenvi-ronmental conditions cause IL-4 to favor a Th2 vs Th1 inflamma-tory response remains to be determined. By developing a betterunderstanding of the interaction of Th1-associated and Th2-asso-ciated cytokines, more effective therapeutics for chronic inflam-matory diseases can be designed.
AcknowledgmentsWe gratefully acknowledge the skills of T. Bringman, E. Callas,J. Maskrey, J. Vargas, and Dr. J. Cupp in cell sorting; the technical assis-tance of the Schering-Plough Research Institute histology laboratory; Dr.Rene de Waal Malefyt for development and assistance with TaqMan anal-ysis of mRNA; and Dr. Heleen Scheerens for critical reading of themanuscript.
References1. O’Garra, A., and K. Murphy. 1994. Role of cytokines in determining T-lympho-
cyte function.Curr. Opin. Immunol. 6:458.2. Abbas, A. K., K. M. Murphy, and A. Sher. 1996. Functional diversity of helper
T lymphocytes.Nature 383:787.3. O’Garra, A., L. Steinman, and K. Gijbels. 1997. CD41 T-cell subsets in auto-
immunity. Curr. Op. Immunol. 9:872.4. Falcone, M., and N. Sarvetnick. 1999. Cytokines that regulate autoimmune re-
sponses.Curr. Opin. Immunol. 11:670.5. Racke, M. K., A. Bonomo, D. E. Scott, B. Cannella, A. Levine, C. S. Raine,
E. M. Shevach, and M. Rocken. 1994. Cytokine-induced immune deviation as atherapy for inflammatory autoimmune disease.J. Exp. Med. 180:1961.
6. Rocken, M., M. Racke, and E. M. Shevach. 1996. IL-4-induced immune devia-tion as antigen-specific therapy for inflammatory autoimmune disease.Immunol.Today 17:225.
7. Berg, D. J., R. Kuhn, K. Rajewsky, W. Muller, S. Menon, N. Davidson,G. Grunig, and D. Rennick. 1995. Interleukin-10 is a central regulator of theresponse to LPS in murine models of endotoxic shock and the Schwartzmanreaction but not endotoxin tolerance.J. Clin. Invest. 96:2339.
8. Berg, D. J., M. W. Leach, R. Kuhn, K. Rajewsky, W. Muller, N. J. Davidson, andD. Rennick. 1995. Interleukin 10 but not interleukin 4 is a natural suppressant ofcutaneous inflammatory responses.J. Exp. Med. 182:99.
9. Cua, D. J., H. Groux, D. R. Hinton, S. A. Stohlman, and R. L. Coffman. 1999.Transgenic interleukin 10 prevents induction of experimental autoimmune en-cephalomyelitis.J. Exp. Med. 189:1005.
10. Apparailly, F., C. Verwaerde, C. Jacquet, C. Auriault, J. Sany, and C. Jorgensen.1998. Adenovirus-mediated transfer of viral IL-10 gene inhibits murine collagen-induced arthritis.J. Immunol. 160:5213.
11. Ma, Y., S. Thornton, L. E. Duwel, G. P. Boivin, E. H. Giannini, J. M. Leiden,J. A. Bluestone, and R. Hirsch. 1998. Inhibition of collagen-induced arthritis inmice by viral IL-10 gene transfer.J. Immunol. 161:1516.
12. Lubberts, E., L. A. B. Joosten, L. van den Bersselaar, M. M. A. Helsen,A. C. Bakker, J. B. J. van Meurs, F. L. Graham, C. D. Richards, andW. B. van den Berg. 1999. Adenoviral vector-mediated overexpression of IL-4 inthe knee joint of mice with collagen-induced arthritis prevents cartilage destruc-tion. J. Immunol. 163:4546.
13. Finnegan, A., K. Mikecz, P. Tao, and T. T. Glant. 1999. Proteoglycan (aggrecan)-induced arthritis in BALB/c mice is a Th1-type disease regulated by Th2 cyto-kines.J. Immunol. 163:5383.
14. Mueller, R., T. Krahl, and N. Sarvetnick. 1996. Pancreatic expression of inter-leukin-4 abrogates insulitis and autoimmune diabetes in nonobese diabetic(NOD) mice.J. Exp. Med. 184:1093.
15. Mueller, R., L. M. Bradley, T. Krahl, and N. Sarvetnick. 1997. Mechanism un-derlying counterregulation of autoimmune diabetes by IL-4.Immunity 7:411.
16. Cameron, M. J., G. A. Arreaza, P. Zucker, S. W. Chensue, R. M. Strieter,S. Chakrabarti, and T. L. Delovitch. 1997. IL-4 prevents insulitis and insulin-dependent diabetes mellitus in nonobese diabetic mice by potentiation of regu-latory T helper-2 cell function.J. Immunol. 159:4686.
17. Scott, B., R. Liblau, S. Degermann, L. A. Marconi, L. Ogata, A. J. Caton,H. O. McDevitt, and D. Lo. 1994. A role for non-MHC genetic polymorphism insusceptibility to spontaneous autoimmunity.Immunity 1:73.
18. Maron, R., W. W. Hancock, A. Salvin, M. Hattori, V. Kuchroo, and H. L. Weiner.1999. Genetic susceptibility or resistance to autoimmune encephalomyelitis inMHC congenic mice is associated with differential production of pro- and anti-inflammatory cytokines.Int. Immunol. 11:1573.
19. Powrie, F., S. Menon, and R. L. Coffman. 1993. Interleukin-4 and interleukin-10synergize to inhibit cell-mediated immunity in vivo.Eur. J. Immunol. 23:3043.
20. Kitching, A. R., P. G. Tipping, X. R. Huang, D. A. Mutch, and S. R. Holdsworth.1997. Interleukin-4 and interleukin-10 attenuate established crescentic glomeru-lonephritis in mice.Kidney Intl. 52:52.
21. Ramanathan, S., Y. de Kozak, A. Saoudi, O. Goureau, P. H. Van der Meide,P. Druet, and B. Bellon. 1996. Recombinant IL-4 aggravates experimental auto-immune uveoretinitis in rats.J. Immunol. 157:2209.
22. Schuler, T., Z. Qin, S. Ibe, N. Noben-Trauth, and T. Blankenstein. 1999. T helpercell type 1-associated and cytotoxic T lymphocyte-mediated tumor immunity isimpaired in interleukin 4-deficient mice.J. Exp. Med. 189:803.
23. Mencacci, A., G. Del Sero, E. Cenci, C. Fe d’Ostiani, A. Bacci, C. Montagnoli,M. Kopf, and L. Romani. 1998. Endogenous interleukin 4 is required for devel-opment of protective CD41 T helper type 1 cell responses toCandida albicans.J. Exp. Med. 187:307.
24. Bagley, J., T. Sawada, Y. Wu, and J. Iacomini. 2000. A critical role for inter-leukin 4 in activating alloreactive CD4 T cells.Nat. Immun. 1:257.
25. Braegger, C. P., and T. T. MacDonald. 1994. Immune mechanisms in chronicinflammatory bowel disease.Ann. Allergy 72:135.
26. Bhan, A. K., E. Mizoguchi, R. N. Smith, and A. Mizoguchi. 1999. Colitis intransgenic and knockout animals as models of human inflammatory bowel dis-ease.Immunol. Rev. 169:195.
27. Fuss, I. J., M. Neurath, M. Boirivant, J. S. Klein, C. de la Motte, S. A. Strong,C. Fiocchi, and W. Strober. 1996. Disparate CD41 lamina propria (LP) lympho-kine secretion profiles in inflammatory bowel disease.J. Immunol. 157:1261.
28. Blumberg, R., L. J. Saubermann, and W. Strober. 1999. Animal models of mu-cosal inflammation and their relation to human inflammatory bowel disease.Curr. Opin. Immunol. 11:648.
29. Davidson, N. J., M. M. Fort, W. Muller, M. W. Leach, and D. M. Rennick. 2000.Chronic colitis in IL-102/2 mice: insufficient counter regulation of a Th1 re-sponse.Int. Rev. Immunol. 19:91.
30. Powrie, F. 1995. T cells in inflammatory bowel disease: protective and patho-genic roles.Immunity 3:171.
31. Simpson, S. J., S. Shah, M. Comisky, Y. P. de Jong, B. Wang, E. Mizoguchi, A.K. Bhan, and C. Terhorst. 1998. T cell-mediated pathology in two models ofexperimental colitis depends predominantly on the interleukin12/signal trans-ducer and activator of transcription (stat)-4 pathway, but is not conditional oninterferon-g expression by T cells.J. Exp. Med. 187:1225.
32. Takeda, K., B. E. Clausen, T. Kaisho, T. Tsujimura, N. Terada, I. Forster, andS. Akira. 1999. Enhanced Th1 activity and development of chronic enterocolitisin mice devoid of stat3 in macrophages and neutrophils.Immunity 10:39.
33. Powrie, F., J. Carlino, M. W. Leach, S. Mauze, and R. L. Coffman. 1996. Acritical role for transforming growth factor-b but not interleukin 4 in the sup-pression of T helper type 1-meditaed colitis by CD45RBlow CD41 T cells.J. Exp. Med. 183:2669.
34. Hogaboam, C. M., B. A. Vallance, A. Kumar, C. L. Addison, F. L. Graham,J. Gauldie, and S. M. Collins. 1997. Therapeutic effects of interleukin-4 genetransfer in experimental inflammatory bowel disease.J. Clin. Invest. 100:2766.
35. Powrie, F., R. Correa-Oliveira, S. Mauze, and R. L. Coffman. 1994. Regulatoryinteractions between CD4RBhi and CD45RBlow CD41 T cells are important forthe balance between protective and pathogenic cell-mediated immunity.J. Exp.Med. 179:589.
36. Davidson, N. J., S. A. Hudak, R. E. Lesley, S. Menon, M. W. Leach, andD. M. Rennick. 1998. IL-12, but not IFN-g, plays a major role in sustaining thechronic phase of colitis in IL-10-deficient mice.J. Immunol. 161:3143.
37. Powrie, F., M. W. Leach, S. Mauze, S. Menon, L. Barcomb, and R. L. Coffman.1994. Inhibition of Th1 responses prevents inflammatory bowel disease inscidmice reconstituted with CD45RBhi CD41 T cells. Immunity 1:553.
38. Kuhn, R., J. Lohler, D. Rennick, K. Rajewsky, and W. Muller. 1993. Interleukin-10-deficient mice develop chronic enterocolitis.Cell 75:263.
39. Mohrs, M., B. Ledermann, G. Kohler, A. Dorfmuller, A. Gessner, andF. Brombacher. 1999. Differences between IL-4 and IL-4 receptora-deficientmice in chronic leishmaniasis reveal a protective role for IL-13 in receptor sig-naling.J. Immunol. 162:7302.
40. Berg, D. J., N. J. Davidson, R. Kuhn, W. Muller, S. Menon, G. Holland,L. Thompson-Snipes, M. W. Leach, and D. Rennick. 1996. Entercolitis and coloncancer in interleukin-10-deficient mice are associated with aberrant cytokine pro-duction and CD41 TH1-like responses.J. Clin. Invest. 98:1010.
41. Bonecchi, R., G. Bianchi, P. P. Bordignon, D. D’Ambrosio, R. Lang, A. Borsatti,S. Sozzani, P. Allavena, P. A. Gray, A. Mantovani, et al. 1998. Differentialexpression of chemokine receptors and chemotactic responsiveness of type 1 Thelper cells (Th1s) and Th2s.J. Exp. Med. 187:129.
42. Tiffany, H. L., L. L. Lauten, J.-L. Gao, J. Pease, M. Locati, C. Combadiere,W. Modi, T. I. Bonner, and P. M. Murphy. 1997. Identification of CCR8: ahuman monocyte and thymus receptor for the CC chemokine I-309.J. Exp. Med.186:165.
43. Zingoni, A., H. Soto, J. A. Hedrick, A. Stoppacciaro, C. T. Storlazzi,F. Sinigaglia, D. D’Ambrosio, A. O’Garra, D. Robinson, M. Rocchi, et al. 1998.The chemokine receptor CCR8 is preferentially expressed in Th2 but not Th1cells.J. Immunol. 161:547.
2799The Journal of Immunology
by guest on May 17, 2018
http://ww
w.jim
munol.org/
Dow
nloaded from

44. de Vries, J. E., and J. Punnonen. 1996. Interleukin-4 and interleukin-13. InCy-tokine Regulation of Humoral Immunity. C. Snapper, ed. John Wiley & Sons,London, pp. 000–000.
45. de Waal Malefyt, R. 1999. Role of interleukin-10., interleukin-4, and interleu-kin-13 in resolving inflammatory responses. InInflammation: Basic Principlesand Clinical Correlates. G. A. Snyderman, ed. Lippincott-Williams & Wilkins,Philadelphia, pp. 837–849.
46. Chomarat, P., and J. Banchereau. 1998. Interleukin-4 and interleukin-13: theirsimilarities and discrepancies.Int. Rev. Immunol. 17:1.
47. Reinecker, H., and D. Podolsky. 1995. Human intestinal epithelial cells expressfunctional cytokine receptors sharing the commongc chain of the interleukin 2receptor.Proc. Natl. Acad. Sci. USA 92:8353.
48. Terres, G., and R. Coffman. 1998. The role of IL-4 and IL-10 cytokines in con-trolling an anti-tumor response in vivo.Int. Immunol. 10:823.
49. Asseman, C., S. Mauze, M. W. Leach, R. L. Coffman, and F. Powrie. 1999. Anessential role of interleukin 10 in the function of regulatory T cells that inhibitintestinal inflammation.J. Exp. Med. 190:995.
50. Boirivant, M., I. J. Fuss, A. Chu, and W. Strober. 1998. Oxazolone colitis: amurine model of T helper cell type 2 colitis treatable with antibodies to inter-leukin 4.J. Exp. Med. 188:1929.
51. Dohi, T., K. Fujihashi, P. D. Rennert, K. Iwatani, H. Kiyono, and J. R. McGhee.1999. Hapten-induced colitis is associated with colonic patch hypertrophy and Thelper cell 2-type responses.J. Exp. Med. 189:11169.
52. Farber, J. M. 1997. MIG and IP-10: CXC chemokines that target lymphocytes.J. Leukocyte Biol. 61:246.
53. D’Andrea, A., X. Ma, M. Aste-Amezaga, C. Paganin, and G. Trinchieri. 1995.Stimulatory and inhibitory effects of interleukin (IL)-4 and IL-13 on the produc-tion of cytokines by human peripheral blood mononuclear cells: priming forIL-12 and tumor necrosis factora production.J. Exp. Med. 181:537.
54. Fukao, T., S. Matsuda, and S. Koyasu. 2000. Synergistic effects of IL-4 and IL-18on IL-12-dependent IFN-g production of dendritic cells.J. Immunol. 164:64.
55. Striz, I., T. Mio, Y. Adachi, P. Heires, R. A. Robbins, J. R. Spurzem, M. J. Illig,D. J. Romberger, and S. I. Rennard. 1999. IL-4 induces ICAM-1 expression inhuman bronchial epithelial cells and potentiates TNFa. Am. J. Physiol. 277:L58.
2800 IL-4 TREATMENT PROMOTES Th1-TYPE COLITIS
by guest on May 17, 2018
http://ww
w.jim
munol.org/
Dow
nloaded from





























