Embed Size (px)
Citation preview

Human Nuclear Respiratory Factor 2aSubunit cDNA: Isolation, Subcloning,
Sequencing, and In Situ Hybridization ofTranscripts in Normal and Monocularly
Deprived Macaque Visual System
AILI GUO, FENG NIE, AND MARGARET WONG-RILEY*
Department of Cell Biology, Neurobiology, and Anatomy, Medical College of Wisconsin,Milwaukee, Wisconsin 53226
ABSTRACTNuclear respiratory factor 2 (NRF-2) has been shown to contribute to the transcriptional
regulation of a number of subunits of respiratory chain enzymes, including cytochrome c oxidase(CO). Our recent study demonstrated a parallel distribution of the a subunit proteins of NRF-2(NRF-2a) with CO in the monkey striate cortex, and that it can be regulated by neuronal activity.To determine whether this regulation is at the transcriptional level, the present study examinedthe expression of NRF-2a mRNA in normal and monocularly deprived adult monkeys. A partialNRF-2a cDNA was isolated from a human brain cDNA library. Sequence analysis revealed thatit shared 99% identity with the published sequence from human HeLa cells. Riboprobes ofNRF-2a was generated and labeled with digoxigenin-11-UTP for in situ hybridization. Theexpression pattern of NRF-2a mRNA in the normal striate cortex paralleled that of CO activity.It was highly expressed in layers IVC and VI, which contained high levels of CO, and moredensely expressed in puffs of layers II and III than in interpuffs. In monkeys monocularly treatedwith tetrodotoxin for 1 day to 2 weeks, both NRF-2a expression and CO activity were reduced indeprived ocular dominance columns of the visual cortex and in deprived layers of the lateralgeniculate nucleus. These data indicate that, in the normal and visually deprived adult monkeys,NRF-2a is regulated by neuronal activity at the transcriptional level. J. Comp. Neurol. 417:221–232, 2000. © 2000 Wiley-Liss, Inc.
Indexing terms: transcription factor; cytochrome oxidase; tetrodotoxin; visual cortex; lateral
geniculate nucleus
Cytochrome c oxidase (CO, ferrocytochrome C, oxygenoxidoreductase, EC 1.9.3.1), a mitochondrial energy-generating enzyme, catalyzes the transfer of electronsfrom reduced cytochrome c to molecular oxygen, generat-ing adenosine triphosphate (ATP) through the coupledprocess of oxidative phosphorylation (Wikstrom et al.,1981; Hatefi, 1985; Kadenbach et al., 1987). There is atight coupling between neuronal activity and oxidativemetabolism, and neuronal activity controls energy expen-diture (Lowry, 1975). Thus, CO has served as a sensitiveindicator of metabolism and the relative levels of energy-dependent neuronal activity averaged over a period oftime (reviewed in Wong-Riley, 1989; Wong-Riley et al.,1998a, b). The transcriptional regulation of CO biosynthe-sis in response to changing neuronal activity is of partic-ular interest, since the assembly of CO holoenzyme re-
quires a perfect coordination of its 13 subunits encoded byboth the nuclear and mitochondrial genomes. A transcrip-tion factor or factors that can coordinate the bigenomicexpression of CO subunits would be ideal. Several tran-scription factors were studied (Herdegen et al., 1993,1995; Herdegen and Leah, 1997; Chaudhuri et al., 1995),and the significance of nuclear respiratory factor 2
Grant sponsor: NIH; Grant number: R01 EY05439.* Correspondence to: Margaret Wong-Riley, PhD, Department of Cell
Biology, Neurobiology, and Anatomy, Medical College of Wisconsin, 8701Watertown Plank Road, Milwaukee, WI 53226. E-mail: [email protected]
Received 15 June 1999; Revised 26 October 1999; Accepted 27 October1999
THE JOURNAL OF COMPARATIVE NEUROLOGY 417:221–232 (2000)
© 2000 WILEY-LISS, INC.

(NRF-2) in this event has been highlighted by recent find-ings.
NRF-2, a transcription factor of the Ets family, was firstidentified during a mutational analysis of the promoterregion of the rat nuclear-encoded cytochrome c oxidasesubunit IV (RCO IV) (Virbasius and Scarpulla, 1991).NRF-2 is the human homolog of mouse GA-binding pro-tein (GABP) (Gugneja et al., 1995) and recognizes theGGAA core motif (Virbasius et al., 1993). Five distinctpolypeptides have been identified (a, b1, b2, g1, and g2),among which only the a subunit can bind DNA and het-erodimerizes with b or g subunits to form distinct DNA-protein complexes in various promoters (Virbasius andScarpulla, 1991, 1994; Virbasius et al., 1993; Scarpulla,1997). The various b and g subunits are splice variants.Other than the RCO IV gene, four NRF-2 sites arranged intandem were found to be required for maximal expressionof the mouse nuclear-encoded CO Vb promoter (Virbasiuset al., 1993), and functional NRF-2 sites were revealed inthe nuclear-encoded CO VIIaL gene in vitro (Seelan et al.,1996). Not only is it involved in the expression of a numberof nuclear-encoded CO genes, NRF-2 also contributes tothe transcriptional expression of mitochondrial transcrip-tion factor A (mtTFA or Tfam), which is essential for thereplication and transcription of mitochondrial DNA (Fish-er et al., 1987, 1989; Fisher and Clayton, 1988; Parisi andClayton, 1991; Parisi et al., 1993; Shadel and Clayton,1993; Virbasius and Scarpulla, 1994). Thus, NRF-2 mayserve as a coordinator of expressions of CO subunits fromthe two genomes.
To understand how NRF-2 is expressed in vivo andwhether and how it responds to changing neuronal activ-ity for the regulated expression of the respiratory complex,it is of importance to evaluate how neuronal activity,transcriptional regulation, and energy metabolism arelinked. It is known that the distribution of CO activity isnot homogeneous within the primate visual cortex (Wong-Riley, 1989, 1994; Wong-Riley et al., 1989; Nie and Wong-Riley, 1996a,b), and alterations in neuronal activity byexperimental interventions gave rise to changing CO ac-tivity. Monocular blockade of retinal impulse with tetro-dotoxin (TTX) resulted in a down-regulation of CO activityin deprived regions of the monkey striate cortex (Wong-Riley et al., 1989; Hevner and Wong-Riley, 1990) and thelateral geniculate nucleus (LGN) (Wong-Riley and Carroll,1984; Liu and Wong-Riley, 1990; Hevner and Wong-Riley,1993). The regulation of this enzyme is mainly at thetranscriptional level (Hevner and Wong-Riley, 1991;Wong-Riley et al., 1997). Our recent study demonstrated aparallel distribution of the a subunit protein of NRF-2 andCO in the monkey striate cortex and that it was regulatedby neuronal activity (Nie and Wong-Riley, 1999). To de-termine whether the regulation of NRF-2a is at the tran-scriptional level, the present study aimed at isolating apartial cDNA of NRF-2a from a human brain cDNA li-brary, generating riboprobes, and examining the expres-sion of NRF-2a mRNA by in situ hybridization in thestriate cortex and the LGN of normal and monocularlydeprived adult monkeys.
MATERIALS AND METHODS
Animal tissue preparation
Adult macaque (Macaca fascicularis) monkeys wereused. All experiments were carried out in accordance with
the National Institute of Health Guide for the Care andUse of Laboratory Animals (NIH Publications No. 80–23,revised 1978). All efforts were made to minimize animalsuffering, to reduce the number of animals used, and toutilize alternatives to in vivo techniques, when available.
Animals were monocularly injected with TTX as de-scribed previously (Wong-Riley and Carroll, 1984; Wong-Riley et al., 1989). Briefly, animals were anesthetized withketamine HCl (25 mg/kg, IM), and two drops of 0.5%proparacaine hydrochloride were applied topically overthe corneas of the eyes to be injected. Each animal re-ceived an intravitreal injection of TTX (19 mg in 10 ml ofsterile water) into its left eye, close to the limbus. At thisdosage, it was previously found that axoplasmic transportwas not blocked in retinal ganglion cells of cats and ma-caque monkeys, and the effect on cytochrome oxidase ac-tivity was completely reversible even after 4 weeks of TTX(Wong-Riley and Riley, 1983; Wong-Riley et al., 1989;Wong-Riley, 1994). Postoperatively, chloromycetin oph-thalmic ointment (1%) was applied to the conjunctiva asprophylaxis. The pupillary light reflex was used as a con-venient indicator of the effects of TTX in blocking impulsetransmission. The reflex ceased about 15 minutes afterthe injection and did not return for at least 4 days in theadult macaque. Thus, for 1 day of TTX treatment, only oneinjection was given; for 1 or 2 weeks of treatment, TTXwas given twice a week. Six monkeys were injected withTTX and two for each time point. Another two normalmonkeys were used in this and other studies.
At the end of the experiments, the animals were deeplyanesthetized with ketamine hydrochloride (25 mg/kg, IM)and sodium pentobarbital (60 mg/kg, IP) and were sacri-ficed without perfusion. The fresh monkey striate cortexand LGN were quickly frozen with dry ice and stored at280°C. Frozen 10 mm cross sections were cut on a cryo-stat, thaw mounted on Superfrost Plus slides (Fisher Sci-entific, Pittsburgh, PA), and frozen at 280°C until the dayof use. Alternate serial sections were processed for COhistochemistry (Wong-Riley, 1979) and NRF-2a in situhybridization.
Isolation of NRF-2a cDNA
Oligonucleotides were synthesized as primers for isolat-ing NRF-2a from a human brain cDNA library (Strat-agene, La Jolla, CA). Synthesis was performed by OperonTechnologies (Alameda, CA 94501). Primers for NRF-2awere as follows: 59-GAGAATTATGTAGTCTCAACC-39 and59-CACAAGAGTATGAAATATTTATAA-39, correspondingto nucleotides 844–864 at the 59 end and nucleotides1677–1700 at the 39 end, respectively, of the humanNRF-2a cDNA from HeLa cells (Gugneja et al., 1995,GenBank U13044). All primers were designed to have aTm of 58° C. Synthesized primers were desalted and usedin a polymerase chain reaction (PCR) to isolate and am-plify NRF-2a. PCR was done with a hot start and 30 cyclesof denaturation (94°C), anneal (53°C), and elongation(72°C). Amplified PCR products were run on a 1.5% low-melting agarose gel, stained with ethidium bromide forsize verification, excised from the gels, and purified withthe Qiaex kit (Qiagen, Chatsworth, CA).
Subcloning and sequencing
The isolated NRF-2a cDNA was subcloned into theEcoRI site of plasmid pCRII (TA cloning kit, Invitrogen,San Diego, CA), which contained phage SP6 and T7 pro-
222 A. GUO ET AL.

moters. The plasmids contain the lacZ gene for blue/whitecolor selection and the ampicillin-resistant gene. The plas-mids with inserts were amplified in white colonies oftransformed OneShot competent cells (Invitrogen) and pu-rified on a Qiagen plasmid column, according to the man-ufacturer’s instructions. Purified plasmids were digestedwith EcoRI for size confirmation. Insert DNAs were thensequenced by the dideoxy sequencing method of Sanger etal. (1977), using either Vistra DNA Sequencer 725 or ABIPrism 377.
Riboprobe synthesis
Antisense and sense riboprobes for NRF-2a were gen-erated by in vitro transcription using sets of restrictionenzymes and RNA polymerases (EcoRV and Sp6 for thesense strand, and BamHI and T7 for the antisensestrand). Riboprobes were labeled with digoxigenin-11-UTP (Boehringer-Mannheim, Indianapolis, IN) accordingto the manufacturer’s instructions and as described pre-viously (Hevner and Wong-Riley, 1991; Wong-Riley et al.,1997).
In situ hybridization
Protocols for in situ hybridization of cryostat sections,including immunohistochemistry of digoxigenin-labeledprobes, were as described previously (Hevner and Wong-Riley, 1991; Wong-Riley et al., 1997). Solutions for in situhybridization were treated with 0.1% diethylpyrocarbon-ate (DEPC) to destroy RNase activity and autoclaved.Antisense riboprobes were used to hybridize with NRF-2amRNAs, and sense riboprobes were used as controls fornonspecific binding. Adjacent sections were used for pro-cessing NRF-2a in situ hybridization and CO histochem-istry. The specificity and intensity of in situ hybridizationresults was partially related to the fixation as well ashybridization conditions. We found that cryostat sectionspre-fixed with 4% paraformaldehyde for 10 minutes beforehybridization gave good results.
Optical density measurement and statisticalanalysis
To analyze quantitatively changes in NRF-2a tran-scripts and CO activity following different time periods ofmonocular TTX treatment, optical densities of reactionproduct were measured by means of a Zeiss Zonax MPM03 photometer (Zeiss, Thornwood, NY) attached to a Zeisscompound microscope. Two-micron spot-size readings
were taken from the cytoplasm of cells. The mean of tworeadings from different spots of the cytoplasm was takenfor each cell measured. At each survival time, 100 cellswere measured from deprived or non-deprived ocular dom-inance columns within layer IVC of the striate cortex aswell as from each of the deprived (layers 2, 3, and 5) ornon-deprived (layers 1, 4, and 6) layers of the LGN ipsi-lateral to the TTX-injected eye. To ensure uniformity,readings were taken from deprived and non-deprived re-gions of the same sections at a magnification of 2503. Agrid pattern with 10 3 10 squares in a reticule was used,such that cells located at the intersections of lines weremeasured. The background was subtracted by setting zeroover a blank area (without tissue) in each slide, and alllighting conditions, magnifications, and reference pointswere kept constant.
The number of layers in the macaque LGN varies witheccentricity: six for most of central binocular vision (twomagnocellular, four parvocellular), four for peripheralmonocular vision (two for parvocellular, two for magnocel-lular), and two for the monocular crescent (one magnocel-lular, one parvocellular) (Kaas et al., 1972; Malpeli andBaker, 1975; Malpeli et al., 1996). Serial sections of LGNwith six layers used for CO histochemistry and NRF-2 insitu hybridization in the present study represented thecentral binocular portion of the visual field. Nevertheless,data from layers 3 and 5 or from layers 4 and 6 of the LGNwere pooled, as layer 3 is continuous with layer 5 andlayer 4 is with layer 6 (Lee and Malpeli, 1994; Tzonev etal., 1995).
A one-tailed Student’s t-test for non-paired comparisons(two-sample equal variance) was used to analyze differ-ences between deprived and non-deprived layers of theLGN or ocular dominance columns within layer IVC of thestriate cortex. Densitometric readings were normalized toyield a density of 1 for non-deprived columns or layers.Results were expressed as mean 6 SD. A probability of0.05 or less was considered significant.
RESULTS
Specificity of NRF-2a cDNA
Gel electrophoresis of NRF-2a PCR product isolatedfrom human brain cDNA library showed a single band ofthe correct size (data not shown). DNA sequence analysisrevealed that our human brain NRF-2a cDNA (HB in Fig.
Fig. 1. Partial amino acid sequence deduced from our humanbrain-derived (HB) cDNA of nuclear respiratory factor 2a subunit(NRF-2a) is shown in comparison with the published HeLa cells (HH)
sequence (Gugneja et al., 1995, GenBank U13044). Amino acids arenumbered on the right. Substitutions of three amino acids were ob-served as indicated. The Ets domain is underlined.
223HUMAN NRF-2a cDNA IN MONKEY BRAIN

Figure 2
224 A. GUO ET AL.
1) shared 99% identity with the published sequence fromhuman HeLa cells (HH in Fig. 1; GenBank U13044, 1970
bp in length; Gugneja et al., 1995). It was 857 bp in length,corresponding to 844–1700 of the HeLa cells sequence(GenBank U13044) encoding amino acids 221–454 ofNRF-2a. Three nucleotides were found to be different fromthe published cDNA, leading to three amino acid substi-tutions: an A1052T transversion converting arginine toserine, a T1053A transversion converting cysteine toserine, and a T1504C transition converting valine to ala-nine. We are not certain whether these substitutions arebrain specific, or of inter-individual variations, or werePCR-induced, but they were consistent for two separatecolonies isolated.
NRF-2a mRNA: distribution and correlationwith CO activity in the visual cortex
In adjacent cross sections of the normal striate cortex, aclose match between the patterns of CO activity and
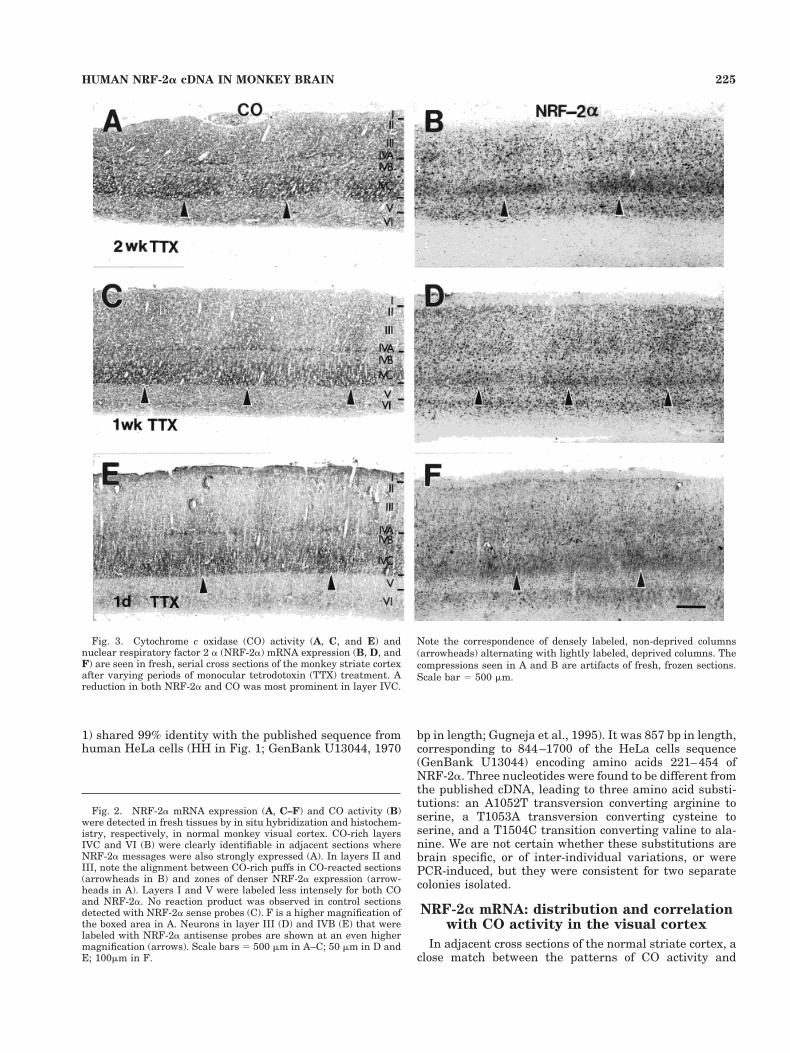
Fig. 3. Cytochrome c oxidase (CO) activity (A, C, and E) andnuclear respiratory factor 2 a (NRF-2a) mRNA expression (B, D, andF) are seen in fresh, serial cross sections of the monkey striate cortexafter varying periods of monocular tetrodotoxin (TTX) treatment. Areduction in both NRF-2a and CO was most prominent in layer IVC.
Note the correspondence of densely labeled, non-deprived columns(arrowheads) alternating with lightly labeled, deprived columns. Thecompressions seen in A and B are artifacts of fresh, frozen sections.Scale bar 5 500 mm.
Fig. 2. NRF-2a mRNA expression (A, C–F) and CO activity (B)were detected in fresh tissues by in situ hybridization and histochem-istry, respectively, in normal monkey visual cortex. CO-rich layersIVC and VI (B) were clearly identifiable in adjacent sections whereNRF-2a messages were also strongly expressed (A). In layers II andIII, note the alignment between CO-rich puffs in CO-reacted sections(arrowheads in B) and zones of denser NRF-2a expression (arrow-heads in A). Layers I and V were labeled less intensely for both COand NRF-2a. No reaction product was observed in control sectionsdetected with NRF-2a sense probes (C). F is a higher magnification ofthe boxed area in A. Neurons in layer III (D) and IVB (E) that werelabeled with NRF-2a antisense probes are shown at an even highermagnification (arrows). Scale bars 5 500 mm in A–C; 50 mm in D andE; 100mm in F.
225HUMAN NRF-2a cDNA IN MONKEY BRAIN

NRF-2a gene expression was seen (Fig. 2A,B). Sense con-trols showed only a low-level, non-specific background la-beling (Fig. 2C). The boxed area in Figure 2A is magnifiedin Figure 2F.
NRF-2a mRNA was present in all layers of the normalvisual cortex but varied in density among the layers (Fig.2A,F). It was highly expressed in layers IVC and VI,moderate in layers II, III, and IVA and rather low inlayers I, IVB, and V. Layer IVC contained a broad band ofdensely packed and mostly well-labeled cells. IVB stainedlightly in general but possessed some densely labeled so-mata. IVA was faintly visible with moderately labeledsomata, but this layer was not as striking as in CO histo-chemistry. In layers II and III, lightly expressed regionswere intermingled with moderately expressed regions,and these corresponded in space to CO-poor interpuffs andCO-rich puffs, respectively. Layer VI had numerous mod-erately to darkly labeled somata and appeared quite dis-tinct, especially for the upper part. Higher magnificationsof layers III and IVB are shown in Figure 2D and E,respectively, indicating that NRF-2a mRNA was localized
in both the cytoplasm and nuclei, but mainly in the cyto-plasm, of neurons.
Layers I and V were less densely labeled with NRF-2amRNA. The former contained sparsely labeled cell somatawith the lowest staining intensity, and the latter waslabeled at a much lower intensity than layer IVC. Thewhite matter had little or no detectable level of NRF-2amRNA.
TTX-induced reduction in NRF-2a mRNAexpression
After 2 weeks of TTX treatment, obvious decreases inNRF-2a transcripts were observed in deprived oculardominance columns of the striate cortex (Fig. 3B), whereCO activity was also reduced (Fig. 3A). These decreaseswere most prominent in layer IVC but were also detect-able in layers II, III, V, and VI. One week (Fig. 3C,D) or 1day (Fig. 3E,F) of TTX treatment induced similar patternsof down-regulation in NRF-2a transcripts and CO activity.Figure 4 shows a higher magnification of cells in non-deprived and deprived layer IV (Fig. 4A,B) and non-
Fig. 4. Higher magnifications of NRF-2a mRNA expression innon-deprived and deprived columns within layers IVC (A and B) andVI (C and D) from the same section of the striate cortex after 1 weekof monocular TTX treatment. Densely labeled cells (arrows) were
easily seen in non-deprived columns of both layers (A and C), whereasmainly lightly to moderately labeled somata (arrows) were observedin deprived columns of both layers (B and D). Scale bar 5 25 mm.
226 A. GUO ET AL.

deprived and deprived layer VI (Fig. 4C,D) after 1 week ofTTX treatment. Decreased expression of NRF-2a mRNAwas visible in neurons of the deprived layers, whereasdensely labeled somata were easily found in non-deprivedlayers (arrows in each figure).
TTX treatment also brought about a reduction inNRF-2a mRNA in deprived layers of the LGN that nor-mally received input from the TTX-treated eye. It wasmuch reduced in deprived layers 2, 3, and 5 comparedwith non-deprived layers 1, 4, and 6 in the ipsilateral LGN
Fig. 5. NRF-2a mRNA levels decreased in the deprived LGN lay-ers 2, 3, and 5 that normally received input from the ipsilateral eyetreated with TTX for 1 week (A) or 1 day (C). In comparison, thenon-deprived layers 1, 4, and 6 (asterisks) that received input fromthe contralateral, untreated eye remained densely labeled for NRF-
2a. CO activity was also down-regulated in the corresponding layersin adjacent sections of the LGN after 1 week (B) or 1 day (D) of TTXtreatment. Arrowheads in A and B denote optic disk representation.Scale bar 5 500 mm.
227HUMAN NRF-2a cDNA IN MONKEY BRAIN
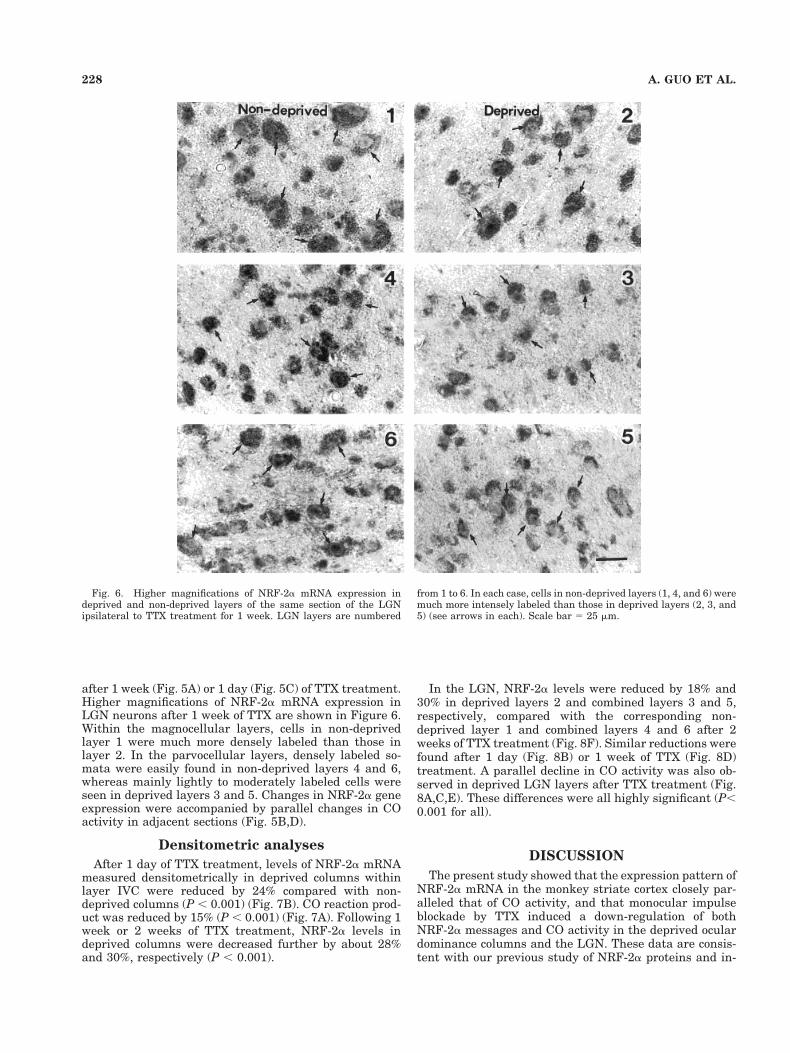
after 1 week (Fig. 5A) or 1 day (Fig. 5C) of TTX treatment.Higher magnifications of NRF-2a mRNA expression inLGN neurons after 1 week of TTX are shown in Figure 6.Within the magnocellular layers, cells in non-deprivedlayer 1 were much more densely labeled than those inlayer 2. In the parvocellular layers, densely labeled so-mata were easily found in non-deprived layers 4 and 6,whereas mainly lightly to moderately labeled cells wereseen in deprived layers 3 and 5. Changes in NRF-2a geneexpression were accompanied by parallel changes in COactivity in adjacent sections (Fig. 5B,D).
Densitometric analyses
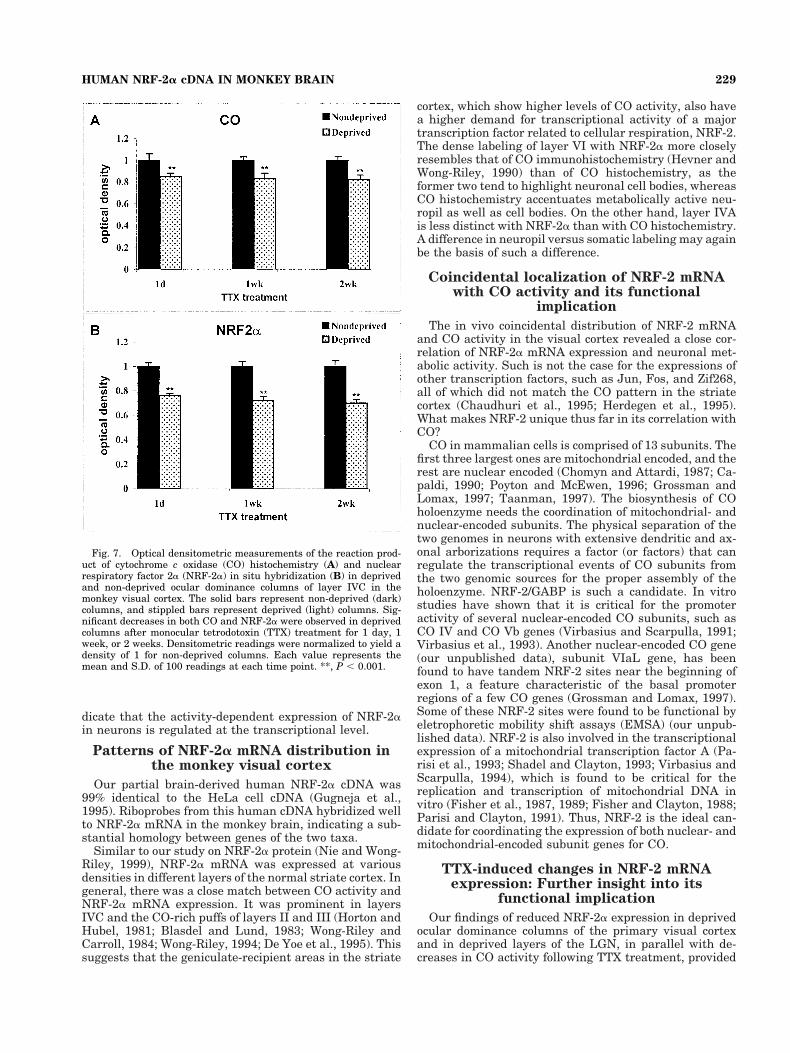
After 1 day of TTX treatment, levels of NRF-2a mRNAmeasured densitometrically in deprived columns withinlayer IVC were reduced by 24% compared with non-deprived columns (P , 0.001) (Fig. 7B). CO reaction prod-uct was reduced by 15% (P , 0.001) (Fig. 7A). Following 1week or 2 weeks of TTX treatment, NRF-2a levels indeprived columns were decreased further by about 28%and 30%, respectively (P , 0.001).
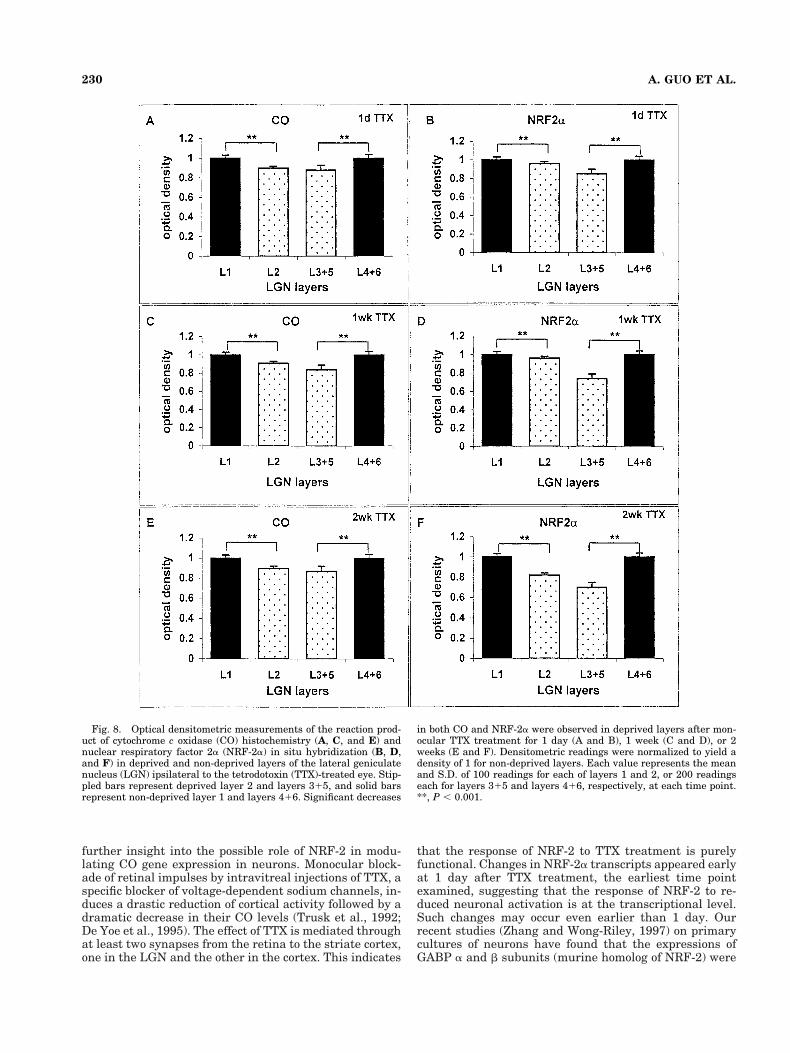
In the LGN, NRF-2a levels were reduced by 18% and30% in deprived layers 2 and combined layers 3 and 5,respectively, compared with the corresponding non-deprived layer 1 and combined layers 4 and 6 after 2weeks of TTX treatment (Fig. 8F). Similar reductions werefound after 1 day (Fig. 8B) or 1 week of TTX (Fig. 8D)treatment. A parallel decline in CO activity was also ob-served in deprived LGN layers after TTX treatment (Fig.8A,C,E). These differences were all highly significant (P,0.001 for all).
DISCUSSION
The present study showed that the expression pattern ofNRF-2a mRNA in the monkey striate cortex closely par-alleled that of CO activity, and that monocular impulseblockade by TTX induced a down-regulation of bothNRF-2a messages and CO activity in the deprived oculardominance columns and the LGN. These data are consis-tent with our previous study of NRF-2a proteins and in-
Fig. 6. Higher magnifications of NRF-2a mRNA expression indeprived and non-deprived layers of the same section of the LGNipsilateral to TTX treatment for 1 week. LGN layers are numbered
from 1 to 6. In each case, cells in non-deprived layers (1, 4, and 6) weremuch more intensely labeled than those in deprived layers (2, 3, and5) (see arrows in each). Scale bar 5 25 mm.
228 A. GUO ET AL.
dicate that the activity-dependent expression of NRF-2ain neurons is regulated at the transcriptional level.
Patterns of NRF-2a mRNA distribution inthe monkey visual cortex
Our partial brain-derived human NRF-2a cDNA was99% identical to the HeLa cell cDNA (Gugneja et al.,1995). Riboprobes from this human cDNA hybridized wellto NRF-2a mRNA in the monkey brain, indicating a sub-stantial homology between genes of the two taxa.
Similar to our study on NRF-2a protein (Nie and Wong-Riley, 1999), NRF-2a mRNA was expressed at variousdensities in different layers of the normal striate cortex. Ingeneral, there was a close match between CO activity andNRF-2a mRNA expression. It was prominent in layersIVC and the CO-rich puffs of layers II and III (Horton andHubel, 1981; Blasdel and Lund, 1983; Wong-Riley andCarroll, 1984; Wong-Riley, 1994; De Yoe et al., 1995). Thissuggests that the geniculate-recipient areas in the striate
cortex, which show higher levels of CO activity, also havea higher demand for transcriptional activity of a majortranscription factor related to cellular respiration, NRF-2.The dense labeling of layer VI with NRF-2a more closelyresembles that of CO immunohistochemistry (Hevner andWong-Riley, 1990) than of CO histochemistry, as theformer two tend to highlight neuronal cell bodies, whereasCO histochemistry accentuates metabolically active neu-ropil as well as cell bodies. On the other hand, layer IVAis less distinct with NRF-2a than with CO histochemistry.A difference in neuropil versus somatic labeling may againbe the basis of such a difference.
Coincidental localization of NRF-2 mRNAwith CO activity and its functional
implication
The in vivo coincidental distribution of NRF-2 mRNAand CO activity in the visual cortex revealed a close cor-relation of NRF-2a mRNA expression and neuronal met-abolic activity. Such is not the case for the expressions ofother transcription factors, such as Jun, Fos, and Zif268,all of which did not match the CO pattern in the striatecortex (Chaudhuri et al., 1995; Herdegen et al., 1995).What makes NRF-2 unique thus far in its correlation withCO?
CO in mammalian cells is comprised of 13 subunits. Thefirst three largest ones are mitochondrial encoded, and therest are nuclear encoded (Chomyn and Attardi, 1987; Ca-paldi, 1990; Poyton and McEwen, 1996; Grossman andLomax, 1997; Taanman, 1997). The biosynthesis of COholoenzyme needs the coordination of mitochondrial- andnuclear-encoded subunits. The physical separation of thetwo genomes in neurons with extensive dendritic and ax-onal arborizations requires a factor (or factors) that canregulate the transcriptional events of CO subunits fromthe two genomic sources for the proper assembly of theholoenzyme. NRF-2/GABP is such a candidate. In vitrostudies have shown that it is critical for the promoteractivity of several nuclear-encoded CO subunits, such asCO IV and CO Vb genes (Virbasius and Scarpulla, 1991;Virbasius et al., 1993). Another nuclear-encoded CO gene(our unpublished data), subunit VIaL gene, has beenfound to have tandem NRF-2 sites near the beginning ofexon 1, a feature characteristic of the basal promoterregions of a few CO genes (Grossman and Lomax, 1997).Some of these NRF-2 sites were found to be functional byeletrophoretic mobility shift assays (EMSA) (our unpub-lished data). NRF-2 is also involved in the transcriptionalexpression of a mitochondrial transcription factor A (Pa-risi et al., 1993; Shadel and Clayton, 1993; Virbasius andScarpulla, 1994), which is found to be critical for thereplication and transcription of mitochondrial DNA invitro (Fisher et al., 1987, 1989; Fisher and Clayton, 1988;Parisi and Clayton, 1991). Thus, NRF-2 is the ideal can-didate for coordinating the expression of both nuclear- andmitochondrial-encoded subunit genes for CO.
TTX-induced changes in NRF-2 mRNAexpression: Further insight into its
functional implication
Our findings of reduced NRF-2a expression in deprivedocular dominance columns of the primary visual cortexand in deprived layers of the LGN, in parallel with de-creases in CO activity following TTX treatment, provided
Fig. 7. Optical densitometric measurements of the reaction prod-uct of cytochrome c oxidase (CO) histochemistry (A) and nuclearrespiratory factor 2a (NRF-2a) in situ hybridization (B) in deprivedand non-deprived ocular dominance columns of layer IVC in themonkey visual cortex. The solid bars represent non-deprived (dark)columns, and stippled bars represent deprived (light) columns. Sig-nificant decreases in both CO and NRF-2a were observed in deprivedcolumns after monocular tetrodotoxin (TTX) treatment for 1 day, 1week, or 2 weeks. Densitometric readings were normalized to yield adensity of 1 for non-deprived columns. Each value represents themean and S.D. of 100 readings at each time point. **, P , 0.001.
229HUMAN NRF-2a cDNA IN MONKEY BRAIN
further insight into the possible role of NRF-2 in modu-lating CO gene expression in neurons. Monocular block-ade of retinal impulses by intravitreal injections of TTX, aspecific blocker of voltage-dependent sodium channels, in-duces a drastic reduction of cortical activity followed by adramatic decrease in their CO levels (Trusk et al., 1992;De Yoe et al., 1995). The effect of TTX is mediated throughat least two synapses from the retina to the striate cortex,one in the LGN and the other in the cortex. This indicates
that the response of NRF-2 to TTX treatment is purelyfunctional. Changes in NRF-2a transcripts appeared earlyat 1 day after TTX treatment, the earliest time pointexamined, suggesting that the response of NRF-2 to re-duced neuronal activation is at the transcriptional level.Such changes may occur even earlier than 1 day. Ourrecent studies (Zhang and Wong-Riley, 1997) on primarycultures of neurons have found that the expressions ofGABP a and b subunits (murine homolog of NRF-2) were
Fig. 8. Optical densitometric measurements of the reaction prod-uct of cytochrome c oxidase (CO) histochemistry (A, C, and E) andnuclear respiratory factor 2a (NRF-2a) in situ hybridization (B, D,and F) in deprived and non-deprived layers of the lateral geniculatenucleus (LGN) ipsilateral to the tetrodotoxin (TTX)-treated eye. Stip-pled bars represent deprived layer 2 and layers 315, and solid barsrepresent non-deprived layer 1 and layers 416. Significant decreases
in both CO and NRF-2a were observed in deprived layers after mon-ocular TTX treatment for 1 day (A and B), 1 week (C and D), or 2weeks (E and F). Densitometric readings were normalized to yield adensity of 1 for non-deprived layers. Each value represents the meanand S.D. of 100 readings for each of layers 1 and 2, or 200 readingseach for layers 315 and layers 416, respectively, at each time point.**, P , 0.001.
230 A. GUO ET AL.

down-regulated by TTX treatment and up-regulated bydepolarizing KCl treatment and were accompanied bysimilar changes in CO activity. Furthermore, this activity-dependent response of GABP occurred prior to changes inCO gene expression, as early as 2 hours for the GABP bsubunit after KCl treatment. These results strongly sup-port the notion that GABP/NRF-2, as a transcriptionalfactor, is important for activity-dependent CO gene ex-pression.
Whether and how NRF-2/GABP works as a critical sen-sor of changes in neuronal activity remains unknown.Scarpulla has proposed that NRFs (NRF-1 and/or NRF-2)and related factors contribute to respiratory chain expres-sion and function by acting on nuclear genes encodingrespiratory subunits, mitochondrial transcription and rep-lication factors, and the rate-limiting heme biosyntheticenzyme (Scarpulla, 1997). A key issue is whether and howNRFs and related factors are able to respond to extra- andintracellular signals to bring about regulated expressionof the respiratory apparatus. Recent data have providedinsight into this issue. First, trans-activation and DNAbinding functions are on different NRF-2 subunits(Gugneja et al., 1996), which raises the prospect thatregulation happens via subunit association. Second, pre-or post-transcriptional regulation may come into play. Ourpresent data are consistent with a transcriptional regula-tion. Third, NRF-2/GABP was implicated as a sensor inaltering the expression of its target genes in response tothe redox state of the cell (Martin et al., 1996). Ourpresent in vivo data are consistent with this proposal, asthe redox state of cortical cells is likely to be altered byafferent impulse blockade. Finally, NRF-2 may be in-volved in the modulation of neuronal metabolism in vivoby regulating CO expression in response to environmentalsignals. In this regard, greater energy consumption infunctionally active regions in response to environmentalstimuli leads to a local increase in the adenosinediphosphate/adenosine triphosphate (ADP/ATP) ratio,which may subsequently affect the expression and/or thebinding ability of this transcription factor (Banks et al.,1969; Erecinska and Silver, 1989) and regulate CO geneexpression. The fact that phosphorylation of NRF-1 accel-erates its DNA binding activity in vitro supported thisconcept. Such modifications have not yet been reported forNRF-2. Taken together, we can postulate that changes infunctional activity of neurons may lead to changes in thelocal expression of NRF-2/GABP (and/or related factors),which, in turn, may regulate the transcriptional activitiesof its target genes, such as CO nuclear subunits andmtTFA, via the possible mechanisms mentioned above.Consequently, CO is expressed at a higher level in areaswith greater energy demands.
Overall, the present study showed that the gene expres-sion of NRF-2a in the primary visual cortex and LGNclosely correlates with that of CO activity. Moreover, neu-ronal activity regulates local contents of NRF-2/GABP inneurons at the transcriptional level.
LITERATURE CITED
Banks P, Mangnall D, Mayor D. 1969. The re-distribution of cytochromeoxidase, noradrenaline and adenosine triphosphate in adrenergicnerves constricted at two points. J Physiol 200:745–762.
Blasdel GG, Lund JS. 1983. Termination of afferent axons in macaquestriate cortex. J Neurosci 3:1389–1413.
Capaldi RA. 1990. Structure and function of cytochrome c oxidase. AnnuRev Biochem 59:569–596.
Chaudhuri A, Matsubara JA, Cynader MS. 1995. Neuronal activity inprimate visual cortex assessed by immunostaining for the transcriptionfactor Zif268. Vis Neurosci 12:35–50.
Chomyn A, Attardi G. 1987. Mitochondrial gene products. In: Lee CP,editor. Current topics in bioenergetics, vol 15. New York: AcademicPress. p 295–329.
De Yoe EA, Trusk TC, Wong-Riley MT. 1995. Activity correlates of cyto-chrome oxidase-defined compartments in granular and supragranularlayers of primary visual cortex of the macaque monkey. Vis Neurosci12:629–639.
Erecinska M, Silver IA. 1989. ATP and brain function. J Cereb Blood FlowMetab 9:2–19.
Fisher RP, Clayton DA. 1988. Purification and characterization of humanmitochondrial transcription factor 1. Mol Cell Biol 8:3496–3509.
Fisher RP, Topper JN, Clayton DA. 1987. Promoter selection in humanmitochondria involves binding of a transcription factor to orientation-independent upstream regulatory elements. Cell 50:247–258.
Fisher RP, Parisi MA, Clayton DA. 1989. Flexible recognition of rapidlyevolving promoter sequences by mitochondrial transcription factor 1.Genes Dev 3:2202–2217.
Grossman LI, Lomax MI. 1997. Nuclear genes for cytochrome c oxidase.Biochim Biophys Acta 1352:174–192.
Gugneja S, Virbasius JV, Scarpulla RC. 1995. Four structurally distinct,non-DNA-binding subunits of human nuclear respiratory factor 2 sharea conserved transcriptional activation domain. Mol Cell Biol 15:102–111.
Gugneja S, Virbasius CM, Scarpulla RC. 1996. Nuclear respiratory factors1 and 2 utilize similar glutamine-containing clusters of hydrophobicresidues to activate transcription. Mol Cell Biol 16:5708–5716.
Hatefi Y. 1985. The mitochondrial electron transport and oxidative phos-phorylation system. Annu Rev Biochem 54:1015–1069.
Herdegen T, Leah JD. 1997. Inducible and constitutive transcription fac-tors in the mammalian nervous system: control of gene expression byJun, Fos and Krox, and CREB/ATF proteins. Brain Res Brain Res Rev28:370–490.
Herdegen T, Sandkuhler J, Gass P, Kiessling M, Bravo R, Zimmermann M.1993. JUN, FOS, KROX, and CREB transcription factor proteins in therat cortex: basal expression and induction by spreading depression andepileptic seizures. J Comp Neurol 333:271–288.
Herdegen T, Kovary K, Buhl A, Bravo R, Zimmermann M, Gass P.1995.Basal expression of the inducible transcription factors c-Jun, Jun-B,Jun-D, c-Fos, Fos-B, and Krox-24 in the adult rat brain. J Comp Neurol354:39–56.
Hevner RF, Wong-Riley M. 1990. Regulation of cytochrome oxidase proteinlevels by functional activity in the macaque monkey visual system.J Neurosci 10:1331–1340.
Hevner RF, Wong-Riley MTT. 1991. Neuronal expression of nuclear andmitochondrial genes for cytochrome oxidase (CO) subunits analyzed byin situ hybridization: comparison with CO activity and protein. J Neu-rosci 11:1942–1958.
Hevner RF, Wong-Riley MT. 1993. Mitochondrial and nuclear gene expres-sion for cytochrome oxidase subunits are disproportionately regulatedby functional activity in neurons. J Neurosci 13:1805–1819.
Horton JC, Hubel DH. 1981. Regular patchy distribution of cytochromeoxidase staining in primary visual cortex of macaque monkey. Nature292:762–764.
Kaas JH, Guillery RW, Allman JM. 1972. Some principles of organizationin the dorsal lateral geniculate nucleus. Brain Behav Evol 6:253–259.
Kadenbach B, Kuhn-Nentwig L, Buge U. 1987. Evolution of a regulatoryenzyme: cytochrome c oxidase (complex IV). In: Lee CP, editor. Currenttopics in bioenergetics, vol 15. New York: Academic Press. p 113–161.
Lee D, Malpeli JG. 1994. Global form and singularity: modeling the blindspot’s role in lateral geniculate morphogenesis. Science 263:1292–1294.
Liu S, Wong-Riley M. 1990. Quantitative light- and electron-microscopicanalysis of cytochrome-oxidase distribution in neurons of the lateralgeniculate nucleus of the adult monkey. Vis Neurosci 4:269–287.
Lowry OH. 1975. Energy metabolism in brain and its control. In: IngvarDH, Lassen NA, editors. Brain work. Alfred Benzon Symposium VII.New York: Academic Press, p 48–64.
Malpeli JG, Baker FH. 1975. The representation of the visual field in thelateral geniculate nucleus of Macaca mulatta. J Comp Neurol 161:569–594.
231HUMAN NRF-2a cDNA IN MONKEY BRAIN

Malpeli JG, Lee D, Baker FH. 1996. Laminar and retinotopic organizationof the macaque lateral geniculate nucleus: magnocellular and parvo-cellular magnification functions. J Comp Neurol 375:363–377.
Martin ME, Chinenov Y, Yu M, Schmidt TK, Yang XY. 1996. Redoxregulation of GA-binding protein-alpha DNA binding activity. J BiolChem 271:25617–25623.
Nie F, Wong-Riley MT.1996a. Metabolic and neurochemical plasticity ofgamma-aminobutyric acid-immunoreactive neurons in the adult ma-caque striate cortex following monocular impulse blockade: quantita-tive electron microscopic analysis. J Comp Neurol 370:350–366.
Nie F, Wong-Riley MT. 1996b. Mitochondrial- and nuclear-encoded sub-units of cytochrome oxidase in neurons: differences in compartmentaldistribution, correlation with enzyme activity, and regulation by neu-ronal activity. J Comp Neurol 373:139–155.
Nie F, Wong-Riley MTT. 1999. Nuclear respiratory factor-2 subunit protein:correlation with cytochrome oxidase and regulation of functional activityin the monkey primary visual cortex. J Comp Neurol 404:310–320.
Parisi MA, Clayton DA. 1991. Similarity of human mitochondrial tran-scription factor 1 to high mobility group proteins. Science 252:965–969.
Parisi MA, Xu B, Clayton DA. 1993. A human mitochondrial transcrip-tional activator can functionally replace a yeast mitochondrial HMG-box protein both in vivo and in vitro. Mol Cell Biol 13:1951–1961.
Poyton RO, McEwen J. 1996. Crosstalk between nuclear and mitochondrialgenomes. Annu Rev Biochem 65:563–607.
Sanger F, Nicklen S, Coulson AR. 1977. DNA sequencing with chain-terminating inhibitors. Proc Natl Acad Sci USA 74:5463–5467.
Scarpulla RC. 1997. Nuclear control of respiratory chain expression inmammalian cells. J Bioenerg Biomembr 29:109–119.
Seelan RS, Gopalakrishnan L, Scarpulla RC, Grossman LI. 1996. Cyto-chrome c oxidase subunit VIIa liver isoform. Characterization andidentification of promoter elements in the bovine gene. J Biol Chem271:2112–2120.
Shadel GS, Clayton DA. 1993. Mitochondrial transcriptional initiation,variation and conservation. J Biol Chem 268:16083–16086.
Taanman, J.W. 1997. Human cytochrome c oxidase: structure, function,and deficiency. J Bioenerg Biomembr 29:151–163.
Trusk TC, Wong-Riley M, DeYoe EA. 1992. Changes in cytochrome oxidaseand neuronal activity in V1 induced by monocular TTX blockade inmacaque monkeys. Soc Neurosci Abstr 18:298.
Tzonev S, Schulten K, Malpeli JG. 1995. Morphogenesis of the lateralgeniculate nucleus: how singularities affect global structure. In: Te-sauro G, Touretzky D, Leen T, editors. Advances in neural informationprocessing systems: proceedings of the 1994 Conference, vol 17. Cam-bridge, MA: MIT Press, p 133–140.
Virbasius JV, Scarpulla RC. 1991. Transcriptional activation through ETSdomain binding sites in the cytochrome c oxidase subunit IV gene. MolCell Biol 11:5631–5638.
Virbasius JV, Scarpulla RC. 1994. Activation of the human mitochondrialtranscription factor A gene by nuclear respiratory factors: a potentialregulatory link between nuclear and mitochondrial gene expression inorganelle biogenesis. Proc Natl Acad Sci USA 91:1309–1313.
Virbasius JV, Virbasius CA, Scarpulla RC. 1993. Identity of GABP withNRF-2, a multisubunit activator of cytochrome oxidase expression,reveals a cellular role for an ETS domain activator of viral promoters.Genes Dev 7:380–392.
Wikstrom M, Krab K, Saraste M. 1981. Cytochrome oxidase: a synthesis.New York: Academic Press.
Wong-Riley M. 1979. Changes in the visual system of monocularly suturedor enucleated cats demonstrable with cytochrome oxidase histochem-istry. Brain Res 171:11–28.
Wong-Riley MTT. 1989. Cytochrome oxidase: an endogenous metabolicmarker for neuronal activity. Trends Neurosci 12:94–101.
Wong-Riley MTT. 1994. Primate visual cortex: dynamic metabolic organi-zation and plasticity revealed by cytochrome oxidase. In Peters A,Rockland KS, editors. Cerebral cortex, vol 10, Primary visual cortex inprimates. New York: Plenum Press, p 141–200.
Wong-Riley MTT, Carroll EW. 1984. The effect of impulse blockage oncytochrome oxidase activity in the monkey visual system. Nature 307:262–264.
Wong-Riley M, Riley DA. 1983. The effect of impulse blockage on cyto-chrome oxidase activity in the cat visual system. Brain Res 261:185–193.
Wong-Riley MT, Trusk TC, Tripathi SC, Hoppe DA. 1989. Effect of retinalimpulse blockage on cytochrome oxidase-rich zones in the macaquestriate cortex: II. Quantitative electron-microscopic (EM) analysis ofneuropil. Vis Neurosci 2:499–514.
Wong-Riley MTT, Mullen MA, Huang, Z, Guyer C. 1997. Brain cytochromeoxidase subunit complementary DNAs: isolation, subcloning, sequenc-ing, light and electron microscopic in situ hybridization of transcripts,and regulation by neuronal activity. Neuroscience 76:1035–1055.
Wong-Riley MTT, Nie F, Hevner RF, Liu S. 1998a. Brain cytochromeoxidase: functional significance and bigenomic regulation in the CNS.In: Gonzalez-Lima F, editor. Cytochrome oxidase in neuronal metabo-lism and Alzheimer’s Disease. New York: Plenum Press, p 1–53.
Wong-Riley MTT, Huang ZF, Liebl W, Nie F, Xu HY, Zhang CY. 1998b.Neurochemical organization of the macaque retina: effect of TTX onlevels and gene expression of cytochrome oxidase and nitric oxidesynthase and on the immunoreactivity of Na1 K1 ATPase and NMDAreceptor subunit I. Vision Res 38:1455–1477.
Zhang C, Wong-Riley. 1997. Effect of depolarization on cytochrome oxidasegene expression in primary neuronal culture of rat cortex. Soc NeurosciAbstr 23:89.
232 A. GUO ET AL.





























