Embed Size (px)
Citation preview

1
2
3
High-efficiencyoptogeneticsilencingwithsoma-targetedanion-4
conductingchannelrhodopsins5
6
7
8
MathiasMahn,LihiGibor,KatayunCohen-KashiMalina,PritishPatil,YoavPrintz,ShirOring,RivkaLevy,9
IlanLamplandOferYizhar10
11
DepartmentofNeurobiology,WeizmannInstituteofScience,Rehovot,Israel12
13
14
15
Correspondingauthor:16
OferYizhar,Ph.D.17
DepartmentofNeurobiology18
WeizmannInstituteofScience19
234Herzlst,Rehovot761000120
Israel21
Tel.+972-8-934695722
peer-reviewed) is the author/funder. All rights reserved. No reuse allowed without permission. The copyright holder for this preprint (which was not. http://dx.doi.org/10.1101/225847doi: bioRxiv preprint first posted online Dec. 8, 2017;

Abstract24
Optogenetic silencing allows time-resolved functional interrogation of defined neuronal populations.25
However, the limitationsof inhibitoryoptogenetic tools impose stringent constraintsonexperimental26
paradigms.Thehighlightpowerrequirementoflight-drivenionpumpsandtheireffectsonintracellular27
ion homeostasis pose unique challenges, particularly in experiments that demand inhibition of a28
widespreadneuronalpopulationinvivo.Guillardiathetaanion-conductingchannelrhodopsins(GtACRs)29
are promising in this regard, due to their high single-channel conductance and favorable photon-ion30
stoichiometry.However,GtACRsshowpoormembranetargetinginmammaliancells,andtheactivityof31
suchchannelscancausetransientexcitationintheaxonduetoanexcitatorychloridereversalpotential32
inthiscompartment.Hereweaddressbothproblemsbyenhancingmembranetargetingandsubcellular33
compartmentalization of GtACRs. The resulting GtACR-based optogenetic tools show improved34
photocurrents,greatlyreducedaxonalexcitation,highlightsensitivityandrapidkinetics,allowinghighly35
efficientinhibitionofneuronalactivityinthemammalianbrain.36
37
Introduction38
Perturbationofneuronalactivityisafundamentalaspectofneuroscienceresearch,oftenusedtogain39
insight into the functional rolesofparticularbrain regions, circuits and cell types 1.Optogenetic tools40
havegreatlyenhancedtheprecisionwithwhichsuchmanipulationscanbeperformed2,providingboth41
temporal precision and cell-type specificity to experiments aimed at defining the roles of individual42
neuralcircuitcomponents inneuralcomputationoranimalbehavior.Currentoptogeneticapproaches43
forsilencingofneuronsaremainlybasedonthelight-activatedmicrobialrhodopsinshalorhodopsin3,4,44
archaerhodopsin 5andcruxhalorhodopsin 6.Theseproteinspump ionsacross theneuronalmembrane45
with millisecond kinetics, independently of the electrochemical gradient. These tools allow neuronal46
silencingwithprecisetemporalonsetandoffset5,7,8.However,ion-pumpingrhodopsinspossessseveral47
characteristics that impose substantial constraints on the experimental paradigm and complicate the48
interpretation of experimental outcomes. These limitations become evenmore pronounced in cases49
whereneuronalsilencingisrequiredforextendedperiodsoftime.Theunfavorablestoichiometryofone50
transportedionforeachabsorbedphotonnecessitatescontinuousilluminationathighlightpower.The51
resultingtissueheating9,10andphototoxicity11restricttheopticallyaddressablebrainvolumethatcan52
beefficientlysilenced.Furthermore,ion-pumpingmicrobialrhodopsinsexhibitadeclineinphotocurrent53
peer-reviewed) is the author/funder. All rights reserved. No reuse allowed without permission. The copyright holder for this preprint (which was not. http://dx.doi.org/10.1101/225847doi: bioRxiv preprint first posted online Dec. 8, 2017;

amplitudesofupto90%withinaminuteofillumination,leadingtoreducedsilencingefficacyovertime5412,8,13.Becauseoftheirinsensitivitytoelectrochemicalgradients,ion-pumpingmicrobialrhodopsinscan55
shift the concentrationsof intracellular ions tonon-physiological levels. In the caseof halorhodopsin,56
this can lead toaccumulationof chloride in theneuron, inducing changes in the reversal potential of57
GABAergic synapses 14. While in the case of archaerhodopsin this can increase the intracellular pH,58
inducing action potential-independent Ca2+ influx and elevated spontaneous vesicle release 13.59
Furthermore,thehyperpolarizationmediatedbyion-pumpingactivitytogetherwiththefastoffkinetics60
canleadtoanincreasedfiringrateuponterminationoftheillumination6,13. 61
Anion-conducting channelrhodopsins (ACRs), a newly established set of optogenetic tools 15,16,17, are62
distinctfromion-pumpingrhodopsinsintwomajoraspects:first,theycanconductmultipleionsduring63
each photoreaction cycle. This increased photocurrent yield per photon makes channelrhodopsins64
superiorintermsoftheiroperationallight-sensitivity.Second,conductingionsaccordingtothereversal65
potential, ACRs are more likely to avoid non-physiological changes in ion concentration gradients. A66
light-gatedchlorideconductancewillshuntmembranedepolarization,whichcanbeusedtoeffectively67
clamp the neuronal membrane potential to the reversal potential of chloride, given that the ion68
permeabilityissufficientlyhigh.Anion-conductingchannelrhodopsinscouldthereforerelieveconstrains69
imposed by ion-pumping rhodopsins. The naturally-occurring anion-conducting channelrhodopsins70
(nACRs) from the cryptophyte algaGuillardia theta 16 are particularly interesting in this regard. These71
channelrhodopsins,namedGtACR1andGtACR2,havenear-perfectanionselectivityandproducelarge72
photocurrentsinmammaliancells,owingtoahighersingle-channelconductancethanthatoftheknown73
cation-conductingchannelrhodopsins16,18.WhileGtACRswereshowntoinhibitbehaviorinthefruitfly7419,20and larval zebrafish 21, theyhavenotyetbeenapplied tomammaliansystems,most likelydue to75
poor membrane targeting and complex activity in the axonal compartment. To overcome these76
limitationsofGtACRsandtherebyofoptogeneticinhibitioningeneral,wegeneratedseveralmembrane77
targeting-enhanced GtACR variants, converging onto soma-targeted GtACR2 (stGtACR2), a fusion78
construct thatcombinesGtACR2withaC-terminal targetingmotif fromthesoma-localizedpotassium79
channelKv2.122.WedemonstrateherethatstGtACR2showsincreasedmembranetargeting,extremely80
high anion photocurrents and reduced axonal excitation, making it the most effective tool for81
optogeneticinhibitionatthecellsomatodate.82
83
peer-reviewed) is the author/funder. All rights reserved. No reuse allowed without permission. The copyright holder for this preprint (which was not. http://dx.doi.org/10.1101/225847doi: bioRxiv preprint first posted online Dec. 8, 2017;

Results84
GtACR2efficientlysilencesneuronalactivityinvitroandinvivo85
To determine the utility of ACRs for silencing of neurons we first expressed the three previously-86
described blue light-activated ACRs, GtACR2 16, iC++ 17, and iChloC 15, in cultured rat hippocampal87
neurons by adeno associated virus (AAV)-mediated gene transfer.Whole-cell patch-clamp recordings88
fromGtACR2-expressingneurons showed reliableoutwardphotocurrents (Fig. 1a) in response to47089
nmfullfieldlightpulses.Thephotocurrentafter1sofcontinuousillumination(stationaryphotocurrent)90
ofGtACR2expressingneuronsclampedto-35mVwassignificantlyhigherthanthatoftheengineered91
ACRs(eACRs)iC++andiChloC(628.5±61.8pA,330.2±37.9pA,and136.3±21.4pA,respectively;Fig.92
1b). Given the poormembrane targeting and intracellular accumulation ofGtACR2 (Fig. 1c), the high93
single-channelconductanceofGtACR216 is likelythecauseforthehighphotocurrentsobservedinthe94
whole-cell recordings.While thenativeGtACR2 seems tooutperform thepreviously-describedeACRs,95
the above findings suggest that improved membrane targeting of GtACR2 would greatly facilitate96
silencing of neuronal activity. Importantly, this should allow efficient silencing by using significantly97
lowerlightpower,enablingoptogeneticcontrolofalargerbrainvolume.98
To verify this prediction, we characterized the efficiency of GtACR2-mediated inhibition in awake,99
behavingmicewithextracellularrecordings(Fig.1d-g).Toquantifytheefficiencyofsilencinginalarge100
cortical volume, we recorded from mice expressing GtACR2 in pyramidal neurons in the medial101
prefrontal cortex (mPFC). Mice were implanted with movable fiberoptic-coupledmicrowire arrays in102
whichelectrodeswereplaced500µmbelowtheopticalfibertip(Fig.1d).Usinganalyticalmodelingof103
lightscatteringandabsorptioninbraintissue23weestimatedthelightpowerdensityatthepositionof104
the extracellular recording site to be 0.11% of the light power density exiting the optical fiber105
(SupplementaryFig.1).Single-unitrecordingsshowedasignificantreductioninneuronalfiringratesin106
responsetoa5s460nm lightpulse (Fig.1e-h).Theserecordingsshowedthatoutof100singleunits107
recorded (n = 2mice, 12 recording sites), 43% and 35% significantly reduced their firing rate at the108
secondhighestandhighesttestedlightpowerdensities(0.5and1mWmm-2,respectively;Fig.1f).Half109
ofthesilencedunitsshowedasignificantfiringratereductionalreadyatthelowesttestedlightpower110
density(125µWmm-2),withsomeunitscompletelysilenced(Fig.1e,h).Atthehighest light intensity,111
silencedunitsshowedpronouncedreboundactivityupon lightpulsetermination (Fig.1g).Notably,all112
lightpowerdensitiesusedwerewellbelowthenecessarylightpowersforinvivooptogeneticinhibition113
peer-reviewed) is the author/funder. All rights reserved. No reuse allowed without permission. The copyright holder for this preprint (which was not. http://dx.doi.org/10.1101/225847doi: bioRxiv preprint first posted online Dec. 8, 2017;

usingmicrobialion-pumps8,6,indicatingthatGtACRscanserveaspotentinhibitoryoptogenetictoolsfor114
somaticsilencinginmammalianneurons.115
ActivationofACRsintheaxonalcompartmentinducesantidromicactionpotentials116
We have previously shown that GtACR1 can induce vesicle release from thalamocortical projection117
neuronsuponilluminationoftheiraxonalterminalsintheacutebrainslice13.Malyshevandcolleagues118
24demonstratedasimilareffectinGtACR2-expressingcorticalpyramidalneuronsinacutebrainslices.119
ToverifythatthisexcitatoryactionofGtACRsisnotanartifactoftheacutebrainslicepreparation,we120
evaluatedtheexcitatoryeffectofACRsintwoseparatepreparations:inculturedhippocampalneurons121
and in awake, freely moving mice. Whole-cell patch-clamp recordings in current-clamp mode from122
GtACR2-expressing cultured neurons revealed that during a 100 ms long illumination pulse, GtACR2123
reliably inhibited action potential (AP) generation (Fig. 2a). However, this was often associated with124
what appeared to be an attenuated AP shortly after light onset (‘escaped AP’, Fig. 2a, inset).When125
recordedinvoltage-clampmode,escapedAPsweremeasuredin6outof12testedGtACR2-expressing126
neuronsinresponseto1mslightpulsesatsaturatinglightpower(4.5mWmm-2).Theseescapedspikes127
occurredevenwhentherecordedphotocurrentwasanoutwardcurrent,expectedtohyperpolarizethe128
somaticmembrane(Fig.2b,upperleft).Wethenaskedwhethertheselight-evokedantidromicAPsare129
specific to naturally-occurring chloride-channels, or a general feature of light-evoked chloride130
conductance in the axon. We therefore tested two engineered anion-conducting channelrhodopsins131
underthesameexperimentalconditions.APswereevokedinresponseto1msand1slonglightpulses132
in 2 / 13 and 7 / 12 iC++ expressing neurons, respectively, showing that this effect is not specific to133
GtACRs(Fig.2b,middle).NoAPswereevokediniChloCexpressingcells(n=22andn=15,for1msand134
1s longlightpulses,respectively;Fig.2bright), likelybecauseoftheoverallsmallerphotocurrentswe135
observed incellsexpressingthisconstruct (Fig.1b).To furthertest thehypothesis thatACRactivation136
mightbedepolarizing in someneuronal compartments,weapplied spatially-restricted laserpulses to137
thesomaorneuritesofculturedhippocampalneuronsduringwhole-cellpatch-clamprecordings.Light138
pulsesdirectedat the somausingagalvanometricmirror system (seeOnlineMethods) inducedsmall139
hyperpolarizingordepolarizingphotocurrents,while lightpulsesdirected toneuritesof the samecell140
evokedantidromicspikes(Fig.2c).141
Theserecordingswereperformedinculturedneuronsafterat least14days invitro,astageatwhich142
intracellular chloride concentrations should reach theadult state 25 as theexpressionof theneuronal143
potassiumchlorideco-transporterKCC226isfullyup-regulated27.Nevertheless,chloridehomeostasisof144
peer-reviewed) is the author/funder. All rights reserved. No reuse allowed without permission. The copyright holder for this preprint (which was not. http://dx.doi.org/10.1101/225847doi: bioRxiv preprint first posted online Dec. 8, 2017;

neuronalculturesmaydifferduetoreducedconcentrationsofKCC2regulatorssuchasinsulin28inthe145
culturemedium.WethereforetestedwhetheractivationofGtACR2could leadtoaxonalexcitation in146
vivo.Werecorded fromGtACR2-expressingmice (Fig.1d)usingmovable fiberoptic-coupledmicrowire147
drivesatthesiteofAAVinjection(Fig.1d-h).Inthesameanimals,weimplantedasecondopticalfiber148
terminatingatthenucleusaccumbens(NAc),aprominentprojectiontargetofthemPFC29 (Fig.2d). In149
responsetobrieflightpulsestothemPFC(5mspulsewidth,460nmat1mWmm-2,whichcorresponds150
to28.8mWatthefibertip),APswereevoked(Fig.2e,g)inthesameAAV-expressingregionthatshowed151
significantsilencingduring5s lightpulses (Fig.1d-h).Moreover, similar5ms lightpulsesdeliveredto152
the NAc led to the induction of short-latency APs in themPFC (Fig. 2f-g), presumably due to axonal153
excitationandantidromicpropagation.154
Insummary,lightstimulationofGtACR2-expressingaxonsledtoAPsinhippocampalneuronsinvitro,in155
thalamocorticalprojectionneuronsinacutebrainslices13andinstriatum-projectingcorticalneuronsin156
awake, behaving mice. We therefore concluded that chloride-mediated axonal depolarization is a157
generalphenomenonthatcouldconfoundtheanalysisofoptogeneticsilencingexperiments.158
OverexpressionofKCC2reducesGtACR2-mediatedantidromicactionpotentials 159
Based on these findings, we reasoned that if GtACR-mediated antidromic spiking is indeed due to a160
positivelyshiftedchloridereversalpotential in theaxon,decreasing theaxonalchlorideconcentration161
should reduce the probability of antidromic spike generation. The chloride extruder KCC2 26 is up-162
regulated inneuronsduringdevelopment, leading tohighendogenousKCC2protein levels in somatic163
and dendritic membranes 30, but does not localize to axons 31,32,33. We first tested whether164
overexpression of KCC2 leads to its localization to the axonal compartment in cultured hippocampal165
neurons.We co-transfected neurons with expression vectors encoding the green fluorescent protein166
mNeonGreen (GFP) and KCC2, or with GFP alone as control. We then labeled the neurons with167
antibodies against the dendrite-specific microtubule-associated protein-2 (MAP2) and KCC2.168
Overexpression of KCC2 led to strong KCC2 immunoreactivity in dendrites and somata of transfected169
neurons(Fig.3a),comparedtoendogenousexpressionlevels(Fig.3a,whitearrow).Toquantifyaxonal170
KCC2 levels,axonsweredetectedasneurites thatareGFP-positiveandMAP2-negative (Fig.3a, zoom171
in).WhilemeanaxonalKCC2intensitywasnotsignificantlydifferentbetweenyoung(7daysinvitro)and172
mature(16daysinvitro)hippocampalcultures,KCC2overexpressionledtoa6.6±1.1foldhigheraxonal173
KCC2signal(Fig.3b,ctrlyoungvs.ctrlmature:P=0.20;ctrlyoungvs.KCC2:P=5*10-7;ctrlmaturevs.174
KCC2:P=9.8*10-3).KCC2expressiondidnotsignificantlyshiftthechloridereversalpotentialmeasured175
peer-reviewed) is the author/funder. All rights reserved. No reuse allowed without permission. The copyright holder for this preprint (which was not. http://dx.doi.org/10.1101/225847doi: bioRxiv preprint first posted online Dec. 8, 2017;

inthesoma(Fig.3c)ortheactionpotentialinitiationthreshold(rheobase,Fig.3d),butindeedledtoa176
significantreductionofGtACR2-evokedantidromicspiking(1mslightpulseswidth,470nmat4.5mW177
mm-2;Fig.3e).Thisresultprovidesfurthersupportforthenotionthatthechloridereversalpotentialin178
theaxonisdepolarizingunderphysiologicalconditions.179
180
Soma-targetingofGtACR2increasessomaticphotocurrentsandreducesaxonalexcitation181
ToovercomethetwomaincaveatsofGtACRsinrespecttotheirutilityasoptogeneticinhibitorytools,182
namelytheirpoormembranetargetingandtriggeringofantidromicspikesintheaxonalcompartment,183
we designed several new GtACR2 variants with altered membrane targeting sequences (Fig. 4a).184
Addition of the ER export and trafficking signals from the mammalian inward rectifying potassium-185
channelKir2.1waspreviouslyshowntoreduceintracellularaggregationofthechloride-pumpNpHR34,35.186
Indeed,fusionofthesesequencestoGtACR2(eGtACR2)ledtoreducedintracellularaccumulationinvivo187
(Fig. 4c, 2nd column). To reduce antidromic spike generation, GtACR2 should be removed from the188
axonalcompartment.Toachievesoma-specificlocalizationofGtACR2,wereplacedtheERexportsignal189
with the soma-targetingmotif of the soma and proximal dendrite localized voltage-gated potassium-190
channel Kv2.1 36,37, which was previously shown to enhance soma-localized expression of191
channelrhodopsin-2 38,22.Hypothesizingthatdestabilizing theproteinbyaddingaproteindegradation-192
promotingproline(P),glutamicacid(E),serine(S),andthreonine(T)richsequence(PEST39)couldlimit193
the effective lifetime of membrane-resident channels diffusing along the axon, thereby potentially194
further restricting GtACR2 protein levels outside the somatic compartment. To test these new viral195
constructs, we infused AAVs encoding them unilaterally into the mPFC of mice, leading to strong196
fluorescence at the injection site aswell as sparse labeling along the injection needle track (Fig. 4b).197
Soma-targetedGtACR2(stGtACR2;Fig.4c,3rdcolumn)aswellasthedestabilizedstGtACR2-PEST(Fig.4c198
,4thcolumn)showedimprovedmembranetargeting,strongsoma-associatedfluorescenceandreduced199
neuritefluorescence(Fig.4d).Functionalcharacterizationofthesoma-targetedconstructsbywhole-cell200
patch-clamprecordings intheacutebrainsliceshoweda2.6-fold increase instationaryphotocurrents201
comparedtountargetedGtACR2,leadingtoaveragephotocurrentsofmorethan2nAwhencellswere202
clamped to -35mV (Fig. 4e). Improvedmembrane targeting alone strongly increased the antidromic203
spikegenerationprobability(Fig.5a;eGtACR2),whilesoma-targetingnotonlyincreasedphotocurrents204
(Fig. 4e) but also decreased the probability of inducing antidromic spikes in cultured hippocampal205
neurons(Fig.5a).DestabilizingstGtACR2usingthePESTsequenceledtoalesspronouncedreductionin206
peer-reviewed) is the author/funder. All rights reserved. No reuse allowed without permission. The copyright holder for this preprint (which was not. http://dx.doi.org/10.1101/225847doi: bioRxiv preprint first posted online Dec. 8, 2017;

antidromicspikegenerationcomparedtostGtACR2(Fig.5a).Photocurrentswerequantifiedinthesame207
neuronstoverifythatthereducedprobabilityofantidromicspikegenerationisnotduetodifferencesin208
peakphotocurrentsofthedifferentconstructsinculturedneurons(Fig.5b).Incontrasttothestationary209
photocurrentsinacutebrainsliceexperiments(Fig.4e)peakphotocurrentsinculturedneuronsdidnot210
differ significantly between constructs, pointing to a lowermembrane targeting efficiency in cultured211
neuronsoraninfluenceoftheshortervirusincubationtime.Nevertheless,itfollowsthatthedramatic212
reductioninantidromicspikingforstGtACR2isnotduetolowerphotocurrents.213
We next asked whether stGtACR2 and stGtACR2-PEST would show improved performance through214
reducedlight-evokedsynapticreleasefromlong-rangeprojectingaxons.WeinjectedAAVsencodingthe215
GtACR2variantstogetherwithasecondAAVencodingacell-fillingfluorophoreunilaterallytothemPFC,216
to allow for visualization of the axons of cortico-cortical projection neurons in the contralateral217
hemisphere (Fig. 5c). During acute brain slice preparation from thesemice, the corpus callosumwas218
severed, separating the somata of the transduced cortico-cortical projecting neurons from their axon219
terminals in the contralateralmPFC. Conductingwhole-cell patch-clamp recordings frompostsynaptic220
neuronsinareaswithfluorescently-labeledaxonscontralateraltotheinjectionsitethereforeallowedus221
tocharacterizetheisolatedeffectofGtACR2activationontheaxonalcompartment.Bluelightpulsesled222
to reliably evoked EPSCs in slices expressing thenon-targetedGtACR2 (Fig. 5d). In contrast, the EPSC223
amplitudewasdramaticallyreducedinslicesexpressingthesoma-restrictedstGtACR2(Fig.5d,e;473.4±224
153.4pAvs.34.1±9.5pAforGtACR2andstGtACR2,respectively).Theincreasedsomaticphotocurrents225
of stGtACR2, together with the near-elimination of antidromic spiking and neurotransmitter release,226
makeitahighlyefficienttoolforoptogeneticinhibition.227
stGtACR2mediatedBLAinhibitionpreventsfearextinctionlearning228
ToverifytheutilityofstGTACR2-mediatedoptogeneticinhibitioninawake,behavinganimals,wechose229
touse this tool for suppressingbasolateral-amygdala (BLA) activityduring extinctionof auditory-cued230
fearconditioning,awell-establishedformofassociative learning40.TheBLAplaysacentralrole inthe231
acquisition aswell as extinction of the conditioned freezing response. Based on previouswork 41,we232
hypothesizedthattemporally-preciseinhibitionoftheBLAduringthedeliveryofconditionedstimuliin233
extinction training would suppress the formation of extinctionmemory.We bilaterally injectedmice234
withAAVencodingstGtACR2orafluorophore-onlycontrolvectorintotheBLAandimplanted200µm-235
diameteroptical fibers above the injection sites (Fig. 6a; Supplementary Fig. 2). Following3weeksof236
recovery,miceunderwentfearconditioningincontextA(Fig.6b).Bothgroups(stGtACR2,n=8;control,237
peer-reviewed) is the author/funder. All rights reserved. No reuse allowed without permission. The copyright holder for this preprint (which was not. http://dx.doi.org/10.1101/225847doi: bioRxiv preprint first posted online Dec. 8, 2017;

n=8)showedincreasedfreezingduringacquisition(ctrl:from3.8±1.9%to39.5±8.1%;stGtACR2:from238
3.5±1.5% to29.6±3.5%, ScheirerRayHare testH=52.91,P=3.5*10-10)withno significantdifference239
between groups (ctrl vs. stGtACR2: Scheirer Ray Hare test H = 0.11, P = 0.74), suggesting that BLA240
activity isnotalteredmerelybyexpressionofstGtACR2(Fig.6c).Totestforfearrecallandextinction,241
miceunderwentextinctiontrainingtwodays later inadifferentcontext fromthat inwhichtheywere242
fearconditioned(contextB).Theextinctionprotocolconsistedof twenty30s tonepresentationsthat243
werepairedwithblue lightdelivery(447nm;5mWfromeachfibertip).Micewerethentested inan244
extinctionretrievaltestthefollowingdayinwhichtheyweresubjectedtotwentyCSpresentations,but245
no light was delivered (Fig. 6b, right). During this test, stGtACR2 mice showed higher freezing rates246
duringCSpresentation (ctrl vs. stGtACR2: ScheirerRayHare testH = 4.30, P = 3.8*10-2), but freezing247
levels were indistinguishable from control mice during the inter-tone intervals (ctrl vs. stGtACR2:248
ScheirerRayHaretestH=3.6*10-2,P=0.85),indicatingthatfearextinctionwaspreventedbystGtACR2249
mediatedBLA inhibitionduringCS presentation. Sierra-Mercado and colleagues 41, previously showed250
that inhibition of the BLA by muscimol injection prior to fear extinction interfered with extinction251
learning. Our results extend these findings, demonstrating that temporally-precise inhibition of BLA252
activityonlyduringCSpresentationusingstGtACR2caninterferewithextinctionlearning.Insummary,253
ourexperiments indicate that stGtACR2 isapowerful inhibitoryoptogenetic tool, allowing temporally254
precisesilencingofneuronalpopulationsinvivo.255
Discussion256
We took amembrane targeting approach to allow the utilization of the high-conductanceGuillardia257
thetaanion-conductingchannelrhodopsins16asanoptogenetictoolinmammalianneurons.Whilethese258
naturally-occurringchannelrhodopsinsshowedgreatpromiseduetotheirhighlyefficientphotocurrents259
and light sensitivity 16, andhaveproveneffective in silencingdrosophila and zebrafish neurons 19,20,21,260
they have seen little use in mammalian neuroscience applications. This was mainly due to poor261
membranetargetingandtocomplexeffectsonaxonalexcitability13,24.Ourfindingsindicatedthateven262
in its non-targeted form, GtACR2 can efficiently silence neurons in the medial prefrontal cortex of263
behaving mice. These results were consistent with the high photocurrent amplitudes recorded in264
neuronsexpressingGtACR2,comparedwithcellsexpressingtheengineeredACRs iC++andiChloC17,15.265
Given the high single channel conductance, favorable photon-ion stoichiometry, and high light266
sensitivity, the light power density for neuronal inhibition with GtACR2 is at least one order of267
magnitude lowerthanthatofothersilencingopsins8,42.However,despite itsapparenthighefficacy,a268
peer-reviewed) is the author/funder. All rights reserved. No reuse allowed without permission. The copyright holder for this preprint (which was not. http://dx.doi.org/10.1101/225847doi: bioRxiv preprint first posted online Dec. 8, 2017;

significant portion of the protein seemed to reside in intracellular compartments, where it cannot269
contribute to functional photocurrents. Furthermore, activationofGtACR2 inour recordingswas also270
associatedwithantidromicspikingatlightonsetwhenilluminatingboththeproximalanddistalaxons.271
WehavepreviouslyobservedGtACR2-mediatedtriggeringofsynapticrelease inthalamocorticalaxons27213, consistentwith recent reports of antidromic spiking in layer 2/3 pyramidal neurons 24 in the slice273
preparation. In this study,weobservedGtACR2-mediatedantidromic spiking in culturedhippocampal274
neurons,incortico-corticalneuronsintheacutesliceandincortico-striatalaxonsofbehavingmice.Our275
findingsindicatethataxonalexcitationbyachlorideconductanceisageneralphenomenon,andcould276
reflectadepolarizedreversalpotentialforchlorideintheaxonalcompartment.Whilesucheffectshave277
been previously reported, for example in hippocampalmossy fibers 43,44, cerebellar 45 and brain-stem278
axons 46,47, systematic evaluation of the phenomenon has been previously restricted to axons that279
naturallyexpressGABA-Areceptors.280
To determine whether elevated chloride concentration in the axon could indeed lead to GtACR2-281
mediated axonal excitation,we co-expressed the KCC2 transporterwithGtACR2 in cultured neurons.282
The endogenous KCC2 transporter, which is expressed in mature neurons and is known to be283
responsible for extruding chloride from the somatodendritic compartment 48, is known to be absent284
from theaxon 31,32,33, potentially permitting ahigher chloride concentration in this compartment.Our285
findingthatoverexpressionofKCC2resultedinasignificantdecreaseinlight-inducedantidromicspiking286
indicatesthatACR-mediatedantidromicspikingcouldindeedbetheresultofasmallerchloridegradient287
in the axon, even in adult neurons. While this antidromic spiking phenotype would probably not288
interferewithlong-terminhibitionexperiments(minutesandupward),itmightbeaconfoundingfactor289
whentemporally-precise (millisecond-scale) inhibition is required.FutureworkcouldcombineGtACR2290
stimulation with red-shifted chloride indicators to directly examine changes in chloride levels in the291
axonalcompartmentduringACR-mediatedchlorideconductance.292
Most importantly,ourstudydemonstrates that thesoma-targetedvariantsofGtACR2show improved293
membraneexpressioninthesomatodendriticcompartment,andoffersuperioranionphotocurrentsfor294
high-efficiency optogenetic silencing of neurons in the mammalian brain. Current optogenetic295
experimentsoften involveasparsely labeledpopulationofneuronsthataredistributedacrossa large296
brain tissue volume 49,50. Efficient silencing of such widely-distributed neuronal populations require297
continuous activity of the inhibitory optogenetic tool 51, placing considerable constraints related to298
tissueheatingandphotodamage52,9.OurdataindicatethatstGtACR2canprovideaneffectivemeansof299
peer-reviewed) is the author/funder. All rights reserved. No reuse allowed without permission. The copyright holder for this preprint (which was not. http://dx.doi.org/10.1101/225847doi: bioRxiv preprint first posted online Dec. 8, 2017;

performingsuchchallengingexperiments,dueto its intrinsicallyhighconductance,which increases its300
effective light sensitivity in expressing neurons. With increasing distance from the fiber tip, the301
wavelength dependent transmittance becomes increasingly relevant. For instance, multiplying the302
actionspectraofGtACR1andGtACR216with theanalyticallymodeled light transmittancecurve 23 for303
braintissue(SupplementaryFig.1)revealedthatexcitationofGtACR2with480nmorofGtACR1with304
510nmwouldprovideoptimal light-mediatedsilencingat500µmdistancefromtheopticfibertip. In305
experiments that require optogenetic manipulation of functionally- but not anatomically-segregated306
neuronalpopulations,stGtACR2mightbecombinedwithred-shiftedtoolssuchasC1V153,Chrimson54307
orReaChR55.Red-shiftedcalciumsensors56,57couldalsobeusedincombinationwithstGtACR2dueto308
itsminimalresponsivityatwavelengthsabove560nm.Notably,bothstGtACR1andstGtACR2arealso309
highly advantageous for multiphoton single-cell silencing experiments 58,59 owing to their somatic310
restriction22andhigh-amplitudephotocurrents.311
In summary,we have demonstrated thatmembrane targeting and somatodendritic restriction of the312
naturally-occurring anion-conducting GtACR2 address two independent constraints of this313
channelrhodopsin, greatly improving photocurrents and minimizing axonally-generated antidromic314
action potentials. We were able to achieve high-efficiency neuronal silencing with the optimized315
stGtACR2 and demonstrated its efficacy for temporally-precise inhibition of amygdala activity during316
extinctionlearning.317
Acknowledgments318
We thank M. Segal (Weizmann Institute of Science) for reagents. We thank R. Zwang for help with319cloning.We thank theYizhar laboratorymembers forcommentson themanuscript.Weacknowledge320support(toO.Y.)fromtheHumanFrontierScienceProgram,aEuropeanResearchCouncilstartinggrant321(ERC-2013-StG OptoNeuromod 337637), the Adelis Foundation, the Lord Sieff of BrimptonMemorial322Fund and the Candice Appleton Family Trust. O.Y. is supported by the Gertrude and Philip Nollman323CareerDevelopmentChair.324
AuthorContributions325
M.M.andO.Y.designedthestudywithinputfromI.L.;M.M.designedtheconstructs,performedinvitro326electrophysiology and imaging experiments. K.CKM. performed in vitro neuronal recordings. L.G.327performedandanalyzed invivo electrophysiology recordings.P.P.performedandanalyzedbehavioral328experimentsundertheguidanceofY.P.andM.M.;S.O.performedhistologyandimagingonbehavioral329mice. R.L. prepared neuronal cultures and viral vectors.M.M. andO.Y. analyzed and interpreted the330resultsandwrotethemanuscript.331
peer-reviewed) is the author/funder. All rights reserved. No reuse allowed without permission. The copyright holder for this preprint (which was not. http://dx.doi.org/10.1101/225847doi: bioRxiv preprint first posted online Dec. 8, 2017;

References332
1.Wurtz,R.H.,Usingperturbationstoidentifythebraincircuitsunderlyingactivevision.Phil.Trans.R.Soc.B370,20140205(2015).
2.Deisseroth,K.,Optogenetics.Naturemethods8,26-29(2011).
3.Han,X.&Boyden,E.S.,Multiple-coloropticalactivation,silencing,anddesynchronizationofneuralactivity,withsingle-spiketemporalresolution.PloSone2,e299(2007).
4.Zhang,F.etal.,Multimodalfastopticalinterrogationofneuralcircuitry.Nature446,633-639(2007).
5.Chow,B.Y.etal.,High-performancegeneticallytargetableopticalneuralsilencingbylight-drivenprotonpumps.Nature463,98-102(2010).
6.Chuong,A.S.etal.,Noninvasiveopticalinhibitionwithared-shiftedmicrobialrhodopsin.Natureneuroscience17,1123-1129(2014).
7.Inoue,K.etal.,Alight-drivensodiumionpumpinmarinebacteria.Naturecommunications4,1678(2013).
8.Mattis,J.etal.,Principlesforapplyingoptogenetictoolsderivedfromdirectcomparativeanalysisofmicrobialopsins.Naturemethods9(2),159-172(2011).
9.Stujenske,J.M.,Spellman,T.&Gordon,J.A.,ModelingtheSpatiotemporalDynamicsofLightandHeatPropagationforInVivoOptogenetics.Cellreports12(3),525-534(2015).
10.Arias-Gil,G.,Ohl,F.W.,Takagaki,K.&Lippert,M.T.,Measurement,modeling,andpredictionoftemperatureriseduetooptogeneticbrainstimulation.Neurophotonics3(4),045007(2016).
11.Cardin,J.A.etal.,Targetedoptogeneticstimulationandrecordingofneuronsinvivousingcell-type-specificexpressionofChannelrhodopsin-2.Natureprotocols5,247-254(2010).
12.Groma,G.I.&Dancshazy,Z.,HowManyMFormsarethereintheBacteriorhodopsinPhotocycle?Biophysicaljournal50(2),357-366(1986).
13.Mahn,M.,Prigge,M.,Ron,S.,Levy,R.&Yizhar,O.,Biophysicalconstraintsofoptogeneticinhibitionatpresynapticterminals.Natureneuroscience19(4),554-556(2016).
14.Raimondo,J.V.,Kay,L.,Ellender,T.J.&Akerman,C.J.,Optogeneticsilencingstrategiesdifferintheireffectsoninhibitorysynaptictransmission.Natureneuroscience15,1102-1104(2012).
15.Wietek,J.etal.,Animprovedchloride-conductingchannelrhodopsinforlight-inducedinhibitionofneuronalactivityinvivo.Scientificreports5,14807(2015).
16.Govorunova,E.G.,Sineshchekov,O.A.,Janz,R.,Liu,X.&Spudich,J.L.,Naturallight-gatedanionchannels:Afamilyofmicrobialrhodopsinsforadvancedoptogenetics.Science349,647-650(2015).
peer-reviewed) is the author/funder. All rights reserved. No reuse allowed without permission. The copyright holder for this preprint (which was not. http://dx.doi.org/10.1101/225847doi: bioRxiv preprint first posted online Dec. 8, 2017;

17.Berndt,A.etal.,Structuralfoundationsofoptogenetics:Determinantsofchannelrhodopsinionselectivity.ProceedingsoftheNationalAcademyofSciences113,822-829(2016).
18.Sineshchekov,O.A.,Govorunova,E.G.,Li,H.&Spudich,J.L.,Gatingmechanismsofanaturalanionchannelrhodopsin.ProceedingsoftheNationalAcademyofSciences112,14236-14241(2015).
19.Mohammad,F.etal.,Optogeneticinhibitionofbehaviorwithanionchannelrhodopsins.Naturemethods14(3),271-274(2017).
20.Mauss,A.S.,Busch,C.&Borst,A.,OptogeneticNeuronalSilencinginDrosophiladuringVisualProcessing.ScientificReports7,13823(2017).
21.Mohamed,G.A.etal.,Opticalinhibitionoflarvalzebrafishbehaviourwithanionchannelrhodopsins.BMCbiology15,103(2017).
22.Baker,C.A.,Elyada,Y.M.,Parra,A.&Bolton,M.M.,Cellularresolutioncircuitmappingwithtemporal-focusedexcitationofsoma-targetedchannelrhodopsin.eLife5(2016).
23.Yona,G.,Meitav,N.,Kahn,I.&Shoham,S.,Realisticnumericalandanalyticalmodelingoflightscatteringinbraintissueforoptogeneticapplications.eneuro3,ENEURO--0059(2016).
24.Malyshev,A.Y.etal.,ChlorideconductinglightactivatedchannelGtACR2canproducebothcessationoffiringandgenerationofactionpotentialsincorticalneuronsinresponsetolight.Neuroscienceletters640,76-80(2017).
25.Ganguly,K.,Schinder,A.F.,Wong,S.T.&Poo,M.-m.,GABAitselfpromotesthedevelopmentalswitchofneuronalGABAergicresponsesfromexcitationtoinhibition.Cell105,521-532(2001).
26.Payne,J.A.,Stevenson,T.J.&Donaldson,L.F.,MolecularcharacterizationofaputativeK-ClcotransporterinratbrainAneuronal-specificisoform.JournalofBiologicalChemistry271,16245-16252(1996).
27.Khirug,S.etal.,DistinctpropertiesoffunctionalKCC2expressioninimmaturemousehippocampalneuronsincultureandinacuteslices.EuropeanJournalofNeuroscience21,899-904(2005).
28.Kelsch,W.etal.,Insulin-likegrowthfactor1andacytosolictyrosinekinaseactivatechlorideoutwardtransportduringmaturationofhippocampalneurons.TheJournalofneuroscience:theofficialjournaloftheSocietyforNeuroscience21(21),8339-8347(2001).
29.Gabbott,P.L.A.,Warner,T.A.,Jays,P.R.L.,Salway,P.&Busby,S.J.,Prefrontalcortexintherat:Projectionstosubcorticalautonomic,motor,andlimbiccenters.TheJournalofComparativeNeurology492,145-177(2005).
30.Zhu,L.,Lovinger,D.&Delpire,E.,CorticalneuronslackingKCC2expressionshowimpairedregulationofintracellularchloride.Journalofneurophysiology93(3),1557-1568(2005).
31.Hübner,C.A.etal.,DisruptionofKCC2revealsanessentialroleofK-Clcotransportalreadyinearly
peer-reviewed) is the author/funder. All rights reserved. No reuse allowed without permission. The copyright holder for this preprint (which was not. http://dx.doi.org/10.1101/225847doi: bioRxiv preprint first posted online Dec. 8, 2017;

synapticinhibition.Neuron30(2),515-524(2001).
32.Szabadics,J.etal.,ExcitatoryeffectofGABAergicaxo-axoniccellsincorticalmicrocircuits.Science(NewYork,N.Y.)311(5758),233-235(2006).
33.Báldi,R.,Varga,C.&Tamás,G.,DifferentialdistributionofKCC2alongtheaxo-somato-dendriticaxisofhippocampalprincipalcells.TheEuropeanjournalofneuroscience32(8),1319-1325(2010).
34.Gradinaru,V.,Thompson,K.R.&Deisseroth,K.,eNpHR:aNatronomonashalorhodopsinenhancedforoptogeneticapplications.Braincellbiology36(1-4),129-139(2008).
35.Gradinaru,V.etal.,Molecularandcellularapproachesfordiversifyingandextendingoptogenetics.Cell141(1),154-165(2010).
36.Trimmer,J.S.,ImmunologicalidentificationandcharacterizationofadelayedrectifierK+channelpolypeptideinratbrain.ProceedingsoftheNationalAcademyofSciencesoftheUnitedStatesofAmerica88(23),10764-10768(1991).
37.Lim,S.T.,Antonucci,D.E.,Scannevin,R.H.&Trimmer,J.S.,AnoveltargetingsignalforproximalclusteringoftheKv2.1K+channelinhippocampalneurons.Neuron25(2),385-397(2000).
38.Wu,C.,Ivanova,E.,Zhang,Y.&Pan,Z.-H.,rAAV-mediatedsubcellulartargetingofoptogenetictoolsinretinalganglioncellsinvivo.PloSone8(6),e66332(2013).
39.Rogers,S.,Wells,R.&Rechsteiner,M.,Aminoacidsequencescommontorapidlydegradedproteins:thePESThypothesis.Science(NewYork,N.Y.)234(4774),364-368(1986).
40.Tovote,P.,Fadok,J.P.&Lüthi,A.,Neuronalcircuitsforfearandanxiety.Naturereviews.Neuroscience16(6),317-331(2015).
41.Sierra-Mercado,D.,Padilla-Coreano,N.&Quirk,G.J.,Dissociablerolesofprelimbicandinfralimbiccortices,ventralhippocampus,andbasolateralamygdalaintheexpressionandextinctionofconditionedfear.Neuropsychopharmacology:officialpublicationoftheAmericanCollegeofNeuropsychopharmacology36(2),529-538(2011).
42.Wietek,J.etal.,Anion-conductingchannelrhodopsinswithtunedspectraandmodifiedkineticsengineeredforoptogeneticmanipulationofbehavior.ScientificReports7,14957(2017).
43.Alle,H.&Geiger,J.R.P.,GABAergicspill-overtransmissionontohippocampalmossyfiberboutons.JournalofNeuroscience27,942-950(2007).
44.Ruiz,A.,Campanac,E.,Scott,R.S.,Rusakov,D.A.&Kullmann,D.M.,PresynapticGABAAreceptorsenhancetransmissionandLTPinductionathippocampalmossyfibersynapses.Natureneuroscience13,431-438(2010).
45.Pugh,J.R.&Jahr,C.E.,AxonalGABAAreceptorsincreasecerebellargranulecellexcitabilityandsynapticactivity.TheJournalofneuroscience:theofficialjournaloftheSocietyforNeuroscience31(2),
peer-reviewed) is the author/funder. All rights reserved. No reuse allowed without permission. The copyright holder for this preprint (which was not. http://dx.doi.org/10.1101/225847doi: bioRxiv preprint first posted online Dec. 8, 2017;

565-574(2011).
46.Turecek,R.&Trussell,L.O.,Presynapticglycinereceptorsenhancetransmitterreleaseatamammaliancentralsynapse.Nature411(6837),587-590(2001).
47.Price,G.D.&Trussell,L.O.,Estimateofthechlorideconcentrationinacentralglutamatergicterminal:agramicidinperforated-patchstudyonthecalyxofHeld.JournalofNeuroscience26,11432-11436(2006).
48.Kaila,K.,Price,T.J.,Payne,J.A.,Puskarjov,M.&Voipio,J.,Cation-chloridecotransportersinneuronaldevelopment,plasticityanddisease.NatureReviewsNeuroscience15,637-654(2014).
49.Liu,X.etal.,Optogeneticstimulationofahippocampalengramactivatesfearmemoryrecall.Nature(2012).
50.Kim,C.K.,Adhikari,A.&Deisseroth,K.,Integrationofoptogeneticswithcomplementarymethodologiesinsystemsneuroscience.NatureReviewsNeuroscience18,222-235(2017).
51.Wiegert,J.S.,Mahn,M.,Prigge,M.,Printz,Y.&Yizhar,O.,Silencingneurons:tools,applications,andexperimentalconstraints.Neuron95,504-529(2017).
52.Lee,J.H.etal.,GlobalandlocalfMRIsignalsdrivenbyneuronsdefinedoptogeneticallybytypeandwiring.Nature465(7299),788-792(2010).
53.Yizhar,O.etal.,Neocorticalexcitation/inhibitionbalanceininformationprocessingandsocialdysfunction.Nature477(7363),171-178(2011).
54.Klapoetke,N.C.etal.,Independentopticalexcitationofdistinctneuralpopulations.Naturemethods11(3),338-346(2014).
55.Lin,J.Y.,Knutsen,P.M.,Muller,A.,Kleinfeld,D.&Tsien,R.Y.,ReaChR:ared-shiftedvariantofchannelrhodopsinenablesdeeptranscranialoptogeneticexcitation.Natureneuroscience16(10),1499-1508(2013).
56.Dana,H.etal.,Sensitiveredproteincalciumindicatorsforimagingneuralactivity.eLife5(2016).
57.Inoue,M.etal.,Rationaldesignofahigh-affinity,fast,redcalciumindicatorR-CaMP2.Naturemethods12(1),64-70(2015).
58.Prakash,R.etal.,Two-photonoptogenetictoolboxforfastinhibition,excitationandbistablemodulation.NatureMethods9,1171-1179(2012).
59.Packer,A.M.etal.,Two-photonoptogeneticsofdendriticspinesandneuralcircuits.NatureMethods9,1202-1205(2012).
60.Liu,Y.etal.,OptogenSIM:a3DMonteCarlosimulationplatformforlightdeliverydesigninoptogenetics.Biomedicalopticsexpress6(12),4859-4870(2015).
peer-reviewed) is the author/funder. All rights reserved. No reuse allowed without permission. The copyright holder for this preprint (which was not. http://dx.doi.org/10.1101/225847doi: bioRxiv preprint first posted online Dec. 8, 2017;

61.Chugh,B.P.etal.,Measurementofcerebralbloodvolumeinmousebrainregionsusingmicro-computedtomography.NeuroImage47(4),1312-1318(2009).
62.Grimm,D.,Kay,M.A.&Kleinschmidt,J.A.,Helpervirus-free,opticallycontrollable,andtwo-plasmid-basedproductionofadeno-associatedvirusvectorsofserotypes1to6.Moleculartherapy:thejournaloftheAmericanSocietyofGeneTherapy7(6),839-850(2003).
63.Graham,F.L.&Eb,A.J.,Anewtechniquefortheassayofinfectivityofhumanadenovirus5DNA.Virology52(2),456-467(1973).
64.Anikeeva,P.etal.,Optetrode:amultichannelreadoutforoptogeneticcontrolinfreelymovingmice.Natureneuroscience15(1),163-170(2011).
65.Grünwald,D.,Shenoy,S.M.,Burke,S.&Singer,R.H.,Calibratingexcitationlightfluxesforquantitativelightmicroscopyincellbiology.Natureprotocols3(11),1809-1814(2008).
66.Pugh,J.R.&Jahr,C.E.,ActivationofaxonalreceptorsbyGABAspilloverincreasessomaticfiring.TheJournalofneuroscience:theofficialjournaloftheSocietyforNeuroscience33(43),16924-16929(2013).
333
334
peer-reviewed) is the author/funder. All rights reserved. No reuse allowed without permission. The copyright holder for this preprint (which was not. http://dx.doi.org/10.1101/225847doi: bioRxiv preprint first posted online Dec. 8, 2017;
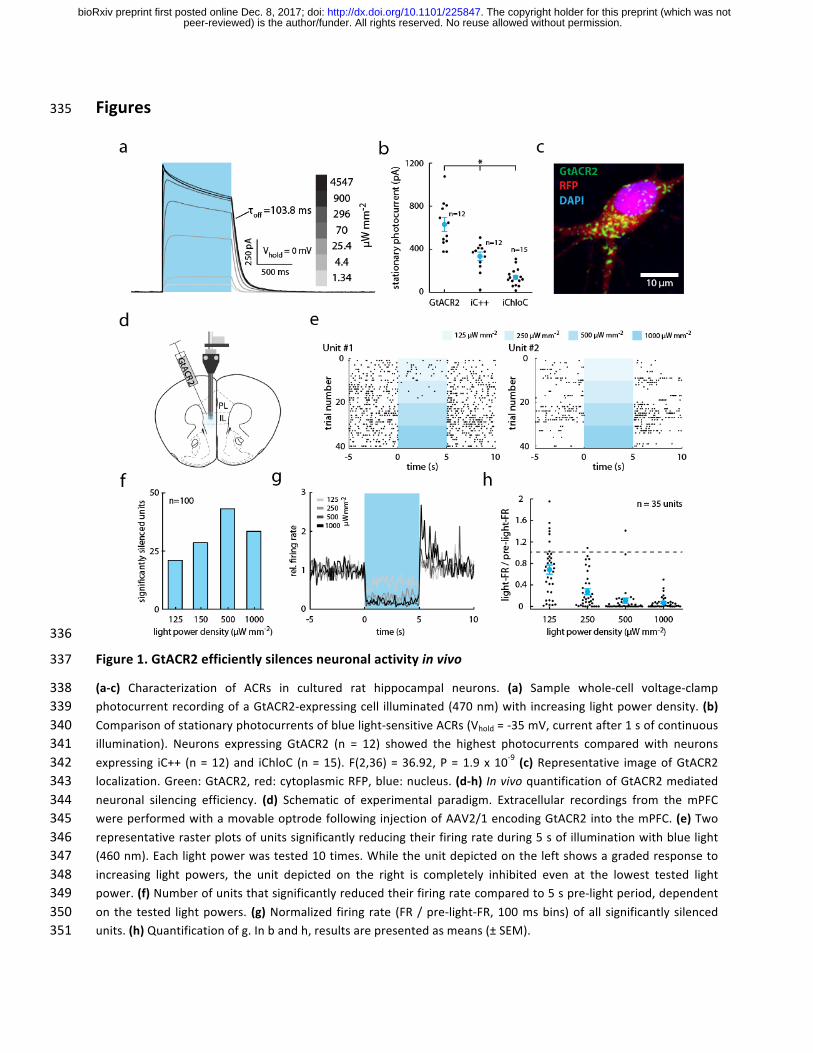
Figures 335
336
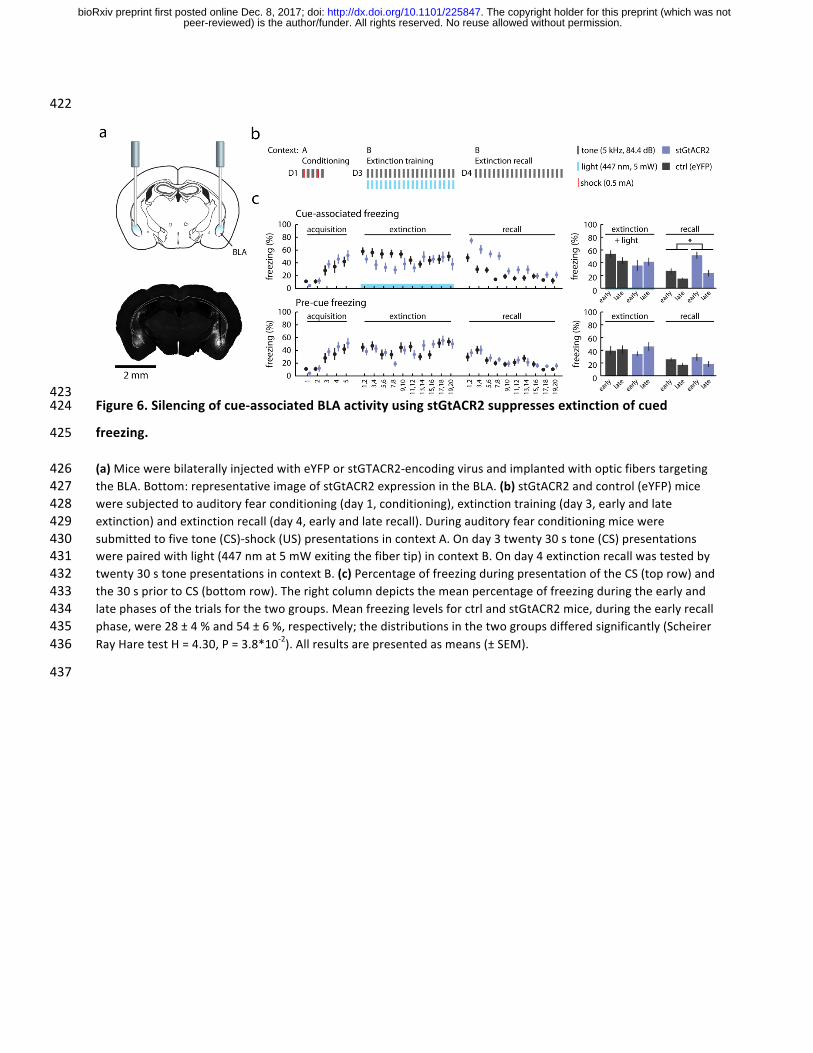
Figure1.GtACR2efficientlysilencesneuronalactivityinvivo337
(a-c) Characterization of ACRs in cultured rat hippocampal neurons. (a) Sample whole-cell voltage-clamp338photocurrentrecordingofaGtACR2-expressingcell illuminated(470nm)with increasinglightpowerdensity.(b)339Comparisonofstationaryphotocurrentsofbluelight-sensitiveACRs(Vhold=-35mV,currentafter1sofcontinuous340illumination). Neurons expressing GtACR2 (n = 12) showed the highest photocurrents compared with neurons341expressing iC++ (n=12)and iChloC (n=15). F(2,36)=36.92,P=1.9x10-9 (c)Representative imageofGtACR2342localization.Green:GtACR2,red:cytoplasmicRFP,blue:nucleus.(d-h) InvivoquantificationofGtACR2mediated343neuronal silencing efficiency. (d) Schematic of experimental paradigm. Extracellular recordings from the mPFC344wereperformedwithamovableoptrodefollowinginjectionofAAV2/1encodingGtACR2intothemPFC.(e)Two345representativerasterplotsofunitssignificantlyreducingtheirfiringrateduring5sofilluminationwithbluelight346(460nm).Eachlightpowerwastested10times.Whiletheunitdepictedontheleftshowsagradedresponseto347increasing light powers, the unit depicted on the right is completely inhibited even at the lowest tested light348power.(f)Numberofunitsthatsignificantlyreducedtheirfiringratecomparedto5spre-lightperiod,dependent349on the tested lightpowers. (g)Normalized firing rate (FR/pre-light-FR,100msbins)ofall significantly silenced350units.(h)Quantificationofg.Inbandh,resultsarepresentedasmeans(±SEM).351
peer-reviewed) is the author/funder. All rights reserved. No reuse allowed without permission. The copyright holder for this preprint (which was not. http://dx.doi.org/10.1101/225847doi: bioRxiv preprint first posted online Dec. 8, 2017;

352
Figure2.ActivationofGtACR2intheaxonalcompartmentinducesactionpotentialsinvitroandin353vivo354
(a-b)Characterizationoflight-evokedspikinginACR-expressingculturedhippocampalneurons(470nmat4.5mW355mm-2). (a) Representative whole-cell current-clamp recording of a GtACR2-expressing cell silenced by light356application. Inset: strongly attenuated spike occurring shortly after light onset. (b) Representative whole-cell357voltage-clamprecordingofescapedactionpotentialsinresponseto1mslightpulses.Piechartsdepictthenumber358of neuronswith induced spikes for the three tested light-gated chloride channels:GtACR2, iC++ and iChloC. (c)359Illuminationofdistalneurites inducesspiking inculturedneurons.Schematicdepicting theoutlineofaGtACR2-360expressing neuron overlaid with the locations of laser illumination spots. Shown are whole-cell voltage-clamp361responses to spatially-restricted illumination at the indicated locations. (d-g) In vivo extracellular recording362followingGtACR2expressioninthemPFC.(d)SchematicoftheimplantationallowingforilluminationoftheNAc,a363downstreamtargetofthemPFC,whilerecordinginthemPFC.(e)SingleunitsrecordedinthemPFC,showingrapid364light-evokedresponsesduringa20mstime-windowstartingwitha5mslightpulse(1mWmm-2
,correspondingto36528.8mWatthefibertip).Unitsarearrangedfromtoptobottomaccordingtotheirmeanfirstspikelatencyacross36620trials.Piechartsdepictthenumberofneuronswithsignificantlyincreasedspikerates.(f)mPFCunitsshowing367significantlyincreasedfiringratesinresponsetoilluminationoftheNAc.Unitsaresortedbymeanspikelatency.368Lightpoweratthefibertip:28.8mW.Piechartasin(e).(g)Percentofunitswithincreasedfiringrateinresponse369to5mslightpulsesofincreasinglightpower.370
371
peer-reviewed) is the author/funder. All rights reserved. No reuse allowed without permission. The copyright holder for this preprint (which was not. http://dx.doi.org/10.1101/225847doi: bioRxiv preprint first posted online Dec. 8, 2017;
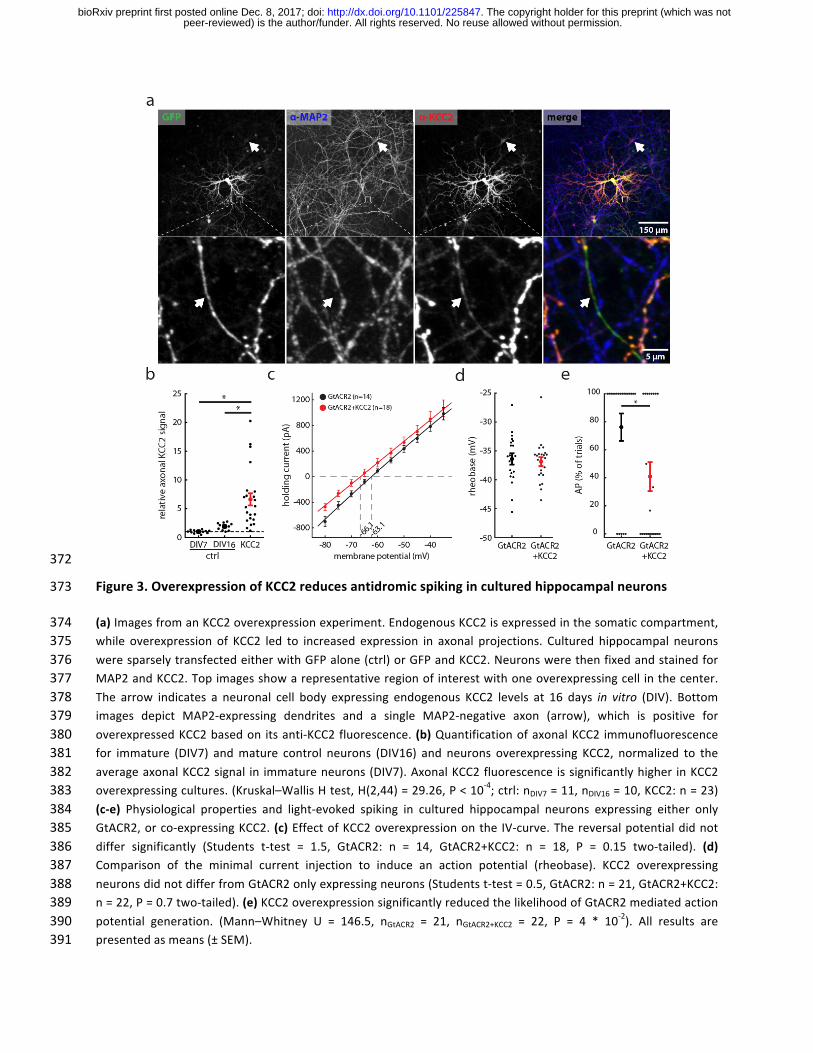
372
Figure3.OverexpressionofKCC2reducesantidromicspikinginculturedhippocampalneurons373
(a)ImagesfromanKCC2overexpressionexperiment.EndogenousKCC2isexpressedinthesomaticcompartment,374while overexpression of KCC2 led to increased expression in axonal projections. Cultured hippocampal neurons375weresparselytransfectedeitherwithGFPalone(ctrl)orGFPandKCC2.Neuronswerethenfixedandstainedfor376MAP2andKCC2.Topimagesshowarepresentativeregionofinterestwithoneoverexpressingcell inthecenter.377The arrow indicates a neuronal cell body expressing endogenous KCC2 levels at 16 days in vitro (DIV). Bottom378images depict MAP2-expressing dendrites and a single MAP2-negative axon (arrow), which is positive for379overexpressedKCC2basedon itsanti-KCC2fluorescence. (b)QuantificationofaxonalKCC2 immunofluorescence380for immature (DIV7) andmature control neurons (DIV16) and neurons overexpressing KCC2, normalized to the381averageaxonalKCC2signal in immatureneurons(DIV7).AxonalKCC2fluorescence issignificantlyhigher inKCC2382overexpressingcultures.(Kruskal–WallisHtest,H(2,44)=29.26,P<10-4;ctrl:nDIV7=11,nDIV16=10,KCC2:n=23)383(c-e) Physiological properties and light-evoked spiking in cultured hippocampal neurons expressing either only384GtACR2,orco-expressingKCC2.(c)EffectofKCC2overexpressiononthe IV-curve.Thereversalpotentialdidnot385differ significantly (Students t-test = 1.5, GtACR2: n = 14, GtACR2+KCC2: n = 18, P = 0.15 two-tailed). (d)386Comparison of the minimal current injection to induce an action potential (rheobase). KCC2 overexpressing387neuronsdidnotdifferfromGtACR2onlyexpressingneurons(Studentst-test=0.5,GtACR2:n=21,GtACR2+KCC2:388n=22,P=0.7two-tailed).(e)KCC2overexpressionsignificantlyreducedthelikelihoodofGtACR2mediatedaction389potential generation. (Mann–Whitney U = 146.5, nGtACR2 = 21, nGtACR2+KCC2 = 22, P = 4 * 10
-2). All results are390presentedasmeans(±SEM). 391
peer-reviewed) is the author/funder. All rights reserved. No reuse allowed without permission. The copyright holder for this preprint (which was not. http://dx.doi.org/10.1101/225847doi: bioRxiv preprint first posted online Dec. 8, 2017;
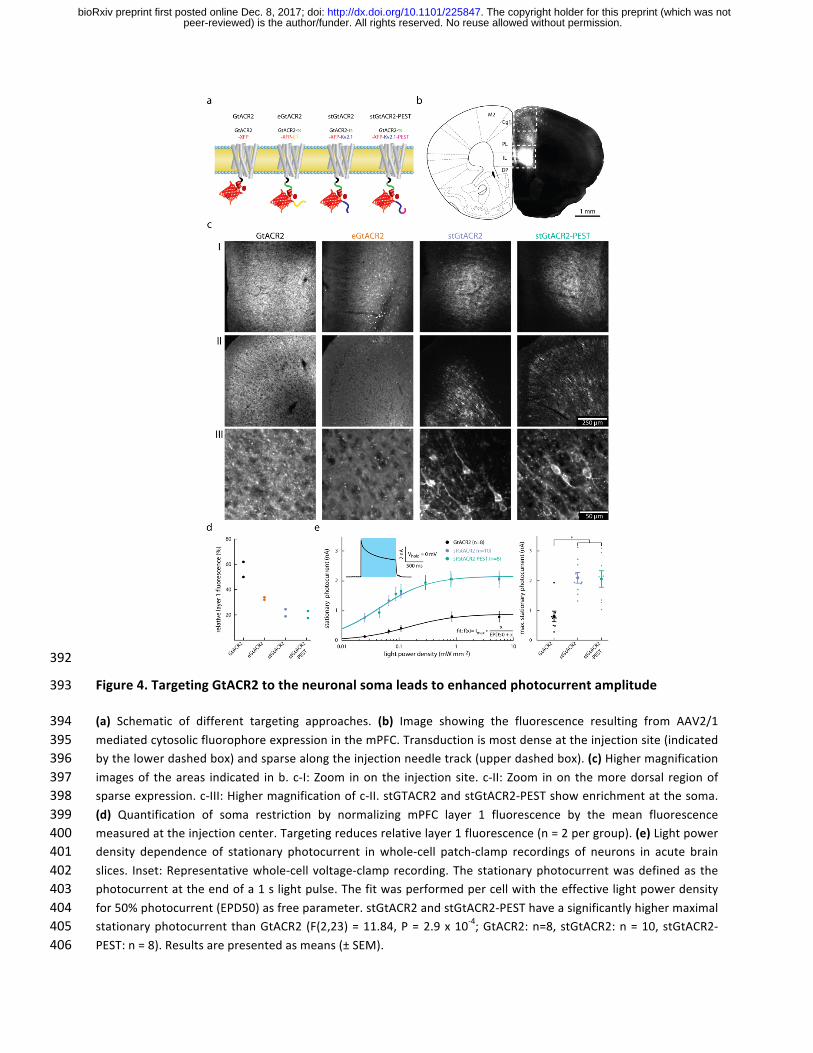
392
Figure4.TargetingGtACR2totheneuronalsomaleadstoenhancedphotocurrentamplitude393
(a) Schematic of different targeting approaches. (b) Image showing the fluorescence resulting from AAV2/1394mediatedcytosolicfluorophoreexpressioninthemPFC.Transductionismostdenseattheinjectionsite(indicated395bythelowerdashedbox)andsparsealongtheinjectionneedletrack(upperdashedbox).(c)Highermagnification396imagesof theareas indicated inb. c-I:Zoom inon the injectionsite. c-II:Zoom inon themoredorsal regionof397sparseexpression.c-III:Highermagnificationofc-II.stGTACR2andstGtACR2-PESTshowenrichmentatthesoma.398(d) Quantification of soma restriction by normalizing mPFC layer 1 fluorescence by the mean fluorescence399measuredattheinjectioncenter.Targetingreducesrelativelayer1fluorescence(n=2pergroup).(e)Lightpower400density dependence of stationary photocurrent inwhole-cell patch-clamp recordings of neurons in acute brain401slices. Inset:Representativewhole-cellvoltage-clamprecording.Thestationaryphotocurrentwasdefinedas the402photocurrentattheendofa1slightpulse.Thefitwasperformedpercellwiththeeffectivelightpowerdensity403for50%photocurrent(EPD50)asfreeparameter.stGtACR2andstGtACR2-PESThaveasignificantlyhighermaximal404stationaryphotocurrent thanGtACR2 (F(2,23)=11.84,P=2.9x10-4;GtACR2:n=8,stGtACR2:n=10,stGtACR2-405PEST:n=8).Resultsarepresentedasmeans(±SEM).406
peer-reviewed) is the author/funder. All rights reserved. No reuse allowed without permission. The copyright holder for this preprint (which was not. http://dx.doi.org/10.1101/225847doi: bioRxiv preprint first posted online Dec. 8, 2017;
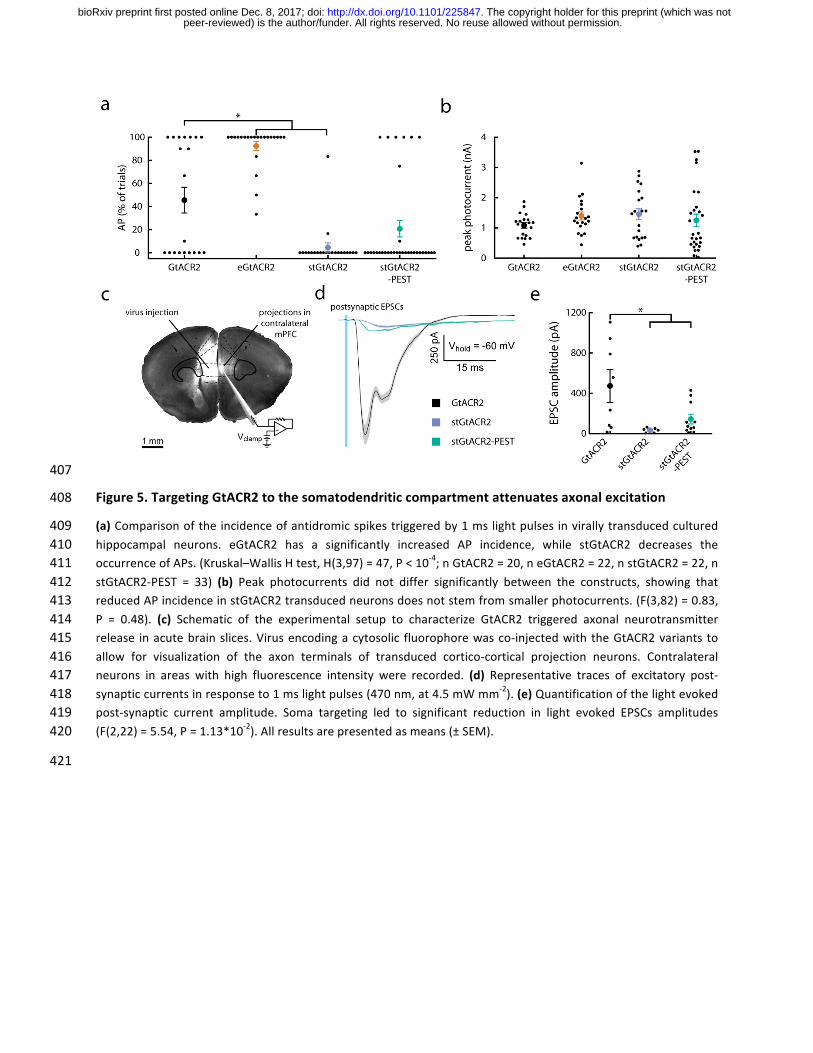
407
Figure5.TargetingGtACR2tothesomatodendriticcompartmentattenuatesaxonalexcitation408
(a)Comparisonoftheincidenceofantidromicspikestriggeredby1mslightpulsesinvirallytransducedcultured409hippocampal neurons. eGtACR2 has a significantly increased AP incidence, while stGtACR2 decreases the410occurrenceofAPs.(Kruskal–WallisHtest,H(3,97)=47,P<10-4;nGtACR2=20,neGtACR2=22,nstGtACR2=22,n411stGtACR2-PEST = 33) (b) Peak photocurrents did not differ significantly between the constructs, showing that412reducedAPincidenceinstGtACR2transducedneuronsdoesnotstemfromsmallerphotocurrents.(F(3,82)=0.83,413P = 0.48). (c) Schematic of the experimental setup to characterize GtACR2 triggered axonal neurotransmitter414release inacutebrainslices.Virusencodingacytosolic fluorophorewasco-injectedwiththeGtACR2variants to415allow for visualization of the axon terminals of transduced cortico-cortical projection neurons. Contralateral416neurons in areas with high fluorescence intensity were recorded. (d) Representative traces of excitatory post-417synapticcurrentsinresponseto1mslightpulses(470nm,at4.5mWmm-2).(e)Quantificationofthelightevoked418post-synaptic current amplitude. Soma targeting led to significant reduction in light evoked EPSCs amplitudes419(F(2,22)=5.54,P=1.13*10-2).Allresultsarepresentedasmeans(±SEM).420
421
peer-reviewed) is the author/funder. All rights reserved. No reuse allowed without permission. The copyright holder for this preprint (which was not. http://dx.doi.org/10.1101/225847doi: bioRxiv preprint first posted online Dec. 8, 2017;
422
423Figure6.Silencingofcue-associatedBLAactivityusingstGtACR2suppressesextinctionofcued424
freezing.425
(a)MicewerebilaterallyinjectedwitheYFPorstGTACR2-encodingvirusandimplantedwithopticfiberstargeting426theBLA.Bottom:representativeimageofstGtACR2expressionintheBLA.(b)stGtACR2andcontrol(eYFP)mice427weresubjectedtoauditoryfearconditioning(day1,conditioning),extinctiontraining(day3,earlyandlate428extinction)andextinctionrecall(day4,earlyandlaterecall).Duringauditoryfearconditioningmicewere429submittedtofivetone(CS)-shock(US)presentationsincontextA.Onday3twenty30stone(CS)presentations430werepairedwithlight(447nmat5mWexitingthefibertip)incontextB.Onday4extinctionrecallwastestedby431twenty30stonepresentationsincontextB.(c)PercentageoffreezingduringpresentationoftheCS(toprow)and432the30spriortoCS(bottomrow).Therightcolumndepictsthemeanpercentageoffreezingduringtheearlyand433latephasesofthetrialsforthetwogroups.MeanfreezinglevelsforctrlandstGtACR2mice,duringtheearlyrecall434phase,were28±4%and54±6%,respectively;thedistributionsinthetwogroupsdifferedsignificantly(Scheirer435RayHaretestH=4.30,P=3.8*10-2).Allresultsarepresentedasmeans(±SEM).436
437
peer-reviewed) is the author/funder. All rights reserved. No reuse allowed without permission. The copyright holder for this preprint (which was not. http://dx.doi.org/10.1101/225847doi: bioRxiv preprint first posted online Dec. 8, 2017;
438
439
440
441
442
443
444
445
446
447
448
449
450
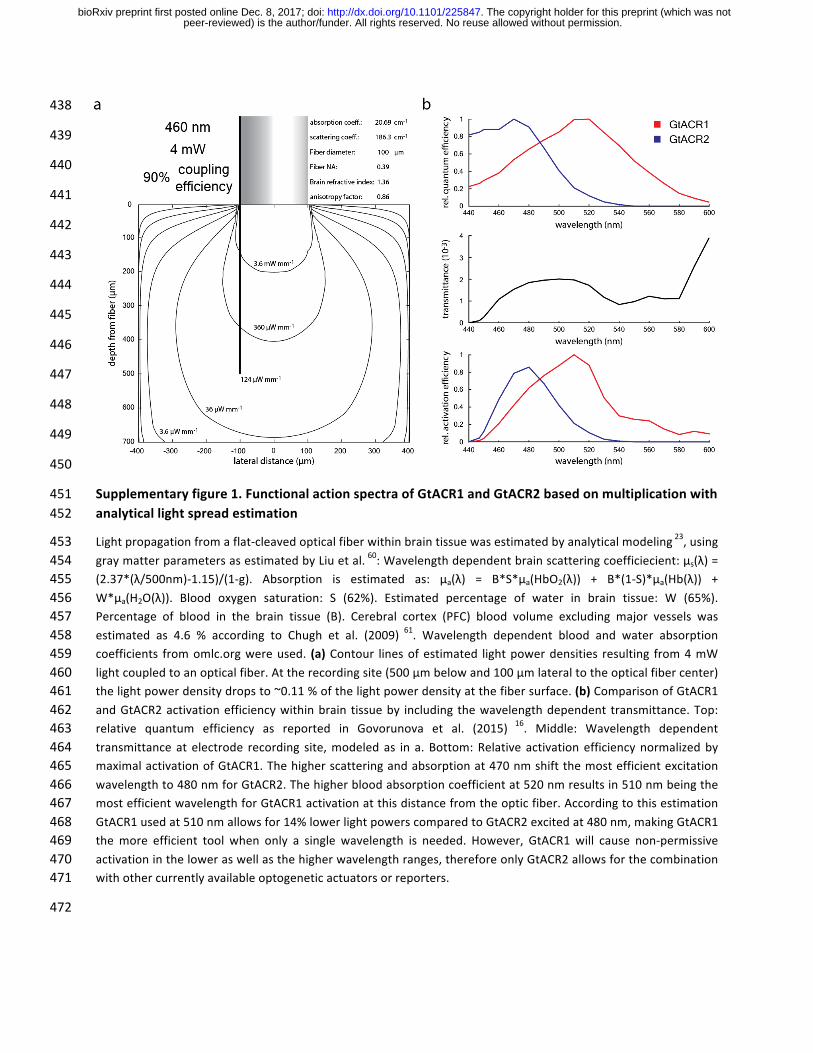
Supplementaryfigure1.FunctionalactionspectraofGtACR1andGtACR2basedonmultiplicationwith451analyticallightspreadestimation452
Lightpropagationfromaflat-cleavedopticalfiberwithinbraintissuewasestimatedbyanalyticalmodeling23,using453graymatterparametersasestimatedbyLiuetal.60:Wavelengthdependentbrainscatteringcoefficiecient:µs(λ)=454(2.37*(λ/500nm)-1.15)/(1-g). Absorption is estimated as: µa(λ) = B*S*µa(HbO2(λ)) + B*(1-S)*µa(Hb(λ)) +455W*µa(H2O(λ)). Blood oxygen saturation: S (62%). Estimated percentage of water in brain tissue: W (65%).456Percentage of blood in the brain tissue (B). Cerebral cortex (PFC) blood volume excluding major vessels was457estimated as 4.6 % according to Chugh et al. (2009) 61. Wavelength dependent blood and water absorption458coefficients fromomlc.orgwereused. (a)Contour linesofestimated lightpowerdensities resulting from4mW459lightcoupledtoanopticalfiber.Attherecordingsite(500µmbelowand100µmlateraltotheopticalfibercenter)460thelightpowerdensitydropsto~0.11%ofthelightpowerdensityatthefibersurface.(b)ComparisonofGtACR1461andGtACR2activationefficiencywithinbrain tissueby including thewavelengthdependent transmittance.Top:462relative quantum efficiency as reported in Govorunova et al. (2015) 16. Middle: Wavelength dependent463transmittanceat electrode recording site,modeledas in a. Bottom:Relative activationefficiencynormalizedby464maximalactivationofGtACR1.Thehigherscatteringandabsorptionat470nmshiftthemostefficientexcitation465wavelengthto480nmforGtACR2.Thehigherbloodabsorptioncoefficientat520nmresultsin510nmbeingthe466mostefficientwavelengthforGtACR1activationatthisdistancefromtheopticfiber.Accordingtothisestimation467GtACR1usedat510nmallowsfor14%lowerlightpowerscomparedtoGtACR2excitedat480nm,makingGtACR1468themore efficient tool when only a single wavelength is needed. However, GtACR1 will cause non-permissive469activationintheloweraswellasthehigherwavelengthranges,thereforeonlyGtACR2allowsforthecombination470withothercurrentlyavailableoptogeneticactuatorsorreporters.471
472
peer-reviewed) is the author/funder. All rights reserved. No reuse allowed without permission. The copyright holder for this preprint (which was not. http://dx.doi.org/10.1101/225847doi: bioRxiv preprint first posted online Dec. 8, 2017;
473
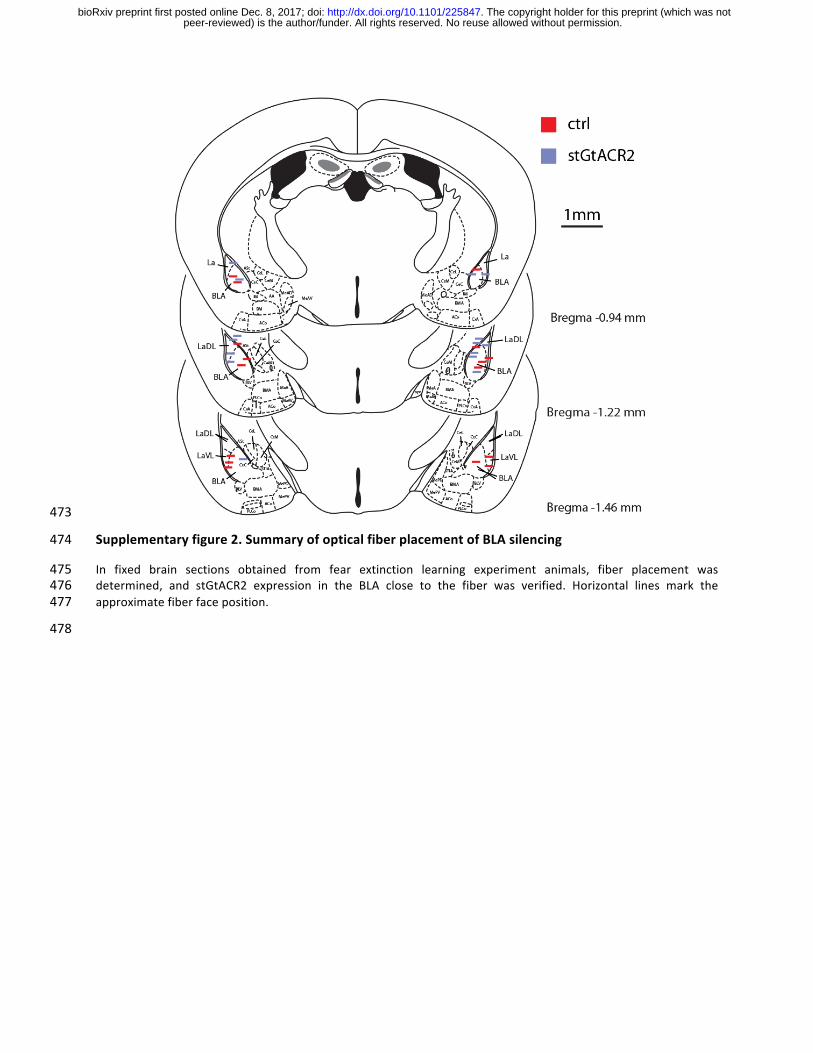
Supplementaryfigure2.SummaryofopticalfiberplacementofBLAsilencing474
In fixed brain sections obtained from fear extinction learning experiment animals, fiber placement was475determined, and stGtACR2 expression in the BLA close to the fiber was verified. Horizontal lines mark the476approximatefiberfaceposition.477
478
peer-reviewed) is the author/funder. All rights reserved. No reuse allowed without permission. The copyright holder for this preprint (which was not. http://dx.doi.org/10.1101/225847doi: bioRxiv preprint first posted online Dec. 8, 2017;

ExperimentalProcedures479
ProductionofrecombinantAAVvectors480
HEK293cellswereseededat25%-35%confluence.Thecellsweretransfected24hlaterwithplasmids481
encodingAAVrep,capandavectorplasmidfortherAAVcassetteexpressingtherelevantDNAusingthe482
PEImethod62.Cellsandmediumwereharvested72haftertransfection,pelletedbycentrifugation(300483
g), resuspended in lysis solution ([mM]: 150NaCl, 50 Tris-HCl; pH8.5withNaOH) and lysedby three484
freeze-thawcycles. The crude lysatewas treatedwith250Ubenzonase (Sigma)per 1mlof lysate at485
37°Cfor1.5htodegradegenomicandunpackagedAAVDNAbeforecentrifugationat3000gfor15min486
to pellet cell debris. The virus particles in the supernatant (crude virus)were purified using heparin-487
agarose columns, eluted with soluble heparin, washed with phosphate buffered saline (PBS) and488
concentratedbyAmiconcolumns.Viralsuspensionwasaliquotedandstoredat–80°C.Viraltiterswere489
measuredusing real-timePCR.AAVvectorsused for intracranial injectionshadgenomic titers ranging490
between8.6*1010and2*1011genomecopiespermilliliter(gc/ml).Wheredirectlycomparedvirustiters491
were matched by dilution to the lowest concentration. AAV vectors used for neuronal culture492
transductionwereadded4daysaftercellseeding.Thetiterwasmatchedtofinalmediumconcentration493
of1.1*108gc/ml.AlloftheAAVexpressionconstructsdescribedinthisstudywillbeavailablefreelyon494
Addgenetofacilitatetheutilizationofthesenewtoolsbytheneurosciencecommunity.495
Thefollowingviruseswereusedinthisstudy:496
AAV2/1.hSyn1.GtACR2-eGFP.WPRE,AAV2/1.CamKIIα.GtACR2-ts-Fred-Kv2.1.WPRE,497
AAV2/1.CamKIIα.GtACR2-ts-Fred-ER.WPRE,AAV2/1.CamKIIα.GtACR2-ts-Fred-Kv2.1-PEST.WPRE,498
AAV2/1.CamKIIα.TagRFP-T.WPRE,AAV2/1.CamKIIα.eYFP.WPRE,AAV2/1.CamKIIα.iC++-eYFP.WPRE,499
AAV2/1.hSyn.iChlOC-eGFP.WPRE.500
Primaryhippocampalneuronculture501
Primary cultured hippocampal neuronswere prepared frommale and female P0 Sprague-Dawley rat502
pups(Envigo).CA1andCA3wereisolated,digestedwith0.4mgml−1papain(Worthington),andplated503
ontoglass coverslipspre-coatedwith1:30Matrigel (Corning).Culturedneuronsweremaintained ina504
5% CO2 humidified incubator with Neurobasal-A medium (Invitrogen) containing 1.25% fetal bovine505
serum(FBS,BiologicalIndustries),4%B-27supplement(Gibco),2mMGlutamax(Gibco)andplatedon506
coverslips in a24-well plateat adensityof 65,000 cells perwell. To inhibit glial overgrowth,200µM507
fluorodeoxyuridine(FUDR,Sigma)wasaddedafter4daysofinvitroculture(DIV).508
peer-reviewed) is the author/funder. All rights reserved. No reuse allowed without permission. The copyright holder for this preprint (which was not. http://dx.doi.org/10.1101/225847doi: bioRxiv preprint first posted online Dec. 8, 2017;

Calciumphosphatetransfectionofculturedneurons509
Neurons were transfected using the calcium phosphate method 63. Briefly, the medium of primary510
hippocampal neurons cultured in a 24well platewas collected and replacedwith 400 µl serum-free511
MEMmedium(ThermoFisherscientific).30µltransfectionmix(2µgplasmidDNAand250µMCaCl2in512
HBSatpH7.05)wereaddedperwell.After1hincubationthecellswerewashed2timeswithMEMand513
themediumwaschangedbacktothecollectedoriginalmedium.Culturedneuronswereusedbetween514
14–17DIVforexperiments.515
Thefollowingplasmidswereusedinthisstudy:516
pAAV_hSyn1_GtACR2-eGFP_WPRE(basedonAddgene85463),pAAV_CamKIIα_mNeonGreen_WPRE,517
pAAV_CamKIIα(0.4kb)_mScarlet_WPRE,pCITF_KCC2-tdTomato(Addgene61404).518
Animals519
All experimental procedures were approved by the Institutional Animal Care and Use Committee520
(IACUC)attheWeizmannInstituteofScience.Six-week-oldC57BL/6mice(P35–45)wereobtainedfrom521
Envigo.Upto5maleorfemaleC57BL/6micewerehousedinacageinalight-dark(12h-12h)cyclewith522
foodandwateradlibitum.Micewerehousedfor6-12weeksfollowingsurgerytoallowforrecoveryand523
virusexpression.524
Stereotacticinjectionofviralvectors525
Six-week-oldC57BL/6mice(P35–45)wereinitiallyinducedwithketamine(80mgkg−1)andxylazine(10526
mg kg−1) and placed into a stereotaxic frame (David Kopf Instruments), before isoflurane anesthesia527
(~1% in O2, v/v). A craniotomy (∼1 mm in diameter) was made above the injection site. Virus528
suspensionswere slowly injected (100 nlmin–1) using a 34G beveled needle (Nanofil syringe,World529
Precision Instruments). After injection, the needlewas left in place for an additional 5min and then530
slowly withdrawn. The surgical procedure was either continued with optic fiber or optrode drive531
implantations (describedbelow),or the surgical incisionwasclosedwith tissueglueand0.05mgkg−1532
Buprenorphinewassubcutaneously injectedforpost-surgicalanalgesia. Injectionstargetingthemedial533
prefrontalcortex(mPFC)weremade1.8mmanterior,0.3mmlateraland2.53mmventraltobregma.534
Basolateralamygdala(BLA) injectioncoordinateswere1.15mmposterior,3.0mmlateraland5.0mm535
ventral to bregma. For mPFC injections, 1 µl of the indicated virus was injected. For fear extinction536
peer-reviewed) is the author/funder. All rights reserved. No reuse allowed without permission. The copyright holder for this preprint (which was not. http://dx.doi.org/10.1101/225847doi: bioRxiv preprint first posted online Dec. 8, 2017;

experiments mice were bilaterally injected with 500 nl AAV2/1.CamKIIα.stGtACR2-Fred.WPRE or537
AAV2/1.CamKIIα.eYFP.WPREwithagenomictiterintherangeof2-3x1011vpml-1.538
OpticfiberandOptrodedriveimplantation539
Forfiberopticimplantation,acraniotomy(∼1 mmindiameter)wasmadeabovetheimplantationsite540
and a ferrule-terminated optical fiber (ThorLabs) was placed at the desired coordinates using a541
stereotaxicframe(DavidKopfInstruments).ForbilateralBLAtargeting,thefibertipwasplaced1.15mm542
posterior, 3.0 mm lateral and 4.8 mm ventral to bregma. For nucleus accumbens, the fiber was543
implantedata45°anglewiththeferrulepointingposteriortoallowforoptrodedriveplacementabove544
themPFCinthesameanimals.Thefibertipwasaimedtoterminate1.42mmanterior,1mmlateraland545
5mmventraltobregma.TheopticalfiberwassecuredtotheskullusingMetabond(Parkell)anddental546
acrylic. In mice trained for fear extinction learning additional dental acrylic was applied in a second547
session under isoflurane anesthesia (~1% in O2, v/v) after fear learning (day 2). For optrode drive548
implantation,themovabledrivewasloweredtoaninitialrecordingpositionabovethePL(AP:1.8mm,549
ML:0.3mm,DV:–2.3mm).Priortothepermanentattachmentoftheoptrodetotheskull,theoptrode550
guidewasprotectedwithKwik-Kastsiliconeelastomer(WorldPrecisionInstruments)andsecuredusing551
dentalacrylic.Micewereallowedtorecoverforat least6weeksbeforeexperiments.Thelocationsof552
implantedopticalfibersandoptrodeswerevalidatedhistologicallyforallexperimentalmice.553
Acutebrainslicepreparation554
Mice were injected intraperitoneally with pentobarbital (130 mg kg−1, i.p.) and perfused with555
carbogenated (95% O2, 5% CO2) ice-cold slicing solution ([mM] 2.5 KCl, 11 glucose, 234 sucrose, 26556
NaHCO3, 1.25 NaH2PO4, 10MgSO4, 2 CaCl2; 340mOsm). After decapitation, 300 µm coronalmPFC557
sliceswereprepared in carbogenated ice-cold slicing solutionusinga vibratome (LeicaVT1200S)and558
allowed to recover for 20 min at 33°C in carbogenated high-osmolarity artificial cerebrospinal fluid559
(high-OsmACSF; [mM]3.2KCl, 11.8 glucose, 132NaCl, 27.9NaHCO3, 1.34NaH2PO4, 1.07MgCl2, 2.14560
CaCl2;320mOsm)followedby40minincubationat33°CincarbogenatedACSF([mM]3KCl,11glucose,561
123NaCl,26NaHCO3,1.25NaH2PO4,1MgCl2,2CaCl2;300mOsm).Subsequently,sliceswerekeptatRT562
incarbogenatedACSFuntiluse.TherecordingchamberwasperfusedwithcarbogenatedACSFatarate563
of2mlmin–1andmaintainedat32°C.564
Electrophysiologicalmethodsforcellcultureandacutebrainslicerecordings565
peer-reviewed) is the author/funder. All rights reserved. No reuse allowed without permission. The copyright holder for this preprint (which was not. http://dx.doi.org/10.1101/225847doi: bioRxiv preprint first posted online Dec. 8, 2017;

Whole-cellpatchclamprecordingswereperformedundervisualcontrolusingobliqueilluminationona566
two-photon laser scanningmicroscope (Ultima IV, Bruker) equipped with a 12 bit monochrome CCD567
camera (QImagingQIClick-R-F-M-12). Borosilicate glass pipettes (Sutter Instrument BF100-58-10)with568
resistancesrangingfrom3–7MΩwerepulledusingalasermicropipettepuller(SutterInstrumentModel569
P-2000).Forhippocampalneuroncultures,electrophysiologicalrecordingsfromneuronswereobtained570
inTyrode’smedium ([mM]150NaCl,4KCl,2MgCl2,2CaCl2,10D-glucose,10HEPES;320mOsm;pH571
adjustedto7.35withNaOH),AcOHTyrode’smedium([mM]125NaCl,25AcOH,4KCl,2MgCl2,2CaCl2,572
10 D-glucose, 10 HEPES; 320 mOsm; pH adjusted to 7.35 with NaOH). The recording chamber was573
perfused at 0.5 ml min–1 and maintained at 29°C. Pipettes were filled using standard intracellular574
solution([mM]135K-gluconate,4KCl,2NaCl,10HEPES,4EGTA,4MgATP,0.3NaGTP;280mOsmkg–1;575
pHadjustedto7.3withKOH)oranintracellularsolutionallowingforEPSCandIPSCrecording([mM]120576
Cs-gluconate,11CsCl,1MgCl2,1CaCl2,10HEPES,11EGTA,5QX-314;280mOsmkg–1;pHadjustedto577
7.3 with CsOH). Whole-cell voltage clamp recordings were performed using a MultiClamp 700B578
amplifier,filteredat8kHzanddigitizedat20kHzusingaDigidata1440Adigitizer(MolecularDevices).579
Invivoopticalsilencingandelectrophysiology580
Allelectrophysiologicalrecordingsinawake,freelymovingmicewereperformedusinganoptrodedrive581
consisting of an electrode bundle of 16 microwires (25 μm diameter straightened tungsten wires;582
Wiretronic Inc.) attached to an 18 pin electrical connector, concentrically arranged around an optical583
fiber inamechanicallyadjustabledrive (Anikeevaetal., 2011 64). Extracellularwaveformsignalswere584
collectedusingtheDigitalLynxintegratedhardwareandsoftwaresystem(NeuralynxInc.).Theelectrical585
signalwasfiltered(600–6,000Hz),amplifiedusingaHS-18-CNR-LEDunity-gainhead-stageamplifierand586
digitized at 32 kHz. The electrode–fiber assembly was lowered using themechanical drive to a new587
recording site at the end of each recording session, leaving at least 1.5 h before the next session to588
ensure stable recordings. Optical stimulation was applied through a ferrule-terminated optical fiber589
(ThorLabs)attachedtothepatch-chordbyazirconiasleeve(ThorLabs).ForopticalsilencingofmPFC,we590
used a blue diode laser (λ = 460 nm,OmicronNanoTechnology). Light transmission for each optrode591
drivewasmeasuredwithacalibratedpowermeter(ThorLabs)atthetipoftheopticalfiberattheendof592
theexperiment.Lightpowerwasmeasureddailybeforeexperimentsatthetipoftheopticalpatchcord.593
Neural data were sorted manually using Off-Line Spike Sorter 3.2.4 (OFSS, Plexon) and analyzed in594
Matlab(MathWorks).595
Invivooptogeneticsilencinginmiceduringextinctiontraining596
peer-reviewed) is the author/funder. All rights reserved. No reuse allowed without permission. The copyright holder for this preprint (which was not. http://dx.doi.org/10.1101/225847doi: bioRxiv preprint first posted online Dec. 8, 2017;

Mice in both the stGtACR2 and control group (eYFP expressing)were placed in the fear conditioning597
chamber(MedAssociates)incontextA.MicewerepresentedwithfivepairingsoftheCS(50mslong5598
kHz84.4dBtones,deliveredat10Hzfor30s)andUS(continuous0.5mAfootshockfor1s).EachCS599
coterminatedwiththeUS,witha60sintervalbetweenCS–USpairings.Onday3,micewereconnected600
totheopticalpatchchordandthenplacedinadifferentchamber(contextB).ContextBdifferedfrom601
contextAinthefollowingaspects:odor(A:1%Aceticacidvs.B:70%EtOH),lighting(A:IRvs.B:IR+602
white light), box size (A: small, B: large), floor texture (A: grid, B: plain),wall texture (A:metal vs. B:603
Plexiglas),andbackgroundnoise(A:nonevs.B:fan).Micewereallowed10minofhabituationandthen604
presentedwith20repetitionsoftheCS,separatedby60sintervals.TheCSwaspairedwith5mWblue605
light(447nm)administeredbilaterallyfromthefibertipinbothgroups.Totestextinctionlearning,mice606
were placed in context B on day 4 and presented with 20 repetitions of the CS, separated by 60 s607
intervals.Moviesrecordedat25framespersecondwereautomaticallyscoredforfreezingonday1and608
4 by EthoVision XT 11.5 (Noldus) and by a custom written OpenCV-Python script. The number of609
changedpixelscomparedtothelastframewasquantifiedandfilteredbyaGaussianfilterwith3frames610
standarddeviation.Whenmicewereconnectedtoopticalpatchcords,onlychangedpixelsaroundthe611
mousebodywereconsidered,todiscardpatchcordmotion.Amousewasconsideredtobefreezingif612
38consecutivevalues(1.5s)werebelow983pixels(0.5%ofallpixels,EthoVision)or100pixels(within613
theROIaroundthemouse,OpenCV-Pythonscript).614
Immunofluorescenceandmicroscopy615
Hippocampalneuronalcultureswerefixedfor15minwith4%paraformaldehydeinPBS.Coverslipswere616
washedthreetimesinPBS,incubatedinblockingsolutionfor45min(10%normaldonkeyserum(NDS)617
with0.1%Triton inPBS)and thenexposedovernightat4°C tomonoclonalmouseanti-KCC2primary618
antibody (diluted1:1500 in5%NDS,PBS; catalog#167594S1-12;USBiological)and rabbitanti-MAP2619
(diluted1:1000in5%NDS,PBS;catalog#4542S;CellSignalingTechnology).Following3washesinPBS,620
coverslipswere incubated for2hat room temperature (RT)witha Cy5DonkeyAnti-Rabbit IgG (H+L)621
(diluted1:500in5%NDS,PBS;catalog#711-175-152;JacksonImmunoResearch)andCy3DonkeyAnti-622
Mouse IgG (H+L) (diluted1:1000 in 5%NDS, PBS; catalog # 715-165-151; Jackson ImmunoResearch).623
Coverslipswerethenwashed2timeswithPBS,dippedbrieflyintodouble-distilledwaterandembedded624
inDABCOmountingmedium (Sigma). Immunostainedneuronswere imagedwith a confocal scanning625
microscope(LSM700,CarlZeiss)usinga20xobjectiveforoverviewimages(NA0.8;CarlZeiss)anda63626
xoil immersionobjective(NA1.40;CarlZeiss) forquantification.Miceweredeeplyanesthetizedusing627
peer-reviewed) is the author/funder. All rights reserved. No reuse allowed without permission. The copyright holder for this preprint (which was not. http://dx.doi.org/10.1101/225847doi: bioRxiv preprint first posted online Dec. 8, 2017;

pentobarbital (0.4mg g−1 bodyweight) and perfused transcardially with ice-cold phosphate buffered628
saline (PBS, pH 7.4) followed by a solution of 4% paraformaldehyde (PFA) in PBS. After overnight629
postfixation at 4 °C, brainswere removed from the skull and incubated overnight in 4% PFA in PBS.630
Brainswerestoredinto30%sucroseinPBSforatleast24horuntilsectioning.Coronalsections(30μm631
or50µm)werecutonamicrotome(LeicaMicrosystems)andcollectedincryoprotectantsolution(25%632
glycerol, 30% ethylene glycol in PBS pH 6.7). Free-floating sections weremounted on gelatin-coated633
slides,dehydratedandembeddedinDABCOmountingmedium(Sigma). Imageswereacquiredusinga634
virtual slide scanner V (Olympus). Acquisition settingswere kept constantwithin each experiment to635
allowforcomparisonbetweenmice.636
Invitroilluminationanddrugapplication637
Whole-fieldillumination invitrowasperformedusinga470nmlightemittingdiode(29nmbandwidth638
LED; M470L2-C2; Thorlabs) delivered through the microscope illumination path including a custom639
dichroicinordertoreflectthe470nmactivationwavelength.Lightpowerdensitieswerecalculatedby640
measuring the light transmitted through the objective using a power meter (Thorlabs PM100A with641
S146C sensor) and dividing by the illumination area, calculated from the microscope objective field642
number andmagnification 65. D-AP5 (25 µM; ab120003; Abcam) and CNQX (10 µM; C-141, Alomone)643
werebathappliedduringallcultureexperiments.Forspatially-restrictedilluminationofneuronalsoma644
orneurites,a473nmdiodelaser(Bruker)wasdirectedtotheimagingplanewithgalvanometricmirrors,645
yieldingadiffraction-limitedspotof light thatprovidedbrief lightpulses (1ms)ateach location,with646
500msinter-pulseintervalsbetweennonadjacentlocations.647
Dataanalysisandstatisticalmethods648
During whole-cell patch-clamp recordings, pClamp 10 software (Molecular Devices) was used for649
acquisition. Data was analyzed using custom scripts written in Matlab (Mathworks). To quantify650
postsynapticcurrentamplitudesinresponsetolightpulses,holdingcurrenttraceswerefilteredwitha651
Savitzky-Golay11point,secondorder,Welchwindowfunctionfilterandthemaximalchangeinholding652
currentwithin20ms (EPSCs) after lightdeliverywasdetermined. Fiji (basedon ImageJ2;USNational653
InstitutesofHealth)wasusedforimmunofluorescenceimageanalysis.IntheKCC2immunofluorescence654
experimentallnumbers(n)refertothenumberofimagedaxons,inthetargetinghistologynrefersto655
thenumberofmice,andinelectrophysiologicalrecordings,nreferstothenumberofrecordedneurons656
/ units. To detect significantlymodulated units during in vivo silencing experiments, a paired-sample657
peer-reviewed) is the author/funder. All rights reserved. No reuse allowed without permission. The copyright holder for this preprint (which was not. http://dx.doi.org/10.1101/225847doi: bioRxiv preprint first posted online Dec. 8, 2017;

student’s t-testwasperformed comparing thenumberofdetectedactionpotentials between the5 s658
pre-lightperiodand the5 s lightperiodduring10 trialsper lightpower.Todetectantidromic spiking659
units, a paired-sample student’s t-test was performed comparing the number of detected action660
potentialsbetweenthe20mspre-lightperiodand the20ms lightperiodstartingwith the5ms light661
pulse.Allvaluesareindicatedasmean±SEM.Significancewasdeterminedatasignificancelevelof0.05662
withTukey'shonestlysignificantdifference(HSD)posthoctestusedtocorrectformultiplecomparisons.663
Incaseofnon-normaldatadistributionnon-parametrictestswereused:Mann–WhitneyUtestwasused664
forasinglecomparisons,theKruskal-WallisHtestforone-wayanalysisofvariance,andtheScheirerRay665
Haretestfortwo-wayanalysisofvariance.Nostatisticaltestswereruntopredeterminesamplesize,but666
sample sizeswere similar to those commonly used in the field. Blinding and randomizationwere not667
performed;howeverautomatedanalysiswasusedwheneverpossible.668
peer-reviewed) is the author/funder. All rights reserved. No reuse allowed without permission. The copyright holder for this preprint (which was not. http://dx.doi.org/10.1101/225847doi: bioRxiv preprint first posted online Dec. 8, 2017;





























