Embed Size (px)
Citation preview

Fort Hays State University Fort Hays State University
FHSU Scholars Repository FHSU Scholars Repository
Master's Theses Graduate School
Spring 2010
Herpetofaunal response to prescribed burning on Matagorda Herpetofaunal response to prescribed burning on Matagorda
Island, Texas: with emphasis on Texas horned lizard (Phrynosoma Island, Texas: with emphasis on Texas horned lizard (Phrynosoma
cornutum) cornutum)
Ashley Inslee Fort Hays State University
Follow this and additional works at: https://scholars.fhsu.edu/theses
Part of the Biology Commons
Recommended Citation Recommended Citation Inslee, Ashley, "Herpetofaunal response to prescribed burning on Matagorda Island, Texas: with emphasis on Texas horned lizard (Phrynosoma cornutum)" (2010). Master's Theses. 171. https://scholars.fhsu.edu/theses/171
This Thesis is brought to you for free and open access by the Graduate School at FHSU Scholars Repository. It has been accepted for inclusion in Master's Theses by an authorized administrator of FHSU Scholars Repository.

HERPETOFAUNAL RESPONSE TO PRESCRIBED BURNING ON MATAGORDA
ISLAND, TEXAS: WITH EMPHASIS ON TEXAS HORNED LIZARD
(PHRYNOSOMA CORNUTUM)
being
A Thesis Presented to the Graduate Faculty
of the Fort Hays State University in
Partial Fulfillment of the Requirements for
the Degree of Master of Science
by
Ashley A. Inslee
B.S., Fort Hays State University
Date____________________ Approved__________________________________ Major Professor
Approved__________________________________ Chair, Graduate Council

i
This thesis for the Master of Science Degree
By
Ashley A. Inslee
has been approved
________________________________________
Chair, Supervisory Committee
________________________________________
Supervisory Committee
________________________________________
Supervisory Committee
________________________________________
Supervisory Committee
_______________________________________
Chair, Department of Biology

ii
ABSTRACT
Texas horned lizard (Phrynosoma cornutum) populations have been in steady
decline over the past twenty years due to habitat loss, pesticide use, the pet trade, and
invasion of the red imported fire ant (Solenopsis invicta). At Aransas National Wildlife
Refuge the P. cornutum population has declined, except for on Matagorda Island. A
prescribed burn regime was initiated to maintain the coastal prairie habitat. This closed
environment was unique in its composition of arid adapted and wetland species. There
was minimal human disturbance, and a single road bisects the length of the island. The
island was invaded by S. invicta around 1993. I conducted an extensive survey of the
island herpetofauna to determine the distribution of P. cornutum, as well as to quantify
the influence of the burn regime on P. cornutum. I also investigated the effect of road-
side habitat on both P. cornutum and habitat structure, as well as the microhabitat
associations of P. cornutum.
Herpetofaunal populations were investigated in six distinct habitat types on
Matagorda Island by using drift-fence arrays with funnel traps, random 50-m walks, and
systematic road cruising. In 2009, individual P. cornutum were captured along the road
and individuals were tracked by using a fluorescent powder method. I also sampled
habitat structure and ant mounds along a 60-m transect perpendicular to Middle Road.
Habitat among burn treatments exhibited significant differences in percent ground
cover, standing stem density, and soil compaction in 2008 and 2009. Eleven
herpetofaunal species were captured in the drift fence arrays. The most abundant species

iii
were 55 six-lined racerunners (Aspidoscelis sexlineata) and 16 slender glass lizards
(Ophisaurus attenuatus); however, only three P. cornutum were captured with drift-fence
arrays, of which I observed high mortality rates by S. invicta. I did not observe any P.
cornutum on the random 50-m searches. I captured 41 individuals in 2008 and 128
individuals in 2009 along the road, and estimated the population size at 138 individuals
by using the Schnabel index. Two distinct distributions in the population were isolated
by a recent burn in both 2008 and 2009. A majority of P. cornutum activity occurs
within 5 m of the roadside, which accounts for only 2% of the grassland habitat. Ant
species also were not randomly distributed on the island. Solenopsis invicta was
observed throughout the island, along with cone ants (Dorymyrmex sp.). Acrobat ants
(Crematogaster sp.) were concentrated on the north and south portions of the island. No
Pogonomyrmex sp. were observed.
Phrynosoma cornutum were associated positively with older burn treatments and
the road-side habitat, and were negatively associated with recent burns. Habitat variation
was not the only factor that influenced distribution. Solenopsis invicta occurred in high
abundance in Burn 1 treatments. The distribution of Crematogaster sp. was congruent
with the distribution of P. cornutum. In areas with the most heterogeneous habitat, I
observed high diversity of ant genera. Phrynosoma cornutum were observed in
alarmingly low abundance and warrant monitoring and augmented management
strategies.

iv
PREFACE
I have written this thesis in two chapters and formatted each to fulfill the
requirements of the author guidelines for The Southwestern Naturalist. I will submit
chapters one and two to the aforementioned journal shortly after the final thesis draft is
accepted.

v
ACKNOWLEDGMENTS
I would never have imagined myself leaving a small island in Kenya, to find
myself on another small island off the coast of Texas. But this was the case because two
of my undergraduate professors saw potential in a naive and aspiring wildlife biologist.
Very special thanks to Dr. Bill Stark for taking on the arduous task of being my graduate
advisor, and to Dr. Greg Farley who was able to see possibilities for me that I could not
see for myself.
I am incredibly grateful to have had the opportunity to work under Dr. Bill Stark.
I admire his profound amount of knowledge and his dedication to education as a
profession. He has gone out of his way on numerous occasions to help shape me into a
professional biologist and storyteller. I thank my undergraduate advisor and graduate
committee member, Dr. Greg Farley, for pushing me to think critically and for providing
valuable insights to my project and writing. I truly appreciate Dr. Rob Channell, who
provided me with statistical support and applicable ecological knowledge. I thank Curtis
Wolf for stepping up into a vacant committee member position. His inspiration and
active involvement in both my undergraduate and graduate careers has been invaluable.
I appreciate Chad Stinson for hiring me as a biological technician at Aransas
National Wildlife Refuge. He was a key contributor in the development, implementation,
and success of this project. I also value the hard work and dedication of every employee
in the U.S. Fish and Wildlife Service, especially those at Aransas National Wildlife
Refuge for allowing me to use their facilities and equipment. I particularly appreciate
Carey Strobel and Vicki Muller for preoccupying me while I was off the island, and for

vi
their insight, support, and friendship. The Turtle Patrol volunteers for Matagorda Island
were an added gift to this project. I treasure their support, friendship, food, and stories.
Matagorda Island was a beautiful study site, but at moments could be a ruthless
and ugly place to conduct field work. For this reason, I am grateful for my persistent, yet
patient and very positive field assistants, Megan Hughes and Georgina Jacquez. They
bared their teeth and stuck it out through heat and humidity, mosquitoes, hurricanes, and
break downs (both mental and with vehicles). I hope you cherish that experience.
I also thank Chad Tillberg for his insight into the great world of Formicidae. His
expertise and ability to identify ants was of great value to this project. A special thanks
to the Fleharty family for their financial contribution to this project through the Fleharty
Fellowship. Also, to my fellow peers at Fort Hays State University, thank you for all of
the weekend rendezvous, late nights in the office, and assistance with this project,
especially Lizz Waring, Georgina Jacquez, Megan Hughes, Ryan Pinkall, and Elita
Baldridge.
I am grateful for my family, who supported my decision to continue my education
and for their encouragement. Tremendous thanks to Chris Jones, a true friend, field
assistant, and confidant. Your unconditional love, patience, and support gave me the
motivation to complete this task. I will forever be in your debt. Also to Cooper, whose
walks and playful antics made unbearable days seem bearable.
Many people were responsible for the completion of this project, several of whom
might not be mentioned in the preceding yet whom I truly appreciate.

vii
TABLE OF CONTENTS
Page
SIGNATURE PAGE ........................................................................................................... i
ABSTRACT ........................................................................................................................ ii
PREFACE .......................................................................................................................... iv
ACKNOWLEDGMENTS ...................................................................................................v
TABLE OF CONTENTS .................................................................................................. vii
LIST OF TABLES ............................................................................................................. ix
LIST OF FIGURES ........................................................................................................... xi
CHAPTER 1. HERPETOFAUNAL RESPONSE TO HABITAT MANIPULATION ON
MATAGORDA ISLAND, TEXAS: WITH EMPHASIS ON THE TEXAS
HORNED LIZARD (PHRYNOSOMA CORNUTUM). ...........................................1
INTRODUCTION .........................................................................................................1
STUDY AREA ........................................................................................................7
METHODS ....................................................................................................................9
DRIFT-FENCE ARRAYS .......................................................................................9
VEGETATION SURVEYS ...................................................................................11
ROAD SURVEYS .................................................................................................11
CATCH PER UNIT EFFORT ...............................................................................12
STATISTICAL ANALYSIS .................................................................................12
RESULTS ....................................................................................................................13
VEGETATION ......................................................................................................13

viii
DRIFT-FENCE ARRAYS .....................................................................................14
ROAD CAPTURES ...............................................................................................15
ASSESSMENT OF EFFORT TYPE .....................................................................16
DISCUSSION ..............................................................................................................16
LITERATURE CITED ................................................................................................21
CHAPTER 2. MICROHABIT ASSOCIATIONS OF TEXAS HORNED LIZARD
(PHRYNOSOMA CORNUTUM) WITH INFERENCE FROM POWDER
TRACKING. ..........................................................................................................39
INTRODUCTION .......................................................................................................39
METHODS ..................................................................................................................46
HABITAT ..............................................................................................................46
TRACKING ...........................................................................................................46
ANTS .....................................................................................................................48
STATISTICAL ANALYSIS .................................................................................49
RESULTS ....................................................................................................................51
HABITAT ..............................................................................................................51
TRACKING ...........................................................................................................52
ANTS .....................................................................................................................54
DISCUSSION ..............................................................................................................55
FUTURE RESEARCH AND MANAGEMENT RECOMMENDATIONS .........62
LITERATURE CITED ................................................................................................64

ix
LIST OF TABLES
Chapter 1
TABLE PAGE
1 Abundance of species captured in drift-fence arrays within specified habitat types
from June-August on Matagorda Island, Texas. Significant G-test results are
denoted with an asterisk (G = 24.028, df = 5, p < 0.001). .....................................27
2 Range and mean snout-vent length, total length, and body mass of males, females,
and juveniles by road captures of P. cornutum from June-August 2008 on
Matagorda Island, Texas. .......................................................................................28
3 Chi-square analysis of road captures of P. cornutum showing a significant difference
in habitat use in adjacency to three types of burn regimes within the 50% isopleths
from June-August on Matagorda Island, TX. Percent is the percent of available
adjacent habitat combinations. ...............................................................................29
Chapter 2
TABLE PAGE
1 Range and mean snout-vent length, total length, and body mass of males, females,
and juveniles for road captures of P. cornutum from June-August 2009 on
Matagorda Island, Texas. .......................................................................................71
2 Chi-square analysis of road captures of adult P. cornutum exhibiting a significant
difference in habitat use in adjacency to three types of burn treatments from June-
August 2009 on Matagorda Island, TX. Percent is the percent of available
adjacent habitat combinations ................................................................................72

x
3 Chi-square analysis of road captures of juvenile P. cornutum exhibiting a significant
difference in habitat use in adjacency to three types of burn treatments from June-
August 2009 on Matagorda Island, TX. Percent is the percent of available
adjacent habitat combinations ................................................................................73
4 Microhabitat variables identified by a backward (Wald) logistic regression that are
most influential in predicting the presence of P. cornutum on Matagorda Island,
TX (χ2 = 219.015, df = 3, p < 0.001). ....................................................................74
5 Frequency of occurrence of ant mounds (genera) along thirty 60 m transects
systematically distributed along Middle Road from May-June 2009 on Matagorda
Island, TX.. ............................................................................................................75

xi
LIST OF FIGURES
Chapter 1
FIGURE PAGE
1 Design of a Y-shaped drift-fence array ........................................................................30
2 Design of a drift-fence array for Road-side habitat. ....................................................31
3 Variation in mean percent canopy cover among six habitat types during June and July
2008 on Matagorda Island, Texas. A Kruskal-Wallis followed by a multiple
comparisons test detected significant differences among groups of habitats (H =
223.36, df = 5, p < 0.001). Habitat types that share a letter designation are not
significantly different. ............................................................................................32
4 Variation in mean vegetation compression index (cm) among six habitat types during
June and July 2008 on Matagorda Island, Texas. A Kruskal-Wallis followed by a
multiple comparisons test detected significant difference among groups of
habitats (H = 183.32, df = 5, p < 0.001). Habitat types that share a letter
designation are not significantly different. ............................................................33
5 Variation in mean soil compaction at 7.6 cm (psi) among six habitat types during
June and July 2008 on Matagorda Island, Texas. A Kruskal-Wallis followed by a
multiple comparisons test detected significant differences among groups of
habitats (H = 149.84, df = 5, p < 0.001). Habitat types that share a letter
designation are not significantly different. ............................................................34
6 Mean species richness among six habitat types during June and July 2008 on
Matagorda Island, Texas. A Kruskal-Wallis followed by a multiple comparisons

xii
test detected significant differences among groups of habitats (H = 13.800, df = 5,
p = 0.032). Habitat types that share a letter designation are not significantly
different. .................................................................................................................35
7 Road captures of P.cornutum from June through July on Matagorda Island, Texas.
Chi-square test of independence detected a nonrandom distribution, and a 50%
isopleth outline the distributions. ...........................................................................36
8 Differences among observed numbers of P. cornutum and expected numbers within
adjacent burn-unit combinations along Middle Road on Matagorda Island, Texas
(χ2 = 21.279, df = 5, p < 0.05). The thick line along the zero axis is the calculated
expected value for a random distribution among habitat types. The dashed line
along the x-axis represents middle road and the combination of adjacent habitat
types .......................................................................................................................37
9 Catch per unit effort of P. cornutum from June through August on Matagorda Island,
Texas. Effort types are measured in hours with the exception of Trap Night, and
are defined as the following: Cruise is actively searching from a vehicle at 0-24
kph, Vehicle is actively searching at 24-48 kph, Passive Observations is not
actively searching from >48 kph, Walking is actively searching while walking
from the road to the drift-fence arrays, Transect is actively searching while
walking in a random direction for 50 m. Trap Night is the number of P. cornutum
captured over the total number of traps opened throughout the study per night ...38

xiii
Chapter 2
FIGURE PAGE
1 Locations of systematic surveys of habitat structure within the prescribed burn regime
on Matagorda Island, Texas. ..................................................................................76
2 Discriminant function analysis of nine distance-interals oriented perpendicular to
Middle Road. The distance-intervals separated into two groups (road-side areas
and all others) based on characteristics of habitat structure (Eigenvalue=8.351,
Wilks λ=0.107, df = 13, p < 0.001). .......................................................................77
3 Captures of adult P.cornutum from June-July 2009 on Matagorda Island, Texas. Chi-
square test of independence detected a nonrandom distribution, and a 50%
isopleth outlines the isolated distributions .............................................................78
4 Captures of juvenile P.cornutum from June-July 2009 on Matagorda Island, Texas.
Chi-square test of independence detected a nonrandom distribution, and a 50%
isopleth outlines the isolated distributions .............................................................79
5 Differences among observed numbers of adult P. cornutum and expected numbers
relative to the proportion of available combinations of burn treatments along
Middle Road, Matagorda Island, Texas (χ2 = 58.686, df = 4, p < 0.05). The thick
line along the zero x-axis is the expected value for a random distribution of P.
cornutum on the island. The dashed line along the x-axis title represents Middle
Road and the combination of adjacent burn treatments .........................................80
6 Differences among observed numbers of juvenile P. cornutum and expected numbers
relative to the proportion of available combinations of burn treatments along

xiv
Middle Road, Matagorda Island, Texas (χ2 = 155.952, df = 4, p < 0.05). The thick
line along the zero x-axis is the expected value for a random distribution of P.
cornutum on the island. The dashed line along the x-axis title represents Middle
Road and the combination of adjacent burn treatments .........................................81
7 Relative abundance (number of mounds) for ants (genera) observed along 120-m
transects systematically distributed along Middle Road, Matagorda Island, TX ..82

1
CHAPTER 1
HERPETOFAUNAL RESPONSE TO HABITAT MANIPULATION ON
MATAGORDA ISLAND, TEXAS: WITH EMPHASIS ON THE
TEXAS HORNED LIZARD (PHRYNOSOMA CORNUTUM)
Keywords: Matagorda Island, Phrynosoma cornutum, prescribed burning, reptile
response
INTRODUCTION
Speciation and extinction are evolutionary processes that have been operating
since the origin of life and affecting the number and types of organisms present on Earth.
The fossil record and current species lists suggest that only 2-4% of species that have
ever existed still survive today (Hilton-Taylor, 2007). However, with the increase in
human population, increases in anthropogenically-caused extinctions also have been
observed. The World Conservation Union (IUCN) estimated rate of extinction is 1,000
to 10,000 times greater than the expected natural rate (Hilton-Taylor, 2007). Of the 8,163
reptile species recognized by the IUCN, only 488 species have been evaluated and 61%
of those species were listed as threatened (Baillie et al., 2004). In 1996, 353 species of
reptiles were listed as threatened or endangered by the IUCN; that number increased to
515 species by 2006 (Hilton-Taylor, 2007). Amphibians have experienced more
precipitous declines that have gained public attention and led to political action (Gibbons
et al., 2000). The IUCN have described and evaluated 5,743 amphibian species with 32%
listed as threatened (Baillie et al., 2004). Population declines and extinctions of

2
herpetofauna primarily occur because of habitat loss or degradation and invasive species
(Gibbons et al., 2000; Butchart, 2008).
Other factors contributing to the decline of reptiles and amphibians include
pollution, disease, parasitism, unsustainable use, global climate change, or combinations
of these factors (Gibbons et al., 2000; Hilton-Taylor, 2007; Butchart, 2008). Actions to
address these factors are warranted; however, there are many issues that influence
conservation efforts. These issues include, but are not limited to, monetary cost,
perceived species importance, level of threat, and the time it would take to achieve
conservation goals (McCarthy et al., 2008).
At the turn of the twentieth century, Americans began to recognize the value of
wildlife, and concern was reinforced by the decline of iconic species like bison (Bison
bison) and passenger pigeon (Ectopistes migratorius) populations (Decker et al., 2001).
A few laws including the Lacey Act of 1900, the Migratory Bird Conservation Act of
1929, the Bald Eagle Protection Act of 1940, and the Endangered Species Preservation
Act of 1966 have been enacted in response to the declines in populations of particular
species (Coggins, 1974). A paradigm shift during this time, led by individuals such as
Gifford Pinchot and Aldo Leopold, moved conservation efforts toward a more scientific
and ecological approach to studying wildlife management (Decker et al., 2001).
However, it was not until the Endangered Species Act (ESA) of 1973 that a law was
enacted because ―various species of fish, wildlife, and plants in the United States have
been rendered extinct as a consequence of economic growth and development
untempered by adequate concern and conservation‖ (16 USC Sec. 1531, ESA). The ESA
provided a foundation for habitat protection, land mitigation, and public recognition of

3
threatened species that ultimately increased American populations of species such as the
threatened brown pelican (Pelecanus occidentalis), American alligator (Alligator
mississippiensis), bald eagle (Haliaeetis leucocephalus), whooping crane (Grus
americana), and gray wolf (Canis lupus).
Natural resource managers must consider the biology, ecology, and physiology of
organisms for appropriate management. For example, the thermal ecology of an
ectotherm can be radically different from an endotherm is. Ectotherms generally are
more energy efficient in low productivity environments, and can dedicate more energy
towards growth and reproduction instead of maintaining a constant body temperature as
in endotherms. Yet ectotherms are limited both spatially and temporally in their activity
patterns, because they rely on external heat sources to increase their body temperature.
Most reptiles and amphibians are active in warmer seasons and at lower latitudes (Cowles
and Bogert, 1944; Prieto and Whitford, 1971; Huey and Stevenson, 1979). Reptiles and
amphibians differ in their thermal ecology because radiation is more influential in
reptiles, whereas evaporation has a larger effect on amphibians. Once an ectotherm
reaches its optimal thermal range, it is more effective at capturing prey, escaping
predators, thermoregulating, defending resources, producing healthier offspring, among
other activities (Huey and Stevenson, 1979). Effective management should recognize
these fundamental processes.
Improving long-term management and conservation strategies are continuous
challenges for wildlife managers. Restoring or maintaining biodiversity and the stability
of an ecosystem is a common goal. Two concepts offer guidance for managers in
developing sustainable ecological management plans, 1) past processes and conditions as

4
a context to mimic, and 2) the role of disturbance-driven spatial and temporal variability
in maintaining ecological stability (Landres et al., 1999). Prescribed burn regimes are
often used to suppress the growth of woody vegetation in grasslands (Bragg and Hulbert,
1976). However, it can have substantial effects on vegetative structure and species
composition (Collins, 1992; Van Dyke et al., 2007). Prescribed burning also can increase
habitat heterogeneity when burns occur at the appropriate frequency spatially and
temporally, thus indirectly benefitting overall biodiversity (Collins, 1992; Fuhlendorf et
al., 2006). Prescribed burning indirectly affects the herpetofaunal community by
changing food availability (i.e. arthropod community- Siemann et al., 1998; Benson et al.,
2007; small mammal community- Kaufman et al., 1990), protective vegetative cover
(Benson et al., 2007), and microclimate (Burrow et al., 2001). Moreover, burning can
potentially injure or fatally wound members of the herpetofaunal community do to direct
and indirect exposure to the fire (Prieto, 1995; Cavitt, 2000; Durbian, 2005).
Moeller (2000) reported that prescribed burning was beneficial to Texas horned
lizards (Phrynosoma cornutum) in thornscrub savanna, and attributed this response to the
positive effect that burning had on harvester ants (Pogonomyrmex sp.), the primary prey
of P. cornutum. Similar responses of P. cornutum to prescribed burning in various other
habitats also have been observed (Fair and Henke, 1997b; Burrow et al., 2001).
Thermoregulation also is influenced by vegetative composition and structure (Burrow et
al., 2001). Russell (1997) determined body temperatures of P. cornutum were more
similar to substrate temperatures than air temperatures, and P. cornutum regulated body
temperatures within a range of 34.9-38.6°C. As environmental temperatures rise above

5
the critical thermal limit of P. cornutum, the lizards thermoregulate within their thermal
preference by selecting appropriate microhabitats (Russell, 1997).
Both temperature and habitat structure are influenced by a latitudinal gradient.
The distribution of P. cornutum ranges latitudinally from northern Kansas to northern
Mexico, and longitudinally from western Louisiana to eastern Arizona (Price, 1990);
therefore, temperature and habitat vary within this distribution. According to Henke and
Montemayor (1998), in southern Texas, P. cornutum is most active from April-August,
and is encountered most frequently from May-June. They restrict activity to the morning
from June-August (Henke and Montemayor, 1998).
Matagorda Island, a barrier island along the coastal bend of Texas, is managed by
the United States Fish and Wildlife Service (FWS) based in Aransas National Wildlife
Refuge (Aransas NWR) (McAlister, 2004). In 1992, a burn regime was implemented on
the island by Aransas NWR staff to suppress woody vegetation and manage invasive
species. The island was divided into burn units that were burned primarily in the winter
on a 3-5 year rotation (Chad Stinson, pers. comm.). Most burns were conducted from
October to February due to staff availability and favorable weather conditions (Chad
Stinson, pers. comm.). The land cover consisted of salt marsh along the bay-side and
beach along the gulf-side of the island, which were intermittently inundated with water.
Grassland occurred throughout the interior of the island. Therefore, the herpetofaunal
community is distinct in its composition of arid and wetland adapted species. In addition,
a well documented record of land management is available to assess current and
historical vegetative composition and overall community structure. Matagorda Island
provided a study site with minimal immigration or emigration of terrestrial organisms,

6
and minimal human impact. In the early 1990s, Donaldson et al. (1994) surveyed the
state of Texas for the distribution of P. cornutum. In Aransas County, the southern
mainland county adjacent to Matagorda Island, they captured and marked ten P.
cornutum; however, in Calhoun County, the northern county adjacent to Matagorda
Island, they surveyed for P. cornutum but none were observed (Donaldson et al., 1994).
From 2003-2006, Aransas NWR staff monitored four 50-m long drift-fence arrays each
month on the mainland adjacent to the southern portion of Matagorda Island, and never
captured or observed P. cornutum (Chad Stinson, pers. comm.). Therefore, it is unlikely
that P. cornutum could emigrate from the mainland.
From 1992-1995, Prieto (1995) conducted a monthly survey of small terrestrial
vertebrates and their responses to seasonal prescribed burns on Matagorda Island by
using drift-fence arrays. Prieto (1995) concluded that winter burns provided more
suitable habitat and higher diversity than summer or unburned treatments. However,
Prieto (1995) did not quantify vegetative differences among burn treatments. Prieto
(1995) captured 16 reptile species, 3 amphibian species, and 2 mammal species, and
made several qualitative observations. He rated the species as uncommon, common,
abundant, and seasonal; and although P. cornutum were captured rarely, Prieto (1995)
labeled them as common. Between 1995-2007, Aransas NWR staff suspected a decline
in the population of P. cornutum on the island and has tried to avoid the decline observed
in the mainland population. In 2007, Aransas NWR staff conducted a one day survey for
P. cornutum on the southern half of Matagorda Island and observed eight individuals. On
two additional occasions that same year, two more individuals were observed. However,
refuge staff never kept a detailed record of the capture effort or number of individual

7
horned lizards captured. My study will provide baseline quantitative evaluation of
populations of P. cornutum on Matagorda Island.
My objectives were to 1) conduct a herpetofaunal survey of Matagorda Island, 2)
quantify differences in macrohabitat types on Matagorda Island, 3) attempt to estimate
the abundance and distribution of P. cornutum on Matagorda Island, and 4) to identify the
habitat used by P. cornutum on Matagorda Island relative to the scale of current
management manipulations.
I hypothesized that species richness will be highest in a treatment that was burned
one year prior, along Road-sides and Runways, which might facilitate thermoregulation.
I also hypothesized that the burn treatments will differ in plant structure. According to
Burrow et al. (2001), P. cornutum prefer areas with a matrix of bare ground and
herbaceous vegetation. Therefore, I hypothesized that the distribution of P. cornutum on
the island will be closely associated with first year burns and the road to meet
thermoregulation requirements.
Study Area
I conducted this project on Matagorda Island, a unit of the Aransas NWR
Complex, located on the coastal bend of Texas in Calhoun County, Texas. This island
was located on the eastern edge of the range for P. cornutum. Matagorda Island was
approximately 61 km long and ranged from 1-7 km wide (McAlister and McAlister
2006). It encompassed approximately 500 ha of beach, 10,500 ha of tidal salt marsh, and
9,200 ha of upland grassland. The island was composed of deep shell sand and had a
mild climate that received approximately 102 cm of rainfall annually (Handbook of
Texas Online, http://www.tshaonline.org/handbook/online/articles/CC/hcc2.html).

8
McAlister and McAlister (2006) summarized the history of landuse and
ownership on Matagorda Island. In the late 1800s, the southern 14.5 km of the island
were used for ranching. In 1942, the northern 45 km of the island were acquired by the
federal government as a practice bombing range for the United States Army Air Force,
later the United States Air Force. In 1982 the Texas General Land Office placed Texas
Parks and Wildlife Department (TPWD) in charge of public use, and the Department of
Interior, through the United States Fish and Wildlife Service (USFWS) at Aransas NWR,
was to manage all the natural resources. By 1986, land rights for the southern portion of
Matagorda Island were sold to The Nature Conservancy. The Nature Conservancy began
selling the land, in portions, to the Department of Interior. Currently, the USFWS is
responsible for wildlife and habitat management on the island. The federal and state
governments jointly manage the area and designated it the Matagorda Island National
Wildlife Refuge and State Natural Area (McAlister and McAlister, 2006).
Matagorda Island is unique within the Texas barrier island chain in that it is
uninhabited and has minimal public access. A single road bisects the length of the island
and is used by TPWD, petroleum exploration vehicles, and the USFWS. Exterior
portions of the island are intermittently inundated with water as part of salt marshes, tidal
flats, and beach. The interior portion, and a majority of the island land area, consisted of
flat, rolling grasslands and sand dunes (McAlister and McAlister, 2006). I identified
common plant species and used common names according to coastal Texas field guides
by Richardson (2002) and Lehman et al. (2005), and species included: bermudagrass
(Cynodon spp.), big bluestem (Andropogon gerardii), bushy bluestem (Andropogon
glomeratus), firewheel (Gaillardia pulchella), marshhay cordgrass (Spartina patens),

9
glasswort (Salicornia bigelowvii and Sesuvium spp.), gulfdune paspulum (Paspalum
monostachyum), Johnson grass (Sorghum halepense), mesquite (Prosopis glandulosa),
powderpuff (Mimosa strigillosa), prickly pear cactus (Oputia engelmanii), ragwort
(Senecio riddellii), sandburs (Cenchrus incertus), seacoast bluestem (Schizachyrium
scoparium), sea oats (Uniola paniculata), silverleaf sunflower (Helianthus argophyllus),
snoutbean (Rhynchosia americana), sombrerillo (Hydrocotyle bonariensis), Texas
frogfruit (Phyla nodiflora), western ragweed (Ambrosia psilostachya), and white topped
umbrella grass (Dichromena colorata).
METHODS
Drift-fence Arrays
I identified six macrohabitat types that comprised the majority of area habitable
by upland herpetofauna. Three of the macrohabitat types were semi-permanent and
included secondary Dunes, Road-sides, and Runways from the former Air Force base.
The remaining three macrohabitat types were maintained by prescribed burning. These
habitats were defined as a burn that occurred within the previous year (Burn 1), a burn
two years prior (Burn 2), and a burn three or more years prior (Burn 3).
I placed Y-shaped drift-fence arrays with funnel traps (Figure 1) in four replicates
of each of the six habitat types. The arrays were constructed of hardware clothe and 9-
gauge wire, and stabilized with rebar. Each array consisted of three 7.6-m long fences
radiating from the center point at 120° angles. Two 1.2-m funnel traps with openings at
both ends were placed on both sides of each fence near the midpoint. Terminal funnel
traps were placed at the hub and apices of each array.

10
Trap sites were selected randomly within treatments by using Hawthstools
(Beyer, 2004) with Geographic Information System (GIS) (ArcGIS 9.2, Environmental
Systems Research Institute (ESRI), Redlands, CA) with the following limitations: 1)
Whitford and Bryant (1979) and Fair and Henke (1999) both reported an average daily
movement range of P. cornutum from 9 to 91 m; therefore, a 100-m buffer was placed
inside each treatment to avoid dispersal bias from adjacent macrohabitat types, 2) to
avoid flooding in the burn regimes, drift fence sites were placed on the high plot of land
nearest to the actual random point. The secondary Dunes were selected by randomly
plotting four points on the beach and then walking inland, perpendicular to the surf line,
until reaching the first secondary dune structure. Runway habitats were mapped as
polygon shapefiles with GIS ArcMap 9.2 to randomly select four sites; such that the
nearest plot of soil was selected as the runway site. Road-sides also were mapped as a
polygon shapefile to randomly select four sites. Once Road-side locations were selected,
trap legs equivalent to one drift-fence array were placed alternately on each side of the
road (Figure 2).
The traps were opened four consecutive days per month during June, July, and
August 2008. For each species trapped, I recorded species, date, time, and trap number.
The snout-vent length (cm), total length (cm), mass (g) (Pesola Precision Scales, Baar,
Switzerland), and sex were recorded for P. cornutum. A toe clip was collected from the
third digit of the rear, right foot to mark individuals, which complied with Herpetological
Animal Care and Use Committee guidelines (Guidelines for Use of Live Amphibians and
Reptiles in Field and Laboratory Research, http://www.asih.org/files/hacc-final.pdf).
Male horned lizards were identified by the presence of femoral pores and swelling on

11
both sides of the base of the tail near the cloaca (Moeller et al., 2005). Phrynosoma
cornutum with a snout-vent length <60 mm were considered juveniles and their sex could
not be determined with certainty (Lane, 2008). I also walked a 50-m transect in a random
direct away from each array to search specifically for P. cornutum.
Vegetation Surveys
Vegetation surveys were conducted at the 24 arrays in late June and late July
2008. Nine samples were taken around the perimeter of each array. I used a Daubenmire
frame to quantify percentage canopy cover and identified vegetation cover type (forb,
grass, bare ground, prostrate shrub, shrub, water, and litter). I measured litter depth in the
center of the Daubenmire frame, and soil compaction at 7.6 cm (DICKEY-john
Corporation, Auburn, IL) in the center of the Daubenmire frame. A compression disk
meter (Sharrow, 1984) was used to generate an index of stem density. I also recorded the
most common plant species around the drift-fence arrays.
Road Surveys
Road cruising consisted of a two person team (driver and recorder) that searched
for P. cornutum. At each capture location the same capture information was recorded as
with the drift-fence captures, and a vegetation survey was conducted. In an effort to
assess distribution pattern on the island, I used the road capture locations to construct a
50% isopleth by using Home Range Tools (Rodgers and Carr, 2002) with GIS ArcMap
9.2 (ESRI, Redlands, CA). An isopleth outlined the home range area where 50% of the
total population is located. Six combinations of adjacent burn units occur along Middle
Road and the relative proportions of each were determined.

12
Catch Per Unit Effort
I recorded the effort type for each lizard encounter to evaluate the efficiency of
each effort type. Three search types occurred from a motor vehicle, and were defined as
Cruising (0-24 kph), Vehicle (24-48 kph), and Passive Observation (>48 kph). Two
other search types were defined as Walking, when I was walking from the road to the
drift-fence arrays, and Transect, when I walked the 50-m transect in a random direction.
For funnel traps, effort type was measured by trap night. I compared the effect of capture
methods on number of lizard captures per effort hour.
Statistical Analysis
A Kruskal-Wallis test followed by a test of multiple comparisons (Conover, 1980)
was used to compare differences among herpetofaunal richness, diversity, catch per unit
effort (CPUE), and variation of habitat structure among habitat types. Percentage of
canopy cover, compression index, and soil compaction were used to characterize habitat
types. A G-test of independence was used to evaluate species-habitat association. I used
a chi-square test of independence to assess if P. cornutum are randomly distributed on the
island or if there is a preference for a certain burn regime.
RESULTS
Vegetation
A Kruskal-Wallis detected significant differences in percent canopy cover among
treatments (H = 223.363, df = 5, p < 0.001). Runways and Burn 1 treatments differed
from each other (46.2% and 31.0%, respectively) and exhibited the lowest percent ground

13
cover of any treatment type (Figure 3). Ground cover in the Dune habitat type was
intermediate at 76.0%, and was significantly different from all other habitat types. Burn
2 (97.3%), Burn 3 (94.9%) and Road-sides (92.7%) exhibited nearly complete ground
cover and were significantly different from all other treatments.
A Kruskal-Wallis analysis detected significance of estimates of standing stem
density, as indicated by a compression index, which exhibited a similar pattern to percent
canopy cover among treatments (H = 183.319, df = 5, p < 0.001). Burn 1 and Runway
treatments were similar with the lowest compression indices (Figure 4). Dune and
Roads-side treatments had intermediate values that were significantly different from both
Burn 1 and Runways, and Burn 2 and Burn 3. Burn 2 and Burn 3 were similar and had
significantly higher compression index values than other habitat types.
Significance also was detected by a Kruskal-Wallis analysis among estimates of
soil compaction (H = 149.845, df = 5, p < 0.001). Dunes had the lowest mean
compaction reading, followed by Burn 1 (Figure 5). Burn 2, Burn 3, and Road-side
treatments were statistically similar at an intermediate level and Runways had the highest
compaction reading.
Drift-fence Arrays
Eighteen species were captured in the drift-fence arrays (Table 1). Six of these
were non-target species (not members of the herpetofaunal community). I did not
observe any negative interactions among the non-target and target species. The fulvous
harvest mouse (Reithrodontomys fulvous) and the cotton rat (Sigmodon hispidus) were
the most frequently captured mammals, 12 and 42, respectively; and were evenly
distributed among habitat types. Two Northern bobwhite chicks (Colinus virginianus)

14
were caught in a Burn 1 trap. Three genera of crab were caught in the traps: a land crab
(Gecarcinus sp.), two small land crabs (Sesarma sp.), and 61 ghost crabs (Ocypode sp.).
Ocypode sp. was only captured in Dune habitat.
The Southern leopard frog (Lithobates sphenocephalus) was the only amphibian
captured. Eleven species of reptiles were captured in drift-fence arrays. The most
frequently captured lizard species were six-lined racerunners (Aspidoscelis sexlineata)
and slender glass lizards (Ophisaurus attenuatus), 55 and 16 captures, respectively.
Fifteen coachwhips (Coluber flagellum) also were captured. Aspidoscelis sexlineata, O.
attenuatus, and C. flagellum are vagile species that were observed in nearly every habitat
type. Gulf Coast ribbon snakes (Thamnophis proximus) were distributed nonrandomly
among habitat types with a positive association with the Road-side habitat (G = 24.028,
df = 5, p < 0.001). I captured fifteen T. proximus which were restricted to Burn 2, Burn
3, and Road-side habitats. L. sphenocephalus, T. proximus, and the cottonmouth
(Agkistrodon piscivorus) are mesic adapted species that were captured in Burn 2, Burn 3,
and Road-side habitats usually near some standing water. Eight massasagua rattlesnakes
(Sistrurus catenatus), four Western diamondback rattlesnakes (Crotalus atrox), two
speckled kingsnakes (Lampropeltis getula holbrooki), an Eastern hognose snake
(Heterodon platirhinos), and a Louisiana milksnake (Lampropeltis triangulum) were
captured sporadically among habitat types. Three Texas horned lizards (P. cornutum)
also were captured. Each was captured in a different habitat: Dune, Burn 2, and Road-
side habitats. Accordingly, habitat preference for P. cornutum could not be determined
from this data.

15
Mean species richness was highest in Burn 2, Burn 3, and Road-side habitats, at
an intermediate level in the Dune, and lowest in Burn 1 and Runway habitats (H =
13.801, df = 5, p = 0.032) (Figure 6). The most species rich habitat was the Road-side
with 8 species, and the least species rich was Burn 1 habitat with two species. Although I
observed similar patterns in species diversity and catch per array night, significant
differences could not be detected due to high variability and low sample sizes.
Road Captures
Forty-one P. cornutum were captured along the road from May-July, and I
documented 11 recaptures. Captures comprised 25 females, 12 males, and 4 juveniles.
Average snout-vent length of adults was 79.3 cm and ranged from 58.0-92.5 cm (Table
2). The average total length of adults was 117.5 cm and ranged from 92.0-132.0 cm.
Body mass for adults ranged from 32.0-72.0 g with an average of 33.0 g.
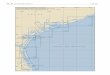
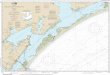
The 50% isopleth indicates two disjunct populations within which 50% of the
observations occurred (Figure 7). The two 50% isopleths were separated by
approximately 10 km. This indicated a nonrandom distribution on the island, which led
me to investigate distribution at the treatment scale.
A chi-square test of independence indicated that observations of P. cornutum did
not occur in the same proportion as available treatment combinations (χ2 = 21.279, df = 5,
p < 0.05) (Table 3). Fewer observations occurred than expected when Burn 1 treatments
were adjacent to both sides of the road and when Burn 3 treatments were adjacent to both
sides of the road. Fewer observations also occurred than expected when Burn 1
treatments were adjacent to Burn 3 treatments. Conversely, observations exceeded

16
expectations when Burn 2 treatments were adjacent to either another Burn 2 treatment or
Burn 3 treatment (Figure 8).
Assessment of Effort Type
Catch per unit effort was calculated for each sampling effort type: Cruising,
Vehicle, Passive Observation, Walking, Transects (walking), and Array Nights (Figure
9). Cruising was the most efficient effort type at 0.3960 captures per hour (cap/hr),
closely followed by Vehicle (0.2429 cap/hr). Passive Observations (0.1481 cap/hr) and
Walking (0.0231 cap/hr) were more efficient than Transects in which no P. cornutum
were captured. Trap nights were the second least efficient effort type with 0.0014 P.
cornutum caught per trap night. Drift fences were the most labor intensive and expensive
capture type, and resulted in the fewest overall captures except for walking random
transects. Mortality rates were also high among all species in the drift-fence arrays due to
depredation by the red imported fire ant (Solenopsis invicta).
DISCUSSION
Reptiles are ectotherms and rely on their environment to optimally thermoregulate
(Cowles and Bogert, 1944; Russell, 1997). Accordingly, I might have anticipated high
numbers of individuals associated with Burn 1, Runway, and Road-side habitats. While
this hypothesis is supported for the Road-side, a weak negative association was observed
among the herpetofauna and Burn 1 and Runway habitat types. Runways had similar
vegetative characteristics to Burn 1 habitats. This might be attributed to the majority of
the runway complex being burned in the previous winter. Three of the drift-fence arrays
in the runway complex were within an area recently burned, while the fourth was in an

17
area burned two seasons ago. The strong negative association observed between P.
cornutum and Burn 1 and Runway habitat types might be attributed to a delay in
recruitment or increased mortality after disturbance (Nichols and Menke, 1984).
Burning temporarily reduces biomass and overall productivity (Chang, 1996), in
that it results in reduced cover used for predator avoidance (Burrow et al., 2001).
Precipitation levels were below average for the spring and summer and might account for
low recolonization rates. Prieto (1995) proposed that recolonization rates increase with
precipitation due to higher productivity.
Because ectotherms are more active in warmer months, there is a higher risk of
mortality due to prescribed burning in summer months (Prieto, 1995; Cavitt, 2000). Male
P. cornutum move more in April and May to intercept receptive females (Stark et al.,
2005). Cavitt (2000) and Prieto (1995) suggest that winter burns reduce mortality rates
of herpetofauna compared to summer burning; therefore, the prescribed winter burns that
current management implemented likely benefit the herpetofauna observed in the
grassland on Matagorda Island. However, Fair and Henke (1997b) propose that high
intensity winter burns might have a negative effect on horned lizards that hibernate near
the soil surface. My study supports the findings by Burrow et al. (2001) that little cover
was available to protect P. cornutum from predators, and the vegetation might not
provide enough structure for thermoregulative behaviors.
The species richness I observed was similar to that of Prieto (1995). However,
my trapping methods focused on P. cornutum, and were not appropriate for trapping all
grassland herpetofauna. In addition to the species I captured, Prieto (1995) also captured
Texas brown snake (Storeria dekayi texana), ground skink (Scincella lateralis), ornate

18
box turtle (Terrapene ornata), yellow mud turtle (Kinosternon flavescens), red-eared
slider (Trachemys scripta elegans), Eastern narrowmouth toad (Gastrophryne
carolinensis), and Gulf Coast toad (Ollotis nebulifer). I observed S. lateralis, T. ornata,
and T. scripta elegans along the road-side, and the carapace and plastron of K. flavescens,
but I did not observe the other species. Prieto (1995) suggested that some of these
species are seasonally abundant, which might explain their absence from my observation.
Road cruising produced the highest capture rate and most salient information, and
was consistent with the study by Fair and Henke (1997a). I only observed one P.
cornutum mortality along the road. Along with the possibility of thermoregulation, the
road might be used as a corridor to more optimal foraging areas, better nesting habitat, or
more protective cover. The fact that P. cornutum is found in two disjunct populations
separated by Burn 1 habitat is noteworthy. The present data suggested that if burning
was prescribed without consideration for distribution and dispersal of P. cornutum the
species could become locally extinct. Phrynosoma cornutum is not as vagile as other
reptiles on the island, and has longer recolonization time than other lizard species on the
island (Fair and Henke, 1999).
In 2006 and 2007, the Aransas NWR staff conducted P. cornutum surveys via
road cruising (Chad Stinson, Aransas NWR, pers. comm.). They estimated one P.
cornutum every 8.05 km for both years. These surveys, however, did not consider
recaptures; therefore, the 2006 and 2007 estimates are likely inflated. Also, during the
2006 survey, Aransas patrol teams spent approximately 48 man hours in the field actively
searching on foot for P. cornutum, but none were observed. Because of the cryptic
coloration of P. cornutum, there is the potential for observer bias on transect walks, and I

19
accounted for this bias by placing drift-fence arrays within the habitats. Nonetheless, the
results in 2006 and 2007 were consistent with the current findings in which walking
transects and passive drift-fence arrays yielded no or few observations, and road surveys
produced the most observations.
There are several potential reasons for low capture rate, including: 1) the traps
were not effective for capturing P. cornutum, 2) I sampled the wrong habitat types, or 3)
the species is rare within these habitats. Trap openings were large enough to
accommodate the cephalic ornamentation of P. cornutum, and on one occasion I
witnessed the behavior associated with a capture. The individual did not appear hesitant
to move into the funnel trap. With regard to habitat types, undoubtedly, there is finer
scale variation within the habitat types, but this initial investigation focused on providing
insight into management practices, resulting in my choice of habitat type. Finally, given
the relative amount of effort and diversity of methods expended to capture the
herpetofauna and P. cornutum in particular, it seems reasonable to conclude that P.
cornutum is uncommon in these habitats and perhaps rare on the island. Although drift
fences resulted in a number of captures and provided a reasonably unbiased method of
sampling the herpetofauna, the overall capture rate was low, the cost in human resources
was high, and mortality rates for all species were high due to depredation by S. invicta.
Accordingly, while this method might be justified periodically, it is not an effective
means of annual monitoring particularly in the case of P. cornutum.
These data suggested a high association between a number of reptiles, including
P. cornutum, with the Road-side and adjacent habitat. Most species exhibit a positive
association with the increase in vegetative structure observed in Burn 2, Burn 3, and

20
Road-side habitats. P. cornutum exhibited a more complex pattern, perhaps because
more data were available for analysis, but nonetheless the species exhibited a strong
negative association with both Burn 1 and Burn 3 habitats. Based on these data, a
reevaluation of habitat manipulation strategies might be necessary. Certainly,
fragmenting the grassland habitat with uniform burn units from dunes to salt marsh could
be detrimental to a species with low vagility and relatively low recolonization rates.
A system of burn unit where units run parallel with the coast in narrow strips of
two to three units would be optimal. This would allow P. cornutum to avoid fire or
indirect factors with negative effects and provide a corridor for recolonization. The
positive association with Burn 2 treatments was further reinforced by the apparent
mitigating effect Burn 2 has when paired with Burn 1 and Burn 3 (Figure 8).
Alternatively, the present conclusions might be based on observations at a scale
too large to determine the actual factors influencing the distribution and abundance of P.
cornutum. Within the 50% isopleths (Figure 7) there are clumped observations. A finer-
grained investigation might reveal more specific associations that are only indirectly
associated with fire regime. For example, P. cornutum might be associated with a
specific plant or invertebrate community that is only in part influenced by fire regime.
Vegetation response to burning can be slow to recover (Collins, 1992; Chang, 1996; Van
Dyke et al., 2007), which influences the recruitment of the arthropod community (Benson
et al., 2007; Siemann et al., 1998). The distribution of food resources (harvester ants) and
nest predators (fire ants) might be more influential. Populations throughout the range of
this species have been declining primarily due to habitat loss, the introduction of S.
invicta, and the use of pesticides (Donaldson et al., 1994). Accordingly, future research

21
at a finer scale seems prudent prior to the consideration of a full-scale modification of a
landscape level management plan.
LITERATURE CITED
BAILLIE, J. E. M., C. HILTON-TAYLOR, AND S. N. STUART (editors). 2004. 2004 IUCN red
list of threatened species: a global species assessment. IUCN, Gland, Switzerland
and Cambridge, UK.
BENSON, T. J., J. J. DINSMORE, AND W. L. HOHMAN. 2007. Responses of plants and
arthropods to burning and disking of riparian habitats. Journal of Wildlife
Management 71: 1949-1957.
BEYER, H. L. 2004. Hawth's Analysis Tools for ArcGIS (version 3.27). Available at
http://www.spatialecology.com/htools.
BRAGG, T. B. AND L. HULBERT. 1976. Woody plant invasion of unburned Kansas
bluestem prairie. Journal of Range Management 29:19-23.
BURROW, A. L., R. KAZMAIER, E. HELLGREN, AND D. RUTHVEN, III. 2001. Microhabitat
selection by Texas horned lizards in southern Texas. Journal of Wildlife Management
65:645-652.
BUTCHART, S. H. M. 2008. Red List indices to measure the sustainability of species use
and impacts of invasive alien species. Bird Conservation International 18:245-262.
CAVITT, J. F. 2000. Fire and a tallgrass prairie reptile community: effects on relative
abundance and seasonal activity. Journal of Herpetology 34:12-20.

22
CHANG, C. 1996. Ecosystem responses to fire and variations in fire regimes. In Sierra
Nevada Ecosystem Project: Final report to Congress Assessments and scientific basis
for management options. Davis: University of California, Centers for Water and
Wildland Resources.
COGGINS, G. C. 1974. Conserving wildlife resources: An overview of the endangered
species act of 1973. North Dakota Law Review 51:315-340.
COLLINS, S. L. 1992. Fire frequency and community heterogeneity in tallgrass prairie
vegetation. Ecology 73:2001-2006.
CONOVER, W.J. 1980. Practical Nonparametric Statistics, second edition. John Wiley &
Sons, New York.
COWLES, R. B. AND C. M. BOGERT. 1944. A preliminary study of the thermal
requirements of desert reptiles. Bulletin of the American Museum of Natural History
83:261-269.
DECKER, D. J., T. L. BROWN, AND W. F. SIEMER. 2001. Evolution of people-wildlife
relations. Pages 3-22 in Human dimensions of wildlife management in North
America (D. J. Decker, T. L. Brown, and W. F. Siemer editors). The Wildlife
Society, Bethesda, Maryland.
DONALDSON, W., A. H. PRICE, AND J. MORSE. 1994. The current status and future
prospects of the Texas horned lizard (Phrynosoma cornutum) in Texas. Texas
Journal of Science 46:97-113.

23
DURBIAN, F. E. 2005. Effects of mowing and summer burning on the massasauga
(Sistrurus catenatus). American Midland Naturalist 155:329-334.
FAIR, W. S. AND S. HENKE. 1997a. Efficacy of capture methods for a low density
population of Phrynosoma cornutum. Herpetological Review 28:135-137.
FAIR, W. S. AND S. HENKE. 1997b. Effects of habitat manipulations on Texas horned
lizards and their prey. The Journal of Wildlife Management 61:1366-1370.
FAIR, W. S. AND S. HENKE. 1999. Movements, home ranges, and survival of Texas
horned lizards (Phrynosoma cornutum). Journal of Herpetology 33:517-525.
FUHLENDORF, S. D., W. C. HARRELL, D. M. ENGLE, R. G. HAMILTON, C. A. DAVIS, AND D.
M. LESLIE, JR. 2006. Should heterogeneity be the basis for conservation?
Grassland bird response to fire and grazing. Ecological Applications 16:1706-
1716.
GIBBONS, L. W., D. E. SCOTT, T. J. RYAN, K. A. BUHLMANN, T. D. TUBERVILLE, F. S.
METTS, J. L. GREENE, T. MILLS, Y. LEIDEN, S. POPPY, AND C. T. WINNE. 2000.
The global decline of reptiles, Déjà vu amphibians. Bioscience 50:653-666.
HENKE, S. E. AND M. MONTEMAYOR. 1998. Diel and monthly variations in capture
success of Phrynosoma cornutum via road cruising in Southern Texas.
Herpetological Review 29:148-150.
HILTON-TAYLOR, C. 2007. Species Extinction. IUCN Species Survival Commission.
Gland, Switzerland.

24
HUEY, R. B. AND R. D. STEVENSON. 1979. Integrating thermal physiology and ecology
of ectotherms: A discussion of approaches. American Zoologist 19: 357-366.
KAUFMAN, D. W., E. J. FINCK, AND G. A. KAUFMAN. 1990. Small mammals and
grassland fires. In S. L. Collins and L. L. Wallace (eds.), Fire in North American
Tallgrass Prairies, pp. 46-80. University of Oklahoma Press, Norman.
LANDRES, P.B., P. MORGAN, AND F. J. SWANSON. 1999. Overview of the use of natural
variability concepts in managing ecological systems. Ecological Applications, 9:
1179-1188.
LANE, J. S. 2008. Assessing diet and daily movement of Texas horned lizards
(Phrynosoma cornutum) in Southern Texas. Unpublished Thesis. West Texas A&M
University.
LEHMAN, R. L., R. O’BRIEN, AND T. WHITE. 2005. Plants of the Texas Coastal Bend.
Texas A&M University Press, College Station, TX.
MCALISTER, W. H. 2004. Life on Matagorda Island. Texas A&M University Press,
Texas.
MCALISTER, W. H. AND M. MCALISTER. 2006. Guidebook to the Aransas National
Wildlife Refuge. Mince Country Press, Victoria, Texas.
MCCARTHY, M. A., C. THOMPSON, S. GARNETT. 2008. Optimal investment in
conservation of species. Journal of Applied Ecology 45:1428-1435.

25
MOELLER, B. A. 2000. Conservation ecology of the Texas horned lizard (Phrynosoma
cornutum): Comparative effects of summer and winter burning. Unpublished Thesis,
Auburn University, Alabama.
MOELLER, B. A., E. C. HELLGREN, D. C. RUTHVEN, R. T. KAZMAIER, AND D. R.
SYNATZSKE. 2005. Temporal differences in activity patterns of male and female
Texas horned lizards (Phrynosoma cornutum) in southern Texas. Journal of
Herpetology 39:336-339.
NICHOLS, R. AND J. MENKE. 1984. Effects of chaparral shrubland fire on terrestrial
wildlife. In Shrublands in California: Literature review and research needed for
management, edited by J. J. DeVries, 74-96. Contribution 191. Davis: University of
California, Davis, Water Resources Center.
PRICE, A. H. 1990. Phrynosoma cornutum (Harlan) Texas horned lizard. Catalogue of
American Amphibians and Reptiles 469:1-7.
PRIETO, A. A., JR. AND W. WHITFORD. 1971. Physiological responses to temperature in
horned lizards, Phrynosoma cornutum and Phrynosoma douflassii. Copeia 1971:498-
504.
PRIETO, F. G. 1995. Terrestrial small vertebrate response to seasonal prescribed burns on
Matagorda Island - A Texas coastal barrier island. Unpublished data.
RICHARDSON, A. 2002. Wildflowers and other plants of Texas beaches and islands.
University of Texas Press, Austin, TX.

26
RODGERS, A. R. AND A. P. CARR. 2002. Home range extensions. Ontario Ministry of
Natural Resources’ Centre fro Northern Forest Ecosystem Research, Thunder Bay,
Ontario, Canada.
RUSSELL, T. A. 1997. Thermal ecology of the Texas horned lizard (Phrynosoma
cornutum). Unpublished Thesis, University of Central Oklahoma, Oklahoma.
SHARROW, S. H. 1984. A simple disc meter for measurement of pasture height and
forage bulk. Journal of Range Management 37:94-95.
SIEMANN, E., D. TILMAN, J. HAARSTAD, AND M. RITCHIE. 1998. Experimental tests of
the dependence of arthropod diversity on plant diversity. The American Naturalist
152:738-750.
STARK, R. C., S. FOX, AND D. M. LESLIE, JR. 2005. Male Texas horned lizards increase
daily movements and area covered in spring: A mate searching strategy? Journal of
Herpetology 39:169-173.
VAN DYKE, F., J. SCHMELING, S. STARKENBURG, S. HEUN YOO, AND P. STEWART. 2007.
Responses of plant and bird communities to prescribed burning in tallgrass prairies.
Biodiversity and Conservation 16:827-839.
WHITFORD, W. G. AND M. BRYANT. 1979. Behavior of a predator and its prey: the
horned lizard (Phrynosoma cornutum) and harvester ants (Pogonomyrmex spp.).
Ecology 60:686-694.

27
Run
way
6 1 4 - - - - - - - 1 3 1 - - - -
Roa
d 1 7 3 3 7 2 3 - 1 1 - - 9 5 - - 1 -
Bur
n 3
2 7 5 3 6 2 1 2 - - 1 - 6 1 - - - -
Bur
n 2
2 17
4 1 2 1 3 1 1 - - - 13
3 - - 1 -
Bur
n 1
11
- 1 - - - - - - - - 3 2 2 - - -
Dun
e
7 3 3 - 3 - 1 1 1 - - 8 - - 1 - 61
Tot
al #
5 55
16
15
15
8 7 4 3 2 1 1 42
12 2 1 2 61
Scie
ntifi
c N
ame
Lith
obat
es sp
heno
ceph
alus
Aspi
dosc
elis
sexl
inea
ta
Oph
isau
rus a
ttenu
atus
C
olub
er fl
agel
lum
Th
amno
phis
pro
xim
us
Sist
ruru
s cat
enat
us
Agki
stro
don
pisc
ivor
us
Cro
talu
s atr
ox
Phry
noso
ma
corn
utum
La
mpr
opel
tis g
etul
a ho
lbro
oki
Lam
prop
eltis
tria
ngul
um
Het
erod
on p
latir
hino
s
Sigm
odon
his
pidu
s Re
ithro
dont
omys
fulv
esce
ns
Col
inus
vir
gini
anus
Gec
arci
nus s
p.
Sesa
rma
sp.
Ocy
pode
sp.
Com
mon
Nam
e A
mph
ibia
ns
So
uthe
rn L
eopa
rd F
rog
Rep
tiles
Six-
lined
Rac
erun
ner
Sl
ende
r Gla
ss L
izar
d
Coa
chw
hip
*G
ulf C
oast
Rib
bon
Snak
e
Mas
sass
agua
Cot
tonm
outh
Wes
tern
Dia
mon
dbac
k
Texa
s Hor
ned
Liza
rd
Sp
eckl
ed K
ings
nake
Loui
sian
a M
ilk S
nake
East
ern
Hog
nose
Sna
ke
Mam
mal
s
Cot
ton
Rat
Fulv
ous H
arve
st M
ouse
A
ves
N
orth
ern
Bob
whi
te
Inve
rteb
rate
s
Land
Cra
b
Smal
l Lan
d C
rab
G
host
Cra
b
TAB
LE 1
—A
bund
ance
of s
peci
es c
aptu
red
in d
rift-f
ence
arr
ays w
ithin
spec
ified
hab
itat t
ypes
from
June
-Aug
ust o
n M
atag
orda
Isla
nd, T
exas
. Sig
nific
ant G
-test
resu
lts a
re d
enot
ed w
ith a
n as
teris
k (G
= 2
4.02
8, d
f = 5
, p <
0.0
01).

28
Snout-Vent Length (mm)
Total Length (mm)
Body Mass (g)
N Range Mean Range Mean Range Mean
Female 25 66.0 - 92.5 82.4
98.0 - 132.0 120.1
37.0 - 72.0 35.8
Male 12 58.0 - 82.0 72.8
92.0 - 129.0 112.1
32.0 - 53.0 27.2
Juvenile 4 29.0 – 59.0 43.0
42.0 – 90.0 64.0
1.25 – 14.0 24.8
Total 41 29.0 - 92.5 75.7 42.0 - 136.0 112.3 15.3 - 72.0 48.1
TABLE 2—Range and mean snout-vent length, total length, and body mass of males,
females, and juveniles by road captures of P. cornutum from June-August 2008 on
Matagorda Island, Texas.

29
Adjacent Habitats Percent Expected Observed
Burn1-Burn1 11.8 5.882 0
Burn1-Burn2 5.9 2.941 2
Burn1-Burn3 9.8 4.902 1
Burn2-Burn2 15.7 7.843 14
Burn2-Burn3 33.3 16.667 26
Burn3-Burn3 23.5 11.765 7
21.279χ2calc>11.070χ2
crit
TABLE 3—Chi-square analysis of road captures of P. cornutum showing a
significant difference in habitat use in adjacency to three types of burn regimes
within the 50% isopleths from June-August on Matagorda Island, TX. Percent
is the percent of available adjacent habitat combinations.

30
FIG. 1—Design of a Y-shaped drift-fence
array.
120°
120° 120°
Funnel Trap
I I I
, I
, ,

31
FIG. 2—Design of a drift-fence array for
Road-side habitat.
M
I
D
D
L
E
R
O
A
D
50 m
50 m
-~---------- ------} --------~----------
------------ } --------- . --------

32
Fig. 3—Variation in mean percent canopy cover among six habitat types during June and
July 2008 on Matagorda Island, Texas. A Kruskal-Wallis followed by a multiple
comparisons test detected significant differences among groups of habitats (H = 223.36,
df = 5, p < 0.001). Habitat types that share a letter designation are not significantly
different.
76.0 31.0 97.3 94.9 92.7 46.20
102030405060708090
100
Dune Burn 1 Burn 2 Burn 3 Road Runway
PER
CE
NT
GR
OU
ND
CO
VE
R
HABITAT TYPE
B
A
CCC
A

33
Fig. 4—Variation in mean vegetation compression index (cm) among six habitat types
during June and July 2008 on Matagorda Island, Texas. A Kruskal-Wallis followed by a
multiple comparisons test detected significant difference among groups of habitats (H =
183.32, df = 5, p < 0.001). Habitat types that share a letter designation are not
significantly different.
13.35 9.27 26.21 31.25 18.85 10.480
5
10
15
20
25
30
35
Dune Burn 1 Burn 2 Burn 3 Road Runway
ME
AN
CO
MPR
ESS
ION
(cm
)
HABITAT TYPE
ABA
AB
C
C
A

34
Fig. 5—Variation in mean soil compaction at 7.6 cm (psi) among six habitat types during
June and July 2008 on Matagorda Island, Texas. A Kruskal-Wallis followed by a
multiple comparisons test detected significant differences among groups of habitats (H =
149.84, df = 5, p < 0.001). Habitat types that share a letter designation are not
significantly different.
31.55 43.33 102.71 79.38 122.43 178.210
20
40
60
80
100
120
140
160
180
200
Dune Burn 1 Burn 2 Burn3 Road Runway
ME
AN
SO
IL C
OM
PAC
TIO
N (p
si)
HABITAT TYPE
A
C
BC
BBC
A

35
Fig. 6—Mean species richness among six habitat types during June and July 2008 on
Matagorda Island, Texas. A Kruskal-Wallis followed by a multiple comparisons test
detected significant differences among groups of habitats (H = 13.800, df = 5, p = 0.032).
Habitat types that share a letter designation are not significantly different.
1.25 0.58 1.67 1.5 1.92 0.670
0.5
1
1.5
2
2.5
Dune Burn 1 Burn 2 Burn 3 Road Runway
ME
AN
SPE
CIE
S R
ICH
NE
SS
HABITAT TYPE
AB
A
C
BCBC
A

36
FIG. 7--Road captures of P.cornutum from June through July on Matagorda Island, Texas. Chi-square test of independence detected a nonrandom distribution, and a 50% isopleth outline the distributions.
N
1,egend 0 }dalllf,O{lla \s~d B(lllll.daIY o nurnunits - }&(Idle itoad - 1l\Jllv,,a.y Cl capture;
- nurn 1 'ldlf pooe - Bllfl\ 2 '{dlfS prlOC Ollfl\ 3 oc }MJCC 'l~ pooc .~-- CJ so¾ ,sap\dh

37
Fig. 8—Differences among observed numbers of P. cornutum and expected numbers
within adjacent burn-unit combinations along Middle Road on Matagorda Island, Texas
(χ2 = 21.279, df = 5, p < 0.05). The thick line along the zero axis is the calculated
expected value for a random distribution among habitat types. The dashed line along the
x-axis represents middle road and the combination of adjacent habitat types.
-5.9 -0.9 -3.9
9.3 9.3
-4.8
-8.0
-6.0
-4.0
-2.0
0.0
2.0
4.0
6.0
8.0
10.0
12.0
Burn 1
Burn 1
Burn 1
Burn 2
Burn 1
Burn 3
Burn 2
Burn2
Burn 2
Burn 3
Burn 3
Burn 3
OB
SER
VE
D -
EX
PEC
TE
D V
AL
UE
S
ADJACENT BURN UNITS

38
Fig. 9—Catch per unit effort of P. cornutum from June through August on Matagorda
Island, Texas. Effort types are measured in hours with the exception of Trap Night, and
are defined as the following: Cruise is actively searching from a vehicle at 0-24 kph,
Vehicle is actively searching at 24-48 kph, Passive Observations is not actively searching
from >48 kph, Walking is actively searching while walking from the road to the drift-
fence arrays, Transect is actively searching while walking in a random direction for 50 m.
Trap Night is the number of P. cornutum captured over the total number of traps opened
throughout the study per night.
0.3963
0.2429
0.1481
0.0231 0.0000 0.00140.00
0.05
0.10
0.15
0.20
0.25
0.30
0.35
0.40
0.45
0.50
Cruise(hr)
Vehicle(hr)
Pass. Obs.(hr)
Walking(hr)
Transect(hr)
Trap Night(# of open
traps)
CAT
CH
/ E
FFO
RT
EFFORT TYPE

39
CHAPTER 2
MICROHABITAT ASSOCIATIONS OF TEXAS HORNED LIZARD (PHRYNOSOMA
CORNUTUM) WITH INFERENCE FROM POWDER TRACKING
Keywords: fluorescent powder tracking, Formicidae, landscape ecology, microhabitat,
Phrynosoma cornutum, reptile movement
INTRODUCTION
Environments are patchy relative to scale, and this patchiness strongly influences
the distribution of organisms, their interactions, and their adaptations (Wiens, 1976).
Large heterogeneous habitats tend to support more species (Collins, 1992; Turner et al.,
2001; Fuhlendorf et al., 2006). Larger patches contain local environmental variability,
such as microclimate differences and plant structure, while smaller patches have a greater
perimeter-to-area ratio (Turner et al., 2001). Human disturbance often simplify these
perimeters from complex shapes to straight lines (Krummel et al., 1987; Turner et al.,
2001). The spatial distribution of resources in these heterogeneous landscapes can affect
the growth, reproduction, and dispersal of organisms (Hellgren et al., 2010; Turner et al.,
2001). However, animal movements and foraging patterns also can affect spatial
heterogeneity (Turner et al., 2001).
Within a heterogeneous landscape, the potential exists for separate populations to
be linked by immigration, also referred to as metapopulations (Levins, 1969). Organisms
need suitable habitat for movement from areas where natality exceeds mortality (sources)
to areas where mortality exceeds natality (sinks) (Pulliam, 1988). The connectivity of

40
suitable habitat often can be a limiting factor in the spatial distribution of a population
rendering some areas accessible and others inaccessible (Turner et al., 2001).
Characterizing suitable habitat for food resources, thermoregulation, shelter, and
reproduction within the heterogeneous landscape is necessary to determine if it is
spatially connected relative to unsuitable habitat. Yet, a suitable habitat patch for one
organism might not be suitable for another organism making heterogeneity both scale and
species specific (Turner et al., 2001). Although the vagility of a species influences
dispersal distances, the distance and recolonization rates can be limited by the size and
connectivity of these patches (Turner et al., 2001). Therefore, corridors, narrow patches
of habitat that connect similar patches but differ from the surrounding matrix, can benefit
the dispersal of organisms (Turner et al., 2001).
Disturbance is another process that affects landscape heterogeneity and might be
essential to the maintenance of community structure and ecosystem function (Turner et
al., 2001). Similar to heterogeneity, disturbance is a scale dependent process that might
be disruptive at one level, yet stabilizing at another level (Turner et al., 2001). White and
Pickett (1985) define a disturbance as a relatively discrete event that disrupts the structure
of an ecosystem, community, or population and changes resource availability or the
physical environment. In grasslands, fire is a natural process and source of heterogeneity
that maintain the landscape. This disturbance alters resource abundance, which can
influence the movement to and recolonization of an area.
In certain environments, spatial heterogeneity can limit the extent and severity of
a disturbance and increase the rate of recovery by providing refuges for populations that

41
will recolonize the disturbed area (Risser et al., 1984). Disturbances that occur within a
particular cover type might be slowed by increased landscape heterogeneity; however, a
disturbance that spreads among cover types or is enhanced by edge effects will be
augmented by increased landscape heterogeneity (Turner et al., 2001). The number of
residual species that remain after the disturbance will influence recovery. Even if the
number of residuals is low, the number of propagules input from the surrounding matrix
can support recovery. However, the density of popagule input decreases as disturbance
patch size increases (Turner et al., 2001).
Understanding how organisms move through heterogeneous landscapes can
benefit management and conservation strategies. Currently, the most common techniques
of tracking animal movement are direct observation, capture-mark-recapture, thread
trailing, radiotelemetry, and fluorescent powder tracking (Blankenship et al., 1990). The
oldest technique for studying movement is by direct observation. The advantages of this
technique are that handling the organism is minimized, movements are not impeded by
equipment, and direct observations are made on where individuals are located and their
behaviors (Sanderson, 1966). However, direct observations are time consuming and few
individuals can be studied at one time (Sanderson, 1966). This technique is difficult with
small or cryptic organisms, especially in dense vegetation (Fellers and Drost, 1989).
Additionally, the proximity of the observer might influence the movement of the
organism, which contradicts the ultimate objective.
Indirect observation techniques compensate for the time commitment of one
observer for very few individuals, but increases the handling time of the organism and

42
might affect movement. Capture-mark-recapture techniques have been used to study
recruitment (Pradel, 1996) and estimate home ranges (Lemen and Freeman, 1985), but
can be labor intensive with low capture success in areas of low abundance (Fisher and
Muth, 1995; Eggert, 2002; Inslee, Chapter 1). In general, capture-mark-recapture
techniques yield coarse movement information (Lemen and Freeman, 1985).
The thread trailing technique is less labor intensive than the capture-mark-
recapture technique, and produces more accurate estimates of individual movement
information (Mendel and Vieira, 2003). Several variations of this technique have been
employed with the primary disadvantages being 1) mechanical failure (Heath, 1961;
Mendel and Vieira, 2003), 2) the weight and bulk of the apparatus can hinder movement
(Wilson, 1994), 3) tracking is limited by the length of the thread (Anderson et al., 1988;
Blankenship et al., 1990), and 4) fabricating a unique design for each organism (Heath,
1961; Anderson et al., 1988; Mendel and Vieira, 2003).
Radiotelemetry is a point-data technique that is commonly used across taxa to
study animal movements, especially since the size and mass of the transmitters have
decreased with improved technology (Reinert, 1984; Blankenship et al., 1990; Fisher and
Muth, 1995, Eggert, 2002). The battery life and size of transmitter can be a limiting
factor in the length of the study and the size of the target species, respectively (Fellers
and Drost, 1989; Eggert, 2002). The temporal precision of radiotelemetry data also is
directly related to the number of times the animal is relocated; therefore, it can be time-
consuming (Eggert, 2002). Although this technique is commonly used in microhabitat

43
studies (Reinert, 1984), there can be potentially valuable information that lie between the
relocation points (Eggert, 2002).
A primary limitation of the previously mentioned techniques is that they do not
give 24-hour trails that are precise in the absence of an observer (Sanderson, 1966). The
aforementioned techniques also can be expensive and require substantial time
commitments (Sanderson, 1966; Fellers and Drost, 1989; Blankenship et al., 1990). As
an alternative, I used powder tracking that provides accurate spatial movement (Fellers
and Drost, 1989; Eggert, 2002); precise estimates of edge effect, corridor use, and
foraging activity (Eggert, 2002); is inexpensive (Fellers and Drost, 1989; Blankenship et
al., 1990); and can be used on small animals (Lemen and Freeman, 1985; Eggert, 2002).
In addition, powder tracking allows for vertical habitat use to be quantified (Fellers and
Drost, 1989; Eggert, 2002). The fluorescent powder is resistant to degradation and works
well in diverse habitats including dense vegetation, bare ground, and is even detectible in
wet environments (Halfpenny, 1992; Eggert, 2002). However, this can limit the length of
a study because individuals that stay in the same place lay overlapping trails that can be
difficult to interpret (Stark and Fox, 2000, Inslee, pers. obs.). Although this is a non-
invasive procedure, the handling time can cause stress for the organism that might affect
behavior (Dodd, 1992).
In the past, horned lizards (Phrynosoma sp.) were placed into bags and shaken
with powder to coat the entire body with the fluorescent dye. This can be hazardous to
individuals by reducing their cryptic coloration, which places them at higher risk of
predation (Fellers and Drost, 1989). P. cornutum scales also do not offer enough surface

44
area to hold the dye and release it over a 24-hour period (Fellers and Drost, 1989,
Blankenship et al., 1990). The Powdered Fur Patch (PFP) method was developed to
reduce these limitations (Lane, 2008), and effectively track P. cornutum throughout their
habitat.
Texas horned lizards (Phrynosoma cornutum) require a matrix of bare ground and
herbaceous vegetative structure to thermoregulate within their optimal range (Russell,
1997; Burrow et al., 2001). The upland area of Matagorda Island was covered in coastal
tall grass prairie, and the primary disturbance on the island that alters vegetative structure
was prescribed burning (Inslee, Chapter 1). Burn patches of different ages on Matagorda
Island exhibited differences in standing stem density, canopy cover, and soil compaction
(Inslee, Chapter 1). On Matagorda Island, the grassland landscape was divided into 40
block-shaped burn units ranging from 92-558 ha. Burn patches and Middle Road, which
bisects the length of the island, introduced simplified edges into a naturally complex
system of habitat structure. The landscape of Matagorda Island was disrupted by Middle
Road, and Inslee (Chapter 1) suggests that it might influence dispersal of organisms and
habitat use.
Habitat fragmentation, one of the primary threats to populations of P. cornutum,
focuses investigations on the amount of suitable habitat and its isolation from other
suitable patches (Donaldson et al., 1994; Turner et al., 2001). In 2008, Inslee (Chapter 1)
identified two disjunct populations of P. cornutum, one on each end of Matagorda Island.
Results from Inslee (Chapter 1) suggested that areas burned during the previous or
current year were unsuitable habitats for P. cornutum; however, they did not explain the

45
spatial distribution of individuals. Spatial patterns that also might influence P. cornutum
include road structure and networks (Fair and Henke, 1997a; Henke and Montemayor,
1998, Sherbrooke, 2002), vegetation (Burrow et al., 2001; Hellgren et al., 2010), and the
distribution and relative abundance of ants (Munger, 1984).
Patterns that occur at scales other than the burn patch level, such as the
distribution of ants (Moeller, 2000), might affect the spatial distribution of P. cornutum.
The red imported fire ant (Solenopsis invicta) was first observed on Matagorda Island in
the early 1990s (Chad Stinson, pers. comm.). Invasions of S. invicta have significant
negative ecological impacts on invertebrates, birds, mammals, and herpetofaunal
populations (Wojcik et al., 2001; Allen et al., 2004). Indirect impacts might include
reduced survival and growth rate, behavior changes, changes in foraging pattern, reduced
food availability, and changes in habitat use (Allen et al., 2004). Native ant richness and
abundance can be negatively impacted by S. invicta invasions. In central Texas,
Morrison (2002) observed that 12 years after an initial S. invicta invasion, ant diversity
returned to pre-invasion levels, but experienced lower abundance and the loss of some
native ant species.
The objectives of this study were to 1) characterize habitat structure relative to a
fire disturbance, and a major landscape feature, Middle Road (Inslee, Chapter 1), 2)
quantify suitable microhabitat relative to P. cornutum by analyzing their movements
through the landscape, 3) document the distribution of genera of Formicidae and P.
cornutum throughout Matagorda Island, 4) gain insights into the influence of S. invicta on

46
the distribution of P. cornutum on Matagorda Island, and 5) provide an evaluation on
current management practices relative to the conservation of P. cornutum.
METHODS
Habitat
I conducted a systematic habitat survey along the length of Middle Road on
Matagorda Island to quantify available microhabitat. I used transects to quantify habitat
structure and ground cover at 1-km intervals along the entire length of Middle Road
(Figure 1); 52 transects total. Transects were oriented perpendicular to the road and
extended 60 m on each side of the road. Measurements were made at intervals of 0 m
(road-side vegetation), 5 m, 10 m, 15 m, 20 m, 30 m, 40 m, 50 m, and 60 m from the road
edge. I used a Daubenmire frame to examine percentage ground cover and identified
percentage of the predetermined ground-cover types (forb, grass, bare ground, prostrate
shrub, shrub, water, and litter). Thatch depth (cm) and soil compaction (DICKEY-john
Corporation, Auburn, IL) were estimated. A compression disk meter (Sharrow, 1984)
was used to generate an index of stem density. I also recorded the year from last burn
(Burn 1, Burn 2, and Burn 3) (Inslee, Chapter 1).
Tracking
Phrynosoma cornutum were captured from May through the beginning of August
in 2009. Road cruising was used to capture P. cornutum by using the same methods as
2008 (Inslee, Chapter 1). However, P. cornutum with a snout-vent length <40 mm were
considered juveniles and their sex could not be determined with certainty. Middle Road

47
and the north runway complex on Matagorda Island were systematically surveyed and
equal sampling effort was maintained. The time of day was separated into two sampling
periods: sunrise to 1200h and 1600h to sunset to coincide with greatest activity periods.
When P. cornutum were captured, the date, time, snout-vent length, total length, mass,
and sex were recorded, and a toe clip was collected from the third digit of the rear, left
foot to mark individuals. To assess distribution pattern of P. cornutum on Matagorda
Island, I used road capture locations to construct a 50% isopleth by using Home Range
Tools (Rodgers and Carr, 2002) with GIS ArcMap 9.2 (ESRI, Redlands, CA) (Inslee,
Chapter 1).
I used the PFP method for tracking the movement of individuals over a 24-hr
period (Lane, 2008). After processing the lizard, we would cut a piece of rabbit fur to fit
the ventral surface of the organism without hindering appendage movement, obstructing
the cloaca, and without being visible from the dorsal view. The fur patch was adhered to
the ventral scales with a few drops of cyanoacrelate gel adhesive (Superglue Gel; Henkel
Consumer Adhesives, Inc., Avon, Ohio, USA). A shop towel with a hole in it the size of
the fur patch was wrapped around the lizard only exposing the fur patch. This confined
the powder only to the fur patch. I then held the lizard upside-down in the palm and used
a small paint brush to coat the fur patch with a fluorescent powder dye (Redwop Powder,
Lightning Powder Company, Inc., Jacksonville, FL). The fur was parted back every few
millimeters and the powder dye was coated within the partition throughout the patch.
Any hairs from the fur patch that were visible from the dorsal side of the lizard were
trimmed with scissors. The lizard was then released at the site of capture.

48
The evening of the capture and release day, P. cornutum were relocated by using
a high-intensity ultraviolet light. The powder trail highlighted the path P. cornutum took
throughout the day and was marked with fiberglass stakes every 5 meters. Coordinates
also were recorded for the beginning and end points of the powder trail. When the lizard
was relocated on the first night, powder was reapplied to the fur patch, and the lizard was
placed back in its resting position. Typically, the horned lizard would remain inactive
throughout this process, but if it ran a distance from the original resting area we would
begin tracking from the new resting place. If the lizard was relocated a second night,
then the fur patch was carefully removed.
The day after all tracking was completed a habitat survey was conducted. The
same habitat measurements as the systematic survey were recorded every 5 m along the
powder trail (Stark et al., 2005). I also measured the absolute distance the lizard moved
(cm) (actual distance) and relative distance the lizard moved (cm) (straight-line distance
between beginning and end point).
Ants
Along every fourth habitat transect, I surveyed ant mounds. This survey entailed
cutting a half meter swath of vegetation along the transect line and systematically
searching for ant mounds up to a quarter of a meter on either side of the transect. When
we found an ant mound, we recorded the GPS coordinates and collected approximately
10 individuals with the aid of an aspirator. Ants were stored in 5 ml plastic transport
tubes and frozen as soon as possible. Subsequently, ants were sorted from soil and
vegetation and stored in tubes to be identified at a later time. Ants were identified to

49
genus with the aid of Fisher and Cover (2007) and an Olympus SZX16 stereomicroscope.
A subset of voucher specimens was sent to Dr. Chad Tillberg, Linfield College, to be
verified. Another subset of voucher specimens was catalogued at the Sternberg Museum
of Natural History.
Statistical Analysis
A Kruskal-Wallis test followed by a test of multiple comparisons (Conover, 1980)
was used to evaluate differences among burn treatments. Similar to 2008, percentage of
canopy cover, compression index, and soil compaction at a 7.6 cm depth were used to
characterize burn treatments (Inslee, Chapter 1).
I used mark-recapture data to estimate population size by using the Schnabel
index. This index assumes no births or deaths and a closed population during the
sampling period. Only adult captures were used in this analysis. I used a chi-square
analysis to assess if P. cornutum were randomly distributed on the island or if there was a
preference for a specific burn treatment (Inslee, Chapter 1).
A discriminant function analysis (DFA) predicts group membership from a set of
variables (Tabachnick and Fidell, 2007). I used a DFA to identify similarities of habitat
characteristics from the systematic habitat survey among nine distance intervals from
Middle Road. The nine distances intervals were used as groups from each of the 52
transects. This allowed me to discern differences in habitat structure at distances from
Middle Road and along the entire length of Matagorda Island.
I used a logistic regression to predict the presence or absence of P. cornutum
based on habitat variables measured in both systematic surveys and tracked surveys

50
(Tabachnick and Fidell, 2007). By using the presences of tracked habitat data, the
logistic regression predicted what habitat variables, or microhabitat, P. cornutum were
most likely to be associated. To improve logistic regression performance, the systematic
habitat points were resampled to equal the number of tracked points. Habitat data were
arcsin transformed, and a backward (Wald) logistic regression was used to identify the
habitat variables that best predict suitable microhabitat for P. cornutum.
A Spearman correlation was used to determined correlations between independent
variables in both systematic and tracked habitat datasets. This was used to explain
variation in the logistic regression and DFA.
A randomization technique was used to test if the distribution of P. cornutum was
influenced by S. invicta (Resampling Stats™) (Blank, 2004). The Euclidean distance was
calculated between the individual capture locations of P. cornutum and locations of S.
invicta mounds and native ant mounds. I calculated the observed distances. Then the
association of ant mounds was randomized among the individual capture locations of P.
cornutum and resampled without replacement. Population distributions were generated
by using the average Euclidean distance, and I recorded the average expected values
10,000 times. A Z-test was used to compare the observed mean distances of P. cornutum
to invasive ant mounds and native ant mounds to the expected distances.
RESULTS
Habitat
Percent canopy cover was significantly different among burn treatments (H =
226.404, df = 2, p < 0.001). The Burn 1 treatment exhibited the lowest percent canopy

51
cover, and Burn 3 treatment had the highest percent canopy cover. Estimates of standing
stem density, as indicated by a compression index, exhibited a pattern similar to canopy
cover among treatments (H = 265.027, df = 2, p < 0.001). Significance was detected
among estimates of soil compaction at a 7.6 cm depth (H = 28.563, df = 2, p < 0.001).
However, soil compaction in Burn 1 and Burn 3 treatments were statistically similar and
less compact than Burn 2 treatments.
Habitat structure along the road-side was different from 5 m from the road-side
and further (Figure 2). The DFA discriminated among nine distance-intervals oriented
perpendicular to Middle Road and regrouped them into two distinct groups. Road-side
habitat structure was characterized by high soil compaction that averaged 295 psi, low
percent canopy cover that averaged 40%, and high bare ground that averaged 76%.
Habitat structure 5 m from the road-side and further had an average soil compaction of 75
psi, percent canopy cover of 90%, and percent bare ground of 50%. These variables
loaded positively in the DFA and explained a significant amount of the variation in
habitat structure (Wilks λ = 0.200, df = 120, p < 0.001), (eigenvalue = 2.140).
Tracking
A total of 128 P. cornutum were captured along the road from May-August 2009,
and I had 23 recaptures from the current year and one from 2008. Captures comprised 44
females, 26 males and 58 juveniles. Mean snout-vent length of adults was 71.5 cm, and
ranged from 40.0-93.0 cm (Table 1). Juvenile average snout-vent length was 26.7 cm
and ranged from 20.5-36.5 cm. The average total length of adults was 106.5 cm, ranging
from 58.0-132.0 cm, while the average total length of juveniles was 38.8 cm, and ranged

52
from 31.0-53.5 cm. Average body mass for adults was 25.1 g, and ranged from 4.0-48 g.
Juvenile body mass ranged from 0.5-4.0 g, with an average of 1.1 g.
Using the Schnabel index, I estimated 138 P. cornutum present on the island, with
a 95% confidence interval of 132-143 adults. Two disjunct populations where 50% of
the observations of adult P. cornutum occurred were located at opposite ends of the
island (Figure 3). The two 50% isopleths were separated by approximately 10 km. The
distribution of juveniles was similar but more restricted and the 50% isopleths for
juveniles were separated by approximately 27 km (Figure 4).
The percent of available treatment types according to adjacent combinations was
dominated by Burn 1 treatments adjacent to Burn 3 treatments at 45% (Tables 3 and 4).
Burn 1 treatments adjacent to another Burn 1, and Burn 3 treatments adjacent to Burn 2
and another Burn 3 were at 13%, 21%, and 17%, respectively. Only 5% of the available
adjacent treatments were Burn 2 adjacent to Burn 2. There were no Burn 1 treatments
adjacent to Burn 2 treatments due to a wildfire, eliminating that combination of adjacent
treatments.
A chi-square test indicated that observations of adult P. cornutum did not occur in
the same proportion as available treatment combinations (χ2 = 58.686, df = 4, p < 0.05)
(Table 2). Relative to Middle Road, fewer observations occurred than expected when
Burn 1 treatments were adjacent to another Burn 1 or Burn 3 treatment (Figure 5).
Observations exceeded expectations when Burn 3 treatments were adjacent to one
another. Observations were near expected values for adults when Burn 2 treatments were
adjacent to another Burn 2 or a Burn 3 treatment.

53
Juvenile P. cornutum also were distributed nonrandomly across Matagorda Island
as indicated by a chi-square analysis (χ2 = 155.952, df = 4, p < 0.05) (Table 3). Juveniles
were observed less than expected in all adjacent treatment types except for when Burn 3
was adjacent to Burn 3 (Figure 6).
Twenty P. cornutum were tracked, 16 females and 6 males, throughout the length
of Matagorda Island in all three burn treatments. Only 19 tracks were used in analysis of
habitat associations because one of the trails overlapped and the path could not be
discerned. Eight P. cornutum were relocated and powdered a second night, one of which
was also tracked a third night. The average absolute distance was 67.50 m and ranged
from 7.68-155.50 m. Relative distance averaged 36.28 m, and ranged from 7.60-113.30
m. Most of the individuals followed the most obvious and open path of travel, whether
along Middle Road, bare ground, or a game trail. I sampled habitat characteristics at 5-m
intervals along each path; 243 points total. Among these points, 76% occurred within 5
m of the road and runway. The individual tracked for three nights accounted for 26% of
the points 5 m or more off the road.
The initial logistic regression was significant and reduced the number of variables
used in assessing microhabitat associations with the presence of P. cornutum from 13
variables to 5 variables. However, one of the remaining 5 variables was not significant;
therefore, another backward (Wald) logistic regression was conducted without that
variable. This final predictive model was significant (χ2 = 219.015, df = 3, p < 0.001),
and correctly predicted the location of 80% of the tracked lizard locations, and 48% of
the variation could be explained with 3 variables (Nagelkerke R2 = 0.485). Percent

54
canopy cover, percent basal cover, and soil compaction at a 7.6 cm depth were the three
variables that remained in the logistic equation (Table 4).
The results of both the DFA and logistic regression need to be interpreted with
caution as a few of the variables are highly correlated. Percent basal cover was positively
correlated with percent canopy cover (rho = 0.844, df = 934, p < 0.001), and percent bare
ground was negatively correlated with percent litter cover (rho = -0.782, df = 934, p <
0.001).
Ants
A total of 276 ant mounds was sampled across 15 120 m long transects that were.
I averaged 18.4 mounds per transect. However, no ant mounds were observed on the
north side of Middle Road at transect 32 (Figure 1). Nine ant genera were identified on
Matagorda Island (Table 5), and were unevenly distributed throughout the island (Figure
7). S. invicta was the most frequently observed ant and occurred throughout all transects
except for the northernmost transect. The highest frequency of S. invicta was observed
throughout the center of the island in Burn 1 treatments. Dorymyrmex sp. was observed
in all transects but two; however, they were in lower frequency than S. invicta.
Crematogaster sp. was only observed at opposite ends of Matagorda Island. The most
genera rich transect was 16, which was near the intersection of Burn 1, Burn 2, and Burn
3 treatments. The randomization detected a significant difference in the distance of P.
cornutum to native and invasive ant mounds (Z = -20.522, df = 179, p < 0.001). P.
cornutum was positively associated with native ant mounds, and negatively associated
with invasive ant mounds.

55
DISCUSSION
In the early 1990s, a decline in populations of P. cornutum was becoming
apparent. Donaldson et al. (1994) conducted a Texas-wide survey for P. cornutum and
captured-and-marked ten individuals in Aransas County, a mainland area adjacent to
Matagorda Island. They also observed S. invicta and Pogonomyrmex sp. throughout
Central and Southern Texas (Donaldson et al., 1994). On Matagorda Island, Prieto
(1995) used drift-fence arrays within prescribed burn treatments to characterize the
herpetofauna, and categorized P. cornutum as a common species. Prieto (1995) also
noted that the herpetofauna had recovered to pre-burn levels by the second year post burn
and that P. cornutum were consistently found on the road-side. From 2003-2006, refuge
staff at Aransas NWR monitored four 50-m long drift-fence arrays monthly in Aransas
County, and never captured P. cornutum (Chad Stinson, pers. comm.). In 2008, P.
cornutum was one of the least frequently captured herpetofaunal species on Matagorda
Island (Inslee, Chapter 1).
Fire disturbance, as manipulated by the USFWS, on Matagorda Island promoted
heterogeneity. Canopy cover and standing stem density increased following burns, there
is a transition from forbs to grasses after two growing seasons, and soil is more
compacted after two years relative to other burn treatments. Middle Road has a
significant impact on habitat structure throughout Matagorda Island. The soil was more
compact with less vegetative cover within 5 m of the road-side and transitions to more
densely covered and loosely compacted soil further off the road. The estimated
population size of P. cornutum on Matagorda Island was 138 individuals. Both PFP

56
tracking and mark-recapture data indicated a close association of P. cornutum with road-
side habitat in Burn 2 and Burn 3 treatments. Literature suggests that P. cornutum was a
dietary specialist of Pogonomyrmex sp. (Pianka and Parker, 1975; Lane, 2008). Many
hours were spent systematically searching for native ants; however, I did not locate any
Pogonomyrmex sp. Native ants were more commonly associated with Burn 2 and 3
treatments, while the invasive S. invicta were closely associated with Burn 1 habitat.
Road captures of P. cornutum occurred in high abundance at both ends of
Matagorda Island and estimated home ranges suggest two disjunct populations. This
pattern was similar to the pattern observed in 2008 and in both years the populations were
separated by an extensive area of Burn 1 treatment. However, the northern population
was more restricted and the southern population expanded compared to 2008 (Inslee,
Chapter 1) (Figure 3). Juvenile distributions were more restricted than adults from both
2008 and 2009 (Figure 4).
My data supported the growing literature that prescribed burning generated
suitable habitat for P. cornutum. The distribution on Matagorda Island strongly suggests
that P. cornutum avoid Burn 1 treatments (Inslee, Chapter 1) (Figures 5 and 6). Burrow
et al. (2001), Moeller (2000), and Hellgren et al. (2010) suggest that Burn 1 does not
provide adequate protective cover, food resources, or thermoregulative structure. In
2008, I observed greater than expected numbers of P. cornutum in Burn 2 treatments.
But in 2009, I observed only the expected number, possibly due to the limited availability
of that treatment (< 6%). In 2008, there were fewer observations than expected in Burn 3
adjacent to Burn 3 treatments (Inslee, Chapter 1). However, the low number of

57
observations in Burn 3 habitat was reversed in 2009, possibly because there was little
road-side habitat not associated with Burn 1 treatments.
The PFP method was a very efficient and effective way to monitor the movement
of P. cornutum. Two of the tracked individuals that were not relocated with the
ultraviolet light were recaptured on road surveys within a week of being released. The
fur patch had been removed and the body mass of the individual had not decreased. By
not tracking from June-August, I reduced the effect on male mate-searching strategies
(Stark et al., 2005). A majority of P. cornutum tended to follow the path of the road or
small animal trails; however, similar to the results of Stark et al. (2005), individuals were
not hindered by dense vegetation in contradiction to Whiting et al. (1993). I observed
that tracked individuals would crawl on top of grasses, which Stark et al. (2005) suggests
is an adaptive behavior to thermoregulate, or for predator avoidance from birds of prey.
A few of the relocated individuals were observed sleeping in the open along the road-
side.
The prescribed burn regime on Matagorda Island generated habitat patches.
Heterogeneity was increased throughout the island within each of the patches by this
disturbance. Some variables associated with this heterogeneous habitat structure are
percent canopy cover, standing stem density, and soil compaction. Higher canopy cover
and standing stem density occurred in Burn 3 treatments. This is intuitive, because
multiple growing seasons since the last fire allow for more dense vegetative cover.
However, higher soil compaction occurred in areas that were burned two years prior.

58
Altering the proportion of the landscape occupied by different serial stages also
influences the stability of a population and community (Turner et al., 2001). The pattern
of heterogeneity at the island level is altered every year depending on the prescribed burn
schedule. For example, burned patches in the adjacent landscape across Middle Road
were not as evenly distributed across Matagorda Island in 2009 as they were in 2008
(Inslee, Chapter 1). In 2009, there was no location on the island where Burn 2 treatments
were adjacent to Burn 1, while in 2008 6% of the island was Burn 2 adjacent to Burn 1.
Decreases in proportions of available habitat from 2008 to 2009 also were observed in
Burn 2 adjacent to Burn 2 and Burn 3, as well as Burn 3 adjacent to Burn 3. Yet, the
amount of Burn 1 adjacent to Burn 3 treatments increased by 32% from 2008 to 2009.
Burn 1 adjacent to Burn 1 remained relatively stable at 11% and 13%. One of the reasons
the proportion of available habitat differed between years is a wildfire burned 2,000 ha on
the central portion of the island in mid-April 2009. The potential extent of this natural
disturbance was reduced by the heterogeneity of the prescribed burn regime, but also by
the natural heterogeneity of the adjacent salt marsh. I considered this wildfire as a Burn 1
treatment in all my analyses.
I focused habitat analysis on a structurally simple yet influential feature, Middle
Road, and observed its effect on habitat structure throughout the length of Matagorda
Island. Habitat along the road-side up to 5 m off the road was significantly different than
habitat more distant from the road-side. The road-side habitat was very homogeneous,
while habitat further off the road transitioned to a variable habitat structure and
composition that was on average greater in canopy cover and standing stem density with

59
more loosely compacted soil (Figure 2). Percent litter cover, percent bare ground, and
soil compaction explain most of the variation among burn treatments. Although there is
minimal traffic along Middle Road, the traffic that does use the road consists of heavy
machinery and vehicles, which explains the high compaction along road-sides. At the
northern portion of the island was an abandoned Air Force base where heavy machinery
was used and contributed to soil compaction. This might explain the higher variation
observed at the north end of the island in the DFA. A higher proportion of bare ground
was present along the road-sides, most likely due to the high soil compaction that restricts
vegetative growth. Deeper litter was present further off the road. Burrow et al. (2001)
described the importance of microhabitat selection of P. cornutum throughout the day.
Bare ground was important in the morning and evenings while litter was selected in the
afternoon (Burrow et al., 2001). The close proximity of bare ground and litter along
Middle Road might be suitable habitat for thermoregulatory behaviour.
Growth, reproduction, and dispersal of organisms can be strongly affected by the
spatial distribution of resources in a heterogeneous landscape (Turner et al., 2001). I
analyzed habitat associations to assess the cause of the disjunct populations of P.
cornutum on Matagorda Island (Figure 3 and 4). A logistic regression identified three
variables that significantly influenced the presence of individuals. P. cornutum is
strongly associated with habitat characteristics consistent with that observed along the
side of Middle Road. The model predicted P. cornutum to occur less often in areas with
high percent canopy cover. However, in areas with little canopy cover, P. cornutum
occurred in areas with higher basal cover. Burrow et al. (2001) demonstrated that habitat

60
structure is valuable for thermoregulation. Phrynosoma cornutum were associated with
areas of higher soil compaction. Phrynosoma cornutum burrow to avoid solar radiation,
to hibernate, and to dig out nests (Russell, 1997; Sherbrooke, 2003). The soil depth
required for these behaviors varies depending on environmental conditions and soil type
(Fair and Henke, 1997b; Sherbrooke, 2003). To avoid solar radiation they will burrow to
depths of 1.3-7.6 cm, while nest depths range from 15.2-20.3 cm (Sherbrooke, 2003).
The higher soil compaction in Burn 2 treatment would allow for burrowing depths
consistent with behavior to avoid solar radiation (Inslee, Chapter 1). Burn 3 treatments,
however, have more loosely compacted soil that would be easier to dig to appropriate
nests (Sherbrooke, 2003). I observed high abundance of juveniles in Burn 3 treatments.
Distribution of food resources might influence the observed distributions of P.
cornutum (Whiting et al., 1993). Eleven genera of ants were recorded on Matagorda
Island. Phrynosoma cornutum is considered a dietary specialist, and Pogonomyrmex sp.
can constitute up to 80% of their diet (Pianka and Parker, 1975; Lane, 2008). In 2007,
Chad Stinson (pers. comm.) observed Pogonomyrmex sp. at the north end of the island
near the runways. However, after systematically surveying Matagorda Island, I did not
observe any Pogonomyrmex sp. or any evidence of their presence. DeMers (1993)
suggested that Pogonomyrmex occidentalis used road ditches as a corridor for dispersal.
I speculate that as the population of Pogonomyrmex sp. was declining on the island, their
distribution constricted to road-side habitat. It would not be unreasonable for P.
cornutum to then follow their preferred food source and remain there even after

61
Pogonomyrmex sp. went locally extinct. This might explain why P. cornutum are
observed frequently along the road-side.
Solenopsis invicta have been present on Matagorda Island at least since 1993
(Chad Stinson, pers. comm.) and are currently the dominant ant species (Table 6).
Similar to other biological invaders, S. invicta tend to invade and dominate entire habitat
patches (Holway et al., 2002, Suarez and Case, 2002) by scavenging on invertebrates and
small vertebrates (Allen et al., 2004). Although I never observed an interaction of P.
cornutum with S. invicta, it is likely that S. invicta is a predator of P. cornumtum on
Matagorda Island. S. invicta have been known to attack and prey upon incubating eggs
and hibernating adult P. cornutum (Allen et al., 2004). I also observed S. invicta feeding
on fauna, including P. cornutum, in funnel traps (Inslee, Chapter 1).
It is evident based on the two populations of P. cornutum on either end of the
island (Figures 3 and 4) that they are not associated with the high frequency of S. invicta
at the center of the island (Figure 7). Although I determined that the distance of P.
cornutum from invasive fire ants was significantly greater than the distance from P.
cornutum to native ants, one of the assumptions of this procedure is that all organisms
(lizards and ants) were evenly distributed throughout the island. The distribution of S.
invicta was unevenly distributed across Matagorda Island, and highly associated with
Burn 1 treatments. I observed higher abundance of juvenile P. cornutum in Burn 3
treatments, which were isolated at both ends of the island. Beside the fact that soil
compaction in these treatments are conducive for building nests, these areas are also
furthest away from S. invicta, a potential predator. I observed an uneven sex ratio in

62
these treatments (1F:0.57M). I suggest that female P. cornutum are avoiding areas with
S. invicta when selecting nesting sites. In addition, the distribution of P. cornutum,
especially juveniles is congruent with the distribution of Crematogaster sp. on both ends
of the island. In California, Suarez and Case (2002) observed that juvenile coastal horned
lizards (Phrynosoma coronatum) fed on the smaller Crematogaster sp. of ants in areas
invaded by non-native ants.
Future Research and Management Recommendations
Recovery and species responses following a disturbance depend on patch size,
heterogeneity, distance from undisturbed sites, and life history, which are all influenced
by the intensity, size, and frequency of disturbance (Turner et al., 2001). Fire was a
manipulated disturbance on the island, while S. invicta changed the context of the
landscape. These processes appear to be closely interrelated, but I could not isolate them
statistically.
Although roads are often considered major contributors to habitat fragmentation,
Middle Road might be considered a corridor linking the two metapopulations of P.
cornutum on Matagorda Island. For a corridor to be effective, Rosenberg et al. (1997)
recorded these common patterns: 1) organisms will select pathways with similar elements
of their preferred habitat when confronted with the choice, 2) the relative use of the
matrix for movement depends on its contrast with the suitable habitat of the organism,
and 3) in areas of less favorable habitat the behavior of the organism might change.
Powder tracking suggests that most of the movement occurs within 5 m of either side of
the road.

63
Organism movements can be better understood by using a combination of
tracking methods (Stapp, 1997; Eggert, 2002; Mendel and Vieira, 2003). A more
comprehensive understanding of microhabitat associations through the day and through
seasons could be achieved by incorporating radiotelemetry (Burrow et al., 2001; Eggert,
2002). Higher connectivity of patches and smaller burned patch sizes might benefit P.
cornutum because this might increase recolonization rates of P. cornutum, and reduce
relative abundance of S. invicta relative to native ants, assuming that S. invicta is
influenced by fire. Soil compaction was identified as a key variable influencing P.
cornutum distribution and habitat characteristics by both the logistic regression model
and the DFA. In both 2008 and 2009, soil compaction was highest in Burn 2 treatments.
These data suggested that heterogeneity of soil compaction was required by P. cornutum.
Further investigation of movement throughout different seasons would provide better
understanding of where P. cornutum is mating, nesting, and hibernating and at what soil
depths (Fair and Henke, 1997b).
On Matagorda Island, P. cornutum were observed in alarmingly low abundance
and warrant monitoring and augmented management strategies. Phrynosoma cornutum
were observed within 5 m of the road-side, which only constituted 2% of the available
grassland habitat. The pattern of prescribed burn patch is highly influential on the
distribution of P. cornutum. Management should consider burning in smaller patch sizes
to allow P. cornutum to move more easily among suitable habitat with minimal
interaction with S. invicta.

64
LITERATURE CITED
ALLEN, C. R., D. M. EPPERSON, AND A. S. GARMESTANI. 2004. Red imported fire ant
impacts of wildlife: a decade of research. American Midland Naturalist 152:88-103.
ANDERSON, T. J. C., A. J. BERRY, J. N. AMOS, AND J. M COOK. 1988. Spool-and-line
tracking of the New Guinea spiny bandicoot, Echymipera kalubu (Marsupialia,
Peramelidae). Journal of Mammalogy 69:114-120.
BLANK, S. 2004. Resampling Stats™ for Excel WinVersion 2.0 – Internal RNG,
Resampling Stats, Inc.
BLANKENSHIP, E. L., T. W. BRYAN, AND S. P. JACOBSEN. 1990. A method for tracking
tortoises using fluorescent powder. Herpetological Review 21:88-89.
BURROW, A. L., R. KAZMAIER, E. HELLGREN, AND D. RUTHVEN, III. 2001. Microhabitat
selection by P. cornutum in southern Texas. Journal of Wildlife Management 65:645-
652.
COLLINS, S. L. 1992. Fire frequency and community heterogeneity in tallgrass prairie
vegetation. Ecology 73:2001-2006.
CONOVER, W.J. 1980. Practical Nonparametric Statistics, second edition. John Wiley &
Sons, New York.
DEMERS, M. 1993. Roadside ditches as corridors for range expansion of the western
harvester ant (Pogonomyrmex occidentalis Cresson). Landscape Ecology 8:93-102.

65
DODD, C. K. 1992. Fluorescent powder is only partially successful in tracking
movements of the six-lined racerunner (Cnemidophorus sexlineatus). Florida Field
Naturalist 20:8-14.
DONALDSON, W., A. H. PRICE, AND J. MORSE. 1994. The current status and future
prospects of the Texas horned lizard (Phrynosoma cornutum) in Texas. Texas
Journal of Science 46:97-113.
EGGERT, C. 2002. Use of fluorescent pigments and implantable transmitters to track a
fossorial toad (Pelobates fuscus). Herpetological Journal 12:69-74.
FAIR, W. S. AND S. HENKE. 1997a. Efficacy of capture methods for a low density
population of Phrynosoma cornutum. Herpetological Review 28:135-137.
FAIR, W. S. AND S. HENKE. 1997b. Effects of habitat manipulations on Texas horned
lizards and their prey. The Journal of Wildlife Management 61:1366-1370.
FELLERS, G. M. AND C. A. DROST. 1989. Fluorescent powder: a method for tracking
reptiles. Herpetological Review 20:91-92.
FISHER, B. L. AND S. P. COVER. 2007. Ants of North America: a guide to the genera.
University of California Press, Berkely, California.
FISHER, M. AND A. MUTH. 1995. A backpack method for mounting radio transmitters to
small lizards. Herpetological Review 26:139-140.
FUHLENDORF, S. D., W. C. HARRELL, D. M. ENGLE, R. G. HAMILTON, C. A. DAVIS, AND D.
M. LESLIE, JR. 2006. Should heterogeneity be the basis for conservation?

66
Grassland bird response to fire and grazing. Ecological Applications 16:1706-
1716.
HALFPENNY, J. C. 1992. Environmental impacts of powder tracking using fluorescent
pigments. Journal of Mammalogy 73:680-682.
HEATH, W. G. 1961. A trailing device for small animals designed for field study of the
gila monster (Heloderma suspectum). Copeia 1961:491-492.
HELLGREN, E. C., A. L. BURROW, R. T. KAZMAIER, AND D. C. RUTHVEN, III. 2010. The
effects of winter burning and grazing on resources and survival of Texas horned
lizards in a thornscrub ecosystem. The Journal of Wildlife Management 71:300-309.
HENKE, S. E. AND M. MONTEMAYOR. 1998. Diel and monthly variations in capture
success of Phrynosoma cornutum via road cruising in Southern Texas.
Herpetological Review 29:148-150.
HOLWAY, D. A., L. LACH, A. V. SUAREZ, N. D. TSUTSUI, AND T. J. CASE. 2002. The
causes and consequences of ant invasions. Annual Review of Ecological Systems
33:181-233.
KRUMMEL, J. R., R. H. GARDNER, G. SUGIHARA, R. V. O’NEILL, AND P. R. COLEMAN.
1987. Landscape patterns in a disturbed environment. Oikos 48:321-324.
LANE, J. S. 2008. Assessing diet and daily movement of Texas horned lizards
(Phrynosoma cornutum) in Southern Texas. Unpublished Thesis. West Texas A&M
University.

67
LEMEN, C. A. AND P. W. FREEMAN. 1985. Tracking mammals with fluorescent pigments:
a new technique. Journal of Mammalogy 66:134-136.
LEVINS, R. 1969. Some demographic and genetic consequences of environmental
heterogeneity for biological control. Bulletin of the Entomological Society of
America 15: 237-240.
MENDEL, S. M. AND M. V. VIEIRA. 2003. Movement distances and density estimation of
small mammals using the spool-and-line technique. Acta Theriologica 48:289-300.
MOELLER, B. A. 2000. Conservation ecology of the Texas horned lizard (Phrynosoma
cornutum): comparative effects of summer and winter burning. Unpublished Thesis,
Auburn University, Alabama.
MORRISON, L. W. 2002. Long-term impacts of an arthropod-community invasion by the
imported fire ant, Solenopsis invicta. Ecology 83:2337-2345.
MUNGER, J. C. 1984. Optimal Foraging? Patch use by horned lizards (Iguanidae:
Phrynosoma). The American Naturalist 123:654-680.
PIANKA, E. R. AND W. S. PARKER. 1975. Ecology of horned lizards: a review with special
reference to Phrynosoma platyrhinos. Copeia 1:141-162.
PRADEL, R. 1996. Utilization of capture-mark-recapture for the study of recruitment and
population growth rate. Biometrics 52:703-709.
PRIETO, F. G. 1995. Terrestrial small vertebrate response to seasonal prescribed burns on
Matagorda Island- A Texas coastal barrier island. Unpublished data.

68
PULLIAM, H. R. 1988. Sources, Sinks, and Population Regulation. The American
Naturalist 132:652-661.
REINERT, H. K. 1984. Habitat separation between sympatric snake populations. Ecology
65:478-486.
RISSER, P. G., J. R. KARR, AND R. T. T. FORMAN. 1984. Landscape ecology: directions
and approaches. Special Publication Number 2. Illinois Natural History Survey,
Champaign, Illinois, USA.
ROSENBERG, D. K., B. R. NOON, AND E. C MESLOW. 1997. Biological corridors: form,
function, and efficacy. BioScience 47:677-687.
RUSSELL, T. A. 1997. Thermal ecology of the Texas horned lizard (Phrynosoma
cornutum). Unpublished Thesis, University of Central Oklahoma, Oklahoma.
SANDERSON, G. C. 1966. The study of mammal movements: a review. The Journal of
Wildlife Management 30:215-235.
SHARROW, S. H. 1984. A simple disc meter for measurement of pasture height and
forage bulk. Journal of Range Management 37:94-95.
SHERBROOKE, W. C. 2002. Seasonally skewed sex-ratios of road-collected Texas horned
lizards (Phrynosoma cornutum). Herpetological Review 33:21-24.
SHERBROOKE, W. C. 2003. Introduction to horned lizards of North America. University
of California Press, Berkeley, California.

69
STAPP, P. 1997. Habitat selection by an insectivorous rodent: patterns and mechanisms
across multiple scales. Journal of Mammalogy 78:1128-1143.
STARK R. C. AND S. F. FOX. 2000. Use of fluorescent powder to track horned lizards.
Herpetological Review 31:230-231.
STARK, R. C., S. FOX, AND D. M. LESLIE, JR. 2005. Male Texas horned lizards increase
daily movements and area covered in spring: A mate searching strategy? Journal of
Herpetology 39:169-173.
SUAREZ, A. V. AND T. J. CASE. 2002. Bottom-up effects of persistence of a specialist
predator: ant invasions and horned lizards. Ecological Applications 12:291-298.
TABACHNICK, B. G. AND L. S. FIDELL. 2007. Using Multivariate Statistics (fifth ed.).
Allyn and Bacon, Boston, MA.
TURNER, M. G., GARDNER R. H., AND R. V. O’NEILL (editors). 2001. Landscape ecology
in theory and practice: pattern and process. Springer Science+Business Media, LLC,
United States.
WHITE, P. S. AND S. T. A. PICKETT. 1985. Natural disturbance and patch dynamics: an
introduction. Pages 3-13 in The ecology of natural disturbance and patch dynamics
(S. T. A. Pickett and P. S. White, editors). Academic Press, New York.
WHITING, M. J., J. R. DIXON, AND R. C. MURRAY. 1993. Spatial distribution of a
population of Texas horned lizards (Phrynosoma cornutum: Phrynosomatidae)
relative to habitat and prey. The Southwestern Naturalist 38:150-154.

70
WIENS, J. A. 1976. Population responses to patchy environments. Annual Review of
Ecology and Systematics 7:81-120.
WILSON, D. S. 1994. Tracking small animals with thread bobbins. Herpetological
Review 25:13-14.
WOJCIK, D. P., C. R. ALLEN, R. J. BRENNER, E. A. FORYS, D. P. JOUVENAZ, AND R. S.
LUTZ. 2001. Red imported fire ants: impact on biodiversity. American Entomologist
47:16-23.

71
TABLE 1—Range and mean snout-vent length, total length, and body mass of males,
females, and juveniles for road captures of P. cornutum from June-August 2009 on
Matagorda Island, Texas.
Snout-Vent Length (mm)
Total Length (mm)
Body Mass (g)
N Range Mean
Range Mean
Range Mean
Female 44 40.0 - 93.0 72.3
58.0 - 132.0 104.6
4.0 - 48.0 25.2
Male 26 58.0 - 84.0 70.1
83.0 - 132.0 109.7
12.0 - 36.0 25.0
Juvenile 58 20.5 - 36.5 26.7
31.0 – 53.5 38.8
0.5 – 4.0 1.1
Total 128 20.5 - 93.0 51.2
31.0 - 132.0 75.8
0.5 - 48.0 14.3

72
TABLE 2—Chi-square analysis of road captures of adult P. cornutum exhibiting a
significant difference in habitat use in adjacency to three types of burn treatments from
June-August 2009 on Matagorda Island, TX. Percent is the percent of available adjacent
habitat combinations.
Adjacent Habitats Percent Expected Observed
Burn1-Burn1 13.5 10 2
Burn1-Burn3 42.3 30 15
Burn2-Burn2 5.8 4 5
Burn2-Burn3 21.2 15 14
Burn3-Burn3 17.3 13 36
58.686χ2calc > 9.488χ2
crit

73
TABLE 3—Chi-square analysis of road captures of juvenile P. cornutum exhibiting a
significant difference in habitat use in adjacency to three types of burn treatments from
June-August 2009 on Matagorda Island, TX. Percent is the percent of available adjacent
habitat combinations.
Adjacent Habitats Percent Expected Observed
Burn1-Burn1 13.5 7 0
Burn 1-Burn3 42.3 24 4
Burn2-Burn2 5.7 3 2
Burn2-Burn3 21.1 12 5
Burn3-Burn3 17.3 10 45
155.952χ2calc > 9.488χ2
crit

74
TABLE 4—Microhabitat variables identified by a backward (Wald) logistic regression that
are most influential in predicting the presence of P. cornutum on Matagorda Island, TX
(χ2 = 219.015, df = 3, p < 0.001).
Variables B Wald DF Sig. % Canopy Cover -3.582 26.035 1 <0.001
% Basal Cover 3.407 15.433 1 <0.001
Soil Compaction 0.010 74.964 1 <0.001
Constant -1.027 8.963 1 0.003

75
TABLE 5—Frequency of occurrence of ant mounds (genera) along thirty 60 m transects
systematically distributed along Middle Road from May-June 2009 on Matagorda Island,
TX.
Ant Genera Frequency
Atta sp. 0.006
Crematogaster sp. 0.157
Cyphomyrmex sp. 0.003
Dorymyrmex sp. 0.229
Forelius sp. 0.075
Paratrechina sp. 0.018
Pheidole sp. 0.030
Solenopsis invicta 0.456
Trachymyrmex sp. 0.024

76
FIG. 1—Locations of systematic surveys of habitat structure within the prescribed burn regime on Matagorda Island, Texas.
N
A 0 2.S 5 10 ~ lom1ter1
Legend X Tran mt Numbor --- Middlo Road
D MataaordahlandBoundary Runway
- Burn 1 yoar prior - Burn 2 yoar1 prior - Burn 3 or moro yoar1 prior

77
-4.00 -2.00 0.00 2.00 4.00 6.00
Discriminant Function 1
0
10
20
30
40
50
Tran
sect
Alo
ng M
iddl
e Roa
d
Distance from Middle Road
0 m (road-side)
5 m
10 m
15 m
20 m
30 m
40 m
50 m
60 m
FIG. 2—Discriminant function analysis of nine distance-interals oriented perpendicular to
Middle Road. The distance-intervals separated into two groups (road-side areas and all
others) based on characteristics of habitat structure (Eigenvalue=8.351, Wilks λ=0.107, df
= 13, p < 0.001).
a
0 ·.\·
.rtJ. _. e • • i: '• . •• i
• 0 • D ... ... V
*

78
FIG. 3—Captures of adult P.cornutum from June-July 2009 on Matagorda Island, Texas. Chi-square test of independence detected a nonrandom distribution, and a 50% isopleth outlines the isolated distributions.
N
A 0 2,S s lOKilometm
Legend D MataaordahhmdBoundary D BurnUnit1 - Middle Road
- RunWi'f
A Male Road Capture• • Female Road Captum
- Burn 1 Year Prior - Burn 2 Year1 Prior - Burn 3 or Moro Yoar, Prior
D SO½I1opleth

79
FIG. 4—Captures of juvenile P.cornutum from June-July 2009 on Matagorda Island, Texas. Chi-square test of independence detected a nonrandom distribution, and a 50% isopleth outlines the isolated distributions.
N
A 0 2,S S 10 I<il om oton
Legend CJ Mataaorda hi and Boundary CJ Burn Unit, - - Middlo Road - Runway
• juvonilo, N
- Burn 1 Yoar Prior - Burn 2 Yoar, Priot ~ . - Burn 3 or Moro Yan Prior
D 50% leopleth

80
FIGURE 5—Differences among observed numbers of adult P. cornutum and expected
numbers relative to the proportion of available combinations of burn treatments along
Middle Road, Matagorda Island, Texas (χ2 = 58.686, df = 4, p < 0.05). The thick line
along the zero x-axis is the expected value for a random distribution of P. cornutum on
the island. The dashed line along the x-axis title represents Middle Road and the
combination of adjacent burn treatments.
-7.7 -15.5
0.8
-1.2
23.5
-25.0
-20.0
-15.0
-10.0
-5.0
0.0
5.0
10.0
15.0
20.0
25.0
Burn 1
Burn 1
Burn 1
Burn 3
Burn 2
Burn2
Burn 2
Burn 3
Burn 3
Burn 3
Obs
erve
d-E
xpec
ted
Valu
es
Adjacent Burn Treatments

81
FIGURE 6—Differences among observed numbers of juvenile P. cornutum and expected
numbers relative to the proportion of available combinations of burn treatments along
Middle Road, Matagorda Island, Texas (χ2 = 155.952, df = 4, p < 0.05). The thick line
along the zero x-axis is the expected value for a random distribution of P. cornutum on
the island. The dashed line along the x-axis title represents Middle Road and the
combination of adjacent burn treatments.
-7.0 -20.0 -1.0 -7.0
35.0
-35.0
-25.0
-15.0
-5.0
5.0
15.0
25.0
35.0
Burn 1
Burn 1
Burn 1
Burn 3
Burn 2
Burn2
Burn 2
Burn 3
Burn 3
Burn 3
Obs
erve
d -E
xpec
ted
Valu
es
Adjacent Burn Treatments

82
FIG. 7—Relative abundance (number of mounds) for ants (genera) observed along 120-m transects systematically
distributed along Middle Road, Matagorda Island, TX.