Embed Size (px)
Citation preview

1 3
Planta (2014) 239:107–126DOI 10.1007/s00425-013-1964-3
ORIGINAL ARTICLE
Identification and characterization of NF‑Y transcription factor families in Canola (Brassica napus L.)
Mingxiang Liang · Xiangzhen Yin · Zhongyuan Lin · Qingsong Zheng · Guohong Liu · Gengmao Zhao
Received: 25 August 2013 / Accepted: 16 September 2013 / Published online: 6 October 2013 © Springer-Verlag Berlin Heidelberg 2013
homologs in Arabidopsis, a model dicot plant, but differ-ent from those in the model monocot plant rice, suggest-ing that plant NF-Ys diverged before monocot and dicot plants differentiated. Spatial-tempo expression patterns, as determined by qRT-PCR, showed that most BnNF-Ys were widely expressed in different tissues throughout the canola life cycle and that several closely related BnNF-Y subunits had similar expression profiles. Based on these findings, we predict that BnNF-Y proteins have functions that are conserved in the homologous proteins in other plants. This study provides the first extensive evaluation of the BnNF-Y family, and provides a useful foundation for dissecting the functions of BnNF-Y.
Keywords BnNF-Y (NUCLEAR FACTOR-Y) · Canola (Brassica napus) · Gene structure · HAP (Heme Activator Protein) · Phylogeny
AbbreviationsNF-Y NUCLEAR FACTOR-YHAP Heme Activator ProteinCBF CCAAT-binding FactorLEC1 LEAFY COTYLEDON 1L1L LEAFY COTYLEDON1-LIKELEC2 LEAFY COTYLEDON 2EST Expressed sequenceORF Open reading frameCDS Coding sequenceUTR Untranslated regionbZIP The basic domain-leucine zipperFT FLOWERING LOCUS TSOC1 SUPPRESSOR OF OVEREXPRESSION OF
CO 1Ehd1 Early heading date1Hd3a Heading date 3a
Abstract NF-Y (NUCLEAR FACTOR-Y), a heterotrim-eric transcription factor, is composed of NF-YA, NF-YB, and NF-YC proteins in yeast, animal, and plant systems. In plants, each of the NF-YA/B/C subunit forms a multi-member family. NF-Ys are key regulators with important roles in many physiological processes, such as drought tolerance, flowering time, and seed development. In this study, we identified, annotated, and further characterized 14 NF-YA, 14 NF-YB, and 5 NF-YC proteins in Bras-sica napus (canola). Phylogenetic analysis revealed that the NF-YA/B/C subunits were more closely clustered with the Arabidopsis thaliana (Arabidopsis) homologs than with rice OsHAP2/3/5 subunits. Analyses of the con-served domain indicated that the BnNF-YA/B/C subfami-lies, respectively, shared the same conserved domains with those in other organisms, including Homo sapiens, Sac-charomyces cerevisiae, Arabidopsis, and Oryza sativa (rice). An examination of exon/intron structures revealed that most gene structures of BnNF-Y were similar to their
X. Yin, Z. Lin, and Q. Zheng contributed equally to this work.
Electronic supplementary material The online version of this article (doi:10.1007/s00425-013-1964-3) contains supplementary material, which is available to authorized users.
M. Liang (*) · X. Yin · Z. Lin · Q. Zheng · G. Zhao College of Resources and Environmental Sciences, Nanjing Agricultural University, Tongwei Road 6, Xuanwu District, Nanjing 210095, Jiangsu Province, Chinae-mail: [email protected]
M. Liang · X. Yin · Z. Lin · Q. Zheng · G. Zhao Jiangsu Key Lab of Marine Biology, Nanjing, China
G. Liu Shandong Hongda Group, Laiyang, China

108 Planta (2014) 239:107–126
1 3
RFT1 RICE FLOWERING LOCUS 1LHCB Light-harvesting Chlorophyll a/b-binding
proteinqRT-PCR Quantitative real-time PCRDAP Days after planting
Introduction
NUCLEAR FACTOR-Y (NF-Y), also known as heme acti-vator protein (HAP) or CCAAT-binding factor (CBF), is a group of transcription factors found in yeast (Hahn and Guarente 1988; McNabb et al. 1995), mouse (Hooft van Huijsduijnen et al. 1990), rat (Maity et al. 1990), human (Becker et al. 1991), plants (Edwards et al. 1998; Stephen-son et al. 2007; Thirumurugan et al. 2008; Cao et al. 2011), and other eukaryotes. It includes at least three subunits: NF-YA (HAP2 or CBF-B), NF-YB (HAP3 or CBF-A), and NF-YC (HAP5 or CBF-C), all of which are required for CCAAT binding and downstream gene transcription (Maity and de Crombrugghe 1998). During transcriptional acti-vation, NF-YA, NF-YB, and NF-YC form a heterotrimer (Kim et al. 1996). Whereas each NF-Y subunit in yeast and animals is encoded by a single gene, each NF-Y subunit in plants is encoded by a multi-gene family (Edwards et al. 1998; Stephenson et al. 2007; Thirumurugan et al. 2008; Cao et al. 2011). In Arabidopsis, there are at least 10 anno-tated members in each NF-Y family (Gusmaroli et al. 2001, 2002; Siefers et al. 2009). Rice harbors at least 10 NF-YA genes, 12 NF-YB genes, and 8 NF-YC genes (Thirumuru-gan et al. 2008). In Triticum aestivum, 10 NF-YA, 11 NF-YB, and 14 NF-YC have been characterized (Stephenson et al. 2007). In Brachypodium distachyon, a model mono-cot plant, 7 NF-YA, 17 NF-YB, and 12 NF-YC have been identified (Cao et al. 2011). The occurrence of multi-mem-ber families in plants could suggest genetic redundancy or functional divergence (Cai et al. 2007; Kumimoto et al. 2008; Liang et al. 2012). Currently, the NF-Ys identified in plants are mainly involved in processes such as embryo development (Lotan et al. 1998; Kwong et al. 2003), fatty acid synthesis (Mu et al. 2008; Tan et al. 2011), chloroplast biogenesis (Miyoshi et al. 2003; Stephenson et al. 2011), nodule development (Combier et al. 2006), stress response (Kreps et al. 2002; Nelson et al. 2007; Li et al. 2008), absci-sic acid (ABA) perception (Warpeha et al. 2007; Yamamoto et al. 2009), root elongation (Ballif et al. 2011), and flower-ing regulation (Wenkel et al. 2006; Cai et al. 2007; Kumi-moto et al. 2008; Cao et al. 2011; Liang et al. 2012).
The finding that NF-Y proteins from animals and yeast share high homology with those form plants allows us to use reverse genetics methods to study NF-Y families in plants (Gusmaroli et al. 2001, 2002; Stephenson et al. 2007; Thirumurugan et al. 2008; Cao et al. 2011). The core
domain of NF-YA consists of two parts, one of which regu-lates the interaction with NF-YB or NF-YC and the other of which is required for DNA binding (Xing et al. 1993, 1994; Mantovani et al. 1994). Both NF-YB and NF-YC are structurally similar to histone subunits H2B and H2A, due to the presence of the highly conserved histone-fold motif (HFM) (Mantovani 1999). NF-YB and NF-YC both con-tain separate domains for DNA contact and mutual interac-tion (Romier et al. 2003).
Although NF-Y members have been identified in sev-eral plant species, particularly model plants, their roles in canola were hitherto unclear. Canola (Brassica napus) plants constitute the third largest vegetable oil source in the world. Although NF-Y members were reported in canola around 20 years ago, only a few NF-Y family members have been described in detail. To achieve higher oil con-tent, molecular strategies were employed to identify regu-lators of plant embryo development that have been found in several gene classes to be involved in this process, such as LEC1 (LEAFY COTYLEDON 1), L1L (LEAFY COTYLE-DON1-LIKE), LEC2 (LEAFY COTYLEDON 2), and WRIN-KLED1 (Lotan et al. 1998; Focks and Benning 1998; Stone et al. 2001; Kwong et al. 2003). Of these, LEC1 and L1L, which play central roles in embryogenesis, were best stud-ied (Lotan et al. 1998; Kwong et al. 2003; Lee et al. 2003; Yazawa and Kamada 2007; Alemanno et al. 2008). LEC1 is a member of the NF-YB family and can induce embryo development even in vegetative cells (Lotan et al. 1998). Mutant plants of lec1 exhibit abnormal cotyledons and des-iccation-intolerant embryos, while constitutive overexpres-sion of LEC1 in wild-type Arabidopsis plants or lec1 plants results in embryo-like structures on the leaves. Kwong et al. (2003) reported that another NF-YB in Arabidopsis, L1L, which showed high sequence identity to LEC1, could par-tially replace LEC1 in embryo development. However, L1L and LEC1 showed different temporal expression patterns in plants, suggesting that those two genes may function dif-ferently during embryogenesis. To circumvent the seedling lethality that is common in transgenic lines constitutively expressing LEC1 or L1L. Mu et al. (2008) constructed a transgenic line in which LEC1 or L1L expression was driven by an estradiol-inducible promoter, and observed that LEC1 or L1L plays a role in fatty acid synthesis. Over-expressor plants accumulated increased levels of fatty acid species and lipids. Furthermore, BnLEC1 and BnL1L were found to have similar functions in regulating the lipid path-way as their Arabidopsis homologs. A recent follow-up study by the same group also revealed that the conditional expression of BnLEC1 and BnL1L in canola increased seed oil content without having any obvious developmen-tal effect, a finding which may have agricultural applica-tions (Tan et al. 2011). Moreover, the developing seeds and silique walls of transgenic plants expressing BnLEC1

109Planta (2014) 239:107–126
1 3
exhibit enhanced expression of several genes involved in sucrose synthesis and transport. Apart from LEC1 and L1L, CBF-B, which is orthologous to AtNF-YA9, is another NF-Y subunit reported in canola (Albani and Robert 1995). The RNA of this NF-YA was detected in sepals, petals, and siliques, while an alternative splicing transcript of this protein was detected by RNA blot analysis. A subsequent study by the same group demonstrated that the expression of an antisense construct of BnNF-YA under the tapetal-specific promoter BnOlnB;4 in transgenic canola plants reduced both male and female viability (Lévesque-Lemay et al. 2003).
Even though canola has been extensively studied due to the high nutritional value of the fatty acids in the seeds and its suitability for biodiesel production, little is known about the NF-Y members in this plant. This is at least partly due to the large size of the canola genome. Canola is an allo-tetraploid that contains genomes of the species Brassica rapa and Brassica oleracea. Recently, the entire sequence of the B. rapa genome (Accession chiifu-401-42), which is over 280 Mb, was released and there are plans to sequence the genomes of other Brassicaceae species (Wang et al. 2011). Interestingly, the difference in genome size between B. rapa and A. thaliana (125 Mb) is largely caused by transposable elements as opposed to protein coding genes, since more than 93 % of gene families are shared by these two species (Wang et al. 2011).
In the current study, we aimed to use sequence infor-mation from the model dicotyledonous plant Arabidopsis to identify all of the NF-Y genes in canola species and to determine their transcriptional profiles. Furthermore, we analyzed the evolutionary relationship between these genes and their homologs in other plant species and examined the exon/intron organization of the canola NF-Y gene families. Definitely, some canola NF-Ys would be missing in our study due to incompleteness of canola EST database and defect of our methodology, but the results of this study will provide an entry point for investigating the roles of each canola NF-Y gene in regulating plant growth.
Materials and methods
Identification and verification of BnNF-Y family members in canola
The full-length cDNA or CDS of each of the 30 predicted Arabidopsis NF-Ys (Siefers et al. 2009; Petroni et al. 2012) was used to sequentially search the oilseed rape database (http://www.compbio.dfci.harvard.edu/cgi-bin/tgi/gimain.pl?gudb=oilseed_rape) (Release 5.0). Arabidopsis NF-Y subunit sequences were retrieved from the Arabidopsis Information Resource (http://www.arabidopsis.org). The
BLASTN program was used with an E value cut-off of 8.2e−05 to identify both assembled oil rape EST sequences and EST singletons. All Arabidopsis NF-Ys corresponded with at least one homolog in the oil rape database. Each BnNF-Y sequence was retrieved from the database and primers used to amplify the CDSs were designed with Primer Premier 5.0. The PCR products obtained from both cDNA and genomic DNA amplification were cloned into the pMD®19-T cloning vector (TaKaRa Code: D102A), and then transformed into DH5α Escherichia coli cells. Due to initial sequencing errors in the database and DNA polymorphisms in canola cultivars, nucleotide sequences with 98 % identity over their length were considered as the same gene. All cloned BnNF-Y sequences were already submitted to the NCBI website (Supplementary Table S1).
Phylogenetic trees, conservation domains, and exon/intron gene structure analyses
The phylogenetic trees of BnNF-Y, AtNF-Y, and OsHAP subunit families were constructed using the neighbor-join-ing method in Molecular Evolutionary Genetics Analy-sis (MEGA; version 4.1) with the following parameters: Jones–Taylor–Thornton (JTT) model and 1,000 bootstrap replicates. Multiple sequence alignments were first per-formed using the amino acid sequences of the conserved region and full-length protein by ClustalX software and then crested with CLC Sequence Viewer 6.8 software (http://www.clcbio.com/products/clc-sequence-viewer/). The sequences of AtNF-Ys and OsHAPs were retrieved from TAIR (http://www.arabidopsis.org/) and OrygenesDB (http://orygenesdb.cirad.fr/), respectively. The exon/intron gene structures were constructed by comparing the CDSs with their corresponding genomic DNA (without UTR) sequences and analyzed using the Gene Structure Display Serve tool (http://gsds.cbi.pku.edu.cn/).
Plant material and growth conditions
Plants of oilseed rape (Brassica napus ‘Nanyanyou 1’) were grown in thirty pots (30-cm diameter) containing organic cultivation matrix (Zhenjiang Xing Nong Organic Fertilizer Co., Ltd.) in late September. The pots were first placed in a greenhouse for 75 days, and then leaves, stems, and roots were harvested (before vernalization). The same plants were then transferred to an open room for vernalization. Leaves, stems, and roots were col-lected at 153 DAP (Day After Planting). At 177 DAP, the inflorescence had just become visible, and leaves, stems, roots, and inflorescences were collected again. The same tissue types were collected at 202 DAP, when numerous flowers were present. Finally, leaves, roots, and siliques were collected at 220 DAP, during seed maturation. The

110 Planta (2014) 239:107–126
1 3
collected samples were immediately dipped in liquid nitrogen and stored at −80 °C until used for gene expres-sion analysis. All experiments were performed in biologi-cal triplicate.
Genomic DNA extraction, total RNA isolation, and primary cDNA synthesis
Prior to extraction of genomic DNA and total RNA, sam-ples were grounded in pre-frozen mortars. DNA was extracted from oilseed rape using a method from Rogers and Bendich’s (1985) CTAB-based protocol. Total RNA of each sample was extracted using an E.Z.N.A. Plant RNA Kit (Omega Biotek, Cat#R6827), following the manufacturer’s instructions. The quality and quantity of extracted DNA and RNA from all samples was confirmed by both agarose gel visualization and spectrophotometry (Thermo Scientific, NanoDropTM 1000). Prior to reverse transcription, total RNA samples were pretreated with an RNase-Free DNase Set (Omega Biotek, Cat#E1091) at 25–30 °C for 15 min to eliminate any contaminating genomic DNA. Primary cDNA was synthesized using a PrimeScript® 1st Strand cDNA Synthesis Kit (TaKaRa Code: D6110A), according to the manufacturer’s instructions.
Analysis of gene expression by quantitative real-time PCR (qRT-PCR)
QRT-PCR was performed to quantify the transcription lev-els of BnNF-Y genes expressed in different developmental stages and various tissues using the 2−ΔΔCт method, since the deviation error of the amplification efficiency between target genes and the reference gene was less than 10 % according to our trial experiments. 18S rRNA was used as an internal control. Total RNA was prepared as above. Briefly, 3 μg of total RNA was used for the synthesis of first-strand cDNA using oligo (dT) as a primer. Real-time PCR was performed on an Applied Biosystems 7500 real-time PCR system using the SYBR® Premix ExTaqTM Kit (TaKaRa Code:DRR041A). Each 20.0-μl qRT-PCR reac-tion included 10.0 μl SYBR® premix ExTaqTM (2×), 0.4 μl Forward Primer (10 μM), 0.4 μl Reverse Primer (10 μM), 0.4 μl ROX Reference DyeII (50×), and 2.0 μl cDNA diluted ten times as template. The reaction was initi-ated at 95 °C for 30 s, followed by 40 cycles of 95 °C for 3 s and 60 °C for 34 s with a final dissociation stage. The specificity of primers designed for real-time RT-PCR was confirmed by separating the products on agarose gels and clone sequencing. The data presented for RT-PCR experi-ments are the average relative quantities from three biologi-cal replicates, where every biological replicate is the mean of three technical repeats.
Results
Identification and annotation of canola BnNF-Y
Full-length and conserved regions of each of the 30 reported Arabidopsis NF-Ys (Siefers et al. 2009; Petroni et al. 2012) were used to sequentially BLAST the oilseed rape EST database (http://compbio.dfci.harvard.edu/cgi-bin/tgi/gimain.pl?gudb=oilseed_rape). The sequences with high similarity (E value lower than 8.2e−05) were retrieved from the database. Ninety tentative consen-sus (TC) sequences (40 TC sequences similar to AtNF-YAs, 32 TC sequences similar to AtNF-YBs, and 18 TC sequences similar to AtNF-YCs, data not shown) were obtained in total. Some NF-Y-like ESTs with appar-ently incorrect ORFs (open reading frame) or domains were excluded. More than 10 members showed lit-tle similarities with NF-Y family, but further classi-fied as BnNC2s or BnDbp3/4s, and were not included here as a recent review suggested (Petroni et al. 2012). Thus, 78 TC sequences (30 TC sequences similar to AtNF-YAs, 30 TC sequences similar to AtNF-YBs, and 18 TC sequences similar to AtNF-YCs; Supplementary Table S1) were selected and used as templates to design primers to compare our sequences with ESTs identi-fied in the canola database. These sequenced TC mem-bers were then aligned with AtNF-Ys to identify the putative coding sequences (CDSs, including the stop codon) in DNASTAR (Lasergene. v7.1), respectively. Only partial CDSs were available for a few TCs. To obtain the full-length CDSs for these sequences, addi-tional sequences were retrieved from the NCBI (Sup-plementary Table S1). Primers were designed according to sequences flanking the putative CDSs. Particularly, the primers of BnNF-YA3, BnNF-YA13, and BnNF-YA14 were individually designed according to Brassica rapa subsp. pekinensis sequences retrieved from the NCBI, and BnNF-YA2 was designed based on five B. napus sequences (CN731758, EV101783, EV101871, EV103401, and FG556290) retrieved from the NCBI (Supplementary Table S1). For BnNF-YB6, only a par-tial CDS was available for the sequence with the high-est similarity, GR449214. In contrast to TC173425, the 5′ end of GR449214 was found to be highly similar to that of TC173425, and so we attempted to amplify this sequence using the forward primer of TC173425 and the reverse primer of GR449214. Either canola cDNA or genomic DNA was used as template. As a result, both sequences were successfully amplified. Sequencing revealed that these sequences appeared to be full-length sequences and contained the original partial GR449214 sequence. All primer pairs used to amplify genomic DNA and cDNA of BnNF-Ys are listed in Supplementary Table
111Planta (2014) 239:107–126
1 3
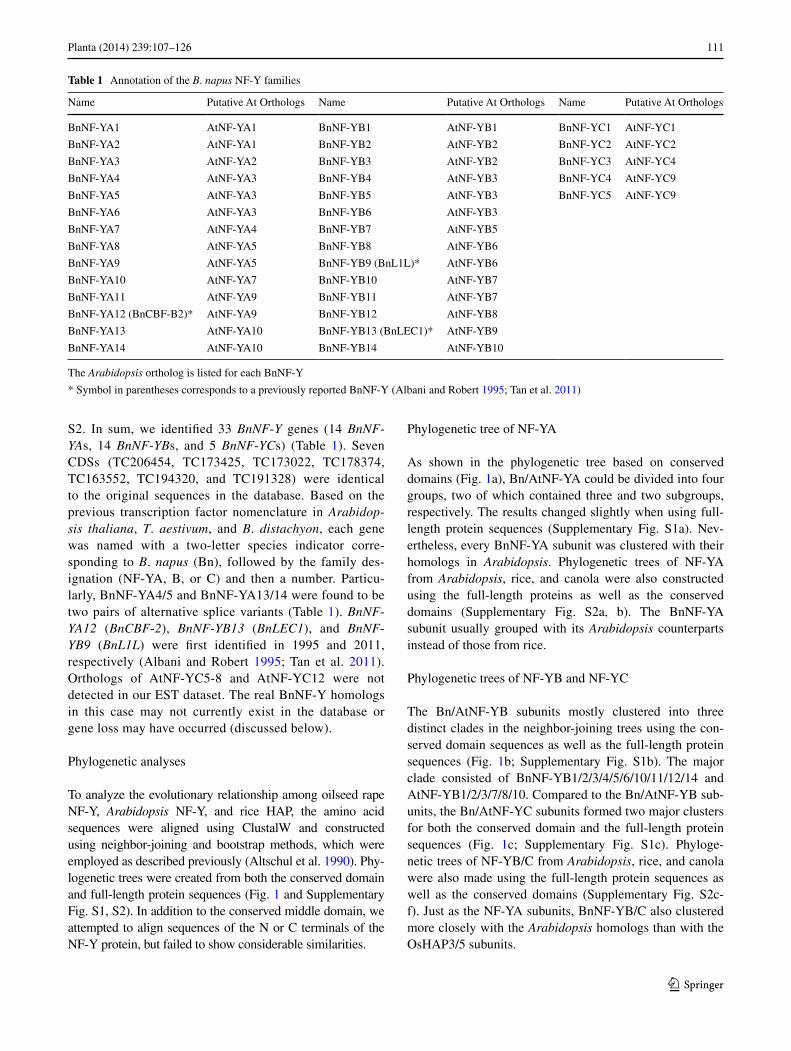
S2. In sum, we identified 33 BnNF-Y genes (14 BnNF-YAs, 14 BnNF-YBs, and 5 BnNF-YCs) (Table 1). Seven CDSs (TC206454, TC173425, TC173022, TC178374, TC163552, TC194320, and TC191328) were identical to the original sequences in the database. Based on the previous transcription factor nomenclature in Arabidop-sis thaliana, T. aestivum, and B. distachyon, each gene was named with a two-letter species indicator corre-sponding to B. napus (Bn), followed by the family des-ignation (NF-YA, B, or C) and then a number. Particu-larly, BnNF-YA4/5 and BnNF-YA13/14 were found to be two pairs of alternative splice variants (Table 1). BnNF-YA12 (BnCBF-2), BnNF-YB13 (BnLEC1), and BnNF-YB9 (BnL1L) were first identified in 1995 and 2011, respectively (Albani and Robert 1995; Tan et al. 2011). Orthologs of AtNF-YC5-8 and AtNF-YC12 were not detected in our EST dataset. The real BnNF-Y homologs in this case may not currently exist in the database or gene loss may have occurred (discussed below).
Phylogenetic analyses
To analyze the evolutionary relationship among oilseed rape NF-Y, Arabidopsis NF-Y, and rice HAP, the amino acid sequences were aligned using ClustalW and constructed using neighbor-joining and bootstrap methods, which were employed as described previously (Altschul et al. 1990). Phy-logenetic trees were created from both the conserved domain and full-length protein sequences (Fig. 1 and Supplementary Fig. S1, S2). In addition to the conserved middle domain, we attempted to align sequences of the N or C terminals of the NF-Y protein, but failed to show considerable similarities.
Phylogenetic tree of NF-YA
As shown in the phylogenetic tree based on conserved domains (Fig. 1a), Bn/AtNF-YA could be divided into four groups, two of which contained three and two subgroups, respectively. The results changed slightly when using full-length protein sequences (Supplementary Fig. S1a). Nev-ertheless, every BnNF-YA subunit was clustered with their homologs in Arabidopsis. Phylogenetic trees of NF-YA from Arabidopsis, rice, and canola were also constructed using the full-length proteins as well as the conserved domains (Supplementary Fig. S2a, b). The BnNF-YA subunit usually grouped with its Arabidopsis counterparts instead of those from rice.
Phylogenetic trees of NF-YB and NF-YC
The Bn/AtNF-YB subunits mostly clustered into three distinct clades in the neighbor-joining trees using the con-served domain sequences as well as the full-length protein sequences (Fig. 1b; Supplementary Fig. S1b). The major clade consisted of BnNF-YB1/2/3/4/5/6/10/11/12/14 and AtNF-YB1/2/3/7/8/10. Compared to the Bn/AtNF-YB sub-units, the Bn/AtNF-YC subunits formed two major clusters for both the conserved domain and the full-length protein sequences (Fig. 1c; Supplementary Fig. S1c). Phyloge-netic trees of NF-YB/C from Arabidopsis, rice, and canola were also made using the full-length protein sequences as well as the conserved domains (Supplementary Fig. S2c-f). Just as the NF-YA subunits, BnNF-YB/C also clustered more closely with the Arabidopsis homologs than with the OsHAP3/5 subunits.
Table 1 Annotation of the B. napus NF-Y families
The Arabidopsis ortholog is listed for each BnNF-Y
* Symbol in parentheses corresponds to a previously reported BnNF-Y (Albani and Robert 1995; Tan et al. 2011)
Name Putative At Orthologs Name Putative At Orthologs Name Putative At Orthologs
BnNF-YA1 AtNF-YA1 BnNF-YB1 AtNF-YB1 BnNF-YC1 AtNF-YC1
BnNF-YA2 AtNF-YA1 BnNF-YB2 AtNF-YB2 BnNF-YC2 AtNF-YC2
BnNF-YA3 AtNF-YA2 BnNF-YB3 AtNF-YB2 BnNF-YC3 AtNF-YC4
BnNF-YA4 AtNF-YA3 BnNF-YB4 AtNF-YB3 BnNF-YC4 AtNF-YC9
BnNF-YA5 AtNF-YA3 BnNF-YB5 AtNF-YB3 BnNF-YC5 AtNF-YC9
BnNF-YA6 AtNF-YA3 BnNF-YB6 AtNF-YB3
BnNF-YA7 AtNF-YA4 BnNF-YB7 AtNF-YB5
BnNF-YA8 AtNF-YA5 BnNF-YB8 AtNF-YB6
BnNF-YA9 AtNF-YA5 BnNF-YB9 (BnL1L)* AtNF-YB6
BnNF-YA10 AtNF-YA7 BnNF-YB10 AtNF-YB7
BnNF-YA11 AtNF-YA9 BnNF-YB11 AtNF-YB7
BnNF-YA12 (BnCBF-B2)* AtNF-YA9 BnNF-YB12 AtNF-YB8
BnNF-YA13 AtNF-YA10 BnNF-YB13 (BnLEC1)* AtNF-YB9
BnNF-YA14 AtNF-YA10 BnNF-YB14 AtNF-YB10

112 Planta (2014) 239:107–126
1 3
Conservation domains analyses
The genes of three BnNF-Y subfamilies encode putative transcription factors that are variable in length (143–316 amino acids). Each member of the NF-Y family contains evolutionarily conserved domains that are responsible for heterodimerization, heterotrimerization, and DNA interac-tions at the CCAAT sites. To investigate the evolutionarily conserved domains of BnNF-Y, multiple sequence align-ments for each NF-Y family member were performed in ClustalX (Thompson et al. 1997) using full-length protein sequences and then crested with CLC Sequence Viewer 6.8 software (http://www.clcbio.com/products/clc-sequence-viewer/). The multiple alignments showed the conserved domains for each NF-Y family member, with their inter-action domains highlighted (Xing et al. 1994; Kim et al. 1996; Sinha et al. 1996; McNabb et al. 1997; Romier et al. 2003) (Fig. 2). A comparison of amino acid sequences of the conserved domain showed that BnNF-Y proteins are highly similar to mammalian (Such as Homo sapiens and mouse), Saccharomyces cerevisiae, Arabidopsis, and rice NF-Y proteins (Supplementary Fig. S3).
BnNF-YA subunits
The conserved core region of BnNF-YA is ~53 amino acids in length, which is approximately the length suggested by Maity and de Crombrugghe (1992). In mammalian and S. cerevisiae NF-YA proteins, three histidine (H) and three arginine (R) residues are absolutely conserved, and are essential for DNA binding (Xing et al. 1993). These resi-dues are all conserved in the 14 BnNF-YAs. The same con-served residues were often identified in Arabidopsis, rice, T. aestivum (wheat), and Brachypodium NF-YA members (Siefers et al. 2009; Thirumurugan et al. 2008; Stephenson et al. 2007; Cao et al. 2011). This domain comprises two subdomains, an A1 domain for the NF-YB/C interaction and an A2 domain for DNA contact (Fig. 2a and Supplementary Fig. S3a) (Xing et al. 1993, 1994; Mantovani et al. 1994). All of these sequences include 21 absolutely conserved amino acids (Fig. 2a). Therefore, the specific function of this region appears to be highly conserved in all BnNF-YA proteins. In addition, the two pairs of alternative splice variants, BnNF-YA4/5 and BnNF-YA13/14, were found to contain the same conserved domain sequence. However, the conserved core residues of BnNF-YA still have some differ-ences with those from other species. For example, BnNF-YA13 and BnNF-YA14 contain the amino acid sequence LKN, while AtNF-YA10 does not. Furthermore, the V, I, and S residues from BnNF-YA13 and BnNF-YA14 occur at position 21, 26, and 28, respectively; whereas the E, L, and R residues occur at the same positions in the AtNF-YA10 amino acid sequence (Supplementary Fig. S3a).
AtNF-YA1 BnNF-YA2 BnNF-YA1
AtNF-YA7 BnNF-YA10
AtNF-YA4 BnNF-YA7
BnNF-YA3 BnNF-YA14 BnNF-YA13
AtNF-YA2 AtNF-YA10
BnNF-YA4 BnNF-YA5
AtNF-YA3 BnNF-YA6
AtNF-YA8 BnNF-YA8 AtNF-YA5
BnNF-YA9 AtNF-YA6
BnNF-YA12 BnNF-YA11
AtNF-YA9 MouseNF-YA
88
5060
99
6899
98
90
80
36
85
29
39
48
6091
74
95
6485
3067
0.1
(a)
BnNF-YB5 BnNF-YB6
AtNF-YB3 BnNF-YB4
AtNF-YB2 BnNF-YB2 BnNF-YB3
BnNF-YB11 AtNF-YB7
BnNF-YB10 AtNF-YB1
BnNF-YB1 BnNF-YB14
AtNF-YB10 AtNF-YB8
BnNF-YB12 AtNF-YB5
BnNF-Y7 AtNF-YB4
AtNF-YB9 BnNF-YB13
BnNF-YB9 AtNF-YB6 BnNF-YB8
MouseNF-YB76
100
99
100
9989
995099
4479
87
42
63
99
48
27
61
66
7478
0.1
(b)
AtNF-YC3 AtNF-YC9 BnNF-YC4 BnNF-YC5 AtNF-YC2
BnNF-YC2 BnNF-YC1 BnNF-YC3 AtNF-YC1 AtNF-YC4
AtNF-YC6 AtNF-YC12
AtNF-YC7 AtNF-YC5
AtNF-YC8 MouseNF-YC
9940
4680
91
99
50
47
99
0.1
(c)
Fig. 1 Phylogenetic trees of canola and Arabidopsis NF-Y families based on the conserved domains. The phylogenetic trees were con-structed using the conserved amino acid sequences by the neighbor-joining method implemented by MEGA software, version 4.1. The numbers at each branch point represent the bootstrap scores (1,000 replicates). A branch with a bootstrap score below 50 was usually considered unreliable. Sequences of the mouse NF-Y subunits were used to root the trees, respectively. a Phylogenetic tree of canola and Arabidopsis NF-YA families based on the conserved domains. b Phy-logenetic tree of canola and Arabidopsis NF-YB families based on the conserved domains. c Phylogenetic tree of canola and Arabidop-sis NF-YC families based on the conserved domains

113Planta (2014) 239:107–126
1 3
Fig. 2 Brassica napus NF-Y family member alignments. Amino acid sequences correspond to the conserved regions in all BnNF-Y proteins. Hs, Homo sapiens; Mm, Mus musculus; Sc, S. cerevisiae. Numbers in parentheses correspond to the actual positions of the first amino acids of the conserved domains within each protein; num-bers above the alignment were used as reference points in the text; and numbers to the right correspond to the amount of each amino acid presented here. In the consensus line, uppercase letters repre-sent identity in more than 50 % of sequences and X represents less than 50 % identity. a BnNF-YA family alignment. To eliminate some gaps resulting from nonhomologous sequences, the Q residues were removed from BnNF-YA13 and BnNF-YA14 between the K and P residues at position 31/32. The amino acid sequence VLD between the A and Q residues at 22/23 and the F between the K and R resi-dues at position 29/30 from BnNF-YA3 were removed. The NF-YA conserved regions were composed of two alpha-helices: A1 mediates the NF-YB–NF-YC interaction and A2 is responsible for CCAAT
binding (Xing et al. 1993, 1994; Mantovani et al. 1994). b BnNF-YB family alignment. The secondary structures, alpha-helices (solid blue rectangles) and coils (black lines), are represented on the top of the alignment, based on (Romier et al. 2003). The DNA-binding and subunit-binding domains are represented as black and colored bars, respectively (Romier et al. 2003; Sinha et al. 1996). The NF-YC interaction domain extends across two independent regions and partly overlaps with the DNA-binding and NF-YA interaction domains. c BnNF-YC family alignment. The secondary structures, alpha-helices (solid blue rectangles) and coils (black lines), are rep-resented on the top of the alignment, based on (Romier et al. 2003). The DNA-binding and subunit-binding domains are represented by black and colored bars, respectively (Romier et al. 2003; Kim et al. 1996; McNabb et al. 1997). The NF-YA interaction domain extends across two separate regions. The DNA-binding domain in BnNF-YC consists of two amino acids, AR

114 Planta (2014) 239:107–126
1 3
BnNF-YB and BnNF-YC subunits
The BnNF-YB subunit core region is ~95 amino acids in length, which is close to that found in other species (Maity and de Crombrugghe 1992). This region contains the domains for DNA binding, protein–protein interac-tions (Sinha et al. 1996; Romier et al. 2003). As shown in Fig. 2b and Supplementary Fig. S3b, this region has structural similarities with the histone-fold motif (HFM) of the core histone H2B (Mantovani 1999). The BnNF-YC subunit core region is ~77 amino acids long, which is shorter than the average length of NF-YC subunits reported by Maity and de Crombrugghe (1992). Like BnNF-YB proteins, BnNF-YC proteins are also charac-terized by a core histone, but the core histone is more similar to H2A than H2B (Mantovani 1999) (Fig. 2c and Supplementary Fig. S3c).
Identity analyses
Molecular analysis of the deduced BnNF-Y polypeptides by the ClustalW method using MegAlign in DNASTAR (Lasergene.v7.1) identified high levels of similarity between members of the BnNF-Y families (Table 2). Generally, the conserved domains of individual BnNF-Y members within each family showed much higher levels of similarity than did the full-length CDSs, suggesting that fragments of the BnNF-Ys outside of the conserved domain have low identities. In the BnNF-YA family, the closely related genes, BnNF-YA1 and BnNF-YA2, BnNF-YA4/5 and BnNF-YA6, BnNF-YA8 and BnNF-YA9, and BnNF-YA11 and BnNF-YA12, were shown to share much higher levels of sequence identity (>96.1 % at the conserved domain level and >80.4 % at the full-length protein level), respectively, compared to the other putative paralogs (Table 2a). The alternative splice vari-ants, BnNF-YA4 and BnNF-YA5, were found to share 100 % sequence identity by the ClustalW method, but BnNF-YA4 was 11 amino acids (QANFSFHYADP) shorter than BnNF-YA5. In the BnNF-YB family, the closely related genes, BnNF-YB2 and BnNF-YB3, con-tained the same conserved domain sequences (Table 2b; 100 % sequence identity for both conserved domains and 92.2 % at the full-length protein level). The three closely related genes, BnNF-YB4, BnNF-YB5, and BnNF-YB6, were shown to have 96.8–98.9 % sequence similari-ties at the conserved domain level (Table 2b). The con-served domain sequence identities for the closely related genes, BnNF-YB8/9 and BnNF-YB10/11, were 98.9 and 97.9 %, respectively. In addition, the conserved domain sequences of BnNF-YB2 and BnNF-YB3 were found to be highly similar to those of BnNF-YB4, BnNF-YB5, and BnNF-YB6 (93.7–94.7 %, Table 2b). In Arabidopsis,
AtNF-YB2 was found to be highly similar to AtNF-YB3 (93.7 % identity for the conserved domain sequence). The level of sequence identity between BnNF-YB12 and BnNF-YB14 was found to be 91.6 % for their conserved domain sequences (Table 2b), while in Arabidopsis, the level of sequence identity between their orthologs AtNF-YB8 and AtNF-YB10 was found to be 94.7 %. In the BnNF-YC family, the level of sequence identity for the conserved domains of BnNF-YC1 and BnNF-YC3 was found to be 100 %, as it was for BnNF-YC4 and BnNF-YC5 (Table 2c).
Exon/intron gene structure analysis
Analysis of gene structure may provide some information about the evolutionary mechanisms underlying the genesis of gene families. To investigate the evolutionary conserva-tion and sequence divergence of NF-Ys between canola and the dicot Arabidopsis or the monocot rice, we deter-mined their corresponding exon/intron gene structures. The genomic DNA and cDNA sequences of BnNF-Ys were cloned and compared. All of these NF-Y genes were interrupted by introns, except for some NF-YB/C subfam-ily members. The gene structures without the untranslated regions (UTRs) were constructed for BnNF-Ys, AtNF-Ys, and OsHAPs using the Gene Structure Display Serve tool (http://gsds.cbi.pku.edu.cn/) (Fig. 3 and Supplemen-tary Fig. S4). In addition, all introns except the last one of OsHAP3A follow the GT-AG splicing rule, and each of the last exons includes a stop codon.
Gene structures of BnNF-YAs
As shown in Fig. 3, most of the members of the BnNF-YA family are in agreement with those of their putative orthologs in Arabidopsis. Generally, BnNF-YA or AtNF-YA members contain three to five introns in their cod-ing regions, with the exception of AtNF-YA5, which has only two introns. The introns of the BnNF-YA and AtNF-YA members range in length from 65 to 486 bp (base pair). Particularly, the third exon of BnNF-YA5 contains a partial sequence of the second intron of BnNF-YA4. The alternative splice variants, BnNF-YA13 and BnNF-YA14, share the first three exons and introns, and the fourth exon and part of the fourth intron of BnNF-YA14 form the fourth exon of BnNF-YA13. Nevertheless, both BnNF-YA8 and BnNF-YA9 have one more exon than their Arabidopsis putative ortholog, AtNF-YA5, while both of their first exons had clearly split into two exons in oil rape. Rice NF-YA members share two to five introns, with a very long intron in OsHAP2E (1633 bp), OsHAP2I (7115 bp), and OsHAP2J (1055 bp) (Supplementary Fig. S4).

115Planta (2014) 239:107–126
1 3
Tabl
e 2
Iden
tity
mat
rix
of th
e B
nNF-
Y f
amily
for
the
full-
leng
th p
rote
in a
nd th
e co
nser
ved
dom
ain
sequ
ence
s
(a)
Am
ino
acid
iden
tify
for
full-
leng
th p
rote
in
BnN
F-
YA
1B
nNF-
Y
A2
BnN
F-
YA
3B
nNF-
Y
A4
BnN
F-
YA
5B
nNF-
Y
A6
BnN
F-
YA
7B
nNF-
Y
A8
BnN
F-
YA
9B
nNF-
Y
A10
BnN
F-
YA
11B
nNF-
Y
A12
BnN
F-
YA
13B
nNF-
YA
14
Am
ino
acid
iden
tify
for
cons
erve
d do
mai
n
BnN
F-Y
A1
–80
.524
.127
.727
.626
.926
.527
.629
.026
.844
.640
.823
.122
.8
BnN
F-Y
A2
96.2
–22
.826
.326
.325
.327
.925
.927
.627
.143
.041
.824
.723
.9
BnN
F-Y
A3
67.3
65.4
–24
.524
.625
.227
.925
.227
.329
.522
.022
.465
.560
.9
BnN
F-Y
A4
67.3
65.4
67.9
–10
0.0
87.5
34.7
40.8
43.5
37.0
25.4
25.9
23.0
24.9
BnN
F-Y
A5
67.3
65.4
67.8
100.
0–
87.9
34.7
39.6
42.9
37.0
25.3
25.7
23.2
25.0
BnN
F-Y
A6
67.3
65.4
69.8
98.1
98.1
–34
.741
.045
.238
.724
.424
.823
.725
.6
BnN
F-Y
A7
63.5
65.4
67.9
73.6
73.6
75.5
–34
.432
.864
.626
.626
.029
.529
.8
BnN
F-Y
A8
73.1
71.2
67.9
88.7
88.7
88.7
77.4
–81
.440
.324
.623
.925
.025
.8
BnN
F-Y
A9
69.2
67.3
66.0
86.8
86.8
86.8
73.6
96.2
–38
.929
.627
.927
.428
.4
BnN
F-Y
A10
71.2
69.2
73.6
77.4
77.4
79.2
84.9
81.1
77.4
–30
.729
.829
.529
.1
BnN
F-Y
A11
78.8
78.8
67.9
75.5
75.5
77.4
67.9
73.6
71.7
73.6
–92
.922
.223
.1
BnN
F-Y
A12
76.9
76.9
66.0
73.6
73.6
75.5
66.0
71.7
69.8
71.7
98.1
–21
.322
.1
BnN
F-Y
A13
65.4
67.3
87.0
60.4
60.4
62.3
67.9
60.4
58.5
67.9
66.0
64.2
–95
.5
BnN
F-Y
A14
65.4
67.3
87.0
60.4
60.4
62.3
67.9
60.4
58.5
67 9
66.0
64.2
100.
0–
(b)
Am
ino
acid
iden
tify
for
full-
leng
th p
rote
in
BnN
F-
YB
1B
nNF-
Y
B2
BnN
F-
YB
3B
ILN
F-
YB
4B
nNF-
Y
B5
BnN
F-
YB
6B
nNF-
Y
B7
BnN
F-
YB
8B
nNF-
Y
B9
BnN
F-Y
B10
BnN
F-Y
B11
BnN
F-Y
B12
BnN
F-Y
B13
BnN
F-Y
B14
Am
ino
acid
iden
tify
for
cons
erve
d do
mai
n
BnN
F-Y
B2
–64
.063
.865
.765
.563
.858
.148
.346
.551
.451
.471
.849
.770
.6
BnN
F-Y
B2
82.1
–92
.272
.875
.670
.959
.141
.243
.749
.149
.456
.345
.759
.8
BnN
F-Y
B3
82.1
100.
10–
71.3
73.4
70.1
57.7
42.0
43.4
47.8
47.5
55.0
46.5
59.2
BnN
F-Y
B4
83.2
93.7
93.7
–95
.591
.058
.546
.346
.951
.552
.559
.350
.361
.6
BnN
F-Y
B5
81.1
93.7
93.7
96.8
–91
.758
.146
.848
.752
.253
.261
.351
.063
.2
BnN
F-Y
B6
82.1
94.7
94.7
97.9
98.9
–57
.343
.245
.151
.852
.159
.647
.362
.0
BnN
F-Y
B7
72.6
78.9
78.9
75.8
74.7
75.8
–42
.342
.359
.459
.754
.045
.353
.8
BnN
F-Y
B8
65.3
68.4
68.4
69.5
69.5
69.5
62.1
–84
.731
.834
.043
.151
.840
.2
BnN
F-Y
B9
63.8
70.2
70.2
71.3
71.3
71.3
63.8
98.9
–33
.234
.942
.156
.240
.9
BnN
F-Y
B10
75.8
84.2
84.2
89.5
87.4
88.4
76.8
63.2
64.9
–89
.849
.133
.049
.1
BnN
F-Y
B11
74.7
83.2
83.2
88.4
86.3
87.4
75.8
63.2
64.9
97.9
–50
.333
.748
.0
BnN
F-Y
B12
85.3
82.1
82.1
83.2
83.2
84.2
69.5
68.4
67.0
78.9
77.9
–41
.781
.4
BnN
F-Y
B13
63.2
66.3
66.3
67.4
67.4
67.4
62.1
83.2
84.0
65.3
63.2
61.1
–40
.5

116 Planta (2014) 239:107–126
1 3
Gene structures of BnNF-YBs and BnNF-YCs
The gene structures of all the BnNF-YB and BnNF-YC members resemble their putative homologs in Arabidop-sis. Particularly, the third exons of BnNF-YB1 and AtNF-YB1 are only six nucleotides long (i.e., 5′-TTGGAG-3′). All of the third exons of BnNF-YB12/14 and their putative Arabidopsis ortholog, AtNF-YB8/10, are six nucleotides long (i.e., 5′-ATGGAG-3′). The sequence of the last exon of BnNF-YC1, BnNF-YC3, and AtNF-YC4 is consistently 5′-TTGA-3′, while the corresponding exon sequence in AtNF-YC1 is 5′-TTAA-3′. In rice, as for OsHAP2, most of OsHAP3/5 has markedly different gene structures from NF-YB/C of Arabidopsis and canola (Supplementary Fig. S4).
In addition, all BnNF-Y introns, except for the last intron of BnNF-YA7, differ in length from the correspond-ing ones in the AtNF-Ys, which indicates that introns of the BnNF-Y and AtNF-Y genes may be under loose selective pressure. The similarity in gene structures of BnNF-Ys and their putative orthologs, AtNF-Ys, implies the conservation of gene functions between BnNF-Ys and AtNF-Ys. Most gene structures of OsHAPs are quite different from those of AtNF-Ys and BnNF-Ys, suggesting that plant NF-Ys diverged before the monocot–dicot split.
Tissue- and development-specific BnNF-Y expression patterns
To investigate the functions of BnNF-Y genes, we exam-ined their gene expression profiles. Systematic analysis of the expression patterns of members of the BnNF-Y fam-ily may reveal their physiological roles in different growth processes in B. napus. We performed quantitative real-time PCR (qRT-PCR) to quantify the relative transcription lev-els of BnNF-Y genes expressed in different developmental stages and tissues. To avoid cross-amplification among the different members of the BnNF-Y family, all primers used for qRT-PCR were designed based on regions outside the conserved domains. We then examined gene expression in leaves, stems, roots, inflorescences, and siliques of dif-ferent developmental stages for all 33 BnNF-Y members. The expression level of each gene in one of three leaves sampled at 75 DAP was arbitrarily set as 1.0. To compare gene expression levels between different NF-Ys in canola, expression level of every NF-Y gene in 75-DAP leaf sam-ples was further semi-quantified at two given cycles (30/35 cycles; Supplementary Fig. S5).
BnNF-YA expression
Most BnNF-YA members were detected in all tissues and had the highest expression at 202 DAP (Fig. 4). The a
Iden
tity
mat
rix
of th
e B
nNF-
YA
fam
ily, b
iden
tity
mat
rix
of th
e B
nNF-
YB
fam
ily, c
iden
tity
mat
rix
of th
e B
nNF-
YC
fam
ily
Tabl
e 2
con
tinue
d
(b)
Am
ino
acid
iden
tify
for
full-
leng
th p
rote
in
BnN
F-
YB
1B
nNF-
Y
B2
BnN
F-
YB
3B
ILN
F-
YB
4B
nNF-
Y
B5
BnN
F-
YB
6B
nNF-
Y
B7
BnN
F-
YB
8B
nNF-
Y
B9
BnN
F-Y
B10
BnN
F-Y
B11
BnN
F-Y
B12
BnN
F-Y
B13
BnN
F-Y
B14
BnN
F-Y
B14
82.1
86.3
86.3
85.3
85.3
86.3
71.6
67.4
68.1
80.0
80.0
91.6
62.1
–
(c)
Am
ino
acid
iden
tify
for
full-
leng
th p
rote
in
B11
NF-
YC
1B
nNF-
YC
2B
nNF-
YC
3B
nNF-
Y
C4
BnN
F-
YC
5
Am
ino
acid
iden
tify
for
cons
erve
d do
mai
n
BnN
F-Y
C1
–62
.589
.159
.559
.2
BnN
F-Y
C2
88.3
–58
.062
.663
.9
BnN
F-Y
C3
100.
088
.3–
55.5
54.1
BnN
F-Y
C4
89.6
90.9
89.6
–94
.7
BnN
F-Y
C5
89.6
90.9
89.6
100.
0–

117Planta (2014) 239:107–126
1 3
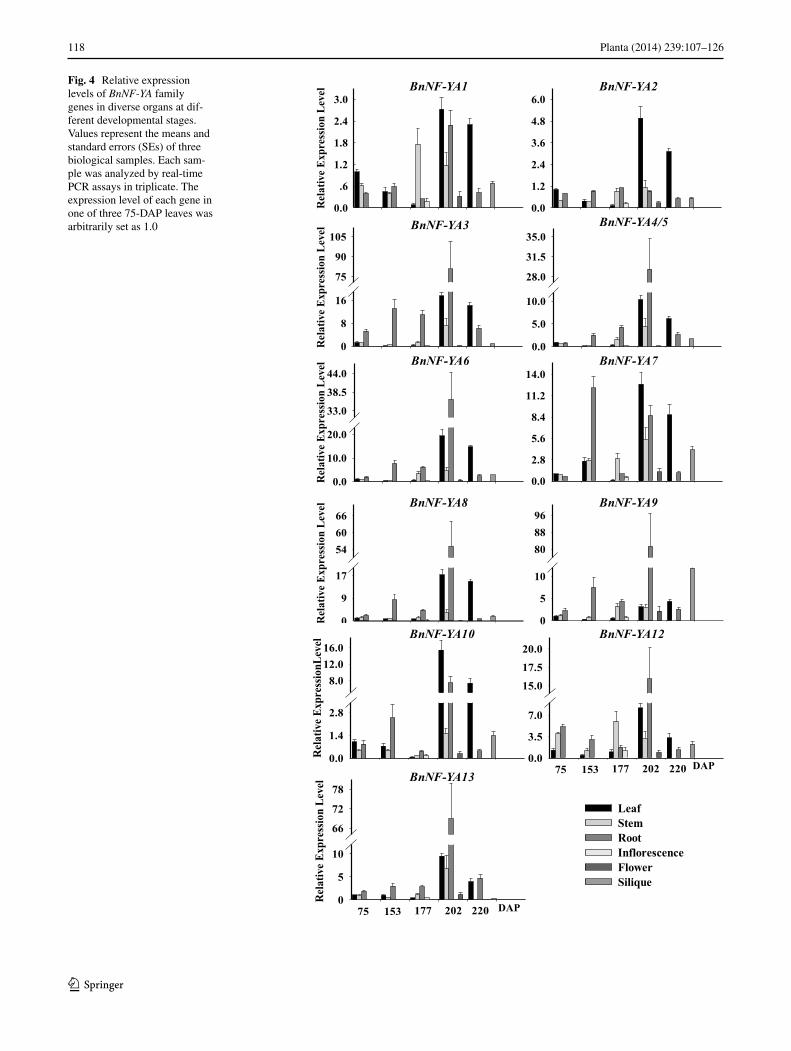
expression level of BnNF-YA1 and BnNF-YA2 decreased slowly before flowering (177 DAP) and then increased rapidly until flowering (202 DAP). The transcript level of BnNF-YA1 was relatively higher in the leaf, stem, and root samples at 202 DAP, while that of BnNF-YA2 was mostly detected in leaves at 202 DAP or 220 DAP. Whereas the expression of both BnNF-YA1 and BnNF-YA2 was elevated
in stem and root tissues at 202 DAP, there were no nota-ble changes in other organs or at different developmen-tal stages. Before 202 DAP, BnNF-YA3 expression was stronger in roots than in other organs at the same stage. BnNF-YA3 expression levels in leaves, stems, and roots at 202 DAP were higher than at other stages, and lev-els were highest in the roots (almost 80-fold higher than
Fig. 3 Exon/intron structure of BnNF-Ys and AtNF-Ys. Black boxes denote exons within coding regions and the lines connecting them represent introns. The length of boxes and lines represents the size (bp) of the corresponding exon and intron, respectively, as do the
numbers in black boxes or above the lines. The gene structures with-out the untranslated regions (UTRs) were constructed using the Gene Structure Display Serve tool (http://gsds.cbi.pku.edu.cn/)
118 Planta (2014) 239:107–126
1 3
Fig. 4 Relative expression levels of BnNF-YA family genes in diverse organs at dif-ferent developmental stages. Values represent the means and standard errors (SEs) of three biological samples. Each sam-ple was analyzed by real-time PCR assays in triplicate. The expression level of each gene in one of three 75-DAP leaves was arbitrarily set as 1.0

119Planta (2014) 239:107–126
1 3
baseline). BnNF-YA4/5 and BnNF-YA6 show similarly uni-form expression patterns. Their expression levels peaked in roots at 202 DAP, while levels were significantly increased in leaves at 202 or 220 DAP. BnNF-YA7 expression was strongest in the roots at 153 DAP and in the leaves at 202 DAP, while its expression levels were very low in other tis-sues at all stages, except for the stems, flowers, and roots at 220 DAP. BnNF-YA8 and BnNF-YA9 showed similar expression patterns, but expression differed in leaves at 202 DAP and in siliques at 220 DAP. The expression of these genes was highest in roots at 202 DAP. BnNF-YA10 expression was strongest in leaves at 202 DAP, and low-est in all tissues at 177 DAP. BnNF-YA12 transcripts, which were most abundant in roots at 202 DAP, were detected in all organs, while BnNF-YA11 transcripts were not detected in any organ at any developmental stage analyzed (data not shown). BnNF-YA13 expression was higher in all organs at 202 DAP than at any other stage. The transcript level of BnNF-YA14 was so low that it was not detectable in any organ at any developmental stage analyzed (data not shown).
BnNF-YB expression
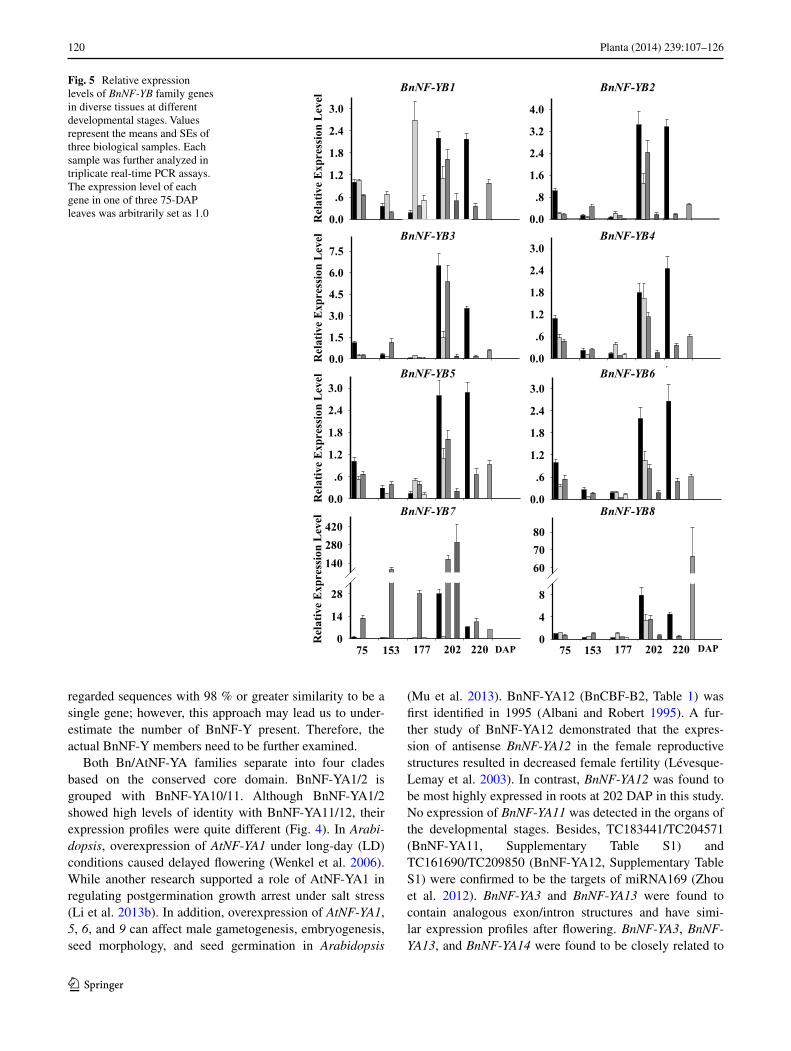
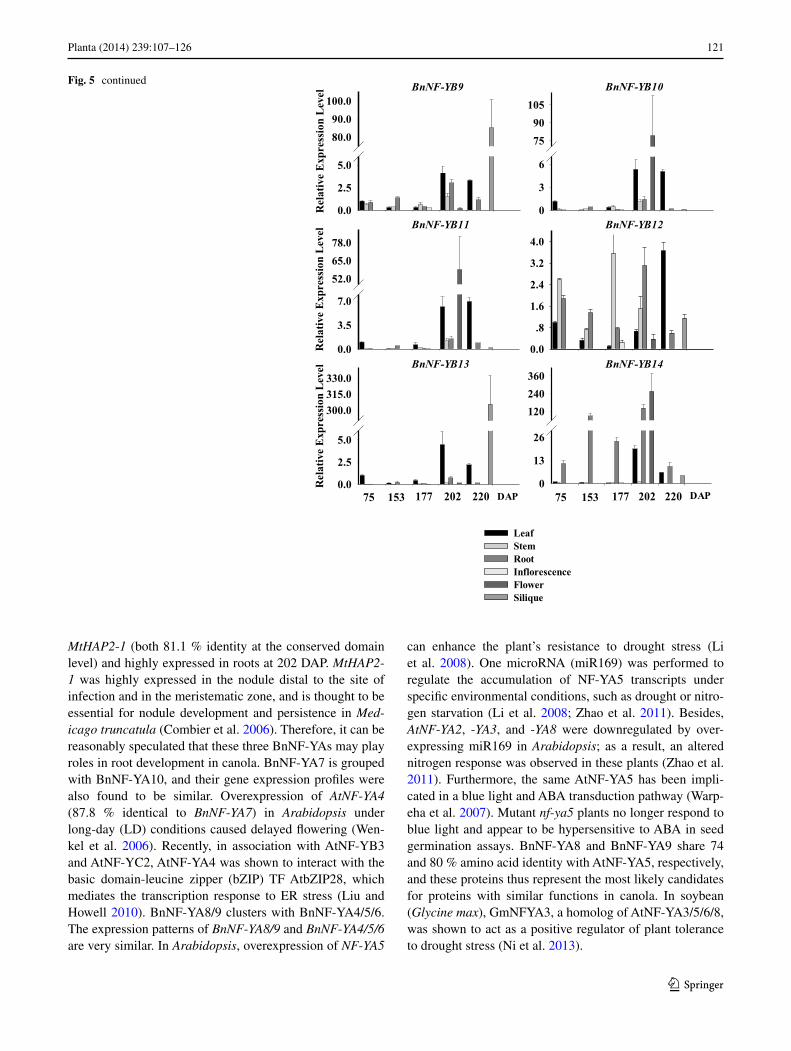
BnNF-YB1 transcript levels decreased slightly before flow-ering and then increased in leaves, and were relatively high in stems at 177 DAP (Fig. 5). BnNF-YB2 and BnNF-YB3 had similar expression profiles, as did BnNF-YB4, BnNF-YB5, and BnNF-YB6. Expression levels of BnNF-YB4/5/6 were modestly up-regulated in leaves at 202 or 220 DAP. Similar to BnNF-YB1, the expression levels of these five genes were mildly down-regulated in leaves before flower-ing. Low levels of BnNF-YB7 were detected in stems, inflo-rescences, and siliques, while high levels were found in flowers. Expression of this transcript was variable in roots throughout the growth process, but remained relatively high compared to that in other tissues. The expression of BnNF-YB8, BnNF-YB9, and BnNF-YB13 was higher in siliques than in other tissues, and almost absent at 75, 153, or 177 DAP, respectively. BnNF-YB10 and BnNF-YB11 transcripts were most abundant in flowers. BnNF-YB12 was ubiqui-tously expressed in all tissues examined, and their levels decreased before flowering and then gradually increased in leaves. BnNF-YB14 was strongly expressed in roots at all stages before the mature period and had a similar expres-sion profile as BnNF-YB5.
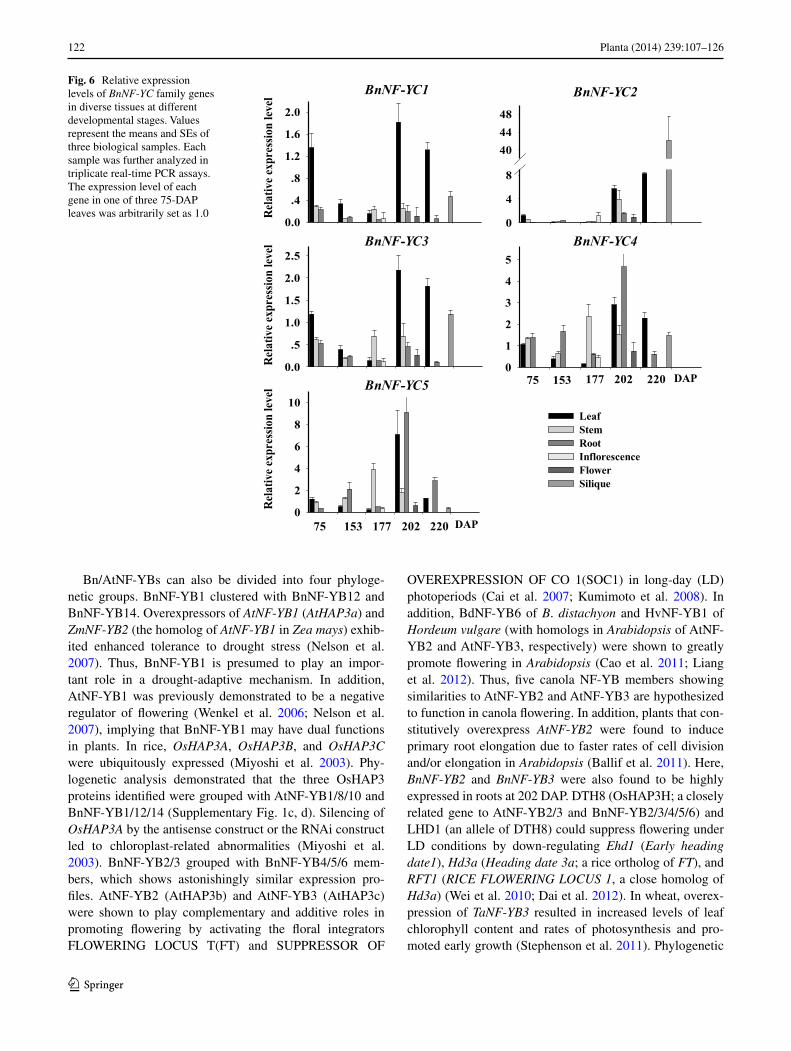
BnNF-YC expression
BnNF-YC1 and BnNF-YC3 were closely related and had similar expression patterns (Fig. 6). The expression lev-els of BnNF-YC1 and BnNF-YC3 were low at all stages, with the exception of leaf tissue. BnNF-YC2 expression
appeared to be lower in the first three stages analyzed than at other stages. During flowering and the developing silique stage (202–220 days), BnNF-YC2 had relatively high lev-els of expression. The transcript level of BnNF-YC2 was most abundant in siliques during the whole growth sea-son. BnNF-YC1 and BnNF-YC3 were strongly induced in leaves. In contrast, BnNF-YC4 and BnNF-YC5 expression levels were higher in roots at 202 DAP than in other tissues.
Discussion
Multi-gene NF-Y families in plants have recently generated much interest due to their important roles in plant devel-opment or stress regulation. In contrast to yeast and ani-mal systems, plants have evolved multiple genes for each NF-Y subfamily. In this study, we cloned 14 BnNF-YAs, 14 BnNF-YBs, and 5 BnNF-YCs. Since the genome sequence of oilseed rape is not available, the number of BnNF-Ys identified is likely an underestimate. Brassica napus is an allotetraploid species with 2n = 38 chromosomes and two genomes (AA derived from B. rapa and CC from B. olera-cea). Given that the B. rapa genome, which has been fully sequenced (Wang et al. 2011), has 19 NF-YAs, 20 NF-YBs, and 6 NF-YCs based on our preliminary bioinformatics analysis (Supplementary Table S3), we conclude that the number of NF-Ys in the canola genome could indeed be higher than that reported here. Interestingly, five of the pre-dicted AtNF-Ys, that is, AtNF-YC5, AtNF-YC6, AtNF-YC7, AtNF-YC8, and AtNF-YC12, seem not to have correspond-ing homologs in the B. rapa genome (Supplementary Table S3). Although VRN2 of wheat can interact with Arabidop-sis NF-YC7 to repress flowering in vitro (Li et al. 2011), the functions of the above-mentioned NF-YC members are largely unknown. Why B. rapa contains around twice as many AtNF-YA/B homologs as the number of AtNF-YA/B proteins present in Arabidopsis, but fewer AtNF-YC homologs is unknown. Currently, the oil rape EST database contains more than 82,000 ESTs, which is even fewer than the number of Arabidopsis ESTs present in the DFCI database (more than 110,000). Some canola NF-Y transcripts may not be represented in the current database. However, considering that the hexaploid B. rapa contains around 41,000 genes as compared to the 30,000 present in Arabidopsis, the number of NF-Ys in B. napus may not be double that of B. rapa NF-Y members, due to genome loss during evolution. Even though canola was the product of crosses between A and C owner genomes in Brassica, extensive gene loss and fractionation of the genome were suggested to occur, and indeed was also observed in Arabi-dopsis itself. Arabidopsis thaliana is predicted to have undergone a 30 % reduction in genome size as compared to ancient A. lyrata. In our analysis, for simplification, we
120 Planta (2014) 239:107–126
1 3
regarded sequences with 98 % or greater similarity to be a single gene; however, this approach may lead us to under-estimate the number of BnNF-Y present. Therefore, the actual BnNF-Y members need to be further examined.
Both Bn/AtNF-YA families separate into four clades based on the conserved core domain. BnNF-YA1/2 is grouped with BnNF-YA10/11. Although BnNF-YA1/2 showed high levels of identity with BnNF-YA11/12, their expression profiles were quite different (Fig. 4). In Arabi-dopsis, overexpression of AtNF-YA1 under long-day (LD) conditions caused delayed flowering (Wenkel et al. 2006). While another research supported a role of AtNF-YA1 in regulating postgermination growth arrest under salt stress (Li et al. 2013b). In addition, overexpression of AtNF-YA1, 5, 6, and 9 can affect male gametogenesis, embryogenesis, seed morphology, and seed germination in Arabidopsis
(Mu et al. 2013). BnNF-YA12 (BnCBF-B2, Table 1) was first identified in 1995 (Albani and Robert 1995). A fur-ther study of BnNF-YA12 demonstrated that the expres-sion of antisense BnNF-YA12 in the female reproductive structures resulted in decreased female fertility (Lévesque-Lemay et al. 2003). In contrast, BnNF-YA12 was found to be most highly expressed in roots at 202 DAP in this study. No expression of BnNF-YA11 was detected in the organs of the developmental stages. Besides, TC183441/TC204571 (BnNF-YA11, Supplementary Table S1) and TC161690/TC209850 (BnNF-YA12, Supplementary Table S1) were confirmed to be the targets of miRNA169 (Zhou et al. 2012). BnNF-YA3 and BnNF-YA13 were found to contain analogous exon/intron structures and have simi-lar expression profiles after flowering. BnNF-YA3, BnNF-YA13, and BnNF-YA14 were found to be closely related to
Fig. 5 Relative expression levels of BnNF-YB family genes in diverse tissues at different developmental stages. Values represent the means and SEs of three biological samples. Each sample was further analyzed in triplicate real-time PCR assays. The expression level of each gene in one of three 75-DAP leaves was arbitrarily set as 1.0
121Planta (2014) 239:107–126
1 3
MtHAP2-1 (both 81.1 % identity at the conserved domain level) and highly expressed in roots at 202 DAP. MtHAP2-1 was highly expressed in the nodule distal to the site of infection and in the meristematic zone, and is thought to be essential for nodule development and persistence in Med-icago truncatula (Combier et al. 2006). Therefore, it can be reasonably speculated that these three BnNF-YAs may play roles in root development in canola. BnNF-YA7 is grouped with BnNF-YA10, and their gene expression profiles were also found to be similar. Overexpression of AtNF-YA4 (87.8 % identical to BnNF-YA7) in Arabidopsis under long-day (LD) conditions caused delayed flowering (Wen-kel et al. 2006). Recently, in association with AtNF-YB3 and AtNF-YC2, AtNF-YA4 was shown to interact with the basic domain-leucine zipper (bZIP) TF AtbZIP28, which mediates the transcription response to ER stress (Liu and Howell 2010). BnNF-YA8/9 clusters with BnNF-YA4/5/6. The expression patterns of BnNF-YA8/9 and BnNF-YA4/5/6 are very similar. In Arabidopsis, overexpression of NF-YA5
can enhance the plant’s resistance to drought stress (Li et al. 2008). One microRNA (miR169) was performed to regulate the accumulation of NF-YA5 transcripts under specific environmental conditions, such as drought or nitro-gen starvation (Li et al. 2008; Zhao et al. 2011). Besides, AtNF-YA2, -YA3, and -YA8 were downregulated by over-expressing miR169 in Arabidopsis; as a result, an altered nitrogen response was observed in these plants (Zhao et al. 2011). Furthermore, the same AtNF-YA5 has been impli-cated in a blue light and ABA transduction pathway (Warp-eha et al. 2007). Mutant nf-ya5 plants no longer respond to blue light and appear to be hypersensitive to ABA in seed germination assays. BnNF-YA8 and BnNF-YA9 share 74 and 80 % amino acid identity with AtNF-YA5, respectively, and these proteins thus represent the most likely candidates for proteins with similar functions in canola. In soybean (Glycine max), GmNFYA3, a homolog of AtNF-YA3/5/6/8, was shown to act as a positive regulator of plant tolerance to drought stress (Ni et al. 2013).
Fig. 5 continued
122 Planta (2014) 239:107–126
1 3
Bn/AtNF-YBs can also be divided into four phyloge-netic groups. BnNF-YB1 clustered with BnNF-YB12 and BnNF-YB14. Overexpressors of AtNF-YB1 (AtHAP3a) and ZmNF-YB2 (the homolog of AtNF-YB1 in Zea mays) exhib-ited enhanced tolerance to drought stress (Nelson et al. 2007). Thus, BnNF-YB1 is presumed to play an impor-tant role in a drought-adaptive mechanism. In addition, AtNF-YB1 was previously demonstrated to be a negative regulator of flowering (Wenkel et al. 2006; Nelson et al. 2007), implying that BnNF-YB1 may have dual functions in plants. In rice, OsHAP3A, OsHAP3B, and OsHAP3C were ubiquitously expressed (Miyoshi et al. 2003). Phy-logenetic analysis demonstrated that the three OsHAP3 proteins identified were grouped with AtNF-YB1/8/10 and BnNF-YB1/12/14 (Supplementary Fig. 1c, d). Silencing of OsHAP3A by the antisense construct or the RNAi construct led to chloroplast-related abnormalities (Miyoshi et al. 2003). BnNF-YB2/3 grouped with BnNF-YB4/5/6 mem-bers, which shows astonishingly similar expression pro-files. AtNF-YB2 (AtHAP3b) and AtNF-YB3 (AtHAP3c) were shown to play complementary and additive roles in promoting flowering by activating the floral integrators FLOWERING LOCUS T(FT) and SUPPRESSOR OF
OVEREXPRESSION OF CO 1(SOC1) in long-day (LD) photoperiods (Cai et al. 2007; Kumimoto et al. 2008). In addition, BdNF-YB6 of B. distachyon and HvNF-YB1 of Hordeum vulgare (with homologs in Arabidopsis of AtNF-YB2 and AtNF-YB3, respectively) were shown to greatly promote flowering in Arabidopsis (Cao et al. 2011; Liang et al. 2012). Thus, five canola NF-YB members showing similarities to AtNF-YB2 and AtNF-YB3 are hypothesized to function in canola flowering. In addition, plants that con-stitutively overexpress AtNF-YB2 were found to induce primary root elongation due to faster rates of cell division and/or elongation in Arabidopsis (Ballif et al. 2011). Here, BnNF-YB2 and BnNF-YB3 were also found to be highly expressed in roots at 202 DAP. DTH8 (OsHAP3H; a closely related gene to AtNF-YB2/3 and BnNF-YB2/3/4/5/6) and LHD1 (an allele of DTH8) could suppress flowering under LD conditions by down-regulating Ehd1 (Early heading date1), Hd3a (Heading date 3a; a rice ortholog of FT), and RFT1 (RICE FLOWERING LOCUS 1, a close homolog of Hd3a) (Wei et al. 2010; Dai et al. 2012). In wheat, overex-pression of TaNF-YB3 resulted in increased levels of leaf chlorophyll content and rates of photosynthesis and pro-moted early growth (Stephenson et al. 2011). Phylogenetic
Fig. 6 Relative expression levels of BnNF-YC family genes in diverse tissues at different developmental stages. Values represent the means and SEs of three biological samples. Each sample was further analyzed in triplicate real-time PCR assays. The expression level of each gene in one of three 75-DAP leaves was arbitrarily set as 1.0

123Planta (2014) 239:107–126
1 3
analysis indicated that TaNF-YB3 was closely related to At/BnNF-YB2 and At/BnNF-YB3 (>92.5 % identity in the conserved domain). Therefore, the five BnNF-YB2/3/4/5/6 genes may play similar roles in photosynthesis. BnNF-YB8 and BnNF-YB9 are grouped with BnNF-YB13 and have similar expression profiles. A previous study indi-cated that LEC1 (AtNF-YB9) regulates late embryogenesis (West et al. 1994). Following this report, numerous stud-ies have focused on LEC1 and its related LEC1-like gene (L1L, NF-YB6), and clearly show that these two genes play essential, but complementary roles during embry-onic development (Lotan et al. 1998; Kwong et al. 2003; Lee et al. 2003; Yazawa and Kamada 2007; Alemanno et al. 2008). In this study, BnNF-YB8/9 and BnNF-YB13 had the highest levels of expression in the mature siliques of all tissues examined, while levels of expression were very low at other stages. Furthermore, NF-YB6 and NF-YB9 were both found to inhibit the ABA-mediated delay in seed germination (Warpeha et al. 2007), while NF-YB9 was also assigned roles in activating the transcription of LHCB (light-harvesting chlorophyll a/b-binding protein) (Warpeha et al. 2007) and mediating the up-regulation of the auxin synthesis gene YUCCA10 (Junker et al. 2012). In addition, in rice, OsL1L (OsHAP3E) was shown to affect the development of leaves, panicles, spikelets, and flow-ers, and to regulate meristem identity determination during both vegetative and reproductive development (Ito et al. 2011). BnNF-YB10 and BnNF-YB11 transcripts were most abundant in flowers, which suggesting that the two genes may have functions related to flower.
Two major clades are identified in the Bn/AtNF-YC families. The five identified BnNF-YCs are grouped with AtNF-YC1/2/3/4/9 (Fig. 1c; Supplementary Fig. 1c). In Arabidopsis, AtNF-YC1, -YC4, and -YC9 were shown to be expressed in 6-day-old etiolated seedlings (Warpeha et al. 2007). Further assays showed that AtNF-YC4 was involved in inhibiting the ABA-mediated delay in seed germination. Previous researches showed that AtNF-YC2 was normally expressed only at low levels during seedling development, but up-regulated during seed development (Siefers et al. 2009; Liu and Howell 2010). In our study, the BnNF-YC2 transcript was detected at very high levels in siliques. Considering that plant NF-YA/B/C subunits may need to form a complex to be functional, BnNF-YC2 is an excellent candidate for a protein with a role in seed devel-opment. Recently, it was reported that overexpression of Arabidopsis NF-YC1 or NF-YC2 resulted in early flowering (Hackenberg et al. 2012). It appears that these genes pro-moted flowering through FT, since the FT transcript level was increased in transgenic plants overexpressing NF-YC1 or NF-YC2. Direct evidence for a role for NF-YC in flower-ing time control comes from the nf-yc3 nf-yc4 nf-yc9 triple mutant in Arabidopsis, which showed significantly delayed
flowering under LD conditions (Kumimoto et al. 2010). These three most closely related BnNF-YC members may function as their Arabidopsis homologs in promoting flow-ering. In wheat, TaNF-YC11, one ortholog of AtNF-YC1/4, was shown to be a potential regulator of photosynthesis-related genes (Stephenson et al. 2007, 2010). Although BnNF-YC1 and BnNF-YC3, two homologs of AtNF-YC1/4, were highly expressed in leaves at 75, 202, and 220 DAP, it remains to be determined if BnNF-YC1 and BnNF-YC3 have similar functions. In Picea wilsonii, PwHAP5, which has high levels of sequence identity with AtNF-YC3/9, was shown to be strongly expressed in germinating pollen and to play a role in determining the orientation of pollen tube growth (Yu et al. 2011). A following research revealed that PwHAP5 plays positive roles in response to salinity, osmotic and ABA stress at different developmental stages in plants, respectively, via possibly regulating stress-related genes (Li et al. 2013a). In overexpression and RNAi exper-iments in Phaseolus vulgaris, PvNF-YC1 was shown to positively regulate nodule development, possibly by acti-vating G2-M cell cycle genes, and to have a positive influ-ence on the infection process of bean roots by symbiotic Rhizobia (Zanetti et al. 2010). Sequence analysis indicated that PvNF-YC1 displayed 67.0, 64.1, 67.4, and 66.2 % amino acid identities with AtNF-YC3, AtNF-YC9, BnNF-YC4, and BnNF-YC5 at the full-length protein level, respectively. Furthermore, PvNF-YC1 was found to have 97.4 % sequence identity with all of the above-mentioned four genes at the conserved domain level. In this study, both BnNF-YC4 and BnNF-YC5, two paralogs of AtNF-YC3/9, were shown to be highly expressed in roots at 202 DAP.
The finding that the BnNF-YA/B/C subunits have similar expression profiles may suggest that they potentially form a regulatory complex. For example, BnNF-YC2 was shown to have similar expression profiles to BnNF-YB8/9/13. BnNF-YB9 (L1L) and BnNF-YB13 (LEC1) were found to play important roles in regulating the lipid pathway (Mu et al. 2008; Shen et al. 2010; Tan et al. 2011). Thus, BnNF-YC2 and BnNF-YB8/9/13 are excellent candidates for mol-ecules with a role in seed development.
Conclusions
This study provides the first extensive evaluation of the BnNF-Y family by characterizing the phylogenetic rela-tionships, exon/intron structures, and spatial–temporal expression patterns of its members. The results presented here could offer a useful foundation for dissecting the func-tions of BnNF-Y (We sum up some reported plant NF-Ys function in Supplementary Table S4). Several subunits in every family showed not only structural analogies, but also

124 Planta (2014) 239:107–126
1 3
similar expression profiles, which suggests redundancy among these genes. Members of different families were found to have similar expression patterns, which may pro-vide a basis for determining if those BnNF-Ys could form a complex. Plants, as sessile organisms, need dynamic devel-opmental strategies to respond to external environments. The presence of multiple BnNF-Y members may generate flexibility in regulatory mechanisms and thereby increase the chance of survival.
Acknowledgments We thank Prof. Zhaopu Liu from Nanjing Agricultural University for kindly supplying the canola (Nanyanyou 1) seeds. We are grateful to Dr. Zengrong Huang and Ms Jin Wang from Nanjing Agricultural University for advice for qRT-PCR. We also thank Kathleen Farquharson for valuable comments on the manuscript revision. This research was supported by grants from the Natural Science Foundation of Jiangsu province (BK2011635), the State Key Laboratory of Crop Genetics and Germplasm Enhance-ment (ZW2010004), a China Postdoctoral Science Foundation-funded project (20110491441), a Jiangsu Postdoctoral Science Foundation-funded project (1101013B), Fundamental Research Funds for the Central Universities (KYZ201206) and the Priority Academic Pro-gram Development of Jiangsu Higher Education Institutions (RAPD program (809001), and the Technological Innovation Foundation for Young Scientists of Nanjing Agricultural University (Y201058).
Conflict of interest The authors declare that they have no compet-ing interests.
References
Albani D, Robert LS (1995) Cloning and characterization of a Bras-sica napus gene encoding a homologue of the B subunit of a het-eromeric CCAAT-binding factor. Gene 167:209–213
Alemanno L, Devic M, Niemenak N, Sanier C, Guilleminot J, Rio M, Verdeil JL, Montoro P (2008) Characterization of leafy cotyle-don1-like during embryogenesis in Theobroma cacao L. Planta 227:853–866
Altschul SF, Gish W, Miller W, Myers EW, Lipman DJ (1990) Basic local alignment search tool. J Mol Biol 215:403–410
Ballif J, Endo S, Kotani M, MacAdam J, Wu Y (2011) Over-expres-sion of HAP3b enhances primary root elongation in Arabidopsis. Plant Physiol Biochem 49:579–583
Becker DM, Fikes JD, Guarente L (1991) A cDNA-encoding a human CCAAT-binding protein cloned by functional complementation in yeast. Proc Natl Acad Sci USA 88:1968–1972
Cai X, Ballif J, Endo S, Davis E, Liang M, Chen D, Dewald D, Kreps J, Zhu T, Wu Y (2007) A putative CCAAT-binding transcription factor is a regulator of flowering timing in Arabidopsis. Plant Physiol 145:98–105
Cao S, Kumimoto RW, Siriwardana CL, Risinger JR, Holt III BF (2011) Identification and characterization of NF-Y transcription factor families in the monocot model plant Brachypodium dis-tachyon. PloS ONE 6:e21805
Combier JP, Frugier F, de Billy F, Boualem A, El-Yahyaoui F, Moreau S, Vernie T, Ott T, Gamas P, Crespi M, Niebel A (2006) MtHAP2-1 is a key transcriptional regulator of symbiotic nodule develop-ment regulated by microRNA169 in Medicago truncatula. Genes Dev 20:3084–3088
Dai X, Ding Y, Tan L, Fu Y, Liu F, Zhu Z, Sun X, Sun X, Gu P, Cai H (2012) LHD1, an allele of DTH8/Ghd8, controls late heading
date in common wild rice (Oryza rufipogon). J Integr Plant Biol 54:790–799
Edwards D, Murray JAH, Smith AG (1998) Multiple genes encod-ing the conserved CCAAT-box transcription factor complex are expressed in Arabidopsis. Plant Physiol 117:1015–1022
Focks N, Benning C (1998) wrinkled1: a novel, low-seed-oil mutant of Arabidopsis with a deficiency in the seed-specific regulation of carbohydrate metabolism. Plant Physiol 118:91–101
Gusmaroli G, Tonelli C, Mantovani R (2001) Regulation of the CCAAT-binding NF-Y subunits in Arabidopsis thaliana. Gene 264:173–185
Gusmaroli G, Tonelli C, Mantovani R (2002) Regulation of novel members of the Arabidopsis thaliana CCAAT-binding nuclear factor Y subunits. Gene 283:41–48
Hackenberg D, Keetman U, Grimm B (2012) Homologous NF-YC2 subunit from Arabidopsis and tobacco is activated by photooxi-dative stress and induces flowering. Int J Mol Sci 13:3458–3477
Hahn S, Guarente L (1988) Yeast HAP2 and HAP3: transcriptional activators in a heteromeric complex. Science 240:317–321
Hooft van Huijsduijnen R, Li XY, Black D, Matthes H, Benoist C, Mathis D (1990) Co-evolution from yeast to mouse: cDNA clon-ing of the two NF-Y (CP-1/CBF) subunits. EMBO J 9:3119–3127
Ito Y, Thirumurugan T, Serizawa A, Hiratsu K, Ohme-Takagi M, Kurata N (2011) Aberrant vegetative and reproductive develop-ment by overexpression and lethality by silencing of OsHAP3E in rice. Plant Sci 181:105–110
Junker A, Mönke G, Rutten T, Keilwagen J, Seifert M, Thi TMN, Renou JP, Balzergue S, Viehöver P, Hähnel U (2012) Elongation-related functions of LEAFY COTYLEDON1 during the develop-ment of Arabidopsis thaliana. Plant J 71:427–442
Kim IS, Sinha S, de Crombrugghe B, Maity SN (1996) Determina-tion of functional domains in the C subunit of the CCAAT-bind-ing factor (CBF) necessary for formation of a CBF-DNA com-plex: CBF-B interacts simultaneously with both the CBF-A and CBF-C subunits to form a heterotrimeric CBF molecule. Mol Cell Biol 16:4003–4013
Kreps JA, Wu YJ, Chang HS, Zhu T, Wang X, Harper JF (2002) Tran-scriptome changes for Arabidopsis in response to salt, osmotic, and cold stress. Plant Physiol 130:2129–2141
Kumimoto RW, Adam L, Hymus GJ, Repetti PP, Reuber TL, Marion CM, Hempel FD, Ratcliffe OJ (2008) The nuclear factor Y subu-nits NF-YB2 and NF-YB3 play additive roles in the promotion of flowering by inductive long-day photoperiods in Arabidopsis. Planta 228:709–723
Kumimoto RW, Zhang Y, Siefers N, Holt BF III (2010) NF-YC3, NF-YC4 and NF-YC9 are required for CONSTANS-mediated, photoperiod-dependent flowering in Arabidopsis thaliana. Plant J 63:379–391
Kwong RW, Bui AQ, Lee H, Kwong LW, Fischer RL, Goldberg RB, Harada JJ (2003) LEAFY COTYLEDON1-LIKE defines a class of regulators essential for embryo development. Plant Cell 15:5–18
Lee H, Fischer RL, Goldberg RB, Harada JJ (2003) Arabidopsis LEAFY COTYLEDON1 represents a functionally specialized subunit of the CCAAT binding transcription factor. Proc Natl Acad Sci USA 100:2152–2156
Lévesque-Lemay M, Albani D, Aldcorn D, Hammerlindl J, Keller W, Robert LS (2003) Expression of CCAAT-binding factor antisense transcripts in reproductive tissues affects plant fertility. Plant Cell Rep 21:804–808
Li WX, Oono Y, Zhu JH, He XJ, Wu JM, Iida K, Lu XY, Cui XP, Jin HL, Zhu JK (2008) The Arabidopsis NFYA5 transcription factor is regulated transcriptionally and posttranscriptionally to promote drought resistance. Plant Cell 20:2238–2251
Li CX, Distelfeld A, Comis A, Dubcovsky J (2011) Wheat flowering repressor VRN2 and promoter CO2 compete for interactions with NUCLEAR FACTOR-Y complexes. Plant J 67:763–773

125Planta (2014) 239:107–126
1 3
Li L, Yu Y, Wei J, Huang G, Zhang D, Liu Y, Zhang L (2013a) Homol-ogous HAP5 subunit from Picea wilsonii improved tolerance to salt and decreased sensitivity to ABA in transformed Arabidopsis. Planta, pp 1–12
Li Y-J, Fang Y, Fu Y-R, Huang J-G, Wu C-A, Zheng C–C (2013b) NFYA1 is involved in regulation of postgermination growth arrest under salt stress in Arabidopsis. PLoS ONE 8:e61289
Liang M, Hole D, Wu J, Blake T, Wu Y (2012) Expression and func-tional analysis of NUCLEAR FACTOR-Y, subunit B genes in barley. Planta, pp 1–13
Liu J-X, Howell SH (2010) bZIP28 and NF-Y transcription factors are activated by ER stress and assemble into a transcriptional complex to regulate stress response genes in Arabidopsis. Plant Cell 22:782–796
Lotan T, Ohto M, Yee KM, West MAL, Lo R, Kwong RW, Yamagi-shi K, Fischer RL, Goldberg RB, Harada JJ (1998) Arabidopsis LEAFY COTYLEDON1 is sufficient to induce embryo develop-ment in vegetative cells. Cell 93:1195–1205
Maity SN, de Crombrugghe B (1992) Biochemical analysis of the B subunit of the heteromeric CCAAT-binding factor. J Biol Chem 267:8286–8292
Maity SN, de Crombrugghe B (1998) Role of the CCAAT-binding pro-tein CBF/NF-Y in transcription. Trends Biochem Sci 23:174–178
Maity SN, Vuorio T, de Crombrugghe B (1990) The B-subunit of a rat heteromeric CCAAT-binding transcription factor shows a striking sequence identity with the yeast HAP2 transcription factor. Proc Nati Acad Sci USA 87:5378–5382
Mantovani R (1999) The molecular biology of the CCAAT-binding factor NF-Y. Gene 239:15–27
Mantovani R, Li X-Y, Pessara U, van Huisjduijnen RH, Benoist C, Mathis D (1994) Dominant negative analogs of NF-YA. J Biol Chem 269:20340–20346
McNabb DS, Xing YY, Guarente L (1995) Cloning of yeast HAP5: a novel subunit of a heterotrimeric complex required for CCAAT binding. Genes Dev 9:47–58
McNabb DS, Tseng K, Guarente L (1997) The Saccharomyces cer-evisiae Hap5p homolog from fission yeast reveals two conserved domains that are essential for assembly of heterotetrameric CCAAT-binding factor. Mol Cell Biol 17:7008–7018
Miyoshi K, Ito Y, Serizawa A, Kurata N (2003) OsHAP3 genes regu-late chloroplast biogenesis in rice. Plant J 36:532–540
Mu JY, Tan HL, Zheng Q, Fu FY, Liang Y, Zhang JA, Yang XH, Wang T, Chong K, Wang XJ, Zuo JR (2008) LEAFY COTYLEDON1 is a key regulator of fatty acid biosynthesis in Arabidopsis. Plant Physiol 148:1042–1054
Mu J, Tan H, Hong S, Liang Y, Zuo J (2013) Arabidopsis transcription factor genes NF-YA1, 5, 6, and 9 play redundant roles in male gametogenesis, embryogenesis, and seed development. Mol Plant 6:188–201
Nelson DE, Repetti PP, Adams TR, Creelman RA, Wu J, Warner DC, Anstrom DC, Bensen RJ, Castiglioni PP, Donnarummo MG, Hinchey BS, Kumimoto RW, Maszle DR, Canales RD, Kro-likowski KA, Dotson SB, Gutterson N, Ratcliffe OJ, Heard JE (2007) Plant nuclear factor Y (NF-Y) B subunits confer drought tolerance and lead to improved corn yields on water-limited acres. Proc Natl Acad Sci USA 104:16450–16455
Ni Z, Hu Z, Jiang Q, Zhang H (2013) GmNFYA3, a target gene of miR169, is a positive regulator of plant tolerance to drought stress. Plant Mol Biol 82:113–129
Petroni K, Kumimoto RW, Gnesutta N, Calvenzani V, Fornari M, Tonelli C, Holt BF III, Mantovani R (2012) The promiscuous life of plant NUCLEAR FACTOR Y transcription factors. Plant Cell 24:4777–4792
Rogers S, Bendich A (1985) Extraction of DNA from milligram amounts of fresh, herbarium and mummified plant tissues. Plant Mol Biol 5:69–76
Romier C, Cocchiarella F, Mantovani R, Moras D (2003) The NF-YB/NF-YC structure gives insight into DNA binding and tran-scription regulation by CCAAT factor NF-Y. J Biol Chem 278:1336–1345
Shen B, Allen WB, Zheng P, Li C, Glassman K, Ranch J, Nubel D, Tarczynski MC (2010) Expression of ZmLEC1 and ZmWRI1 increases seed oil production in maize. Plant Physiol 153:980–987
Siefers N, Dang KK, Kumimoto RW, Bynum WE, Tayrose G, Holt BF III (2009) Tissue-specific expression patterns of Arabidopsis NF-Y transcription factors suggest potential for extensive combi-natorial complexity. Plant Physiol 149:625–641
Sinha S, Kim IS, Sohn KY, de Crombrugghe B, Maity SN (1996) Three classes of mutations in the a subunit of the CCAAT-binding factor CBF delineate functional domains involved in the three-step assembly of the CBF-DNA complex. Mol Cell Biol 16:328–337
Stephenson TJ, McIntyre CL, Collet C, Xue GP (2007) Genome-wide identification and expression analysis of the NF-Y family of tran-scription factors in Triticum aestivum. Plant Mol Biol 65:77–92
Stephenson TJ, McIntyre CL, Collet C, Xue GP (2010) TaNF-YC11, one of the light-upregulated NF-YC members in Triticum aesti-vum, is co-regulated with photosynthesis-related genes. Funct Integr Genomics 10:265–276
Stephenson TJ, McIntyre CL, Collet C, Xue GP (2011) TaNF-YB3 is involved in the regulation of photosynthesis genes in Triticum aestivum. Funct Integr Genomics 11:327–340
Stone SL, Kwong LW, Yee KM, Pelletier J, Lepiniec L, Fischer RL, Goldberg RB, Harada JJ (2001) LEAFY COTYLEDON2 encodes a B3 domain transcription factor that induces embryo develop-ment. Proc Nati Acad Sci USA 98:11806–11811
Tan HL, Yang XH, Zhang FX, Zheng X, Qu CM, Mu JY, Fu FY, Li JA, Guan RZ, Zhang HS, Wang GD, Zuo JR (2011) Enhanced seed oil production in canola by conditional expression of Bras-sica napus LEAFY COTYLEDON1 and LEC1-LIKE in develop-ing seeds. Plant Physiol 156:1577–1588
Thirumurugan T, Ito Y, Kubo T, Serizawa A, Kurata N (2008) Identi-fication, characterization and interaction of HAP family genes in rice. Mol Genet Genomics 279:279–289
Thompson JD, Gibson TJ, Plewniak F, Jeanmougin F, Higgins DG (1997) The CLUSTAL_X windows interface: flexible strategies for multiple sequence alignment aided by quality analysis tools. Nucl Acids Res 25:4876–4882
Wang X, Wang H, Wang J, Sun R, Wu J, Liu S, Bai Y, Mun J-H, Ban-croft I, Cheng F (2011) The genome of the mesopolyploid crop species Brassica rapa. Nat Genet 43:1035–1039
Warpeha KM, Upadhyay S, Yeh J, Adamiak J, Hawkins SI, Lapik YR, Anderson MB, Kaufman LS (2007) The GCR1, GPA1, PRN1, NF-Y signal chain mediates both blue light and abscisic acid responses in Arabidopsis. Plant Physiol 143:1590–1600
Wei X, Xu J, Guo H, Jiang L, Chen S, Yu C, Zhou Z, Hu P, Zhai H, Wan J (2010) DTH8 suppresses flowering in rice, influencing plant height and yield potential simultaneously. Plant Physiol 153:1747–1758
Wenkel S, Turck F, Singer K, Gissot L, Le Gourrierec J, Samach A, Coupland G (2006) CONSTANS and the CCAAT box binding complex share a functionally important domain and interact to regulate flowering of Arabidopsis. Plant Cell 18:2971–2984
West MA, Yee KM, Danao J, Zimmerman JL, Fischer RL, Goldberg RB, Harada JJ (1994) LEAFY COTYLEDON1 is an essential reg-ulator of late embryogenesis and cotyledon identity in Arabidop-sis. Plant Cell 6:1731–1745
Xing Y, Fikes JD, Guarente L (1993) Mutations in yeast HAP2/HAP3 define a hybrid CCAAT box binding domain. EMBO J 12:4647–4655
Xing Y, Zhang S, Olesen JT, Rich A, Guarente L (1994) Subu-nit interaction in the CCAAT-binding heteromeric complex is

126 Planta (2014) 239:107–126
1 3
mediated by a very short α-helix in HAP2. Proc Natl Acad Sci USA 91:3009–3013
Yamamoto A, Kagaya Y, Toyoshima R, Kagaya M, Takeda S, Hattori T (2009) Arabidopsis NF-YB subunits LEC1 and LEC1-LIKE activate transcription by interacting with seed-specific ABRE-binding factors. Plant J 58:843–856
Yazawa K, Kamada H (2007) Identification and characterization of carrot HAP factors that form a complex with the embryo-specific transcription factor C-LEC1. J Exp Bot 58:3819–3828
Yu YL, Li YZ, Huang GX, Meng ZD, Zhang D, Wei J, Yan K, Zheng CC, Zhang LY (2011) PwHAP5, a CCAAT-binding transcription factor, interacts with PwFKBP12 and plays a role in pollen tube growth orientation in Picea wilsonii. J Exp Bot 62:4805–4817
Zanetti ME, Blanco FA, Beker MP, Battaglia M, Aguilar OM (2010) A C subunit of the plant nuclear factor NF-Y required for rhizo-bial infection and nodule development affects partner selec-tion in the common bean–Rhizobium etli symbiosis. Plant Cell 22:4142–4157
Zhao M, Ding H, Zhu JK, Zhang F, Li WX (2011) Involvement of miR169 in the nitrogen-starvation responses in Arabidopsis. New Phytol 190:906–915
Zhou ZS, Song JB, Yang ZM (2012) Genome-wide identification of Brassica napus microRNAs and their targets in response to cad-mium. J Exp Bot 63:4597–4613





























