Embed Size (px)
Citation preview

Identification of a set of calcium-binding proteins in reticuloplasm, the
luminal content of the endoplasmic reticulum
DARRYL R. J. MACER and GORDON L. E. KOCH
Laboratory of Molecular Hiology, Hills Road, Cambridge CB2 2QII, luigland
Summary
A procedure was developed for the isolation ofreticuloplasm, the luminal material of the endo-plasmic reticulum (ER). A reticuloplasm-rich ex-tract was prepared from a murine plasmacytomacell line that contains large amounts of ER, by firstextracting the cytoplasmic contents using hypotoniclysis to yield ER-rich 'shells' followed by mechan-ical lysis to release the ER contents. The extractcontains five major proteins with apparent molecu-lar weights of 100, 75, 60, 58 and 55 (xlO3)Mr bySDS— polyacrylamide gel electrophoresis. The 100,75 and 58 (Xl03)Mr species were identified as theknown ER proteins endoplasmin, BiP and PD1,respectively. The ER association of the 60 and 55(X103) Mr proteins was confirmed by conf ocal fluor-escence microscopy with affinity-purified anti-bodies.
Equilibrium dialysis with isolated reticuloplasmgave a calcium-binding capacity of 300 nmoles cal-cium per mg protein with half-maximal binding at3 mM-Ca2+. Purified endoplasmin bound 280 nmolescalcium per mg protein at a calcium concentrationof 5mM-Ca2+.
A calcium overlay test revealed that, in additionto endoplasmin, reticuloplasm contained at leastthree other calcium-binding proteins: i.e. BiP, PDIand the 55xlO3Mr protein, respectively, withendoplasmin and the 5 5 x 1 0 ^ , protein (CRP55)accounting for the major proportion of the calcium-binding activity.
Treatment of cells with calcium ionophore led tothe specific over-expression of the major calcium-binding reticuloplasmins endoplasmin, BiP andCRP55.
These studies show that the lumen of the ERcontains a family of proteins with the capacity tobind significant amounts of calcium in the millimo-lar range and thereby to confer upon the ER theability to perform a calcium storage function anal-ogous to that of the sarcoplasmic reticulum inmuscle cells.
Key words: endoplasmic reticulum, reticuloplasm, calcium-binding proteins, endoplasmin, BiP, protein disulphideisomerase, CRP55.
Introduction
Calcium ions play a vital role in a variety of cellularprocesses. This is achieved through a stringent control ofcalcium levels within cells (Carafoli, 1987; Somlyo,1984). One of the major mechanisms for maintainingintracellular calcium in the sub-micromolar range is theCa2+-ATPase system, which pumps calcium actively outof the cytoplasm through the plasma membrane (Camp-bell, 1983). However, it is also known that intracellularmembrane systems can sequester significant amounts ofcalcium and release this for use in intracellular signalling(Hasselbach & Makinose, 1961; Somlyo, 1984; Baker,1986; Streb et al. 1983). One of the best-characterizedintracellular systems operating under normal physiologi-cal conditions is the sarcoplasmic reticulum (SR) inmuscle, which has been clearly established as the system
Journal of Cell Science 91, 61-70 (1988)Printed in Great Britain © The Company of Biologists Limited 1988
for removing Ca2+ from muscle cytoplasm to levels thatwould induce dissociation from troponin (MacLennan &Holland, 1975).
It is now widely accepted that in non-muscle systemsthe mitochondria play a relatively minor role in calciumhomeostasis and attention has been focussed on the roleof the endoplasmic reticulum (ER) in the regulation ofcytosolic calcium levels (Henkart, 1975; Somlyo, 1984;Baker, 1986; Streb et al. 1983; Berridge & Irvine, 1984).In this respect it has been suggested that the ER might beable to store and release calcium in a manner analogous tothat involved in the sarcoplasmic reticulum (Baker,1986). Thus, it has been shown directly that the calciumconcentration within the ER is relatively high and thatthe calcium can be released by agents such as inositoltriphosphate and GTP (Streb et al. 1984; Gill et al.1986). Isolated microsomes can accumulate calcium in an
61

ATP-dependent process and release it under the in-fluence of the above-mentioned agents (Spat et al. 1987).The existence of a Ca2+-ATPase activity capable ofpumping calcium into the ER has also been suggested(Moore & Klaus-Freedmann, 1983). However, one majordeficiency in this area is a lack of knowledge about thenature of the stored calcium in the ER. By analogy withSR one would expect the ER in non-muscle cells tocontain one or more proteins in the luminal space orreticuloplasm that could carry out a role analogous to thatof calsequestrin (MacLennan & Holland, 1975). Indeed,it has recently been established that the reticuloplasm is aprotein-rich medium that contains a family of abundantresident proteins called reticuloplasmins (Koch, 1987),one of which is a major glycoprotein called endoplasmin,which is able to bind significant amounts of calcium in themillimolar range (Koch et al. 1986a).
In this study we have carried out a systematic study ofreticuloplasm with a view to identifying the repertoire ofcalcium-binding proteins associated with the ER. Toachieve this, we have developed a general procedure forisolating reticuloplasm, identified and characterized themajor abundant proteins in the isolate and examinedthese for calcium-binding activity.
Materials and methods
Growth and preparation of cellsMOPC-315 cells (Eisen et al. 1968) were grown in Dulbecco'smodified Eagle's medium (DMEM) with 10% foetal calf serum,100000 units ml" ' penicillin/streptomycin and 4 mM-pyruvateto a density of ~5X 105 cells ml" ' , harvested by centrifugationat 300if and washed thoroughly with phosphate-buffered saline(PBS).
Preparation of reticuloplasmWashed MOPC-315 cells were suspended at IX 107 cells min"'in lOmM-Tris-HC1, p H 7 5 , on ice and allowed to swell andlyse completely. The resultant 'shells' were centrifuged out at1000£, suspended in PBS at 1 X 10s cells equivalent per ml anddisrupted by syringing through an 18 gauge needle. The celldebris was pelletted by centrifugingat 100 000g for 30 min. Thesupernatant was collected and protease inhibitors (Koch et al.1986a) added. The supernatant was stored at 4°C. For longerstorage at —20°C glycerol was added to 50 % to preventfreezing, and dialysed out before use.
Preparation of sarcoplasmic reticulum vesiclesThe method used was based on that of MacLennan (1970), andwas used to obtain vesicles from rabbit and munne skeletalmuscle. Muscle tissue was homogenized in an equal volume of120niM-NaCl, 5 mM-imidazole, pH7-4. It was spun at 1000gfor 5 min, and the pellet was re-extracted with buffer. Thecombined superantants were squeezed through four layers ofmuslin, and the pH adjusted to 7 4 . The suspension was spun at10 000 revs min"1 in a SS-34 rotor for IS min. The supernatantwas squeezed through four layers of muslin, and centrifuged at16 000 revs min"1 in a SS-34 rotor for 50 min. The pellet is thepreparation of sarcoplasmic reticulum vesicles.
Purification of calsequestrinSarcoplasmic reticulum vesicles were suspended in PBS and
PS LYS SH RET
*«
Fig. 1. Isolation of enriched reticuloplasm from a murineplasmacytoma cell line (MOPC-315). PS, protein standards(from top): glycogen phosphorylase (95X 10"'il/r); bovineserum albumin (67X103); ovalbumin (45X103); carbonicanhydrase (30X103); Q--lactalbumin (17X103). LYS, lysatefrom MOPC-315 cells; SH, hypotonic shells from MOPC-315cells; RET, enriched reticuloplasm extracted from MOPC-315 shells. The major proteins enriched in the reticuloplasmpreparation are arrowed.
saponin was added to a final concentration of 001 %. After10 min on ice the membranes were centrifuged out at 100 000 £•for 50 min. The supernatant contains calsequestrin at >95 %purity.
Preparation of antibodies to individual reticuloplasminsRabbits (Dutch White or New Zealand White) were immunizedwith 1-2 mg of purified reticuloplasm. The first injection waswith Freund's complete adjuvant and subsequent injectionswere at monthly intervals without adjuvant. Serum was col-lected by standard procedures and stored in 0 0 2 % sodiumazide.
Monospecific antibodies to particular proteins in the purifiedreticuloplasm were prepared, as described (Koch et al. 1986a).A sample of reticuloplasm was fractionated on a preparativeSDS-polyacrylamide gel and the proteins were transferred to a
62 D. R. J. Macer and G. L. E. Koch

.EP
P6O fDIP55
EP BiP
P60 PDl P55Fig. 2. Monospecific antibodies to the major reticuloplasmins from MOPC-315 cells. Antibodies were prepared as described inMaterials and methods, and tested by immunoblotting analysis on two-dimensional gels of lysates from MOPC-315 cells (thesamples had been partially depleted for endoplasmin (Koch el al. 1986o) with ConA-agarose). Panel L shows the proteinpattern from the lysate; and panels EP, BiP, P60, PDl and P55 show immunoblots developed with antibodies to endoplasmin,BiP, P60, PDl and CRPS5, respectively (see text).
nitrocellulose sheet by electroblotting (see below). The nitro-cellulose was stained with O'Ol % Ponceau Red in 5% aceticacid and the strips corresponding to each protein componentwere carefully excised. The strips were then immersed in theundiluted immune serum and mixed by rotation at 4°C for atleast 24 h. The strips were removed, washed thoroughly withPBS and the antibodies eluted with O2M-glycine-HCl, pH2(lOmin at 4°C). The eluate was immediately neutralized top H 7 5 with 1 M-Tris-base and tested for antibodies by lmmu-noblotting on cell lysates. Strips can be used repeatedly for theextraction of antibody from unfractionated serum.
Gel eiectmphoresis and immunobloltingSDS—polyacrylamide gel electrophoresis was carried out ac-cording to the method of Laemmli (1970) and two-dimensionalgel electrophoresis by the method of O'Farrell (1975). Immuno-blotting was as described by Towbin el al. (1979). Rabbitantibodies were developed with IZ5I-labelled Protein A (Amer-sham).
Immunojiiiorescence microscopyImmunofluorescence microscopy was carried out as described
(Koch el al. 1987) using the confocal microscope described byWhite el al. (1987). MOPC-315 cells were fixed in 3-5%formaldehyde in PBS (5min), permeabilized with 1 % NonidetP40 (5 min), labelled with the appropriate antibody (5 min) anddeveloped with fluorescein-conjugated goat anti-rabbit Ig.
Purification of pmteins fivrn reticuloplasmEndoplasmin was selectively extracted from purified reticulo-plasm with Con A-Sepharose as described (Koch el al. 1986«).The non-bound fraction was dialysed against lOmM-Tns- HC1,p H 7 5 , and made up to 50% saturation with solid ammoniumsulphate. The precipitate was centrifuged out and the super-natant made up to 85 % saturation with solid ammoniumsulphate. The precipitate was collected by centrifugation,dissolved in lOmM-Tris- HC1, pH 7-5, and dialysed against thesame buffer. The sample was applied to DEAE-Sephadexequilibriated with lOmM-Tris-HCI, p H 7 5 , and eluted with alinear gradient (10 mM to 70mM-Tris- HCI, pH 7 5). Fractionswere monitored by SDS-polyacrylamide gel electrophoresisand immunoblotting and those containing (in order of elution)P58, BiP, PDl and CRP55 were pooled and concentrated. Eachsample was fractionated by gel filtration on Sephacryl S200 to
Calcium-binding reticuloplasmins 63

Fig. 3. Immunofluorescence localization of the major reticuloplasmins from MOPC-315 cells. Samples were prepared andstained, as described (Koch et al. 1987). A. Endoplasmin; B, P60; C, PD1; D, CRPSS.
give the purified proteins. All of the purified preparations were>90% pure for the respective protein (Macer, 1988).
Results
Purification of reticuloplasmThe approach used for the isolation of the luminalcontents of the ER, referred to as reticuloplasm, wasbased on previous studies on the major glycoproteinendoplasmin (Koch et al. 1986a, 1987). We have pre-viously established (Koch et al. 1988) that endoplasmin isa major component of reticuloplasm and can therefore beused a marker for the isolation of the contents of the ER.It was found that the exposure of murine plasmacytomacells (in this study we used the MOPC-315 line but anyplasmacytoma cell line can be employed) to hypotonic
conditions resulted in the extraction of all the cvtoplasmicproteins, but endoplasmin itself was retained in the shellsproduced by the lysis. Fig. 1 shows that, in addition toendoplasmin, the major 100xl03/V/r protein in suchpreparations, several other major proteins are selectivelyretained in the shells. When the shells were disruptedmechanically by repeated syringing through a hypoder-mic needle and the particulate material was pclletted byhigh-speed centrifugation, the supernatant material con-tained five major proteins with apparent molecularweights (on SDS-polyacrylamide gels) of 100, 75, 60, 58and 55(XlO3).'l/r, respectively.
Characterization of the major proteins in isolatedreticuloplasmThe major proteins in the soluble extract from the shells
64 D. R. J. Macer and G. L. E. Koch
300 a b c d e
-o
16~5
100
4 6[Ca] (ITIM)
10
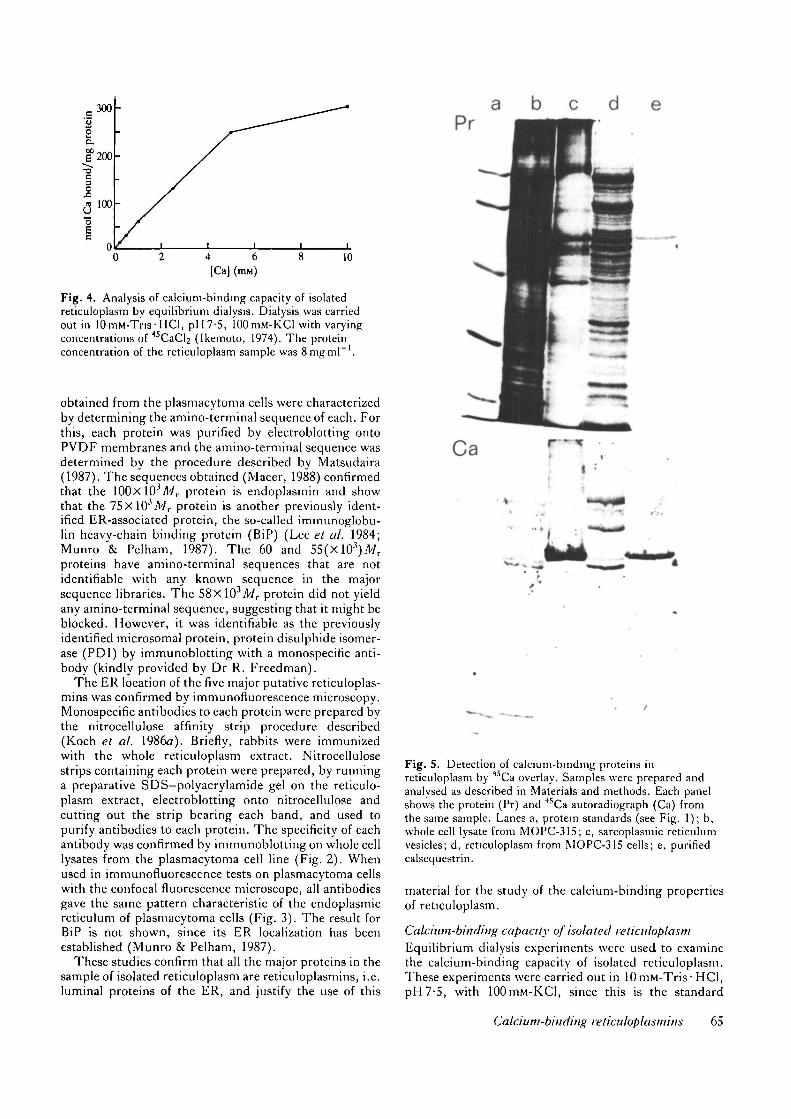
Fig. 4. Analysis of calcium-binding capacity of isolatedreticuloplasm by equilibrium dialysis. Dialysis was carriedout in 10niM-Tris-HCl, pH7-5, 100mM-K.Cl with varyingconcentrations of 45CaCl2 (Ikemoto, 1974). The proteinconcentration of the reticuloplasm sample was 8mgml~' .
obtained from the plasmacytoma cells were characterizedby determining the amino-terminal sequence of each. Forthis, each protein was purified by electroblotting ontoPVDF membranes and the amino-terminal sequence wasdetermined by the procedure described by Matsudaira(1987). The sequences obtained (Macer, 1988) confirmedthat the 100xl03iV/r protein is endoplasmin and showthat the 75xl03iV/r protein is another previously ident-ified ER-associated protein, the so-called immunoglobu-lin heavy-chain binding protein (BiP) (Lee et al. 1984;Munro & Pelham, 1987). The 60 and 55(XlO3)Mr
proteins have amino-terminal sequences that are notidentifiable with any known sequence in the majorsequence libraries. The 58xl03yV/r protein did not yieldany amino-terminal sequence, suggesting that it might beblocked. However, it was identifiable as the previouslyidentified microsomal protein, protein disulphide isomer-ase (PDI) by immunoblotting with a monospecific anti-body (kindly provided by Dr R. Freedman).
The ER location of the five major putative reticuloplas-mins was confirmed by immunofluoreseence microscopy.Monospecific antibodies to each protein were prepared bythe nitrocellulose affinity strip procedure described(Koch et al. 1986a). Briefly, rabbits were immunizedwith the whole reticuloplasm extract. Nitrocellulosestrips containing each protein were prepared, by runninga preparative SDS-polyacrylamide gel on the reticulo-plasm extract, electroblotting onto nitrocellulose andcutting out the strip bearing each band, and used topurify antibodies to each protein. The specificity of eachantibody was confirmed by immunoblotting on whole celllysates from the plasmacytoma cell line (Fig. 2). Whenused in immunofluoreseence tests on plasmacytoma cellswith the confocal fluorescence microscope, all antibodiesgave the same pattern characteristic of the endoplasmicreticulum of plasmacytoma cells (Fig. 3). The result forBiP is not shown, since its ER localization has beenestablished (Munro & Pelham, 1987).
These studies confirm that all the major proteins in thesample of isolated reticuloplasm are reticuloplasmins, i.e.luminal proteins of the ER, and justify the use of this
Fig. S. Detection of calcium-binding proteins inreticuloplasm by 4SCa overlay. Samples were prepared andanalysed as described in Materials and methods. Each panelshows the protein (Pr) and 45Ca autoradiograph (Ca) fromthe same sample. Lanes a, protein standards (see Fig. 1); b,whole cell lysate from MOPC-315; c, sarcoplasmic reticulumvesicles; d, reticuloplasm from MOPC-315 cells; e, purifiedcalsequestrin.
material for the study of the calcium-binding propertiesof reticuloplasm.
Calcium-binding capacity of isolated reticuloplasmEquilibrium dialysis experiments were used to examinethe calcium-binding capacity of isolated reticuloplasm.These experiments were carried out in lOmM-Tris- HC1,pH7-5, with lOOmM-KCl, since this is the standard
Calcium-binding reticuloplasmins 65

Ca Pr
Fig. 6. Identification of calcium-binding proteins in enriched reticuloplasm. Enriched reticuloplasm was purified, fractionatedon two-dimensional polyacrylamide gels (O'Farrell, 1975), transferred to nitrocellulose (Towbin el al. 1979) and developed with45Ca (Koch et al. 1986). The name spots were identified with the corresponding monospecific antibodies (Fig. 2). The samplewas partially depleted for endoplasmin with ConA-agarose (Koch et al. 1986a). Spots 1, endoplasmin; 2, BiP; 3, PD1; 4,CRP55.
buffer used in the study of calcium-binding by sarcoplas-mic reticulum vesicles and the major calcium-storageprotein calsequestrin (Campbell et al. 1983). Fig. 4shows that calcium-binding saturates in the millimolarrange, half-maximal binding occurring at about 2-5 raM-calcium. At lOmM-calcium the isolated reticuloplasmbinds ~300nmoles calcium per mg protein.
Previous studies have shown that endoplasmin is one ofthe major calcium-binding proteins in plasmacytoma cells(Koch et al. 1986a). However, the binding studies werenot performed under equilibrium conditions so they wererepeated here using equilibrium dialysis, as describedabove. At 5 mM-calcium native endoplasmin bound~280nmoles calcium per mg protein, supporting theprevious suggestions that endoplasmin is a major cal-cium-binding protein of the ER.
Calcium-binding proteins in reticuloplasmThe repertoire of calcium-binding proteins in reticulop-lasm was examined by the SDS-polyacrylamide gel/elec-troblotting/4SCa overlay technique used previously
(Koch et al. 19866). The advantage of this technique,apart from its intrinsic convenience, is that it can be usedon complex mixtures as well as pure samples of proteins(Fig. 5) and with a sample of purified sarcoplasmicreticulum vesicles (Fig. 5). In the synthesis mixture,significant calcium-binding is observed with only theknown calcium-binding protein a'-lactalbumin. In thesample of SR vesicles, the major calcium-binding specieshas an apparent molecular weight of ~55xlO3 and co-migrates with pure calsequestrin, which itself bindssubstantial amounts of calcium under the conditionsused. We therefore conclude that the 4SCa overlay tech-nique was suitable for the detection of calcium-bindingproteins of the calsequestrin-type, that is, high-capacityand low-affinity.
Fig. 5 shows that there are several calcium-bindingproteins in a whole cell lysate derived from the plasma-cytoma cell. The major binding species have apparentmolecular weights of 100, 90, 75, 58 and 55X103,respectively. Fig. 5 shows that the 100, 75, 58 and55(XlO3)Mr calcium-binding species co-purify with re-
66 D. R. y. Macer and G. L. E. Koch
I 0 I I I
o 0 10
16
12
10[Cal (M)
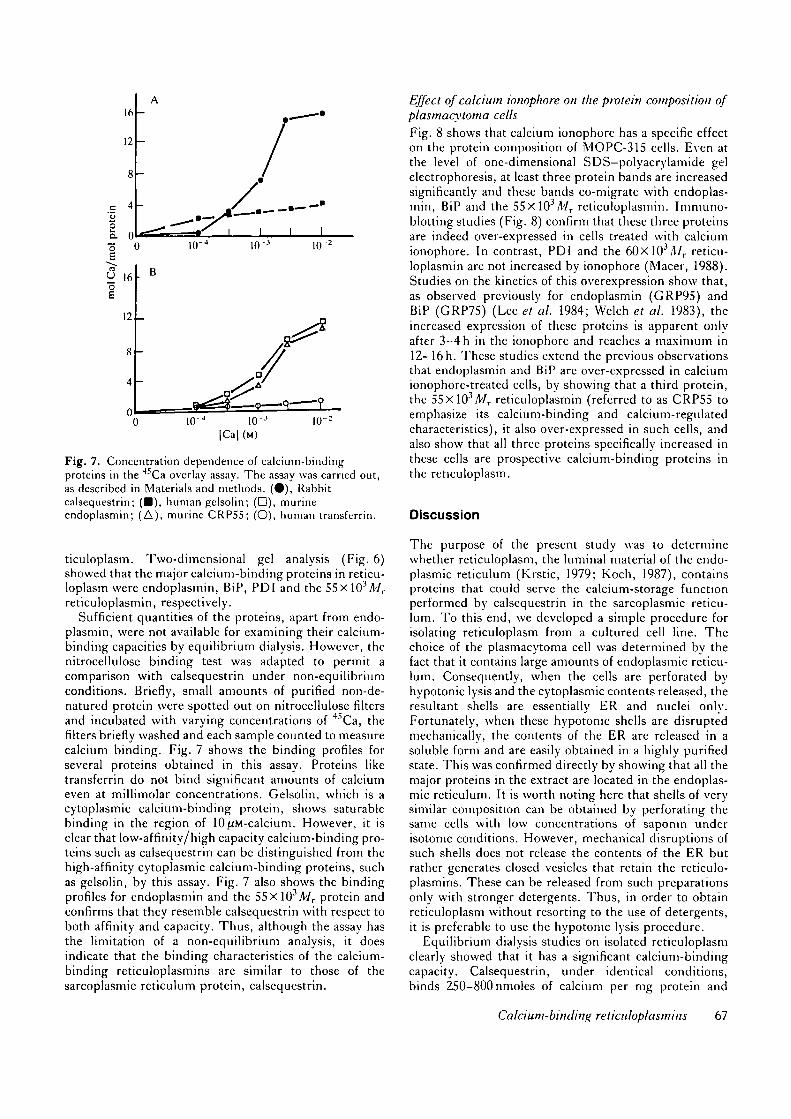
Fig. 7. Concentration dependence of calcium-bindingproteins in the 45Ca overlay assay. The assay was carried out,as described in Materials and methods. (•) , Rabbitcalsequestrin; (•) , human gelsolin; (D), murineendoplasmin; (A), murine CRPS5; (O), human transferrin.
ticuloplasm. Two-dimensional gel analysis (Fig. 6)showed that the major calcium-binding proteins in reticu-loplasm were endoplasmin, BiP, PDI and the 55xl03Mr
reticuloplasmin, respectively.Sufficient quantities of the proteins, apart from endo-
plasmin, were not available for examining their calcium-binding capacities by equilibrium dialysis. However, thenitrocellulose binding test was adapted to permit acomparison with calsequestrin under non-equilibriumconditions. Briefly, small amounts of purified non-de-natured protein were spotted out on nitrocellulose filtersand incubated with varying concentrations of 4SCa, thefilters briefly washed and each sample counted to measurecalcium binding. Fig. 7 shows the binding profiles forseveral proteins obtained in this assay. Proteins liketransferrin do not bind significant amounts of calciumeven at millimolar concentrations. Gelsolin, which is acytoplasmic calcium-binding protein, shows saturablebinding in the region of 10^M-calcium. However, it isclear that low-affinity/high capacity calcium-binding pro-teins such as calsequestrin can be distinguished from thehigh-affinity cytoplasmic calcium-binding proteins, suchas gelsolin, by this assay. Fig. 7 also shows the bindingprofiles for endoplasmin and the 55xl03il7r protein andconfirms that they resemble calsequestrin with respect toboth affinity and capacity. Thus, although the assay hasthe limitation of a non-equilibrium analysis, it doesindicate that the binding characteristics of the calcium-binding reticuloplasmins are similar to those of thesarcoplasmic reticulum protein, calsequestrin.
Effect of calcium ionophore on the protein composition ofplasmacytoma cellsFig. 8 shows that calcium ionophore has a specific effecton the protein composition of MOPC-315 cells. Even atthe level of one-dimensional SDS-polyacrylamide gelelectrophoresis, at least three protein bands are increasedsignificantly and these bands co-migrate with endoplas-min, BiP and the 55xlO3Jl7r reticuloplasmin. Immuno-blotting studies (Fig. 8) confirm that these three proteinsare indeed over-expressed in cells treated with calciumionophore. In contrast, PDI and the 60xl03/l/ r reticu-loplasmin are not increased by ionophore (Macer, 1988).Studies on the kinetics of this overexpression show that,as observed previously for endoplasmin (GRP95) andBiP (GRP75) (Lee et al. 1984; Welch et al. 1983), theincreased expression of these proteins is apparent onlyafter 3-4 h in the ionophore and reaches a maximum in12-16 h. These studies extend the previous observationsthat endoplasmin and BiP are over-expressed in calciumionophore-treated cells, by showing that a third protein,the 55xl03yV/r reticuloplasmin (referred to as CRP55 toemphasize its calcium-binding and calcium-regulatedcharacteristics), it also over-expressed in such cells, andalso show that all three proteins specifically increased inthese cells are prospective calcium-binding proteins inthe reticuloplasm.
Discussion
The purpose of the present study was to determinewhether reticuloplasm, the luminal material of the endo-plasmic reticulum (Krstic, 1979; Koch, 1987), containsproteins that could serve the calcium-storage functionperformed by calsequestrin in the sarcoplasmic reticu-lum. To this end, we developed a simple procedure forisolating reticuloplasm from a cultured cell line. Thechoice of the plasmacytoma cell was determined by thefact that it contains large amounts of endoplasmic reticu-lum. Consequently, when the cells are perforated byhypotonic lysis and the cytoplasmic contents released, theresultant shells are essentially ER and nuclei only.Fortunately, when these hypotonic shells are disruptedmechanically, the contents of the ER are released in asoluble form and are easily obtained in a highly purifiedstate. This was confirmed directly by showing that all themajor proteins in the extract are located in the endoplas-mic reticulum. It is worth noting here that shells of verysimilar composition can be obtained by perforating thesame cells with low concentrations of saponin underisotonic conditions. However, mechanical disruptions ofsuch shells does not release the contents of the ER butrather generates closed vesicles that retain the reticulo-plasmins. These can be released from such preparationsonly with stronger detergents. Thus, in order to obtainreticuloplasm without resorting to the use of detergents,it is preferable to use the hypotonic lysis procedure.
Equilibrium dialysis studies on isolated reticuloplasmclearly showed that it has a significant calcium-bindingcapacity. Calsequestrin, under identical conditions,binds 250-800 nmoles of calcium per mg protein and
Calcium-binding reticuloplasmins 67

Fig. 8. Increased expression of calcium-bindingreticuloplasmins in calcium inophore-treated cells.MOPC-315 cells were incubated with ( + ) andwithout ( —) 5;tM-A23187 for 16h and analysed onSDS-polyacrylamide gels and immunoblottingusing monospecific antibodies. The antibodies, toendoplasmin (EP), BiP and CRP55 (P55), wereused successively on the same sample. A. Proteinstain; arrows show proteins with increasedexpression in lonophore, which co-migrate withendoplasmin (EP), BiP and CRP55 (P55),respectively. B. Immunoblots showing, from thetop, endoplasmin, BiP and CRP55.
most of this binding occurs in the millimolar range (Cala& Jones, 1983; Campbell et al. 1983; Mawere/ al. 1985).In the case of reticuloplasm, half-maximal binding is at2-5 mM and at lOmM-calcium the binding capacity is300nmoles per mg protein. Thus, the calcium-bindingactivity in reticuloplasm is comparable to that of calse-questrin. The implication is that reticuloplasm containsproteins that have the ability to bind calcium with highcapacity in the millimolar range. However, in prelimi-nary studies with an antibody to calsequestrin, no evi-dence was obtained for a calsequestrin-like protein associ-ated with the ER itself. Therefore, it was necessary todetermine which proteins actually performed the cal-cium-binding function in reticuloplasm.
The procedure used to identify the calcium-bindingproteins in reticuloplasm was to fractionate the reticulo-plasm by gel electrophoresis, transfer the proteins tonitrocellulose and probe directly for calcium-bindingwith 45Ca. This approach has been used previously toidentify proteins that bind calcium in the micromolarrange (Koch et al. 1986a,6; Maruyama et al. 1984).However, using calsequestrin as a model, we have shownthat the method also works for low-affinity/high-capacity
calcium-binding proteins. This is probably not surpris-ing, since the calcium-binding sites in such proteinsappear to consist of clusters of negative charges, ratherthan any specific fold, such as the E-F hand found in thehigh-affinity calcium-binding proteins. It is also worthemphasizing that the test is quite specific, in that it clearlydistinguishes between proteins that are known to bindsignificant amounts of calcium and those that do not. Inthe case of endoplasmin it was directly confirmed byequilibrium dialysis that the protein does bind significantamounts of calcium under the conditions used in thecalsequestrin binding assay. In view of this, we concludethat endoplasmin, BiP, PDI and CRP55 are all signifi-cant calcium-binding proteins of the reticuloplasm. Ofthese, CRP55 appears to have a similar calcium-bindingcapacity and affinity to calsequestrin, whilst endoplasminis of moderate capacity. BiP and PDI bind smalleramounts of calcium but their capacity appears significantenough to make a contribution to the total calcium-binding capacity of reticuloplasm.
The main conclusion to emerge from these studies isthat the lumen of the ER does contain a number ofproteins that could participate in the calcium storage
68 D. R. jf. Macer and G. L. E. Koch

function attributed to this organelle in non-muscle cells.Since the proteins concerned are known to exist in the ERof most, and probably all, eukaryotic cells (Macer, 1988),it follows that the ER can function as a general calciumstorage organelle.
The obvious question that arises now concerns thedemonstration that the proteins actually perform thestorage function in vivo. This is not easy to do and evenin the case of calsequestrin, it is based only on circum-stantial evidence. However, at least one line of evidencedoes exist for this in the case of the ER proteins. It hasbeen known for some time that calcium ionophores have avery specific effect on the protein composition of cells.Previous studies showed that endoplasmin and BiP wereover-expressed in ionophore-treated cells (Lee, 1987). Inthis study we have confirmed that a third protein, the55xl03iV/r reticuloplasmin identified in this study(CRP5S), is also over-expressed in ionophore-treatedcells. Thus, the specific effect of altered calcium metab-olism in cells is to induce the increased production ofthree ER proteins that have the capacity to bind calcium.It is reasonable to speculate that such a response suggestsa role for the calcium-binding, calcium-regulated pro-teins in calcium homeostasis. One possibility is that theresponse leads to an increase in the intrinsic calciumstorage capacity of the ER and thereby permits the cell tocounteract the damaging effects of increased cytosoliccalcium caused by the ionophore. Such an explanation isalso consistent with the established phenomenon that avariety of stresses can also induce the synthesis of thecalcium-binding reticuloplasmins. It is believed that oneof the common events following the exposure of cells tostress is an alteration in calcium levels in the cytosol(Trump et at. 1981). If not corrected, this can lead to celldeath. The observed increased synthesis of the prospec-tive calcium storage proteins in the ER could be one wayof preventing this calcium-induced catastrophe.
In this study, we have been concerned with thequestion of whether the ER lumen does contain proteinsthat could carry out a calcium-storage function and weconclude that it does. However, it should be emphasizedthat, unlike the sarcoplasmic reticulum, the ER performsseveral other functions and the function of the relevantcalcium-binding proteins may not be confined to calcium-storage.
References
BAKER, P. F. (1986). GTP and calcium release. Nature, Loud. 320,395.
BERRIDGE, M. J. & IRVINE, R. F. (1984). Inositol triphosphate, anovel second messenger in cellular signal transduction. Nature,Lond 312, 315-321.
CALA, S. E. & JONES, L. R. (1983). Rapid purification ofcalsequestrin from cardiac muscle sarcoplasmic reticulum by Ca2+
dependent clution from phcnyl-Sepharosc. J. biol. Chew 258,11932-11936.
CAMPBELL, A. K. (1983). lntracelhdar Calcium, its Universal Roleas Regulator. New York: Wiley.
CAMPBELL, K. P., MACLENNAN, D. H., JORGENSEN, A. O. &
MlNTZEL, M. C. (1983). Purification and characterisation ofcalsequestrin from canine cardiac sarcoplasmic reticulum and
identification of the 53 000 dalton glycoprotein. J. biol Chem. 258,1197-1204.
CARAFOLI, E. (1987). Intracellular calcium homeostasis. A. Rev.Diochem. 56, 395-433.
EISEN, H. N., SIMMS, E. S. & POTTER, M. (1968). Mouse myelomaproteins with anti hapten antibody activity. The protein producedby plasma cell tumour MOPC-315. Biochemistry 7, 4126-4133.
GILL, D. L., MEDA, T., CHEUCH, S. J. & N O E L . M . \V. (1986).
Ca2+ release from the endoplasmic reticulum is mediated by aguanine nucleotide regulatory mechanism. Nature, Loud. 320,461-464.
HASSELBACH, W. & MAKINOSE, M. (1961). Die calcium pumpe dcrenschaffungs grana. Biochem. Z. 333, 518-528.
HENKART, M. (1975). Endoplasmic reticulum sequesters calcium inthe squid giant axon. Biophys. J. 15, 267a.
IKEMOTO, N. (1974). The calcium binding sites involved in theregulation of the purified adenosine triphosphatase of thesarcoplasmic reticulum. J. biol. Chem. 249, 649-651.
KRSTIC, R. V. (1979). infrastructure of the Mammalian Cell. Berlin:Spnnger-Verlag.
KOCH, G. L. E. (1987). Reticuloplasmins, a novel group of proteinsin the endoplasmic reticulum. J. Cell Sci. 87, 491-492.
KOCH, G. L. E., MACER, D. R. J. & SMITH, M. J. (1987).
Visualization of the intact endoplasmic reticulum byimmunofluorescence with antibodies to the major ER glycoprotein,cndoplasm. J. Cell Sa 87,535-542.
KOCH, G. L. E., MACER, D. R. J. & WOODING, F. B. P. (1988).
Endoplasmin is a reticuloplasmin. J. Cell Set. 90, 485-491.KOCH, G., SMITH, M., MACER, D., WEBSTER, P. & MORTARA, R.
(1986a). Endoplasmic reticulum contains a common abundantcalcium-binding glycoprotein endoplasmin. J. Cell Set. 86,217-232.
KOCH, G., SMITH, M., TWENTYMAN, P. & WRIGHT, K. (19866).
Identification of a novel calcium-binding protein (CP22) in multi-drug resistant murine and hamster cells. FEBS Lett. 195, 275—279.
LEE, A. S. (1987). Co-ordinated regulation of a set of genes byglucose and calcium ionophores in mammalian cells. TrendsBiochem. Sci. 12, 20-23.
LEE, A. S., BELL, J. & TING, J. (1984). Biochemical characterisationof the 94- and 78-kilodalton glucose regulated proteins in hamsterfibroblasts. J. biol Chem. 259, 4616-4621.
LAEMMLI, U. K. (1970). Cleavage of structural proteins during theassembly of the head of bacteriophage T4. Nature, Land. 227,680-685.
MACER, D. R. J. (1988). Identification and analysis of proteins of thereticuloplasm. Ph.D. thesis, University of Cambridge.
MACLENNAN, D. H. (1970). Purification and properties of anadenosine triphosphatase from sarcoplasmic reticulum. J. biol.Chem. 245, 4508-4518.
MACLENNAN, D. H. & HOLLAND, P. C. (1975). Calcium transport insarcoplasmic reticulum. A. Rev Biophys Bioengng. 4, 377—404.
MARUYAMA, K., MIKAWA, T. & EBASHI, S. (1984). Detection of
calcium binding protein by 45Ca autoradiography on nitrocellulosemembranes after SDS clectrophoresis. J. Biochem. 95, 511-519.
MATSUDAIRA, P. (1987). Sequence from picomole quantities ofproteins electroblotted onto polyvinyhdcne fluoride membranes. .7.biol. Chem. 262, 10035-10038."
MAWER, A., TANAKA, M., OZAWA, T. & FLEISCHER, S. (1985).
Purification and crystallisation of the calcium binding protein ofthe sarcoplasmic reticulum from skeletal muscle. Proc. natn Acad.Sci. U.SA. 82, 4036-4040.
MOORE, P. B. & KLAUS-FREEDMAN, N. (1983). Hepatic microsomalCa2+ -dependent ATPase. Biochem J 214, 69-75.
MUNRO, S. & PELHAM, H. R. B. (1987). A C-terminal signalprevents the secretion of luminal ER proteins. Cell 48, 899-907.
O'FARRELL, P. H. (1975). High resolution two-dimensionalelectrophoresis of proteins. J . biol. Chem. 250, 4007-4021.
SOMLYO, A. P. (1984). Cellular site of calcium regulation. Nature,Land. 309, 516-517.
SPAT, A., LUKACS, G. L., EBERHARDT, I., KIESEL, L. &
RUNNEBAUM, B. (1987). Binding of inositol phosphates andinduction of Ca2+ release from pituitary microsomal fractions.Biochem. J. 244, 493-496.
STREB, H., IRVINE, R. F., BERRIDGE, M. J. & SCHULZ, I. (1983).
Calcium-binding reticuloplasmins 69

Release of calcium from a non-mitochondrial store in pancreaticacinar cells by inositol-l,4,5,-triphosphate. Nature, Land. 306,67-69.
TOWBIN, H., STAEHLIN, T. & GORDON, J. (1979). Electrophoretic
transfer of proteins from polyacrylamidc gels to nitrocellulosesheets; procedure and some applications. Proc. naln. Acad. Sci.L'.SA. 76, 4350-4354.
TRUMP, B. F., BEREGESKY, 1. K. & OSORNIS-VARGAS, A. R. (1981).
In Cell Death m Biolog\' and Pathology (ed. I. D. Bowen & R. A.Lockshin), pp. 209-242. London, New York: Chapman & Hall.
WELCH, W. J., GARRELLS, J. 1., THOMAS, G. P., LIN, J. J. C. &
FERAMISCO, J. R. (1983). Biochemical characterisation of themammalian stress proteins and identification of two stresa proteinsas glucose- and calcium-ionophore regulated proteins, jf. biol.Chew. 256, 5309-5312.
WHITE, J. G., AMOS, \V. B. & FORDHAM, M. (1987). An evaluation
of confocal versus conventional imaging of biological structures byfluorescence light microscopy. J . Cell Biol. 105, 41-48.
(Received 14 April I9SS - Accepted 2 June I9SS)
Note added in proof
45Ca overlay experiments using nitrocellulose filterswere carried out in 25mM-Hepes, pH7-2, 100mM-KCl,10mM-MgCl2, using the procedures described by Kochetal. (1986a).
70 D. R. J. Macer and G. L. E. Koch







![Calcium [Ca2+]i very low ~50-100 nM –Many calcium binding proteins = high buffering capacity Divalent cation forms ionic bridges –Glutamic acid –Aspartic](https://img.pdfslide.us/doc/110x75/5697bf921a28abf838c8efef/calcium-ca2i-very-low-50-100-nm-many-calcium-binding-proteins-high.jpg)





















