Embed Size (px)
Citation preview

Eur. J. Biochem. 17 (1970) 171-177
Hormone Action on Liver Adenyl Cyclase Activity The Effects of Glucagon and Fluoride on a Particulate Preparation
from Rat and Mouse Liver
K. Dieter HEPP, Roswitha EDEL, and Otto WIELAND Institut fiir Diabetesforschung, Munchen
(Received July 6/August 21, 1970)
Adenyl cyclase activity was measured by conversion of [32P]ATP to adenosine 3' : 5'-cyclic [32P]phosphate in a washed particle suspension from mouse and rat liver. The test contained an ATP regenerating system and 10 mM unlabeled Ado-3' : 5'-P to trap the labeled Ado-3' : 5'-P formed. Decreased activities were observed when methyl xanthines were used in place of un- labeled Ado-3':5'-P. The reaction was stimulated by Mg++ which increased Vmax without changing the apparent K , towards ATP. Adenyl cyclase activity was stimulated by synthetic glucagon over the range of 0.1-20 pg/ml. Removal of Ca++ by ethylene glycol-bis-(B-amino- ethyl ether)-N,N'-tetraacetic acid (EGTA) strongly augmented glucagon action, while 1 mM Ca++ was inhibitory. The data suggest enhanced transmission of the hormone signal from the (hypothetical) receptor to the catalytic enzyme unit by chelation of membrane bound calcium.
In contrast to the hyperbolic dose response curve observed with glucagon, the curve with fluoride (1 - 10 mM) was sigmoidal in shape, the effect being inhibited by 1 mM pyrophosphate. Although inhibition by Ca++ was observed, EGTA did not augment the effect of fluoride. The effects of fluoride and glucagon a t maximal concentrations were not additive, indicating stimula- tion of the same adenyl cyclase system.
Since the discovery of adenosine 3' : 5'-cyclic monophosphate (Ado-3': 5'-P) as the mediator sub- stance of the glycogenolytic effect of epinephrine and glucagon in liver by Sutherland and Rall [l], and the subsequent demonstration of adenyl cyclase activity in liver [Z], the use of labeled ATP has con- siderably improved assay systems measuring the activity of this enzyme in homogenate preparations. The recently developed method of Krishna et al. [3] in which labeled ATP is converted to Ado-3':5'-P which is then separated by ion exchange chromato- graphy and a BaOH, and ZnSO, coprecipitation, has been successfully employed by several groups [3-91. Birnbaumer et al. [4,5], and Rodbell et al. [6] have used this procedure in their thorough studies on the properties of adenyl cyclase activity in fat cell membranes and on the action of several hormones on this receptor-enzyme system.
In contrast to the work with fat cell membranes, relatively little information is available on the prop- erties of adenyl cyclase in liver preparations. I n short communications Bitensky et al. [lo] and Marinetti et al. [ll] have reported on the distribution of epi- nephrine and glucagon sensitive systems in liver,
Unusual Abbreviations. EGTA, ethylene glycol-bis- (/Laminoethyl ether) -N,N'-tetraacetic acid; Ado-3': 5'-P, adenosine 3': 5'-cyclic monophosphate.
Enzymes. Pyruvic kinase or ATP: pyruvate phospho- transferase (EC 2.7.1.40).
and Pohl et al. [12] have described glucagon and fluoride sensitivity in the plasma membrane of hepatic parenchymal cells. The present report de- scribes the adaptation of the method of Krishna to the investigation of the properties of adenyl cyclase activity in particulate preparations of mouse and rat- liver and its stimulation by glucagon and fluoride as part of a study on hormone action upon this organ.
EXPERIMENTAL PROCEDURE
Materials Synthetic glucagon was a gift of Dr. Wunsch
(Max-Planck-Institut fur EiweiB und Lederforschung, Munich). ATP (disodium salt), Ado-3' :5'-P, phos- phoenol-pyruvic acid (mono-potassium salt), ATP : pyruvate phosphotransferase were obtained from Boehringer Mannheim GmbH (Mannheim, Germany). Dowex-BOW-Xf3 (100-200 mesh, H+-Form) was from Serva, (Heidelberg, Germany). All other chemicals were from E. Merck (Darmstadt, Germany). [3H]Ado-3' : 5'-P was purchased from the New Eng- land Nuclear Corp. (Dreieichenhain, Germany). [OI-~~PIATP was from Radiochemical Centre (Amer- sham, England). [OI-~~PIATP had to be purified ac- cording to Taunton et al. [7] from an unidentified impurity which gave blank values of about 1200 counts/min under the test conditions as described below.

Eur. J. Biochem. 172 Hormone Action on Liver Adenyl Cyclase
Adenyl Cyclase Assay Adenyl cyclase activity was assayed by measuring
the conversion of [OI-~~PIATP to Ado-3' : 5'-[3zP]P, in the presence of an ATP-regenerating system and unlabeled Ado-3': 5'-P. The standard system con- tained 0.2 M Tris-C1 pH 7.5, 0.9 mM MgCl,, 0.45 mM [OL-~~PIATP (5 pC), 6 mM potassium P-pyruvate, 5 pg pyruvate kinase and enzyme preparation con- taining 100-600 pg protein. Incubations were car- ried out in 1.5 ml plastic cups (Netheler and Hinz, Hamburg-Eppendorf, Germany) in a final volume of 0.1 ml. Standard incubation time was 10 min, the temperature 33". The reaction was initiated by ad- dition of the particle suspension with an Eppendorf micropipet (Netheler and Hinz, Hamburg-Eppendorf, Germany). The incubation was stopped by heating for 3 min at 97" after addition of a mixture of ATP, and [3H]Ado-3': 5'-P [13]. Ad0-3':5'-[~~PlP was then isolated by chromatography on Dowex W-X8 and two Ba(OH),-ZnSO, coprecipitations according to Rodbell [S3] who followed essentially the method of Krishna et al. [3]. Recovery of the added [3H]Ado- 3':5'-P was between 35 and 500/,. Blank values, to which the homogenate preparation was added shortly before boiling, were counted to less than 100 counts/ min with the purified [s2P]ATP. Radioactivity was measured in a Tri-Carb scintillation spectrometer (Model 3003, Packard Instruments, Chicago), ap- propriate quench corrections were made with the use of an external standard.
Preparation of Particle Suspensions Male swiss albino mice, about 40g, were fed a
normal laboratory chow ad libitum. They were killed by cervical dislocation and the liver was rapidly excised, minced and homogenized for 3 times 10 sec, in 5 volumes of 0.2 M Tris-C1, pH 7.4, with a high speed mechanical homogenizer (Ultraturrax, Janke and Kunkel, Staufen, Germany). The homogenate waa filtered through a nylon mesh and centrifuged for 20 min at 40000 x g in a refrigerated centrifuge. After completely decanting the supernatant, the first pellet contained 500/, of the total liver protein and more than 900/, of the total particulate material (the 155 000 x g x 20 min precipitate). The precipitate was then resuspended and washed three times in the same volume of buffer. The final suspension was prepared by hand in a glasslteflon homogenizer, with 0.02 M Tris-C1. The protein content of this prepara- tion was about 40°/, of the total protein. Protein concentration was adjusted to approx. I0 mglml. In some experiments, the livers of male Sprague-Dawley rats, fed ad libitum were used and prepared ac- cordingly. Protein was determined by a modification of the biuret method which was adapted for mem- branous material [13a] : 50 p1 homogenate suspension were shaken mechanically for 5 min with 1 ml of
dioxane. After centrifugation for 1 min a t 12000 xg, the supernatant was decanted and replaced by 1 ml 0.3 M trichloroacetic acid, which again was shaken, centrifuged, and decanted. The precipitate was dis- solved in 1 ml of biuret reagent under shaking for 30 min. Absorbance was measured in a 1-cm cuvette at 546 nm. Crystalline albumin (Behringwerke-Mar- burg, Germany), served as protein standard.
Calcium content of particle suspensions was measured in a Leitz-Unicam atomic absorption spectrometer after precipitation of protein with 5 01,
trichloracetic acid.
RESULTS Studies on the Assay System
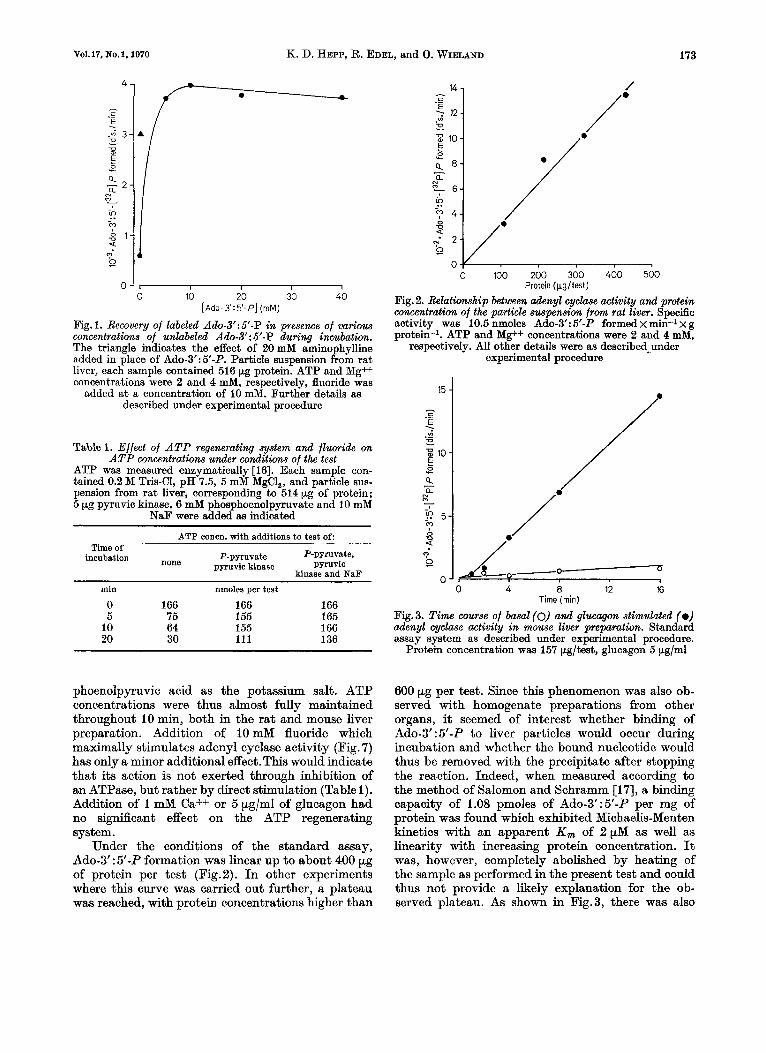
Although Pohl et al. [12] have demonstrated a hormone sensitive adenyl cyclase of high specific activity in the plasma membrane of rat liver, it is not known whether there are other subcellular com- ponents with adenyl cyclase activity in this organ. The particle preparation employed in the present work was therefore chosen to comprise most of the particulate material of the liver after washing away the soluble protein which contains most of the phos- phodiesterase activity [14]. Preliminary studies of different homogenization techniques and different buffers showed the highest yield in adenyl cyclase activity after high speed mechanical homogenization in 0.2 M Tris-C1 at pH 7.4. In comparison with the total homogenate, a six-fold increase in specific en- zyme activity was observed. In spite of the extensive washing, a residual phosphodiesterase activity re- mained with this particle preparation which cor- responded with specific activities of particulate fractions of rat liver as shown in an earlier report from this laboratory [14]. In order to prevent break- down of labeled Ado-3':5'-P in the reaction, theo- phylline, or the better soluble aminophylline were added to the incubation system at concentrations of 10 and 20 mM, respectively. However, further ex- periments which included high concentrations of un- labeled Ado-3' : 5'-P in the assay system [15], revealed that both methyl xanthines were either inhibiting the adenyl cyclase (see also [12]) or not completely blocking phosphodiesterase activity in the present system. Thus, as derived from Fig.1, 10 mM Ado- 3' : 5'-P seemed the appropriate concentration and was routinely used in the test. The figure shows furthermore, that there was no product inhibition with concentrations up to 40 mM Ado-3':5'-P. Under the conditions of the test, ATP is rapidly lost as shown in Table 1. In order to maintain ATP concen- trations throughout the incubation period, an ATP regenerating system as previously employed by several other groups [4, SO, 12,151 had to be included. Such regeneration of ATP from ADP was achieved by addition of 5 pg pyruvic kinase and 6 mM phos-
Vol.17, No.l.1970 K. D. HEPP. R. EDEL, and 0. WIELAND
5 12- ." U - P 10- E 8 Q 8 - a
L
- 6 -
In
? 4 - V
-2 2 -
173
I I I 1 0 10 20 30 40
[Ado-3' :5 ' -P] (mM)
Fig. 1. Recovery of labeled Ado-3': 5'-P in presence of various concentrations of unlabeled Ado-3': 5'-P during incubation. The triangle indicates the effect of 20 mM aminophylline added in place of Ado-3':5'-P. Particle suspension from rat liver, each sample contained 516 pg protein. ATP and Mg++ coiicentrations were 2 and 4 mM, respectively, fluoride was
added at a concentration of 10 mM. Further details as described under experimental procedure
Table 1. Effect of A T P regenerating system and fluoride on A T P concentrations under conditions of the test
ATP was measured enzymatically [16]. Each sample con- tained 0.2 M Tris-C1, pH 7.5, 5 mM MgCI,, and particle sus- pension from rat liver, corresponding to 514 pg of protein; 5 pg pyruvic kinase, 6 mM phosphoenolpyruvate and 10 mM
NaF were added as indicated
ATP concn. with additions to test o f
none pyruvic kinase pyruvic
Time of incubation P-pyruvate P-pyruvate,
kinase and NaF
min nmoles per test
0 166 166 166 5 75 155 165
10 64 155 166 20 30 111 136
phoenolpyruvic acid as the potassium salt. ATP concentrations were thus almost fully maintained throughout I0 min, both in the rat and mouse liver preparation. Addition of 10 mM fluoride which maximally stimulates adenyl cyclase activity (Fig. 7) has only a minor additional effect. This would indicate that its action is not exerted through inhibition of an ATPase, but rather by direct stimulation (Table 1). Addition of I mM Ca++ or 5 pg/ml of glucagon had no significant effect on the ATP regenerating system.
Under the conditions of the standard assay, Ado-3' : 5'-P formation was h e a r up to about 400 pg of protein per test (Fig.2). I n other experiments where this curve was carried out further, a plateau was reached, with protein concentrations higher than
Y------ 0 0 100 200 300 400 500
Protein (pgltest)
Pig. 2. Relationship between adenyl cyclme activity and protein concentration of the particle suapenaion from rat liver. Specific activity was 10.5 nmoles Ado-3': 5'-P formed x min-l x g protein-1. ATP and Mg++ concentrations were 2 and 4 mM,
respectively. All other details were as described,.under experimental procedure
0 J p ( b _ , - - o Y 0 4 8 12 16 I
Time (min)
Pig.3. Time wurse of basal (0) and q l w q o n stimulated (0) adenyl cyclase activity in mouse liver preparation. Standard assay system as described under experimental procedure.
Protein concentration waa 157 pgltest, glucagon 5 pg/ml
600 pg per test. Since this phenomenon was also ob- served with homogenate preparations from other organs, it seemed of interest whether binding of Ado-3':5'-P to liver particles would occur during incubation and whether the bound nucleotide would thus be removed with the precipitate after stopping the reaction. Indeed, when measured according to the method of Salomon and Schramm [17], a binding capacity of 1.08 pmoles of Ado-3':5'-P per mg of protein was found which exhibited Michaelis-Menten kinetics with an apparent K , of 2pM as well as linearity with increasing protein concentration. It was, however, completely abolished by heating of the sample as performed in the present test and could thus not provide a likely explanation for the ob- served plateau. As shown in Fig.3, there was also
174 Hormone Action on Liver Adenyl Cyclase Em. J. Biochem.
100
. - 50
- 2 4 6 l/[ATP] (rnM-')
I
4 6 l/[ATP] (rnM-')
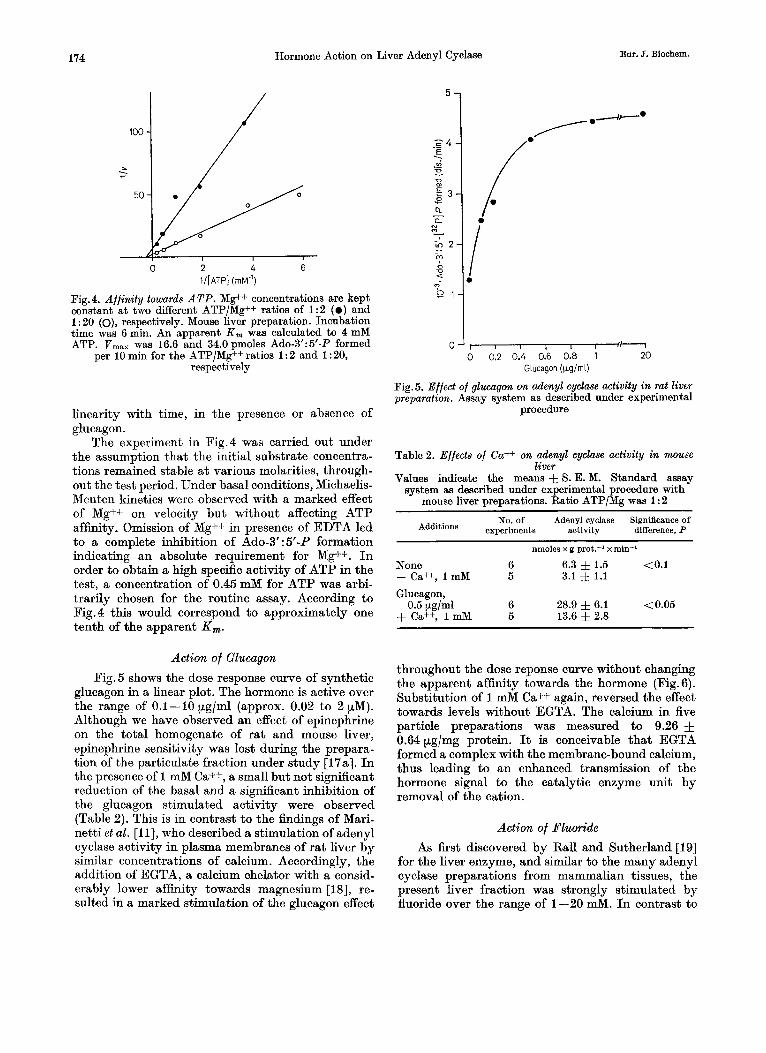
Big.4. Affinity towards ATP. Mg++ concentrations are kept constant at two different ATP/Mg++ ratios of 1 : 2 ( 0 ) and 1 : 20 (o), respectively. Mouse liver preparation. Incubation time was 6 min. An apparent K , was calculated to 4 mM ATP. Vmax was 16.6 and 34.0pmoles Ado-3':5'-P formed
per 10 min for the ATP/Mg++ ratios 1 : 2 and 1 : 20, respectively
linearity with time, in the presence or absence of glucagon.
The experiment in Fig.4 was carried out under the assumption that the initial substrate concentra- tions remained stable a t various molarities, through- out the test period. Under basal conditions, Michaelis- Menten kinetics were observed with a marked effect of Mg++ on velocity but without affecting ATP affinity. Omission of Mg++ in presence of EDTA led to a complete inhibition of Ado-3':5'-P formation indicating an absolute requirement for Mg++. I n order to obtain a high specific activity of ATP in the test, a concentration of 0.45 mM for ATP was arbi- trarily chosen for the routine assay. According to Fig. 4 this would correspond to approximately one tenth of the apparent K,.
Action of Glucagon Fig. 5 shows the dose response curve of synthetic
glucagon in a linear plot. The hormone is active over the range of 0.1-10 pglml (approx. 0.02 to 2 pM). Although we have observed an effect of epinephrine on the total homogenate of rat and mouse liver, epinephrine sensitivity was lost during the prepara- tion of the particulate fraction under study [17a]. I n the presence of 1 mM Ca++, a small but not significant reduction of the basal and a significant inhibition of the glucagon stimulated activity were observed (Table 2). This is in contrast to the findings of Mari- netti et al. [ill, who described a stimulation of adenyl cyclase activity in plasma membranes of rat liver by similar concentrations of calcium. Accordingly, the addition of EGTA, a calcium chelator with a consid- erably lower affinity towards magnesium [18], re- sulted in a marked stimulation of the glucagon effect
I l l 1 I I d * 0 0.2 0.4 0.6 0.8 1 20
Glucagon (Wglrnl)
Fig.5. Effect of glucagon on adenyl cyclme activity in rat liver preparation. Assay system as described under experimental
procedure
Table 2. Effects of Ca++ on adenyl cyclase activity in m u s e liver
Values indicate the means f S. E. M. Standard assay system as described under experimental procedure with
mouse liver preparations. Ratio ATP/Mg was 1 : 2
Additions experiments activity difference, P No. of Adenyl cyclase Significance of
nmoles x g prot.-' x min-'
None 6 6.3 f 1.5 <0.1 + Ca++, 1 mM 5 3.1 -J= 1.1
Glucagon,
+ Ca++, 1 mM 5 13.6 f 2.8 0.5 pg/ml 6 28.9 & 6.1 < 0.05
throughout the dose reponse curve without changing the apparent affinity towards the hormone (Fig. 6). Substitution of 1 mM Ca++ again, reversed the effect towards levels without EGTA. The calcium in five particle preparations was measured to 9.26 f 0.64pg/mg protein. It is conceivable that EGTA formed a complex with the membrane-bound calcium, thus leading to an enhanced transmission of the hormone signal to the catalytic enzyme unit by removal of the cation.
Action of Fluoride As first discovered by Rall and Sutherland [19]
for the liver enzyme, and similar to the many adenyl cyclase preparations from mammalian tissues, the present liver fraction was strongly stimulated by fluoride over the range of 1-20 mM. I n contrast to
Vol. 17, No. 1, 1970 K. D. HEPP, R. EDEL, and 0. WIELAND 175
2o 1
i
0 0.1 0.2 0.5 1.0 Glucagon ( L g l m l )
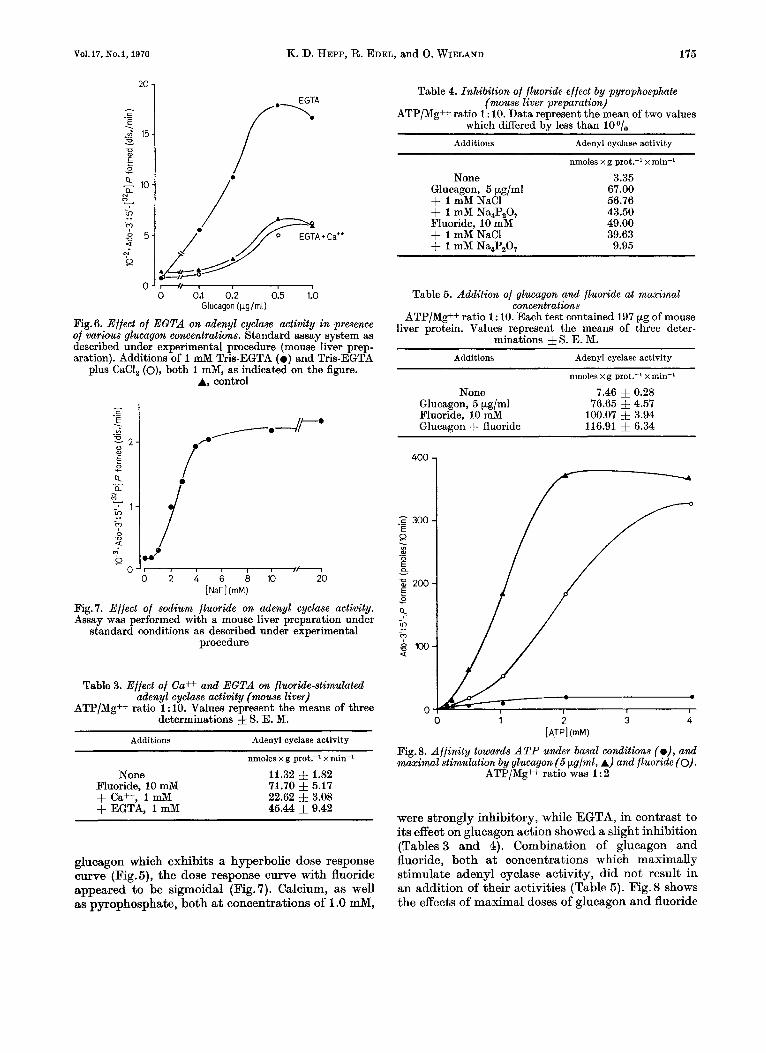
Fig.6. Effect of E G T A on adenyl cyclase activity in presence of various glucagon concentrations. Standard assay system as described under experimental procedure (mouse liver prep- aration). Additions of 1 mM Tris-EGTA ( 0 ) and Tris-EGTA
plus CaCI, (o), both 1 mM, as indicated on the figure. A, control
? .. Fig. 7. Effect of sodium fluoride on adenyl cyclase activity. Assay was performed with a mouse liver preparation under
standard conditions as described under experimental procedure
Table 3. Effect of Ca++ and E G T A on fluoride-stimulated d e n y 1 cyclase activity ( m u s e liver)
ATP/Mg++ ratio 1: 10. Values represent the means of three determinations f S. E. M.
Additions Adenyl cyclase activity
nmoles x g prot.-' x min-'
None 11.32 f 1.82 71.70 f 5.17 22.62 f 3.08 45.44 f 9.42
Fluoride, 10 mM + Ca++, 1 mM + EGTA, 1 mM
glucagon which exhibits a hyperbolic dose response curve (Fig.5), the dose response curve with fluoride appeared to be sigmoidal (Fig.7). Calcium, as well as pyrophosphate, both at concentrations of 1.0 mM,
Table 4. Inhibition of fluoride effect by pyrophosphate (mouse liver preparation)
ATP/Mg++ ratio 1 : 10. Data represent the mean of two values which differed bv less than loo/.
Additions Adcnyl cyclase activity
None Glucagon, 5 pg/ml + 1 mM NaCl + 1 mM Na,P,O, Fluoride, 10 mM + 1 mM NaCl + 1 mM Na,P,O,
nmoles xg prot.? x min-'
3.35 67.00 56.76 43.50 49.00 39.63
9.95
Table 5. Addition of glucugon and fluoride at maximal concentrations
ATP/Mg++ ratio 1 : 10. Each test contained 197 pg of mouse liver protein. Values represent the means of three deter-
minations & S. E. M. Additions Adenyl cyclase activity
nmoles x g prot.-' x min-a
None 7.46 f 0.28 76.65 5 4.57
100.07 & 3.94 116.91 f 6.34
Glucagon, 5 pg/ml Pluoride, 10 mM Glucagon + fluoride
400
._ F 300 E 0 . v)
0 - a2 E - z 200 E z - .?-
S r n -5 ?
0 0 1 2 3 4
[ATPI (mM)
Fig.8. Affinity towards ATP under basal conditions (e), and maximal stimulation by glucagon (5 pg/ml, A) and fluoride (0).
ATP/Mg++ ratio was 1 : 2
were strongly inhibitory, while EGTA, in contrast to its effect on glucagon action showed a slight inhibition (Tables 3 and 4). Combination of glucagon and fluoride, both at concentrations which maximally stimulate adenyl cyclase activity, did not result in an addition of their activities (Table 5). Fig. 8 shows the effects of maximal doses of glucagon and fluoride
176 Hormone Action on Liver Adenyl Cyclase Eur. J. Biochem.
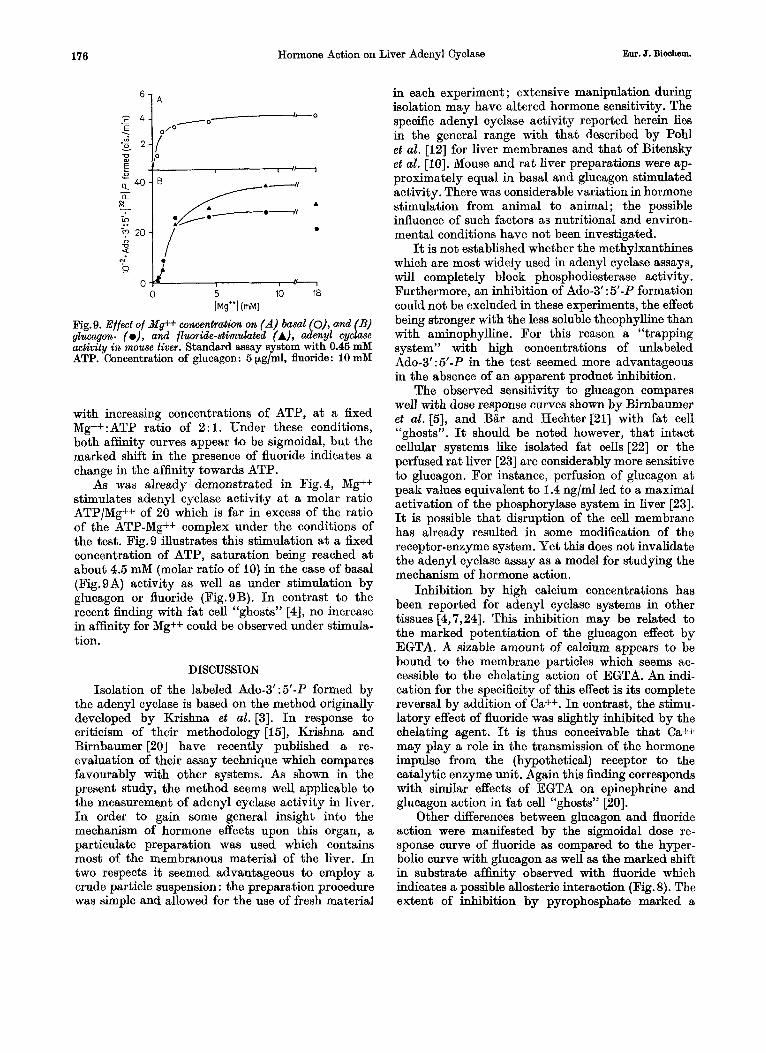
6 1 A
0 5 10 18 h+’] (mM)
Fig.9. Effect of Mg++ concentration on (A) basal (O), and (B) glucagon- (o), and fluoride-stimulated (A), adenyl cyclase activity in mouse liver. Standard assay system with 0.46 mM ATP. Concentration of glucagon: 5 pg/ml, fluoride: 10 mM
with increasing concentrations of ATP, a t a fixed Mg++:ATP ratio of 2 : l . Under these conditions, both affinity curves appear to be sigmoidal, but the marked shift in the presence of fluoride indicates a change in the affinity towards ATP.
As was already demonstrated in Fig.4, Mg++ stimulates adenyl cyclase activity a t a molar ratio ATP/Mg++ of 20 which is far in excess of the ratio of the ATP-Mg++ complex under the conditions of the test. Fig.9 illustrates this stimulation a t a fixed concentration of ATP, saturation being reached a t about 4.5 mM (molar ratio of 10) in the case of basal (Fig.9A) activity as well as under stimulation by glucagon or fluoride (Fig.9B). I n contrast to the recent h d i n g with fat cell “ghosts” [4], no increase in affinity for Mg++ could be observed under stimula- tion.
DISCUSSION
Isolation of the labeled Ado-3’ :5’-P formed by the adenyl cyclase is based on the method originally developed by Krishna et al. [3]. In response to criticism of their methodology [S], Krishna and Birnbaumer [20] have recently published a re- evaluation of their assay technique which compares favourably with other systems. As shown in the present study, the method seems well applicable to the measurement of adenyl cyclase activity in liver. I n order to gain some general insight into the mechanism of hormone effects upon this organ, a particulate preparation was used which contains most of the membranous material of the liver. In two respects it seemed advantageous to employ a crude particle suspension : the preparation procedure was simple and allowed for the use of fresh material
in each experiment; extensive manipulation during isolation may have altered hormone sensitivity. The specific adenyl cyclase activity reported herein lies in the general range with that described by Pohl et al. [i2] for liver membranes and that of Bitensky et al. [lo]. Mouse and rat liver preparations were ap- proximately equal in basal and glucagon stimulated activity. There was considerable variation in hormone stimulation from animal to animal; the possible influence of such factors as nutritional and environ- mental conditions have not been investigated.
It is not established whether the methylxanthines which are most widely used in adenyl cyclase assays, will completely block phosphodiesterase activity. Furthermore, an inhibition of Ado-3’ : 5’-P formation could not be excluded in these experiments, the effect being stronger with the less soluble theophylline than with aminophylline. For this reason a “trapping system” with high concentrations of unlabeled Ado-3‘ : 5’-P in the test seemed more advantageous in the absence of an apparent product inhibition.
The observed sensitivity to glucagon compares well with dose response curves shown by Birnbaumer et al. [5], and Bar and Hechter [21] with fat cell “ghosts”. It should be noted however, that intact cellular systems like isolated fat cells [22] or the perfused rat liver [23] are considerably more sensitive to glucagon. For instance, perfusion of glucagon a t peak values equivalent to 1.4 ng/ml led to a maximal activation of the phosphorylase system in liver [23]. It is possible that disruption of the cell membrane has already resulted in some modification of the receptor-enzyme system. Yet this does not invalidate the adenyl cyclase assay as a model for studying the mechanism of hormone action.
Inhibition by high calcium concentrations has been reported for adenyl cyclase systems in other tissues [4,7,24]. This inhibition may be related to the marked potentiation of the glucagon effect by EGTA. A sizable amount of calcium appears to be bound to the membrane particles which seems ac- cessible to the chelating action of EGTA. An indi- cation for the specificity of this effect is its complete reversal by addition of Ca++. I n contrast, the stimu- latory effect of fluoride was slightly inhibited by the chelating agent. It is thus conceivable that Ca++ may play a role in the transmission of the hormone impulse from the (hypothetical) receptor to the catalytic enzyme unit. Again this h d i n g corresponds with similar effects of EGTA on epinephrine and glucagon action in fat cell “ghosts” [20].
Other differences between glucagon and fluoride action were manifested by the sigmoidal dose re- sponse curve of fluoride as compared to the hyper- bolic curve with glucagon as well as the marked shift in substrate affinity observed with fluoride which indicates a possible allosteric interaction (Fig. 8). The extent of inhibition by pyrophosphate marked a

Vo1.17, So.1, 1970 K. D. HEPP, R. EDEL, and 0. WIELAND 177
further distinction between glucagon and fluoride action. Maximal stimulation by the two compounds was not additive, indicating that both substances act eventually on the same catalytic enzyme unit. These phenomena would be consistent with the concept [25] of a regulatory subunit comprising the hormone re- ceptor coupled to a catalytic subunit which is sub- ject to direct stimulation by fluoride.
The many similar characteristics of adenyl cyclase activity in fat cells [4--6,211 adrenal tissue [7] heart 1241 and liver are quite striking and would point to some general properties of mammalian adenyl cyclase systems.
The authors wish to thank Dr. G. Loffler for his advice in the computer program for the calculation of the adenyl cyclase data. This work was supported by the Deutsche Forschungsgemeinschaft.
REFERENCES 1. Sutherland, E. W., and Rall, T. W., J . Biol. Chem. 232
2. Sutherland, E. W., Rall, T. W., and Menon, T., J . Biol.
3. Krishna, G., Weiss, B., and Brodie, B. B., J . Pharmacol.
4. Birnbaumer. L.. Pohl. S. E.. and Rodbell. M.. J . Biol.
(1958) 1077.
Chem. 237 (1962) 1220.
Exp. Ther. 163 (1968) 379.
Chem. 244 (1969) 3468. '
(1969) 3477. 5. Birnbaumer, L., and Rodbell, IT., J . Biol. Chew. 244
6. Rodbell. M.. Birnbaumer, L.. and Pohl, S. L., J . Biol. . . Chem: 245 (1970) 718.
244 (1969) 247. 7. Taunton, 0. D., Roth, J., and Pastan, I., J . Biol. Chenz.
8. Levey, G. S., and Epstein, S. E., J . Clin. Invest. 48 (1969) 1663.
9. Wolff, J., and Jones, A. B., Proc. Nut. Acud. Sci. U.S. A .
10. Bitenskv. M. W.. Russell. V.. and Robertson. W.. Bio- 65 (1970) 454.
chem."hiophys: Res. Commun. 31 (1968) 706.
Biophys. Res. Commun. 36 (1969) 185.
164 (1969) 566.
'
11. Marinetti, G. V., Ray, T. K., and Tomasi, V., Biochem.
12. Pohl, S. L., Birnbaumer, L., and Rodbell, M., Science,
13. Rodbell, M., J . Biol. Chem. 242 (1967) 5744. 13a. Weiss, L., personal communication. 14. Menahan, L. A., Hepp, K. D., and Wieland, O., Bur. J .
15. Bar, H. P., and Hechter, O., Anal. Biochem. 29 (1969) Biochem. 8 (1969) 435.
476.
311 (1958) 245. 16. Lamprecht, W., and Trautschold, I., 2. Physiol. Chenz.
17. Salomon. Y.. and Schramm. M., Biochem. Biovhus. Res. A - , ,
Commun. 38 (1970) 106. 17a. Hepp, K. D., unpublished results. 18. Chabarek, S., and Martell, A. E., Organic Sequestering
19. Rall, T. W., and Sutherland, E. W., J . Biol. Chem. 202
20. Krishna, G., and Birnbaumer, L., Anal. Riochem. 35
21. Bar. H. P.. and Hechter, 0.. Proc. Nut. Acad. Sci. TI . S . A .
Agents, John Wiley, New York 1959, p. 577.
(1958) 1065.
(1970) 393. . ,
63 (1969) 350. 22. Hepp, K. D., Menahan, L. A., Wieland, O., and Willi-
ams, R. H., Biochim. Biophys. Acta, 184 (1969) 554. 23. Mackrell, D. J., and Sokal, J. E., Diabetes, 18 (1969) 724. 24. Drummond, G. I., and Duncan, L., J . Biol. Chem. 245
25. Robison, G. A., Butcher, R. W., and Sutherland, E. W., (1970) 976.
Ann. N . Y. Acad. Sci. 139 (1967) 703.
K. D. Hepp, R. Edel and 0. Wieland Institut fur Diabetesforschung BRD-8000 Miinchen 23, Kolner Platz 1, Germany
12 Eiw. J. Biochem., Vo1.17





























