Embed Size (px)
Citation preview

Journal of Applied Microbiology 1998, 84, 293–303
Heterogeneity of plant-associated streptococci ascharacterized by phenotypic features and restrictionanalysis of PCR-amplified 16S rDNA
A. Ulrich and T. Mu¨ llerCentre for Agricultural Landscape and Land Use Research Muncheberg, Institute of Microbial Ecology and SoilBiology, Muncheberg, Germany
6104/02/97: received 12 February 1997, revised 19 May 1997 and accepted 22 May 1997
A. ULRICH AND T. MULLER. 1998. Thirty-five strains of streptococci isolated from foragegrasses were examined by restriction analysis of PCR-amplified 16S rDNA. Using a set ofseven endonucleases, five 16S rDNA genotypes were obtained. The isolates could beidentified as belonging to the species Enterococcus faecium, Ent. mundtii, Ent. faecalis, Ent.casseliflavus and Lactococcus lactis ssp. lactis, respectively. To assign the isolates to oneof these species, digestion with the endonuclease Hinf I was sufficient. Data obtained fromwhole-cell protein analysis were in accordance with the species identification.Additionally, three of the five species found were subdivided into two related subgroups.Both methods are suitable for a rapid and unequivocal identification of plant-associatedstreptococci. The phenotypic characters showed a high diversity and were insome cases contrary to the expected features usually given in genus and speciesdescriptions. The results demonstrated the close relatedness of the determined species incontrast to their high phenotypic heterogeneity.
INTRODUCTION species (for review see Schleifer et al. 1995). Although entero-cocci are predominantly inhabitants of the intestines of
Among lactic acid bacteria colonizing the leaf surface of veg-warm-blooded animals and humans, plant-associated strepto-
etable, cereal and forage plants, streptococci usually occur atcocci also belong to this genus (Mundt 1986). Species of
high frequency (Coggin et al. 1962 ; Mundt and Hammerenterococci, primarily Ent. faecalis, are of medical importance
1968 ; Ruser 1989 ; Lin et al. 1992). They exist on plants inbecause they cause several infections in humans and animals
a truly epiphytic relationship, are spread by seeds and are(Moellering 1992 ; Devriese et al. 1995). They are considered
able to reproduce on growing plants as demonstrated byto have sanitary significance as water quality indicators (Sin-
Mundt et al. (1962). Streptococci participate substantially inton et al. 1993b). Therefore, enterococci of aquatic eco-
the initiation of a spontaneous fermentation process of plantsystems, or of human and veterinary medical origins, unlike
material (Woolford 1984 ; Daeschel et al. 1987).those colonizing plant surfaces, have been well investigatedAs a result of the application of DNA : rRNA hybrid-and characterized. Additionally, many of the enterococci iso-ization, determination of 16S rRNA sequences, and sero-lated from plants have been reported to differ physiologicallylogical studies, streptococci were divided into the threefrom strains originating from warm-blooded animals (Mundtgenetically distinct genera Streptococcus, Enterococcus and1976 ; Devriese et al. 1992 ; Sinton et al. 1993b). Therefore,Lactococcus (for reviews see Schleifer 1987 ; Devriese et al.Mundt (1976) called these bacteria ‘plant type faecalis or1993 ; Sinton et al. 1993a). The genus Enterococcus wascasseliflavus biotype’.proposed for the faecal group of streptococci (Schleifer and
The significance of the genus Lactococcus is based mainlyKilpper-Balz 1984) and currently consists of at least 18on its large-scale use as a starter culture in the dairy industry.Lactococci normally occur in milk and, unlike enterococci,
Correspondence to : Dr A. Ulrich, Centre for Agricultural Landscape andthey are not usually found in faecal materials and have neverLand Use Research Muncheberg, Institute of Microbial Ecology and Soilbeen identified as causes of infectious diseases (Teuber et al.Biology, Eberswalder Str. 84, D-15374 Muncheberg, Germany (e-mail :
[email protected]). 1992). Nevertheless, they also occur on green plant material
© 1998 The Society for Applied Microbiology

294 A. ULRICH AND T. MULLER
(Teuber et al. 1992 ; Salama et al. 1995). Hence, the surface were enriched overnight in MRS broth and purified twice bystreaking on MRS agar.of plant material is a habitat occupied by Enterococcus as
well as Lactococcus. The ability to produce proteinaceous The following characteristics were considered in order toselect streptococci within the isolates: Gram-positive, cat-compounds with bactericidal activity (bacteriocins) is very
common in both genera (De Vuyst 1994a, b). As dem- alase-negative, homofermentative fermentation pathway, andproduction of L(¦)-lactic acid and spherical or ovoid cellsonstrated in a recent study (Muller et al. 1996), a bacteriocin-
like substance produced by an Ent. faecium strain isolated in short, medium or long chains. The procedures for thedetermination of these features were specified by Muller andfrom grass was very effective in inhibiting lactic acid bacteria
as well as other plant-associated bacteria. Although data Lier (1994). Thirty-five isolates selected by these charac-teristics (Table 1) were chosen for further investigation.obtained in this habitat are limited, it could be concluded
that strains with such antagonistic activity possibly have anecological function in the phyllosphere. A correct identi-
Strainsfication of the bacteria investigated in this habitat is theprerequisite for such ecological studies. Identification of The reference strains used in this study are the type strainsEnterococcus appears to be difficult at least on the basis of of the respective species and were obtained from the Germanphenotypic features (Devriese et al. 1993). There is a lack of Collection of Micro-organisms and Cell Cultures (DSMZ) :knowledge on the genetic variability and the natural diversity Enterococcus mundtii DSM 4838 (NCDO 2375), Ent. hiraewithin the genus Lactococcus (Salama et al. 1995). DSM 20160 (ATCC 8043), Ent. faecium DSM 20477 (NCTC
Comparison of 16S rRNA or rDNA nucleotide sequences 7171), Ent. faecalis DSM 20478 (NCDO 581), Ent. cas-can be applied to identify and classify isolates at species seliflavus DSM 20680 (NCDO 2372), Ent. durans DSMand higher levels (Woese 1987 ; Cilia et al. 1996), but these 20633, Lactococcus lactis spp. lactis DSM 20481 (NCDO 604)methods are too time-consuming for application in routine and L. lactis spp. cremoris DSM 20069 (NCDO 607). For allidentifications of isolates. Variations in the 16S rRNA genes reference strains listed above, total or nearly total 16S rDNAshould also be reflected in a restriction fragment length poly- sequences are known (Williams et al. 1991 ; accession no.morphism (RFLP). Thus, restriction analysis of amplified M58837, M58836).16S rDNA has been used successfully for differentiation andclassification of several bacterial species (Jayaro et al. 1991 ;Ralph et al. 1993 ; Laguerre et al. 1994). Phenotypic characterization
This study was carried out mainly to determine a way ofThe environmental isolates were tested for their ability torapidly and unequivocally identifying plant-associated strep-grow in MRS broth at 10°C and 45°C, at pH values adjustedtococci, which should be practicable in further ecologicalto 9·2 and 9·6 and with NaCl supplemented to final con-studies. Both well known classical methods of phenotypiccentrations of 4 and 6·5%, respectively. These tests were readcharacterization, and applied molecular-biological methods,after 5 d, except for growth at 10°C (8 d) and at 45°C (2 d).were used for this relatively neglected group of phylloplanePigmentation was examined by assessing the colour of thebacteria. By so doing, an insight into the heterogeneity ofsediments of liquid cultures. Their capacity for hydrolysingstreptococci isolated from grasses was also gained.arginine was analysed as described by Schade et al. (1986).Unless further specified, all tests were incubated at 30°C. To
MATERIALS AND METHODS select enterococci, the isolates were spread on membrane-filter agar according to Slanetz and Bartley (1957). In addition
Isolation of plant-associated streptococci to growth, reduction of triphenyl-tetrazolium chloride wasassessed by the colony colours (pink or dark-red indicatedLactic acid bacteria were isolated from different plantEnterococcus). The plates were incubated under aerobic con-material as shown in Table 1. For detaching the bacteria 30ditions for 2 d at 37°C.g samples of herbage were cut into pieces of about 5 mm
and homogenized for 2 min in 270 ml sterile water using aStomacher lab blender (Seward Medical 400, London, UK).
Antibiotic resistance testsAn aliquot of a logarithmic dilution of the homogenizedsample in quarter-strength Ringer’s solution was transferred Overnight cultures of each strain in MRS broth were spread
on MRS agar containing 15 mg ml−1 each of the antibioticsto Lactobaccillus MRS agar (De Man et al. 1960) (incubationfor 3 d at 30°C), Lactobacillus selective Rogosa agar (Rogosa actinomycin C (Ac), chloramphenicol (Cm), erythromycin
(Em), kanamycin (Km), penicillin G (Pn) and tetracyclineet al. 1951) (3d at 30°C), or Enterococcus selective SB agar(Slanetz and Bartley 1957) (2 d at 37°C) for enumeration of (Tc) (Sigma-Aldrich Chemie GmbH, Deisenhofen,
Germany). The plates were read for growth after 5 d.lactic acid bacteria. Cells of colonies picked out of the agar
© 1998 The Society for Applied Microbiology, Journal of Applied Microbiology 84, 293–303

PLANT-ASSOCIATED STREPTOCOCCI 295
Table 1 Isolates of plant-associatedstreptococci*
—–––––––––––––––––––––––––––––––––––––––––––––––––––––––––––––––––––––––––
Medium usedStrains Plant material for isolation—–––––––––––––––––––––––––––––––––––––––––––––––––––––––––––––––––––––––––P 194 Orchard grass MRSP 5250, P 5251, P 5252, Orchard grass SBP 5253, P5254, P 6318P 207 Meadow fescue MRSP 423 Pasture grass mixture MRSP 1955 Pasture grass mixture SBP 1905, P 7515 Pasture grass mixture ROGOSAP 5255 Timothy SBP 3911, P 6182, P 6187, Timothy ROGOSAP 6200, P 6284P 5385 Perennial ryegrass SBP 5977, P 5997, P 6025, Perennial ryegrass ROGOSAP 6056, P 6089P 7516, P 7517, P 7518 Litter of pasture grass SBP 7266, P 7297, P 7300, Litter of pasture grass ROGOSAP 7304, P 7306, P7416P 7341 Roots of pasture grass ROGOSAP 1109 Grass silage MRS—–––––––––––––––––––––––––––––––––––––––––––––––––––––––––––––––––––––––––
* Streptococci were isolated from pure stands of Dactylis glomerata (orchard grass),Festuca pratensis (meadow fescue), Phleum pratense (timothy), Lolium perenne(perennial ryegrass), as well as from a mixed cultivation of various grasses, using MRSagar, ROGOSA agar selective for lactic acid bacteria or SB agar described as selective forEnterococcus.
Polymerase chain reaction (PCR) Analysis of 16S rDNA fragments
Aliquots (2–8 ml) of the PCR products were digested withFor template preparation, single colonies grown on solidmedium were picked, resuspended in 20 ml 25 mmol l−1 each of the following enzymes : CfoI, HaeIII, AluI, HinfI,
MspI, Sau3A and ScrFI (Boehringer Mannheim, Germany ;NaOH/0·25% SDS and heated for 15 min at 95°C. Fromthis lysate, 0·2 ml were directly used for PCR without further New England Biolabs, Beverly, MA, USA). In order to detect
small differences between bands, the DNA fragments werepurification. Reference strains of potentially pathogenicEnterococcus species were only received as heat-inactivated separated in 2–3·5% Metaphor agarose gels (FMC Bio-
products, Rockland, ME, USA) depending on the fragmentcell material. However, these cells could be treated similarly.Primers fD1 and rD1 (Weisburg et al. 1991) used in this sizes to be distinguished. The DNA molecular weight mar-
kers V and VI, respectively, were used as size standardstudy are homologous to the consensus sequence of the 16SrRNA genes and are capable of amplifying nearly the com- (Boehringer Mannheim). The gels were stained with ethi-
dium bromide and documented with a video camera imageplete 16S rDNA. Routinely, a 50 ml reaction containing 10pmol of each primer in a standard buffer (Gibco BRL Life system (EasyImage plus, Herolab, Wiesloch, Germany). The
fraction of shared DNA fragments (F) was used to estimateTechnology Inc., Gaithersburg, MD, USA) was used. Theamplifications were performed in a GeneAmp PCR System the relative sequence divergence by the method of Nei and
Li (1979). Pairwise comparison resulted in a matrix which2400 (Perkin-Elmer Corporation, Norwalk, CT, USA) withthe following protocol : initial denaturation at 95°C for 2 min ; was the basis for calculation with the unweighted pair-group
method using arithmetic averages (UPGMA) (Sneath and30 cycles of 40 s at 94°C, 40 s at 54°C, 1 min at 72°C ; asingle final extension at 72°C for 3 min ; and a final soak at Sokal 1973). Further, algorithms according to Ward (1963)
and the weighted pair-group method using arithmetic aver-4°C (Weisburg et al. 1991 ; Laguerre et al. 1994 ; modified).After the reaction, 3 ml of the PCR products were examined ages (WPGMA) (Sneath and Sokal 1973) were used to con-
struct phylogenetic trees.in a 1% agarose gel.
© 1998 The Society for Applied Microbiology, Journal of Applied Microbiology 84, 293–303

296 A. ULRICH AND T. MULLER
SDS-PAGE contradictions were established in strains which grew wellwith pink colonies on SB agar but failed to grow in broth
To analyse the patterns of soluble cellular proteins, the iso-with 6·5% NaCl, at pH 9·6 and at 45°C (P 7341 and P 7516),
lates were grown in MRS bouillon at 30°C. Cells of stationaryand in strains which grew well with 6·5% NcCl and at pH
phase were harvested by centrifugation (11 000 g for 10 min9·6, but showed no or only weak growth on SB agar and no
at 4°C) and washed three times in quarter-strength Ringer’sgrowth at all at 45°C (P 7266, P 7304 and P 7306). Therefore,
solution. The cells were resuspended in 0·3 ml 50 mmol l−1
a classification of these strains on the basis of these phenotypicTris-HCl, pH 7·0 supplemented by 40 g l−1 human serum
characteristics was not possible.albumin (Serva), and disrupted in the presence of 0·6 g glassbeads (diameter 0·25–0·5 mm) using a bead mill (type MM-2, Retsch, Haan, Germany). After centrifugation (37 000 g Restriction analysis of amplified 16S rDNAfor 30 min at 4°C) the supernatant fluids were treated with
rDNA of all 35 strains isolated could be amplified with theNOVEX Tris-Glycine Sample Buffer (NOVEX, San Diego,
primers described by Weisburg et al. (1991) and showed aCA, USA) according to the manufacturers’ instructions. The
characteristic single band of about 1500 bp. Digestion of thesesamples were denatured at 85°C for 2 min and directly applied
PCR products with the seven endonucleases used resulted into SDS-PAGE using an Xcell IITM mini-cell electrophoresis
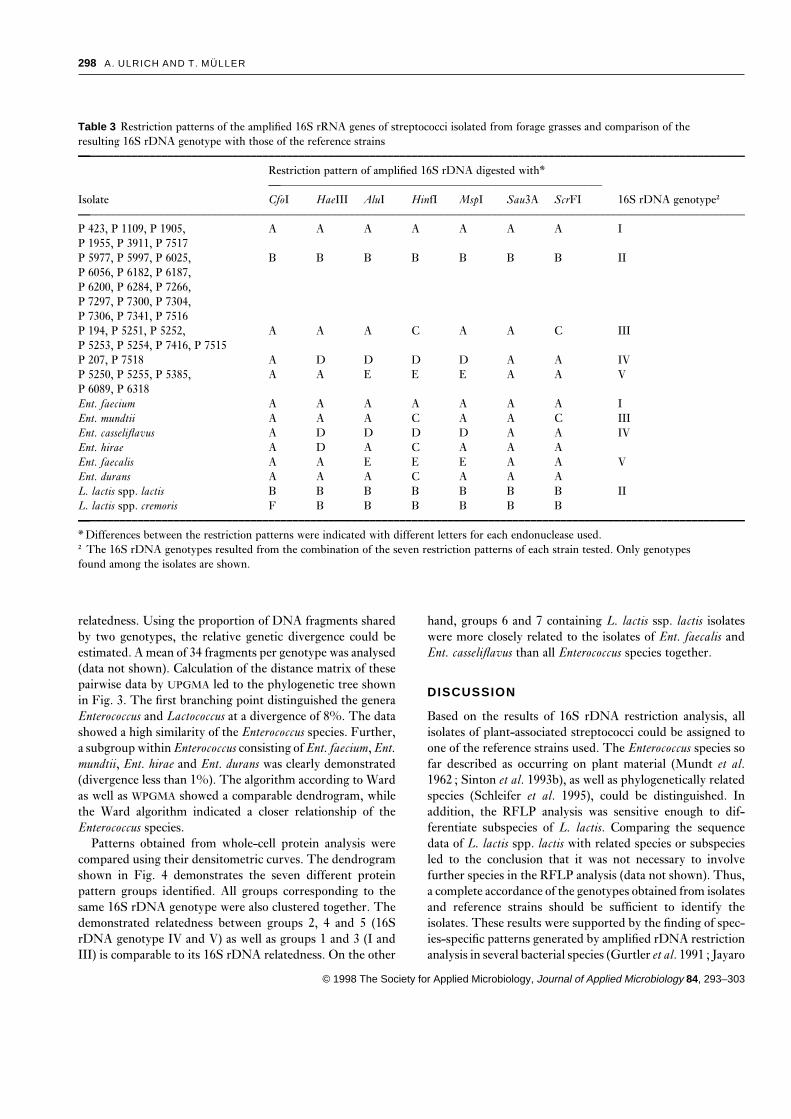
two to five different restriction patterns per enzyme (Tablesystem (NOVEX). SDS-PAA gels had a concentration gradi-
3, Fig. 1). DNA fragments smaller than 70 bp were notent of 4–20%. The gels were stained with Coomassie brilliant
well resolved in electrophoresis and therefore not used forblue and documented with a video camera image system
comparison of patterns. Summarizing all restriction patterns(EasyImage plus). To determine the relationship between the
led to five different 16S rDNA genotypes (I–V, Table 3). Twopatterns, the software program GelComparTM 3·0 (Applied
restriction patterns of one representative of each genotype areMaths BVBA, Kortrijk, Belgium) was used. Calculation par-
demonstrated in Fig. 1. It could be shown that use of theameters were as described by Pot et al. (1994). The patterns
endonuclease HinfI is sufficient to assign the isolatesof single tracks of the gels were analysed using densitometric
unequivocally to one of the genotypes.track curves. The calculation involved standardization of each
Further, all reference strains used in this study could bepattern to the protein marker pattern, background substrac-
amplified and distinguished by RFLP analysis (Table 3). Totion, and equalization of the total protein quantity per pattern.
differentiate these eight species and subspecies, at least fourThe dendrograms were clustered using UPGMA (Sneath and
enzymes (HinfI, HaeIII, ScrFI and CfoI) were required. TheSokal 1973).
presence of identical restriction patterns within the genotypesclearly differentiated both genera. The five genotypes (I–V)obtained from the isolates were identical to those of Ent.
RESULTSfaecium, L. lactis spp. lactis, Ent. mundtii, Ent. casseliflavusand Ent. faecalis, respectively. Therefore, all isolates of plant-
Phenotypic characteristicsassociated streptococci could be identified as belonging to oneof these species. The relation of Lactococcus to EnterococcusThirty-five streptococcal isolates of the phyllosphere were
investigated to assess their phenotypic diversity and to assign isolates was found to be 15 : 20. However, the Enterococcusisolates obtained were much more diverse in species than thethem, in a preliminary way, to either Enterococcus or Lacto-
coccus. Most of the phenotypic features shown in Table 2 are Lactococcus isolates which only belonged to one subspecies. Asregards the phenotypic characteristics, only the two isolatestypical for one of these two genera. However, the isolates were
of high phenotypic diversity. Summarizing all characters, 18 belonging to Ent. casseliflavus were completely homogeneous,whereas the isolates of all other species differed in one ordifferent phenotypic profiles were obtained which resulted in
several contradictions with the combinations of characters as more phenotypic features (Tables 2 and 3). Furthermore, theonly feature analysed which was typical for all isolates of adescribed in the taxonomic literature (Table 2). Two profiles
(strains P 423, P 1109, P 1905, P 7517 and strains P 5254, P species or a genus was pigmentation. All isolates of Ent.mundtii produced a yellow pigment.7416, respectively) could be unambiguously assigned to the
genus Enterococcus and only one profile to the genus Lac-tococcus (strains P 5977, P 5997, P 6025, P 6284, P 7297 and
Whole-cell protein analysisP 7300). It should be noted that growth on SB agar alone isnot a criterion for enterococci. However, the reduction of Each of the isolated strains was analysed by SDS-PAGE of
soluble cell proteins. All 35 isolates could be clustered intotriphenyl-tetrazolium chloride, which is manifest in reddish-coloured colonies, is much more definitive (Slanetz and Bar- one of seven groups of protein patterns (Table 2). No visible
differences between patterns of the isolates belonging to thetley 1957). Lactococci as well as enterococci are reported togrow at 10°C, but 17 strains failed to do so. The most striking same group could be detected. Conversely, patterns of the
© 1998 The Society for Applied Microbiology, Journal of Applied Microbiology 84, 293–303

PLANT-ASSOCIATED STREPTOCOCCI 297
Table 2 Phenotypic characteristics and groups of protein PAGE patterns of streptococci isolated from forage grasses—––––––––––––––––––––––––––––––––––––––––––––––––––––––––––––––––––––––––––––––––––––––––––––––––––––––––––––––––
Growth†—––––––––––––––––––––––––––––––––––––––––SB 4% 6·5% pH pH Hydrol. Presumed PAGE
Isolate Pigments* agar 10 °C 45 °C NaCl NaCl 9·2 9·6 arg.‡ genus§ ABR> pattern ¶—––––––––––––––––––––––––––––––––––––––––––––––––––––––––––––––––––––––––––––––––––––––––––––––––––––––––––––––––P 194 y ¦, dr − ¦w ¦ ¦ ¦ ¦ ¦ E.a − 1P 5251, P 5253 y ¦, p − ¦ ¦ ¦ ¦ ¦ ¦ E.a − 1P 5252 y ¦, p − ¦w ¦ ¦ ¦ ¦ ¦ E.a − 1P 5254, P 7416 y ¦, p ¦ ¦ ¦ ¦ ¦ ¦ ¦ E. − 1P 7515 y ¦, p − − ¦ ¦ ¦ ¦ ¦ E.a − 1P 1109, P 1905 − ¦, p ¦ ¦ ¦ ¦ ¦ ¦ ¦ E. − 1P 423, P 7517 − ¦, p ¦ ¦ ¦ ¦ ¦ ¦ ¦ E. − 3P 1955 − ¦, p − ¦ ¦ ¦ ¦ ¦ ¦ E.a Em 1P 3911 − ¦, p − ¦ ¦ ¦ ¦ ¦ ¦ E.a Em 3P 5250 − ¦, dr − ¦w ¦ ¦ ¦ ¦ − E.a − 2P 6089 − ¦, dr − ¦w ¦ ¦ ¦ ¦ − E.a − 4P 5255, P 5385 − ¦, dr − ¦ ¦ ¦ ¦ ¦ ¦ E.a − 4P 6318 − ¦, dr − ¦ ¦ ¦ ¦ ¦ − E.a − 4P 207, P 7518 − ¦, p − − ¦ ¦ ¦ ¦ − E.a − 5P 5977, P 5997, P 6025 − ¦w ¦ − ¦ − ¦ − ¦ L. − 6P 6284, P 7297, P 7300P 6056 − ¦w ¦ ¦w ¦ − ¦ − ¦ L.a − 6P 6182, P 6187, P 6200 − − − − ¦ − ¦ − ¦ L.a − 6P 7341 − ¦, p ¦ − ¦ − ¦ − ¦ − − 6P 7516 − ¦, p ¦ − ¦ − ¦ − − − − 6P 7266, P 7306 − ¦w ¦ − ¦ ¦ ¦ ¦ ¦ − − 7P 7304 − − ¦ − ¦ ¦ ¦ ¦ ¦ − − 7—––––––––––––––––––––––––––––––––––––––––––––––––––––––––––––––––––––––––––––––––––––––––––––––––––––––––––––––––
* y, Yellow pigment.† ¦w, weak positive result. SB agar can be used for selective growing of Enterococcus (see Materials and Methods).dr, p, Dark red and pink reaction, respectively.‡ Ability to hydrolyse arginine is species-specific in Lactococcus (Teuber et al. 1992) as well as in Enterococcus (Devriese et al. 1992).§ Summarizing the phenotypic characteristics resulted in the presumption of the genus Enterococcus or Lactococcus. Strains of bothgenera are described to grow at 10 °C, in a broth with 4% NaCl and in media adjusted to pH 9·2. Typical for Enterococcus isthe ability to grow also at 45 °C with 6·5% NaCl and at pH 9·6 (Devriese et al. 1993), whereas Lactococcus does not possess theseproperties (Teuber et al. 1992). a indicates isolates differing in one or more phenotypic characteristic from those described above.> ABR, Antibiotic resistance. The following antibiotics were screened : Em, Km, Tc, Cm, Ac, Pn.¶ Identical patterns obtained with SDS-PAGE of the soluble proteins are indicated by the same group number.
isolates from different groups are clearly distinguishable (Fig. lactis showed clearly different phenotypic features. All isolatesof group 6 were unable to grow in a medium containing 6·5%2). Each of the seven different patterns showed at least one
characteristic protein band. In contrast to the other pheno- NaCl and in a medium adjusted to pH 9·6. Conversely, allisolates of group 7 grew well in these media which is said totypic features, the groups of protein patterns were in con-
formity with the identified 16S rDNA genotypes. Some 16S be atypical for the genus Lactococcus (Schleifer 1987 ; Teuberet al. 1992).rDNA genotypes were further divided into two groups of
protein patterns. Thus, genotype I corresponded to groups 1and 3, genotype V to groups 2 and 4, and genotype II to
Relationship within 16S rDNA genotypes and withingroups 6 and 7. Only group 1 contained isolates from more
groups of protein patternthan one genotype, but both corresponding genotypes rep-resented the related species Ent. faecium and Ent. mundtii. All 16S rDNA genotypes obtained from the isolates and
reference strains were used to estimate their phylogeneticThe SDS-PAGE groups 6 and 7 representing L. lactis spp.
© 1998 The Society for Applied Microbiology, Journal of Applied Microbiology 84, 293–303
298 A. ULRICH AND T. MULLER
Table 3 Restriction patterns of the amplified 16S rRNA genes of streptococci isolated from forage grasses and comparison of theresulting 16S rDNA genotype with those of the reference strains—––––––––––––––––––––––––––––––––––––––––––––––––––––––––––––––––––––––––––––––––––––––––––––––––––––––––––––––––
Restriction pattern of amplified 16S rDNA digested with*—––––––––––––––––––––––––––––––––––––––––––––––––––––––––
Isolate CfoI HaeIII AluI HinfI MspI Sau3A ScrFI 16S rDNA genotype†—––––––––––––––––––––––––––––––––––––––––––––––––––––––––––––––––––––––––––––––––––––––––––––––––––––––––––––––––P 423, P 1109, P 1905, A A A A A A A IP 1955, P 3911, P 7517P 5977, P 5997, P 6025, B B B B B B B IIP 6056, P 6182, P 6187,P 6200, P 6284, P 7266,P 7297, P 7300, P 7304,P 7306, P 7341, P 7516P 194, P 5251, P 5252, A A A C A A C IIIP 5253, P 5254, P 7416, P 7515P 207, P 7518 A D D D D A A IVP 5250, P 5255, P 5385, A A E E E A A VP 6089, P 6318Ent. faecium A A A A A A A IEnt. mundtii A A A C A A C IIIEnt. casseliflavus A D D D D A A IVEnt. hirae A D A C A A AEnt. faecalis A A E E E A A VEnt. durans A A A C A A AL. lactis spp. lactis B B B B B B B IIL. lactis spp. cremoris F B B B B B B—––––––––––––––––––––––––––––––––––––––––––––––––––––––––––––––––––––––––––––––––––––––––––––––––––––––––––––––––
* Differences between the restriction patterns were indicated with different letters for each endonuclease used.† The 16S rDNA genotypes resulted from the combination of the seven restriction patterns of each strain tested. Only genotypesfound among the isolates are shown.
relatedness. Using the proportion of DNA fragments shared hand, groups 6 and 7 containing L. lactis ssp. lactis isolateswere more closely related to the isolates of Ent. faecalis andby two genotypes, the relative genetic divergence could be
estimated. A mean of 34 fragments per genotype was analysed Ent. casseliflavus than all Enterococcus species together.(data not shown). Calculation of the distance matrix of thesepairwise data by UPGMA led to the phylogenetic tree shown
DISCUSSIONin Fig. 3. The first branching point distinguished the generaEnterococcus and Lactococcus at a divergence of 8%. The data Based on the results of 16S rDNA restriction analysis, all
isolates of plant-associated streptococci could be assigned toshowed a high similarity of the Enterococcus species. Further,a subgroup within Enterococcus consisting of Ent. faecium, Ent. one of the reference strains used. The Enterococcus species so
far described as occurring on plant material (Mundt et al.mundtii, Ent. hirae and Ent. durans was clearly demonstrated(divergence less than 1%). The algorithm according to Ward 1962 ; Sinton et al. 1993b), as well as phylogenetically related
species (Schleifer et al. 1995), could be distinguished. Inas well as WPGMA showed a comparable dendrogram, whilethe Ward algorithm indicated a closer relationship of the addition, the RFLP analysis was sensitive enough to dif-
ferentiate subspecies of L. lactis. Comparing the sequenceEnterococcus species.Patterns obtained from whole-cell protein analysis were data of L. lactis spp. lactis with related species or subspecies
led to the conclusion that it was not necessary to involvecompared using their densitometric curves. The dendrogramshown in Fig. 4 demonstrates the seven different protein further species in the RFLP analysis (data not shown). Thus,
a complete accordance of the genotypes obtained from isolatespattern groups identified. All groups corresponding to thesame 16S rDNA genotype were also clustered together. The and reference strains should be sufficient to identify the
isolates. These results were supported by the finding of spec-demonstrated relatedness between groups 2, 4 and 5 (16SrDNA genotype IV and V) as well as groups 1 and 3 (I and ies-specific patterns generated by amplified rDNA restriction
analysis in several bacterial species (Gurtler et al. 1991 ; JayaroIII) is comparable to its 16S rDNA relatedness. On the other
© 1998 The Society for Applied Microbiology, Journal of Applied Microbiology 84, 293–303

PLANT-ASSOCIATED STREPTOCOCCI 299
ture are limited, the species composition found in the phyl-losphere is in contrast to those in clinical material, animals,faeces or waste water. In these habitats, Ent. faecalis, alone ortogether with Ent. hirae or Ent. faecium, is the predominantspecies. On the other hand, the species Ent. mundtii and Ent.casseliflavus are rare (Facklam and Collins 1989 ; Stern et al.1994 ; Blaimont et al. 1995 ; Kaufhold and Klein 1995). Theseobservations indicate the distinct populations in these habi-tats.
In the present study, we found an unexpectedly high diver-sity in phenotypic characters of strains of the same species.This finding was manifested in the failure to categorize allisolates into one of the genera by simple physiological tests.Devriese et al. (1993) have already indicated that an unequivo-cal separation of enterococci from other Gram-positive, cat-alase-negative cocci is not possible on the basis of availablephenotypic criteria. In our study, we found three strains
Fig. 1 Restriction patterns of the amplified 16S rDNA. An belonging to the genus Lactococcus (P 7266, P 7304 and Pexample of each of the five identified genotypes as given in Table 3 7306) which were also able to grow in broth with 6·5% NaCl.is shown. For clear demonstration, only the two bands used for This is in contrast to the suggestion that growth in thegroup differentiation are indicated for HinfI digestion. Lane 1 : P
presence of 6·5% NaCl is one of the best identification charac-1955 (I) ; lane 2 : P 7300 (II) ; lane 3 : P 194 (III) ; lane 4 : P 207ters of the genus Enterococcus (Devriese et al. 1992). This(IV) ; lane 5 : P 6318 (V). The lanes M (PBR328 ; HinfI/BglI) showfurther demonstrated that two, phenotypically clearly dif-the DNA standard markerferent subgroups of L. lactis spp. lactis existed on the plantmaterial. Using SDS-PAGE it is possible to distinguish bothsubgroups.
Yellow pigmentation is an indication for Ent. mundtii andEnt. casseliflavus (Facklam and Collins 1989). This could beconfirmed, in our study at least, for all strains of Ent. mundtiiidentified. As pigments were not found in our Lactococcusstrains and neither have they been reported to occur in thisgenus in the literature, this feature could sometimes be veryhelpful for differentiating plant-associated streptococci, par-ticularly if the portion of Ent. mundtii in the population isrelatively high. Formation of reddish colonies on SB agar dueto the reduction of triphenyl-tetrazolium chloride is a reactionthat can easily be observed and should therefore be consideredas a selective criterion for enterococci exclusively (Slanetzand Bartley 1957). In addition, according to Devriese et al.(1993), strong triphenyl-tetrazolium reducing activity and
Fig. 2 SDS-PAGE patterns of soluble proteins. An example of relatively large colonies are characters for a presumptive spec-each of the different patterns found as given in Table 2 is shown.
ies identification of Ent. faecalis. That was true in the presentLane 1 : P 5252 (1) ; lane 2 : P 5250 (2) ; lane 3 : P 423 (3) ; lane
study for five strains, but not true for another strain (P 194)4 : P 6089 (4) ; lane 5 : P 7518 (5) ; lane 6 : P 5977 (6) ; lane 7 : Pwhich was later identified as Ent. mundtii. However, we found7306 (7). The lane M represents the molecular weight markerthat the selectivity of the SB medium is not as strong as(MARK 12TM, Novex)supposed, because two (P 7341 and P 7516) out of 15 strainsidentified as L. lactis spp. lactis were also able to grow wellon this agar with reduction of the triphenyl-tetrazolium. Thecriteria of growth at 10°C in broth with 4% NaCl and at pHet al. 1991 ; Vaneechoutte et al. 1992 ; Laguerre et al. 1994).
The occurrence of Ent. faecium, Ent. mundtii, Ent. casseliflavus 9·2 were described as typical characters of enterococci as wellas lactococci. However, not all isolates of either genera wereand Ent. faecalis on plant material was in good agreement
with earlier studies (for review see Devriese et al. 1992). able to grow at 10°C. All species determined here have beendescribed as arginine hydrolysers (Teuber et al. 1992 ;Although data available from this study and from the litera-
© 1998 The Society for Applied Microbiology, Journal of Applied Microbiology 84, 293–303

300 A. ULRICH AND T. MULLER
Fig. 3 Phylogenetic tree showing the relationships between the identified 16S rDNA genotypes of the isolates and the reference strains,respectively. A matrix of pairwise genetic distances estimated from the comparison of the restriction fragments by the method of Nei and Li(1979) was used for calculation (UPGMA). The genotype numbers (I to V) are identical to those listed in Table 3
Fig. 4 Dendrogram characterizing the relationships between the classified strain groups based on SDS-PAGE patterns after calculationusing GelComparTM software. In addition to the SDS-PAGE groups (1–7) the corresponding 16S rDNA genotypes are indicated
© 1998 The Society for Applied Microbiology, Journal of Applied Microbiology 84, 293–303

PLANT-ASSOCIATED STREPTOCOCCI 301
Devriese et al. 1993), but some of our isolates were unable to Phylogenetic trees based on 16S rRNA sequences clearlydifferentiate the genera Lactococcus and Enterococcus (Schlei-degrade this amino acid.
Based on the results of screening for antibiotic resistance, fer et al. 1995). The further subdivision of Enterococcusrevealed the presence of some ‘species groups’ (Williams etit seems that this property is not very common in plant-
associated enterococci. Only three out of the 35 strains tested al. 1991) within the genus, such as the Ent. faecium group.The dendrogram using genetic distances estimated fromwere resistant to one out of the six antibiotics supplemented.
Conversely, antibiotic resistance is much more common in RFLP data showed the same branching points and hence wasin good agreement with these findings. The genetic distancesenterococci isolated from pork, water and clinical sources,
which were observed to have higher resistance rates par- obtained from data of restriction analysis and the 16S rRNAsequences were also comparable. However, the finding thatticularly to erythromycin and tetracycline (Knudtson and
Hartman 1993). This finding should be considered as further in some cases, no more than 38% shared fragments could befound indicates the limitation of this method for classificationindication of phenotypic differences between strains of strep-
tococci occurring in different habits, even though they rep- within and between related genera.The results of the present study revealed a high diversityresent the same species.
SDS-PAGE analysis of whole-cell protein patterns is in the phenotypic features of plant associated streptococciwhich prevented an unambiguous assignment into the generauseful for clearly discriminating a multitude of species of
lactic acid bacteria including Lactococcus and Enterococcus Enterococcus and Lactococcus in most cases. However, bothSDS-PAGE of the whole-cell proteins and RFLP analysis of(Descheemaeker et al. 1994). It is equally possible to dif-
ferentiate and identify Enterococcus species (Merquior et al. the 16S rDNA, respectively, are rapid and reproducible toolsfor identifying plant-associated streptococci. Diversity within1994). This could be confirmed by our results showing a high
agreement between data obtained from analysis of the 16S and between species occurring in this habitat can be clearlyindicated. This information might be helpful for inves-rDNA genotypes and SDS-PAGE patterns, respectively. In
addition, we could show a further subdivision of the isolates tigations of ecological interactions in the phyllosphere.of a species by SDS-PAGE. Descheemaeker et al. (1994)could assign L. lactis strains to their respective subspecies
ACKNOWLEDGEMENTSby SDS-PAGE. However, a high similarity was observedbetween L. lactis spp. lactis and ssp. cremoris strains. On the This work was supported by the Bundesministerium furother hand, we were able to find two electrophoretic groups Ernahrung, Landwirtschaft und Forsten and the Ministeriumwithin L. lactis spp. lactis demonstrating the heterogeneity of fur Ernahrung, Landwirtschaft und Forsten des Landesthe isolates in the subspecies. Teixera et al. (1995) showed Brandenburg. Mrs Liane Linkg, Karin Luttich, Kerstinthat protein patterns could also be used for accurate charac- Umann and Sigune Weinert are thanked for technical assist-terizations of physiologically atypical strains of Ent. faecium. ance, and Ole Wendroth for critical reading of the manu-This is in accordance with our results showing that physio- script.logically different isolates could be classified to their Enter-ococcus species using SDS-PAGE. However, in some cases,
REFERENCESstrains of the related species Ent. mundtii and Ent. faecium arenot unambiguously separable by their protein pattern (SDS- Blaimont, B., Charlier, J. and Wauters, G. (1995) ComparativePAGE pattern group 1). On the other hand, strains of Ent. distribution of Enterococcus species in faeces and clinical samples.faecium form a further electrophoretic group. This dem- Microbial Ecology in Health and Disease 8, 87–92.onstrates the strong relationship of both species in contrast Cilia, V., Lafay, B. and Christen, R. (1996) Sequence heterogeneitiesto the high heterogeneity within the species Ent. faecium. among 16S ribosomal RNA sequences, and their effect on phylo-
genetic analyses at the species level. Molecular Biology and Evol-The pattern obtained by SDS-PAGE has been shown to beution 13, 451–461.reliable for comparing and grouping closely related species and
Coggin, J.H., Mundt, J.O. and Johnson, L.F. (1962) Growth andstrains (Pot et al. 1994 ; Vauterin et al. 1996). Its correlationproliferation of Streptococcus faecalis var. liquefaciens on plants.with DNA-DNA hybridization also indicates the value ofBacteriological Proceedings 62, 22.phylogenetic studies (Costas 1992 ; Teixera et al. 1995). We
Costas, M. (1992) Classification, identification, and typing of bac-could cluster SDS-PAGE pattern groups of one species as wellteria by the analysis of their one-dimensional polyacrylamide gel
as related species in accordancewith their 16S rDNAgenotype. electrophoretic protein patterns. Advances in Electrophoresis 5,However, the genera Lactococcus and Enterococcus would not 351–408.be correctly categorized. This confirms the conclusion of Van- Daeschel, M.A., Andersson, R.E. and Fleming, H.P. (1987)damme et al. (1996) that the use of SDS-PAGE for classi- Microbial ecology of fermenting plant materials. FEMS Micro-fication is limited by the fact that it yields only discriminative biology Reviews 46, 357–367.
De Man, J.C., Rogosa, M. and Sharpe, M.E. (1960) A medium forinformation at or below the species level.
© 1998 The Society for Applied Microbiology, Journal of Applied Microbiology 84, 293–303

302 A. ULRICH AND T. MULLER
the cultivation of lactobacilli. Journal of Applied Bacteriology 23, Moellering, R.C. (1992) Emergence of Enterococcus as a significantpathogen. Clinical Infectious Diseases 14, 1173–1178.130–135.
Muller, M. and Lier, D. (1994) Fermentation of fructans by epi-De Vuyst, L. (1994a) Bacteriocins produced by Lactococcus lactisphytic lactic acid bacteria. Journal of Applied Bacteriology 76, 406–strains. In Bacteriocins of Lactic Acid Bacteria : Microbiology, Gen-411.etics and Applications ed. De Vuyst, L. and Vandamme, E.J. pp.
Muller, T., Behrendt, U. and Muller, M. (1996) Antagonistic143–149. London : Chapman and Hall.activity in plant-associated lactic acid bacteria. MicrobiologicalDe Vuyst, L. (1994b) Bacteriocins of Enterococcus. In Bacteriocins ofResearch 151, 63–70.Lactic Acid Bacteria : Microbiology, Genetics and Applications ed.
Mundt, J. O. (1976) Streptococci in dried and frozen foods. JournalDe Vuyst, L. and Vandamme, E.J. pp. 511–513. London : Chap-of Milk and Food Technology 39, 413–416.man and Hall.
Mundt, J. O. (1986) Enterococci. In Bergey’s Manual of SystematicDescheemaeker, P., Pot, B., Ledeboer, A.M., Verrips, T. andBacteriology ed. Sneath, P.H.A., Mair, N.S., Sharpe, M.E. andKersters, K. (1994) Comparison of the Lactococcus lactis dif-Hold, J.G. pp. 1063–1065. Baltimore : Williams & Wilkins.ferential medium (DCL) and SDS-PAGE of whole-cell proteins
Mundt, J. O. and Hammer, J.L. (1968) Lactobacilli on plants.for the identification of lactococci to subspecies level. SystematicApplied Microbiology 16, 1326–1330.and Applied Microbiology 17, 459–466.
Mundt, J. O., Coggins, J.H. and Johnson, L.F. (1962) Growth ofDevriese, L.A., Collins, M.D. and Wirth, R. (1992) The genusStreptococcus faecalis var. liquefaciens on plants. Applied Micro-Enterococcus. In The Procaryotes 2nd edn. ed. Balows, A., Truper,biology 10, 552–555.H.G., Dworkin, M., Harder, W. and Schleifer, K.-H. pp. 1465–
Nei, M. and Li, W.-H. (1979) Mathematical model for studying1481. New York : Springer Verlag.genetic variation in terms of restriction endonucleases. ProceedingsDevriese, L.A., Pot, B. and Collins, M.D. (1993) Phenotypic identi-of the National Academy of Sciences of USA 76, 5269–5273.fication of the genus Enterococcus and differentiation of phylo-
Pot, B., Vandamme, P. and Kersters, K. (1994) Analysis of elec-genetically distinct enterococcal species and species groups.trophoretic whole-organism protein fingerprints. In ModernJournal of Applied Bacteriology 75, 399–408.Microbial Methods. Chemical Methods in Procaryotic SystematicsDevriese, L.A., Chiers, K., De Herrdt, P. et al. (1995) Enterococcused. Goodfellow, M. and O’Donnell, A.G. pp. 493–521. Chich-hirae infections in psittacine birds : epidemiological, pathologicalester : John Wiley & Sons Ltd.and bacteriological observations. Avian Pathology 24, 523–531.
Ralph, D., McClelland, M., Welsh, J., Baranton, G. and Perolat,Facklam, R.R. and Collins, M.D. (1989) Identification of Entero-P. (1993) Leptospira species categorized by arbitrarily primedcoccus species isolated from human infection by a conventionalpolymerase chain reaction (PCR) and by mapped restriction poly-test scheme. Journal of Clinical Microbiology 27, 731–734.morphisms in PCR-amplified rRNA genes. Journal of BacteriologyGurtler, V., Wilson, V.A. and Mayall, B.C. (1991) Classification of175, 973–981.
medically important clostridia using restriction endonuclease siteRogosa, M., Mitchell, J.A. and Wiseman, R.F. (1951) A selective
differences of PCR-amplified 16S rDNA. Journal of Generalmedium for the isolation of oral and faecal lactobacilli. Journal of
Microbiology 137, 2673–2679.Bacteriology 62, 132–133.
Jayaro, B.M., Dore, J.J.E., Baumbach, G.A., Matthews, K.R. andRuser, B. (1989) Erfassung und Identifizierung des epiphytischen
Oliver, S.P. (1991) Differentiation of Streptococcus uberis from Milchsaurebakterienbesatzes auf Gras and Mais in AbhangigkeitStreptococcus parauberis by polymerase chain reaction and restric- von Standort, Sorte, Entwicklungsstadium sowie Ernte- und Kli-tion fragment length polymorphism analysis of 16S ribosomal maeinflussen. Landbauforschung Volkenrode, Sonderheft 103.DNA. Journal of Clinical Microbiology 29, 2774–2778. Salama, M.S., Musafija-Jeknic, T., Sandine, W.E. and Giovannoni,
Kaufhold, A. and Klein, R. (1995) Species identification and anti- S.J. (1995) An ecological study of lactic acid bacteria : isolation ofbiotic susceptibility of enterococci isolated from clinical speci- new strains of Lactococcus including Lactococcus lactis subspeciesmens of hospitalized patients. International Journal of Medical cremoris. Journal of Dairy Science 78, 1004–1017.Microbiology, Virology, Parasitology and Infectious Diseases 282, Schade, W., Wunschel, R. and Jahrig, A. (1986) Kurzschema zur507–518. Gattungsdiagnostik von in Bier auftretenden Bakterien. Berichte
Knudtson, L.M. and Hartman, P.A. (1993) Antibiotic resistance der Humboldt-Universitat Berlin 6, 11–37.among enterococcal isolates from environmental and clinical sour- Schleifer, K.-H. (1987) Recent changes in the taxonomy of lacticces. Journal of Food Protection 56, 489–492. acid bacteria. FEMS Microbiology Reviews 46, 201–203.
Laguerre, G., Allard, M.R., Revoy, F. and Amarger, N. (1994) Schleifer, K.-H. and Kilpper-Balz, R. (1984) Transfer of Strep-Rapid identification of rhizobia by restriction fragment length tococcus faecalis and Streptococcus faecium to the genus Enterococcuspolymorphism analysis of PCR-amplified 16S rRNA genes. nom. rev. as Enterococcus faecalis comb. nov. and EnterococcusApplied and Environmental Microbiology 60, 56–63. faecium comb. nov. International Journal of Systematic Bacteriology
Lin, C., Bolsen, K.K., Brent, B.E. et al. (1992) Epiphytic microflora 34, 31–34.on alfalfa and whole-plant corn. Journal of Dairy Science 75, 2484– Schleifer, K.-H., Ehrmann, M., Beimfohr, C., Brockmann, E., Lud-2493. wig, W. and Amann, R. (1995) Application of molecular methods
Merquior, V.L.C., Peralta, J.M., Facklam, R.R. and Teixeira, L.M. for the classification and identification of lactic acid bacteria.(1994) Analysis of electrophoretic whole-cell protein profiles as a International Dairy Journal 5, 1081–1094.tool for characterization of Enterococcus species. Current Micro- Sinton, L.W., Donnison, A.M. and Hastie, C.M. (1993a) Faecal
streptococci as faecal pollution indicators : a review. Part I : Tax-biology 28, 149–153.
© 1998 The Society for Applied Microbiology, Journal of Applied Microbiology 84, 293–303

PLANT-ASSOCIATED STREPTOCOCCI 303
onomy and enumeration. New Zealand Journal of Marine and Vandamme, P., Pot, B., Gillis, M., de Vos, P., Kersters, K. andSwings, J. (1996) Polyphasic taxonomy, a consensus approach toFreshwater Research 27, 101–115.
Sinton, L.W., Donnison, A.M. and Hastie, C.M. (1993b) Faecal bacterial systematics. Microbiological Reviews 60, 407–438.Vaneechoutte, M., Rossau, R., De Vos, P. et al. (1992) Rapid identi-streptococci as faecal pollution indicators : a review. Part II :
Sanitary significance, survival, and use. New Zealand Journal of fication of bacteria of the Comamonadaceae with amplified riboso-mal DNA-restriction analysis (ARDRA). FEMS MicrobiologyMarine and Freshwater Research 27, 117–137.
Slanetz, L.W. and Bartley, C.H. (1957) Numbers of enterococci in Letters 93, 227–234.Vauterin, L., Swings, J. and Kerters, K. (1996) Protein elec-water, sewage, and faeces determined by the membrane filter
technique with an improved medium. Journal of Bacteriology 74, trophoresis and classification. In Handbook of New Bacterial Sys-tematics ed. Goodfellow, M. and O’Donnell, A.G. pp. 251–280.591–595.
Sneath, P.H.A. and Sokal, R.R. (1973) Numerical Taxonomy. San London : Academic Press Ltd.Ward, J.H. (1963) Hierarchical grouping to optimize an objectiveFrancisco : W.H. Freeman & Co.
Stern, C.S., Carvalho, M.D.S. and Teixeira, L.M. (1994) Charac- function. Journal of the American Statistical Association 58, 236–244.terization of enterococci isolated from human and nonhuman
sources in Brazil. Diagnostic Microbiology and Infectious Disease Weisburg, W.G., Barns, S.M., Pelletier, D.A. and Lane, D.J. (1991)16S ribosomal DNA amplification for phylogenetic study. Journal20, 61–67.
Teixeira, L.M., Facklam, R.R., Steigerwalt, A.G., Pigott, N.E., of Bacteriology 173, 697–703.Williams, A.M., Rodrigues, U.M. and Collins, M.D. (1991) Intrag-Merquior, V.L.C. and Brenner, D.J. (1995) Correlation between
phenotypic characteristics and DNA relatedness within Enter- eneric relationship of enterococci as determined by reversetranscriptase sequencing of small-subunit rRNA. Research inococcus faecium strains. Journal of Clinical Microbiology 33, 1520–
1523. Microbiology 142, 67–74.Woolford, M.K. (1984) The Silage Fermentation. New York : MarcelTeuber, M., Geis, A. and Neve, H. (1992) The genus Lactococcus.
In The Procaryotes, 2nd edn ed. Balows, A., Truper, H.G., Dwor- Dekker Inc.Woese, C.R. (1987) Bacterial evolution. Microbiological Reviews 51,kin, M., Harder, W. and Schleifer, K.-H. pp. 1482–1500. New
York : Springer Verlag. 221–271.
© 1998 The Society for Applied Microbiology, Journal of Applied Microbiology 84, 293–303





























