Embed Size (px)
Citation preview

Molecular Membrane Biology, 2010; Early Online, 1–12
High plasma membrane lipid order imaged at the immunologicalsynapse periphery in live T cells
DYLAN M. OWEN1, STEPHANE ODDOS1, SUNIL KUMAR1, DANIEL M. DAVIS2,MARK A. A. NEIL3, PAUL M. W. FRENCH3, MICHAEL L. DUSTIN5,ANTHONY I. MAGEE4 & MAREK CEBECAUER4
1Chemical Biology Centre, 2Division of Cell and Molecular Biology, 3Department of Physics, 4National Heart and LungInstitute, Imperial College, London, UK, and 5Skirball Institute of Biomolecular Medicine, New York University School ofMedicine, New York, USA
(Received 18 March 2010; and in revised form 12 May 2010)
AbstractCholesterol- and glycosphingolipid-enriched membrane lipid microdomains, frequently called lipid rafts, are thought to playan important role in the spatial and temporal organization of immunological synapses. Higher ordering of lipid acyl chains wassuggested for these entities and imaging of membrane order in living cells during activation can therefore help to understandthe mechanisms responsible for the supramolecular organization of molecules involved in the activation of T cells. Here, weemploy the phase-sensitive membrane dye di-4-ANEPPDHQ together with a variety of spectrally-resolved microscopytechniques, including 2-channel ratiometric TIRF microscopy and fluorescence lifetime imaging, to characterize membraneorder at the T cell immunological synapse at high spatial and temporal resolution in live cells at physiological temperature. Wefind that higher membrane order resides at the immunological synapse periphery where proximal signalling through theimmunoreceptors and accessory proteins in microclusters has previously been shown to take place. The observed spatialpatterning of membrane order in the immunological synapse depends on active receptor signalling.
Keywords: Lipid rafts, membrane lipid order, the immunological synapse, T cells
Introduction
The immunological synapse (IS) is the junctionformed between immune cells (e.g. T, B and NKcells) and their targets (e.g., antigen-presenting cells,APCs) [Monks et al. 1998, Grakoui et al. 1999,Bromley et al. 2001]. It is formed during the processof immune cell activation and also during inhibition[Davis et al. 1999]. Previously, it has been shownthat, within this junction, signalling and adhesionproteins connected to cell activation are patternedinto a system of concentric Supra-Molecular Activa-tion Clusters (SMACs) and mobile protein micro-clusters [Monks et al. 1998, Grakoui et al. 1999,Yokosuka et al. 2005, Varma et al. 2006]. It wasfurther demonstrated that the peripheral and distalregions of the IS are important areas for adhesionand proximal signal transduction events such asprotein tyrosine phosphorylation [Lee et al. 2003,
Varma et al. 2006]. For the central area of the IS,the cSMAC, regulatory and signal enhancing roleshave been proposed [Lee et al. 2003].The appearance of certain proteins and lipids in
specific areas of the membrane led to the hypothesisthat cholesterol- and sphingolipid-enriched, highlyordered membrane lipid microdomains (also knownas lipid rafts, LRs) may play an important role in theorganization of molecules at the cell plasma mem-brane [Simons and Ikonen 1997]. Some proteinshave been shown to accumulate in, and others to beexcluded from, these entities using various ‘raft mar-kers’ [Day and Kenworthy 2009]. Labelling raftsoften includes multivalent cholera toxin subunit Bor antibodies, raising the possibility of aggregation ofsmaller structures. New non-intrusive sensing meth-ods for membrane lipids’ physico-chemical pro-perties are required for direct observation of thesedomains in living cells at physiological temperatures
Correspondence: Dr Marek Cebecauer, National Heart and Lung Institute, Imperial College, London, UK. Tel: +44 79087 58786. Fax: +44 20759 43100.E-mail: [email protected] Dylan M. Owen is currently at the Centre for Vascular Research, University of New South Wales, Sydney, Australia.
ISSN 0968-7688 print/ISSN 1464-5203 online � 2010 Informa UK LtdDOI: 10.3109/09687688.2010.495353
Mol
Mem
br B
iol D
ownl
oade
d fr
om in
form
ahea
lthca
re.c
om b
y N
yu M
edic
al C
ente
r on
06/
29/1
0Fo
r pe
rson
al u
se o
nly.

[Cebecauer et al. 2009]. Indirect measurements byFörster resonance energy transfer [Sharma et al.2004], fluorescence correlation spectroscopy (FCS)[Lenne et al. 2006], and single particle tracking[Suzuki et al. 2007], demonstrated transient confine-ment of putative raft components in nanodomains inthe plasma membrane of resting cells. In recent yearsthe LR hypothesis has evolved into one with a moretransient, dynamic picture; ‘LRs are now thought to besmall (10–200 nm), heterogeneous, dynamic domainsthat compartmentalize cellular processes and can bestabilized to form larger platforms’ [Pike 2006]. Datacombining super-resolution imaging with FCS con-firm this view [Eggeling et al. 2009]. Experimentswith artificial membranes have demonstrated thatdomainsmimickingLRs inbiologicalmembranes havehigher lipid ordering (liquid-ordered phase) than thesurrounding ‘non-raft’ area (liquid-disordered phase)[Sengupta et al. 2007]. Two strategies have beendeveloped for imaging lipid ordering in cellular mem-branes using fluorescence: phase-partitioning dyes[Burack et al. 2002] and environmentally-sensitivefluorescent probes which can report on their localmolecular surroundings [Parasassi et al. 1990,Gaus et al. 2003]. The size and short average lifespanof membrane lipid microdomains poses a significantchallenge to optical microscopy techniques which arelimited in their spatial resolution and minimum imageacquisition times. The use of a single environmentally-sensitive probe that can change its fluorescent proper-ties depending on whether it is residing in ordered ordisordered membranes is a potentially powerfulapproach for improving the spatio-temporal resolutionof membrane studies. A prime example of this type ofimaging is the use of the phase-sensitive, membrane-partitioning dye LAURDAN which shows a 50 nmshortening of its peak fluorescence emission wave-length between the liquid-disordered and liquid-ordered phases [Gaus et al. 2006].Membrane lipid microdomains have been impli-
cated as having an important role in the organizationof signalling events at the IS of T cells [Janes et al.1999, Tavano et al. 2004]. Gaus and colleagues usedLAURDAN generalized polarization (GP) values cal-culated from two spectral channels to image thedistribution of membrane order in T cells. Theyfound that higher order membrane regions accumu-late in the contact site between T cells and APCsforming a relatively diffuse peripheral ring in the IS[Gaus et al. 2005]. Subsequently, they used thecholesterol analogue, 7-ketocholesterol (7KC), todisrupt membrane order and showed an impact onT cell activation [Rentero et al. 2008]. LAURDANimages in these studies were acquired using fixedcells at 25�C with potentially unphysiological effects
on the highly dynamic membranes of living cells[Magee et al. 2005]. In addition, two-photon excita-tion is required for LAURDAN imaging whichresults in a decrease in spatial resolution and, owingto the low two-photon excitation cross-section,longer acquisition times are required to achieve agiven signal-to-noise level compared to single-photon-excited probes.Recently, a new probe sensitive to membrane
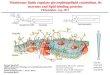
order, di-4-ANEPPDHQ, has been developed[Obaid et al. 2004, Jin et al. 2006] which can be exci-ted in the blue spectral region with single-photonexcitation providing increased spatial resolution anddecreased acquisition times. More importantly, itallows the use of widefield illumination and detec-tion thus opening the door to high-speed imagingand total internal reflection fluorescence (TIRF)microscopy. Di-4-ANEPPDHQ exhibits a 60 nmspectral blue-shift between the disordered andordered lipid phases (Figure 1a) [Jin et al., 2006],is readily soluble in lipids and aqueous solutionsand shows minimal fluorescence when not lipid-associated, presumably due to a high non-radiativefluorescence decay rate in aqueous environment. Thecombination of various imaging techniques togetherwith the advantages of di-4-ANEPPDHQ allowed usto study the distribution of membrane lipid order inliving cellular systems at physiological temperatureand high spatial and temporal resolution, including inlive primary T cells. All presented results were achie-ved at physiological temperature, and therefore movethe discussion of the role of membrane lipid organi-zation in the activation processes of T cells closer tothe natural state.
Materials and methods
2-channel ratiometric TIRF microscopy
TIRF microscopy was performed on a custom-built microscope with excitation at 473 nm from adiode-pumped solid-state laser (Blues 50, Cobolt,Solna, Sweden) delivered via a single mode opticalfiber (SMF) and a rotatable mirror to adjust the TIRFangle. Excitation is delivered into the back-port of aninverted epifluorescence microscope (IX71, Olym-pus, Tokyo, Japan) equipped with an X60, 1.45NAoil-immersion TIRF objective. Fluorescence was col-lected on an electron-multiplying CCD camera(EMCCD) (iXon, Andor, Belfast, UK) in the range500–593 nm and 600–680 nm using a 2-channelimager (Dual-View, Optical Insights) in order todistinguish high and low order of the lipid environ-ment (see Figure 1a). Fluorescence above 680 nmwas
2 D.M. Owen et al.
Mol
Mem
br B
iol D
ownl
oade
d fr
om in
form
ahea
lthca
re.c
om b
y N
yu M
edic
al C
ente
r on
06/
29/1
0Fo
r pe
rson
al u
se o
nly.

not collected due to high background fluorescence(data not shown). Data were processed using customsoftware (LabVIEW, National Instruments, Austin,TX, USA). Calculated GP values (Equation 1) foreach pixel were then pseudo-coloured (from highmembrane order – red, to low membrane order –
blue; see colour bars for Figures 2–4). The imageswere merged with the fluorescence intensity imagesuch that the result displays both order and structuralinformation. The sample was maintained at 37�Cusing a stage and objective heater (PeCon, Erbach,Germany). Quantitation of image data was performedby measuring average GP values in the central region
(defined from the centre to half way to the synapseedge) and the remaining peripheral region.
GP =I II I
500 600
500 600
− −
− −
−+
593 680
593 680
1( )
Confocal microscopy
Single-photon excitation confocal microscopy wasundertaken using an inverted, epi-fluorescence laser-scanning confocal microscope (TCS SP5, LeicaMicrosystems Ltd, Wetzlar, Germany) with a 63�,1.25NA oil-immersion objective lens. Excitation wasat 488 nm from an argon-ion laser. Fluorescencedetection was in the wavelength bands 500–580 nmand 620–750 nm (Figure 1a) using internal photo-multiplier tubes. z-slices were acquired every 700 nm.GP values were calculated as for TIRF images. 3Dreconstructions and en face views were generated usingVolocity software (Improvision, Waltham, MA, USA).
Fluorescence lifetime imaging
For FLIM, fluorescence was collected in the range540–750 nm using a PMT and TCSPC electronics(SPC-830, Becker and Hickl GmbH, Berlin, Ger-many) with single-photon excitation at 460 nmfrom a frequency-doubled Ti:Sapphire laser (Tsu-nami, Spectra-Physics, Mountain View, CA, USA).Fluorescence decays were fitted to a single-expo-nential function and pseudo-coloured as for the GPimages (experimental details available as Supple-mentary Material online).
Optical tweezing
Cell and IS orientation by optical tweezing was per-formed using an infrared (980 nm) optical trappingbeam as previously described [Oddos et al. 2008].Confocal fluorescence imaging was then performed inthe same way as for the 3D confocal microscopy.
Sample preparation
Jurkat T cells (clone E6.1) and Raji B cells werecultured in RPMI1640 medium (PAA, Pasching,Austria) supplemented with 10% foetal calf serum(FCS) at 37�C in a 5% CO2 atmosphere. Raji B cellswere loaded (or not) with the superantigen Staphylo-coccal enterotoxin E (SEE) for 90 min at 37�C. HumanCD4+ T cells were isolated using a MACS kit fornegative selection (Miltenyi Ltd) after separation of
500 550
a
b
600
Inte
nsity
(a.
u.)
Wavelength (nm)
DOPC
DPPC7ChoI3
EMCCD
SampleSMF
Rotatablemirror
Collectinglens
ND
Laser
1-45NAObj
650 700 750
Figure 1. (a) The fluorescence emission spectra of di-4-ANEPPDHQ in ordered (green) and disordered (red) modelmembranes. The unilamellar vesicles were formed from dioleoylphosphatidylcholine (DOPC) (disordered phase) or a mixture ofdipalmitoyl phosphatidylcholine (DPPC) and cholesterol (Chol) inthe ratio 7:3 (ordered phase). The 60 nm blue-shift in fluorescenceemission between the disordered and ordered phases was detectedusing spectrofluorimetry. The shaded regions show the spectralwindows that were subsequently acquired for construction of theGP value images. (b) Schematic of the 2-channel ratiometric TIRFmicroscope. This Figure is reproduced in colour in the onlineversion of Molecular Membrane Biology.
Membrane order at the immunological synapse 3
Mol
Mem
br B
iol D
ownl
oade
d fr
om in
form
ahea
lthca
re.c
om b
y N
yu M
edic
al C
ente
r on
06/
29/1
0Fo
r pe
rson
al u
se o
nly.

peripheral blood mononuclear cells on a Ficoll gra-dient (GE Healthcare). One hour prior to imaging,the cells were resuspended in HEPES-buffered saltsaline (HBSS) supplemented with 1 mMCa2+, 2 mMMg2+, 1% gelatin and 5 mM di-4-ANEPPDHQ (Invi-trogen, Carlsbad, CA, USA). The use of serum-freemedium containing gelatin dramatically reduces thefluorescence background from residual dye whichwould otherwise swamp the red wavelength channel(Figure S1, Supplementary Material online).Coverslip chambers (Lab-Tek, Brendale, Australia)
coated with poly-L-lysine followed by 10 mg/mlantibodies to CD3 (OKT3 mAb) in phosphate-buffered saline (PBS) and blocked by incubation at37�C with PBS supplemented with 1% gelatin for30 min were used for activatory surface stimulation ofT cells.Supported planar bilayers with GPI-anchored
forms of I-Ek and ICAM-1 were prepared in FCS2flow cells (Bioptechs, Butler, PA, USA) and used forstimulation of AND T cells as described before[Grakoui et al. 1999]. The I-Ek molecules on thebilayers were loaded with 100 mM MCC88-103-peptide in citrate buffer for 48 h. The density ofthe GPI-anchored proteins in the bilayers was: I-Ek
20 sites/mm2 and ICAM-1 150 sites/mm2 (see Supple-mentary Material online for details).For ordered phase disruption using 7KC, 15 mg/ml
7KC and cholesterol (both from Avanti Lipids,Alabaster, AL, USA) in ethanol were combined inthe ratio 2:1. Over 30 min, these were then added to asolution of 50 mg/ml methyl-b-cyclodextrin (Sigma-Aldrich, St Louis, MO, USA) in PBS at 80�C to afinal sterol concentration of 1.5 mg/ml. 15 ml of thelipid solution were then added to 1 ml of cell mediumcontaining 5 � 105 cells at 37�C for 30 min.
Results
Membrane lipid order: Live T cell – APC conjugates
IS junctions between T cells and APCs create afavourable molecular organization for processes invol-ved in the integration and regulation of activatorysignals and for effector functions. To test whetherthe phase-sensitive dye, di-4-ANEPPDHQ, will allowus to image membrane order within this structure inreal time we first incubated living T cells with APCsunder physiological temperature of 37�C and app-lied conventional 2-colour confocal microscopy. Weobserved the accumulation of higher lipid order inthe IS periphery and relatively lower order in thecentral region (Figure 2c and f). Very low orderwas observed in intracellular membrane structures,
as has been reported previously [Owen et al. 2006](Figure 2b and e). These distributions agree wellwith previously published data acquired usingLAURDAN and multi-photon microscopy on fixedcells at room temperature [Gaus et al. 2005]. Largemacromolecular rearrangements in the IS and for-mation of a peripheral adhesive ring and a centralreceptor-rich area in activated T cells were shown tobe antigen signalling-dependent [Monks et al. 1998].We imaged the distribution of membrane order inT cells forming conjugates with APC in the absenceof antigenic stimulus. Again, increased ordering oflipids was observed at the contact site between T cellsand APC but to a much lower level than in thepresence of the antigenic stimulus. Little or no segre-gation into highly ordered peripheral ring and centralarea with relatively lower lipid ordering was observed(Figure 2g–l).Live cell imaging of T cell-APC conjugates is
associated with a low probability that the IS willform in the imaging plane of the microscope thusrequiring 3D z-stacks to be acquired. This limitsspatial resolution as confocal microscopes typicallyhave axial resolutions of around 700 nm and the 3Drendering required to generate en face images of theIS further degrades the image quality. In order toovercome this limitation, we applied optical tweezersto reorientate the IS into the confocal imaging plane(Figure 2m) [Oddos et al. 2008]. This permits the ISto be directly imaged at the higher lateral resolutionof approximately 200 nm. Again, a broad ring of highmembrane order at the IS periphery was observedwith decreased order towards the centre (Figure 2n).The membrane order distribution, as shown in theGP value plot (Figure 2o), was statistically significant(p < 0.05) as indicated by the quantitation of thecentral and peripheral regions (Figure 2p).
Membrane lipid order: Live T cells activated onantibody-coated coverslips
Because conventional confocal fluorescence micros-copy enables imaging of fluorescent objects in thefocal plane with an axial resolution (sectioningstrength) of around 700 nm, the signal detectedmay include contributions from other intracellularstructures within the focal volume as well as theplasma membrane. In order to overcome this con-straint, we used 2-channel ratiometric TIRF micros-copy (see Materials and methods and Figure 1b) toimage the membrane order of T cells activatedon coverslips with immobilized anti-CD3e antibodies.Figure 3a–c shows a representative brightfieldtransmitted light image, TIRF intensity image and
4 D.M. Owen et al.
Mol
Mem
br B
iol D
ownl
oade
d fr
om in
form
ahea
lthca
re.c
om b
y N
yu M
edic
al C
ente
r on
06/
29/1
0Fo
r pe
rson
al u
se o
nly.

0.25o p 0.3
0.2
0.1
0
-0.1
-0.2
Central Periphery
0.2
0.15
0.1
0.05
GP
val
ue
GP
val
ue
00 5
Distance (µm)10
a b c
d e f
g h i
j k
m n
l
Figure 2.
Membrane order at the immunological synapse 5
Mol
Mem
br B
iol D
ownl
oade
d fr
om in
form
ahea
lthca
re.c
om b
y N
yu M
edic
al C
ente
r on
06/
29/1
0Fo
r pe
rson
al u
se o
nly.

a membrane order (GP) map, respectively, of a liveJurkat T cell synapse imaged approximately 10 minafter the contact with the activating surface. The cellshows a large, spread interface with the antibody-coated coverslip mimicking the IS formed byT cells spreading over the surface of an APC. A higherlipid order accumulation in the IS periphery withreduced order distributed towards the central regionconfirms the results observed during live imaging ofT cells conjugated with APCs. As no 3D reconstruc-tion is involved it is possible to perform accuratequantitative analysis of the data. Figure 3h showsGP values from central and peripheral regionsacquired from 20 individual cells (p = 0.10) and aline profile of the GP values from one, representativecase. The relatively small difference in GP values forcentral and peripheral regions of the IS is probablycaused by contact shape variability and the poorlydefined border between these two regions. Similarly,poorly defined formation of a central area enrichedin receptors has previously been observed in T cellson an activatory surface [Bunnell et al. 2001, 2002a).It is known that cultured cells (e.g., Jurkat T cells)
do not retain all the properties of primary cells. Wetherefore imaged the distribution of membraneorder obtained from primary human CD4+ T cellsactivated on an anti-CD3e antibody-coated coverslip(Figure 3d). The image shows a similar distributionof higher membrane order in the periphery with anindication of the multi-focality observed in synapsesformed between primary T cells and dendritic cellspresenting the antigen on their surface [Brossard et al.2005].The higher signal-to-noise level, lower background,
increased sectioning strength and shorter imageacquisition times of TIRF microscopy enabled usto observe some finer substructures within a largerfield of high order at the IS periphery. Most strikingly,the peripheral ring of higher membrane order repro-ducibly showed discontinuities or small areas of lowerorder suggesting possible sub-compartmentalizationof cellular membranes (Figure 3c and d). As TIRFimages can be acquired at high frame-rates, it is pos-sible to obtain a time-course of the order distribution
as a movie. Selected frames of a representative movie(Supplementary Movie S1, available online) are dis-played in Figure 3e. The general order distributionwas observed to be stable over the imaged time-scale.However, we have been able to monitor points oflower order (blue) diffusing in the higher order back-ground (Figure 3e; arrow). Small mobile areas ofhigh, intermediate and low order, which may repre-sent different vesicular sub-synaptic structures, wereobserved in cells where plasma membrane di-4-ANEPPDHQ dye was reduced by a quick wash(Supplementary Movies S2 and S3, available online,and selected frames in Figure 3f and 3g). We hypothe-size that different vesicle populations may be impor-tant in trafficking of different membrane proteins,based on their affinity to membranes of various com-position and lipid order. Of note, the resolution hereis approximately 300 � 300 � 100 nm (x, y, z) whichis higher than has previously been possible whenimaging membrane order.
Membrane lipid order: The effect of 7-ketocholesterol
Having established the capability to image membraneorder at the IS at high spatial and temporal resolution,we investigated whether changes in membrane order,for example due to pharmacological treatment, can bestudied using our technique. The cholesterol analo-gue 7KC was previously described as an effectivesubstance to reduce plasma membrane order of cul-tured T cells without significant effect on their via-bility and proximal signalling [Rentero et al. 2008]. Asseen in Figure 4a and 4b (imaged by 2-channelratiometric confocal microscopy), higher order ofthe plasma membrane was eliminated by 7KC pre-treatment of T cells. The effect on intracellular mem-branes was less dramatic but the order was stillreduced, as expected. The cells maintained normalmorphology even 90 min after treatment with 7KC.Using TIRF microscopy, the high membrane orderring observed in the IS periphery was absent in 7KC-treated cells activated on anti-CD3-coated coverslips(Figure 4c and d) and quantitation supported the
Figure 2. Membrane order at the synapse of live T cell-APC conjugates. (a) brightfield, (b) GP image and (c) 3D reconstruction showing enface view of the cell-cell interface (dashed line viewed from the direction of the arrow on b; APC position is indicated by dashed ring) of livedi-4-ANEPPDHQ-stained Jurkat T cells forming synapses with unstained APCs (SEE-loaded Raji cells). (d, e and f) second example showingsimilar order distribution. APC (not shown) position in f is indicated with dashed circle and the other T cell is not shown for clarity(representative of n = 8). (g, h and i) and (j, k and l) show two examples of the synapses formed in the absence of antigenic stimulus (SEE;representative of n = 16). The total acquisition time for the 3D stack was approximately 30 sec with both images acquired approximately 10 minafter synapse formation. In all images, red colour represents higher and green/blue lower order membranes. Scale bars 10 mm. T cell synapsesimaged using optical tweezers: Brightfield (m) and GP image (n) of the activatory conjugate formed between Jurkat T cell and APC (as in a)with the IS oriented in the focal plane using optical tweezers. Representative line profile across the synapse (o) and quantitation of the GP valuesin the central and peripheral regions (n = 8, p < 0.05) (p). Scale bar 10 mm. This Figure is reproduced in colour in the online version ofMolecularMembrane Biology.
6 D.M. Owen et al.
Mol
Mem
br B
iol D
ownl
oade
d fr
om in
form
ahea
lthca
re.c
om b
y N
yu M
edic
al C
ente
r on
06/
29/1
0Fo
r pe
rson
al u
se o
nly.

dramatic reduction of membrane order in the IS of7KC-treated activated T cells (Figure 4e). Reducedmembrane order did not, however, affect the ability ofT cells to spread over the activating surface. Of note,treatment of Jurkat T cells with 7KC caused nosignificant changes (< 10%) in the surface expressionof CD3e (Rentero et al. [2008] and data not shown).
Membrane lipid order: Live primary T cells on supportedplanar bilayers
Antibodies immobilized on the glass surface of cover-slips are likely to reduce the diffusional behaviour of
membrane proteins. Incorporating ligands for TCR(peptide-MHC) and adhesion molecules (ICAM-1)into supported planar lipid bilayers represents a morephysiological activating surface for primary T cells[Groves and Dustin 2003]. Supported bilayers allowligands lateral freedom and therefore do not restrictmobility of membrane proteins on T cells. We foundthat di-4-ANEPPDHQ dye in medium shows highaffinity for T cells but also for the planar bilayer.Standard 2-channel intensity imaging is not sensitiveenough to image membrane order of T cells in thepresence of a large disordered-phase backgroundfrom DOPC in the planar bilayer. We have previously
Central Peripheral Position (a.u.)
GP
val
ue
GP
val
ue
0.480.445
0.44
0.435
0.43
0.425
0.42
0.46
0.44
0.42
0.40
e
f
g
b c da
h
Figure 3. Order distribution at the T cell IS formed with activating antibody-coated glass coverslips. (a) Brightfield image showing the maincell body (outlined red) and the large synaptic interface (outlined yellow). (b) TIRF intensity image of the interface. (c) GP image at the synapseshowing higher order (red) and lower order (green to blue) (representative of n = 20). (d) GP image of the interface formed by a primary humanT cell (n = 6). Presented images are of a T cell approximately 10 min after formation of contact with activatory surface. (e) GP images showingdynamics of the observed order distribution (see also Supplementary Movie 1, available online). (f and g) GP images showing low- and high-order putative sub-synaptic vesicles (white arrows) diffusing in the plasma membrane background (see also Supplementary Movies 2 and 3,available online). Acquisition time was 1 sec per frame. (h) Quantitative analysis of GP values in the centre and periphery of Jurkat cell synapses(n = 20, p = 0.10) and a representative line profile across the contact site. Scale bars 5 mm. This Figure is reproduced in colour in the onlineversion of Molecular Membrane Biology.
Membrane order at the immunological synapse 7
Mol
Mem
br B
iol D
ownl
oade
d fr
om in
form
ahea
lthca
re.c
om b
y N
yu M
edic
al C
ente
r on
06/
29/1
0Fo
r pe
rson
al u
se o
nly.

demonstrated that fluorescence lifetime imaging(FLIM) offers enhanced contrast when imagingdi-4-ANEPPDHQ in live cells [Owen et al. 2006].We therefore applied FLIM using single-photon exci-tation and time-correlated single-photon counting(TCSPC) on a scanning confocal microscope to imp-rove the contrast when imaging membrane order ofactivated T cells on supported planar bilayers. Itshould be noted that, due to the dynamic nature ofprimary T cell-bilayer synapses and the longer acqui-sition times required for FLIM, both the spatialand temporal resolution are reduced relative to theantibody-coated coverslip data (Figure 3).Figure 5a–d show brightfield and FLIM images of
primary T cell synapses formed on supported planarbilayers. The low order background (blue) is due to
DOPC present in the bilayer, for which we found asimilar fluorescence lifetime to previously publishedresults [Owen et al. 2006]. Despite the lower spatialand temporal resolution of the measurements, theobserved order distribution at the synapse remainsgenerally consistent with the previous results. Wehypothesize that the discontinuous nature of thehigh order membrane regions around the peripherymay represent membrane sub-compartmentalization.Treatment of primary murine T cells with 7KC(Figure 5e and 5f) disrupted higher order membraneregions that accumulated primarily in the periphery ofthe formed synapses. All these data support the highordering of membrane lipids in the periphery ofthe IS, the area maintained by the dynamic actincytoskeleton and associated with active signalling
0.450.4
0.350.3
0.250.2
0.150.1
0.050
-0.05Untreated
GP
val
ue
Treated
a b
c d
e
Figure 4. Effect of 7KC treatment on membrane order. (a and b) GP images of di-4-ANEPPDHQ-stained Jurkat T cells in suspension imagedusing confocal microscopy without and after 7KC treatment, respectively. (c and d) TIRF GP images of the IS formed on an anti-CD3e antibody-coated coverslip ~ 10 min after contact formation without and after 7KC treatment, respectively. (e) Quantitation ofmembrane order in untreated and 7KC-treated T cell synapses (n = 20) showing reduced order after 7KC treatment (p < 0.001). Scale bars5 mm. This Figure is reproduced in colour in the online version of Molecular Membrane Biology.
8 D.M. Owen et al.
Mol
Mem
br B
iol D
ownl
oade
d fr
om in
form
ahea
lthca
re.c
om b
y N
yu M
edic
al C
ente
r on
06/
29/1
0Fo
r pe
rson
al u
se o
nly.

processes responsible for T cell stimulation [Dustin2009].
Discussion
The formation of large assemblies composed of sig-nalling molecules is thought to improve the efficiencyand robustness of signalling processes in the crowdedand dynamic environment of cells [Cebecauer et al.2010]. In T cells, microclusters containing receptorsand signalling proteins as well as higher order macro-molecular assemblies (such as supramolecular acti-vatory clusters) in the IS play an important role in theinitiation and regulation of processes leading to theiractivation and stimulation of effector functions[Dustin et al. 1998, Monks et al. 1998, Grakoui et al.1999, Bunnell et al. 2002b, Campi et al. 2005,Yokosuka et al. 2005]. The involvement of lipids inthe formation and stabilization of such assemblies hasbeen suggested based on the presence of crucial clustercomponents in isolated detergent-resistant mem-branes [Montixi et al. 1998, Xavier et al. 1998,Drevot et al. 2002] and on co-localization studies ofTCR complex constituents with LR markers inactivated T cells [Janes et al. 1999]. The concept ofLRs’ importance for T cell activation has also been
disputed and protein-protein interactions weresuggested to be the sole force responsible for drivinghigher organization of signalling molecules in the IS[Munro 2003, Douglass and Vale 2005]. Live cellimaging of membrane organization in the IS and itsdependence on lipid composition can thereforeimprove our understanding of molecular processesleading to the T cell activation.Previous work suggested the polarized distribution
of membrane order in T cells forming conjugateswith APC in fixed cells at 25�C [Gaus et al. 2005].Here, we have imaged the distribution of membraneorder at the IS at higher spatial and temporal resolu-tion than has been achieved previously, and for thefirst time in living T cells at physiological temperature.This latter point is crucial because reducing theambient temperature of cells even over a few degreesCelsius has been shown to induce membrane micro-domain aggregation and T cell signalling [Magee et al.2005]. Hence, we have observed slightly increasedtotal GP values in the plasma membrane of unstimu-lated fixed (GP = 0.20 ± 0.05; n = 10) compared to liveJurkat T cells (GP = 0.14 ± 0.02; n = 10). These values,even though not statistically significant (p ~ 0.2),indicate that fixation and lower temperature cancause subtle changes in the organization of plasma
a
e f
b c d
Figure 5. Representative order distributions at the IS formed by primary murine T cells activated on supported planar bilayers (n = 10). (a–d)Brightfield and FLIM images of di-4-ANEPPDHQ-stained primary T cells forming synapses on planar bilayers. (e and f) Brightfield and FLIMimages of T cell synapses after treatment with 7KC. Colour bar represents the mean fluorescence lifetime fitted in each pixel (seeSupplementary Material, available online). Scale bars 5 mm. This Figure is reproduced in colour in the online version of Molecular MembraneBiology.
Membrane order at the immunological synapse 9
Mol
Mem
br B
iol D
ownl
oade
d fr
om in
form
ahea
lthca
re.c
om b
y N
yu M
edic
al C
ente
r on
06/
29/1
0Fo
r pe
rson
al u
se o
nly.

membrane which may shift the fine balance of inter-molecular forces involved in the processes leading tothe activation of T cells.We applied a variety of fluorescence microscopy
techniques including 3D 2-channel confocal imagingand optical tweezer-oriented live cell conjugates aswell as 2-channel ratiometric TIRF microscopy usingactivating antibody-coated glass coverslips. In allcases tested, higher plasma membrane order wasdemonstrated to accumulate preferentially in theperipheral regions of the synapse with lower lipidordering in the central region. The observed distri-bution is in agreement with previously published dataacquired at lower spatial and temporal resolutionusing the dye LAURDAN [Gaus et al. 2005]. Sincesimilar membrane order distribution is observed withLAURDAN, a chemically different molecule, it is alsopossible to exclude potential artefactual protein-dye interactions. Further, high membrane order wasremoved by treatment with the cholesterol oxide,7KC, indicating a cholesterol dependence ratherthan exclusively the effect of membrane proteinsand their binding partners in the stabilization ofhigher order membranes.We have observed partial accumulation of highly
ordered membranes in the junction formed betweenT cells and APCs in the absence of antigen. Thisincrease in membrane order is probably caused by theinteractions of T cell adhesive receptors, such asCD2 and LFA-1 (in low-affinity state), and theirligands on the surface of the APC (CD58 andICAM-1) and the underlying actin cytoskeleton[Springer et al. 1987]. No large-scale segregationinto high order peripheral ring and lower membraneorder central zone has been observed in the absence ofthe antigen. Such uniform distribution of membranelipid order across the junction is similar to the pre-viously reported random localization of surface recep-tors (e.g., LFA-1 and TCR) in T cells formingconjugates with APC in the absence of antigen[Monks et al. 1998].Previously, only steady-state images of membrane
order distribution have been achieved. Here, nochanges in the overall general pattern of a high mem-brane order forming peripheral ring of the IS withreduced lipid ordering in the central zone have beendetected over 1 min using live cell imaging in TIRFmode. With this technique, it was possible to observewhat we hypothesize to be sub-synaptic vesicles oflow, medium and high membrane lipid order whichhave not been previously reported for any cellularsystem. The co-existence of sub-synaptic vesicleswith various levels of membrane order is indicatedin movies of cells where the plasma membranedi-4-ANEPPDHQ dye concentration was reduced
by washing. Formation of sub-membranous vesicularstructures of different properties supports the obser-vation of the presence of proximal T cell signallingmolecules, LAT and SLP-76, in non-overlapping,dynamic clusters [Purbhoo et al. 2010].We have succeeded in imaging membrane order in
live primary CD4+ human T cells using 2-channelTIRF microscopy and have used di-4-ANEPPDHQwith FLIM to image membrane order at the primarymurine T cell synapse formed with activating sup-ported planar bilayers. These results represent a sig-nificant advance in terms of physiological relevancesince many cultured cells of tumour origin (e.g.,Jurkat T cells) have modified membrane lipid com-position and planar bilayers represent a model forAPCs with unrestricted mobility of ligand/receptorpairs, therefore reducing the probability of increasedmembrane order due to restrictions caused by immo-bilized antibodies.All presented data demonstrate formation of an
often discontinuous wide ring of high membrane orderat the IS periphery with reduced lipid ordering in thecentral region. The function of the central synapticregion, the cSMAC, remains controversial since fullyfunctional synapses were demonstrated without anydetectable cSMAC [Lee et al. 2002]. On the otherhand, regulatory and signal enhancing roles wereproposed for the cSMAC based on data acquiredusing weak antigens [Cemerski et al. 2008]. Sinceonly relatively strong activatory signals were appliedin our study, we cannot exclude the possible accumu-lation of more ordered membranes in the centralregion under different stimulatory conditions. Thefunction of highly ordered membranes in peripheraland distal regions of the IS and the mechanism(s) oftheir formation and stabilization remain to beexplored. The accumulation of integrins and associ-ated molecules in the pSMAC supports its generallyaccepted adhesive function [Monks et al. 1998,Grakoui et al. 1999]. Formation of TCR and signal-ling microclusters required for T cell activation wasdemonstrated to occur primarily at the very edge of theIS, the distal region (dSMAC) [Varma et al. 2006].We speculate that lipids forming a higher order mem-brane environment here may help the formation oflarger proteinaceous domains (e.g., TCR microclus-ters) or platforms (e.g., integrin-actin cytoskeletonassemblies during activation) by reducing molecularfluctuations within the plasmamembrane. The latter issupported by the observation of higher membraneorder in the junction between T cells and APCs inthe absence of antigenic stimulus.Weak adhesive inter-actions of individual receptors or small oligomers, butnot of microscopic supramolecular assemblies, areexpected to influence plasma membrane organization
10 D.M. Owen et al.
Mol
Mem
br B
iol D
ownl
oade
d fr
om in
form
ahea
lthca
re.c
om b
y N
yu M
edic
al C
ente
r on
06/
29/1
0Fo
r pe
rson
al u
se o
nly.

ofTcells in the absence ofTCRsignalling [Dustin et al.1987].The IS periphery is also a proximal signalling area
[Varma et al. 2006]. Many signalling proteinsinvolved in immune cell activation were shown tocontain post-translational modifications increasingtheir partitioning to highly ordered domains inmembranes [Lommerse et al. 2006]. Higher ordermembrane environments in the synapse peripherytogether with putative formation of vesicles withdifferent lipid compositions and lipid order (Supple-mentary Movies S1–S3, available online) may presentan elegant way of signalling regulation by com-partmentalization of various signalling pathwaysinto separate substructures.The existence of small (Ø < 40 nm) membrane
microdomains with increased order and thereforeincreased selective molecular confinement wasobserved in live cells indirectly by fluorescence cor-relation spectroscopy [Lenne et al. 2006] and recen-tly directly by super-resolution STED microscopy[Eggeling et al. 2009]. Whether the observed lipidstructure (higher membrane order peripheral ring) isformed by the aggregation of these nanoscopic lipidrafts as has been postulated [Pike 2006] remainsto be demonstrated. The small predicted size ofLRs in resting cells prohibits observation of theirfusion in real time using diffraction-limited imagingtechniques.Taken together, the methods used in this work
have moved the limits of membrane order imagingtowards higher spatial and temporal resolution and tolive cell systems. The observation of high membraneorder at the periphery of the IS where proximalsignalling takes place, and the presence of high andlow order dynamic sub-synaptic structures, under-lines the importance of membrane lipid order inregulating T cell activation.
Acknowledgements
We thank Rajat Varma and Etienne Joly for helpfulsuggestions and critical reading of the manuscript,and Chris Dunsby for helpful comments on the man-uscript and assistance with optical tweezing. PMWFand DMD acknowledge a Wolfson Royal SocietyResearch Merit Award. We also acknowledge theFacility for Imaging by Light Microscopy at ImperialCollege.
Declaration of interest: DMO, SK and SOacknowledge funding from the Engineering and Phys-ical Sciences Research Council via the ChemicalBiology Centre. We acknowledge Medical ResearchCouncil (MRC) grants G0700771 and G0100471 to
AIM and National Institutes of Health grant AI43542to MLD. DMD acknowledges funding from theMRC. The authors report no conflicts of interest.The authors alone are responsible for the content andwriting of the paper.
References
Bromley SK, Burack WR, Johnson KG, Somersalo K, Sims TN,Sumen C, Davis MM, Shaw AS, Allen PM, Dustin ML.2001. The immunological synapse. Annual Rev Immunol19:375–396.
Brossard C, Feuillet V, Schmitt A, Randriamampita C, Romao M,Raposo G, Trautmann A. 2005. Multifocal structure of the Tcell-dendritic cell synapse. Eur J Immunol 35:1741–1753.
Bunnell SC, Hong DI, Kardon JR, Yamazaki T, McGlade CJ,Barr VA, Samelson LE. 2002a. T cell receptor ligation inducesthe formation of dynamically regulated signaling assemblies.J Cell Biol 158:1263–1275.
Bunnell SC, Hong DI, Kardon JR, Yamazaki T, McGlade CJ,Barr VA, Samelson LE. 2002b. T cell receptor ligation inducesthe formation of dynamically regulated signaling assemblies.J Cell Biol 158:1263–1275.
Bunnell SC, Kapoor V, Trible RP, Zhang W, Samelson LE. 2001.Dynamic actin polymerization drives T cell receptor-inducedspreading: A role for the signal transduction adaptor LAT.Immunity 14:315–329.
Burack WR, Lee K-H, Holdorf AD, Dustin ML, Shaw AS. 2002.Cutting edge: Quantitative imaging of raft accumulation in theimmunological synapse. J Immunol 169:2837–2841.
Campi G, Varma R, Dustin ML. 2005. Actin and agonistMHC-peptide complex-dependent T cell receptor microclustersas scaffolds for signaling. J Experim Med 202:1031–1036.
Cebecauer M, Owen DM, Markiewicz A, Magee AI. 2009. Lipidorder and molecular assemblies in the plasma membrane ofeukaryotic cells. Biochem Soc Transact 37:1056–1060.
Cebecauer M, Spitaler M, Serge A, Magee AI. 2010. Signallingcomplexes and clusters: Functional advantages and methodo-logical hurdles. J Cell Sci 123:309–320.
Cemerski S, Das J, Giurisato E, Markiewicz MA, Allen PM,Chakraborty AK, Shaw AS. 2008. The balance between Tcell receptor signaling and degradation at the center of theimmunological synapse is determined by antigen quality. Immu-nity. 29:414-422.
Davis DM, Chiu I, Fassett M, Cohen GB, Mandelboim O,Strominger JL. 1999. The human natural killer cell immunesynapse. Proc Nat Acad Sci USA 96:15062–15067.
Day CA, Kenworthy AK. 2009. Tracking microdomain dynamicsin cell membranes. Biochimic Biophys Acta 1788:245–253.
Douglass AD, Vale RD. 2005. Single-molecule microscopy revealsplasma membrane microdomains created by protein-proteinnetworks that exclude or trap signaling molecules in T cells.Cell 121:937–950.
Drevot P, Langlet C, Guo X-J, Bernard A-M, Colard O,Chauvin J-P, Lasserre R, He H-T. 2002. TCR signal initiationmachinery is pre-assembled and activated in a subset of mem-brane rafts. EMBO J 21:1899–1908.
Dustin ML. 2009. The cellular context of T cell signaling. Immu-nity 30:482–492.
Dustin ML, Olszowy MW, Holdorf AD, Li J, Bromley S, Desai N,Widder P, Rosenberger F, Anton van der Merwe P,Allen PM, et al. 1998. A novel adaptor protein orchestratesreceptor patterning and cytoskeletal polarity in T-cell contacts.Cell 94:667–677.
Membrane order at the immunological synapse 11
Mol
Mem
br B
iol D
ownl
oade
d fr
om in
form
ahea
lthca
re.c
om b
y N
yu M
edic
al C
ente
r on
06/
29/1
0Fo
r pe
rson
al u
se o
nly.

Dustin ML, Sanders ME, Shaw S, Springer TA. 1987. Purifiedlymphocyte function-associated antigen-3 binds to CD2 andmediates lymphocyte-T adhesion. J ExperimMed 165:677–692.
Eggeling C, Ringemann C, Medda R, Schwarzmann G,Sandhoff K, Polyakova S, Belov VN, Hein B,von Middendorff C, Schonle A, et al. 2009. Direct observationof the nanoscale dynamics of membrane lipids in a living cell.Nature 457:1159–1163.
Gaus K, Chklovskaia E, Fazekas de St Groth B, Jessup W,Harder T. 2005. Condensation of the plasma membrane atthe site of T lymphocyte activation. J Cell Biol 171:121–131.
Gaus K, Gratton E, Kable EPW, Jones AS, Gelissen I,Kritharides L, Jessup W. 2003. Visualizing lipid structure andraft domains in living cells with two-photon microscopy. ProcNat Acad Sci USA 100:15554–15559.
Gaus K, Zech T, Harder T. 2006. Visualizing membrane micro-domains by Laurdan 2-photon microscopy. Molec Memb Biol23:41–48.
Grakoui A, Bromley SK, Sumen C, Davis MM, Shaw AS,Allen PM, Dustin ML. 1999. The immunological synapse:A molecular machine controlling T cell activation. Science285:221–227.
Groves JT, Dustin ML. 2003. Supported planar bilayers in studieson immune cell adhesion and communication. J ImmunologMeth 278:19–32.
Janes PW, Ley SC, Magee AI. 1999. Aggregation of lipid raftsaccompanies signaling via the T cell antigen receptor. J Cell Biol147:447–461.
Jin L, Millard AC, Wuskell JP, Dong X, Wu D, Clark HA,Loew LM. 2006. Characterization and application of a newoptical probe for membrane lipid domains. Biophys J 90:2563–2575.
Lee K-H, Dinner AR, Tu C, Campi G, Raychaudhuri S, Varma R,Sims TN, Burack WR, Wu H, Wang J, et al. 2003. Theimmunological synapse balances T cell receptor signaling anddegradation. Science 302:1218–1222.
Lee K-H, Holdorf AD, Dustin ML, Chan AC, Allen PM,Shaw AS. 2002. T cell receptor signaling precedes immuno-logical synapse formation. Science 295:1539–1542.
Lenne P-F, Wawrezinieck L, Conchonaud F, Wurtz O, Boned A,Guo X-J, Rigneault H, He H-T, Marguet D. 2006. Dynamicmolecular confinement in the plasma membrane by microdo-mains and the cytoskeleton meshwork. EMBO J 25:3245–3256.
Lommerse PHM, Vastenhoud K, Pirinen NJ, Magee AI,Spaink HP, Schmidt T. 2006. Single-molecule diffusion revealssimilar mobility for the Lck, H-Ras, and K-Ras membraneanchors. Biophys J 91:1090–1097.
Magee AI, Adler J, Parmryd I. 2005. Cold-induced coalescence ofT-cell plasma membrane microdomains activates signallingpathways. J Cell Sci 118:3141–3151.
Monks CRF, Freiberg BA, Kupfer H, Sciaky N, Kupfer A. 1998.Three-dimensional segregation of supramolecular activationclusters in T cells. Nature 395:82–86.
Montixi C, Langlet C, Bernard AM, Thimonier J, Dubois C,Wurbel MA, Chauvin JP, Pierres M, He HT. 1998. Engage-ment of T cell receptor triggers its recruitment to low-densitydetergent-insoluble membrane domains. EMBO J 17:5334–5348.
Munro S. 2003. Lipid rafts: Elusive or illusive? Cell 115:377–388.
Obaid AL, Loew LM, Wuskell JP, Salzberg BM. 2004. Novelnaphthylstyryl-pyridinium potentiometric dyes offer advan-tages for neural network analysis. J NeuroscMeth 134:179–190.
Oddos S, Dunsby C, Purbhoo MA, Chauveau A, Owen DM,Neil MAA, Davis DM, French PMW. 2008. High-speedhigh-resolution imaging of intercellular immune synapses usingoptical tweezers. Biophys J 95:L66–68.
Owen DM, Lanigan PMP, Dunsby C, Munro I, Grant D,Neil MAA, French PMW, Magee AI. 2006. Fluorescencelifetime imaging provides enhanced contrast when imagingthe phase-sensitive dye di-4-ANEPPDHQ in model membranesand live cells. Biophys J 90:L80–82.
Parasassi T, De Stasio G, d’Ubaldo A, Gratton E. 1990. Phasefluctuation in phospholipid membranes revealed by Laurdanfluorescence. Biophys J 57:1179–1186.
Pike LJ. 2006. Rafts defined: A report on the Keystone symposiumon lipid rafts and cell function. J Lipid Res 47:1597–1598.
Purbhoo MA, Liu H, Oddos S, Owen DM, Neil MA, Pageon SV,French PM, Rudd CE, Davis DM. 2010. Dynamics of sub-synaptic vesicles and surface microclusters at the immunologicalsynapse. Sci Signal. 3:ra36.
Rentero C, Zech T, Quinn CM, Engelhardt K, Williamson D,Grewal T, Jessup W, Harder T, Gaus K. 2008. Functionalimplications of plasma membrane condensation for T cellactivation. PLoS ONE 3:e2262.
Sengupta P, Baird B, Holowka D. 2007. Lipid rafts, fluid/fluidphase separation, and their relevance to plasma membranestructure and function. Semin Cell Develop Biol 18:583–590.
Sharma P, Varma R, Sarasij RC, Ira Gousset K,Krishnamoorthy G, Rao M, Mayor S. 2004. Nanoscale orga-nization of multiple GPI-anchored proteins in living cell mem-branes. Cell 116:577–589.
Simons K, Ikonen E. 1997. Functional rafts in cell membranes.Nature 387:569–572.
Springer TA, Dustin ML, Kishimoto TK, Marlin SD. 1987. Thelymphocyte function-associated LFA-1, CD2 and LFA-3 mole-cules: Cell adhesion receptors of the immune system. AnnualRev Immunol 5:223–252.
Suzuki KGN, Fujiwara TK, Sanematsu F, Lino R, Edidin M,Kusumi A. 2007. GPI-anchored receptor clusters transientlyrecruit Lyn and G{alpha} for temporary cluster immobilizationand Lyn activation: Single-molecule tracking study 1. J Cell Biol177:717–730.
Tavano R, Gri G, Molon B, Marinari B, Rudd CE, Tuosto L,Viola A. 2004. CD28 and lipid rafts coordinate recruitment ofLck to the immunological synapse of human T lymphocytes.J Immunol 173:5392–5397.
Varma R, Campi G, Yokosuka T, Saito T, Dustin ML. 2006. T cellreceptor-proximal signals are sustained in peripheral microclus-ters and terminated in the central supramolecular activationcluster. Immunity 25:117–127.
Xavier R, Brennan T, Li Q, McCormack C, Seed B. 1998. Mem-brane compartmentation is required for efficient T cell activa-tion. Immunity 8:723–732.
Yokosuka T, Sakata-Sogawa K, Kobayashi W, Hiroshima M,Hashimoto-Tane A, Tokunaga M, Dustin ML,Saito T. 2005. Newly generated T cell receptor microclustersinitiate and sustain T cell activation by recruitment ofZap70 and SLP-76. Nature Immunol 6:1253–1262.
Supplementary material available online
Supplementary informationMovies 1, 2 and 3.
12 D.M. Owen et al.
Mol
Mem
br B
iol D
ownl
oade
d fr
om in
form
ahea
lthca
re.c
om b
y N
yu M
edic
al C
ente
r on
06/
29/1
0Fo
r pe
rson
al u
se o
nly.
![Lipid assembly into cell membranes - IJSbio.ijs.si/~krizaj/group/Predavanja 2011/Biochemistry Lipids... · membrane lipid asymmetry are found in the red blood cell membrane [3], and](https://img.pdfslide.us/doc/110x75/5e324dd387dca6413522f348/lipid-assembly-into-cell-membranes-krizajgrouppredavanja-2011biochemistry-lipids.jpg)