Embed Size (px)
Citation preview

lable at ScienceDirect
Soil Biology & Biochemistry 68 (2014) 419e428
Contents lists avai
Soil Biology & Biochemistry
journal homepage: www.elsevier .com/locate/soi lb io
Hemiparasitic litter additions alter gross nitrogen turnover intemperate semi-natural grassland soils
Andreas Demeya,*, Tobias Rütting b, Dries Huygens c,d,e, Jeroen Staelens a,c, Martin Hermy f,Kris Verheyen a, Pascal Boeckx c
a Forest & Nature Lab (ForNaLab), Ghent University, Geraardsbergsesteenweg 267, B-9090 Gontrode, BelgiumbDepartment of Earth Sciences, University of Gothenburg, Box 460, 405 30 Gothenburg, Swedenc Isotope Bioscience Laboratory (ISOFYS), Ghent University, Coupure Links 653, 9000 Gent, Belgiumd Institute of Agricultural Engineering and Soil Science, Faculty of Agricultural Sciences, Universidad Austral de Chile, Valdivia, Chilee Instituto Multidisciplinario de Biología Vegetal e IMBIV, Consejo Nacional de Investigaciones Científicas y Técnicas de Argentina, Facultad de CienciasExactas, Físicas y Naturales, Universidad Nacional de Córdoba, Córdoba, ArgentinafDivision Forest, Nature and Landscape (FNL), KULeuven, Celestijnenlaan 200E, 3001 Heverlee, Belgium
a r t i c l e i n f o
Article history:Received 25 March 2013Received in revised form9 October 2013Accepted 16 October 2013Available online 25 October 2013
Keywords:Hemiparasitic plantsLitterN transformationsRhinanthusPedicularisSemi-natural grassland
* Corresponding author. Tel.: þ32 (0)9 264 90 38;E-mail address: [email protected] (A. Dem
0038-0717/$ e see front matter � 2013 Elsevier Ltd.http://dx.doi.org/10.1016/j.soilbio.2013.10.025
a b s t r a c t
Hemiparasitic plants accumulate nutrients in their leaves and therefore produce high-quality litter withfaster decomposition and nutrient release rates compared to non-parasitic litter. Higher levels of plant-available nitrogen (N) in the presence of hemiparasitic plants have been attributed to this ‘litter effect’,but effects on N dynamics in the soil remain unstudied. We tested the hypothesis that litter of Rhinanthusangustifolius and Pedicularis sylvatica increases N transformation rates in the soil more than non-parasiticlitter of a species mix from the same communities. We expected the litter effect to be higher in theoligotrophic Pedicularis soil compared to the mesotrophic Rhinanthus soil. Gross N transformation rateswere quantified using a 15N tracing modeling approach. Differentially 15N labeled NH4Cl þ KNO3 wasadded to two soils with three treatments (control, soil amended with non-parasitic litter, soil amendedwith Rhinanthus or Pedicularis litter) in a laboratory incubation experiment. The concentration and 15Nenrichment of NH4
þ and NO3 in the soil were measured at six time points within one or two weeks(depending on the soil) after label addition. Hemiparasitic litter addition increased the overall cycling ofN more compared to the addition of non-parasitic litter. Relative to the non-parasitic litter, addition ofRhinanthus litter increased the net flux from organic N to NH4
þ by 61% and net (autotrophic) nitrificationby 80%. Addition of Pedicularis litter increased the net flux from organic N to NH4
þ by 28% relative toaddition of non-parasitic litter, while there was no effect on nitrification. Surprisingly, gross minerali-zation of organic N to NH4
þ decreased with litter addition for the Rhinanthus soil (control soil > non-parasitic litter > Rhinanthus litter), while it increased with litter addition in the Pedicularis soil (controlsoil < non-parasitic litter < Pedicularis litter). Our results support the hypothesis that litter from hem-iparasitic plants increases soil N availability more than non-parasitic litter, but contradicts the expec-tation that the hemiparasitic litter effect would be more pronounced in an oligotrophic as compared to amesotrophic system. This litter-induced augmentation in soil fertility provides e in addition to theparasitic suppression of hosts e a second potentially important pathway by which hemiparasitic plantsimpact on plant community composition. However, future research on P and K return via hemiparasiticlitter should be considered.
� 2013 Elsevier Ltd. All rights reserved.
1. Introduction
The net effect of hemiparasitic plants on plant communitystructure and diversity results from both parasitism and litter
fax: þ32 (0)9 264 90 92.ey).
All rights reserved.
pathways (Press, 1998; Spasojevic and Suding, 2011). The para-sitism pathway refers to direct negative effects of hemiparasiticplants on host species and indirect positive effects on non-hostspecies. Most parasitic plants are generalists, but show highlevels of host preference (Press and Phoenix, 2005). Therefore,parasitism changes the competitive relations between preferredand non-preferred hosts in the vegetation with possible effects ondiversity (Gibson and Watkinson, 1991; Matthies, 1996; Press et al.,

A. Demey et al. / Soil Biology & Biochemistry 68 (2014) 419e428420
1999). For example, the decrease of total, graminoid and legumebiomass in grasslands infected with Rhinanthus spp. is thought toalter the species composition in favor of non-leguminous forbs andto increase the local diversity (Ameloot et al., 2005; Davies et al.,1997; Gibson and Watkinson, 1991). The litter pathway operatesvia effects on nutrient cycling. Hemiparasitic plants accumulatenutrients in their tissues, which is thought, in part, to be a result oftheir high transpiration rates (Gauslaa, 1990; Gauslaa and Odasz,1990; Pate, 1995; Phoenix and Press, 2005) and therefore producelitter with high decomposability (Press, 1998; Press et al., 1999;Quested et al., 2003a, 2002; Seel and Press, 1993). There is limitedevidence that hemiparasites can increase the amount of nitrogen(N) inputs to the soil (March and Watson, 2010; Quested et al.,2003a), increase net N mineralization (Bardgett et al., 2006), in-crease the amount of plant-available N (Ameloot et al., 2008), andenhance plant growth (Quested et al., 2003b). In grassland eco-systems, this increase in plant-available N (and likely other nutri-ents) potentially increases graminoid biomass and therefore mightdecrease diversity and change community composition as generallyobserved in nutrient addition experiments (De Schrijver et al.,2011; Hejcman et al., 2007; Silvertown et al., 2006). All together,parasitism is expected to decrease productivity and to impact ondiversity, either positively or negatively, depending on host pref-erence, while litter effects may increase productivity and haveweaker impacts on diversity (Spasojevic and Suding, 2011).
While the impact of hemiparasites on community structure iswell-studied (see the reviews of Ameloot et al., 2005; Press andPhoenix, 2005), studies looking at the litter pathway are scarcer.Press (1998) suggested that litter effects of hemiparasites should beinvestigated both in field and microcosm, through simple speciesmanipulation experiments, coupled to litter and tracer studies.Quested et al. (2005, 2003a,b, 2002) used litter in field and meso-cosm experiments and found that litter of the sub-arctic Bartsiaalpina increased N inputs to the soil, released N faster than co-occurring, non-parasitic species and enhanced plant growth morethan litter of non-parasitic species. March andWatson (2007, 2010)found that the mistletoe Amyema miquelii increased both litterfalland annual litter N (�1.65), phosphorus (P, �3) and potassium(K,�8.5) returns in temperate eucalypt forest. Ameloot et al. (2008)measured 15N abundance in the vegetation four months after a 15Ntracer was added to the soil of plots with and without Rhinanthusminor. A lower 15N enrichment in the vegetation of parasitized plotsindicated that the added tracer was more diluted by 14N from thesoil solution, suggesting higher N availability in the presence ofR. minor. The authors discussed several possible reasons for thisincrease in N availability, of which one is hemiparasitic litter input.In an observational study, Spasojevic and Suding (2011) associatedCastilleja occidentalis presencewith higher productivity and foliar Nconcentrations in co-occurring species in alpine tundra. Based ondecomposition trials, in which C. occidentalis (alone and in mix-tures) lost N much faster compared to the tested non-parasiticspecies, Spasojevic and Suding (2011) speculated that litter effectsoutweigh parasitism effects. However, the authors found no highersoil inorganic N pool associatedwith C. occidentalis. They concludedthat future research should examine N cycling in more detail.
Here, we aimed at understanding better how hemiparasiticlitter influences the bioavailability of N in the soil. Nitrogenbioavailability is primarily regulated by the depolymerization of N-containing organic compounds to monomers (e.g., amino acids),broadly available to bothmicro-organisms and plants. However, themain form of N taken up by plants likely varies depending on the Nstatus of the soil (Schimel and Bennett, 2004). Except for extremeN-poor ecosystems, plants primarily use inorganic N, shifting fromammonium (NH4
þ) to nitrate (NO3�) with increasing N availability.
As N mineralization and immobilization occur simultaneously in
the soil (Bottomley et al., 2012), net mineralization assays areinadequate for evaluating N supply rates and, hence, N bioavail-ability. Especially in N-limited soils, any produced inorganic N mayimmediately be immobilized by the plant or the microbial com-munity due to the strong competition for available inorganic N inthe soil matrix. To study the soil N cycle in detail, numerical modelsare the best available tool (Rütting et al., 2011). Combined with arobust technique for parameter optimization, they allow the esti-mation of several simultaneously occurring gross N trans-formations (Mary et al., 1998; Müller et al., 2007; Myrold andTiedje, 1986). In addition, numerical models correct for re-mineralization of added 15N, which is not the case for analyticalequations (Hart et al., 1994).
For this study, we selected two hemiparasitic species growing inecosystem types with contrasting dynamics: Rhinanthus angustifo-lius C.C. GMEL growing in mesotrophic grasslands and Pedicularissylvatica L. growing in oligotrophic heath-grassland (hereafter Rhi-nanthus and Pedicularis). Using a numerical data analysis based on a15N tracing model (Müller et al., 2007), we compared N trans-formations in (i) control soil, (ii) soil amended with non-parasiticlitter of a species mix and (iii) soil amended with hemiparasiticlitter from the same communities. The first hypothesis we tested isthat hemiparasitic litter addition increases gross soil N trans-formation rates more than addition of non-parasitic litter. Althoughplant uptake is excluded by the experimental design, increases ingross NH4
þ andNO3� forming process rates are expected to increaseN
bioavailability. The second hypothesis we tested is that the relativeeffect of hemiparasitic litter on gross soil N transformation rates ishigher in the oligotrophic (nutrient-poor) Pedicularis soil comparedto themesotrophic (moderately nutrient-rich) Rhinanthus soil. In anoligotrophic soil, the impact of high-quality hemiparasitic litter onNcycling is expected to be large as N cycling is inherently slow. In amesotrophic soil with faster N cycling, in contrast, the impact ofhemiparasitic litter is expected to be less profound.
2. Materials and methods
2.1. Studied vegetation types
For both hemiparasites, a representative site was selected in theeast of Flanders (northern Belgium). These were Doode Bemde(50�4805700N, 4�3805400E) for Rhinanthus and Hooiput (51�1805300N,5�703400E) for Pedicularis (hereafter Rhin-D and Pedi-H). The longterm (1981e2010) mean annual precipitation in the region is852 mm, and the mean annual temperature is 10.5 �C (RoyalMeteorological Institute of Belgium). The most important soilcharacteristics are listed in Table 1. Both sites are semi-naturalgrasslands located in nature reserves. The management consistsof mowing once (Pedi-H) or twice (Rhin-D) a year. The vegetation atRhin-D is particularly diverse with on average 21 � 1 species per0.25 m2, whereas Pedi-H counts 13 � 1 species per 0.25 m2. Basedon cover estimates, most important species at Rhin-D were Juncusacutiflorus EHRH. EX HOFFM. (21% average cover), Lotus uliginosusSCHKUHR (15%), Anthoxanthum odoratum agg. (12%), Ranunculusacris L. (9%) and Lychnis flos-cuculi L. (8%). At Pedi-H these werePotentilla erecta (L.) (20%), Erica tetralix L. (15%), Polygala serpyllifoliaHOSÉ (14%),Molinia caerulea agg. (12%) and Agrostis canina L. (10%).
2.2. Litter collection
Withering Rhinanthus and Pedicularis plants were collected fromboth sites at the end of June 2010. Since the management isinherently connected to the presence of the hemiparasites, only thepart of the plant that returns to the soil as litter under the man-agement regime was used in our experiment: Rhinanthus leaves as

Table 1Soil and litter properties (mean � SE) as well as amounts of 15N label added to threetreatments at two sites (Rhin-D and Pedi-H). All soil numeric data are for the 5e15 cm layer.
Rhin-D Pedi-H
SoilWRB class Eutric fluvisol Gleyic podzolUSDA texture Silt Loamy sandDrainage class (Moderately) poor PoorpH-H2O 5.18 � 0.20 4.59 � 0.01Water content (g g�1 dry soil) 0.51 � 0.01 0.38 � 0.01Bulk density (g cm�3) 0.52 � 0.11 0.72 � 0.06Water-filled pore space (%)a 33 38Total C (%) 4.07 � 0.18 3.12 � 0.76Total N (%) 0.37 � 0.01 0.13 � 0.05C:N 11 24N:P 8 20
Non-parasitic litterC:N 36 31N:P 7 18
Hemiparasitic litterC:N 25 26N:P 8 8
Label addition NH4þeN/NO3
�eN (mg container�1)Control 10.0/36.6 10.0/1.0Non-parasitic litter 10.0/18.1 7.2/1.0Hemi-parasitic litter 10.0/3.1 3.7/1.0
a Calculated with particle density¼ %OM*1.25þ (1� %OM)*2.65 g cm�3, where %OM ¼ 1.72*%C (Boyd, 1995).
A. Demey et al. / Soil Biology & Biochemistry 68 (2014) 419e428 421
they fall of before mowing removes the withered stems, and totalPedicularis shoots as only the tips are removed by mowing. Mix-tures of other, non-parasitic species were gathered by clipping totalshoot biomass within a randomly selected plot (1 m2) from whichthe hemiparasite was removed. This was done mid July (Rhin-D) ormid-august (Pedi-H) prior to the regular mowing managementtook place. The four litter types were oven-dried in the laboratory(25 �C) and ground with an ultra centrifugal mill (mesh size 2 mm)(ZM200, Retsch, Germany).
2.3. Soil collection and preparations
One week prior to 15N label additions, fresh mineral soil (5e15 cm) was collected directly under the sod at both sites from three
Fig. 1. Nitrogen (N) pools and transformations considered in the 15N tracing modelthat was used for data analysis. Included pools are organic N (Norg) in soil and litter,extractable ammonium (NH4
þ) and nitrate (NO3þ) and adsorbed ammonium (NH4
þads,
Pedicularis soil only). Fluxes are mineralization of organic N to ammonium (MNorg),immobilization of ammonium to organic N (INH4), oxidation of ammonium to nitrate(ONH4), oxidation of organic N to nitrate (ONorg), immobilization of nitrate to organic N(INO3), adsorption of ammonium (ANH4) and release of adsorbed ammonium (RNH4a).
randomly selected 20 � 20 cm plots. Additionally, three soil coreswere taken for determination of bulk density. Roots were sorted outby hand in the laboratory. For each soil type 99 polypropylenecontainers (180 mL, r ¼ 2.5 cm, h ¼ 10 cm) were weighed, filledwith 60 g fresh soil, and weighed again. A third of the containerswere amendedwith 100mg hemiparasitic litter, another thirdwereamended with 100 mg non-parasitic litter and the remaining thirdserved as unamended controls. All containers were mixed with aspoon after which soil was pressed to a height of 1.9 cm (Pedi-H) or3.2 cm (Rhin-D) to achieve field bulk density. Containers wereweighed and covered with pierced parafilm (7 pinholes) to allowgas exchange but limit evaporation losses. Containers were pre-incubated for one week in a dark room (20 �C) to recover afterdisruption. Every two days parafilm was removed, containers wereweighed and distilled water was dripped onto the soil tocompensate evaporation losses; that way, field gravimetric mois-ture content at the time of soil collection (see Table 1) was main-tained. Meanwhile, we removed seedlings that germinated fromthe seed bank. One container of each treatment was extracted with120 mL 1 M KCl the day before 15N label addition. Concentrations ofNH4
þ and NO3� in the extracts were determined colorimetrically: for
NH4þ the salycilateenitroprusside method (Mulvaney, 1996) was
used on an autoanalyzer (AA3, Bran and Luebbe, Germany), whileNO3
� was determined on the same autoanalyzer in the form of NO2�
after reduction in a CdeCu column followed by the reaction of theNO2 with N-1-napthylethylenediamine to produce a chromophore.The NO3
� results were corrected for NO2� present in the soil samples.
To detect significant trends in NH4þ and NO3
�, a linear regressionanalysis was performed in SPSS 21.
2.4. 15N tracing experiment
The 15N label (� mg 15N g�1 soil) was introduced by adding aNH4CleKNO3 solution, inwhich one of the two Nmoieties was 15N-labeled (99atom% excess), with the ‘mirrored’ moiety at naturalabundance. Application rates of NH4
þ and NO3� (Table 1) were about
20% of the standing pool measured in the test samples, so thattheoretical 15N enrichments for NH4
þ and NO3� were between 15%
and 33% after label application. The added N species were dissolvedin a 1.4 mL solution to each container by inserting a spinal needle(BD, 18G �90 mm) in the soil through a template with 7 evenlyspaced holes, releasing 0.2 mL of the solution while withdrawingthe needle through the soil. This is essential for a uniform distri-bution of the added 15N label (Hart et al., 1994). Soil temperature(20 �C) and moisture content (Table 1) were kept constant duringthe entire experiment. Three containers per treatment (two for thelast two time steps) were extracted 0.25, 1, 4, 24, 72 and 168 h(Rhin-D soil) or 0.25, 4, 24, 72,168 and 312 h (Pedi-H soil) after labeladditionwith 120mL 1M KCl and shaken for 1 h. The NH4
þ and NO3�
concentrations in the extract were determined as above. The 15Ncontents of NH4
þ and NO3� were analyzed after conversion to N2O
using a trace gas preparation unit (ANCA-TGII, PDZ Europa, UK)coupled to an Isotope Ratio Mass Spectrometer (IRMS) (20-20,SerCon, UK). NH4
þwas converted by addingMgO to soil extracts andabsorbing NH3 into H2SO4, after which N2O was produced by re-action with NaOBr (Hauck, 1982; Saghir et al., 1993). NO3
� wasreduced by CdeCu at pH 4.7 to produce NO2
� and NH2OH as in-termediates of N2O (Stevens and Laughlin, 1994). Whenever NH4
þ orNO3
� concentrations in the KCl extract were too low, they werespiked with an NH4Cl or KNO3 solution at natural abundance.
2.5. 15N tracing model
A numerical 15N tracing model was used to quantify multiplegross N transformation rates for each treatment. The 15N tracing

Fig. 2. Net (black) and gross (grayed out) N transformation rates (mg N g�1 day�1) between the different N pools (mg N g�1, boxes) in Rhin-D soil (aec) and Pedi-H soil (def) withoutlitter addition (a, d), amended with a non-parasitic litter (b, e), and amended with hemiparasitic litter (c, f). The arrow widths of the net transformation rates are drawn to scalewithin both soil types for ease of comparison between the three treatments. Skew lines between white and black parts of a pool’s box indicate that this pool increased during theincubation period.
A. Demey et al. / Soil Biology & Biochemistry 68 (2014) 419e428422
model was originally described by Müller et al. (2004). In thepresent experiment, we applied a modified version that relies on aMarkov chain Monte Carlo algorithm for parameter optimization(Müller et al., 2007; Rütting and Müller, 2007). This model enablesto simultaneously quantify gross rates for a variety of N trans-formations described either as zero or first order kinetics. This isdone byminimizing themisfit function (model optimization) in theform of a quadratic weighted error between the observed data andthe model output. Therefore, the average and standard error of themeasured soil NH4
þ and NO3� concentrations and their respective
15N enrichments are used. The model optimization results in aprobability density function for each model parameter, fromwhichaverage parameter values and standard errors (SE) are calculated(Rütting and Müller, 2007; Staelens et al., 2011). The optimizationalgorithm was programmed in MatLab (Version 7.11, The Math-Works Inc.). This algorithm called the 15N tracing model, which wasseparately setup in Simulink (Version 7.6, The MathWorks Inc.).
The three main N pools considered in the tracing model (Fig. 1)were organic N (Norg) in soil organic matter (SOM) and litter, aswell as extractable ammonium (NH4
þ) and nitrate (NO3�). For the
Pedi-H soil, we included an additional pool of adsorbed ammo-nium (NH4
þads), accounting for non-extractable N, which is
assumed to be adsorbed or immobilized quickly to negativelycharged soil particles (e.g. clay minerals, organic matter) (Huygenset al., 2007; Müller et al., 2004; Rütting et al., 2010). The rapidadsorption of NH4
þ was indicated by the rather low recovery(70 � 8%) of 15NH4
þ 15 min after label addition to the Pedi-H soil. Incontrast, 94 � 14% of the added 15NH4
þ was recovered after 15 minin the Rhin-D soil. The considered N transformations are shown inFig. 1. Several modifications in kinetic settings, considered N poolsand included N transformations were tested to identify the model
that best described the measured soil mineral N concentrationsand 15N contents, governed by the Akaike Information Criterion(AIC). A model with a smaller AIC is more likely to be correct and,hence, only modifications decreasing the AIC value were consid-ered for the final data analysis (Burnham and Anderson, 2002).Starting with a basic setup e including only MNorg, INH4, ONH4 andINO3 e model parameters were added or omitted one by one toexamine whether simpler models could describe the measured Ndynamics and to assess the robustness of the obtained gross Nfluxes (Nelissen et al., 2012; Staelens et al., 2011). To allow easycomparison between treatments, the same set of pools andtransformations was used in all three treatments within both soils.In the final model for the Rhin-D soil, three N pools and fivetransformations were retained (Fig. 2); for the Pedi-H soil weselected four N pools and seven transformations. The trans-formations that were not considered in the final model, based onthe AIC, were likely not occurring in the soils and hence the grossrates can be assumed to be zero. The initial (i.e. at t ¼ 0 h) size and15N content of the NH4
þ and NO3� pools were obtained by back-
wards extrapolation of the first two time steps (0.25 and 1 or 4 hdata) (Müller et al., 2004). The initial value of the NH4
þads pool was
calculated according to Münchmeyer (2001). Based on the finalkinetic settings and model parameters, mean gross N fluxes werecalculated by integrating the rates over the total period and sub-sequent division by the total time (Nelissen et al., 2012; Rüttingand Müller, 2007; Staelens et al., 2011). Because of the highnumber of iterations of the 15N-tracing model, statistical tests areinappropriate for the comparison of results. However, an alter-native to a test for significant differences at a ¼ 0.05 is to testwhether the 85% confidence intervals (85% CI) overlap (Paytonet al., 2000; Rutting et al., 2010).
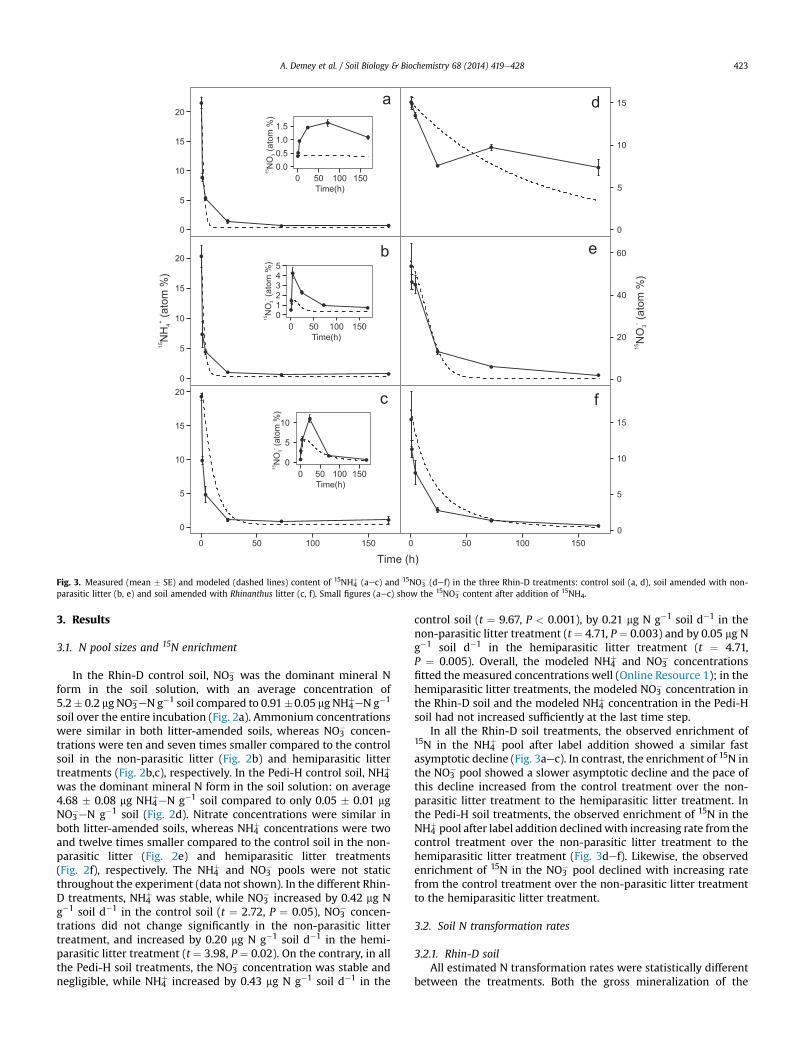
-51
)%
mota(O
N
+51
)%
mota(H
N
)%
mota(O
N)
%mota(
ON
10
15
20
0 50 100 150
0
5
10
0 50 100 150Time(h)
)%
mota(O
N
a
b
c
d
e
f
Fig. 3. Measured (mean � SE) and modeled (dashed lines) content of 15NH4þ (aec) and 15NO3
� (def) in the three Rhin-D treatments: control soil (a, d), soil amended with non-parasitic litter (b, e) and soil amended with Rhinanthus litter (c, f). Small figures (aec) show the 15NO3
� content after addition of 15NH4.
A. Demey et al. / Soil Biology & Biochemistry 68 (2014) 419e428 423
3. Results
3.1. N pool sizes and 15N enrichment
In the Rhin-D control soil, NO3� was the dominant mineral N
form in the soil solution, with an average concentration of5.2� 0.2 mg NO3
�eN g�1 soil compared to 0.91�0.05 mg NH4þeN g�1
soil over the entire incubation (Fig. 2a). Ammonium concentrationswere similar in both litter-amended soils, whereas NO3
� concen-trations were ten and seven times smaller compared to the controlsoil in the non-parasitic litter (Fig. 2b) and hemiparasitic littertreatments (Fig. 2b,c), respectively. In the Pedi-H control soil, NH4
þ
was the dominant mineral N form in the soil solution: on average4.68 � 0.08 mg NH4
þeN g�1 soil compared to only 0.05 � 0.01 mgNO3
�eN g�1 soil (Fig. 2d). Nitrate concentrations were similar inboth litter-amended soils, whereas NH4
þ concentrations were twoand twelve times smaller compared to the control soil in the non-parasitic litter (Fig. 2e) and hemiparasitic litter treatments(Fig. 2f), respectively. The NH4
þ and NO3� pools were not static
throughout the experiment (data not shown). In the different Rhin-D treatments, NH4
þ was stable, while NO3� increased by 0.42 mg N
g�1 soil d�1 in the control soil (t ¼ 2.72, P ¼ 0.05), NO3� concen-
trations did not change significantly in the non-parasitic littertreatment, and increased by 0.20 mg N g�1 soil d�1 in the hemi-parasitic litter treatment (t ¼ 3.98, P ¼ 0.02). On the contrary, in allthe Pedi-H soil treatments, the NO3
� concentration was stable andnegligible, while NH4
þ increased by 0.43 mg N g�1 soil d�1 in the
control soil (t ¼ 9.67, P < 0.001), by 0.21 mg N g�1 soil d�1 in thenon-parasitic litter treatment (t ¼ 4.71, P ¼ 0.003) and by 0.05 mg Ng�1 soil d�1 in the hemiparasitic litter treatment (t ¼ 4.71,P ¼ 0.005). Overall, the modeled NH4
þ and NO3� concentrations
fitted the measured concentrations well (Online Resource 1); in thehemiparasitic litter treatments, the modeled NO3
� concentration inthe Rhin-D soil and the modeled NH4
þ concentration in the Pedi-Hsoil had not increased sufficiently at the last time step.
In all the Rhin-D soil treatments, the observed enrichment of15N in the NH4
þ pool after label addition showed a similar fastasymptotic decline (Fig. 3aec). In contrast, the enrichment of 15N inthe NO3
� pool showed a slower asymptotic decline and the pace ofthis decline increased from the control treatment over the non-parasitic litter treatment to the hemiparasitic litter treatment. Inthe Pedi-H soil treatments, the observed enrichment of 15N in theNH4
þ pool after label addition declinedwith increasing rate from thecontrol treatment over the non-parasitic litter treatment to thehemiparasitic litter treatment (Fig. 3def). Likewise, the observedenrichment of 15N in the NO3
� pool declined with increasing ratefrom the control treatment over the non-parasitic litter treatmentto the hemiparasitic litter treatment.
3.2. Soil N transformation rates
3.2.1. Rhin-D soilAll estimated N transformation rates were statistically different
between the treatments. Both the gross mineralization of the

A. Demey et al. / Soil Biology & Biochemistry 68 (2014) 419e428424
organic N pool to the NH4þ pool (MNorg) and the gross immobiliza-
tion of NH4þ back to organic N (INH4) were highest in the control soil,
about 35% smaller in the non-parasitic litter treatment and about80% smaller in the hemiparasitic litter treatment compared to thecontrol soil (Fig. 2aec, Table 2). In contrast, the net N flux fromorganic N to NH4
þ (MNorgeINH4) was an order of magnitude higher inboth litter addition treatments compared to the control soil, being61% higher in the hemiparasitic litter treatment compared to thenon-parasitic litter treatment. Also the oxidation of NH4
þ to NO3�
(ONH4) was higher in both litter addition treatments compared tothe control soil, and was 80% higher in the hemiparasitic littertreatment compared to the non-parasitic litter treatment. Oxida-tion of organic N direct to NO3
� (ONrec) occurred in the control soil,but nearly ceased in the litter addition treatments. Gross immobi-lization of NO3
� to organic N (INO3) was similar toONrec in the controltreatment and was 60% and 54% smaller in the non-parasitic littertreatment and the hemiparasitic litter treatment respectively. Allthree treatments had a net flux from NO3
� to organic N (INO3eONrec > zero), but this N flux was an order of magnitude higher inthe litter addition treatments compared to the control soil. For thehemiparasitic litter treatment, the net flux from NO3
� to organic Nwas 24% higher compared to the non-parasitic litter treatment.
3.2.2. Pedi-H soilAll estimated transformation rates were statistically different
between the treatments, except for ONrec and INO3 that did not differbetween the control soil and the non-parasitic litter treatment. Thegross mineralization of organic N to NH4
þ (MNorg) increased fromcontrol soil over the non-parasitic litter treatment (þ34%) to thehemiparasitic litter treatment (þ71%) (Fig. 2def, Table 2). Grossimmobilization of NH4
þ back to organic N (INH4) only occurred in thecontrol soil. Therefore, the net flux from organic N to NH4
þ in thelitter addition treatments was substantially higher than in thecontrol soil (þ203% for the non-parasitic litter treatmentand þ287% for the hemiparasitic litter treatment). In the hemi-parasitic litter treatment, this net flux was 28% higher compared tothe non-parasitic litter treatment. Oxidation of NH4
þ to NO3� (ONH4)
was negligible in all three treatments. The adsorption or immobi-lization of extractable NH4
þ to adsorbed NH4þ (ANH4) and the release
Table 2Gross N transformation rates (mean and SD) estimated by the 15N tracing model forthe three treatments for both soils. Rhin-D: soil from site ‘Doode Bemde’ with litterfrom Rhinanthus and its co-occurring, non-parasitic species; Pedi-H: soil from site‘Hooiput’ with litter from Pedicularis and its co-occurring, non-parasitic species.Fluxes are mineralization of organic N to ammonium (MNorg), immobilization ofammonium to organic N (INH4), oxidation of ammonium to nitrate (ONH4), oxidationof organic N to nitrate (ONorg), immobilization of nitrate to organic N (INO3),adsorption of ammonium (ANH4) and release of adsorbed ammonium (RNH4a).
Abbreviation Kinetics N transformation rate (mg N g�1 day�1)
Control Non-parasitic litter Hemiparasitic litter
Mean SD Mean SD Mean SD
Rhin-DMNorg 0 9.70 0.49 6.36 0.38 2.18 0.10INH4 1st 9.66 0.48 6.05 0.38 1.68 0.06ONH4 0 0.10 0.01 0.30 0.02 0.54 0.01ONorg 0 0.98 0.09 0.02 0.02 <0.01 <0.01INO3 1st 1.02 0.10 0.40 0.02 0.47 0.01
Pedi-HMNorg 0 2.06 0.03 2.76 0.11 3.52 0.22INH4 1st 1.15 0.07 e e e e
ONH4 1st 0.02 <0.01 e e e e
ONorg 0 0.16 0.02 0.16 0.03 0.23 0.01INO3 1st 0.18 0.02 0.16 0.03 0.22 0.01ANH4 1st 2.23 0.13 6.51 0.33 6.96 0.58RNH4 1st 1.67 0.15 3.92 0.25 3.47 0.89
� Denotes transformations not considered in the final model.
of adsorbed NH4þ (RNH4a) were higher in the litter addition treat-
ments than in the control soil. The net NH4þ adsorption (ANH4e
RNH4a) was about five times higher in the litter addition treatments,being 35% higher in the hemiparasitic litter treatment compared tothe non-parasitic litter treatment. Direct oxidation of organic N toNO3
� (ONrec) and gross NO3� immobilization to organic N (INO3) were
similarly small in all treatments, with a negligible net flux fromNO3
� to organic N (INO3eONrec).
4. Discussion
4.1. Two contrasting soil types
In a conceptual N model proposed by Schimel and Bennett(2004), the dominant available N form shifts from organic Nmonomers over NH4
þ to NO3� as N availability increases. According
to this model, the oligotrophic Pedi-H soil is situated in the zonewith moderately low N supply with NH4
þ being the most importantDIN form and nitrification being negligible (Schimel and Bennett,2004). On the other hand, the mesotrophic Rhin-D soil is rathersituated in the moderately high zone of N supply, where NH4
þ is stillthe most important DIN form (looking at gross production rates),but also some nitrification occurs.
The gross mineralization rate (MNorg) in the mesotrophic Rhin-Dsoil (9.7 mg g�1 soil, Table 2, Fig. 2a) is moderately high compared tothose generally observed for grassland soils; while measuredvalues in isotope dilution studies range from 0.3 to 44 mg g�1, 86% isbelow 10 mg g�1 soil (Booth et al., 2005). Nearly all mineralized NH4
þ
is immobilized again in microbial biomass (INH4), resulting in asmall net flux from organic N to NH4
þ. The (autotrophic) nitrificationrate (ONH4) in the Rhin-D soil (0.1 mg g�1 soil, Table 2, Fig. 2a) is verysmall compared to the 0.1e7 mg g�1 range observed in other isotopedilution studies for grassland soils (Booth et al., 2005). However, asthe 15N traced in the NO3
� pool after 15NH4þ addition was not
modeled well (Fig. 3a, small figure), the nitrification rate might beunderestimated here. But, as modeled, 91% of NO3
� is produced bydirect oxidation of organic N (ONorg). Microbial immobilization ofNO3
� (INO3) is lower than NO3� production (ONorgþ ONH4). As a result,
the NO3� pool increased throughout the experiment (þ0.42 mg g�1
soil d�1). Altogether, this build-up of DIN in the soil solution in-dicates that N, relative to C, is excessively available for microbialgrowth.
In contrast to the mesotrophic Rhin-D soil, the gross minerali-zation rate in the oligotrophic Pedi-H soil (2.06 mg g�1 soil, Table 2,Fig. 2d) is in the lower range of other studies (Booth et al., 2005). Asonly 56% is immobilized again in microbial biomass, the net fluxfrom organic N to NH4
þ is much higher compared to the Rhin-D soil.Nearly no (autotrophic) nitrification (ONH4) occurs in the Pedi-H soil(0.02 mg g�1 soil, Table 2, Fig. 2d), likely due to the high soil acidity(pH 4.59): below a pH of 4.5, nitrification becomes negligible (Hartet al., 1994). Instead, NH4
þ is rapidly adsorbed or immobilized to theNH4
þads pool (and released again), with a concurrent net adsorption.
This rapid adsorption (or immobilization) of NH4þ is presumably
explained by the high organic matter content of the Pedi-H(96 � 16 mg C g�1) with a high C:N ratio (24 � 1) and could berelated to a high polyphenolics:N ratio (e.g., Hattenschwiler andVitousek, 2000) typically found in acid dwarf-shrub rich vegeta-tions similar to that of the Pedi-H site (Eskelinen et al., 2009). Inaddition, the Pedi-H site is poorly drained, potentially increasingthe polyphenolic content. Like ONH4, the other N fluxes related toNO3
� (ONorg, INO3) are small as well, with concurrent negligible netNO3
� immobilization. Throughout the experiment, the NH4þ pool
increased by a similar rate (0.43 mg g�1 soil d�1) as NO3� did in the
Rhin-D soil. Again, this build-up of DIN in the soil solution indicatesthat N is excessively available for microbial growth. While there

A. Demey et al. / Soil Biology & Biochemistry 68 (2014) 419e428 425
was a build-up of NO3� (and not NH4
þ) in the Rhin-D soil, there was abuild-up of NH4
þ (and not NO3�) in the Pedi-H soil.
Microbial activity reaches its optimum at a soil water-filled porespace (WFPS) in the range of 40e60% (Franzluebbers, 1999). Duringthe incubation, WFPS was slightly lower, being 33% for Rhin-D and38% for Pedi-H (Table 1). The similar values suggest that soilmoisture only had a marginal influence on differences in N cyclingprocesses between both soils.
4.2. Effects of Rhinanthus litter on soil N dynamics
In contrast to expectations, litter addition decreased grossmineralization and NH4
þ immobilization rates (Fig. 2aec, Table 2). Apossible explanation is the discrepancy in C:N ratio between thesoil and the added litter (Table 1). The soil has a C:N ratio (11:1)within the range of 8:1 to 12:1 typical for soil microbes (Wright andColeman, 2000).When (hemiparasitic) litter with a higher C:N ratiois added to the soil, microbes will cope with the increased Cavailability by using available N sources in a more efficient way. Inother words, micro-organisms will increase their nitrogen use ef-ficiency (NUE) in order to enhance their metabolism and maintainhomeostasis (i.e. greater microbial investments in N acquisitionenzymes for N assimilation, reduced microbial investment in Nrespiration) (Griffiths et al., 2012; Mooshammer et al., 2012).Concurrently, they may excrete redundant C by decreasing their Cuse efficiency (overflow mechanism, Manzoni and Porporato,2009). The N is thus, in the short term, tied up into the microbialbiomass, leading to reduced gross N mineralization and immobili-zation rates. In the long term, a further increase in microbialbiomass as a result of litter addition will likely result in enhancedmicrobial turnover, generating labile N inputs that might increase Nbioavailability. The differences between Rhinanthus litter and thenon-parasitic litter may be attributed to the quality of the litter; ifthe higher C:N ratio in the non-parasitic litter is related to a highercontent of secondary compound such as polyphenols and lignin, Cmight actually be less bioavailable compared to the hemiparasiticlitter (Cornwell et al., 2008; Quested et al., 2003a).
In contrast to expectations based on stoichiometry(Mooshammer et al., 2012), litter additionwith a relatively high C:Nratio (compared to soil C:N) increased (autotrophic) nitrification. Apossible explanation is the increased net mineralization, whichleads to an enhanced NH4
þ availability for nitrifiers. Increasednitrification together with a tenfold increase of the net flux fromorganic N to NH4
þ and the net immobilization of NO3� indicates an
increased cycling from organic N over NH4þ and NO3
� back to organicN. This increase in N cycling is higher in the hemiparasitic littertreatment compared to the non-parasitic litter. We suggest thathemiparasitic litter increases N cycling more than the non-parasiticlitter as a result of its higher decomposability leading to highermicrobial activity. Hemiparasitic litter was indeed shown todecompose faster than litter from many e but not all e co-occurring species in sub-arctic and alpine habitats (Quested et al.,2003a; Spasojevic and Suding, 2011). In a previous study (Demeyet al., 2013a), we found that Rhinanthus leaf litter decomposedextremely fast e losing 93% of its mass in eight months. Litter massloss in a similar period (nine months) of graminoids (50%) and non-parasitic forbs (63%) at the same sites was considerably smaller(Ameloot et al. unpublished).
The N that is potentially available for plantemicrobe competi-tion (defined as the gross production of NH4
þ and NO3�) is domi-
nated by the gross mineralization to NH4þ, which is lower in the
hemiparasitic litter treatment compared to the non-parasitic littertreatment. However, the microbial mineralization-immobilizationturnover (MIT, MNorg and INH4) measured by pool dilution experi-ments may reflect microbial cycling and recycling of small pools of
highly labile, N-rich compounds rather than the overall breakdownof soil organic matter (Fierer et al., 2001). Since it is the latter thatregulates overall N cycling (Schimel and Bennett, 2004), microbialMITmay be a poor predictor of N availability to plants. In the case oflitter addition, net mineralization (MNreceINH4eONH4) is not a goodpredictor either, as it is highly dependent on the particular momentof measurement e often with initial net immobilization changingin time to net mineralization (Manzoni et al., 2008). For thesereasons, we propose that the NorgeNH4
þeNO3�eNorg cycling
(Fig. 2bec) e increasing from control over non-parasitic litter toRhinanthus litter e best represents the relative differences in plant-available N between treatments. The increased turnover of NO3
� byhemiparasitic litter in particular might be important for plant up-take, as plants compete more effectively with microbes for themore mobile NO3
�, in contrast to NH4þ (Recous et al., 1988). Together,
our results suggest that considerably more N is available for plantemicrobe competition in soil with Rhinanthus litter compared to soilwith the non-parasitic litter. Moreover, the higher NO3
� immobili-zation (INO3) in the hemiparasitic litter treatment can enhance Nsupply in the long term by avoiding N losses (leakage, denitrifica-tion), and ultimately increases the active organic N pool (Recouset al., 1988). Where NO3
� immobilization was in the pastcommonly thought to be minimal (Myrold and Tiedje, 1986; Tiedjeet al., 1981), NO3
� immobilization rates approaching gross nitrifi-cation rates have been reported for agricultural, grassland andforest soils (Burger and Jackson, 2003; Hatch et al., 2000; Stark andHart, 1997).
4.3. Effects of Pedicularis litter on soil N dynamics
Litter addition increased both gross and net mineralization ratescompared to the control soil (Fig. 2def, Table 2). Since no NH4
þ
immobilization to organic N occurred in the litter-amended soils,the net flux fromorganic N to NH4
þ equaled the grossmineralizationrate; the mineralization rate was 28% higher in the hemiparasiticlitter treatment compared to the non-parasitic litter treatment. 93%and 99% of mineralized N ended up in the adsorbed NH4
þ pool in thenon-parasitic litter and hemiparasitic litter treatment respectively.Since NH4
þ adsorption is thought to occur because of soil organicmatter, we had expected adsorption rates to be similar in the threetreatments or, alternatively, to increase with standing NH4
þ pool. Incontrast, gross and net NH4
þ adsorption rates were higher at lowerNH4
þ levels. While it is possible that some of the NH4þ is adsorbed to
the added litter, it is more likely that the adsorbed NH4þ pool
(NH4þads) also includes rapid microbial MIT related to microbial
cellular processes.The N available for plantemicrobe competition (equal to the
gross production of NH4þ as NO3
� turnover is negligible) is the sum ofthe fluxes from organic N and NH4
þads to NH4
þ (MNorg þ RNH4a). Thisgross production of NH4
þ is significantly higher (6.99 � 0.10) for thePedicularis litter treatment compared to the non-parasitic littertreatment (6.68 � 0.08). However, as discussed above for Rhinan-thus, the fast microbial MIT is related to microbial cellular processesrather than the overall breakdown of organic N and therefore not agood predictor of plant-available N (Fierer et al., 2001; Schimel andBennett, 2004). As the turnover of the NH4
þads in the litter treat-
ments is likely to include microbial MIT in addition to abioticadsorption-release processes, we suggest it is better to consider theNorgeNH4
þeNH4þads turnovere increasing from control soil over the
non-parasitic litter treatment to the hemiparasitic litter treatmente as an indicator for the effect on plant-available N. We can think ofit as a net transition from a more recalcitrant organic N pool (Norg)to a highly labile organic and adsorbed N pool (NH4
þads). Therefore,
N will likely be more available to plants, at least in the long term.Together, our results suggest that N is more available for plant
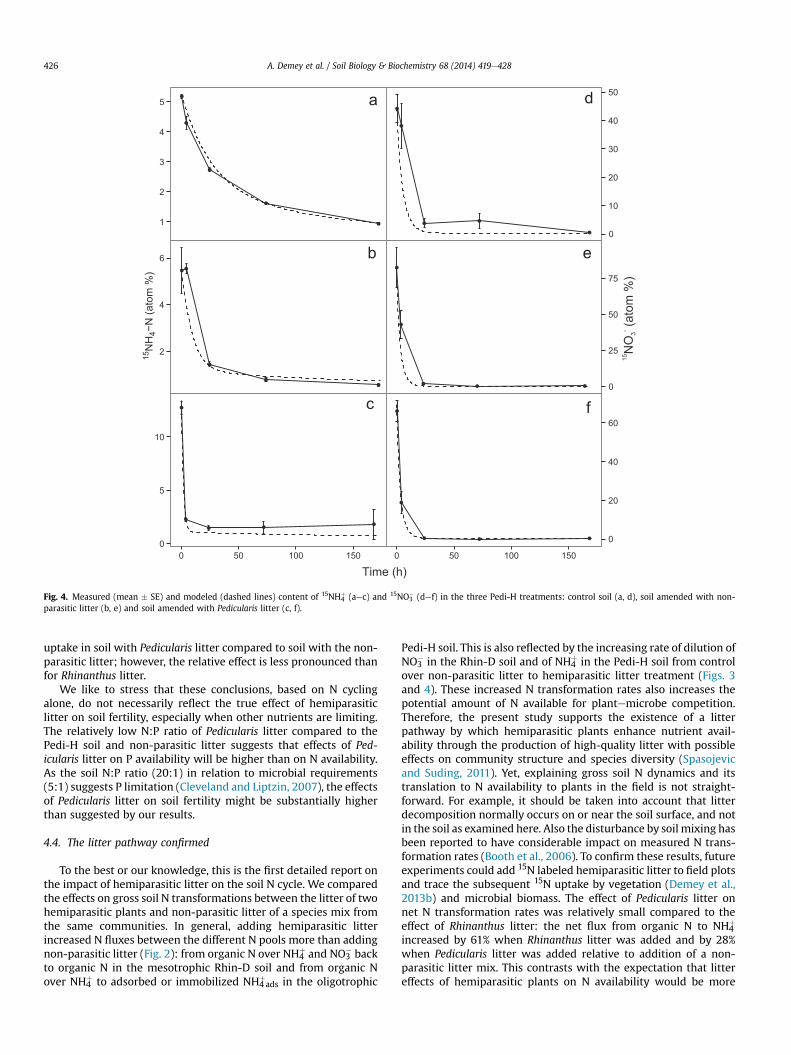
Time (h)
1
2
3
4
5
2
4
6
15N
H4−
N (a
tom
%)
0
5
10
0 50 100 150
0
10
20
30
40
50
-51
)%
mota(O
N3
0
25
50
75
0
20
40
60
0 50 100 150
a d
b
c
e
f
Fig. 4. Measured (mean � SE) and modeled (dashed lines) content of 15NH4þ (aec) and 15NO3
� (def) in the three Pedi-H treatments: control soil (a, d), soil amended with non-parasitic litter (b, e) and soil amended with Pedicularis litter (c, f).
A. Demey et al. / Soil Biology & Biochemistry 68 (2014) 419e428426
uptake in soil with Pedicularis litter compared to soil with the non-parasitic litter; however, the relative effect is less pronounced thanfor Rhinanthus litter.
We like to stress that these conclusions, based on N cyclingalone, do not necessarily reflect the true effect of hemiparasiticlitter on soil fertility, especially when other nutrients are limiting.The relatively low N:P ratio of Pedicularis litter compared to thePedi-H soil and non-parasitic litter suggests that effects of Ped-icularis litter on P availability will be higher than on N availability.As the soil N:P ratio (20:1) in relation to microbial requirements(5:1) suggests P limitation (Cleveland and Liptzin, 2007), the effectsof Pedicularis litter on soil fertility might be substantially higherthan suggested by our results.
4.4. The litter pathway confirmed
To the best or our knowledge, this is the first detailed report onthe impact of hemiparasitic litter on the soil N cycle. We comparedthe effects on gross soil N transformations between the litter of twohemiparasitic plants and non-parasitic litter of a species mix fromthe same communities. In general, adding hemiparasitic litterincreased N fluxes between the different N pools more than addingnon-parasitic litter (Fig. 2): from organic N over NH4
þ and NO3� back
to organic N in the mesotrophic Rhin-D soil and from organic Nover NH4
þ to adsorbed or immobilized NH4þads in the oligotrophic
Pedi-H soil. This is also reflected by the increasing rate of dilution ofNO3
� in the Rhin-D soil and of NH4þ in the Pedi-H soil from control
over non-parasitic litter to hemiparasitic litter treatment (Figs. 3and 4). These increased N transformation rates also increases thepotential amount of N available for plantemicrobe competition.Therefore, the present study supports the existence of a litterpathway by which hemiparasitic plants enhance nutrient avail-ability through the production of high-quality litter with possibleeffects on community structure and species diversity (Spasojevicand Suding, 2011). Yet, explaining gross soil N dynamics and itstranslation to N availability to plants in the field is not straight-forward. For example, it should be taken into account that litterdecomposition normally occurs on or near the soil surface, and notin the soil as examined here. Also the disturbance by soil mixing hasbeen reported to have considerable impact on measured N trans-formation rates (Booth et al., 2006). To confirm these results, futureexperiments could add 15N labeled hemiparasitic litter to field plotsand trace the subsequent 15N uptake by vegetation (Demey et al.,2013b) and microbial biomass. The effect of Pedicularis litter onnet N transformation rates was relatively small compared to theeffect of Rhinanthus litter: the net flux from organic N to NH4
þ
increased by 61% when Rhinanthus litter was added and by 28%when Pedicularis litter was added relative to addition of a non-parasitic litter mix. This contrasts with the expectation that littereffects of hemiparasitic plants on N availability would be more

A. Demey et al. / Soil Biology & Biochemistry 68 (2014) 419e428 427
important in oligotrophic sites (Pedicularis) compared to mesotro-phic sites (Rhinanthus), but note that the soil type and drainage areconfounded with litter types. Future research should also focus onother nutrients such as P and K, in addition to N.
Acknowledgments
We thank Sandy Adriaenssens for assistance with label addi-tions and extractions; Katja Van Nieuland and Jan Vermeulen forthe analyses. This research was funded by the Research Foundatione Flanders (FWO, G.0296.07). AD was supported by a PhD grant ofthe agency for Innovation by Science and Technology (IWT); TRwassupported by the strategic research area Biodiversity andEcosystem services in a Changing Climate (BECC, http://www.cec.lu.se/research/becc); DH was funded as postdoctoral fellow by theEuropean Commission (International Outgoing Fellowships forcareer development (IOF) e Marie Curie Actions, PIOF-GA-2011-301443) and the Research Foundation e Flanders (FWO); JS wasfunded as postdoctoral fellow by the Research Foundation e Flan-ders (FWO).
Appendix A. Supplementary material
Supplementary material related to this article can be found athttp://dx.doi.org/10.1016/j.soilbio.2013.10.025.
References
Ameloot, E., Verheyen, K., Hermy, M., 2005. Meta-analysis of standing crop reduc-tion by Rhinanthus spp. and its effect on vegetation structure. Folia Geobot. 40,289e310.
Ameloot, E., Verlinden, G., Boeckx, P., Verheyen, K., Hermy, M., 2008. Impact ofhemiparasitic Rhinanthus angustifolius and R. minor on nitrogen availability ingrasslands. Plant Soil 311, 255e268.
Bardgett, R.D., Smith, R.S., Shiel, R.S., Peacock, S., Simkin, J.M., Quirk, H., Hobbs, P.J.,2006. Parasitic plants indirectly regulate below-ground properties in grasslandecosystems. Nature 439, 969e972.
Booth, M.S., Stark, J.M., Hart, S.C., 2006. Soil-mixing effects on inorganic nitrogenproduction and consumption in forest and shrubland soils. Plant Soil 289, 5e15.
Booth, M.S., Stark, J.M., Rastetter, E., 2005. Controls on nitrogen cycling in terrestrialecosystems: a synthetic analysis of literature data. Ecol. Monogr. 75, 139e157.
Bottomley, P.J., Taylor, A.E., Myrold, D.D., 2012. A consideration of the relativecontributions of different microbial subpopulations to the soil N cycle. FrontMicrobiol. 3, 373.
Boyd, C.E., 1995. Bottom Soils, Sediment, and Pond Aquaculture. Chapman & Hall,New York, p. 348.
Burger, M., Jackson, L.E., 2003. Microbial immobilization of ammonium and nitratein relation to ammonification and nitrification rates in organic and conven-tional cropping systems. Soil Biol. Biochem. 35.
Burnham, K.P., Anderson, D.R., 2002. Model Selection and Inference: A PracticalInformation-Theoretic Approach. Springer-Verlag, New York.
Cleveland, C.C., Liptzin, D., 2007. C: N: P stoichiometry in soil: is there a “Redfieldratio” for the microbial biomass? Biogeochemistry 85, 235e252.
Cornwell, W.K., Cornelissen, J.H.C., Amatangelo, K., Dorrepaal, E., Eviner, V.T.,Godoy, O., Hobbie, S.E., Hoorens, B., Kurokawa, H., Perez-Harguindeguy, N.,Quested, H.M., Santiago, L.S., Wardle, D.A., Wright, I.J., Aerts, R., Allison, S.D., vanBodegom, P., Brovkin, V., Chatain, A., Callaghan, T.V., Diaz, S., Garnier, E.,Gurvich, D.E., Kazakou, E., Klein, J.A., Read, J., Reich, P.B., Soudzilovskaia, N.A.,Vaieretti, M.V., Westoby, M., 2008. Plant species traits are the predominantcontrol on litter decomposition rates within biomes worldwide. Ecol. Lett. 11,1065e1071.
Davies, D.M., Graves, J.D., Elias, C.O., Williams, P.J., 1997. The impact of Rhinanthusspp. on sward productivity and composition: Implications for the restoration ofspecies-rich grasslands. Biol. Conserv. 82, 87e93.
De Schrijver, A., De Frenne, P., Ampoorter, E., Van Nevel, L., Demey, A., Wuyts, K.,Verheyen, K., 2011. Cumulative nitrogen input drives species loss in terrestrialecosystems. Glob. Ecol. Biogeogr. 20, 803e816.
Demey, A., Ameloot, E., Staelens, J., De Schrijver, A., Verstraeten, G., Boeckx, P.,Hermy, M., Verheyen, K., 2013a. Effects of two contrasting hemiparasitic plantspecies onbiomass production andnitrogenavailability.Oecologia 173, 293e303.
Demey, A., Staelens, J., Baeten, L., Boeckx, P., Hermy, M., Kattge, J., Verheyen, K.,2013b. Nutrient input from hemiparasitic litter favors plant species with a fast-growth strategy. Plant Soil 371, 53e66.
Eskelinen, A., Stark, S., Mannisto, M., 2009. Links between plant communitycomposition, soil organic matter quality and microbial communities in con-trasting tundra habitats. Oecologia 161, 113e123.
Fierer, N., Schimel, J.P., Cates, R.G., Zou, J.P., 2001. Influence of balsam poplar tanninfractions on carbon and nitrogen dynamics in Alaskan taiga floodplain soils. SoilBiol. Biochem. 33, 1827e1839.
Franzluebbers, A.J., 1999. Microbial activity in response to water-filled pore space ofvariably eroded southern Piedmont soils. Appl. Soil Ecol. 11, 91e101.
Gauslaa, Y., 1990. Water relations and mineral nutrients in Melampyrum pratense(Scrophulariaceae) in oligotrophic and mesotrophic boreal forests. Acta Oecol.Int. J. Ecol. 11, 525e537.
Gauslaa, Y., Odasz, A.M., 1990. Water relations, temperatures, and mineral nutrientsin Pedicularis dasyantha (Scrophulariaceae) from Svalbard, Norway. HolarcticEcol. 13, 112e121.
Gibson, C.C., Watkinson, A.R., 1991. Host selectivity and the mediation of compe-tition by the root hemiparasite Rhinanthus minor. Oecologia 86, 81e87.
Griffiths, B., Spilles, A., Bonkowski, M., 2012. C:N:P stoichiometry and nutrientlimitation of the soil microbial biomass in a grazed grassland site underexperimental P limitation or excess. Ecol. Process. 1.
Hart, S.C., Stark, J.M., Davidson, E.A., Firestone, M.K., 1994. Nitrogen mineralization,immobilization, and nitrification. In: Weaver, R.W., Angle, S., Bottomley, P.,Bezdicek, D., Smith, S., Tabatabi, A., Wollum, A. (Eds.), Methods of Soil Analysis;Part 2. Microbiological and Biochemical Properties. Soil Science Society ofAmerica, Madison, WI, pp. 985e1018.
Hatch, D.J., Jarvis, S.C., Parkinson, R.J., Lovell, R.D., 2000. Combining field incubationwith nitrogen-15 labelling to examine nitrogen transformations in low to highintensity grassland management systems. Biol. Fertil. Soils 30.
Hattenschwiler, S., Vitousek, P.M., 2000. The role of polyphenols in terrestrialecosystem nutrient cycling. Trends Ecol. Evol. 15, 238e243.
Hauck, R.D., 1982. Nitrogen isotope ratio analysis. In: Page, A.L., Miller, R.A.,Keeney, D.R. (Eds.), Methods of Soil Analysis, pp. 735e779. Madison, WI.
Hejcman, M., Klaudisová, M., Schellberg, J., Honsová, D., 2007. The Rengen Grass-land Experiment: plant species composition after 64 years of fertilizer appli-cation. Agric. Ecosyst. Environ. 122, 259e266.
Huygens, D., 2007. Refining the paradigm of terrestrial nitrogen cycling intemperate evergreen forests (PhD thesis). Ghent University, Ghent.
Manzoni, S., Jackson, R.B., Trofymow, J.A., Porporato, A., 2008. The global stoichi-ometry of litter nitrogen mineralization. Science 321, 684e686.
Manzoni, S., Porporato, A., 2009. Soil carbon and nitrogen mineralization: theoryand models across scales. Soil Biol. Biochem. 41, 1355e1379.
March, W.A., Watson, D.M., 2007. Parasites boost productivity: effects of mistletoeon litterfall dynamics in a temperate Australian forest. Oecologia 154, 339e347.
March, W.A., Watson, D.M., 2010. The contribution of mistletoes to nutrient returns:evidence for a critical role in nutrient cycling. Aust. Ecol. 35, 713e721.
Mary, B., Recous, S., Robin, D., 1998. A model for calculating nitrogen fluxes in soilusing 15N tracing. Soil Biol. Biochem. 30, 1936e1979.
Matthies, D., 1996. Interactions between the root hemiparasiteMelampyrum arvenseand mixtures of host plants: heterotrophic benefit and parasite-mediatedcompetition. Oikos 75, 118e124.
Mooshammer, M., Wanek, W., Schnecker, J., Wild, B., Leitner, S., Hofhansl, F.,Blochl, A., Hammerle, I., Frank, A.H., Fuchslueger, L., Keiblinger, K.M.,Zechmeister-Boltenstern, S., Richter, A., 2012. Stoichiometric controls of nitro-gen and phosphorus cycling in decomposing beech leaf litter. Ecology 93, 770e782.
Müller, C., Rütting, T., Kattge, J., Laughlin, R.J., Stevens, R.J., 2007. Estimation ofparameters in complex 15N tracing models by Monte Carlo sampling. Soil Biol.Biochem. 39, 715e726.
Müller, C., Stevens, R.J., Laughlin, R.J., 2004. A 15N tracing model to analyse Ntransformations in old grassland soil. Soil Biol. Biochem. 36, 619e632.
Mulvaney, R.L., 1996. Nitrogen-inorganic forms. In: Sparks, D.L. (Ed.), Methods ofSoil Analysis. American Society of Agronomy, Madison, WI, pp. 1123e1184.
Münchmeyer, U., 2001. Zur N-Umsetzung in degradierten Niedermoorböden Nor-dostdeutschlands unter besonderer Berücksichtigung der N-Mineralisierungund des Austrages gasförmiger N-Verbindungen. Beiträge aus Hallen. Pflanze-nernährungsforsch. 5, 1e125.
Myrold, D.D., Tiedje, J.M., 1986. Simultaneous estimation of several nitrogen-cyclerates using N-15 e theory and application. Soil Biol. Biochem. 18.
Nelissen, V., Rütting, T., Huygens, D., Staelens, J., Ruysschaert, G., Boeckx, P., 2012.Maize biochars accelerate short-term soil nitrogen dynamics in a loamy sandsoil. Soil Biol. Biochem. 55, 20e27.
Pate, J.S., 1995. Mineral relationships of parasites and their hosts. In: Press, M.C.,Graves, J.D. (Eds.), Parasitic Plants. Chapman & Hall, London, pp. 80e102.
Payton, M.E., Miller, A.E., Raun, W.R., 2000. Testing statistical hypotheses usingstandard error bars and confidence intervals. Commun. Soil Sci. Plant Anal. 31,547e551.
Phoenix, G.K., Press, M.C., 2005. Linking physiological traits to impacts on com-munity structure and function: the role of root hemiparasitic Orobanchaceae(ex-Scrophulariaceae). J. Ecol. 93, 67e78.
Press, M.C., 1998. Dracula or Robin Hood? A functional role for root hemiparasites innutrient poor ecosystems. Oikos 82, 609e611.
Press, M.C., Phoenix, G.K., 2005. Impacts of parasitic plants on natural communities.New Phytol. 166, 737e751.
Press, M.C., Scholes, J.D., Keay, R.W.J., 1999. Physiological plant ecology. In:Press, M.C., Scholes, J.D., Barker, M.G. (Eds.), Physiological Plant Ecology.Blackwell, Oxford, pp. 175e197.
Quested, H.M., Callaghan, T.V., Cornelissen, J.H.C., Press, M.C., 2005. The impact ofhemiparasitic plant litter on decomposition: direct, seasonal and litter mixingeffects. J. Ecol. 93, 87e98.

A. Demey et al. / Soil Biology & Biochemistry 68 (2014) 419e428428
Quested, H.M., Cornelissen, J.H.C., Press, M.C., Callaghan, T.V., Aerts, R., Trosien, F.,Riemann, P., Gwynn-Jones, D., Kondratchuk, A., Jonasson, S.E., 2003a. Decom-position of sub-arctic plants with differing nitrogen economies: a functionalrole for hemiparasites. Ecology 84, 3209e3221.
Quested, H.M., Press, M.C., Callaghan, T.V., 2003b. Litter of the hemiparasite Bartsiaalpina enhances plant growth: evidence for a functional role in nutrient cycling.Oecologia 135, 606e614.
Quested, H.M., Press, M.C., Callaghan, T.V., Cornelissen, J.H.C., 2002. The hemi-parasitic angiosperm Bartsia alpina has the potential to accelerate decomposi-tion in sub-arctic communities. Oecologia 130, 88e95.
Recous, S., Fresneau, C., Faurie, G., Mary, B., 1988. The fate of labeled N-15 urea andammonium-nitrate applied to a winter-wheat crop. 1. Nitrogen transformationsin the soil. Plant Soil 112.
Rutting, T., Clough, T.J., Muller, C., Lieffering, M., Newton, P.C.D., 2010. Ten years ofelevated atmospheric carbon dioxide alters soil nitrogen transformations in asheep-grazed pasture. Glob. Change Biol. 16, 2530e2542.
Rutting, T., Huygens, D., Staelens, J., Muller, C., Boeckx, P., 2011. Advances in 15N-tracing experiments: new labelling and data analysis approaches. Biochem. Soc.Trans. 39, 279e283.
Rütting, T., Müller, C., 2007. 15N tracing models with a Monte Carlo optimizationprocedure provide new insights on gross N transformations in soils. Soil Biol.Biochem. 39, 2351e2361.
Saghir, N.S., Mulvaney, R.L., Azam, F., 1993. Determination of nitrogen by micro-diffusion in mason jars. 1. Inorganic nitrogen in soil extracts. Commun. Soil Sci.Plant Anal. 24, 1745e1762.
Schimel, J.P., Bennett, J., 2004. Nitrogen mineralization: challenges of a changingparadigm. Ecology 85, 591e602.
Seel, W.E., Press, M.C., 1993. Influence of the host on 3 sub-arctic annual facultativeroot hemiparasites. 1. Growth, mineral accumulation and aboveground dry-matter partitioning. New Phytol. 125, 131e138.
Silvertown, J., Poulton, P., Johnston, E., Edwards, G., Heard, M., Biss, P.M., 2006. ThePark Grass experiment 1856e2006: Its contribution to ecology. J. Ecol. 94, 801e814.
Spasojevic, M., Suding, K., 2011. Contrasting effects of hemiparasites on ecosystemprocesses: can positive litter effects offset the negative effects of parasitism?Oecologia 165, 193e200.
Staelens, J., Rütting, T., Huygens, D., Schrijver, A., Müller, C., Verheyen, K., Boeckx, P.,2011. In situ gross nitrogen transformations differ between temperate decidu-ous and coniferous forest soils. Biogeochemistry 108, 259e277.
Stark, J.M., Hart, S.C., 1997. High rates of nitrification and nitrate turnover in un-disturbed coniferous forests. Nature 385.
Stevens, R.J., Laughlin, R.J., 1994. Determining N-15 in nitrite or nitrate by producingnitrous-oxide. Soil Sci. Soc. Am. J. 58, 1108e1116.
Tiedje, J.M., Sorensen, J., Chang, Y.-Y.L., 1981. Assimilatory and dissimilatory nitratereduction: perspectives and methodology for simultaneous measurement ofseveral nitrogen cycle processes. In: Clark, E., Rosswall, T. (Eds.), TerrestrialNitrogen Cycles e Processes, Ecosystem Strategies and Management Impact,pp. 331e342. Ecol. Bull., Stockholm.
Wright, C.J., Coleman, D.C., 2000. Cross-site comparison of soil microbial biomass,soil nutrient status, and nematode trophic groups. Pedobiologia 44, 2e23.





























