Embed Size (px)
Citation preview

Experimental Hematology 29 (2001) 441–447
0301-472X/01 $–see front matter. Copyright © 2001 International Society for Experimental Hematology. Published by Elsevier Science Inc.PII S0301-472X(01)00618-X
Ibandronate decreases bone disease development and osteoclast stimulatory activity in an in vivo model of human myeloma
Jose C. Cruz
a
, Melissa Alsina
b
, Fiona Craig
c
, Toshiyuki Yoneda
d
, Judith L. Anderson
b
, Mark Dallas
d
, and G. David Roodman
b,e
a
Department of Medicine, Texas Tech University, Lubbock, Tex., USA;
b
Division of Hematology, University of Texas Health Science Center, San Antonio, Tex., USA;
c
Department of Pathology, University of Texas Health Science Center, San Antonio, Tex., USA;
d
Division of Endocrinology, University of Texas Health Science Center, San Antonio,
Tex., USA;
e
General Clinical Research Center and Research Service of the Audie Murphy Veterans Administration Hospital, San Antonio, Tex., USA
(Received 20 September 2000; revised 3 November 2000; accepted 17 November 2000)
Objective.
The benefits of bisphosphonate therapy for multiple myeloma bone disease havebeen clearly documented. However, the effects of bisphosphonates on the osteoclast stimula-tory activity (OSA) that is present in the marrow of patients with multiple myeloma, even be-fore the bone disease is detectable, are unknown. Therefore, we examined the effects of iban-dronate (IB) treatment prior to the development of bone disease in a murine model of humanmyeloma.
Materials and Methods.
Sublethally irradiated severe combined immunodeficient (SCID) micewere transplanted with ARH-77 cells on day 0. These ARH-77 mice were treated daily withsubcutaneous injections of IB started before or at different times after tumor injection as fol-lows: group 1 was started on day
2
7; group 2 on day 0; group 3 on day
1
7; group 4 on day
1
14 after IB administration; and group 5 (control) received no IB. Mice were sacrificed afterthey developed paraplegia.
Results.
The onset of paraplegia was delayed in group 1 vs all other groups (mean day 27 vsday 32;
p
5
0.0098). The number of lytic lesions and the bone surface area of resorption (mm
2
)were significantly decreased in groups 1, 2, and 3, which were treated early with IB, whencompared with groups 4 and 5 (
p
5
0.003 and 0.002, respectively). OSA, as measured by thecapacity of bone marrow plasma from ARH-77 mice to induce osteoclast (OCL) formation inhuman bone marrow cultures, was decreased proportionally to the length of IB treatment.Group 1 had the lowest OSA compared with the other groups (
p
5
0.003). However, all miceeventually developed paraplegia, and at time of sacrifice, tumor burden was not grossly differ-ent among the groups. Interestingly, macroscopic abdominal tumors were more frequent inmice treated with IB.
Conclusion.
These data demonstrate that early treatment of ARH-77 mice with IB prior to de-velopment of myeloma bone disease decreases OSA and possibly retards the development oflytic lesions, but not eventual tumor burden. © 2001 International Society for Experimental
Hematology. Published by Elsevier Science Inc.
Introduction
Multiple myeloma is a malignant disease that results fromthe uncontrolled proliferation of plasma cells in the bonemarrow and other tissues. Osteolytic bone lesions are char-acteristic of multiple myeloma and, along with their compli-cations including severe bone pain, pathologic fractures, spi-
nal cord compression, and hypercalcemia, are a major sourceof morbidity and mortality in these patients [1,2]. The bonedestruction in myeloma is mediated by increased osteoclast(OCL) activity in areas of bone adjacent to myeloma cells.This OCL recruitment is thought to be mediated by OCL ac-tivating factors (OAFs) secreted by myeloma cells and/ormarrow stromal cells in response to the myeloma cells.
Bisphosphonates are a unique class of drugs that recentlyhave been used in the treatment of malignancies that com-monly involve bones, including multiple myeloma [3]. Bis-
Offprint requests to: G. David Roodman, M.D., Ph.D., Research Service(151), Audie Murphy VA Hospital, 7400 Merton Minter Boulevard, SanAntonio, TX 78284; E-mail: [email protected]

442
J.C. Cruz et al./Experimental Hematology 29 (2001) 441–447
phosphonates are characterized by their capacity to inhibitbone resorption, and are effective in the treatment of hyper-calcemia associated with malignancies [4,5]. The use of pa-midronate, a potent second-generation amino-bisphospho-nate, as an adjunct to chemotherapy in patients with stageIII multiple myeloma, has shown efficacy in a large, ran-domized double-blind study. After 21 months, the propor-tion of patients with new skeletal events or new pathologicfractures was significantly lower in the pamidronate-treatedgroup than the placebo group. Although survival was notdifferent between patients receiving pamidronate and pla-cebo, the subgroup of patients treated with pamidronate andsecond-line anti-myeloma chemotherapy lived longer thanthose who received chemotherapy and placebo [6]. In sup-port of this observation, Shipman et al. [7] have reportedthat pamidronate and incandronate can induce cell-cycle ar-rest and apoptosis of human myeloma cell lines in vitro.However, there is no evidence that bisphosphonates pro-duce apoptosis of myeloma cells in patients. More potentthird-generation bisphosphonates are becoming available.These drugs are 100 to 1000 times more potent than pami-dronate in their capacity to inhibit bone resorption, and theymay represent a better treatment option for patients withmyeloma [8]. These new drugs also interfere with other cel-lular metabolic pathways (e.g., the mevalonate pathway)and affect cell processes such as apoptosis and the intracel-lular signaling of key regulatory proteins in tumor cells andin the bone marrow microenvironment [9]. An importantquestion is how the treatment of myeloma with a third-gen-eration bisphosphonate alone (as single therapy) will influ-ence the bone disease and tumor progression in thesepatients. It is noteworthy that the previous studies of bispho-sphonates in multiple myeloma have been done predomi-nantly with patients with stage III disease, whose tumor bur-den and bone destruction are extensive by the time theyenter the study. An attractive option is to treat patients withearlier stages of multiple myeloma with bisphosphonatesbefore extensive bone disease develops. The early use of amore potent bisphosphonate may alter the bone marrow mi-croenvironment and successfully prevent the bone destruc-tion and disease progression, thereby providing a survivaladvantage to these patients. To address this possibility, wetested ibandronate (IB) in an animal model of human multi-ple myeloma bone disease.
Materials and methods
Transplantation of ARH-77 cells into SCID mice
ARH-77 cells, an IgG
k
–secreting human plasma cell leukemiacell line [10], were cultured at 2.5
3
10
5
cells/mL in RPMI-1640(GIBCO, Grand Island, NY, USA) containing 20% fetal calf se-rum (FCS; Hyclone Laboratory, Logan, UT, USA) for 5 days. Fe-male severe combined immunodeficient (SCID) mice (6 to 8weeks old) were irradiated with 400 rads using a Co60 source, and24 hours after irradiation were injected in the tail vein with 10
6
ARH-77 cells (ARH-77 mice) intravenously. Mice were evaluateddaily for signs of disease, and x-rays were obtained at the begin-ning and after sacrificing the mice. After tumor inoculation, wholeblood was collected weekly by retroorbital puncture under meth-oxyflurane for measurement of calcium levels.
Ibandronate administration
IB was generously provided by Boehringer Mannheim (Mann-heim, Germany). IB was diluted in sterile phosphate-buffered sa-line (PBS) at a concentration of 40
m
g/mL for injection into ani-mals. 100
m
L (4
m
g; 160
m
g/kg) of this solution was administereddaily to the mice subcutaneously, until sacrifice. The dose used inthe present study is approximately 100-fold higher than the dosesrequired to inhibit bone resorption in several animal models [11].This dose has been proven to be effective in reducing osteolytic le-sions and tumor burden in bone in a murine model of metastaticbreast cancer [12].
Study groups
Mice were divided into 5 different groups of 5 animals per group.Group 1 started daily injections of IB on day
2
7 (7 days before theARH-77 cell infusion); group 2 started the IB injections on day 0;group 3 started on day
1
7; and group 4 started on day
1
14 afterARH-77 cell infusion. The control group had irradiation and infu-sion of ARH-77 cells, but they did not receive IB. Mice were sacri-ficed 1 or 2 days after becoming paraplegic. Several mice fromeach group were sacrificed when mice in any group in the experi-ment developed paraplegia, in order to evaluate the disease pro-gression in all groups at the same time point. The experiment wasrepeated twice with similar results.
Quantitation of osteolytic bone lesions
After sacrifice, whole-body x-rays of the mice were obtained. Thelesions visible on the radiographs were evaluated by computerizedimage analysis. The number and size (in mm
3
) of the lytic lesionsin the long bones of each mouse were measured.
Histologic analysis
After sacrifice, long bones and vertebral bones were dissected freeof tissue and processed as described below. The liver and spleenwere removed, as well as all abdominal structures that appeared tohave tumor involvement. Soft tissues were fixed in 10% formalinand embedded in paraffin. The sections were stained with hema-toxylin and eosin and were evaluated for the presence of micro-scopic and macroscopic tumor. Apoptosis TUNEL analysis for ap-optotic cells was performed in the soft tissues that containedtumor, as previously described [13]. Bones were fixed in 10% for-malin for 48 hours, decalcified in 14% EDTA for 2 to 3 weeks,processed through several changes of alcohols, and embedded inparaffin. The vertebral bodies and long bones were evaluated forthe presence or absence of tumor infiltration.
Effect of ibandronate on the in vitro growth of ARH-77 cells
The effect of IB on the in vitro growth of ARH-77 cells was testedas follows: ARH-77 cells (2
3
10
5
cells/mL) were cultured in 24-well plates in RPMI-1640 containing 20% FCS. IB was added tothe cultures at different concentrations (10
m
M, 50
m
M, 100
m
M,500
m
M) to evaluate its effect on cell growth in vitro. After incu-bation for 72 hours, cell growth and viability were determined asfollows: The media were removed from the wells, and each wellwas thoroughly washed with 1 mL of PBS. A 10-
m
L aliquot was
J.C. Cruz et al./Experimental Hematology 29 (2001) 441–447
443
taken and mixed with 10
m
L of trypan blue. Viable cells werecounted using a hemocytometer.
Osteoclast stimulatory activity (OSA) determinations
Nonadherent human bone marrow mononuclear cells were pre-pared as previously described [14], and were resuspended in
a
-Minimal Essential Medium (
a
-MEM)-10% horse serum at 10
6
cells/mL. The marrow cells were plated in 96-well plates in quadru-plicate to test the effects of freshly isolated bone marrow plasmafrom ARH-77 mice, treated and not treated with IB. Cultures weremaintained in a humidified atmosphere of 4% CO
2
and air at 37
8
Cfor 3 weeks. The cultures were fed weekly by removing half themedia and replacing it with an equal volume of fresh media. At theend of the culture period, the cells were fixed in 2% formaldehydein PBS, and multinucleated cells that formed were tested for ex-pression of an OCL phenotype by determining their cross-reactivitywith the 23c6 monoclonal antibody, which identifies OCLs (gener-ously provided by Dr. Michael Horton, St. Bartholomew’s Hospi-tal, London) [15]. Reactivity with the 23c6 monoclonal antibodywas determined using biotin-conjugated rabbit anti-mouse IgG cou-pled to alkaline phosphatase (Vector Laboratories; Burlingame,CA, USA) and then counterstaining the cells with methylgreen[16]. We have previously demonstrated that multinucleated cellsthat react with the 23c6 mononuclear antibody (23c6
1
MNC) ex-press calcitonin receptors, contract in response to calcitonin, andform resorption lacunae on calcified matrices, all phenotypic char-acteristics of OCLs [15]. OCL-like cells in each well were countedmicroscopically, without knowledge of the treatment group.
Statistical analysis
Results are expressed as the mean
6
the standard error of the mean(SEM). Differences were compared using Student’s
t
-test and wereconsidered significant for
p
values
,
0.05. Each experiment wasrepeated at least two times for these analyses.
Results
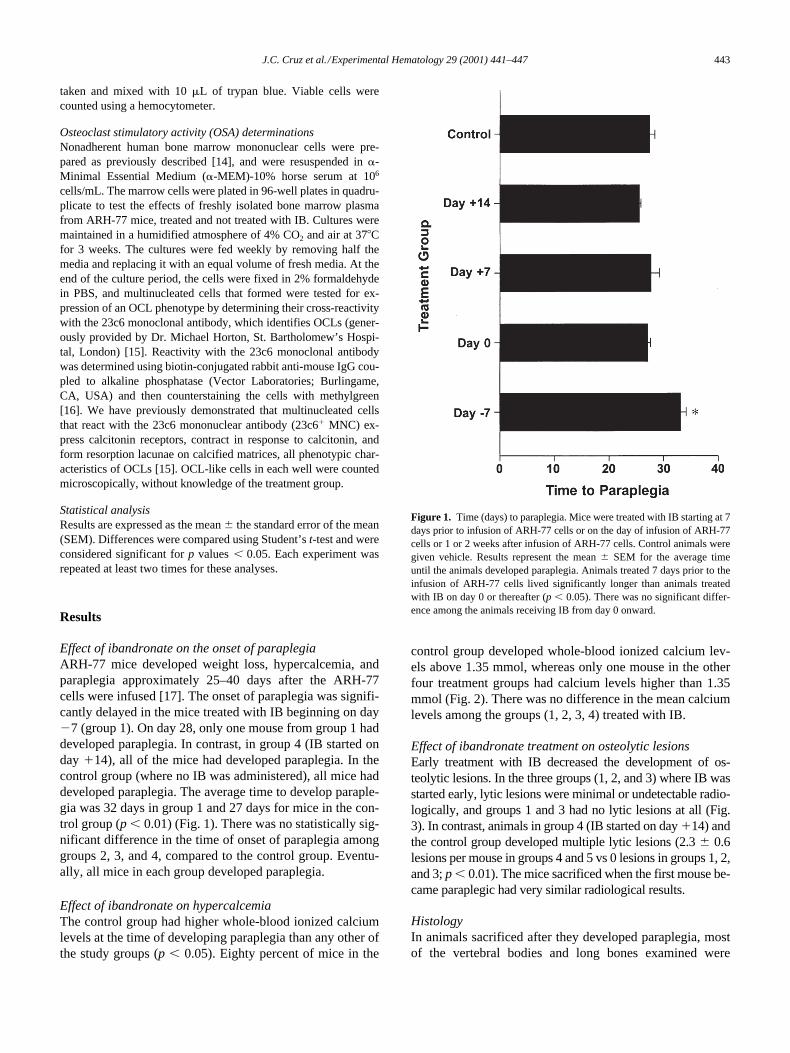
Effect of ibandronate on the onset of paraplegia
ARH-77 mice developed weight loss, hypercalcemia, andparaplegia approximately 25–40 days after the ARH-77cells were infused [17]. The onset of paraplegia was signifi-cantly delayed in the mice treated with IB beginning on day
2
7 (group 1). On day 28, only one mouse from group 1 haddeveloped paraplegia. In contrast, in group 4 (IB started onday
1
14), all of the mice had developed paraplegia. In thecontrol group (where no IB was administered), all mice haddeveloped paraplegia. The average time to develop paraple-gia was 32 days in group 1 and 27 days for mice in the con-trol group (
p
,
0.01) (Fig. 1). There was no statistically sig-nificant difference in the time of onset of paraplegia amonggroups 2, 3, and 4, compared to the control group. Eventu-ally, all mice in each group developed paraplegia.
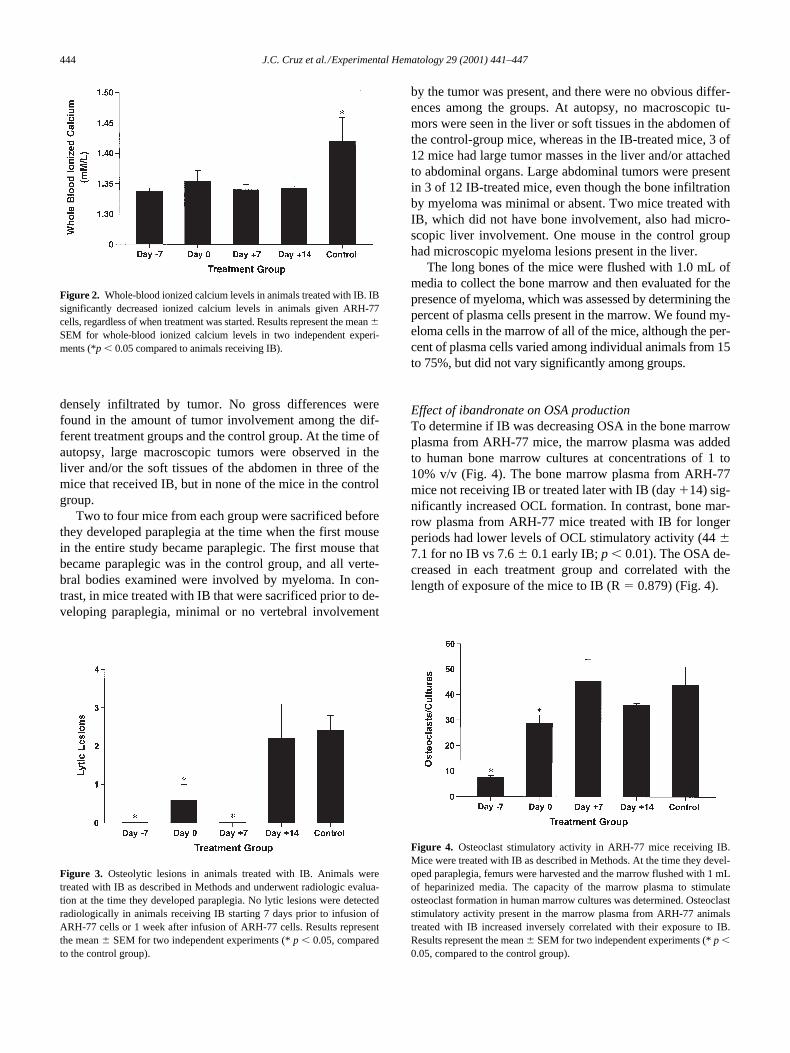
Effect of ibandronate on hypercalcemia
The control group had higher whole-blood ionized calciumlevels at the time of developing paraplegia than any other ofthe study groups (
p
,
0.05). Eighty percent of mice in the
control group developed whole-blood ionized calcium lev-els above 1.35 mmol, whereas only one mouse in the otherfour treatment groups had calcium levels higher than 1.35mmol (Fig. 2). There was no difference in the mean calciumlevels among the groups (1, 2, 3, 4) treated with IB.
Effect of ibandronate treatment on osteolytic lesions
Early treatment with IB decreased the development of os-teolytic lesions. In the three groups (1, 2, and 3) where IB wasstarted early, lytic lesions were minimal or undetectable radio-logically, and groups 1 and 3 had no lytic lesions at all (Fig.3). In contrast, animals in group 4 (IB started on day
1
14) andthe control group developed multiple lytic lesions (2.3
6
0.6lesions per mouse in groups 4 and 5 vs 0 lesions in groups 1, 2,and 3;
p
,
0.01). The mice sacrificed when the first mouse be-came paraplegic had very similar radiological results.
Histology
In animals sacrificed after they developed paraplegia, mostof the vertebral bodies and long bones examined were
Figure 1. Time (days) to paraplegia. Mice were treated with IB starting at 7days prior to infusion of ARH-77 cells or on the day of infusion of ARH-77cells or 1 or 2 weeks after infusion of ARH-77 cells. Control animals weregiven vehicle. Results represent the mean 6 SEM for the average timeuntil the animals developed paraplegia. Animals treated 7 days prior to theinfusion of ARH-77 cells lived significantly longer than animals treatedwith IB on day 0 or thereafter (p , 0.05). There was no significant differ-ence among the animals receiving IB from day 0 onward.
444
J.C. Cruz et al./Experimental Hematology 29 (2001) 441–447
densely infiltrated by tumor. No gross differences werefound in the amount of tumor involvement among the dif-ferent treatment groups and the control group. At the time ofautopsy, large macroscopic tumors were observed in theliver and/or the soft tissues of the abdomen in three of themice that received IB, but in none of the mice in the controlgroup.
Two to four mice from each group were sacrificed beforethey developed paraplegia at the time when the first mousein the entire study became paraplegic. The first mouse thatbecame paraplegic was in the control group, and all verte-bral bodies examined were involved by myeloma. In con-trast, in mice treated with IB that were sacrificed prior to de-veloping paraplegia, minimal or no vertebral involvement
by the tumor was present, and there were no obvious differ-ences among the groups. At autopsy, no macroscopic tu-mors were seen in the liver or soft tissues in the abdomen ofthe control-group mice, whereas in the IB-treated mice, 3 of12 mice had large tumor masses in the liver and/or attachedto abdominal organs. Large abdominal tumors were presentin 3 of 12 IB-treated mice, even though the bone infiltrationby myeloma was minimal or absent. Two mice treated withIB, which did not have bone involvement, also had micro-scopic liver involvement. One mouse in the control grouphad microscopic myeloma lesions present in the liver.
The long bones of the mice were flushed with 1.0 mL ofmedia to collect the bone marrow and then evaluated for thepresence of myeloma, which was assessed by determining thepercent of plasma cells present in the marrow. We found my-eloma cells in the marrow of all of the mice, although the per-cent of plasma cells varied among individual animals from 15to 75%, but did not vary significantly among groups.
Effect of ibandronate on OSA production
To determine if IB was decreasing OSA in the bone marrowplasma from ARH-77 mice, the marrow plasma was addedto human bone marrow cultures at concentrations of 1 to10% v/v (Fig. 4). The bone marrow plasma from ARH-77mice not receiving IB or treated later with IB (day
1
14) sig-nificantly increased OCL formation. In contrast, bone mar-row plasma from ARH-77 mice treated with IB for longerperiods had lower levels of OCL stimulatory activity (44
6
7.1 for no IB vs 7.6
6
0.1 early IB;
p
,
0.01). The OSA de-creased in each treatment group and correlated with thelength of exposure of the mice to IB (R
5
0.879) (Fig. 4).
Figure 2. Whole-blood ionized calcium levels in animals treated with IB. IBsignificantly decreased ionized calcium levels in animals given ARH-77cells, regardless of when treatment was started. Results represent the mean 6SEM for whole-blood ionized calcium levels in two independent experi-ments (*p , 0.05 compared to animals receiving IB).
Figure 3. Osteolytic lesions in animals treated with IB. Animals weretreated with IB as described in Methods and underwent radiologic evalua-tion at the time they developed paraplegia. No lytic lesions were detectedradiologically in animals receiving IB starting 7 days prior to infusion ofARH-77 cells or 1 week after infusion of ARH-77 cells. Results representthe mean 6 SEM for two independent experiments (* p , 0.05, comparedto the control group).
Figure 4. Osteoclast stimulatory activity in ARH-77 mice receiving IB.Mice were treated with IB as described in Methods. At the time they devel-oped paraplegia, femurs were harvested and the marrow flushed with 1 mLof heparinized media. The capacity of the marrow plasma to stimulateosteoclast formation in human marrow cultures was determined. Osteoclaststimulatory activity present in the marrow plasma from ARH-77 animalstreated with IB increased inversely correlated with their exposure to IB.Results represent the mean 6 SEM for two independent experiments (* p ,0.05, compared to the control group).
J.C. Cruz et al./Experimental Hematology 29 (2001) 441–447
445
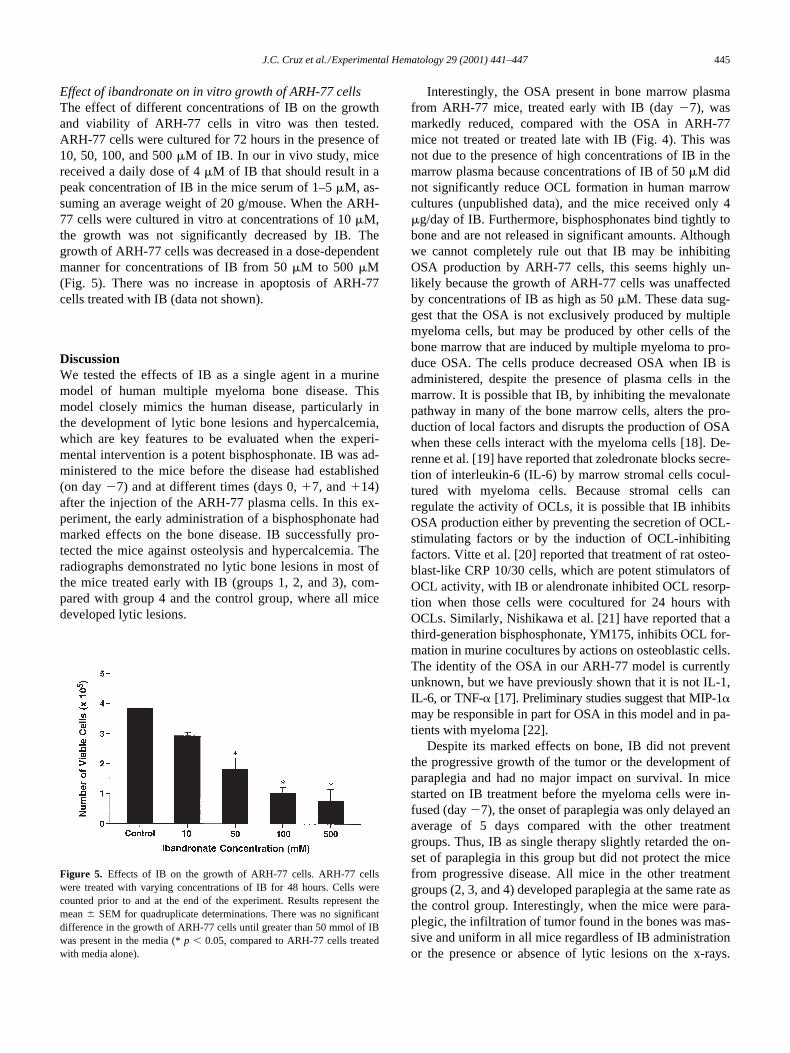
Effect of ibandronate on in vitro growth of ARH-77 cells
The effect of different concentrations of IB on the growthand viability of ARH-77 cells in vitro was then tested.ARH-77 cells were cultured for 72 hours in the presence of10, 50, 100, and 500
m
M of IB. In our in vivo study, micereceived a daily dose of 4
m
M of IB that should result in apeak concentration of IB in the mice serum of 1–5
m
M, as-suming an average weight of 20 g/mouse. When the ARH-77 cells were cultured in vitro at concentrations of 10
m
M,the growth was not significantly decreased by IB. Thegrowth of ARH-77 cells was decreased in a dose-dependentmanner for concentrations of IB from 50
m
M to 500
m
M(Fig. 5). There was no increase in apoptosis of ARH-77cells treated with IB (data not shown).
Discussion
We tested the effects of IB as a single agent in a murinemodel of human multiple myeloma bone disease. Thismodel closely mimics the human disease, particularly inthe development of lytic bone lesions and hypercalcemia,which are key features to be evaluated when the experi-mental intervention is a potent bisphosphonate. IB was ad-ministered to the mice before the disease had established(on day
2
7) and at different times (days 0,
1
7, and
1
14)after the injection of the ARH-77 plasma cells. In this ex-periment, the early administration of a bisphosphonate hadmarked effects on the bone disease. IB successfully pro-tected the mice against osteolysis and hypercalcemia. Theradiographs demonstrated no lytic bone lesions in most ofthe mice treated early with IB (groups 1, 2, and 3), com-pared with group 4 and the control group, where all micedeveloped lytic lesions.
Interestingly, the OSA present in bone marrow plasmafrom ARH-77 mice, treated early with IB (day
2
7), wasmarkedly reduced, compared with the OSA in ARH-77mice not treated or treated late with IB (Fig. 4). This wasnot due to the presence of high concentrations of IB in themarrow plasma because concentrations of IB of 50
m
M didnot significantly reduce OCL formation in human marrowcultures (unpublished data), and the mice received only 4
m
g/day of IB. Furthermore, bisphosphonates bind tightly tobone and are not released in significant amounts. Althoughwe cannot completely rule out that IB may be inhibitingOSA production by ARH-77 cells, this seems highly un-likely because the growth of ARH-77 cells was unaffectedby concentrations of IB as high as 50
m
M. These data sug-gest that the OSA is not exclusively produced by multiplemyeloma cells, but may be produced by other cells of thebone marrow that are induced by multiple myeloma to pro-duce OSA. The cells produce decreased OSA when IB isadministered, despite the presence of plasma cells in themarrow. It is possible that IB, by inhibiting the mevalonatepathway in many of the bone marrow cells, alters the pro-duction of local factors and disrupts the production of OSAwhen these cells interact with the myeloma cells [18]. De-renne et al. [19] have reported that zoledronate blocks secre-tion of interleukin-6 (IL-6) by marrow stromal cells cocul-tured with myeloma cells. Because stromal cells canregulate the activity of OCLs, it is possible that IB inhibitsOSA production either by preventing the secretion of OCL-stimulating factors or by the induction of OCL-inhibitingfactors. Vitte et al. [20] reported that treatment of rat osteo-blast-like CRP 10/30 cells, which are potent stimulators ofOCL activity, with IB or alendronate inhibited OCL resorp-tion when those cells were cocultured for 24 hours withOCLs. Similarly, Nishikawa et al. [21] have reported that athird-generation bisphosphonate, YM175, inhibits OCL for-mation in murine cocultures by actions on osteoblastic cells.The identity of the OSA in our ARH-77 model is currentlyunknown, but we have previously shown that it is not IL-1,IL-6, or TNF-
a
[17]. Preliminary studies suggest that MIP-1
a
may be responsible in part for OSA in this model and in pa-tients with myeloma [22].
Despite its marked effects on bone, IB did not preventthe progressive growth of the tumor or the development ofparaplegia and had no major impact on survival. In micestarted on IB treatment before the myeloma cells were in-fused (day
2
7), the onset of paraplegia was only delayed anaverage of 5 days compared with the other treatmentgroups. Thus, IB as single therapy slightly retarded the on-set of paraplegia in this group but did not protect the micefrom progressive disease. All mice in the other treatmentgroups (2, 3, and 4) developed paraplegia at the same rate asthe control group. Interestingly, when the mice were para-plegic, the infiltration of tumor found in the bones was mas-sive and uniform in all mice regardless of IB administrationor the presence or absence of lytic lesions on the x-rays.
Figure 5. Effects of IB on the growth of ARH-77 cells. ARH-77 cellswere treated with varying concentrations of IB for 48 hours. Cells werecounted prior to and at the end of the experiment. Results represent themean 6 SEM for quadruplicate determinations. There was no significantdifference in the growth of ARH-77 cells until greater than 50 mmol of IBwas present in the media (* p , 0.05, compared to ARH-77 cells treatedwith media alone).

446 J.C. Cruz et al./Experimental Hematology 29 (2001) 441–447
These data demonstrate that the progressive invasion of thebone, spinal cord compression, and death occur even whenthe lytic lesions are not evident on x-ray.
Surprisingly, large tumors were present in the abdomenof 3 of 12 animals treated with IB. None of these animalshad extensive marrow infiltration with myeloma. Interest-ingly, most of the animals terminated early had not as yetdeveloped extensive bone infiltration with myeloma, but theabdomen was invaded with large tumor lesions in three ofthe mice. Macroscopic tumors were not detected in the con-trol mice not receiving IB. This finding differs from thestudies of the effects of IB in breast cancer in mice, whichdemonstrated a marked reduction in tumor burden in thebones, but no significant increase in the soft tissue infiltra-tion by the tumor was seen [12]. Our results suggest thatearly treatment with IB may affect the homing of myelomacells to the bone marrow microenvironment and may favorthe seeding of tumor cells to extraosseus tissues, such as theliver. Eventually, the myeloma cells invade the bone mar-row cavity and compress the spinal canal. However, a muchlarger study would be required to test this possibility. Alter-natively, the development of large abdominal tumors couldbe a particular characteristic of our model, because the hu-man myeloma cells (ARH-77) originally were obtainedfrom a patient with plasma cell leukemia.
Similar to our current study, Dallas et al. [23] adminis-tered IB to mice infused with a murine multiple-myelomacell line. They found IB markedly inhibited myeloma-asso-ciated bone destruction and did not reduce the tumor burdenin the skeleton or total tumor burden, as assessed by IgGlevels, or affect liver and spleen weight. The tumor cellswere able to grow in the bone marrow and completely re-place the marrow cavity, in spite of the OCL inhibition bythe bisphosphonate. These authors did not examine the ef-fects of IB on OSF activity in this model. In contrast to ourresults, the distribution of the tumor in the abdominal or-gans in this study was not different between the mice re-ceiving IB treatment and the control groups.
In our in vitro experiments, the addition of IB to theARH-77 cell culture did not significantly inhibit the cellgrowth at the concentration expected in vivo. There was nosignificant increase in the number of apoptotic cells in theIB-treated mice. The lack of apoptotic effect of IB on my-eloma cells has been reported by other groups [19], al-though other in vitro studies have suggested an increase inapoptosis of myeloma cells exposed to bisphosphonates [7].
In summary, the early administration of ibandronate suc-cessfully retarded the development of lytic lesions in micewith multiple myeloma. IB also affected the bone marrowmicroenvironment and resulted in decreased OSA produc-tion. In contrast, ibandronate did not have a marked effecton survival or tumor growth. Our study suggests that treat-ment of individuals with myeloma bone disease with athird-generation bisphosphonate may decrease the osteo-clast stimulatory activity in the marrow and possibly retard
development of bone lesions, but does not prevent the fataldissemination of the disease in bone and extraosseus tissues.
AcknowledgmentsWe thank Bibi Cates for preparation of this manuscript and thestaff of the General Clinical Research Center, Audie Murphy VAHospital, for their assistance in the care of our myeloma patientsand the collection of samples. This work was supported by re-search funds from the Veterans Administration and National Insti-tutes of Health NCI Grants CA69136 and CA40035.
References1. Roodman GD (1997) Mechanisms of bone lesions in multiple my-
eloma and lymphoma. Cancer 80:15572. Mundy GR, Bertolini DR (1986) Bone destruction and hypercalcemia
in plasma cell myeloma. Semin Oncol 3:2913. Body JJ, Bartl R, et al. (1998) Current use of bisphosphonates in on-
cology. J Clin Onc 16:35904. Berenson J (1997) Bisphosphonates in multiple myeloma. Cancer
(Suppl 80:8):16615. Bilezikian JP (1992) Management of acute hypercalcemia. N Engl J
Med 326:11966. Berenson JR, Lichtenstein A, Porter L, et al. (1998) Long-term pam-
idronate treatment of advanced multiple myeloma patients reducesskeletal events. J Clin Oncol 16(2):593
7. Shipman CM, Rogers MJ, Apperley JF (1997) Bisphosphonates in-duce apoptosis in human myeloma cells; a novel antitumor activity. BrJ Haematol 98:665
8. Fleisch H (2000) Bisphosphonates in bone disease: from the laboratoryto the patient, 4th ed. New York: Academic Press
9. Muhlbauer RC, Bauss F, Janner M, et al. (1991) BM 21.0955, a potentnew bisphosphonate to inhibit bone resorption. J Bone Miner Res6(9):1003
10. Burk KH, Drewindo B, Trujillo JM, Aheram MJ (1978) Establishmentof a human plasma cell line in vitro. Cancer Res 38(8):2508
11. Bauss F (1997) Ibandronate in malignant bone disease and osteoporo-sis: clinical results. Oncologie 20:204
12. Yoneda T, Sasaki A, Dunstan C, et al. (1997) Inhibition of osteolyticbone metastasis of breast cancer by combined treatment with the bis-phosphonate ibandronate and tissue inhibitor of the matrix metallopro-teinase-2. J Clin Invest 99(10):2509
13. Boyce BF, Wright K, Reddy SV, et al. (1995) Targeting Simian virus40 T antigen to the osteoclast in transgenic mice causes osteoclast tu-mors and transformation and apoptosis of osteoclasts. Endocrinology136:5751
14. Kurihara N, Chenu C, Miller M, Civin CI, Roodman GD (1990) Iden-tification of committed mononuclear precursors for osteoclast-likecells formed in long-term marrow cultures. Endocrinology 126:2733
15. Kurihara N, Gluck S, Roodman GD (1990) Sequential expression ofphenotype markers for osteoclasts during differentiation of precursorsfor multinucleated cells formed in long-term human marrow cultures.Endocrinology 127(6):3215
16. Kukita T, McManus LM, Miller M, Civin C, Roodman GD (1989) Os-teoclast-like cells formed in long-term human bone marrow culturesexpress a similar surface phenotype as authentic osteoclasts. Lab In-vest 60:532
17. Alsina M, Boyce B, Devlin R, et al. (1996) Development of an in vivomodel of human multiple myeloma bone disease. Blood 87(4):1495
18. Croucher PI, Shipman CM, Russell RGR, et al. (1999) Bisphospho-nates—mechanisms of action in multiple myeloma [abstract]. Proceed-

J.C. Cruz et al./Experimental Hematology 29 (2001) 441–447 447
ings of International Multiple Myeloma Workshop in Stockhom, Swe-den, #143:69
19. Derenne S, Amiot M, Barille S, et al. (1999) Zoledronate is a potent in-hibitor of myeloma cell growth and secretion of IL-6 and MMP-1 bythe tumoral environment. J Bone Miner Res 14:2048
20. Vitte C, Fleisch H, Guenther HL (1996) Bisphosphonates induce os-teoblasts to secrete an inhibitor of osteoclast-mediated resorption. En-docrinology 137:2324
21. Nishikawa M, Yamamoto M, Murakami T, Akatsu T, Kugai N, NagataN (1998) A third-generation bisphosphonate, YM175, inhibits osteo-
clast formation in murine cocultures by inhibiting proliferation of pre-cursor cells via supporting cell-dependent mechanisms. J Cell Bio-chem 68:186
22. Choi SJ, Cruz JC, Craig F, et al. (2000) Macrophage inflammatoryprotein 1-a (MIP-1a) is a potential osteoclast stimulatory factor inmultiple myeloma. Blood 96:671
23. Dallas SL, Garrett IR, Oyajobi BO, et al. (1999) Ibandronate reducesosteolytic lesions but not tumor burden in a murine model of myelomabone disease. Blood 93:1697





























