Embed Size (px)
Citation preview

Research in Veterinary Science 93 (2012) 168–171
Contents lists available at ScienceDirect
Research in Veterinary Science
journal homepage: www.elsevier .com/locate / rvsc
Identification and phenotypic characterization of cd T cells in rat lymph
Sudhanshu Shekhar ⇑, Simon Milling 1, Chris Jenkins, Gordon MacPhersonSir William Dunn School of Pathology, University of Oxford, South Parks Road, Oxford OX1 3RE, UK
a r t i c l e i n f o
Article history:Received 8 November 2010Accepted 15 July 2011
Keywords:cd T cellsPhenotypeLymphRat
0034-5288/$ - see front matter � 2011 Elsevier Ltd. Adoi:10.1016/j.rvsc.2011.07.014
⇑ Corresponding author. Present address: LaboratorDepartment of Medical Microbiology, Faculty of MedRoom-523, 745 Bannatyne Avenue, Winnipeg, Manit204 789 3298; fax: +1 204 789 3926.
E-mail address: [email protected] (S. Sh1 Present address: Institute for Infection, Immunity
Medical, Veterinary and Life Sciences, University of GGlasgow G12 8TA, UK.
a b s t r a c t
cd T cells represent an unconventional subset of T lymphocytes that are abundant in epithelial tissuesand serve as an early immune defense against microbes. We have, for the first time, identified cd T cellsin steady-state thoracic duct lymph (TDL) from rats. The lymph contains cd T cells expressing CD8 but notCD4, CD25, MHC-II or CD103. The percentage of TDL cd T cells in rats does not change when the mesen-teric lymph nodes (MLN) are surgically removed. Our data suggest that a proportion of cd T cells migratefrom the intestine into rat TDL, under steady-state conditions.
� 2011 Elsevier Ltd. All rights reserved.
cd T cells are unconventional T lymphocytes expressing the cdTCR (T cell receptor). Unlike ab T cells, these cells constitute about1–5% of the lymphocytes in blood and lymphoid tissues of humansand mice, but may comprise up to 50% of T cells in skin epitheliaand intestinal mucosae (Haas et al., 1993). The ability of cd T cellsto recognize conserved non-peptide antigens mainly via MHC(Major histocompatibility complex)-unrestricted mechanism alsodistinguishes them from ab T cells (Carding and Egan, 2002).Furthermore, the abundance of cd T cells in the intraepitheliallayer of the small intestine led to the suggestion that these cellshave a unique role in the maintenance of mucosal barriers(Hayday, 2000). Although they have been demonstrated to play arole in immunosurveillance, immunoregulation and protectionagainst infectious diseases (Girardi, 2006; Hayday and Tigelaar,2003), their functional role is still unclear.
Migration of lymphocytes is crucial for functioning of theimmune system. Naïve T lymphocytes enter secondary lymphoidorgans across high endothelial venules (HEV) and may becomeprimed. The primed T lymphocytes exit the lymph nodes (LN) viaefferent lymph vessels and home to the effector sites (Younget al., 2000). On reaching the effector sites, these lymphocyteseither undergo apoptosis or localize in the tissues or return tothe circulation as effector or memory cells. When migrating backto the circulation, these lymphocytes travel to the LN via afferent
ll rights reserved.
y for Infection and Immunity,icine, University of Manitoba,oba, Canada R3E 0J9. Tel.: +1
ekhar).and Inflammation, College of
lasgow, 120 University Place,
lymph. Then, they exit through efferent lymph vessels and enterthe systemic blood circulation (Young et al., 1993; Smith et al.,1970). cd T cells migrating in the lymph show variation in theexpression of surface markers like CD4 and CD8 among differentspecies. The majority of cd T cells are CD8+CD4� in intestinallymph of pigs, whereas they are uniformly CD4�CD8� in bovinesubscapular lymph (Thielke et al., 2003; Rhijn et al., 2007). How-ever, the phenotype and migration pattern of cd T cells in ratlymph remain unknown. So, the aim of the present study was toidentify and phenotypically characterize steady-state cd T cells inthoracic duct lymph (TDL) of rats. Using thoracic duct cannulationand mesenteric lymphadenectomy (MLNX), we collected TDL cells.TDL cells expressing cd TCR were then identified and analyzed forvarious cell surface markers (CD8, CD4, CD25, MHC-II and CD103).We also compared the percentage of cd T cells from TDL of MLNXrats with the percentage of cd T cells in TDL from rats withoutMLNX (non-MLNX). To our knowledge, this is the first study ofcd T cells in rat TDL, and it therefore contributes to the understand-ing the immunobiology of these cells.
PVG (RT1c) rats were bred and maintained under specific path-ogen-free conditions in the Sir William Dunn School of Pathology,University of Oxford, UK. Rats used were males of 12–24 weeks ofage except the rats used for MLNX, which were 4–6 weeks old. Allprocedures were performed under licences issued by the UK HomeOffice. MLNX and thoracic duct cannulation were performed as de-scribed previously (Milling et al., 2006). In brief, at an age of about4–6 weeks the rats were laparotomized. The intestine was exposedand all visible mesenteric lymph nodes (MLN) were excised byblunt dissection. Because of lymph vessel regeneration during thefollowing period of about 6 weeks, an anastomosis developed be-tween the lymph vessels coming from the gut (afferent) and thelymph vessels leading to the thoracic duct (efferent). The afferent

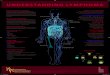
Fig. 1. (A) MLNX TDL samples were stained for cd TCR, ab TCR, CD4, CD8a and CD8b, and analyzed by flow cytometry. The isotype controls were used to gate on different cellpopulations. cd TCR+ cells in the TDL were gated out as shown in (a), and further analyzed for the expression of (b) CD4 and CD8a or (c) CD8a and CD8b. The numbers in theplots represent the cell percentage. (B) MLNX TDL samples were stained for cd TCR, CD25, MHC-II and CD103 and analyzed by flow cytometry. The isotype controls were usedto gate on different cell populations. Expression of (a) CD25, (b) MHC-II and (c) CD103 on cd TCR+ cells (blue line) and isotype controls (red line) were shown. (C) TDL cellswere isolated from MLNX and non-MLNX rats. The cells were stained for cd TCR and analyzed by flow cytometry. Data show the percentage of cd T cells as mean ± SD.Student’s t-test was used to compare the means of cd T cell percentages from MLNX and non-MLNX rats. p value less than 0.05 was considered significant. (For interpretationof the references to colour in this figure legend, the reader is referred to the web version of this article.)
S. Shekhar et al. / Research in Veterinary Science 93 (2012) 168–171 169
lymph obtained from these newly anastomosed vessels is termedas intestinal pseudoafferent lymph (IPL). The thoracic ducts ofMLNX rats were cannulated and TDL was collected on ice.
TDL cells were collected on ice in PBS containing 10 mM EDTAand 20 U/ml heparin. Red blood cells (RBC) were lysed using ACKlysis buffer and the cells were counted using a haemocytometer.Peripheral blood mononuclear cells (PBMC) were isolated fromthe blood using standard Ficoll-Hypaque density gradient centrifu-gation. Spleen and MLN were triturated and filtered through a70 lM Nylon cell strainer. RBC were lysed by incubation in ACKlysis buffer. Analysis of the cells was performed by multi-color flowcytometry. The majority of the monoclonal antibodies (mAb) werepurchased from commercial suppliers, the R73 and OX62 antibod-ies were kindly supplied by the Cellular Immunology Unit, SirWilliam Dunn School of Pathology. The freshly isolated cells fromTDL, spleen, blood or MLN were stained using anti-cd TCR(V65-PE; BD Pharmingen, USA), anti-CD8a (OX8-PerCP; BD Pharm-ingen, UK), or anti-CD8b (341-FITC; BD Pharmingen, UK), anti-CD4(OX35-APC; BD Pharmingen, UK), anti-CD4 (OX35-FITC; BDPharmingen, UK), anti-CD25 (OX39-FITC; BD Pharmingen, UK),anti-MHC-II (OX6-PerCP; BD Pharmingen, UK), anti-ab TCR(R73-Alexa 488; conjugated in house) and anti-CD103(OX62-Alexa 488; conjugated in house) or with respective isotypecontrols (BD Pharmingen, UK). The cells were exposed to the mAbsfor 20 min at 4 �C and washed thrice with FACS buffer (PBS with 5%FCS and 10 mM EDTA). All incubations were carried out at 4 �C inthe dark to prevent photobleaching. Data were collected using a
Cyan (Dako) Flow Cytometer and analyzed using FlowJo software(Tree Star).
To determine whether cd T cells migrate in TDL of rats understeady-state conditions, we collected TDL from MLNX rats on 4occasions, and investigated cd TCR expression on the TDL cells byflow cytometry. On 3 occasions, we used pooled samples fromgroups of 2–3 rats, and measured the percentage of cd T cell. Therewas no statistically significant difference between the cd T cellpercentages in these samples, which were tested using one wayanalysis of variance (p > 0.001). On the 4th occasion, we had asample from an individual animal. We identified a distinct popula-tion of cd T cells in MLNX TDL constituting about 0.5% of the totalTDL cells as shown in Fig. 1A(a). The percentage of cd T cell in theindividual rat TDL was identical to the percentage in pooledsamples, i.e. 0.5%. To better understand the biology of cd T cellrecirculation, we also measured the percentage of cd T cell inTDL samples pooled from groups of 2–3 non-MLNX rats on 3occasions. No statistical significance was observed between the cd T cell percentages from different samples, using one wayanalysis of variance (p > 0.001). We compared the means of the cd T cell percentages calculated from the above mentioned MLNXand non-MLNX experiments, using Student’s t-test. The percent-ages of the cd T cells were expressed as mean ± SD. p value lessthan 0.05 was considered significant. We found that there wasno statistically significant difference between cd T cell percentagesin the TDL of MLNX and non-MLNX rats (Fig. 1C). cd T cells canreach MLN from intestine via intestinal afferent lymph (IAL) ves-

Table 1Percentage of cd T cells in different tissues of rats.
Tissue Mean (%) Standard deviation (SD)
Spleen 0.9433 0.1380TDL 0.5075 0.0464***
MLN 0.5900 0.0100***
Blood 0.5233 0.0416***
Statistical analysis was performed using one way analysis of variance.*** p < 0.001, spleen versus TDL, MLN or blood.
170 S. Shekhar et al. / Research in Veterinary Science 93 (2012) 168–171
sels and from blood via HEV. They exit the MLN and reach TDL viaintestinal efferent lymph (IEL) vessels. Our MLNX versus non-MLNX data, as shown in the Fig. 1C, show that cd T cell percentageis same in the rat TDL irrespective of the presence or absence ofMLN. This clearly indicates that there is no net output of cd T cellsinto the TDL from the MLN. The cd T cells in the TDL must thereforebe derived from the intestine, or from the other lymph nodes thatdrain into the TDL. Because the majority of lymphocytes in the TDLare derived from the efferent output of the MLN (Matsuno et al.,1995), we consider it extremely likely that a significant proportionof the cd T cells in the TDL are derived from the intestine. This viewis consistent with literature in the field that cd T cells comprise 15%of the intestinal intraepithelial lymphocytes (Helgeland et al.,1996), and that the lymphocytes in rat TDL are largely derived fromthe intestinal bed (Mann and Higgins, 1950). We therefore inferthat a large proportion of steady-state cd T cells are likely to enterthe MLN from intestine, rather than from the blood. Consideringthe fact that cd T cells constitute 0.5% of blood cells (Table 1), itseems unlikely that cd T cells that enter the MLN from blood donot contribute to the cd T cell pool. It may be that these cd T cellsare trapped, and eventually undergo apoptosis, within the MLN,and the number of trapped cd T cells is equivalent to the numberof cd T cells that enter the MLN from blood. This is in accordancewith the previous finding that cd T cell percentage in the IAL ofsheep is more than the cd T cell percentage in the IEL (Mackayet al., 1989). Furthermore, homing of the leukocytes to the intes-tine is mediated by CD103 integrin binding to E-cadherin on themucosal cells, and IEL cells in the rat express CD103 (Millinget al., 2007). However, we found no CD103 expression on cd T cellsin rat TDL, as shown in Fig. 1B(c). This appears to contradict theview that cd T cells migrate from the intestine. It may be that ratcd T cells down-modulate CD103 while migrating in TDL. Further-more, recent evidence demonstrates CD103 is not expressed on allcells migrating from the intestine (Milling et al., 2010). Taken to-gether, the data indicate that a proportion of steady-state cd T cellsmigrate from the intestine in TDL, suggesting a role for cd T cells inimmunosurveillance.
The present study also investigated the expression of CD4 andCD8 on rat TDL cd T cells. The TDL was pooled from 2–3 MLNX ratson 3 occasions, and the expression of CD4 and CD8 on TDL cd Tcells was determined using fluorochrome-labeled antibodies, de-tected by flow cytometry. The statistical significance was calcu-lated by one way analysis of variance. p value less than 0.001was considered significant. More than 90% of the cd T cellsexpressed CD8ab heterodimer but not CD8aa homodimer, asshown in Fig. 1A(b and c). To further measure the expression ofthese markers on cd T cells in blood, MLN and spleen, these tissuesamples were harvested from groups of 3 non-MLNX rats on 3occasions, and processed into single cell suspensions. Flowcytometry was performed on single cell suspensions for expressionof cd TCR, CD4, CD8a and CD8b. The percentages of CD8ab+ cd Tcells in blood, MLN and spleen were 91 ± 1.2%, 90 ± 2.5% and90 ± 1.8%, respectively. The statistical significance of the percent-age of CD8ab+ cd T cells in these tissues was determined by oneway analysis of variance and p value less than 0.001 was consid-
ered significant. Kuhnlein et al. (1994) have also reported theexpression of CD8 on about 90% of rat cd T cells from peripherallymphoid organs. Antigen recognition by ab T cells is mediatedby the CD4 or CD8 co-receptors in a MHC-restricted way. Most ofthe human and mouse cd T cells do not express either the CD4or CD8 co-receptors, indicating their lack of MHC restriction. Thebiological significance of the frequent CD8 expression on rat cd Tcells is not understood. It is possible that CD8 could act as a co-receptor for MHC-I mediated antigen recognition by rat cd T cells,analogous to ab T cells. Furthermore, we investigated the expres-sion of CD25 and MHC-II on rat TDL cd T cells. cd T cells have beenshown to up-regulate activation markers like CD25 and MHC-IIupon activation, and function as antigen presenting cells (Brandeset al., 2005). However, cd T cells did not express CD25 or MHC-II inrat TDL, under steady-state conditions, as shown in Fig. 1B(a andb). This lack of expression of CD25 and MHC-II on TDL cd T cellscould be attributed to the steady-state conditions under whichthe cd T cells were studied.
Overall, our data show that there is no significant difference be-tween the steady-state cd T cell percentages in TDL from MLNXand non-MLNX rats and suggest that a proportion of cd T cells mi-grate from the intestine in TDL under steady-state conditions.Migrating cd T cells express CD8ab but not CD103, CD25 orMHC-II. This indicates that CD8ab+ cd T cells may act as a co-receptor for MHC-I mediated antigen recognition.
Conflict of interest
None declared.
Acknowledgment
This study was supported by the Biotechnology and BiologicalSciences Research Council (BBSRC), UK.
References
Brandes, M., Willimann, K., Moser, B., 2005. Professional antigen-presentationfunction by human cd T cells. Science 309, 264–268.
Carding, S.R., Egan, P.J., 2002. cd T cells: functional plasticity and heterogeneity.Nature Review Immunology 2, 336–345.
Girardi, M., 2006. Immunosurveillance and immunoregulation by cd T cells. Journalof Investigative Dermatology 126, 25–31.
Haas, W., Pereira, P., Tonegawa, S., 1993. cd T cells. Annual Review Immunology 11,637–685.
Helgeland, L., Vaage, J.T., Rolstad, B., Midtvedt, T., Brandtzaeg, P., 1996. Microbialcolonization influences composition and T-cell receptor V beta repertoire ofintraepithelial lymphocytes in rat intestine. Immunology 89, 494–501.
Hayday, A.C., 2000. cd T cells: a right time and a right place for a conserved thirdway of protection. Annual Review Immunology 18, 975–1026.
Hayday, A.C., Tigelaar, R., 2003. Immunoregulation in the tissues by cd T cells.Nature Review Immunology 3, 233–242.
Kuhnlein, P., Park, J.H., Herrmann, T., Elbe, A., Hunig, T., 1994. Identification andcharacterization of rat c/d T lymphocytes in peripheral lymphoid organs, smallintestine, and skin with a monoclonal antibody to a constant determinant of thec/d T cell receptor. Journal of Immunology 153, 979–986.
Mann, J.D., Higgins, G.M., 1950. Lymphocytes in thoracic duct intestinal and hepaticlymph. Blood 5, 177–190.
Mackay, C.R., Beya, M., Matzinger, P., 1989. cd T cells express a unique surfacemolecule appearing late during thymic development. European Journal ofImmunology 19, 1477–1483.
Matsuno, K., Kudo, S., Ezaki, T., Miyakawa, K., 1995. Isolation of dendritic cells in therat liver lymph. Transplantation 60, 765–768.
Milling, S.W., Yrlid, U., Jenkins, C., Richards, C.M., Williams, N.A., MacPherson, G.G.,2007. Regulation of intestinal immunity: effects of the oral adjuvant Escherichiacoli heat-labile enterotoxin on migrating dendritic cells. European Journal ofImmunology 37, 87–99.
Milling, S., Jenkins, C., MacPherson, G.G., 2006. Collection of lymph-borne dendriticcells in the rat. Nature Protocol 1, 2263–2270.
Milling, S., Yrlid, U., Cerovic, V., MacPherson, G., 2010. Subsets of migratingintestinal dendritic cells. Immunological Reviews 234, 259–267.
Rhijn, I.V., Rutten, V.P.M.G., Charleston, B., Smits, M., Eden, W., Koets, A.P., 2007.Massive, sustained cd T cell migration from the bovine skin in vivo. Journal ofLeukocyte Biology 81, 968–973.

S. Shekhar et al. / Research in Veterinary Science 93 (2012) 168–171 171
Smith, J.B., Cunningham, A.J., Lafferty, K.J., Morris, B., 1970. The role of the lymphaticsystem and lymphoid cells in the establishment of immunological memory.Australian Journal of Experimental Biology Medical Science 48, 57.
Thielke, K., Moujahid, A., Weisser, C., Waldkirch, E., Pabst, R., Holtmeier, W.,Rothkotter, H.J., 2003. Proliferating intestinal cd T cells recirculate rapidly andare a major source of the cd T cell pool in the peripheral blood. European Journalof Immunology 33, 1649–1656.
Young, A.J., Marston, W.L., Dudler, L., 2000. Subset specific regulation of thelymphatic exit of recirculating lymphocytes in vivo. Journal of Immunology 165,3168–3174.
Young, A.J., Hay, J.B., Mackay, C.R., 1993. Lymphocyte migration in vivo. CurrentTopics in Microbiology Immunology 184, 161.