Embed Size (px)
Citation preview

Copyright © 2005 by the Genetics Society of AmericaDOI: 10.1534/genetics.104.027177
Identification of a Novel Point Mutation of Mouse Proto-Oncogene c-kitThrough N-Ethyl-N-nitrosourea Mutagenesis
Hai-Bin Ruan,* Nian Zhang*,† and Xiang Gao*,‡,1
*Model Animal Research Center, State Key Laboratory of Pharmaceutical Biotechnology, Nanjing University,Nanjing, China 210089, ‡Model Organism Division, E-Institute of Shanghai Unviersity, Shanghai, China
and †Van Andel Research Institute, Grand Rapids, Michigan 49503
Manuscript received February 3, 2004Accepted for publication August 17, 2004
ABSTRACTManipulation of the mouse genome has emerged as an important approach for studying gene function
and establishing human disease models. In this study, the mouse mutants were generated through N-ethyl-N-nitrosourea (ENU)-induced mutagenesis in C57BL/6J mice. The screening for dominant mutationsyielded several mice with fur color abnormalities. One of them causes a phenotype similar to that shownby dominant-white spotting (W) allele mutants. This strain was named Wads because the homozygous mutantmice are white color, anemic, deaf, and s terile. The new mutation was mapped to 42 cM on chromosomefive, where proto-oncogene c-kit resides. Sequence analysis of c-kit cDNA from Wads m/m revealed a uniqueT-to-C transition mutation that resulted in Phe-to-Ser substitution at amino acid 856 within a highlyconserved tyrosine kinase domain. Compared with other c-kit mutants, Wads may present a novel loss-of-function or hypomorphic mutation. In addition to the examination of adult phenotypes in hearingloss, anemia, and mast cell deficiency, we also detected some early developmental defects during germcell differentiation in the testis and ovary of neonatal Wads m/m mice. Therefore, the Wads mutant mayserve as a new disease model of human piebaldism, anemia, deafness, sterility, and mast cell diseases.
N-ETHYL-N-NITROSOUREA (ENU)-induced muta- Human c-kit mutations were also identified in patientswith piebaldism, mastocytosis, gastrointestinal stromal tu-genesis has become a powerful tool for the study
of gene functions and generation of human disease mors (GISTs), acute myeloid leukemia, and germ celltumors (see online Mendelian Inheritance in Man,models recently (Nolan et al. 2000; Hrabe de Angelis
et al. 2000; Herron et al. 2002). The growing mouse OMIM, http://www.ncbi.nlm.nih.gov/entrez/dispomim.cgi?id�164920). Encoded by c-kit, KIT is a type III receptormutant archives provide a rich resource for identifying
the novel disease-related genes, deciphering pathogenic tyrosine kinase that binds to stem cell factor (SCF),which is the product of the mouse Sl locus (Wittermechanisms, and developing new therapies and new drugs
(Balling 2001; Brown and Hardisty 2003). Recently, 1990). Ligand binding activates KIT through dimeri-zation and autophosphorylation (Heldin 1995). Thewe established �40 lines of mutant mice by screening
the ENU-induced dominant mutants in C57BL/6J mice phophorylated KIT then further activates downstreampathways in a variety of cell types (Price et al. 1998;(He et al. 2003). Among them, 11 lines displayed an inter-
esting white spotted coat. One of these lines, Wads, dis- Timokhina et al. 1998; Linnekin 1999; Hou et al. 2000).Unfortunately, mechanisms by which the different KITplayed similar phenotypes to the mouse W/c-kit strain.
Mutations at the mouse W/c-kit locus on chromosome mutations cause various diseases are largely unknown.In this study, we reported the preliminary phenotypicfive can lead to pleiotropic developmental defects, in-
cluding sterility, coat color abnormalities, severe macro- analysis of the Wads mouse, including hearing ability,hematopoiesis, mast cell development, and germ cell dif-cytic anemia, loss of interstitial cells of Cajal (ICC), and
mast cell deficiency (Geissler et al. 1981; Chabot et al. ferentiation. The Wads mutation was also mapped andcloned. The novel point mutation at nucleotide 2567 of1988; Huizinga et al. 1995; Tsujimura 1996). A total of
76 mutant alleles at this locus have been accumulated in the c-kit cDNA results in a substitution of Phe to Ser atKIT amino acid 856 (F856S).mouse up to 1997 (see the Mouse Genome Database,
http://www.informatics.jax.org/searches/mlc.cgi?10603).
MATERIALS AND METHODS
Animals: C57BL/6J mice were obtained from Shanghai Lab-Sequence data from this article have been deposited with the EMBL/oratory Animal Center (Shanghai, China). CAST/Ei mice wereGenBank Data Libraries under accession nos. AY536430 and AY536431.obtained from The Jackson Laboratory (Bar Harbor, ME). Mice1Corresponding author: Model Animal Research Center, Nanjing Uni-were maintained under specific pathogen-free environment.versity, 308 Xuefu Rd., Pukou District, Nanjing 210089, People’s Re-
public of China. E-mail: [email protected] Animal welfare and experimental procedures were carried out
Genetics 169: 819–831 (February 2005)
Dow
nloaded from https://academ
ic.oup.com/genetics/article/169/2/819/6060306 by guest on 13 February 2022

820 H.-B. Ruan, N. Zhang and X. Gao
strictly in accordance with the care and use of laboratory covered the full length were then amplified by PCR with LA-Taq polymerase (TaKaRa, Shiga, Japan) using the followinganimals (National Research Council, 1996) and the related
ethical regulations of Nanjing University. The G1 generation primers: kit1-FOR, 5�-TCAGAGTCTAGCGCAGCCAC-3�; kit1-REV, 5�-GCCTCGTATTCAACAACCAA-3�; kit2-FOR, 5�-TGTAdominant mutant mouse (G1M485) of C57BL/6J background
was generated by ENU mutagenesis (He et al. 2003). This male ACCGATGGAGAAAACG-3�; kit2-REV, 5�-TAAACGAGTCACGCTTCCTT-3�; kit3-FOR, 5�-GCCCTAATGTCGGAACTGAA-3�;mouse was mated to C57BL/6J females for confirming the
inheritance test. Homozygous mutants (Wadsm/m) were gener- and kit3-REV, 5�-GTTTCTGCTCAGGCATCTTC-3�. The threefragments were then subcloned into TA cloning vector pMDated by intercross of the heterozygous mutants (Wadsm/�).
Histological analysis: Skin, testes, or ovaries were dissected 18-T (TaKaRa) and sequenced (Bioasia, Shanghai, China). Allsequences were confirmed by sequencing from both directions.and fixed in Bouin’s fixative. They were dehydrated in rising
concentrations of ethanol, embedded in paraffin, and sec- Single-strand conformation polymorphism: PCR-single-strandconformation polymorphism (SSCP) analysis was performedtioned (7 �m). The sections of testis and ovary were stained
with hematoxylin and eosin according to standard procedures. as described (Strippoli et al. 2001). Briefly, 200 bp of DNAfragment covering the Wads point mutation was amplified byToluidine blue in sodium chloride was used to stain mast cells
in skin tissues. PCR using primers of 5�-GACTGCCCGTGAAGTGGAT-3� and5�-CTTCCAGAGAGGTGGCAAAT-3�. The PCR product wasPeripheral blood analysis: Peripheral blood was collected
using a capillary tube from suborbital veins of 5- to 6-week- mixed with 10� alkali denaturating buffer (NaOH, 500 mm;EDTA, 10 mm) and heated to 94� for 10 min and then quicklyold mice. Blood cell analysis was performed with a Coulter
(Hialeah, FL) STKS hematology analyzer. The parameters cooled on ice for 2 min. Samples were then mixed with loadingbuffer, loaded on an 8% polyacrylamide gel (39:1 acrylamidemeasured included white blood cell counts (WBC); red blood
cell counts (RBC); hemoglobin concentration (HGB); mean to bis-acrylamide; TAE buffer; 0.2% TEMED; 0.05% ammo-nium persulfate 10%), and run at 90 V for �1.5 hr. The gelcorpuscular volume (MCV, the average volume of individual
RBC); mean corpuscular hemoglobin (MCH, the average was stained with 5�g/ml ethidium bromide.weight of hemoglobin in a red blood cell); mean corpuscularhemoglobin concentration (MCHC, the ratio of MCH to MCV);platelet counts (PLT); and mean platelet volume (MPV, the RESULTSaverage volume of individual platelets). The P-values were
Pigmentation defects of Wads mutants: The founderevaluated using an unpaired two-tailed t-test using Prism soft-(G1M485) of the Wads strain had a symmetrical clusterware (GraphPad Software, San Diego) between the blood pa-
rameters of male and female mice and between wild-type mice of white coat on the midline of the abdomen and smalland heterozygous or homozygous mice. white spots on the back (Figure 1, A and B). The coat
Auditory brainstem response test: The auditory brainstem of most of the tail as well as the distal part of the legsresponse (ABR) test was performed with a PowerLab systemalso displayed white color. Half of the progenies (112 of(A&D, Castle Hill, Australia). Mice were anesthetized with212) from the founder showed a similar pigmentationAvertin and the stimulating electrode was embedded in the
mastoid of the right ear, the recording electrode subcutane- abnormality, indicating a single-gene mutation in theously in the vertex, and the earth electrode in the mastoid of Wads strain.the left ear. ABRs were recorded under the sound stimuli of Interestingly, homozygous mutants (Wadsm/m) showed40, 50, and 70 dB sound pressure levels (Zheng et al. 1999).
all white skin and hairs but black eyes (Figure 1C). Be-Cochlear histology and immunohistochemistry: For coch-cause the melanocytes are derived from neural crestlear histology, the mice were killed and temporal bones were
removed from the skull and then fixed in 4% paraformalde- cells (NCCs) whereas retinal pigment epithelium (RPG)hyde at 4� overnight. Tissues were then immersed in decalcify- arises from neural tube epithelium, the lack of func-ing solution (4% EDTA in PBS) for 1 week. The paraffin sec- tional melanocytes suggested that the migration and/ortions were prepared and stained with hematoxylin and eosin.
differentiation of the NCCs were impaired in Wadsm/mFor immunofluorescence, the paraffin sections were dehy-
mice (Zhao et al. 1997).drated and permeabilized with 20 �g/ml proteinase K. Block-The mutation localized at W/c-kit locus: Several signal-ing was carried out with 10% goat serum in PBS. Primary rabbit
antibodies to connexin 26 and 30 (Zymed, San Francisco) were ing pathways, especially the SCF/KIT pathway, havein 2% goat serum/PBS at a dilution of 1:50. A FITC-conjugated been reported to participate in proliferation and differ-goat anti-rabbit secondary antibody (Sigma, St. Louis) was used entiation during melanoblast development (Bennettto detect connexins and the fluorescence was observed under
and Lamoreux 2003; Manova and Bachvarova 1991).a Leica confocal microscope.Therefore, we examined whether the c-kit gene locus,Mutation mapping and cloning: Wads m/� mice of C57BL6/J
were mated to CAST/Ei mice to generate F1. F1 females with 42 cM of chromosome 5, links to the Wads mutation.white spots were backcrossed to CAST/Ei male mice to gener- With the backcrossing protocol for mapping using theate N2. The tail DNA samples of N2 mice were extracted and CAST/Ei strain (Reeves and D’Eustachio 1997), wePCR amplified with D5Mit356 and D5Mit359 microsatellite analyzed the linkage between pigmentation abnormalitymarkers of the Whitehead/MIT database (http://www-genome.
of 60 N2 mice and two microsatellite markers, D5Mit356wi.mit.edu). PCR products were separated by electrophoresisof 41 cM and D5Mit359, of 44 cM on chromosome 5. Wewith 4% agarose II (BioBasic, Markham, ON, Canada) gel.
To examine if the mutation resides in the c-kit gene, total found that the ratio of recombinant/nonrecombinantRNA samples were prepared from the skin of adult C57BL/ phase of D5Mit356 was 2/58, while that of D5Mit3596J and Wads m/m mice using the Trizol kit following the manu- was 5/55. This result strongly suggested that the Wadsfacturer’s instructions (Shenergy, Shanghai, China). Full-
mutation was linked to the W/c-kit locus.length cDNA of c-kit was synthesized with AMV reverse tran-Molecular analysis of the mutation: To examine whetherscriptase (Promega, Madison, WI). Due to the large size of
c-kit cDNA (2925 bp), three overlapping cDNA fragments that the c-kit gene was mutated in Wads mice, wild-type and
Dow
nloaded from https://academ
ic.oup.com/genetics/article/169/2/819/6060306 by guest on 13 February 2022

821Novel Mutation of the Mouse c-kit Gene
Figure 1.—Defects of Wadsmice coat color and mast cells.(A) A Wadsm/� heterozygote withwhite spots on the abdomen.(B) Dorsal view of a Wadsm/� het-erozygote. (C) Black-eyed whiteWads m/m homozygote. (D) Wild-type,(E)Wadsm/�, and(F)Wadsm/m
skin sections were stained withToluidine blue. Black arrowsidentify mast cells (purple) andwhite arrows identify hair folli-cles at telogen phase (blue).
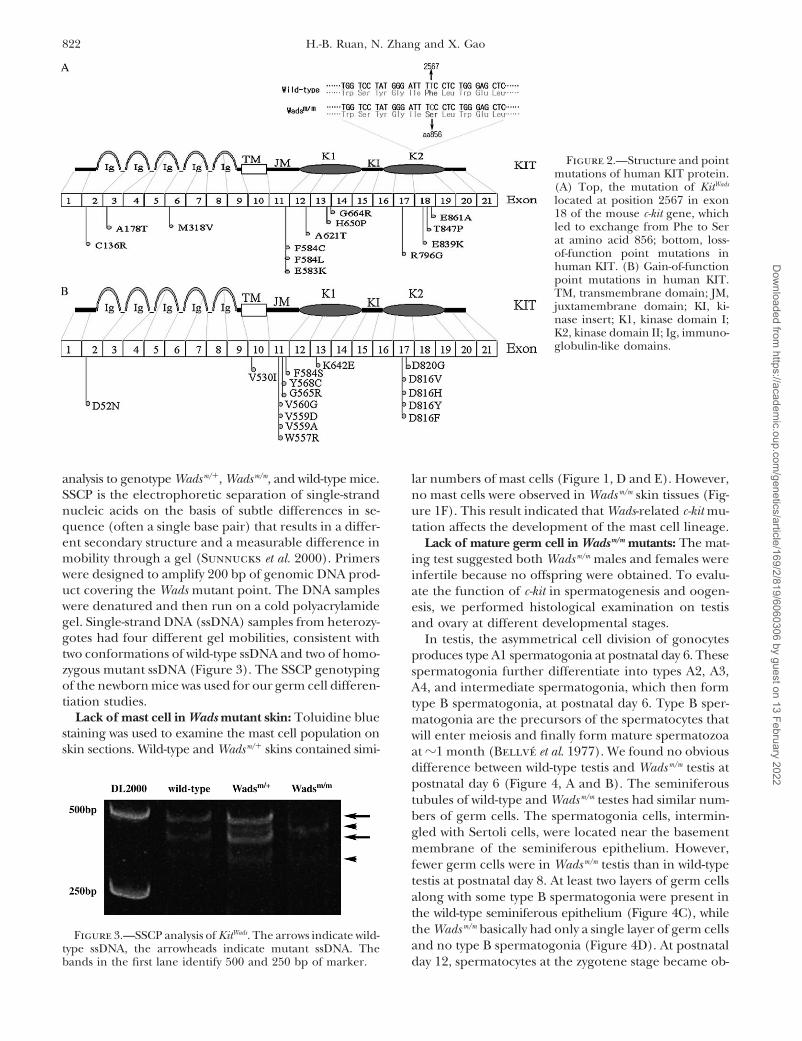
Wadsm/m c-kit cDNA was cloned and sequenced. The se- Ensembl Genome Browser (ENSMUSG00000005672).However, our sequence data differed from the Mus mus-quence data revealed a T-to-C missense transition at
nucleotide 2567 in Wadsm/m coding sequence that re- culus c-kit cDNA sequence listed in GenBank (NM_021099),with a synonymous mutation and two missense muta-sulted in a Phe(F) to Ser(S) change at amino acid 856
(Figure 2A). This mutation position located in the sec- tions. We believe that our sequence data were morereliable, according to the protein sequence alignmentond protein tyrosine kinase (PTK) domain and the Phe
is conserved among all the mammalian KIT proteins. with other mammalian KITs (data not shown).Because the coat abnormality is not available at suchCombined with the phenotypic profiles, Wads may rep-
resent a loss-of-function or hypomorphic c-kit mutation an early stage of animal development, development ofa reliable genotyping method for the Wads mutant is(Reith et al. 1990).
It is also worth pointing out that our wild-type C57BL/ necessary for studying the embryonic and neonatal de-fects. For this reason, we developed a PCR-based SSCP6J cDNA sequence of c-kit was consistent with that of the
Dow
nloaded from https://academ
ic.oup.com/genetics/article/169/2/819/6060306 by guest on 13 February 2022
822 H.-B. Ruan, N. Zhang and X. Gao
Figure 2.—Structure and pointmutations of human KIT protein.(A) Top, the mutation of KitWads
located at position 2567 in exon18 of the mouse c-kit gene, whichled to exchange from Phe to Serat amino acid 856; bottom, loss-of-function point mutations inhuman KIT. (B) Gain-of-functionpoint mutations in human KIT.TM, transmembrane domain; JM,juxtamembrane domain; KI, ki-nase insert; K1, kinase domain I;K2, kinase domain II; Ig, immuno-globulin-like domains.
analysis to genotype Wadsm/�, Wadsm/m, and wild-type mice. lar numbers of mast cells (Figure 1, D and E). However,SSCP is the electrophoretic separation of single-strand no mast cells were observed in Wadsm/m skin tissues (Fig-nucleic acids on the basis of subtle differences in se- ure 1F). This result indicated that Wads-related c-kit mu-quence (often a single base pair) that results in a differ- tation affects the development of the mast cell lineage.ent secondary structure and a measurable difference in Lack of mature germ cell in Wadsm/m mutants: The mat-mobility through a gel (Sunnucks et al. 2000). Primers ing test suggested both Wadsm/m males and females werewere designed to amplify 200 bp of genomic DNA prod- infertile because no offspring were obtained. To evalu-uct covering the Wads mutant point. The DNA samples ate the function of c-kit in spermatogenesis and oogen-were denatured and then run on a cold polyacrylamide esis, we performed histological examination on testisgel. Single-strand DNA (ssDNA) samples from heterozy- and ovary at different developmental stages.gotes had four different gel mobilities, consistent with In testis, the asymmetrical cell division of gonocytestwo conformations of wild-type ssDNA and two of homo- produces type A1 spermatogonia at postnatal day 6. Thesezygous mutant ssDNA (Figure 3). The SSCP genotyping spermatogonia further differentiate into types A2, A3,of the newborn mice was used for our germ cell differen- A4, and intermediate spermatogonia, which then formtiation studies. type B spermatogonia, at postnatal day 6. Type B sper-
Lack of mast cell in Wads mutant skin: Toluidine blue matogonia are the precursors of the spermatocytes thatstaining was used to examine the mast cell population on will enter meiosis and finally form mature spermatozoaskin sections. Wild-type and Wadsm/� skins contained simi- at �1 month (Bellve et al. 1977). We found no obvious
difference between wild-type testis and Wadsm/m testis atpostnatal day 6 (Figure 4, A and B). The seminiferoustubules of wild-type and Wadsm/m testes had similar num-bers of germ cells. The spermatogonia cells, intermin-gled with Sertoli cells, were located near the basementmembrane of the seminiferous epithelium. However,fewer germ cells were in Wadsm/m testis than in wild-typetestis at postnatal day 8. At least two layers of germ cellsalong with some type B spermatogonia were present inthe wild-type seminiferous epithelium (Figure 4C), whilethe Wadsm/m basically had only a single layer of germ cellsFigure 3.—SSCP analysis of KitWads. The arrows indicate wild-and no type B spermatogonia (Figure 4D). At postnataltype ssDNA, the arrowheads indicate mutant ssDNA. The
bands in the first lane identify 500 and 250 bp of marker. day 12, spermatocytes at the zygotene stage became ob-
Dow
nloaded from https://academ
ic.oup.com/genetics/article/169/2/819/6060306 by guest on 13 February 2022

823Novel Mutation of the Mouse c-kit Gene
Figure 4.—Spermatogenesis defects in Wads mutants. Sections of wild-type testis of postnatal day 6 (A), day 8 (C), day 12 (E),and 6 weeks (G) and of Wads m/m testis of day 6 (B), day 8 (D), day 12 (F), 6 weeks (H), and 10 weeks (I) are shown. The typeA and B spermatogonia are indicated by white and black arrows. The Sertoli cells and primary spermatocytes are indicated bywhite and black arrowheads. White stars indicate clusters of cells in the center of abnormal seminiferous tubules and the interstitialspace is enlarged by hyperplastic Ledig cells (black stars). Gross morphology of Bouin’s fixative-fixed adult Wads m/� and Wads m/m
testis is shown (J).
vious in the wild-type seminiferous tubules. In contrast, resembled that of the postnatal day 6 mice. At 6 and10 weeks, only a few germ cells remained at the basalno germ cells entered meiosis in the Wadsm/m testis (Fig-
ure 4, E and F). Overall, the morphology of the seminif- membrane of the seminiferous tubules of the Wadsm/m
testis (Figure 4, H and I). In the adult Wadsm/m testiserous epithelium of the Wadsm/m mice at this stage still
Dow
nloaded from https://academ
ic.oup.com/genetics/article/169/2/819/6060306 by guest on 13 February 2022

824 H.-B. Ruan, N. Zhang and X. Gao
Figure 4.—Continued.
we also observed clusters of cells in the center of the At postnatal day 11, wild-type ovaries showed many de-veloping follicles in the cortex (Figure 5C), comparedseminiferous tubules. They had germ cell characteristics
and it was possible that the disruption of spermatogene- with no follicle in Wadsm/m ovaries (Figure 5D). Only gran-ulose cells remained in adult Wadsm/m ovaries (Figuresis in the Wadsm/m mice caused shedding of germ cells
into the lumen. Moreover, the interstitial space in adult 5F) and the gross size of the Wadsm/m ovary was reduced.Effect of the Wads mutation on peripheral blood:Wadsm/m testis was filled out with overgrowth of Leydig
cells (Figure 4, H and I). This increase of Leydig cell Blood cell analysis of peripheral blood was performedto examine the possible defects in Wads mutant micenumbers was not caused by the reduction of the size of
the seminiferous tubules but by enhanced mitotic activ- because c-kit is expressed and functions in hemopoieticprogenitor cells (Ogawa et al. 1991). As presented inity of Leydig cells (Kissel et al. 2000). And our BrdU as-
say, which showed that the Leydig cells in Wadsm/m testis Table 1, the total values of each parameter of Wadsm/�
and Wadsm/m were compared to those of wild-type mice.were stained positive, was in agreement with this hypoth-esis (data not shown). Finally, the gross size of the adult No differences were seen between wild-type and Wadsm/�
mice, except Wadsm/� mice displayed slight elevation ofWadsm/m testis is much smaller than that of wild types(Figure 4J). MPV (P � 0.5). Interestingly, the RBC counts of Wadsm/m
mice were significant reduced (P � 0.5), whereas theIn the ovary, Kit expression is observed in oocytes fromthe time of birth until ovulation (Horie et al. 1991). Inter- MCV of Wadsm/m was significantly higher (P � 0.0001)
compared with that of wild-type and Wadsm/� mice. Theseaction of Kit with KL/MGF expressed by granulosa cellsis essential for follicle development (Kuroda et al. 1988). results indicated that Wadsm/m mice were suffering from
macrocytic anemia. They were also consistent with ear-In newborn wild-type mouse ovaries, hematoxylin andeosin staining showed many primordial follicles (Figure lier reports that mutations of c-kit or kitl (encoding KIT
ligand SCF) displayed a similar hemopoietic disorder5A). However, no primordial follicles or signal nakedgerm cells were present (Figure 5B) in Wadsm/m ovaries. (Piao and Bernstein 1996). The platelet counts and
Dow
nloaded from https://academ
ic.oup.com/genetics/article/169/2/819/6060306 by guest on 13 February 2022

825Novel Mutation of the Mouse c-kit Gene
Figure 5.—Histological analysis of postnatal ovaries in Wads mice. Sections of ovaries of postnatal day 0 (A and B), day 11 (Cand D), and adult stage (E and F) are shown. (A, C, and E) Wild-type ovaries. (B, D, and F) Wads m/m ovaries. The white arrowsindicate follicles.
mean platelet volume of Wadsm/m mice were also signifi- vascularis of homozygous KitW mutant mice, resultingin endocochlear degeneration, endocochlear potentialcantly elevated compared with that of wild type and
Wadsm/� (P � 0.05 and P � 0.01, respectively). All blood (EP) disappearance, and hearing impairment (Cableet al. 1994, 1995). To examine the hearing ability ofparameters showed no significant difference between
males and females within the same genotype. Wads mice, we carried out an ABR test. Both wild-typeand Wadsm/� mice showed normal waves responding toHearing loss in Wadsm/m mutants: The intermediate
cells that derive from melanoblasts were missing in stria 40, 50, and 70 dB sound pressure levels of stimuli (Figure
Dow
nloaded from https://academ
ic.oup.com/genetics/article/169/2/819/6060306 by guest on 13 February 2022

826 H.-B. Ruan, N. Zhang and X. Gao
6A; data of the Wadsm/� ABR pattern were not shown).However, no ABR response waves were detected inWadsm/m mutant mice 1 month after birth (Figure 6B),indicating that Wads-related c-kit mutation led to hear-ing loss. In wild-type cochlear sections, the stria vascu-laris (SV) was composed of three layers of cells. Theywere marginal cells facing the endolymphatic space,basal cells facing the spiral ligament, and intermediatecells in the middle (Figure 6, C and D). There werealso abundant blood capillaries in the SV. However, theSV of the Wadsm/m cochlea was much thinner than thatof wild type because of the loss of intermediate cellsand defects of blood capillaries (Figure 6, E–H). Mean-while, the organ of Corti and hair cells were degener-ated in the Wadsm/m cochlea (Figure 6, E and G). It wasbelieved that these phenomena were the later pathologi-cal changes of the loss of intermediate cells and EP(Hoshino et al. 1999). Moreover, the SV became moredisorganized and the blood capillaries were depleted inthe SV in the 1-year-old Wadsm/m cochlear sections (Fig-ure 6H).
The gap junction system in the cochlea is critical forion cycling, which is the underlying mechanism of thegeneration of EP. There are widespread gap junctionsbetween basal cells and between basal cells and interme-diate cells. These cells are coupled together as a syncy-tium allowing exchange of intracellular contents suchas K�. Connexins are gap junction proteins that areexpressed in the cochlea (Lautermann et al. 1998; Fig-ure 6, I and K). In an immunohistological study ofWadsm/m compared with the wild-type mice, the connexin26 and connexin 30 expression is disorganized and dis-continuous in SV of Wadsm/m mice, suggesting the gapjunctions in the base of the SV were detrimentally dis-rupted (Figure 6, J and L).
DISCUSSION
In this report we described the phenotypes and ge-nome alternation of a new mutant strain Wads fromENU mutagenesis. Genetic mapping indicated that Wadswas a novel allele of the W/c-kit locus, with a missensepoint mutation of c-kit proto-oncogene (Phe856Ser) in theconservative PTK domain. This mutation led to whitespotting in heterozygotes and black-eyed white color,macrocytic anemia, mast cell deficiency, deafness, andsterility in homozygotes.
The c-kit gene spans �70 kb of DNA and includes 21exons (Vandenbark et al. 1992). The longest transcriptis 5230 bp. Notably, c-kit is among those genes with thehighest spontaneous mutations. Various KIT mutationshave been identified in human, mouse, rat, dog, andpig, etc. (Tsujimura et al. 1991; Pielberg et al. 2002;Zemke et al. 2002). While the actual molecular mecha-nism of this high mutation frequency is still not clear,it may be related to the easy accessibility of the mutagen
TA
BL
E1
Per
iphe
ral
bloo
dce
llan
alys
is
Wild
type
Wad
sm/ �
Wad
sm/m
Mal
eFe
mal
eT
otal
Mal
eFe
mal
eT
otal
Mal
eFe
mal
eT
otal
(n�
4)(n
�4)
(n�
8)(n
�4)
(n�
4)(n
�8)
(n�
4)(n
�4)
(n�
8)
WB
C(�
109 /
L)
7.6
3.
66.
2
1.4
6.9
2.
65.
8
3.0
5.9
2.
65.
8
2.6
7.4
3.
48.
2
3.8
7.8
3.
4R
BC
(�10
9 /L
)7.
3
1.1
7.3
1.
17.
3
1.0
8.4
1.
06.
8
2.5
7.6
2.
06.
4
0.6
5.2
1.
25.
8
1.1*
HG
B(g
/L)
120.
8
15.4
129.
3
25.8
125.
0
20.2
137.
0
11.5
119.
2
39.2
128.
1
28.4
148.
2
27.5
108.
2
40.4
128.
2
38.5
MC
V(fl
)48
.6
1.3
46.2
2.
647
.4
2.3
46.7
1.
246
.4
0.8
46.5
1.
055
.4
4.0
60.8
5.
358
.1
5.2*
**M
CH
(Pg)
16.7
0.
417
.2
0.3
16.9
0.
416
.4
0.6
17.7
2.
317
.0
1.7
16.7
4.
519
.9
1.8
18.3
3.
6M
CH
C(g
/L)
343.
0
5.0
372.
5
21.8
357.
8
21.5
350.
5
10.2
380.
8
52.8
365.
6
38.7
301.
8
83.8
327.
5
22.0
314.
6
58.4
PLT
(�10
9 /L
)71
9.5
26
8.4
755.
5
420.
473
7.5
32
7.1
776.
0
369.
450
4.5
45
4.5
640.
3
410.
099
7.0
28
3.8
1082
.3
124.
010
39.6
20
7.8*
MPV
(fl)
5.1
0.
35.
0
0.2
5.0
0.
25.
8
0.8
5.5
0.
35.
6
0.5*
5.7
0.
65.
6
0.1
5.6
0.
4**
WB
C,
wh
ite
bloo
dce
llco
unts
;R
BC
,re
dbl
ood
cell
coun
ts;
HG
B,
hem
oglo
bin
con
cen
trat
ion
;M
CV
,m
ean
corp
uscu
lar
volu
me;
MC
H,
mea
nco
rpus
cula
rh
emog
lobi
n;
MC
HC
,mea
nco
rpus
cula
rh
emog
lobi
nco
nce
ntr
atio
n;P
LT
,pla
tele
tco
unts
;MPV
,mea
npl
atel
etvo
lum
e.N
odi
ffer
ence
was
seen
betw
een
mal
esan
dfe
mal
esw
ith
inth
esa
me
gen
otyp
e.T
he
aver
age
valu
esof
the
thre
em
ice
wer
eev
alua
ted
usin
gan
unpa
ired
two-
taile
dt-t
est.
*P
0.05
;**
P
0.01
;**
*P�
0.00
01.
to this chromosomal locus as well as the easy recognition
Dow
nloaded from https://academ
ic.oup.com/genetics/article/169/2/819/6060306 by guest on 13 February 2022

827Novel Mutation of the Mouse c-kit Gene
Figure 6.—ABR test and co-chlear histology of B6 and Wadsmutant mice. (A and B) NormalABR waves induced by differentsound pressure level stimuli in awild-type (A) and Wads m/m (B)mouse. (C and E) Cochlear sec-tion of a 2-month-old wild-type(C) and Wads m/m (E) mouse. (G)Section from a 10-month-oldWads m/m mouse. (D, F, and H)Magnification of the stria vascu-laris regions of C, E, and G. Mar-ginal cells (white arrows), basalcells (black arrows), intermediatecells (black arrowheads), andblood capillaries (white arrow-heads) are indicated. (I and H)Stria vascularis immunostainingwith connexin 26 on wild type andWads m/m. (K and L) Stria vascularisimmunostaining with connexin 30on wild type and Wads m/m. Thearrows indicate the margin of theSV. The arrowheads indicate thegap junction between basal cellsand intermediate cells. The starspoint out the disruptions of gapjunction in Wads m/m SV.
of dominant coat abnormality. It is possible that the c-kit Two types of c-kit mutations have been reported inhumans. Loss-of-function mutations, which often takegene is transcriptionally active in dividing germ cells;
therefore the locus is in a constant “open” status. How- place at the tyrosine kinase domains and immuno-globulin-like loops, may cause a deficiency/defect ofever, by examining the expression file of testis, c-kit is
certainly not among the highest-expressing genes, al- melanocyte, mast cell, germ cell, and hematogenic cells(Figure 2A; Spritz 1994; Fleischman et al. 1996; OMIMthough the testis contains mixed types of cells (Su et al.
2002). {*164920}). In contrast, gain-of-function mutations, which
Dow
nloaded from https://academ
ic.oup.com/genetics/article/169/2/819/6060306 by guest on 13 February 2022

828 H.-B. Ruan, N. Zhang and X. Gao
Figure 6.—Continued.
are located in the cytoplasmic juxtamembrane domain KitW-19H) or point mutation (e.g., KitW-37J, KitW-42J) are homo-zygous lethal, while mutations with residual kinase activ-or catalytic domain, were identified as the cause of mas-
tocytosis, acute myeloid leukemia, gastrointestinal stro- ity (e.g., KitW-V, KitW-57J) are homozygous viable (Bern-stein et al. 1991). From these studies we speculatedmal tumors, and germ cell tumors (Figure 2B; Kita-
yama et al. 1996; Tsujimura 1996; Hirota et al. 1998; that KIT of Wadsm/m maintains residual kinase activityalthough further proof is needed. Interestingly, theTian et al. 1999; OMIM {*164920}). The phenotypes of
Wadm/m suggested that the F856S substitution (equal to Wadsm/m mice displayed noticeable reduced prenatal orneonatal viability. Among a total of 207 mice from aF858S in human KIT) is a loss-of-function or hypomor-
phic mutation (Figure 2A). heterozygous intercrossing breed, only 12 of them werehomozygotes, 4 of which died within 10 weeks afterPrevious studies showed that the level of KIT kinase
activity and the severity of the phenotypic expression birth. The proportion of viable homozygous mutantmice was 4.35% and significantly below the expectedfor each W/c-kit allele correlated with the type of muta-
tion. Mutations that abolish activity by deletion (e.g., KitW, 25%. The cause of this lethality is still unknown.
Dow
nloaded from https://academ
ic.oup.com/genetics/article/169/2/819/6060306 by guest on 13 February 2022

829Novel Mutation of the Mouse c-kit Gene
Figure 6.—Continued.
Among all the known c-kit mutants, only KitW-V and whereas the KitWads mutation localized in the kinase do-main II. There are also some phenotypic differencesour KitWads show all the defects that have been reported
in c-kit mutations, including white color, mast cell loss, between these two strains. For instance, The KitW-V het-erozygotes had a slight macrocytic anemia but the KitWadsanemia, hearing loss, and sterility. Nevertheless, KitW-V
is a missense Thr-to-Met point mutation in position 660 heterozygotes did not suffer from anemia (Russell1949). It may reflect that residual kinase activity in KitWads(T660M) of the kinase domain I of the KIT protein,
Dow
nloaded from https://academ
ic.oup.com/genetics/article/169/2/819/6060306 by guest on 13 February 2022

830 H.-B. Ruan, N. Zhang and X. Gao
is more than that in KitW-V mice. Alternatively, the pheno- us a new powerful animal model to comprehensivelyreveal the function of c-kit and most importantly to fig-type variation may result from different mutant sites
in the kinase domain, which might subsequently affect ure out the complex downstream signaling pathwaysand targets of c-kit in related different systems.different downstream signaling pathways. Similar sce-
narios have been reported, where blockage of phospha- We thank Xingxing Gu, Fang He, Zixin Wang, and Haibo Sha fortidylinositol 3�-kinase (PI3K) signaling from KIT did not technical help. This work is supported by the National Natural Science
Fund of China (30300425), the National Gongguan Project of Chinaaffect other KIT downstream responses (Blume-Jensen(2001BA710B), the Joint Research Fund for Overseas Chinese Younget al. 2000; Kissel et al. 2000).Scholars (30228008), and EISU(E03003).The partial viability of the KitWads homozygote really
facilitated the functional analysis of the c-kit gene on latedevelopmental events such as gametogenesis. Previous
LITERATURE CITEDstudies suggested that the migration and/or prolifera-tion of primordial germ cells (PGCs) were impaired in Balling, R., 2001 ENU mutagenesis: analyzing gene function in
mice. Annu. Rev. Genomics Hum. Genet. 2: 463–492.the KitW-V embryos. However, the testes of newborn KitWads
Bellve, A. R., J. C. Cavicchia, C. F. Millette, D. A. O’Brien, Y. M.homozygous mice showed no differences from those of Bhatnagar et al., 1977 Spermatogenic cells of the prepuberalwild-type mice up to postnatal day 6. It is possible that mouse. Isolation and morphological characterization. J. Cell Biol.
74: 68–85.the KitWads mutation affects only postnatal differentiationBennett, D. C., and M. L. Lamoreux, 2003 The color loci ofand maturation of spermatogenesis but not embryonic mice—a genetic century. Pigment. Cell. Res. 16: 333–344.
PGC development. Alternatively, the KitWads mutation did Bernstein, A., L. Forrester, A. D. Reith, P. Dubreuil and R. Rotta-pel, 1991 The murine W/c-kit and Steel loci and the control ofreduce the number of PGCs by affecting their prolifera-hematopoiesis. Semin. Hematol. 28: 138–142.tion and/or survival, but the remaining PGCs were enough Blume-Jensen, P., G. Jiang, R. Hyman, K. F. Lee, S. O’Gorman
to form a relatively normal gonad structure at birth. et al., 2000 Kit/stem cell factor receptor-induced activation ofphosphatidylinositol 3�-kinase is essential for male fertility. Nat.In contrast with testis, the ovary lacks primordial folli-Genet. 24: 157–162.cles in newborn KitWads homozygous females, and this
Brown, S. D., and R. E. Hardisty, 2003 Mutagenesis strategies fordirectly caused the female to be infertile. This suggested identifying novel loci associated with disease phenotypes. Semin.
Cell. Dev. Biol. 14: 19–24.that the c-kit mutation affected the female reproductiveCable, J., D. Huszar, R. Jaenisch and K. P. Steel, 1994 Effects ofsystem before birth. When and how the mutation reacts
mutations at the W locus (c-kit) on inner ear pigmentation andwith female primordial germ cell development remain function in the mouse. Pigment. Cell. Res. 7: 17–32.
Cable, J., I. J. Jackson and K. P. Steel, 1995 Mutations at the Wunclear.locus affect survival of neural crest-derived melanocytes in theIt is interesting to dissect out the differential responsemouse. Mech. Dev. 50: 139–150.
of the KitWads mutation in the male and the female repro- Chabot, B., D. A. Stephenson, V. M. Chapman, P. Besmer and A.Bernstein, 1988 The proto-oncogene c-kit encoding a trans-ductive systems. Transgenic study indicated that the mel-membrane tyrosine kinase receptor maps to the mouse W locus.anocyte stem cell (MSC) could survive independentlyNature. 335: 88–89.
of c-kit signaling transduction. However, MSCs migrated Fleischman, R. A., T. Gallardo and X. Mi, 1996 Mutations in theoutside of the hair follicles in a c-kit-dependent manner ligand-binding domain of the kit receptor: an uncommon site
in human piebaldism. J. Invest. Dermatol. 107: 703–706.(Kunisada et al. 1998). Hematopoietic stem cells ex-Geissler, E. N., E. C. McFarland and E. S. Russell, 1981 Analysispressed c-kit but did not depend on it (Ikuta and Weiss- of pleiotropism at the dominant white-spotting (W) locus of the
man 1992). Similarly, spermatogonia stem cells (SSCs) house mouse: a description of ten new W alleles. Genetics 97:337–361.did not express or depend on c-kit (Schrans-Stassen
He, F., Z. Wang, J. Zhao, J. Bao, J. Ding et al., 2003 Large scaleet al. 1999; Ohta et al. 2003). We suspect only the further screening of disease model through ENU mutagenesis in mice.differentiation of the SSCs requires c-kit gene function. Chin. Sci. Bull. 48: 2665–2671.
Heldin, C. H., 1995 Dimerization of cell surface receptors in signalIn male mice, the differentiation of SSCs and spermato-transduction. Cell 80: 213–223.genesis start �6 days after birth. The meiosis happens Herron, B. J., W. Lu, C. Rao, S. Liu, H. Peters et al., 2002 Efficient
much later. In contrast, the oogonia start meiosis at generation and mapping of recessive developmental mutationsusing ENU mutagenesis. Nat. Genet. 30: 185–189.around embryonic day 13.5 and arrest at the diplotene
Hirota, S., K. Isozaki, Y. Moriyama, K. Hashimoto, T. Nishidastage before birth. We hypothesize that, therefore, KIT et al., 1998 Gain-of function mutations of c-kit in human gastro-function is crucial at the time of stem cell differentiation intestinal stromal tumors. Science 279: 577–580.
Horie, K., K. Takakura, S. Taii, K. Narimoto, Y. Nada et al., 1991and/or meiosis in the reproductive system. Because ofThe expression of c-kit protein during oogenesis and early embry-the different time courses for male and female germ cellonic development. Biol. Reprod. 45: 547–552.
differentiation, the KitWads mutation displayed distinct Hoshino, T., K. Mizuta, J. Gao, S. Araki, K. Araki et al., 1999 Coch-lear findings in the white spotting (Ws) rat. Hear. Res. 140: 145–156.patterns of defects in the male and the female reproduc-
Hou, L., J. J. Panthier and H. Arnheiter, 2000 Signaling andtive systems.transcriptional regulation in the neural crest-derived melanocyte
In this study, we generated and identified a new strain lineage: interactions between KIT and MITF. Development 127:5379–5389.for the mouse c-kit mutant archive. This KitWads mouse
Hrabe de Angelis, M., H. Flaswinkel, H. Fuchs, B. Rathkolb, D.showed almost all the phenotypes associated with proto-Soewartom et al., 2000 Genome-wide, large-scale production
oncogene c-kit mutation. There were detailed differ- of mutant mice by ENU. Nat. Genet. 25: 444–447.Huizinga, J. D., L. Thuneberg, M. Kluppel, J. Malysz, H. B. Mikkel-ences between KitWads and other c-kit alleles. It provided
Dow
nloaded from https://academ
ic.oup.com/genetics/article/169/2/819/6060306 by guest on 13 February 2022

831Novel Mutation of the Mouse c-kit Gene
esen et al., 1995 W/kit gene required for interstitial cells of Cajal Reith, A. D., R. Rottapel, E. Giddens, C. Brady, L. Forresteret al., 1990 W mutant mice with mild or severe developmentaland for intestinal pacemaker activity. Nature 373: 347–349.
Ikuta, K., and I. L. Weissman, 1992 Evidence that hematopoietic defects contain distinct point mutations in the kinase domain ofthe c-kit receptor. Genes Dev. 4: 390–400.stem cells express mouse c-kit but do not depend on steel factor
for their generation. Proc. Natl. Acad. Sci. USA 89: 1502–1506. Russell, E. S., 1949 Analysis of pleiotropism at the W-locus in themouse: relationship between the effects of W and W-v substitutionLautermann, J., W.-J.F. ten Cate, P. Altenhoff, R. Grummer, O.
Traub et al., 1998 Expression of the gap-junction connexin 26 on hair pigmentation and on erythrocytes. Genetics 34: 708–723.Schrans-Stassen, B., H. Van De Kant, D. De Rooij and A. Van Pelt,and 30 in the rat cochlea. Cell Tissue Res. 294: 415–420.
Linnekin, D., 1999 Early signaling pathways activated by c-Kit in 1999 Differential expression of c-kit in mouse undifferentiatedand differentiating type A spermatogonia. Endocrinology 140:hematopoietic cells. Int. J. Biochem. Cell Biol. 31: 1053–1074.
Kissel, H., I. Timakhina, M. P. Hardy, G. Rothschild, Y. Tajima 5894–5900.Spritz, R. A., 1994 Molecular basis of human piebaldism. J. Invest.et al., 2000 Point mutation in Kit receptor tyrosine kinase reveals
essential roles for Kit signaling in spermatogenesis and oogenesis Dermatol. 103: 137S–140S.Strippoli, P., S. Sarchielli, R. Santucci, G. P. Bagnara, G. Brandiwithout affecting other Kit responses. EMBO. J. 19: 1312–1326.
Kitayama, H., T. Tsujimura, I. Matsumura, K. Oritani, H. Ikeda et al., 2001 Cold single-strand conformation polymorphism anal-ysis: optimization for detection of APC gene mutations in patientset al., 1996 Neoplastic transformation of normal hematopoietic
cells by constitutively activating mutations of c-kit receptor tyro- with familial adenomatous polyposis. Int. J. Mol. Med. 8: 567–572.Su, A. I., P. M. Cooke, K. A. Ching, Y. Hakak, J. R. Walker et al.,sine kinase. Blood 88: 995–1004.
Kunisada, T., H. Yoshida, H. Yamazaki, A. Miyamoto, H. Hemmi 2002 Large-scale analysis of the human and mouse transcrip-tomes. Proc. Natl. Acad. Sci. USA 99: 4465–4470.et al., 1998 Transgene expression of steel factor in the basal layer
of epidermis promotes survival, proliferation, differentiation and Sunnucks, P., A. C. Wilson, L. B. Beheregaray, K. Zenger,J. French et al., 2000 SSCP is not so difficult: the applicationmigration of melanocyte precursors. Development 125: 2915–2923.
Kuroda, H., N. Terada, H. Nakayama, K. Matsumoto and Y. Kita- and utility of single-stranded conformation polymorphism in evo-lutionary biology and molecular ecology. Mol. Ecol. 9: 1699–1710.mura, 1988 Infertility due to growth arrest of ovarian follicles
in Sl/Slt mice. Dev. Biol. 126: 71–79. Tian, Q., H. F. Frierson Jr., G. W. Krystal and C. A. Moskaluk,1999 Activating c-kit gene mutations in human germ cell tumors.Manova, K., and R. F. Bachvarova, 1991 Expression of c-kit en-
coded at the W locus of mice in developing embryonic germ Am. J. Pathol. 154: 1643–1647.Timokhina, I., H. Kissel, G. Stella and P. Besmer, 1998 Kit signal-cells and presumptive melanoblasts. Dev. Biol. 146: 312–324.
Nolan, P. M., J. Peters, M. Strivens, D. Rogers, J. Hagan et al., ing through PI3-kinase and Src kinase pathways: an essential rolefor Rac1 and JNK activation in mast cell proliferation. EMBO J.2000 A systematic, genome-wide, phenotype-driven mutagene-
sis programme for gene function studies in the mouse. Nat. 17: 6250–6262.Tsujimura, T., 1996 Role of c-kit receptor tyrosine kinase in theGenet. 25: 440–443.
Ogawa, M., Y. Matsuzaki, S. Nishikawa, S. Hayashi, T. Kunisada development, survival and neoplastic transformation of mast cells.Pathol. Int. 46: 933–938.et al., 1991 Expression and function of c-kit in hemopoietic
progenitor cells. J. Exp. Med. 174: 63–71. Tsujimura, T., S. Hirota, S. Nomura, Y. Niwa, M. Yamazaki et al.,1991 Characterization of Ws mutant allele of rats: a 12-base de-Ohta, H., A. Tohda and Y. Nishimune, 2003 Proliferation and
differentiation of spermatogonial stem cells in the W/Wv mutant letion in tyrosine kinase domain of c-kit gene. Blood 78: 1942–1946.Vandenbark, G. R., C. M. deCastro, H. Taylor, S. Dew-Knightmouse testis. Biol. Reprod. 69: 1815–1821.
Piao, X., and A. Bernstein, 1996 A point mutation in the catalytic and R. E. Kaufman, 1992 Cloning and structural analysis of thehuman c-kit gene. Oncogene 7: 1259–1266.domain of c-kit induces growth factor independence, tumorige-
nicity, and differentiation of mast cells. Blood 87: 3117–3123. Witter, O. N., 1990 Steel locus defines new multipotent growthfactor. Cell 63: 5–6.Pielberg, G., C. Olsson, A.-C. Syvanen and L. Andersson, 2002
Unexpectedly high allelic diversity at the KIT locus causing domi- Zemke, D., B. Yamini and V. Yuzbasiyan-Gurkan, 2002 Mutationsin the juxtamembrane domain of c-KIT are associated with highernant white color in the domestic pig. Genetics 160: 305–311.
Price, E. R., H. F. Ding, T. Badalian, S. Bhattacharya, C. Take- grade mast cell tumors in dogs. Vet. Pathol. 39: 529–535.Zhao, S., L. J. Rizzolo and C. J. Barnstable, 1997 Differentiationmoto et al., 1998 Lineage-specific signaling in melanocytes: c-Kit
stimulation recruits p300/CBP to microphthalmia. J. Biol. Chem. and transdifferentiation of the retinal pigment epithelium. Int.Rev. Cytol. 171: 225–266.273: 17983–17986.
Reeves, R. H., and P. D’Eustachio, 1997 Genetic and comparative Zheng, Q. Y., K. R. Johnson and L. C. Erway, 1999 Assessment ofhearing in 80 inbred strains of mice by ABR threshold analyses.mapping in mice, pp.71–131 in Genome Analysis: A Laboratory Manual,
edited by E. D. Green, B. Birren, S. Klapholz, R. M. Myers and Hear. Res. 130: 94–107.P. Hieter. Cold Spring Harbor Laboratory Press, Cold SpringHarbor, NY. Communicating editor: C. Kozak
Dow
nloaded from https://academ
ic.oup.com/genetics/article/169/2/819/6060306 by guest on 13 February 2022

Dow
nloaded from https://academ
ic.oup.com/genetics/article/169/2/819/6060306 by guest on 13 February 2022




![th Anniversary Special Issues (14): Pancreatic cancer ...€¦ · carcinomas are classified as pancreatic ductal adeno-carcinoma (PDAC)[4]. An activating mutation in a key proto-oncogene](https://img.pdfslide.us/doc/110x75/5f92b3c623023e07b6622eec/th-anniversary-special-issues-14-pancreatic-cancer-carcinomas-are-classified.jpg)
























