Embed Size (px)
Citation preview

HippocampusMajor cellular components:
• dentate gyrus (major source of inputs)
• Ammon’s horn: fields CA1, CA2, CA3/CA4 (integrative role)
• subiculum (major source of efferents)
Major pathways and connections:
• hippocampus receives highly processed sensory information about internal and external
events:
– perforant pathway: hippocampus is reciprocally connected to the entorhinal cortex;
entorhinal cortex has connections with other corticial areas (visual, auditory information)
– fimbria-fornix: hippcampus is also reciprocally connected to septum, thalamus and
hypothalamus
• several pathways that allow for intrahippocampal connections:
– commissural connections: neuronal connections made between neurons in two hippocampi
– Schaffer collaterals: connections between neurons made on one side of hippocampus

HippocampusHippocampus plays an important role in two main processes:
• learning and memory
– especially tasks that involve processes of spatial cues
– evidence for sex differences in hippocampal structure
– also evidence for sex differences in the performance of spatial tasks; gonadal steroids have
been implicated in organizational and activational effects on performance
– in the adult, changes in hippocampal structure accompany hormone changes during estrus
• “brake” on HPA axis
– hippocampus possesses mineralocorticoid and glucocorticoid receptors
– mineralocorticoid receptors are linked to circadian changes in HPA axis
– glucocorticoid receptors are linked to terminating a stress response
– chronic exposure to glucocorticoids can damage the hippocampus leading to higher levels
of glucocorticoids, more hippocampal damage, and so on; damage to the hippocampus has
been linked to memory deficits

HippocampusPatient H.M.:
• H.M. suffered from intractable epilepsy (epileptic seizures)
– an epileptic seizure means that a large collection of neurons in the brain discharge in
abnormal synchrony--seizures can be focal that spread throughout cortex or generalized,
and may involve loss of consciousness as well as contraction of groups of skeletal muscle
– intractable means that his epileptic seizures were resistant to treatment
• to stop his epileptic seizures, heunderwent bilateral hippocampectomy--bilateral
removal of his hippocampi
• following surgery:
– GOOD NEWS: his epilepsy stopped
– BAD NEWS: while he could remember events early in his life, he could not remember
events just prior to surgery (mild form of “retrograde amnesia”), and he was unable to form
new memories (“anterograde amnesia:)

HippocampusPatient H.M.:
• these events in Patient H.M. highlight the important role that the hippocampus plays
in the processes of learning and memory
– mild form of retrograde amnesia and anterograde amnesia indicates that the hippocampus
plays an important role in learning and in the formation of short-term memory (working
memory)
– however, the ability of H.M. to remember early events in his life indicates that the
hippocampus is not the location where long-term memories are stored
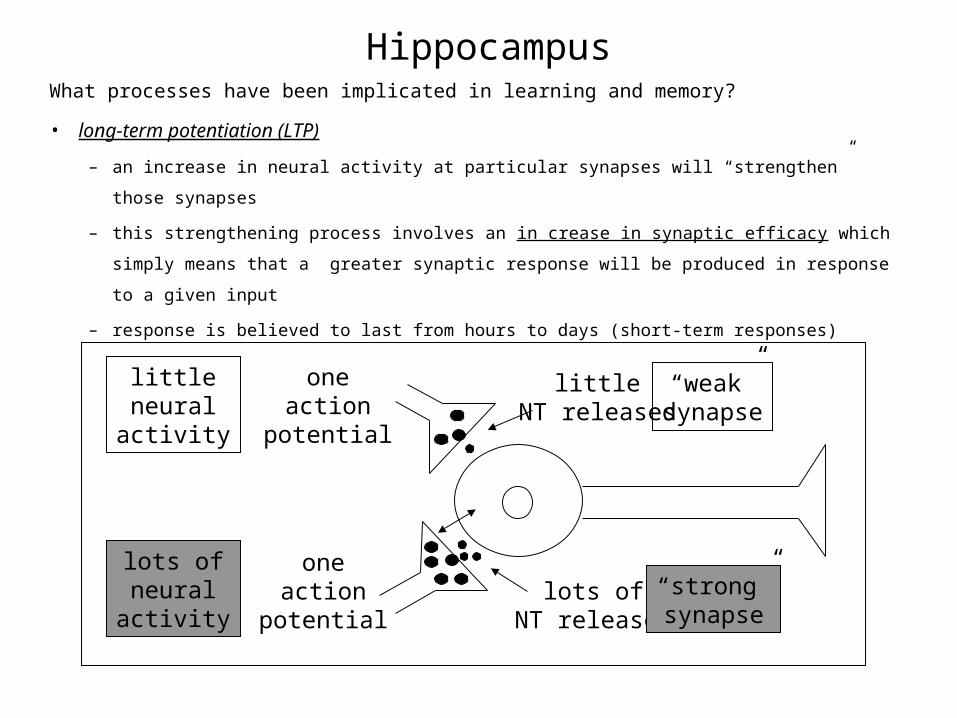
HippocampusWhat processes have been implicated in learning and memory?
• long-term potentiation (LTP)
– an increase in neural activity at particular synapses will “strengthen” those synapses
– this strengthening process involves an in crease in synaptic efficacy which simply means
that a greater synaptic response will be produced in response to a given input
– response is believed to last from hours to days (short-term responses)
oneaction
potential
littleNT released
“weak”synapse
oneaction
potentiallots of
NT released
“strong”synapse
littleneural
activity
lots ofneural
activity

Hippocampus• changes in neuronal morphology
– an increase in neural activity will lead to an increase in neuronal connections
– this strengthening process involves increasing synaptic input and the dendritic complexity
of neurons: 1) more synapses, 2) more dendritic spines, 3) increase in length and branching
of dendrites
– response can last from days to weeks to years (short and long-term responses)
littleneural
activity
lots ofneural
activity

HippocampusLearning and Memory:
• cellular mechanisms are varied but can involve:
– enhanced neurotransmission within synapse--increase in synaptic efficacy; this is thought
to reflect an increase in release of neurotransmitter
– formation of new connections
• these cellular mechanisms of learning and memory have been observed within the
hippocampus
– these changes are thought to reflect learning and formation of short-term memories
especially associated with tasks involving spatial cues
• similar plastic events have also been observed within other brain areas including the
cerebral cortex (e.g., visual cortex) and cerebellum

HippocampusDo gonadal steroids affect neuronal morphology in the hippocampus?
• Answer--yes!
– There is evidence that elevations in estrogen and progesterone can regulate the number of
dendritic spines on neurons in the hippocampus in adult female rats.
Study by Gould et al. (1990):
• Question: Does estrogen and progesterone regulate spine density in neurons within
the hippocampus?
– spine density = number of spines per length of dendrite = number of synapses on spines
dendriticshaft
dendriticspine
Synapses:axosomatic
axodendritic
shaft
spine
axoaxonal
dendrite

Hippocampus• Methods:
– adult female rats: intact, OVX + oil, OVX + estrogen, OVX + estrogen + progesterone
– euthanized animals and stained brain tissue with Golgi technique--silver stain that “fills”
the dendrites, cell bodies and axons of specific neurons
– measured the number of spines per length of apical or basilar dendrites of neurons in CA1,
CA3, and dentate gyrus in female rats
• Results:
– ovariectomy produced a significant decrease in spine density in apical dendrites of neurons
within CA1 region of hippocampus
– administration of estrogen or estrogen plus progesterone produced a significant increase in
spine density in the apical dendrites of CA1 region of hippocampus
– the effect was specific to CA1 region of hippocampus; no change occurred in CA3 region
or in dentate gyrus
– the effect was rapid occurring after only two days of estrogen and 5 hours of progesterone




Hippocampus• Conclusion:
– the levels of estrogen and progesterone can affect the number of spines present within a
select group of neurons within the hippocampus (CA1 region) of adult female rats
• Subsequent studies have shown that such changes in spine density also occur with the
natural fluctuation in hormones that takes place during the estrus cycle.
– high estrogen and progesterone levels = high spine density (late proestrus/early estrus)
– low estrogen and progesterone levels = low spine density
– these changes are occurring in adult female rats every four or five days

HippocampusAre there sex differences in the structure of the hippocampus?
• Answer--yes! There is evidence for a complex interaction between early experience
(rearing), dendritic morphology and sex of individual (rats).
– animals raised in an enriched environment possess neurons that are more complex than
animals raised under normal laboratory conditions; an enriched environment involves the
presence of other animals and various objects to interact with, while normal laboratory
conditions are more plain and animals may be housed alone or in small groups with no
objects to play with
– if you compare males and females housed in the complex environment to rats housed under
normal laboratory conditions, you can see several differences:
– in the apical dendritic tree of CA3 neurons, females housed in the enriched environment
have more dendrites concentrated proximal (close) to the cell body, while males in the
enriched environment had more dendrites concentrated distal (far) from cell body
– in the dentate gyrus, females housed in enriched environments had granule cells with an
increase in dendritic length while males in a similar environment did not show this change


HippocampusAre there sex differences in learning and memory?
• Answer--yes!
• There are numerous examples of differences between males and females in
performance on various tests of learning and memory.
– Males are “better” at passive avoidance learning than females (e.g., males learn more
quickly to not leave a platform because they will get shocked).
– Females are “better” at active avoidance learning than males (e.g., females learn to respond
more quickly to a cue such as a light or tone that signals that they should move to another
part of a chamber to avoid being shocked).
– However, Beatty has argued that such differences may simply reflect sex differences in
activity. That is, females are more active than males and as a consequence they may do
better on active avoidance tasks because of an increased likelihood of making the
association between movement to a given part of a chamber , cue presentation and a
decrease in shock. Females may do more poorly on passive avoidance tasks because of
they can’t sit still.

HippocampusAre there sex differences in learning and memory?
• It is thought that performance on other more complex tasks, such as radial arm maze or
the Morris water maze, may be less influenced by sex differences in activity.
– Maze tasks are considered tests of spatial abilities in rodents because animals solve these
maze tasks by using cues from the surroundings outside of the maze.
– The hippocampus (in rats) is thought to be essential for solving tasks that require the animal
to use its spatial abilities.
• There is evidence that males tend to perform better on spatial tasks than females.
– This sex difference in seen in some species but not all.
– This difference is also somewhat limited--greatest sex differences are observed during
acquisition of the task, and often fewer differences are seen once the task has been learned.
– It has been suggested that males and females used different cues to solve spatial tasks (which
may underlie differences in acquisition), and there is evidence to suggest that exposure to
gonadal steroids during development and in the adult can alter what cues are used to solve a
given task.

HippocampusStudy by Williams et al. (1990)
• Question: Does exposure to androgens or estrogens early in life affect spatial
abilities in adulthood?
• Methods:
– 4 groups: male rats castrated on day 1 (MNC), sham-operated control males (MC), female
rats exposed to estrogen from days 1-10 (FNE), and sham-operated control females treated
with oil (FC)
– at 45 days of age, all groups were gonadectomized (MNC group was already castrated); this
was done to control for any activational effects of on performance
– at 70 days of age, all rats were placed on a food deprivation schedule that kept tham at 85%
of their free-feeding body weight; rats were trained to run down arms of the maze for food
– tested the performance of the rats on locating food pellets when only some of the arms were
baited--12-arm maze, 8 arms were baited with food and 4 arms were not; this relationship
remained constant throughout the experiment

HippocampusStudy by Williams et al. (1990)
• Methods:
– they determined how well animals performed on this task by analyzing number of errors
made until all food pellets were obtained
– 2 types of errors: 1) remembering not to go into unbaited arms, and 2) remembering what
arms were visited on a given day (1 test per day for 18 days)
• Results:
– males and masculinized females showed faster acquisition of maze task than did females or
feminized males
– however, after acquisition, no sex differences in performance were observed


HippocampusStudy by Williams et al. (1990)
• Question: Why are males and females different in acquisition of the radial arm
maze? Do these differences reflect the cues that males and females use to solve the
task?
• Methods:
– similar groups as before: 4 groups: male rats castrated on day 1 (MNC), sham-operated
control males (MC), female rats exposed to estrogen from days 1-10 (FNE), and sham-
operated control females treated with oil (FC); all groups were gonadectomized
– trained the animals on the radial arm maze until high performance levels were obtained
– they changed either landmark cues, geometry or both and tested the performance of the
animals on task
– landmark cues: cues located within or around a maze (table, chair, transport cart); they
manipulated these cues by rearranging items or removing them
– geometric cues: shape of room (corners of room); manipulated geometry by enclosing the
maze within a black circular arena

HippocampusStudy by Williams et al. (1990)
• Results:
– males and androgenized females used primarily geometry to solve the task
– females and feminized males used both geometry and landmarks in performing task
• Conclusions:
– males use fewer cues (geometry) to solve the radial maze than females (geometry and
landmark cues)
– the need to learn fewer cues may explain why males acquire the task more quickly than
females
– enhanced spatial ability in males is promoted by perinatal exposure to gonadal steroids--1)
castration of newborn males decreased rate of acquisition, and 2) administration of estrogen
to newborn females within first 10 days of life increased rate of acquisition
– after acquisition, no sex differences in performance were observed


Hippocampus• Sex differences in maze performance have been associated with sex differences in
brain structure--hippocampus.
• Study by Jacobs et al. (1990) compared the size of the hippocampus in two species of
prarire voles that show sex differences in performance on spatial tasks.
meadowvole
pinevole
Spatial Task Hippocampus
males performbetter than
females
no sexdifference
in performanceon task
11% largerin males than
in females
no sexdifferencein size of
hippocampus

HippocampusPerformance on spatial tasks can also be affected by hormones in the adult.
• In males:
– maximal performance on some spatial tasks are seen in males only after puberty (rise in
testosterone levels)
– increase in spine density in CA1 neurons of male mice observed after puberty
• In females:
– performance on spatial tasks can be altered during estrus cycle

HippocampusStudy by Warren & Juraska (1997)
• Question: Does performance on spatial tasks by females vary with their estrus
cycle?
• Methods:
– several groups of animals were studied, but they focused primarily on females during 2
stages of the estrus cycle in 2 forms of the Morris water maze
– 2 main groups of females: females in late proestrus (elevated estrogen levels) versus
females in late estrus (estrogen levels are low)
– Morris water maze requires that an animal learn to find a platform submerged under water
(water is murky--not a visual task)
– place form of maze--females had to use spatial cues surrounding the maze to find platform
(spatial task)
– cued form of maze--a black ball was suspended above the platform, so females had to learn
to find platform by going toward black ball (cued task)
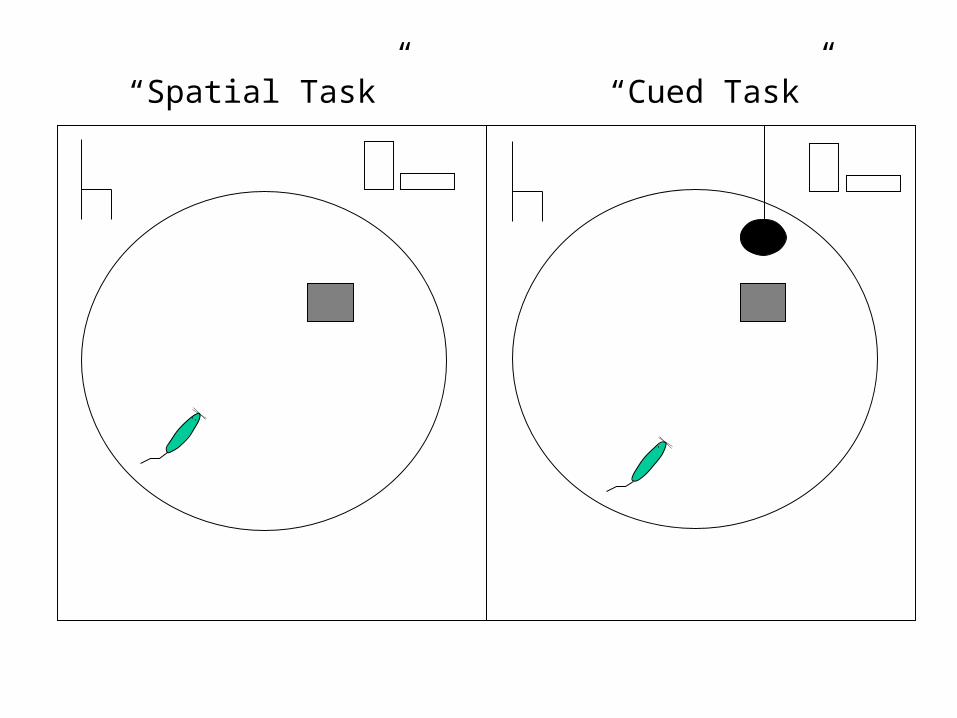
“Cued Task”“Spatial Task”

HippocampusStudy by Warren & Juraska (1997)
• Methods:
– animals received pretraining trials on either the spatial task or the cued task
– on the day of the experiment, females in late proestrus or late estrus received 8 trials, 1 hour
break, followed by 8 additional trials
– how quickly did they find the platform?
• Results:
– on spatial task, late estrus females found platform more quickly than late proestrus females
– on the cued task, late proestrus females found platform more quickly than late estrus
• Conclusions:
– the increase in estrogen during late proestrus (when there is an increase in dendritic spines)
is associated with decreased performance on the spatial task, but increased performance on
the cued task

HippocampusOverall Summary:
• sex differences can be seen in tasks involving spatial learning and memory
• in males:
– increased performance may be associated with the use of fewer cues to learn the task
(geometry)
– gonadal steroids have an organizing effect on spatial ability
– rise in testosterone at puberty may also act to enhance spatial abilities (at least on some
tasks)--activating effect
• in females:
– decreased performance may be associated with learning more and possibly different cues
associated with a spatial task
– in adults, increased estrogen (and associated changes in spine density in the hippocampus)
appears to inhibit performance on tasks requiring use of spatial cues but may enhance
responsiveness to other cues

NEXTSECTION

Hippocampus-Chronic Stress-Memory DeficitsThe pituitary has 2 modes of operation:
• diurnal variations in secretion
– daily rhythm of glucocorticoid secretion
– glucocorticoid levels rise when an individual becomes “active” during the day
– in humans, glucocorticoids are elevated in the morning and are decreased at night
– in rats, glucocorticoids are elevated in the evening (or when lights go out); rats are
nocturnal animals being most active at night
• stress-induced secretion
– glucocorticoid levels rise, in part, to increase blood glucose levels available for fight or
flight reactions

Hippocampus-Chronic Stress-Memory DeficitsThere are two different receptors populations that respond to glucocorticoids:
• mineralocorticoid receptor (MR)
– receptor binds with high-affinity to glucocorticoids (receptor responds to small changes in
glucocorticoid levels)
– MRs are thought to modulate diurnal variations in glucocorticoid secretion
– MRs are found in select brain regions--hippocampus, septum and amygdala
• glucocorticoid receptor (GR)
– receptor that binds with low-affinity to glucocorticoids (receptor responds to large changes
in glucocorticoid levels)
– GRs are thought to modulate stress-induced secretion of glucocorticoids
– GRs are found throughout the brain and body
• the increase in glucocorticoids, either during daily rhythm or during stress, would
activate the appropriate receptor, stimulating neurons within the hippocampus that
would act to inhibit activity of the HPA axis
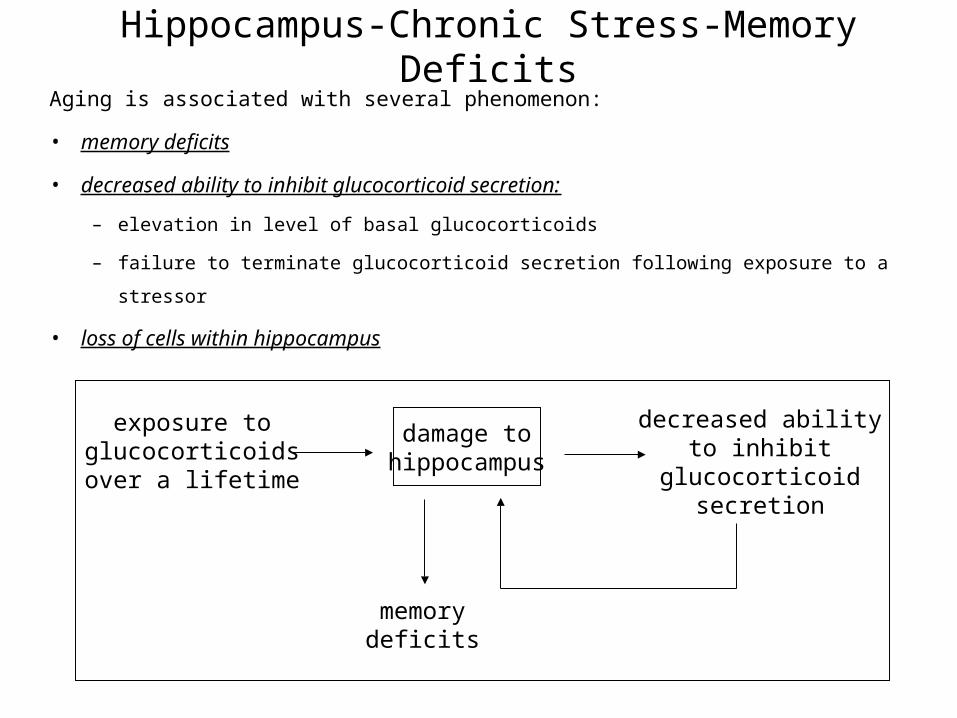
Hippocampus-Chronic Stress-Memory DeficitsAging is associated with several phenomenon:
• memory deficits
• decreased ability to inhibit glucocorticoid secretion:
– elevation in level of basal glucocorticoids
– failure to terminate glucocorticoid secretion following exposure to a stressor
• loss of cells within hippocampus
exposure toglucocorticoidsover a lifetime
damage tohippocampus
decreased abilityto inhibit
glucocorticoidsecretion
memorydeficits


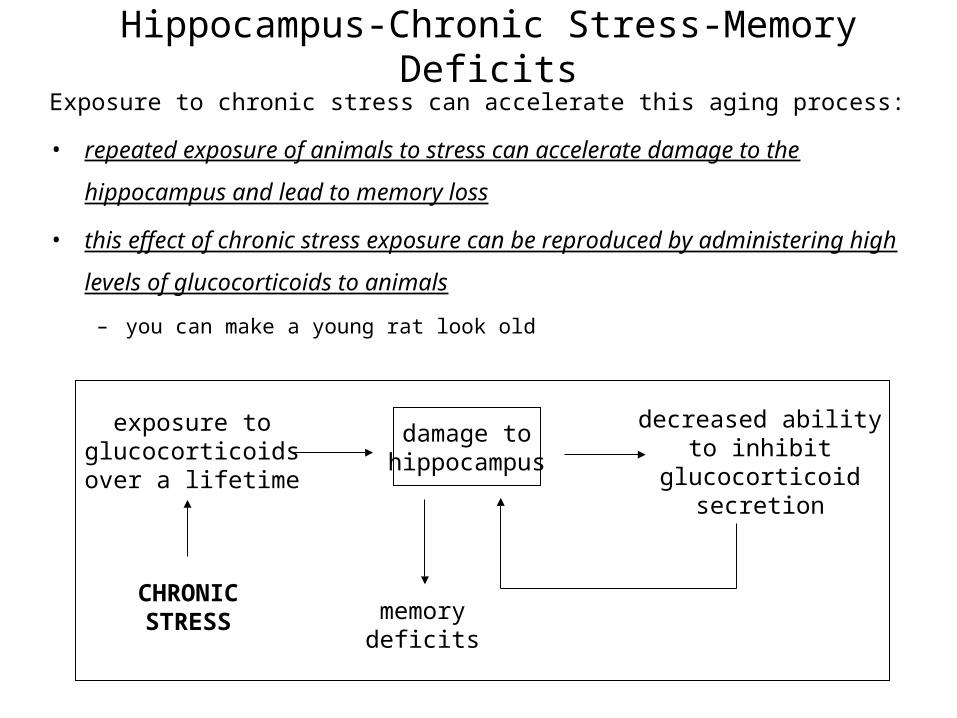
Hippocampus-Chronic Stress-Memory DeficitsExposure to chronic stress can accelerate this aging process:
• repeated exposure of animals to stress can accelerate damage to the hippocampus
and lead to memory loss
• this effect of chronic stress exposure can be reproduced by administering high levels
of glucocorticoids to animals
– you can make a young rat look old
exposure toglucocorticoidsover a lifetime
damage tohippocampus
decreased abilityto inhibit
glucocorticoidsecretion
memorydeficits
CHRONICSTRESS

Hippocampus-Chronic Stress-Memory DeficitsStudy by Arbel et al. (1994)
• Purpose:
– to determine the effects of long-term corticosterone administration on hippocampal morphology
and cognitive performance of middle-aged rats
• Method:
– prescreened rats by testing them on their ability to learn the Morris water maze--learning the
location of a platform submerged under water
– young rats (3 months of age) learn quickly; middle-aged rats (12 months of age) vary--some
learn as quickly as the young rats (‘non-impaired’ rat) and others that show a decrease in
performance (‘impaired” rats)
– a subset of the prescreened rats were killed--the rat’s performance on the Morris water maze was
compared with number of damaged cells within the hippocampus
– a different subset of prescreened rats were tested on the radial arm maze--8-arm maze in which
all of the arms were baited with food; the goal of the task was to obtain all of the food with the
least number of errors; in this task, some of the ‘non-impaired’ rats were given corticosterone

Hippocampus-Chronic Stress-Memory DeficitsStudy by Arbel et al. (1994)
• Results:
– the response of middle-aged rats varied on the Morris water maze--some were ‘non-
impaired’ and showed escape latencies that were as quick as those made by young
rats(escape latency = time to find the platform); other middle-aged rats were ‘impaired’ on
the task taking longer to find the platform
– performance on the Morris water maze task reflected differences in the morphology of
neurons within the hippocampus: 1) young rats and middle-aged ‘non-impaired’ rats had
few cells that were damaged in the hippocampus and showed short latencies(10-50 sec) to
find the platform, 2) middle-aged ‘impaired’ rats had more cells that were damaged in the
hippocampus and they showed longer latencies (81-120 sec) to find the platform
– damaged cells were dark-stained cells having pyknotic nuclei
– administration of corticosterone (9-week period) reduced performance of the ‘non-impaired’
middle-aged rats to that of impaired’ middle-aged rats




Hippocampus-Chronic Stress-Memory DeficitsStudy by Arbel et al. (1994)
• Conclusions:
– decreased performance on the Morris water maze task by ‘impaired’ middle-aged rats was
associated with a greater number of pyknotic of damaged cells in the hippocampus
– administration of corticosterone to ‘non-impaired’ middle-aged rats decreased their
performance on the radial-arm maze

Hippocampus-Chronic Stress-Memory DeficitsHow do glucocorticoids damage the hippocampus?
• complex interaction between transport of glucose into cells, elevated levels of
glutamate at synapse, and elevated levels of calcium within cells
– earlier we noted that glucocorticoids act at intracellular receptors to increase the synthesis
of enzymes that act to increase the process of gluconeogenesis
– evidence that glucocorticoids can also act to increase blood glucose levels by blocking
transport of glucose into some cells of body--a decrease in glucose uptake by cells in the
body allows for more glucose to reach brain, heart and skeletal muscle, however, glucose
uptake can even be reduced in the brain (there is evidence for 25% decreased glucose-
uptake in neurons and glia within the hippocampus)
– decrease in glucose-uptake by neurons and glia is thought to reduce the ability of these
cells to function normally
– of interest, administration of energy substrates, such as glucose, can reduce the damaging
effects of glucocorticoids on hippocampal structure and function

Hippocampus-Chronic Stress-Memory DeficitsHow do glucocorticoids damage the hippocampus?
– in glial cells, the decrease in glucose-uptake following glucocorticoid administration can
inhibit their ability to buffer synapses from excess levels of glutamate
– glutamate is an excitatory neurotransmitter that acts at specific receptors; at the NMDA
receptor, glutamate acts to increase the flow of Ca2+ into cells
– too much glutamate at synapse can lead to too much influx of Ca2+ within neurons
– an inability to regulate intracellular Ca2+ levels can damage cells and ultimately lead to
cell death

NEXTSECTION

NEXTSECTION





























