Embed Size (px)
Citation preview

Proc. Nati. Acad. Sci. USAVol. 83, pp. 917-921, February 1986Biochemistry
Hypothesis about the function of membrane-buried proline residuesin transport proteinsCHRISTOPHER J. BRANDL*t AND CHARLES M. DEBERtt§*Banting and Best Department of Medical Research, C. H. Best Institute, Toronto, M5G 1L6, ON, Canada; tDepartment of Biochemistry, University ofToronto, Toronto, M5S 1A8, ON, Canada; and MResearch Institute, The Hospital for Sick Children, Toronto, M5G 1X8, ON, Canada
Communicated by Elkan R. Blout, October 7, 1985
ABSTRACT In a survey of the bilayer-spanning regions ofintegral membrane proteins, membrane-buried proline resi-dues were found in nearly all transport proteins examined,whereas membrane-buried regions of nontransport proteinswere largely devoid of intramembranous proline residues.When amino acids from the complete sequences of repre-sentative sets of transport and nontransport membrane pro-teins were analyzed for the distribution of proline residuesbetween aqueous vs. membranous domains, proline was shownto be selectively excluded from membranous domains of thenontransport proteins, in accord with expectation from ener-getic and structural considerations. In contrast, proline resi-dues in transport proteins were evenly distributed betweenaqueous and membranous domains, consistent with the notionthat functional membrane-buried proline residues are selec-tively included in transport proteins. As cis peptide bondsinvolving proline arise in proteins and have been implicated inprotein dynamic processes, the cis-trans isomerization of anXaa-Pro peptide bond (Xaa = unspecified amino acid) buriedwithin the membrane-and the resulting redirection of theprotein chain-is proposed to provide the reversible confor-mational change requisite for the regulation (opening/closing)of a transport channel. Parallel to this function, the relativelynegative character of the carbonyl groups of Xaa-Pro peptidebonds may promote their participation as intramembranousliganding sites for positive species in proton/cation transportprocesses.
Proteins that span the lipid bilayers of cellular membranescan be divided into classes based on the structure andfunction of their transmembrane segments (1). Proteins suchas the immunoglobulin M heavy chain (2), influenza virushemagglutinins (3-7), and histocompatibility antigens (8-12)contain a single clearly hydrophobic transmembrane segmentthat serves primarily to anchor the protein within the bilayer(1). Proteins responsible for transport, such as the acetyl-choline receptor (13), the Ca2+,Mg2+-ATPase of sarcoplas-mic reticulum (14), and the lac permease of Escherichia coli(15), are generally believed to traverse the bilayer severaltimes, forming an assembly of conformationally orderedsegments that can create a specific transport channel (16).The higher proportion of polar amino acids noted in mem-brane transport proteins (1) may impart the amphipathiccharacteristics requisite for formation of hydrophilic poreswithin the membrane (17-20). However, regulation of thechannel itself must further involve either the creation oraccessibility of the hydrophilic pore.Recent advances in recombinant DNA technology have
allowed the sequencing of many membrane proteins. Despitethe availability of their sequences, the molecular basis fortransport protein function and regulation is not yet under-stood. Models depicting transport suggest that conformation-
al changes in the protein structure are necessary for substratetranslocation across the lipid bilayer (21-24). Any suchchange responsible for transport would have to be a regulat-ed, reversible dynamic event with specific conformationalconsequences. The cis-trans isomerization of a peptide bondlocated within the membrane and the resulting redirection ofthe local protein chain could provide the conformationalchange necessary for the regulation of a transport channel.Peptide bonds between two amino acids in proteins aregenerally planar and trans (25). However, primarily becauseof destabilization of the trans state caused by steric require-ments of neighboring substituents, the cis form is of compa-rable stability when the imino acid proline (Pro) occurs at thecarboxyl-terminal side of the bond (26). Cis peptide bondsinvolving proline arise in proteins (e.g., see ref. 27) and insynthetic linear and cyclic peptides (28).
After our initial observation that membrane-buried prolineresidues occur widely in transport proteins, we surveyedmembrane-traversing amino acid sequences currently knownfor membrane proteins and compared intramembranous re-gions of transport proteins with those of nontransport pro-teins. Our findings, reported here, lead to the proposal ofspecific functionality of membrane-buried proline residues intransport proteins through (i) channel regulation via cis-transisomerization of Xaa-Pro peptide bonds, where Xaa is anunspecified amino acid, and/or (ii) involvement of Xaa-Procarbonyl groups as intramembranous cation liganding sites.
RESULTS AND DISCUSSIONOccurrence of Membrane-Buried Proline Residues in Trans-
port Proteins. A prerequisite for the general involvement ofproline in transport processes is its occurrence in membra-nous regions of transport proteins. Bilayer-spanning regionsfor which data were available of transport proteins, includingthose that function both as active pumps and as channels, arepresented (bold-face type) in Table 1, whereas intramembra-nous sequences of nontransport membrane proteins are given(bold-face type) in Table 2. A variety of experimentalmethods have been used to deduce these membrane-traver-sing regions, including x-ray crystallography [e.g., bacteri-orhodopsin (50)], hydrophobic-labeling reagents [e.g.,glycophorin (51)], enzymatic digestion {e.g., bovinerhodopsin [reviewed by Hargrave (52)]}, labeling with site-directed antibodies [e.g., lac permease (53)], deletion map-ping using recombinant DNA technology [e.g., acetylcholinereceptor (54)], and determining positions of carbohydratemoieties [e.g., band 3 (36)]. In conjunction with thesemethods, many studies have used the predictive criteria ofamino acid hydrophobicity profiles, such as those presentedby Meek (55), Rose and Roy (56), Hopp and Woods (57),Argos et al. (58), and Kyte and Doolittle (59). These profiles,which are based intrinsically on such data as crystallographic
Abbreviation: Xaa, unspecified amino acid.§To whom correspondence should be addressed at The Hospital forSick Children.
917
The publication costs of this article were defrayed in part by page chargepayment. This article must therefore be hereby marked "advertisement"in accordance with 18 U.S.C. §1734 solely to indicate this fact.

918 Biochemistry: Brandl and Deber
Table 1. Bilayer-spanning regions of membrane proteins involved in transport
Seg- Adjacent AdjacentProtein ment residues Bilayer region residues Ref.
Acetylcholine a QRI PLYFVVNVIIPCLLFSFLTGLVFYLPT DSG 13receptor Ml , QRK PLFYIVYTIIPCILISILAILVFYLPP DAG
y QRK PLFYIINEIAPCVLISSLVVLVYFLPA QAG8 RRK PLFYVINEITPCVLISFLASLAFYLPA ESG
Bacteriorhodopsin 40-63 PDA KKFYAITTLVPAIAFTMYLSMLLG YGL 29, 3077-101 EQN PIYWARYADWLFTTPLLLLDLALLV DADQ172-199 STF KVLRNVTWLWSAYPWWLIGSEGAGIV PLN
Electrophorus electricus Si II LKK WVHFVMMDPFMTLFITLCIILNTLFMSI EHH 31Na+ channel Si IV VQG VVYDIVTQPFEDIFDMALICINMVAMMV ESE
S3 I IGH FTFLPDPWNWLDFSWTMTYIT EFIK+ transporter of E. coliKdpB 587-610 AK YFAIIPAAFAATYPQLNALNIMCL HSPD 32
615-636 PD SAILSAVIFNALIIVFLIPLAL KGG653-682 RRN NLWIYGLGGLLVPFIGIKVIDLLLTVCGLV
Lac carrier of E. coli 10-34 NTNF WMFGLFFFFYFFIMGAYFPFFPIWL HDIN 15, 3346-68 KSDT GHFAAISLFSLLFQPLFGLLS DKL75-99 GLRK YLLWHTGMLVMFAPFFIFIFGPLL QYN101-130 Q YNILVGSIVGGIYLGFCFNAGAPAVEAFIE K260-283 GTR VFGYVTTMGELLNASIMFFAPLH NR312-337 TS ALEVVILKTLHMFEVPFLLVGCFKY ITS
Mitochondrial protonATPase proteolipid 52-78 LRG QLFSYAILGFAFVEAIGLFDLMVALMA KFT 34, 35
Erythrocyte band 3fragment YFPG KLRRVIGDFGVPISILIMVLVDFFI QDTYT 36
Rhodopsin (bovine) 34-64 PWQ FSMLAATMFLLIMLGFPINFLTLYV TVQ 37150-176 ENH AIMGVAFTWVMALACAAPPLV GWS201-234 NES FVIYMFVVHFIIPLIVIFFCYGQLVFTV KEA253-283 VTR MVIIMVIAFLICWLPYAGVAFYIF THQ287-316 FGP IFMTIPAFFAKTSAVYNPVIYIMM NKQ
Yeast cytochrome b 146-169 GAT VITNLFSAIPFVGNDIVSWLWGGF SVS 38179-201 IQR FFALHYLVPFHAAMVIMHLMAL HIH230-252 FKD LVTVFLFMLILALFVFYSPNTLG HPD289-308 PDK LLGVITMFAAILVLLVLPFT DRS348-370 EVP YVLMGQIATFIYFAYFLHVPVI STI
The standard single-letter amino acid code is used, with proline residues in the bilayer region emphasized with superior dots.
determination of "buried" residues in globular proteins, andon parameters arising from thermodynamic properties ofamino acids (e.g., partition coefficients), have been usedwidely to aid in identification of intramembranous sequences.Nevertheless, it emphasized that the boundaries of "mem-brane-transversing" sequences given in Tables 1 and 2-particularly in instances where hydrophobicity profiles havebeen relied upon exclusively by the various cited authors-must be regarded as tentative. The survey in Table 1 revealedthat, with one exception (see below), all contain at least onemembrane-buried proline residue. This finding was in con-trast to the intramembranous sequences of nontransportproteins; a survey of a similar number of bilayer-spanningregions known for proteins in the latter category (Table 2)produced only a few isolated examples of membrane-buriedproline residues. This difference in the proportion of trans-port proteins [eight of nine individual (or families of) proteinsin Table 1] vs. nontransport proteins (3 of 16 in Table 2) withat least one intramembranous proline was statistically signif-icant at the P = 0.0003 level. (Statistical computations wereperformed by using the t test for the difference ofproportionsfrom independent samples with the Microstat package on aCP/M microcomputer.)Many membrane transport proteins traverse the bilayer
several times; for instance, the sodium channel of Electro-phorus electricus has 16 proposed membrane-spanning re-gions (31) of which the three containing proline are given inTable 1. It is reasonable that conformational perturbation atonly one or two sites should be sufficient to regulate the
opening and closing of the protein's ion channel. Note thatmembrane-buried prolines are found conserved in similarpositions in transport proteins that have common functions.Thus, the Ml monomer segments of the four acetylcholinereceptor subunits a, (3, y, and 8 all contain three prolineresidues (Table 1). As Noda et al. (13) indicate, this favors theidea that the M1 segment is of particular functional signifi-cance. Two forms of the Ca2+,Mg2-ATPase of sarcoplasmicreticulum (not included in Table 1) that share 80% amino acidhomology have been observed recently in neonatal rabbitskeletal muscle (60, 61); (all) four proline residues appear tobe conserved within the membrane-spanning regions of thesetwo isozymes.
cis-trans isomerization of a membrane-buried Xaa-Probond could also constitute the basic molecular mechanismresponsible for conformational changes required in othermembrane events, such as receptor-mediated signal trans-duction. In fact, certain membrane-bound receptors may beconsidered to contain a third class of membrane-spanningregion, which functions not to anchor the receptor or mediateion transport but is involved in signal transduction across thebilayer. Thus, both the low density lipoprotein receptor (62)and the insulin receptor (63) contain membrane-buriedproline [although the epidermal growth factor receptor (64)does not].Bovine rhodopsin is also included in Table 1, even though
a transport function for this molecule has not been proven(for a review, see ref. 65); however, as with membrane-boundreceptors, conformational changes of the polypeptide chain
Proc. NatL Acad. Sci. USA 83 (1986)
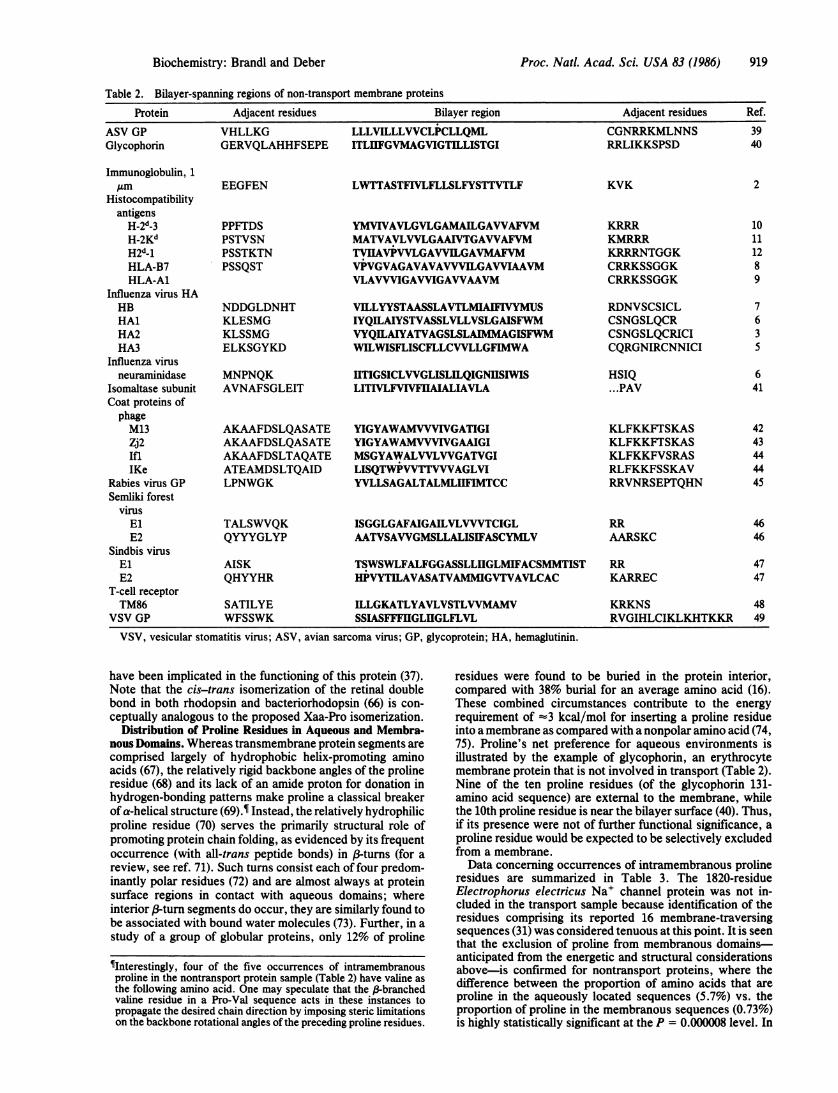
Proc. Natl. Acad. Sci. USA 83 (1986) 919
Table 2. Bilayer-spanning regions of non-transport membrane proteins
Protein Adjacent residues Bilayer region Adjacent residues Ref.
ASV GP VHLLKG LLLVILLLWCLPCLLQML CGNRRKMLNNS 39Glycophorin GERVQLAHHFSEPE ITLIIFGVMAGVIGTILLISTGI RRLIKKSPSD 40
Immunoglobulin, 1,um EEGFEN LWTTASTFIVLFLLSLFYSTTVTLF KVK 2
Histocompatibilityantigens
H-2d-3 PPFTDS YMVIVAVLGVLGAMAILGAWAFVM KRRR 10H-2Kd PSTVSN MATVAVLVVLGAAIVTGAWAFVM KMRRR 11H2d_1 PSSTKTN TVIIAVPVVLGAVVILGAVMAFVM KRRRNTGGK 12HLA-B7 PSSQST VPVGVAGAVAVAWVILGAWIAAVM CRRKSSGGK 8HLA-A1 VLAVVVIGAVVIGAWAAVM CRRKSSGGK 9
Influenza virus HAHB NDDGLDNHT VILLYYSTAASSLAVTLMIAFIVYMUS RDNVSCSICL 7HAl KLESMG IYQILAIYSTVASSLVLLVSLGAISFWM CSNGSLQCR 6HA2 KLSSMG VYQILAIYATVAGSLSLAIMMAGISFWM CSNGSLQCRICI 3HA3 ELKSGYKD WILWISFLISCFLLCVVLLGFIMWA CQRGNIRCNNICI 5
Influenza virusneuraminidase MNPNQK ITIGSICLWGLISLILQIGNIISIWIS HSIQ 6
Isomaltase subunit AVNAFSGLEIT LITIVLFVIVFUAIALIAVLA ...PAV 41Coat proteins ofphageM13 AKAAFDSLQASATE YIGYAWAMVVVIVGATIGI KLFKKFTSKAS 42Zj2 AKAAFDSLQASATE YIGYAWAMVVVIVGAAIGI KLFKKFTSKAS 43Ifl AKAAFDSLTAQATE MSGYAWALVVLWGATVGI KLFKKFVSRAS 44IKe ATEAMDSLTQAID LISQTWPWTTVVVAGLVI RLFKKFSSKAV 44
Rabies virus GP LPNWGK YVLLSAGALTALMLHFIMTCC RRVNRSEPTQHN 45Semliki forest
virusEl TALSWVQK ISGGLGAFAIGAILVLVVVTCIGL RR 46E2 QYYYGLYP AATVSAWGMSLLALISIFASCYMLV AARSKC 46
Sindbis virusEl AISK TSWSWLFALFGGASSLLIGLMIFACSMMTIST RR 47E2 QHYYHR HPVYTILAVASATVAMMIGVTVAVLCAC KARREC 47
T-cell receptorTM86 SATILYE ILLGKATLYAVLVSTLVVMAMV KRKNS 48
VSV GP WFSSWK SSIASFFFIIGLHGLFLVL RVGIHLCIKLKHTKKR 49
VSV, vesicular stomatitis virus; ASV, avian sarcoma virus; GP, glycoprotein; HA, hemaglutinin.
have been implicated in the functioning of this protein (37).Note that the cis-trans isomerization of the retinal doublebond in both rhodopsin and bacteriorhodopsin (66) is con-ceptually analogous to the proposed Xaa-Pro isomerization.
Distribution of Proline Residues in Aqueous and Membra-nous Domains. Whereas transmembrane protein segments arecomprised largely of hydrophobic helix-promoting aminoacids (67), the relatively rigid backbone angles of the prolineresidue (68) and its lack of an amide proton for donation inhydrogen-bonding patterns make proline a classical breakerof a-helical structure (69).¶ Instead, the relatively hydrophilicproline residue (70) serves the primarily structural role ofpromoting protein chain folding, as evidenced by its frequentoccurrence (with all-trans peptide bonds) in /-turns (for areview, see ref. 71). Such turns consist each of four predom-inantly polar residues (72) and are almost always at proteinsurface regions in contact with aqueous domains; whereinterior (-turn segments do occur, they are similarly found tobe associated with bound water molecules (73). Further, in astudy of a group of globular proteins, only 12% of proline
¶Interestingly, four of the five occurrences of intramembranousproline in the nontransport protein sample (Table 2) have valine asthe following amino acid. One may speculate that the /-branchedvaline residue in a Pro-Val sequence acts in these instances topropagate the desired chain direction by imposing steric limitationson the backbone rotational angles of the preceding proline residues.
residues were found to be buried in the protein interior,compared with 38% burial for an average amino acid (16).These combined circumstances contribute to the energyrequirement of -3 kcal/mol for inserting a proline residueinto a membrane as compared with a nonpolar amino acid (74,75). Proline's net preference for aqueous environments isillustrated by the example of glycophorin, an erythrocytemembrane protein that is not involved in transport (Table 2).Nine of the ten proline residues (of the glycophorin 131-amino acid sequence) are external to the membrane, whilethe 10th proline residue is near the bilayer surface (40). Thus,if its presence were not of further functional significance, aproline residue would be expected to be selectively excludedfrom a membrane.Data concerning occurrences of intramembranous proline
residues are summarized in Table 3. The 1820-residueElectrophorus electricus Na' channel protein was not in-cluded in the transport sample because identification of theresidues comprising its reported 16 membrane-traversingsequences (31) was considered tenuous at this point. It is seenthat the exclusion of proline from membranous domains-anticipated from the energetic and structural considerationsabove-is confirmed for nontransport proteins, where thedifference between the proportion of amino acids that areproline in the aqueously located sequences (5.7%) vs. theproportion of proline in the membranous sequences (0.73%)is highly statistically significant at the P = 0.000008 level. In
Biochemistry: Brandl and Deber

920 Biochemistry: Brandl and Deber
Table 3. Distribution of proline residues in membrane proteins
Amino acids Transport Nontransport
Total no. 2673 4609% proline residues 4.3 5.3
Aqueous, no. 1598 4097Proline, no. 81 232% proline residues 5.1 5.7
Membranous, no. 1075 412Proline, no. 35 3% proline residues 3.3 0.73
Data were compiled for 8 of the transport proteins in Table 1 andfor 16 of the nontransport proteins in Table 2. All intramembranousregions (including all bilayer-spanning regions of the transportproteins) are included in the number of "membranous" amino acids.Residues chosen for aqueous or membrane domains are thosedesignated by the authors of the original papers cited in Tables 1 and2.
contrast to this result, proline residues in transport proteinsare distributed rather evenly between aqueous and membra-nous domains; the proportion of proline in aqueously locatedsequences (5.1%) is slightly greater than that for membranesequences (3.3%) (P = 0.02). Recomputation ofP values byusing the paired t test for the difference of means gaveessentially identical results. These findings are consistentwith the notion that membrane-buried proline residues in factare selectively included in transport proteins and, conse-quently, that their presence has functional significance.Environmental Dependence of cis-trans Isomerization
Rates. The cis-trans isomerization of Xaa-Pro bonds hasbeen implicated in dynamic processes of proteins. Brandts etal. (76) have presented arguments indicating that the slowphase of protein renaturation is the result of Xaa-Proisomerization. Further evidence (77-79) has supported thishypothesis, emphasizing the importance of Xaa-Proisomerization in conformational interconversions of pro-teins. Isomerization of Xaa-Pro bonds can be a relativelyslow process (e.g., it has a half-life of 300 sec) for the His-Prodipeptide at room temperature, reflecting the activationenergy of ca. 20 kcal/mol for interconversion between thetwo species (80). Whereas a priori this would seem to be tooslow an event to play a role in the regulation of membraneproteins, which turnover in the order of milliseconds, thehalf-life for Xaa-Pro isomerization is known to be highlydependent on the peptide chain in which the bond is found.For example, in the biologically active decapeptide angio-tensin, the His-Pro bond isomerizes to the cis form with ahalf-life of ca. 3 sec at room temperature (80)-more than100-fold faster than the same bond in the simple His-Prodipeptide. Lin and Brandts (78) have found the isomerizationrate of the Tyr-Pro bond of RNase A to differ significantly,depending on the extent of denaturation of the protein. Smalldeviations from peptide bond planarity, as may arise from thelong-range interactions that stabilize the tertiary structure ofa protein within a membrane, could bring Xaa-Pro isomeriza-tion kinetics into the millisecond range. Thus one couldpostulate that the transport protein domain (whether throughits tertiary structure or through an enzyme-assisted event)acts to catalyze the cis-trans isomerization and to determinethe cis/trans .ratio of a given Xaa-Pro bond, with thetriggering process being directly coupled to the stimulus thatregulates the channel.Xaa-Pro Peptide Bonds as Liganding Sites. Parallel to their
possible role in peptide bond isomerization, further function-ality for intramembranous proline residues may arise fromthe tertiary amide character of the Xaa-Pro bond, whichconfers increased basicity to the attached carbonyl oxygen
atom (81). This situation promotes the involvement of theXaa-Pro carbonyl group in hydrogen bonding (82) and as aliganding site for positively charged species in peptide-cationinteractions (28). The participation of such carbonyl groupsin transmembrane Ca + transport processes has also beensuggested from studies with ionophorous peptides containingimino acids (83). Proteins that transport H+ ions couldsimilarly utilize carbonyl oxygens of membrane-buried Xaa-Pro peptide bonds. Proton translocation may be distinct fromtranslocation of larger substrates in that transport may occurvia transfer ofthe proton along a "chain" ofproton-acceptinggroups without major conformational changes (84). Thus,membrane-buried prolines could contribute to the polarpeptide backbone that comprises part ofthis chain. Ofthe H+translocators listed in Table 1 (including bacteriorhodopsinand yeast cytochrome b), only the mitochondrial H+-ATPaseproteolipid from Neurospora crossa (ref. 34; see also ref. 35)lacks a buried proline residue.
CONCLUSIONLocal structural changes arising from regulated cis-transisomerization of membrane-buried Xaa-Pro bonds of trans-port proteins could result in the realignment of amphipathichelices found within the membrane such that a hydrophilicpore is created or becomes accessible to a substrate ready fortransport across the membrane. The present survey ofmembrane-traversing regions known for membrane transportproteins confirms the selective inclusion of the requisitepotentially functional proline residues. Spectroscopic ap-proaches to the study of these phenomena-e.g., viabiosynthetic incorporation of 13C-labeled amino acids intomembrane proteins of E. coli auxotrophs (e.g., ref. 85)-should aid in the experimental determination of intramem-branous Xaa-Pro conformational state(s) (C.M.D., C.J.B.and P. E. Fraser, unpublished results).
We appreciate the support of Prof. David H. MacLennan, Bantingand Best Department of Medical Research, University of Toronto.We thank Prof. Raisa B. Deber, Department of Health Administra-tion, University of Toronto, for statistical analyses and stimulatingdiscussions. This work was supported in part by grants to C.M.D.from the Medical Research Council of Canada and the NaturalSciences and Engineering Research Council of Canada. C.J.B. holdsa Medical Research Council Studentship.
1. Clement, J. M. (1983) Biochimie 65, 325-338.2. Rogers, J., Early, P., Carter, C., Calame, K., Bond, M.,
Hood, L. & Wall, R. (1980) Cell 20, 303-312.3. Weiner, A. M., Platt, T. & Weber, K. (1972) J. Biol. Chem.
247, 3242-3251.4. Porter, A. G., Barber, C., Carey, M. H., Hallewa, R. A.,
Threlfall, G. & Emtage, J. S. (1979) Nature (London) 282,471-477.
5. Fang, R., Min-Jou, W., Huylebroeck, D., Devos, R. & Fiers,W. (1981) Cell 25, 315-323.
6. Fields, S., Winter, G. & Brownlee, G. G. (1981) Nature(London) 290, 213-217.
7. Krystal, M., Elliott, R. M., Benz, E. W., Jr., Young, J. F. &Palese, P. (1982) Proc. Natl. Acad. Sci. USA 79, 4800-4804.
8. Orr, H. T., Lopez de Castro, J. A., Parham, P., Ploegh, H. L.& Strominger, J. L. (1979) Proc. Natl. Acad. Sci. USA 76,4395-4399.
9. Ploegh, H. L., Orr, H. T. & Strominger, J. L. (1980) Proc.Natl. Acad. Sci. USA 77, 6081-6085.
10. Bregegere, F., Abastado, J. P., Kvist, S., Rask, L., Lalanne,J. L., Garoff, H., Cami, B., Wiman, K., Larhammar, D.,Peterson, P. A., Gachelin, G., Kourilsky, P. & Dobberstein,B. (1981) Nature (London) 292, 78-81.
11. Coligan, J. E., Kindt, T. J., Uehara, H., Martinko, J. &Nathenson, S. G. (1981) Nature (London) 291, 35-39.
12. Kvist, S., Bregegere, F., Rask, L., Cami, B., Garoff, H.,Daniel, F., Wiman, K., Larhammar, D., Abastado, J. P.,
Proc. Natl. Acad. Sci. USA 83 (1986)

Proc. Natl. Acad. Sci. USA 83 (1986) 921
Gachelin, G., Peterson, P. A., Dobberstein, B. & Kowilsky, P.(1981) Proc. Natl. Acad. Sci. USA 78, 2772-2776.
13. Noda, M., Takahashi, H., Tanabe, T., Toyosato, M.,Kikyotani, X., Furutani, Y., Hirose, T., Takashima, H.,Inayama, S., Miyata, T. & Numa, S. (1983) Nature (London)302, 528-532.
14. MacLennan, D. H. & Reithmeier, R. A. F. (1982) in Mem-branes and Transport, ed. Martonosi, A. N. (Plenum, NewYork), pp. 567-571.
15. Foster, D. L., Boublik, M. & Kaback, H. R. (1983) J. Biol.Chem. 258, 31-34.
16. Engelman, D. M. & Steitz, T. A. (1981) Cell 23, 411-422.17. Engelman, D. M. & Zaccai, G. (1980) Proc. Natl. Acad. Sci.
USA 77, 5894-5898.18. Terwilliger, T. C., Weissman, L. & Eisenberg, D. (1981)
Biophys. J. 37, 353-361.19. Argos, P., Rao, J. K. M. & Hargrave, P. A. (1982) Eur. J.
Biochem. 128, 565-575.20. Finer-Moore, J. & Stroud, R. M. (1984) Proc. Natl. Acad. Sci.
USA 81, 155-159.21. Jardetzky, 0. N. (1966) Nature (London) 211, 269-270.22. Singer, S. J. (1974) Annu. Rev. Biochem. 43, 805-833.23. Klingenberg, M. (1981) Nature (London) 290, 449-453.24. Kyte, J. (1981) Nature (London) 292, 201-204.25. Ghelis, C. & Yon, J. (1982) Protein Folding (Academic, New
York).26. Zimmerman, S. S. & Scheraga, H. A. (1976) Macromolecules
9, 408-416.27. Sarkar, S. K., Young, P. E., Sullivan, C. E. & Torchia, D. A.
(1984) Proc. Natl. Acad. Sci. USA 81, 4800-4803.28. Deber, C. M., Madison, V. & Blout, E. R. (1976) Acc. Chem.
Res. 9, 106-113.29. Engelman, D. M., Goldman, A. & Steitz, T. A. (1982) Meth-
ods Enzymol. 88, 81-88.30. Keniry, M. A., Gutowsky, H. S. & Oldfield, E. (1984) Nature
(London) 307, 383-386.31. Noda, M., Shimizu, S., Tanabe, T., Takai, T., Kayano, T.,
Ikeda, T., Takahashi, H., Nakayama, H., Kanaoka, Y.,Minamino, N., Kangawa, K., Matsuo, H., Raftery, M. A.,Hirose, T., Inayama, S., Hayashida, H., Miyata, T. & Numa,S. (1984) Nature (London) 312, 121-127.
32. Hesse, J. E., Wieczorek, L., Altendorf, K., Reicin, A. S.,Dorus, E. & Epstein, W. (1984) Proc. Natl. Acad. Sci. USA81, 4746-4750.
33. Buchel, D. E., Gronenborn, B. & Muller-Hill, B. (1980) Na-ture (London) 283, 541-545.
34. Mao, D., Wachter, E. & Wallace, B. A. (1982) Biochemistry21, 4960-4968.
35. Senior, A. E. (1983) Biochim. Biophys. Acta 726, 81-95.36. Brock, C. J., Tanner, M. J. A. & Kempf, C. (1983) Biochem-
istry 22, 577-586.37. Hargrave, P. A., McDowell, J. H., Curtis, D. R., Wong,
J. K., Juszczak, E., Fong, S.-L., Rao, J. K. M. & Argos, P.(1983) Biophys. Struct. Mech. 9, 235-244.
38. Widger, W. R., Cramer, W. A., Herrmann, R. G. & Trebst,A. (1984) Proc. Natl. Acad. Sci. USA 81, 674-678.
39. Czernilofski, A. P., Levinson, A. D., Varmus, H. E., Bishop,J. M., Tischer, E. & Goodman, H. E. (1980) Nature (London)287, 198-203.
40. Tomita, K., Furthmajr, H. & Marchesi, V. T. (1978) Biochem-istry 17, 4756-4770.
41. Frank, G., Brunner, J., Hauser, H., Wacker, H., Semenza, G.& Zuber, H. (1978) FEBS Lett. 96, 183-188.
42. Von Wezenbeck, P. M. G., Hulsebos, P. M. J. &Schoennmakers, J. G. G. (1980) Gene 11, 129-148.
43. Snell, D. T. & Offord, R. E. (1972) Biochem. J. 127, 167-178.44. Nakashima, Y., Frangione, B., Wiseman, R. & Konigsberg,
W. (1981) J. Biol. Chem. 256, 5792-5797.45. Anilionis, A., Wunner, W. H. & Curtis, P. J. (1981) Nature
(London) 294, 275-278.46. Garoff, H., Frischauf, A. M., Simons, K., Lehrach, H. &
Delius, H. (1980) Nature (London) 288, 236-241.47. Rice, C. M. & Straus, J. H. (1981) Proc. Natl. Acad. Sci. USA
78, 2062-2066.48. Hedrick, S. M., Nielsen, E. A., Kavaler, J., Cohen, D. I. &
Davis, M. M. (1984) Nature (London) 308, 153-158.
49. Rose, J. K., Welch, W. J., Sefton, B. M., Esch, F. S. & Ling,N. C. (1980) Proc. Natl. Acad. Sci. USA 77, 3884-3888.
50. Henderson, R. & Unwin, N. (1975) Nature (London) 257,28-32.
51. Ross, A. H., Radhakrishnan, R., Robson, R. J. & Khorana,H. G. (1982) J. Biol. Chem. 257, 4152-4161.
52. Hargrave, P. A. (1982) Prog. Retinal Res. 1, 1-51.53. Danho, W., Makofske, R., Humiec, F., Gabriel, T. F., Car-
rasco, N. & Kaback, H. R. (1985) in Proceedings of the NinthAmerican Peptide Symposium, eds., Deber, C. M., Hruby,V. J. & Kopple, K. D. (Pierce Chemical, Rockford, IL), pp.59-62.
54. Mishina, M., Tobimatsu, T., Imoto, K., Tanaka, K., Fujita,Y., Fukuda, K., Kurasaki, M., Takahashi, H., Morimoto, Y.,Hirose, T., Inayama, S., Takahashi, T., Kuno, M. & Numa, S.(1985) Nature (London) 313, 364-369.
55. Meek, J. L. (1980) Proc. Natl. Acad. Sci. USA 77, 1632-1636.56. Rose, G. D. & Roy, S. (1980) Proc. Natl. Acad. Sci. USA 77,
4643-4647.57. Hopp, T. P. & Woods, K. R. (1981) Proc. Natl. Acad. Sci.
USA 78, 3824-3828.58. Argos, P., Rao, J. K. M. & Hargrave, P. A. (1982) Eur. J.
Biochem. 128, 565-575.59. Kyte, J. & Doolittle, R. F. (1982) J. Mol. Biol. 157, 105-132.60. Brandl, C. J., Green, N. M., Korczak, B. & MacLennan,
D. H. (1985) Cell, in press.61. MacLennan, D. H., Brandl, C. J., Korczak, B. & Green,
N. M. (1985) Nature (London) 316, 696-700.62. Yamamoto, T., Davis, C. G., Brown, M. S., Schneider, W. J.,
Casey, M. L., Goldstein, J. L. & Russell, D. W. (1984) Cell39, 28-37.
63. Ullrich, A., Bell, J. R., Chen, E. Y., Herrera, R., Petruzzelli,L. M., Dull, T. J., Gray, A., Coussens, L., Liao, Y.-C.,Tzubokawa, M., Mason, A., Seeburg, P. H., Grunfeld, C.,Rosen, 0. M. & Ramachandran, J. (1985) Nature (London)313, 756-761.
64. Ullrich, A., Coussens, L., Hayflick, J. S., Dull, T. J., Gray,A., Tam, A. W., Lee, J., Yarden, Y., Libermann, T. A.,Schlessinger, J., Downward, J., Mayes, E. L. V., Whittle, N.,Waterfield, M. D. & Seeburg, P. H. (1984) Nature (London)309, 418-425.
65. Lewin, R. (1985) Science 227, 500-503.66. Wallace, B. A. (1982) Methods Enzymol. 88, 447-463.67. Chou, P. Y. & Fasman, G. D. (1978) Adv. Enzymol. Relat.
Areas Mol. Biol. 47, 45-148.68. Schultz, G. E. & Schirmer, R. H. (1979) Principles ofProtein
Structure (Springer, New York).69. Madison, V. (1977) Biopolymers 16, 2671-2692.70. Creighton, T. E. (1983) Proteins: Structures and Molecular
Principles (Freeman, New York).71. Smith, J. A. & Pease, L. G. (1980) CRC Crit. Rev. Biochem. 8,
315-399.72. Lewis, P. N., Momany, F. A. & Scheraga, H. A. (1971) Proc.
Natl. Acad. Sci. USA 68, 2293-2297.73. Rose, G. D., Young, W. B. & Gierasch, L. M. (1983) Nature
(London) 304, 654-657.74. Kennedy, S. (1978) J. Membr. Biol. 42, 265-279.75. Steitz, T. A., Goldman, A. & Engelman, D. H. (1982)
Biophys. J. 37, 124-125.76. Brandts, J. F., Halvorson, H. R. & Brennan, M. (1975) Bio-
chemistry 14, 4953-4963.77. Lin, L. N. & Brandts, J. F. (1983) Biochemistry 22, 559-563.78. Lin, L. N. & Brandts, J. F. (1983) Biochemistry 22, 564-573.79. Lin, L. N. & Brandts, J. F. (1983) Biochemistry 22, 553-559.80. Galardy, R. E., Alger, J. R. & Liakopoulou-Kyriakides, M.
(1982) Int. J. Pept. Protein Res. 19, 123-132.81. Veis, A. & Nawrot, C. F. (1970) J. Am. Chem. Soc. 92,
3910-3914.82. Torchia, D. A., deCorato, A., Wong, S. C. K., Deber, C. M.
& Blout, E. R. (1972) J. Am. Chem. Soc. 94, 609-615.83. Deber, C. M. & Lannigan, D. A. (1983) Biophys. J. 41, 380a
(abstr.).84. Singer, S. J. (1977) J. Supramol. Struct. 6, 313-323.85. Henry, G. D., Weiner, J. H. & Sykes, B. D. (1985)Biophys. J.
47, 338a (abstr.).
Biochemistry: Brandl and Deber









![Identification of hydroxylated proline residues in human ... fileand the formation of elastin from tropoelastin is reduced by overhydroxylation of the proline residues [2]. The degree](https://img.pdfslide.us/doc/110x75/5d4b6a0588c99329638bb59d/identification-of-hydroxylated-proline-residues-in-human-the-formation-of-elastin.jpg)



















