Embed Size (px)
Citation preview

80
Induction of cyclooxygenase (COX)-2, which converts arachidonic acid into prostaglandin H2 (PGH
2), confers
cardioprotective effects in the context of I/R injury,1 left ventricular (LV) remodeling after myocardial infarction,2 doxorubicin-induced myocardial injury,3 and acute viral myo-carditis.4,5 Augmentation of cardiac COX-2 activity might thus be a promising tool for cardioprotection in diverse pathologi-cal conditions.
Glucocorticoids are released during the body’s normal protective response to a heart attack, and it act to repress PG biosynthesis in most cell types. Recently, we found
that glucocorticoids markedly upregulate the expression of COX-2 in cardiomyocytes via the glucocorticoid receptor.6 Glucocorticoids also selectively upregulate the gene expres-sion of lipocalin-type PGD synthase (L-PGDS), which converts PGH
2 to PGD
2, and they are highly expressed in
myocardial tissue, including cardiomyocytes.7 Consequently, PGD
2 is the predominant PG induced in vivo in mouse hearts
and in vitro in cultured neonatal rat cardiomyocytes after exposure to glucocorticoids. Dexamethasone (DEX), a glu-cocorticoid receptor–selective agonist, alleviates ischemia–reperfusion (I/R) injury in isolated Langendorff-perfused
Abstract—We recently demonstrated that glucocorticoids markedly upregulate the expression of cyclooxygenase-2 in cardiomyocytes and protect hearts from ischemia–reperfusion (I/R) injury by activating lipocalin-type prostaglandin D (PGD) synthase (L-PGDS)–derived PGD
2 biosynthesis. We examined a downstream mechanism of cardioprotection
elicited by PGD2 biosynthesis. Acute PGD
2 treatment did not protect hearts against I/R injury. We then speculated
that PGD2 and its metabolite 15-deoxy-Δ12,14-PGJ
2 activate gene expression networks to mediate the glucocorticoid-
mediated cardioprotection. Using an unbiased approach, we identified that glucocorticoids induce a number of well-known erythroid-derived 2–like 2 (Nrf2) target genes in the heart in an L-PGDS–dependent manner and that the cardioprotective effect of glucocorticoids against I/R injury was not seen in Nrf2-knockout hearts. We showed relatively low expression of PGD
2 receptors (ie, DP1 and DP2) in the heart but abundant expression of PGF
2α receptor (FP), which binds PGF2α
and PGD2 with equal affinity. Glucocorticoids also failed to induce the expression of L-PGDS–dependent Nrf2 target
genes in FP-knockout hearts. PGD2 acted through its metabolite 15-deoxy-Δ12,14-PGJ
2 in the heart as evidenced by
the glucocorticoid-mediated activation of peroxisome proliferator-activated receptor-γ. In turn, glucocorticoids failed to induce the expression of L-PGDS–dependent Nrf2 target genes in hearts pretreated with peroxisome proliferator-activated receptor-γ antagonist GW9662, and glucocorticoid-mediated cardioprotection against I/R injury was compromised in FP-knockout mice and GW9662-treated mice. In conclusion, PGD
2 protects heart against I/R injury by activating Nrf2
predominantly via FP receptor. In addition, we propose activation of peroxisome proliferator-activated receptor-γ by the dehydrated metabolite of PGD
2 (15-deoxy-Δ12,14-PGJ
2) as another mechanism by which glucocorticoids induce
cardioprotection. (Hypertension. 2014;63:80-87.) • Online Data Supplement
Key Words: 15-deoxyprostaglandin J2 ■ Nrf2 ■ oxidative stress ■ PPAR gamma ■ prostaglandin D2 ■ prostaglandin F
2α receptor
Received May 12, 2013; first decision May 30, 2013; revision accepted September 11, 2013.From the Department of Cardiology (Y.K., S.T., T.M., H.I., X.Y., K.I., S.Y., M.I., K.F., M.S.), Department of Geriatric Medicine (K.S.), Department of
Biochemistry (Y.S., M.S.), and Department of Clinical and Molecular Cardiovascular Research (M.I.), Keio University School of Medicine, Tokyo, Japan; Precursory Research for Embryonic Science and Technology, Japan Science and Technology Agency, Tokyo, Japan (Y.S.); and Department of Molecular and Behavioral Biology, Osaka Bioscience Institute, Suita, Japan (Y.U.).
The online-only Data Supplement is available with this article at http://hyper.ahajournals.org/lookup/suppl/doi:10.1161/HYPERTENSIONAHA. 113.01639/-/DC1.
Correspondence to Motoaki Sano, Department of Cardiology, Keio University School of Medicine, 35 Shinanomachi Shinjuku-ku, Tokyo 160-8582, Japan. E-mail [email protected]
Endogenous Prostaglandin D2 and Its Metabolites Protect the Heart Against Ischemia–Reperfusion Injury
by Activating Nrf2Yoshinori Katsumata, Ken Shinmura, Yuki Sugiura, Shugo Tohyama, Tomohiro Matsuhashi, Hideyuki Ito, Xiaoxiang Yan, Kentaro Ito, Shinsuke Yuasa, Masaki Ieda, Yoshihiro Urade,
Makoto Suematsu, Keiichi Fukuda, Motoaki Sano
© 2013 American Heart Association, Inc.
Hypertension is available at http://hyper.ahajournals.org DOI: 10.1161/HYPERTENSIONAHA.113.01639
See Editorial Commentary, pp 22–23
Heart
by guest on May 17, 2018
http://hyper.ahajournals.org/D
ownloaded from
by guest on M
ay 17, 2018http://hyper.ahajournals.org/
Dow
nloaded from
by guest on May 17, 2018
http://hyper.ahajournals.org/D
ownloaded from
by guest on M
ay 17, 2018http://hyper.ahajournals.org/
Dow
nloaded from
by guest on May 17, 2018
http://hyper.ahajournals.org/D
ownloaded from
by guest on M
ay 17, 2018http://hyper.ahajournals.org/
Dow
nloaded from
by guest on May 17, 2018
http://hyper.ahajournals.org/D
ownloaded from
by guest on M
ay 17, 2018http://hyper.ahajournals.org/
Dow
nloaded from
by guest on May 17, 2018
http://hyper.ahajournals.org/D
ownloaded from
by guest on M
ay 17, 2018http://hyper.ahajournals.org/
Dow
nloaded from
by guest on May 17, 2018
http://hyper.ahajournals.org/D
ownloaded from
by guest on M
ay 17, 2018http://hyper.ahajournals.org/
Dow
nloaded from
by guest on May 17, 2018
http://hyper.ahajournals.org/D
ownloaded from
by guest on M
ay 17, 2018http://hyper.ahajournals.org/
Dow
nloaded from
by guest on May 17, 2018
http://hyper.ahajournals.org/D
ownloaded from
by guest on M
ay 17, 2018http://hyper.ahajournals.org/
Dow
nloaded from
by guest on May 17, 2018
http://hyper.ahajournals.org/D
ownloaded from
Katsumata et al Cardioprotection in PGD2-Enriched Milieu 81
mouse hearts. This cardioprotective effect of DEX could be abolished by either pharmacological inhibition of COX-2 or disruption of the gene encoding L-PGDS.6 Together, these findings indicated that the activation of prostanoid biosyn-thesis (primarily PGD
2) accounts for the major cardiopro-
tective effect of glucocorticoids against I/R injury. Despite numerous investigations to clarify the molecular actions of cardiovascular prostanoids, PGD
2 has received little atten-
tion in studies investigating the mechanisms underlying cardioprotection. Thus, the intracellular signaling path-way downstream of PGD
2 biosynthesis in cardiomyocytes
remains unknown.It is known that PGD
2 interacts with 2 distinct G-protein–
coupled receptors DP1 and DP2.8 The DP1 receptor is a clas-sical prostanoid receptor, whereas the DP2 receptor (formerly known as the chemoattractant receptor-homologous molecule expressed on Th2 cells receptor) is phylogenetically differ-ent from DP1 and other prostanoid receptors and belongs to a family of chemokine receptors.
PGD2 is dehydrated to the J-series of PGs, includ-
ing 15-deoxy-Δ12,14-PGJ2 (15d-PGJ
2). The biological
actions of 15d-PGJ2 are attributed to its ability to form
covalent adducts with thiol residues within specific signal-ing proteins, thus, triggering redox-sensitive cell signal-ing pathways.9 15d-PGJ
2 plays a key role in augmenting
the cellular antioxidant defense capacity and resolution of inflammation by suppressing nuclear factor-κB and activating peroxisome proliferator-activated receptor (PPAR)γ and erythroid-derived 2–like 2 (Nrf2). Whether biologically significant amounts of PGD
2 are converted to
J-series PGs in vivo remains contentious,10 and it is pos-sible that PGD
2 could act through its metabolite 15d-PGJ
2
in cardiomyocytes.In the present study, we investigated the molecular mecha-
nism underlying the cardioprotection afforded by activation of L-PGDS–derived PGD
2 biosynthesis.
Materials and MethodsA detailed, expanded Methods section is available in the online-only Data Supplement.
0
20
40
60
80
100
120Cont (n=6)DEX (n=6)
6050403020100BSL0
20
40
60
80
6050403020100BSL0
150
300
450
600
750
6050403020100BSL
0
1000
2000
3000
4000
5000
6050403020100BSL0
1000
2000
3000
4000
5000
6050403020100BSL
LVD
P (m
mH
g)
Reperfusion (min)
Ischemia
+ LV
dP/d
t (m
mH
g/s)
Reperfusion (min)
- LV
dP/d
t (m
mH
g/s)
Reperfusion (min)
LVE
DP
(mm
Hg)
Reperfusion (min)
Ischemia
HR
(bpm
)
Reperfusion (min)
Ischemia
Tota
l LD
H a
ctiv
ity (I
U)Ischemia Ischemia
n.s.
Con DEX
Nrf2-KO heart
n.s. n.s. n.s. n.s. n.s. n.s.
n.s. n.s. n.s.n.s.
n.s.n.s.
n.s. n.s. n.s.n.s.
n.s. n.s.
n.s.
n.s. n.s. n.s. n.s. n.s.
n.s. n.s.
n.s. n.s. n.s. n.s.
0
4
8
12
16
LVD
P (m
mH
g)
Reperfusion (min)
Ischemia
- LV
dP/d
t (m
mH
g/s)
Reperfusion (min)
Ischemia
+ LV
dP/d
t (m
mH
g/s)
Reperfusion (min)
Ischemia
LVE
DP
(mm
Hg)
Reperfusion (min)
Ischemia
HR
(bpm
)
Reperfusion (min)
Ischemia
Tota
l LD
H a
ctiv
ity (I
U) #
0
20
40
60
80
100
120 Cont (n=7)DEX (n=7)
6050403020100BSL0
20
40
60
80
6050403020100BSL0
150
300
450
600
750
6050403020100BSL
0
1000
2000
3000
4000
5000
6050403020100BSL0
1000
2000
3000
4000
5000
6050403020100BSL Con DEX0
4
8
12
16
## # # # #
# # # #
# #
## #
## #
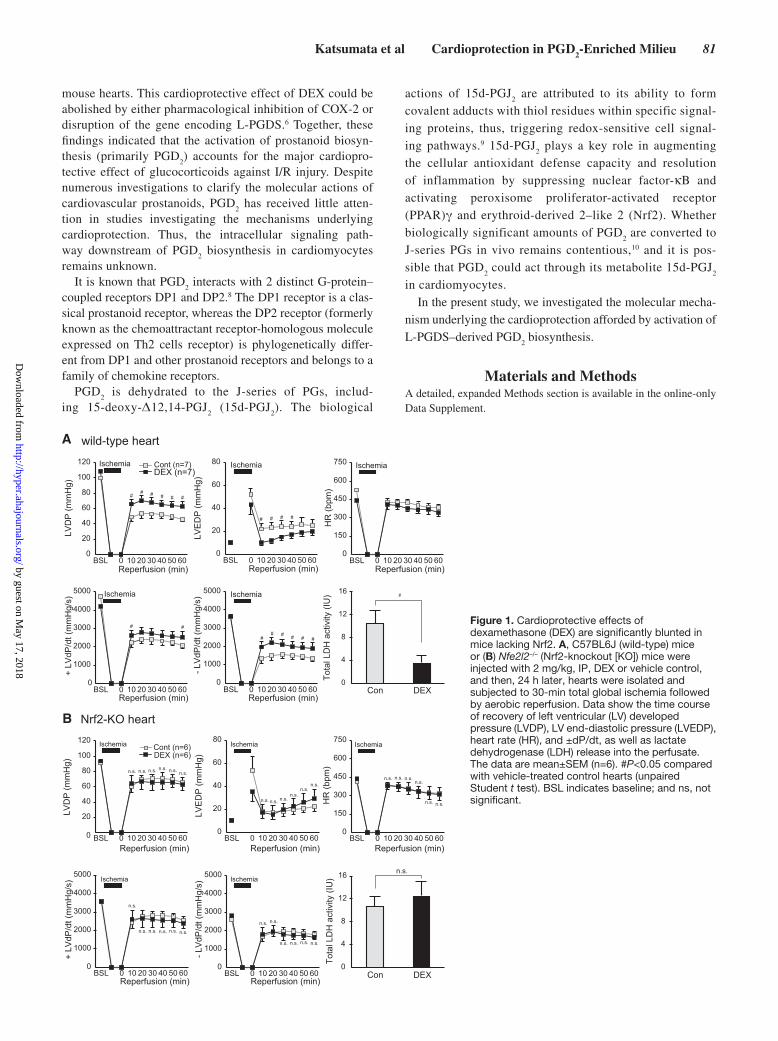
A wild-type heart
B
Figure 1. Cardioprotective effects of dexamethasone (DEX) are significantly blunted in mice lacking Nrf2. A, C57BL6J (wild-type) mice or (B) Nfe2l2–/– (Nrf2-knockout [KO]) mice were injected with 2 mg/kg, IP, DEX or vehicle control, and then, 24 h later, hearts were isolated and subjected to 30-min total global ischemia followed by aerobic reperfusion. Data show the time course of recovery of left ventricular (LV) developed pressure (LVDP), LV end-diastolic pressure (LVEDP), heart rate (HR), and ±dP/dt, as well as lactate dehydrogenase (LDH) release into the perfusate. The data are mean±SEM (n=6). #P<0.05 compared with vehicle-treated control hearts (unpaired Student t test). BSL indicates baseline; and ns, not significant.
by guest on May 17, 2018
http://hyper.ahajournals.org/D
ownloaded from
82 Hypertension January 2014
ResultsGlucocorticoids Induce Nrf2 Target Genes in Heart in an L-PGDS–Dependent MannerAcute PGD
2 treatment did not protect hearts against I/R injury
(Figure S1 in the online-only Data Supplement). We then speculated that PGD
2 and its metabolite PGJ
2 activate gene
expression networks to mediate the later phases of cardiopro-tection. To identify the cardioprotective genes induced by glu-cocorticoids in an L-PGDS–dependent manner, both wild-type and L-PGDS-knockout (L-PGDS-KO) mice were subjected to DEX treatment, and gene expression profiling was examined by DNA microarray analysis. DEX treatment of wild-type, but not L-PGDS-KO, mice upregulated the expression of characteristic Nrf2 target genes. To verify the microarray results, quantitative polymerase chain reaction analysis was performed to compare the time course of changes in mRNA expression of Nrf2 tar-get genes after DEX treatment of wild-type and L-PGDS-KO hearts (Figure S2). In wild-type hearts, the mRNA expression of the well-known Nrf2 target genes Gclc, Gsr, G6pdx, Hmox-1, Gclm, and Cat started to increase between 3 and 12 hours after DEX administration. In L-PGDS-KO hearts, the induction of Gclc, Gsr, and G6pdx expression was significantly blunted after DEX treatment compared with wild-type hearts, whereas there was no difference in the expression of Hmox-1, Gclm, and Cat. DEX treatment had no effect on the expression of Nqo1 in hearts from either wild-type or L-PGDS-KO mice. Interestingly, the expression of Nfe2l2 (Nrf2) itself was increased by DEX treatment in a L-PGDS–dependent manner.
Role of Nrf2 in DEX-Induced CardioprotectionTo examine the putative role of Nrf2 in the cardioprotection afforded by DEX-mediated activation of L-PGDS–driven PGD
2 biosynthesis, C57BL6J (wild-type) and Nrf2-KO mice
were injected with DEX or an equal volume of vehicle (PBS) 24 hours before isolated hearts were subjected to I/R injury. There were no differences in LV function before the induction of ischemia in hearts from Nrf2-KO and wild-type mice.
DEX pretreatment of wild-type mice significantly improved recovery of LV developed pressure and ± dP/dt and attenuated total lactate dehydrogenase (LDH) release into the perfusate during reperfusion compared with the vehicle-treated mice (Figure 1A).
As reported previously,11 LV developed pressure during reperfusion was somewhat higher in hearts from Nrf2-KO compared with wild-type mice (Figure 1B), suggesting that genetic deletion of Nrf2 activates compensatory mechanisms that protect the heart against I/R injury. Baseline expres-sion of L-PGDS was comparable in hearts from wild-type and Nrf2-KO mice, and DEX induced L-PGDS expression in Nrf2-KO hearts to the same extent as in wild-type hearts (Figure S3). Nevertheless, DEX failed to improve functional recovery after I/R injury, and it did not reduce total LDH release into the perfusate during reperfusion in hearts from Nrf2-KO mice. On the basis of these findings, although Nrf2-KO did result in nonspecific cardioprotection in our experimental model, we can state that the Nrf2 activation plays a substantial role in DEX-mediated cardioprotection as a downstream tar-get of L-PGDS–dependent PGD
2 biosynthesis.
B
0
100
200
300
400
0
100
200
300
400
0
100
200
300
1000
hear
t
lung
brai
n
inte
stin
e
kidn
eysp
leen
thym
us
Ptgdr
gene
exp
ress
ion
gene
exp
ress
ion
Gpr44 Ptgfr10-7/18s 10-7/18s 10-7/18s
gene
exp
ress
ion 615
0
25
50
20000
25000
gene
exp
ress
ion
Ptgdr Gpr44 Ptgfr
gene
exp
ress
ion
22810
hear
t
lung
brai
n
inte
stin
e
kidn
eysp
leen
thym
us
hear
t
lung
brai
n
inte
stin
e
kidn
eysp
leen
thym
us
37kDa64kDa
A
heart lungbrain kidney spleenGAPDH
FP
heart lungbrain kidney spleen
wild-type
Rel
ativ
e de
nsity
(
fold
cha
nge)
0
25
50
ES ES-cardiomyocyte (ES-C)
Gpr
44P
tgfr
Ptg
dr
C
10-7/18s
human
0
10
20
30
*
*
D
rat mouse
Ptgds10-5/18s
ES ES-C0
5
10 *
mouse
F
DEX
Intramuscular PGs (pg/tissue mg)
†
0.0 0.3 0.6 0.9 1.2 1.5
PGF2a
PGD2
con
n.s.
E
DEX 0 0
2
4
6
8 WT
(h)
gene
exp
ress
ion
(fo
ld c
hang
e)
6 12 24
Ptgfr
3
##
Gpr
44P
tgfr
Ptg
dr
10-7/18s
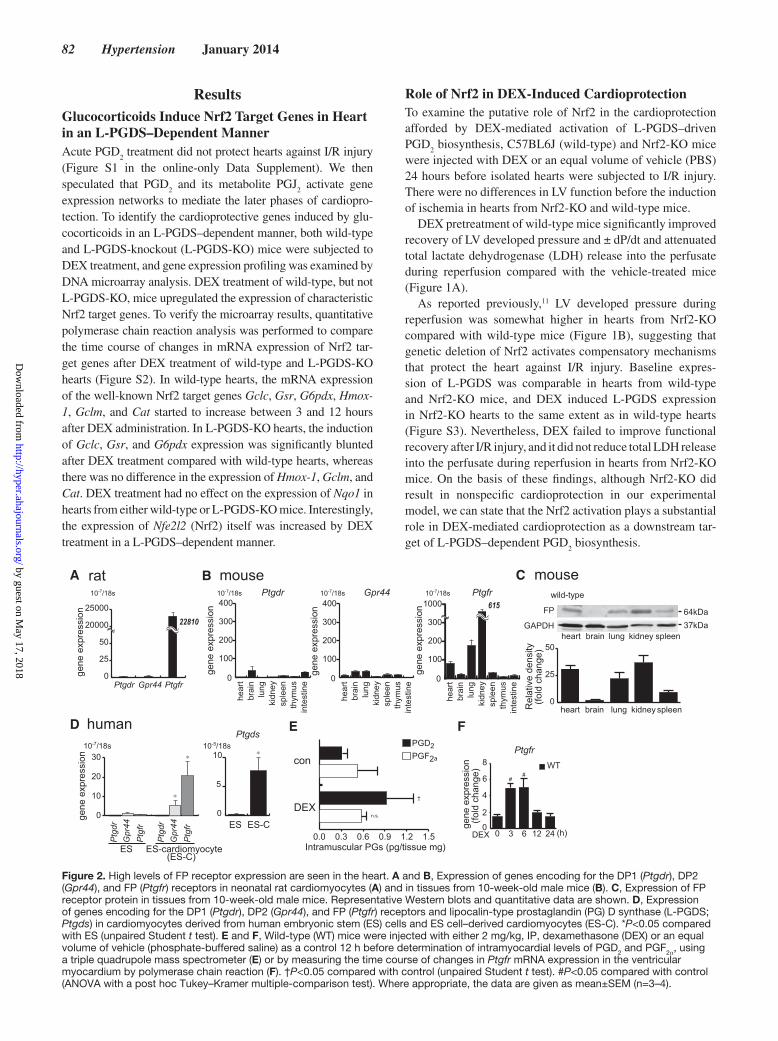
Figure 2. High levels of FP receptor expression are seen in the heart. A and B, Expression of genes encoding for the DP1 (Ptgdr), DP2 (Gpr44), and FP (Ptgfr) receptors in neonatal rat cardiomyocytes (A) and in tissues from 10-week-old male mice (B). C, Expression of FP receptor protein in tissues from 10-week-old male mice. Representative Western blots and quantitative data are shown. D, Expression of genes encoding for the DP1 (Ptgdr), DP2 (Gpr44), and FP (Ptgfr) receptors and lipocalin-type prostaglandin (PG) D synthase (L-PGDS; Ptgds) in cardiomyocytes derived from human embryonic stem (ES) cells and ES cell–derived cardiomyocytes (ES-C). *P<0.05 compared with ES (unpaired Student t test). E and F, Wild-type (WT) mice were injected with either 2 mg/kg, IP, dexamethasone (DEX) or an equal volume of vehicle (phosphate-buffered saline) as a control 12 h before determination of intramyocardial levels of PGD2 and PGF2α, using a triple quadrupole mass spectrometer (E) or by measuring the time course of changes in Ptgfr mRNA expression in the ventricular myocardium by polymerase chain reaction (F). †P<0.05 compared with control (unpaired Student t test). #P<0.05 compared with control (ANOVA with a post hoc Tukey–Kramer multiple-comparison test). Where appropriate, the data are given as mean±SEM (n=3–4).
by guest on May 17, 2018
http://hyper.ahajournals.org/D
ownloaded from
Katsumata et al Cardioprotection in PGD2-Enriched Milieu 83
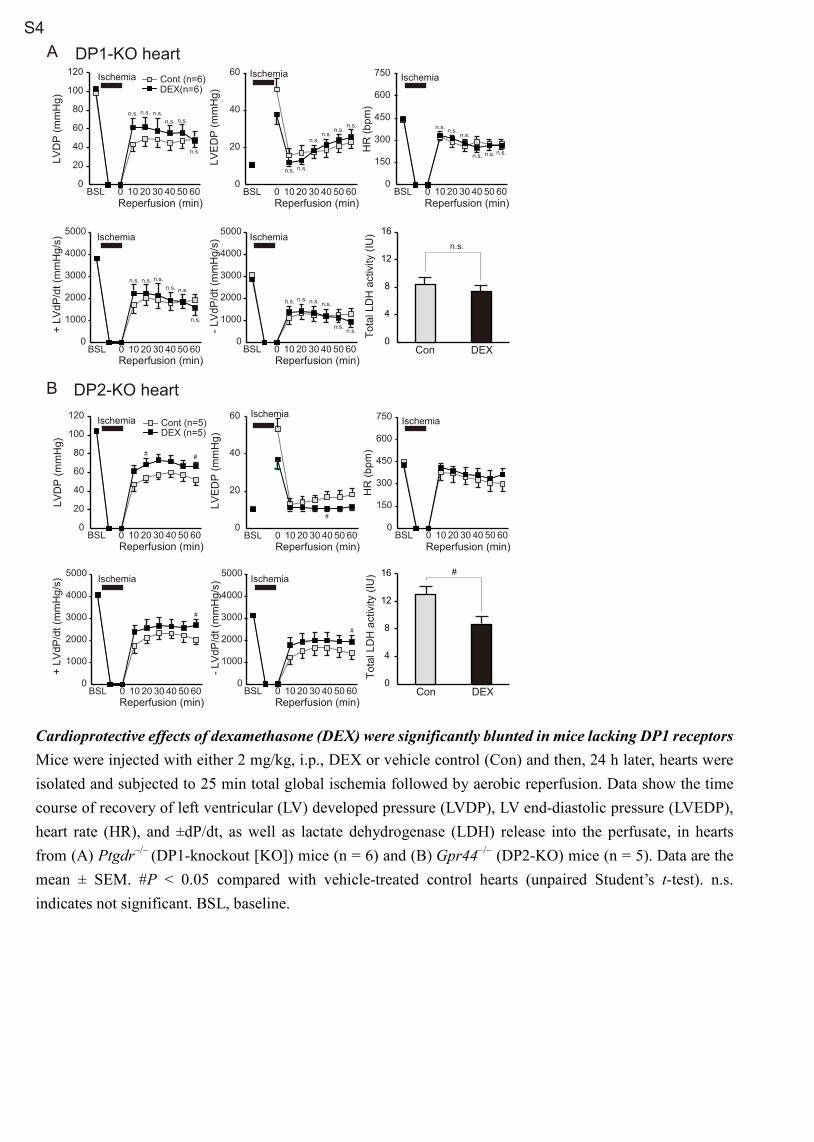
FP Receptor Contributes to PGD2-Mediated Expression of the Nrf2 Target GeneConsistent with a previous observation that DP1, rather than DP2, receptors were implicated in PGD
2-mediated cardioprotection
against anoxia–reoxygenation in cultured neonatal rat cardiomyo-cytes,6 DEX-mediated improvement of functional recovery after I/R injury and reduction of total LDH release into the perfusate during reperfusion were significantly blunted in DP1-KO but not DP2-KO mice (Figure S4). However, the DP1 receptor agonist BW245c failed to increase the mRNA expression of Nrf2 target genes in cultured neonatal rat cardiomyocytes (Figure S5B).
Prostanoids show cross-reactivity with several types of recep-tors. PGD
2 binds with equal affinity to DP1 receptors and the
canonical PGF2α receptor FP,12 whereas the FP receptor binds
PGF2α and PGD
2 with equal, high affinity.13 In the present study,
we found relatively low expression of genes encoding the DP1 (Ptgdr) and DP2 (Gpr44) receptors in cultured neonatal rat car-diomyocytes (Figure 2A), whereas abundant expression was detected for the gene encoding the FP receptor (Ptgfr). Similarly, in adult mouse heart, expression of genes encoding the DP1 and
DP2 receptors was relatively low, whereas the FP receptor gene expression was abundant (Figure 2B and 2C). In addition, the FP receptor expression was higher than that of DP1 and DP2 recep-tors in cardiomyocytes derived from human embryonic stem cells (Figure 2D). In the present study, injection of wild-type mice with DEX resulted in a 3.2-fold increase in intramyocardial PGD
2 levels without any change in PGF
2α levels (Figure 2E). Furthermore, at 3 hours after DEX administration, the mRNA expression of FP receptor (Ptgfr) in the ventricular myocardium of wild-type mice increased (Figure 2F). Together, these results strongly suggested that FP plays a substantial role in PGD
2 sig-
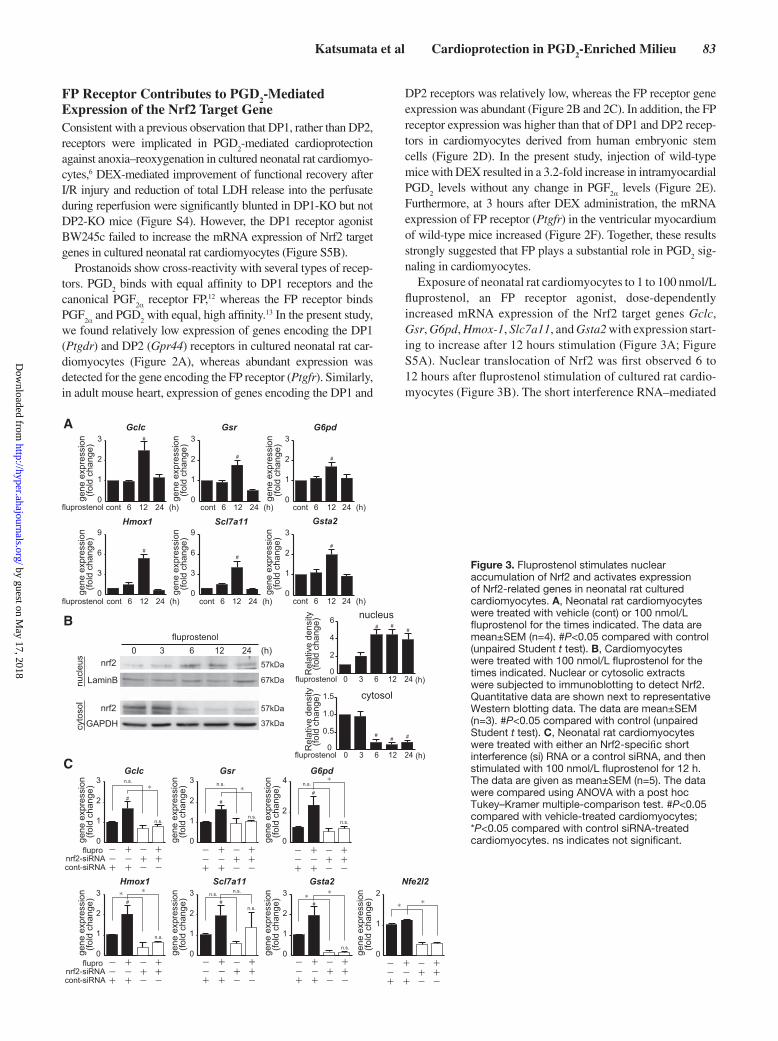
naling in cardiomyocytes.Exposure of neonatal rat cardiomyocytes to 1 to 100 nmol/L
fluprostenol, an FP receptor agonist, dose-dependently increased mRNA expression of the Nrf2 target genes Gclc, Gsr, G6pd, Hmox-1, Slc7a11, and Gsta2 with expression start-ing to increase after 12 hours stimulation (Figure 3A; Figure S5A). Nuclear translocation of Nrf2 was first observed 6 to 12 hours after fluprostenol stimulation of cultured rat cardio-myocytes (Figure 3B). The short interference RNA–mediated
cont 6 12 24 (h) cont 6 12 24 (h)
cont 6 12 24 (h)
cont 6 12 24 (h)
cont 6 12 24 (h) cont 6 12 24 (h)
A
gene
exp
ress
ion
(fo
ld c
hang
e)
0
1
2
3
0
1
2
3
0
1
2
3
0
1
2
3
gene
exp
ress
ion
(fo
ld c
hang
e)
gene
exp
ress
ion
(fo
ld c
hang
e)
gene
exp
ress
ion
(fo
ld c
hang
e)
gene
exp
ress
ion
(fo
ld c
hang
e)
0
3
6
9
0
3
6
9
gene
exp
ress
ion
(fo
ld c
hang
e)
B
LaminB
nrf2
nrf2
GAPDH
57kDa
67kDa
57kDa
37kDa
fluprostenol(h)
nucl
eus
cyto
sol
flupronrf2-siRNA
-- +
++-
cont-siRNA + ++-
- -
-- +
++-
+ ++-
- -
-- +
++-
+ ++-
- -
-- +
++-
+ ++-
- -
flupronrf2-siRNA
-- +
++-
cont-siRNA + ++-
- -
-- +
++-
+ ++-
- -
-- +
++-
+ ++-
- -
0 3 6 12 24
Gclc
Gsta2
Gsr G6pd
fluprostenol
fluprostenol
C
gene
exp
ress
ion
(fo
ld c
hang
e)
0
1
2
3
0
1
2
3
0
2
4
0
1
2
3
gene
exp
ress
ion
(fo
ld c
hang
e)
gene
exp
ress
ion
(fo
ld c
hang
e)
gene
exp
ress
ion
(fo
ld c
hang
e)
gene
exp
ress
ion
(fo
ld c
hang
e)
0
1
2
3
0
1
2
3Nfe2l2
gene
exp
ress
ion
(fo
ld c
hang
e)
gene
exp
ress
ion
(fo
ld c
hang
e)
n.s.*
* *n.s. n.s.
n.s.
n.s.
n.s.
**
Gclc
Gsta2
Gsr G6pd
Hmox1 Scl7a11
Hmox1 Scl7a11
#
##
#
# #
#
#
#
#
##*
*
n.s.*
n.s.*
n.s.
n.s.
n.s.
0
1
2
0 3 6 12 24 (h)
# ##
## #
0
2
4
6
0
0.5
1.0
1.5 cytosol
nucleus
Rel
ativ
e de
nsity
(
fold
cha
nge)
0 3 6 12 24 (h)
Rel
ativ
e de
nsity
(
fold
cha
nge)
fluprostenol
fluprostenol
Figure 3. Fluprostenol stimulates nuclear accumulation of Nrf2 and activates expression of Nrf2-related genes in neonatal rat cultured cardiomyocytes. A, Neonatal rat cardiomyocytes were treated with vehicle (cont) or 100 nmol/L fluprostenol for the times indicated. The data are mean±SEM (n=4). #P<0.05 compared with control (unpaired Student t test). B, Cardiomyocytes were treated with 100 nmol/L fluprostenol for the times indicated. Nuclear or cytosolic extracts were subjected to immunoblotting to detect Nrf2. Quantitative data are shown next to representative Western blotting data. The data are mean±SEM (n=3). #P<0.05 compared with control (unpaired Student t test). C, Neonatal rat cardiomyocytes were treated with either an Nrf2-specific short interference (si) RNA or a control siRNA, and then stimulated with 100 nmol/L fluprostenol for 12 h. The data are given as mean±SEM (n=5). The data were compared using ANOVA with a post hoc Tukey–Kramer multiple-comparison test. #P<0.05 compared with vehicle-treated cardiomyocytes; *P<0.05 compared with control siRNA-treated cardiomyocytes. ns indicates not significant.
by guest on May 17, 2018
http://hyper.ahajournals.org/D
ownloaded from
84 Hypertension January 2014
knockdown of Nrf2 completely abolished the fluprostenol-mediated upregulation of Gclc, Gsr, G6pd, Hmox, and Gsta2 expression but not that of Slc7a11 (Figure 3C).
We examined the role of the FP receptor in the expression of Nrf2 target genes acting as downstream effectors of L-PGDS–mediated PGD
2 biosynthesis. Of the L-PGDS–dependent Nrf2
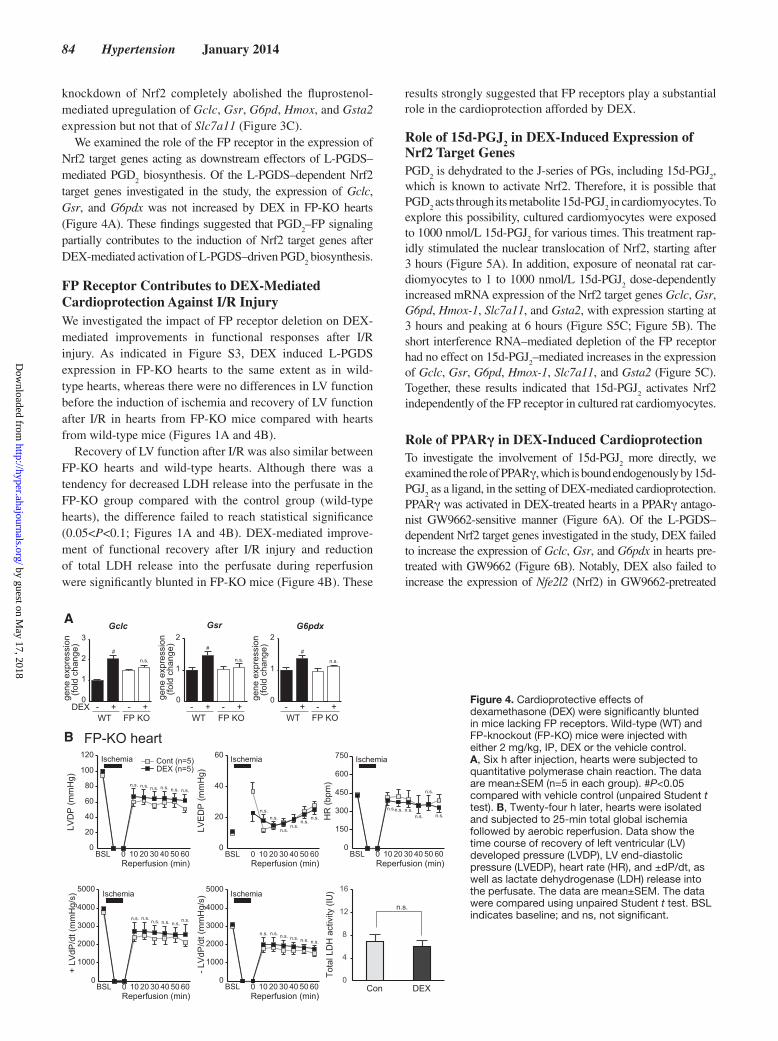
target genes investigated in the study, the expression of Gclc, Gsr, and G6pdx was not increased by DEX in FP-KO hearts (Figure 4A). These findings suggested that PGD
2–FP signaling
partially contributes to the induction of Nrf2 target genes after DEX-mediated activation of L-PGDS–driven PGD
2 biosynthesis.
FP Receptor Contributes to DEX-Mediated Cardioprotection Against I/R InjuryWe investigated the impact of FP receptor deletion on DEX-mediated improvements in functional responses after I/R injury. As indicated in Figure S3, DEX induced L-PGDS expression in FP-KO hearts to the same extent as in wild-type hearts, whereas there were no differences in LV function before the induction of ischemia and recovery of LV function after I/R in hearts from FP-KO mice compared with hearts from wild-type mice (Figures 1A and 4B).
Recovery of LV function after I/R was also similar between FP-KO hearts and wild-type hearts. Although there was a tendency for decreased LDH release into the perfusate in the FP-KO group compared with the control group (wild-type hearts), the difference failed to reach statistical significance (0.05<P<0.1; Figures 1A and 4B). DEX-mediated improve-ment of functional recovery after I/R injury and reduction of total LDH release into the perfusate during reperfusion were significantly blunted in FP-KO mice (Figure 4B). These
results strongly suggested that FP receptors play a substantial role in the cardioprotection afforded by DEX.
Role of 15d-PGJ2 in DEX-Induced Expression of Nrf2 Target GenesPGD
2 is dehydrated to the J-series of PGs, including 15d-PGJ
2,
which is known to activate Nrf2. Therefore, it is possible that PGD
2 acts through its metabolite 15d-PGJ
2 in cardiomyocytes. To
explore this possibility, cultured cardiomyocytes were exposed to 1000 nmol/L 15d-PGJ
2 for various times. This treatment rap-
idly stimulated the nuclear translocation of Nrf2, starting after 3 hours (Figure 5A). In addition, exposure of neonatal rat car-diomyocytes to 1 to 1000 nmol/L 15d-PGJ
2 dose-dependently
increased mRNA expression of the Nrf2 target genes Gclc, Gsr, G6pd, Hmox-1, Slc7a11, and Gsta2, with expression starting at 3 hours and peaking at 6 hours (Figure S5C; Figure 5B). The short interference RNA–mediated depletion of the FP receptor had no effect on 15d-PGJ
2–mediated increases in the expression
of Gclc, Gsr, G6pd, Hmox-1, Slc7a11, and Gsta2 (Figure 5C). Together, these results indicated that 15d-PGJ
2 activates Nrf2
independently of the FP receptor in cultured rat cardiomyocytes.
Role of PPARγ in DEX-Induced CardioprotectionTo investigate the involvement of 15d-PGJ
2 more directly, we
examined the role of PPARγ, which is bound endogenously by 15d-PGJ
2 as a ligand, in the setting of DEX-mediated cardioprotection.
PPARγ was activated in DEX-treated hearts in a PPARγ antago-nist GW9662-sensitive manner (Figure 6A). Of the L-PGDS–dependent Nrf2 target genes investigated in the study, DEX failed to increase the expression of Gclc, Gsr, and G6pdx in hearts pre-treated with GW9662 (Figure 6B). Notably, DEX also failed to increase the expression of Nfe2l2 (Nrf2) in GW9662-pretreated
A
-DEX +WT
- + FP KO
- +WT
- +FP KO
- +WT
- +FP KO
0
1
2
0
1
2Gclc Gsr G6pdx
##
n.s.#
n.s.
gene
exp
ress
ion
(fo
ld c
hang
e)
gene
exp
ress
ion
(fo
ld c
hang
e)
gene
exp
ress
ion
(fo
ld c
hang
e)
0
1
2
3
n.s.
0
20
40
60
80
100
120Cont (n=5)DEX (n=5)
6050403020100BSL0
20
40
60
6050403020100BSL0
150
300
450
600
750
6050403020100BSL
0
1000
2000
3000
4000
5000
6050403020100BSL0
1000
2000
3000
4000
5000
6050403020100BSL
LVD
P (m
mH
g)
Reperfusion (min)
Ischemia
+ LV
dP/d
t (m
mH
g/s)
Reperfusion (min)
- LV
dP/d
t (m
mH
g/s)
Reperfusion (min)
LVE
DP
(mm
Hg)
Reperfusion (min)
Ischemia
HR
(bpm
)
Reperfusion (min)
Ischemia
Tota
l LD
H a
ctiv
ity (I
U)Ischemia Ischemia
Con DEX
n.s.
0
4
8
12
16
n.s. n.s. n.s. n.s. n.s. n.s.
n.s.n.s. n.s.n.s.
n.s.
n.s.n.s.n.s.
n.s.
n.s.n.s.
n.s.
n.s. n.s.n.s. n.s. n.s.
n.s.
n.s. n.s. n.s. n.s. n.s. n.s.
B FP-KO heart
Figure 4. Cardioprotective effects of dexamethasone (DEX) were significantly blunted in mice lacking FP receptors. Wild-type (WT) and FP-knockout (FP-KO) mice were injected with either 2 mg/kg, IP, DEX or the vehicle control. A, Six h after injection, hearts were subjected to quantitative polymerase chain reaction. The data are mean±SEM (n=5 in each group). #P<0.05 compared with vehicle control (unpaired Student t test). B, Twenty-four h later, hearts were isolated and subjected to 25-min total global ischemia followed by aerobic reperfusion. Data show the time course of recovery of left ventricular (LV) developed pressure (LVDP), LV end-diastolic pressure (LVEDP), heart rate (HR), and ±dP/dt, as well as lactate dehydrogenase (LDH) release into the perfusate. The data are mean±SEM. The data were compared using unpaired Student t test. BSL indicates baseline; and ns, not significant.
by guest on May 17, 2018
http://hyper.ahajournals.org/D
ownloaded from
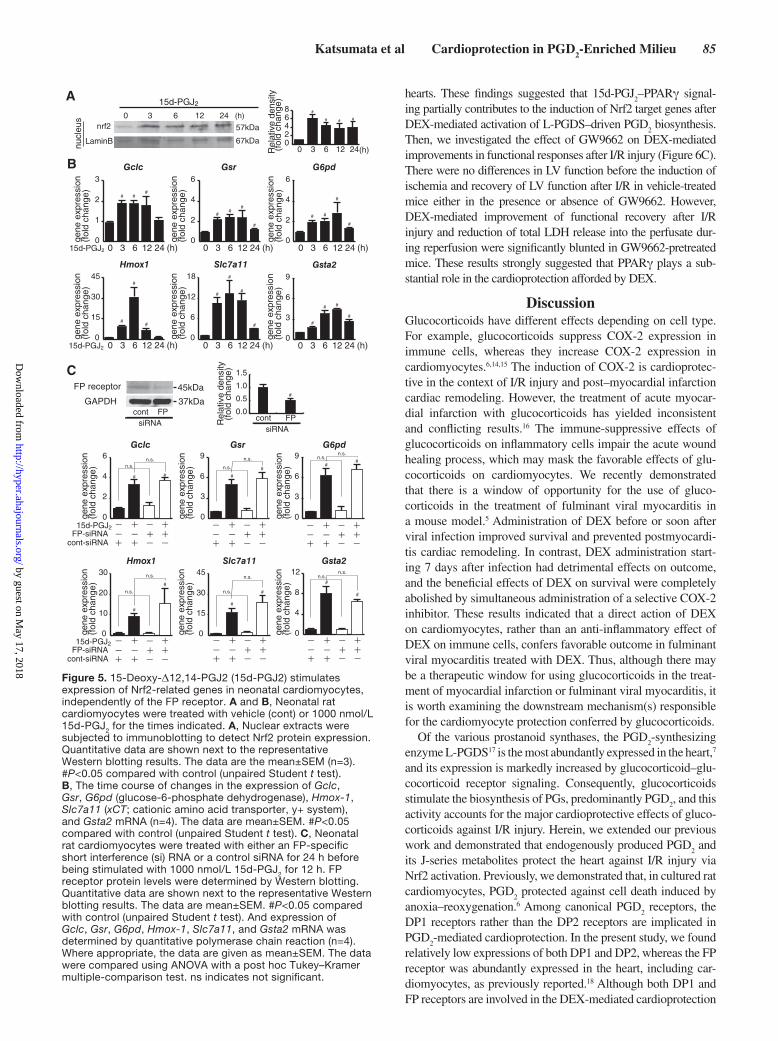
Katsumata et al Cardioprotection in PGD2-Enriched Milieu 85
hearts. These findings suggested that 15d-PGJ2–PPARγ signal-
ing partially contributes to the induction of Nrf2 target genes after DEX-mediated activation of L-PGDS–driven PGD
2 biosynthesis.
Then, we investigated the effect of GW9662 on DEX-mediated improvements in functional responses after I/R injury (Figure 6C). There were no differences in LV function before the induction of ischemia and recovery of LV function after I/R in vehicle-treated mice either in the presence or absence of GW9662. However, DEX-mediated improvement of functional recovery after I/R injury and reduction of total LDH release into the perfusate dur-ing reperfusion were significantly blunted in GW9662-pretreated mice. These results strongly suggested that PPARγ plays a sub-stantial role in the cardioprotection afforded by DEX.
DiscussionGlucocorticoids have different effects depending on cell type. For example, glucocorticoids suppress COX-2 expression in immune cells, whereas they increase COX-2 expression in cardiomyocytes.6,14,15 The induction of COX-2 is cardioprotec-tive in the context of I/R injury and post–myocardial infarction cardiac remodeling. However, the treatment of acute myocar-dial infarction with glucocorticoids has yielded inconsistent and conflicting results.16 The immune-suppressive effects of glucocorticoids on inflammatory cells impair the acute wound healing process, which may mask the favorable effects of glu-cocorticoids on cardiomyocytes. We recently demonstrated that there is a window of opportunity for the use of gluco-corticoids in the treatment of fulminant viral myocarditis in a mouse model.5 Administration of DEX before or soon after viral infection improved survival and prevented postmyocardi-tis cardiac remodeling. In contrast, DEX administration start-ing 7 days after infection had detrimental effects on outcome, and the beneficial effects of DEX on survival were completely abolished by simultaneous administration of a selective COX-2 inhibitor. These results indicated that a direct action of DEX on cardiomyocytes, rather than an anti-inflammatory effect of DEX on immune cells, confers favorable outcome in fulminant viral myocarditis treated with DEX. Thus, although there may be a therapeutic window for using glucocorticoids in the treat-ment of myocardial infarction or fulminant viral myocarditis, it is worth examining the downstream mechanism(s) responsible for the cardiomyocyte protection conferred by glucocorticoids.
Of the various prostanoid synthases, the PGD2-synthesizing
enzyme L-PGDS17 is the most abundantly expressed in the heart,7 and its expression is markedly increased by glucocorticoid–glu-cocorticoid receptor signaling. Consequently, glucocorticoids stimulate the biosynthesis of PGs, predominantly PGD
2, and this
activity accounts for the major cardioprotective effects of gluco-corticoids against I/R injury. Herein, we extended our previous work and demonstrated that endogenously produced PGD
2 and
its J-series metabolites protect the heart against I/R injury via Nrf2 activation. Previously, we demonstrated that, in cultured rat cardiomyocytes, PGD
2 protected against cell death induced by
anoxia–reoxygenation.6 Among canonical PGD2 receptors, the
DP1 receptors rather than the DP2 receptors are implicated in PGD
2-mediated cardioprotection. In the present study, we found
relatively low expressions of both DP1 and DP2, whereas the FP receptor was abundantly expressed in the heart, including car-diomyocytes, as previously reported.18 Although both DP1 and FP receptors are involved in the DEX-mediated cardioprotection
0 6 12 24 (h)
B
gene
exp
ress
ion
(fol
d ch
ange
)
0
3
6
9
0
2
4
6
0
2
4
6
0
1
2
3
gene
exp
ress
ion
(fol
d ch
ange
)
gene
exp
ress
ion
(fol
d ch
ange
)
gene
exp
ress
ion
(fol
d ch
ange
)
gene
exp
ress
ion
(fol
d ch
ange
)
0
6
12
18
0
15
30
45
gene
exp
ress
ion
(fol
d ch
ange
)
Gclc
Gsta2
Gsr G6pd
Hmox1 Slc7a11
3
0 6 12 24 (h)3
0 6 12 24 (h)3 0 6 12 24 (h)3
0 6 12 24 (h)3 0 6 12 24 (h)315d-PGJ2
15d-PGJ2
A
LaminB
nrf2 57kDa
67kDa
(h)
nucl
eus 0 3 6 12 24
15d-PGJ2
##
# #
#
##
#
#
#
#
#
#
#
#
#
#
#
# #
#
#
C
0
4
8
12
0
3
6
9
0
3
6
9
0
2
4
6
0
15
30
45
0
10
20
30Hmox1 Slc7a11
15d-PGJ2FP-siRNA
-- +
++-
cont-siRNA + ++-
- -
-- +
++-
+ ++-
- -
-- +
++-
+ ++-
- -
-- +
++-
+ ++-
- -
15d-PGJ2FP-siRNA
-- +
++-
cont-siRNA + ++-
- -
-- +
++-
+ ++-
- -
gene
exp
ress
ion
(fol
d ch
ange
)
gene
exp
ress
ion
(fol
d ch
ange
)
gene
exp
ress
ion
(fol
d ch
ange
)
gene
exp
ress
ion
(fol
d ch
ange
)
gene
exp
ress
ion
(fol
d ch
ange
)
gene
exp
ress
ion
(fol
d ch
ange
)
n.s.n.s.
Gclc
Gsta2
Gsr G6pd
# #
n.s.n.s.
#
#
n.s.
n.s.
##
n.s.n.s.
##
n.s.
n.s.
#
#n.s.
n.s.
#
#
37kDa
45kDa
GAPDH
FP receptor
FPsiRNA
cont
#
Rel
ativ
e de
nsity
(
fold
cha
nge)
0.0
0.5
1.0
1.5
FPsiRNA
cont
0 3 6 12 24(h)
# # #
Rel
ativ
e de
nsity
(
fold
cha
nge)
02468 #
Figure 5. 15-Deoxy-Δ12,14-PGJ2 (15d-PGJ2) stimulates expression of Nrf2-related genes in neonatal cardiomyocytes, independently of the FP receptor. A and B, Neonatal rat cardiomyocytes were treated with vehicle (cont) or 1000 nmol/L 15d-PGJ2 for the times indicated. A, Nuclear extracts were subjected to immunoblotting to detect Nrf2 protein expression. Quantitative data are shown next to the representative Western blotting results. The data are the mean±SEM (n=3). #P<0.05 compared with control (unpaired Student t test). B, The time course of changes in the expression of Gclc, Gsr, G6pd (glucose-6-phosphate dehydrogenase), Hmox-1, Slc7a11 (xCT; cationic amino acid transporter, y+ system), and Gsta2 mRNA (n=4). The data are mean±SEM. #P<0.05 compared with control (unpaired Student t test). C, Neonatal rat cardiomyocytes were treated with either an FP-specific short interference (si) RNA or a control siRNA for 24 h before being stimulated with 1000 nmol/L 15d-PGJ2 for 12 h. FP receptor protein levels were determined by Western blotting. Quantitative data are shown next to the representative Western blotting results. The data are mean±SEM. #P<0.05 compared with control (unpaired Student t test). And expression of Gclc, Gsr, G6pd, Hmox-1, Slc7a11, and Gsta2 mRNA was determined by quantitative polymerase chain reaction (n=4). Where appropriate, the data are given as mean±SEM. The data were compared using ANOVA with a post hoc Tukey–Kramer multiple-comparison test. ns indicates not significant.
by guest on May 17, 2018
http://hyper.ahajournals.org/D
ownloaded from
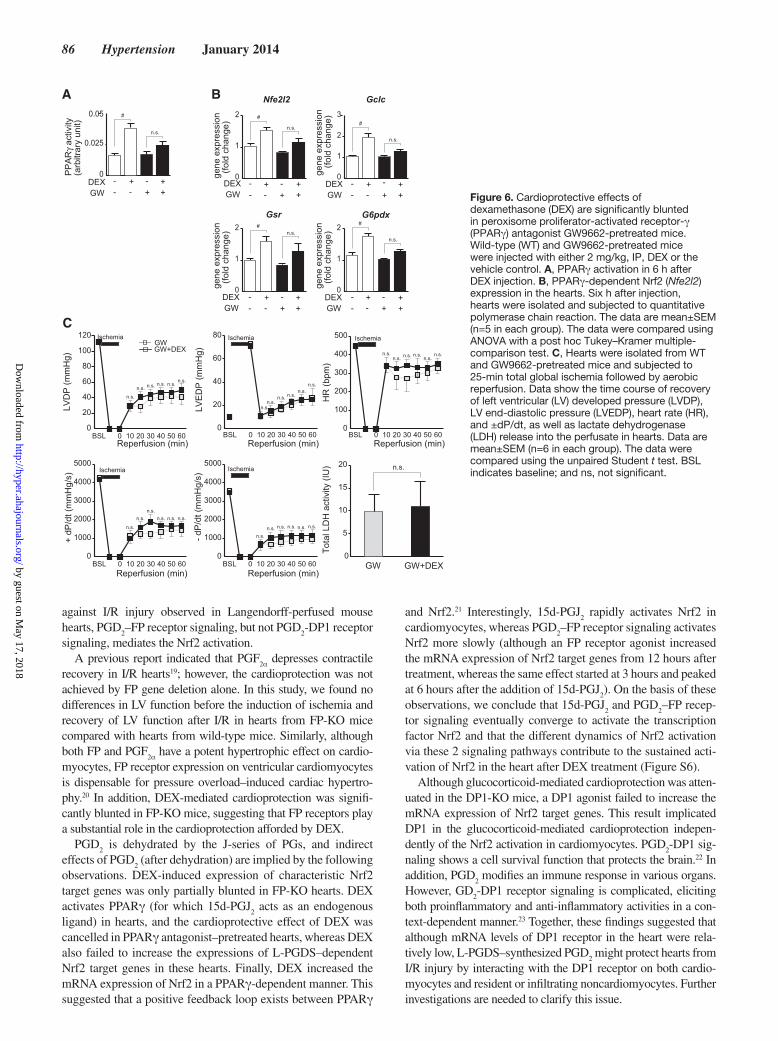
86 Hypertension January 2014
against I/R injury observed in Langendorff-perfused mouse hearts, PGD
2–FP receptor signaling, but not PGD
2-DP1 receptor
signaling, mediates the Nrf2 activation.A previous report indicated that PGF
2α depresses contractile recovery in I/R hearts19; however, the cardioprotection was not achieved by FP gene deletion alone. In this study, we found no differences in LV function before the induction of ischemia and recovery of LV function after I/R in hearts from FP-KO mice compared with hearts from wild-type mice. Similarly, although both FP and PGF
2α have a potent hypertrophic effect on cardio-myocytes, FP receptor expression on ventricular cardiomyocytes is dispensable for pressure overload–induced cardiac hypertro-phy.20 In addition, DEX-mediated cardioprotection was signifi-cantly blunted in FP-KO mice, suggesting that FP receptors play a substantial role in the cardioprotection afforded by DEX.
PGD2 is dehydrated by the J-series of PGs, and indirect
effects of PGD2 (after dehydration) are implied by the following
observations. DEX-induced expression of characteristic Nrf2 target genes was only partially blunted in FP-KO hearts. DEX activates PPARγ (for which 15d-PGJ
2 acts as an endogenous
ligand) in hearts, and the cardioprotective effect of DEX was cancelled in PPARγ antagonist–pretreated hearts, whereas DEX also failed to increase the expressions of L-PGDS–dependent Nrf2 target genes in these hearts. Finally, DEX increased the mRNA expression of Nrf2 in a PPARγ-dependent manner. This suggested that a positive feedback loop exists between PPARγ
and Nrf2.21 Interestingly, 15d-PGJ2 rapidly activates Nrf2 in
cardiomyocytes, whereas PGD2–FP receptor signaling activates
Nrf2 more slowly (although an FP receptor agonist increased the mRNA expression of Nrf2 target genes from 12 hours after treatment, whereas the same effect started at 3 hours and peaked at 6 hours after the addition of 15d-PGJ
2). On the basis of these
observations, we conclude that 15d-PGJ2 and PGD
2–FP recep-
tor signaling eventually converge to activate the transcription factor Nrf2 and that the different dynamics of Nrf2 activation via these 2 signaling pathways contribute to the sustained acti-vation of Nrf2 in the heart after DEX treatment (Figure S6).
Although glucocorticoid-mediated cardioprotection was atten-uated in the DP1-KO mice, a DP1 agonist failed to increase the mRNA expression of Nrf2 target genes. This result implicated DP1 in the glucocorticoid-mediated cardioprotection indepen-dently of the Nrf2 activation in cardiomyocytes. PGD
2-DP1 sig-
naling shows a cell survival function that protects the brain.22 In addition, PGD
2 modifies an immune response in various organs.
However, GD2-DP1 receptor signaling is complicated, eliciting
both proinflammatory and anti-inflammatory activities in a con-text-dependent manner.23 Together, these findings suggested that although mRNA levels of DP1 receptor in the heart were rela-tively low, L-PGDS–synthesized PGD
2 might protect hearts from
I/R injury by interacting with the DP1 receptor on both cardio-myocytes and resident or infiltrating noncardiomyocytes. Further investigations are needed to clarify this issue.
-DEXGW -
+
Nfe2l2 Gclc
Gsr G6pdx
#
gene
exp
ress
ion
(fo
ld c
hang
e)
gene
exp
ress
ion
(fo
ld c
hang
e)
gene
exp
ress
ion
(fo
ld c
hang
e)ge
ne e
xpre
ssio
n (
fold
cha
nge)
0
1
2
3
0
1
2
0
1
2
0
1
2
+
DEXGW
DEXGW
DEXGW
#
--
++
--
++
--
++
LVD
P (m
mH
g)
C
- dP
/dt (
mm
Hg/
s)
Reperfusion (min)
Ischemia
+ dP
/dt (
mm
Hg/
s)
Ischemia
LVE
DP
(mm
Hg)
Reperfusion (min)
Ischemia
HR
(bpm
)
Reperfusion (min)
Ischemia
Tota
l LD
H a
ctiv
ity (I
U)
GW GW+DEX
0
20
40
60
80
100
120GWGW+DEX
Ischemia
Reperfusion (min)6050403020100BSL
0
20
40
60
80
0
100
200
300
400
500
0
1000
2000
3000
4000
5000
0
1000
2000
3000
4000
5000
6050403020100BSL 6050403020100BSL
6050403020100BSLReperfusion (min)
6050403020100BSL
n.s.
n.s. n.s. n.s. n.s. n.s.
n.s.n.s.
n.s. n.s.n.s.
n.s.
n.s.n.s. n.s. n.s.
n.s.n.s.
n.s.
n.s.n.s.
n.s. n.s. n.s.
n.s.n.s. n.s. n.s. n.s. n.s.
A B
PP
AR
γ ac
tivity
(arb
itrar
y un
it)
0
0.025
DEXGW
#
n.s.
-+-
+-+-
+
0.05n.s.
n.s.
#n.s.
#
n.s.
+-
-+
+-
-+
+-
-+
+-
-+
0
5
10
15
20 n.s.
Figure 6. Cardioprotective effects of dexamethasone (DEX) are significantly blunted in peroxisome proliferator-activated receptor-γ (PPARγ) antagonist GW9662-pretreated mice. Wild-type (WT) and GW9662-pretreated mice were injected with either 2 mg/kg, IP, DEX or the vehicle control. A, PPARγ activation in 6 h after DEX injection. B, PPARγ-dependent Nrf2 (Nfe2l2) expression in the hearts. Six h after injection, hearts were isolated and subjected to quantitative polymerase chain reaction. The data are mean±SEM (n=5 in each group). The data were compared using ANOVA with a post hoc Tukey–Kramer multiple-comparison test. C, Hearts were isolated from WT and GW9662-pretreated mice and subjected to 25-min total global ischemia followed by aerobic reperfusion. Data show the time course of recovery of left ventricular (LV) developed pressure (LVDP), LV end-diastolic pressure (LVEDP), heart rate (HR), and ±dP/dt, as well as lactate dehydrogenase (LDH) release into the perfusate in hearts. Data are mean±SEM (n=6 in each group). The data were compared using the unpaired Student t test. BSL indicates baseline; and ns, not significant.
by guest on May 17, 2018
http://hyper.ahajournals.org/D
ownloaded from

Katsumata et al Cardioprotection in PGD2-Enriched Milieu 87
PerspectiveThe global structure and function of the heart are determined by the cross talk between cardiomyocytes and surrounding noncar-diomyocyte cells, including fibroblasts, vascular cells, immune cells, neuronal cells, and cardiac progenitor cells. Each of these cell types exhibits a unique expression pattern for PGD
2 recep-
tors, as well as cell-type specific downstream signaling and transcriptional activity. Thus, the pathophysiological effects of PGD
2 in the diseased heart are undoubtedly even more spatially
and temporally complicated than described herein, and future studies are warranted to investigate this further.
AcknowledgmentsWe thank Y. Miyake, K. Tamaki, and M. Itaya for technical assistance.
Sources of FundingThis work was supported by notable awards for research achievements in science and technology (Takeda Science Foundation), by the Mochida Memorial Foundation for Medical and Pharmaceutical Research, by a PRESTO (Metabolism and Cellular Function) grant from the Japanese Science and Technology Agency (to M.S.), and by a grant from the Ministry of Education, Culture, and Science, Japan (to K.S.), and Japan Science and Technology Agency (JST), Exploratory Research for Advanced Technology (ERATO), Suematsu Gas Biology Project, in Tokyo, 160-8582 (to M.S.), and by the JST-Precursory Research for Embryonic Science and Technology (PRESTO) program (to Y.S.).
DisclosuresNone.
References 1. Shinmura K, Tang XL, Wang Y, Xuan YT, Liu SQ, Takano H, Bhatnagar
A, Bolli R. Cyclooxygenase-2 mediates the cardioprotective effects of the late phase of ischemic preconditioning in conscious rabbits. Proc Natl Acad Sci U S A. 2000;97:10197–10202.
2. Timmers L, Sluijter JP, Verlaan CW, Steendijk P, Cramer MJ, Emons M, Strijder C, Gründeman PF, Sze SK, Hua L, Piek JJ, Borst C, Pasterkamp G, de Kleijn DP. Cyclooxygenase-2 inhibition increases mortality, enhances left ventricular remodeling, and impairs systolic function after myocardial infarction in the pig. Circulation. 2007;115:326–332.
3. Dowd NP, Scully M, Adderley SR, Cunningham AJ, Fitzgerald DJ. Inhibition of cyclooxygenase-2 aggravates doxorubicin-mediated cardiac injury in vivo. J Clin Invest. 2001;108:585–590.
4. Takahashi T, Zhu SJ, Sumino H, Saegusa S, Nakahashi T, Iwai K, Morimoto S, Kanda T. Inhibition of cyclooxygenase-2 enhances myocardial damage in a mouse model of viral myocarditis. Life Sci. 2005;78:195–204.
5. Nakamura H, Kunitsugu I, Fukuda K, Matsuzaki M, Sano M. Diverse stage-dependent effects of glucocorticoids in a murine model of viral myocarditis. J Cardiol. 2013;61:237–242.
6. Tokudome S, Sano M, Shinmura K, et al. Glucocorticoid protects rodent hearts from ischemia/reperfusion injury by activating lipocalin-type
prostaglandin D synthase-derived PGD2 biosynthesis. J Clin Invest. 2009;119:1477–1488.
7. Eguchi Y, Eguchi N, Oda H, Seiki K, Kijima Y, Matsu-ura Y, Urade Y, Hayaishi O. Expression of lipocalin-type prostaglandin D synthase (beta-trace) in human heart and its accumulation in the coronary circulation of angina patients. Proc Natl Acad Sci U S A. 1997;94:14689–14694.
8. Narumiya S. Physiology and pathophysiology of prostanoid receptors. Proc Jpn Acad, Ser B. 2007;83:296–319.
9. Kansanen E, Kivelä AM, Levonen AL. Regulation of Nrf2-dependent gene expression by 15-deoxy-Delta12,14-prostaglandin J2. Free Radic Biol Med. 2009;47:1310–1317.
10. Bell-Parikh LC, Ide T, Lawson JA, McNamara P, Reilly M, FitzGerald GA. Biosynthesis of 15-deoxy-delta12,14-PGJ2 and the ligation of PPARgamma. J Clin Invest. 2003;112:945–955.
11. Zhang Y, Sano M, Shinmura K, Tamaki K, Katsumata Y, Matsuhashi T, Morizane S, Ito H, Hishiki T, Endo J, Zhou H, Yuasa S, Kaneda R, Suematsu M, Fukuda K. 4-hydroxy-2-nonenal protects against cardiac ischemia-reperfusion injury via the Nrf2-dependent pathway. J Mol Cell Cardiol. 2010;49:576–586.
12. Narumiya S, Sugimoto Y, Ushikubi F. Prostanoid receptors: structures, properties, and functions. Physiol Rev. 1999;79:1193–1226.
13. Abramovitz M, Boie Y, Nguyen T, Rushmore TH, Bayne MA, Metters KM, Slipetz DM, Grygorczyk R. Cloning and expression of a cDNA for the human prostanoid FP receptor. J Biol Chem. 1994;269:2632–2636.
14. Yoshikawa N, Nagasaki M, Sano M, Tokudome S, Ueno K, Shimizu N, Imoto S, Miyano S, Suematsu M, Fukuda K, Morimoto C, Tanaka H. Ligand-based gene expression profiling reveals novel roles of glucocor-ticoid receptor in cardiac metabolism. Am J Physiol Endocrinol Metab. 2009;296:E1363–E1373.
15. Sun H, Sheveleva E, Xu B, Inoue H, Bowden TG, Chen QM. Corticosteroids induce COX-2 expression in cardiomyocytes: role of glucocorticoid receptor and C/EBP-beta. Am J Physiol Cell Physiol. 2008;295:C915–C922.
16. Giugliano GR, Giugliano RP, Gibson CM, Kuntz RE. Meta-analysis of corticosteroid treatment in acute myocardial infarction. Am J Cardiol. 2003;91:1055–1059.
17. Smith WL, Urade Y, Jakobsson PJ. Enzymes of the cyclooxygenase path-ways of prostanoid biosynthesis. Chem Rev. 2011;111:5821–5865.
18. Jovanović N, Pavlović M, Mircevski V, Du Q, Jovanović A. An unex-pected negative inotropic effect of prostaglandin F2alpha in the rat heart. Prostaglandins Other Lipid Mediat. 2006;80:110–119.
19. Karmazyn M, Tani M, Neely JR. Effect of prostaglandins I2 (prostacyclin) and F2 alpha on function, energy metabolism, and calcium uptake in isch-aemic/reperfused hearts. Cardiovasc Res. 1993;27:396–402.
20. Hara A, Yuhki K, Fujino T, Yamada T, Takayama K, Kuriyama S, Takahata O, Karibe H, Okada Y, Xiao CY, Ma H, Narumiya S, Ushikubi F. Augmented cardiac hypertrophy in response to pressure overload in mice lacking the prostaglandin I2 receptor. Circulation. 2005;112:84–92.
21. Polvani S, Tarocchi M, Galli A. PPARγ and Oxidative Stress: Con(β) Catenating NRF2 and FOXO. PPAR Res. 2012;2012:641087.
22. Saleem S, Zhuang H, de Brum-Fernandes AJ, Maruyama T, Narumiya S, Doré S. PGD(2) DP1 receptor protects brain from ischemia-reperfusion injury. Eur J Neurosci. 2007;26:73–78.
23. Faveeuw C, Gosset P, Bureau F, Angeli V, Hirai H, Maruyama T, Narumiya S, Capron M, Trottein F. Prostaglandin D2 inhibits the production of inter-leukin-12 in murine dendritic cells through multiple signaling pathways. Eur J Immunol. 2003;33:889–898.
What Is New?•This is the first report on the mechanism behind cardioprotection by
lipocalin-type prostaglandin D (PGD) synthase–derived PGD2.
•This is the first report of the FP receptor significance in PGD2 signaling.
What Is Relevant?• Lipocalin-type PGD
2 synthase is constitutively and highly expressed in
cardiomyocytes and protects the heart under stressful conditions.•Both PGD
2 and the J-series metabolites of PGD
2 activate Nrf2 with differ-
ent temporal dynamics.
SummaryLipocalin-type PGD synthase–derived PGD
2 biosynthesis protects the
heart against I/R injury via the activation of Nrf2. This study is the first to show that the FP receptor serves as a functionally relevant PGD
2
receptor on cardiomyocytes. PGD2–FP receptor signaling participates
in concert with the J-series metabolites of PGD2 to activate Nrf2.
Novelty and Significance
by guest on May 17, 2018
http://hyper.ahajournals.org/D
ownloaded from

Makoto Suematsu, Keiichi Fukuda and Motoaki SanoHideyuki Ito, Xiaoxiang Yan, Kentaro Ito, Shinsuke Yuasa, Masaki Ieda, Yoshihiro Urade,
Yoshinori Katsumata, Ken Shinmura, Yuki Sugiura, Shugo Tohyama, Tomohiro Matsuhashi,Reperfusion Injury by Activating Nrf2
− and Its Metabolites Protect the Heart Against Ischemia2Endogenous Prostaglandin D
Print ISSN: 0194-911X. Online ISSN: 1524-4563 Copyright © 2013 American Heart Association, Inc. All rights reserved.
is published by the American Heart Association, 7272 Greenville Avenue, Dallas, TX 75231Hypertension doi: 10.1161/HYPERTENSIONAHA.113.01639
2014;63:80-87; originally published online October 7, 2013;Hypertension.
http://hyper.ahajournals.org/content/63/1/80World Wide Web at:
The online version of this article, along with updated information and services, is located on the
http://hyper.ahajournals.org/content/suppl/2013/10/07/HYPERTENSIONAHA.113.01639.DC1Data Supplement (unedited) at:
http://hyper.ahajournals.org//subscriptions/
is online at: Hypertension Information about subscribing to Subscriptions:
http://www.lww.com/reprints Information about reprints can be found online at: Reprints:
document. Permissions and Rights Question and Answer this process is available in the
click Request Permissions in the middle column of the Web page under Services. Further information aboutOffice. Once the online version of the published article for which permission is being requested is located,
can be obtained via RightsLink, a service of the Copyright Clearance Center, not the EditorialHypertensionin Requests for permissions to reproduce figures, tables, or portions of articles originally publishedPermissions:
by guest on May 17, 2018
http://hyper.ahajournals.org/D
ownloaded from

Endogenous Prostaglandin D2 and its Metabolites Protect the Heart Against
Ischemia-Reperfusion Injury by Activating Nrf2
Authors:
Yoshinori Katsumata, MD 1
, Ken Shinmura, MD 2
, Yuki Sugiura, PhD 3,5
, Shugo
Tohyama 1
, MD, Tomohiro Matsuhashi, MD 1
, Hideyuki Ito, MD 1
, Xiaoxiang Yan, MD 1,
Kentaro Ito, BS 1, Shinsuke Yuasa, PhD, MD
1, Masaki Ieda, MD
1,4, Yoshihiro Urade,
PhD,6 Makoto Suematsu, MD
3 , Keiichi Fukuda, MD
1 , Motoaki Sano, MD
1,7.
1Department of Cardiology,
2Department of Geriatric Medicine,
3Department of
Biochemistry, 4Department of Clinical and Molecular Cardiovascular Research, Keio
University School of Medicine, Tokyo, Japan 5Precursory Research for Embryonic Science and Technology (PRESTO), Japan
Science and Technology Agency, Tokyo, Japan
6Department of Molecular and Behavioral Biology, Osaka Bioscience Institute, Suita,
Japan 7Address correspondence to: Dr. Motoaki Sano, Department of Cardiology, Keio
University School of Medicine, 35 Shinanomachi Shinjuku-ku, Tokyo, 160-8582,
Japan; FAX: 81-3-5363-3875; Phone: 81-3-5363-3874; e-mail: [email protected]
Short title: Cardioprotection in PGD2-enriched milieu

A detailed, expanded Methods section
Mice
L-PGDS-knockout (KO) mice 1, FP-KO mice
2, DP1-KO mice
3, DP2-KO mice
4, and
Nrf2-KO mice 5 were generated as described previously, all from a C57BL6/J
background. Mice weighing 20-25 g, male, and 10-12 weeks old were used in this study.
All experimental procedures and protocols were approved by the Animal Care and Use
Committees of Keio University.
DEX administration
There were no significant differences in HW, BW, and HW/BW ratio among
wild-type, DP1-KO, DP2-KO, and FP-KO mice. Histological analysis of the heart
revealed no cardiac fibrosis in DP1-KO, DP2-KO, and FP-KO mice prior to
commencing the isolated heart experiments. Echocardiographic examination showed no
difference in LV systolic function among wild-type, DP1-KO, DP2-KO, and FP-KO
mice.
We used dexamethasone (DEX) at a dose of 2 mg/kg body weight in this study based
on our preliminary experiments in mice showing induction of many glucocorticoid
target genes with DEX treatment at that same concentration, which is equivalent to the
dose used in humans 6,7
. Because our preliminary experiments also showed that the
mRNA expression of several well-known Nrf2 target genes started to increase between
3 and 12 h after DEX administration and that the cardioprotective effect of DEX was
maximal when administered 24 h prior to I/R injury, we chose 24 h prior to injury as the
DEX pretreatment administration point.
Cell culture
Neonatal ventricular myocytes from 1−2-day-old Sprague-Dawley rats were subjected
to Percoll gradient centrifugation and differential plating to enrich the cardiac myocyte
population and to deplete non-myocytes, as reported previously 8. Cardiomyocytes were
cultured in a mixture of Dulbecco’s modified Eagle’s medium (DMEM) and M199 with
10% fetal bovine serum (FBS; Biowest, Nuaille, France). After 24 hours of serum
starvation, cardiomyocytes were stimulated with DEX, fluprostenol, or 15d-PGJ2.
Maintenance of human embryonic stem cells
The human embryonic stem cell (ESC) line (KhES-2) was obtained from the
Department of Development and Differentiation, Institute for Frontier Medical Sciences,

Kyoto University and used in conformity with the Guidelines for Derivation and
Utilization of Human Embryonic Stem Cells of the Ministry of Education, Culture,
Sports, Science, and Technology, Japan. We maintained human ESCs on MEFs in
DMEM/nutrient mixture F-12 Ham 1:1 (DMEM-F12; Sigma-Aldrich, St. Louis, MO,
USA) supplemented with 20% knockout serum replacement (KSR; Invitrogen, Carlsbad,
CA, USA), 0.1 mM MEM non-essential amino acids solution (Sigma-Aldrich), 2 mM
L-glutamine (Sigma-Aldrich), 0.1 mM β-mercaptoethanol (Sigma-Aldrich), and 4 ng/ml
basic fibroblast growth factor (bFGF; Wako, Osak-shi, Osaka, Japan).
Differentiation of human pluripotent stem cell-derived cardiomyocytes
We cultured the enzymatically detached undifferentiated colonies of ESCs with MEMα
(Wako) supplemented with 5% FBS and 0.1 mM β-mercaptoethanol in bacterial Petri
dishes to form embryoid bodies (EBs). Fourteen to twenty days later, we observed EBs
containing rhythmically beating cells. Media were changed once a week.
Purification of human ESC-derived cardiomyocytes
Glucose-free DMEM (no glucose, no pyruvate; Invitrogen) supplemented with 4 mM
lactate medium was produced using 1 M lactate stock solution prepared from diluting
10 M lactate (Wako) with sterile 1 M Na-HEPES (Sigma-Aldrich). The human
PSC-derived EBs at differentiation day 20 to 30 were extensively washed with and
exposed to the selection medium. Media were changed every 2 or 3 days to eliminate
dead cells by rapid flushing using 40-μm filters (Becton Dickinson, Franklin Lakes, NJ,
USA), as described previously 9. The resultant human ESC-derived cardiomyocytes
were used after metabolic selection.
Short interference RNA oligonucleotides and transfection
The short interference (si) RNA oligonucleotides used in the present study against rat
Ptgfr and nfel2l genes, as well as control siRNA, were obtained from Ambion. These
siRNA oligonucleotides were transfected into cardiomyocytes using Lipofectamine
RNAiMAX reagent (Invitrogen).
Western blotting
Equal amounts of total protein (15-20 g) from cultured cardiomyocytes and heart
lysates were subjected to sodium dodecyl sulfate–polyacrylamide gel electrophoresis
(SDS-PAGE) and transferred to membranes for immunoblotting. The primary
antibodies used were rabbit anti-FP receptor, anti-L-PGDS (both from Cayman

Chemical, Ann Arbor, MI, USA), and anti-Nrf-2 (Santa Cruz Biotechnology, Santa Cruz,
CA, USA). Protein expression was visualized with horseradish peroxidase-conjugated
secondary antibodies and enhanced chemiluminescence (Amersham Biosciences,
Piscataway, NJ, USA), and was detected using the LAS-3000 luminoimager (Fujifilm
techno products, Ayase-shi, Kanagawa, Japan). Nuclear extracts were prepared as
previously described 10
.
Gene expression
Oligo-DNA microarray analysis was performed using the “3D-Gene” Mouse Oligo chip
24k (Toray Industries, Chuo-ku, Tokyo, Japan) containing 23,522 distinct genes. For
efficient hybridization, this microarray is constructed in three dimensions, with an
additional space between the probes and cylinder stems and with 70-mer
oligonucleotide probes on top. Total RNA was labeled with Cy3 or Cy5 using the
Amino Allyl MessageAMP II aRNA Amplification Kit (Applied Biosystems, Foster, CA,
USA) and the Cy3- or Cy5-labeled RNA samples were pooled in hybridization buffer
before being hybridized for 16 h using the protocols supplied by the manufacturer
(www.3d-gene.com). Hybridization signals were scanned using a ScanArray Express
Scanner (Perkin Elmer, Fremont, CA USA), and processed using GenePixPro ver. 5.0
software (Molecular Devices, Silicon valley, CA, USA). Signals detected for each gene
were normalized using a global normalization method (median Cy3/Cy5 ratio = 1). A
heat map of 22 genes was processed using Cluster 2.0 software
(http://rana.lbl.gov/eisen/).
For quantitative real-time polymerase chain reaction (PCR), total RNA samples from
cultured cardiomyocytes and hearts were prepared using Trizol reagent (Invitrogen),
according to the manufacturer’s instructions. Samples of total RNA (2 μg) were
reverse-transcribed using an RNA PCR Kit (Takara Biotechnology, Otsu, Shiga, Japan),
and the resulting cDNA was used as a PCR template. RNA levels were then determined
by real-time PCR using the ABI PRISM 7700 Sequence Detector (Applied Biosystems).
Predesigned gene-specific primer and probe sets (TaqMan Gene Expression Assays)
were used, and 18s ribosomal RNA was amplified as an internal control. Relative gene
expression levels (i.e., the amount of target gene normalized against that of the internal
control) were calculated using the comparative cycle threshold (Ct) method as 2Ct
.
Quantification of PGs
The amount of PGs in the heart was quantified using the method of Kita et al. 11
. Briefly,
a triple-quadrupole mass spectrometer equipped with an electrospray ionization (ESI)

ion source (LCMS-8030; Shimadzu Corporation, Kyoto, Kyoto, Japan) was used in the
negative-ESI and multiple reaction monitoring (MRM) modes. For accurate
quantification, we used an internal standard consisting of a mixture of
deuterium-labeled PG.
Langendorff perfusion of the heart
In the Langendorff-perfused heart experiment, we excluded 17 mice that showed poor
LV function during the initial perfusion or massive leakage from the aorta from the
further analysis. The table S1 presents the number of mice excluded in each group. We
speculated that technical probems were mainly responsible for these exclusions because
there was no difference in the ratio of excluded mice number to total mice number
assigned to each experiment among all strains examined.
After hearts had been excised from heparinized mice, they were perfused with
modified Krebs-Henseleit buffer (composition [in mmol/L]: NaCl 120; NaHCO3 25;
KCl 5.9; MgSO4 1.2; CaCl2 1.75; glucose 10) gassed with 95% O2 and 5% CO2 at 37°C
according to the Langendorff procedure. Coronary perfusion pressure was maintained at
90 mmHg. A plastic catheter with a polyethylene balloon was inserted into the LV
through the left atrium. Before the induction of ischemia, LV end-diastolic pressure
(LVEDP) was adjusted to 10 mmHg by filling the balloon with water. Indices of LV
function (LV systolic pressure [LVSP], LVEDP, LV developed pressure [LVDP;
calculated as the difference between LVSP and LVEDP], +dP/dt, and –dP/dt) were
recorded as described previously 8. Total lactate dehydrogenase (LDH) activity released
into the perfusate was measured using a commercially available kit (Sigma-Aldrich).
PPARγ activity measurements in muscle
Nuclear extracts were prepared from the heart homogenates using a nuclear extract kit
(Active Motif, Rixensart, Belgium) according to the manufacturer’s instructions. The
protein was measured using a BCA Protein Assay kit (Thermo Fisher Scientific Inc,
Rockford, IL). PPAR-γ activation was assayed using an ELISA-based PPAR-γ
activation TransAM kit (Active Motif, Rixensart, Belgium), according to the
manufacturer’s instructions. Ten micrograms of nuclear protein samples were incubated
for 1 hour in a 96-well plate coated with an oligonucleotide that contains a PPAR
response element domain (5ʹ-AACTAGGTCAAAGGTCA-3ʹ), to which the activated
PPAR-γ contained in nuclear extracts specifically binds. After washing, PPAR-γ
antibody (1:1000 dilutions) was added to these wells and incubated for 1 hour. After
incubation for 1 hour with a secondary horseradish peroxidase-conjugated antibody

(1:1000 dilution), specific binding was detected by colorimetric estimation at 450 nm
with a reference wavelength of 655 nm. PPARγ antagonist GW9662 (3 mg/kg/body
weight) was administered prior to DEX administration and I/R as previously reported 12
.
Statistical analyses
Values are presented as mean ± SEM. The n values represent independent experiments.
The significance of differences between two means was evaluated using two-tailed
Student’s unpaired t-tests. Multiple comparisons involving more than three groups were
made using one-way analysis of variance (ANOVA) with a post hoc Tukey-Kramer
multiple-comparison test. P < 0.05 was considered significant.

Supplement Reference
1. Eguchi Y, Eguchi N, Oda H, Seiki K, Kijima Y, Matsu-ura Y, Urade Y, Hayaishi O.
Expression of lipocalin-type prostaglandin d synthase (beta-trace) in human heart
and its accumulation in the coronary circulation of angina patients. Proc Natl Acad
Sci U S A. 1997;94:14689-14694.
2. Sugimoto Y, Yamasaki A, Segi E, Tsuboi K, Aze Y, Nishimura T, Oida H, Yoshida
N, Tanaka T, Katsuyama M, Hasumoto K, Murata T, Hirata M, Ushikubi F, Negishi
M, Ichikawa A, Narumiya S. Failure of parturition in mice lacking the
prostaglandin f receptor. Science. 1997;277:681-683.
3. Matsuoka T, Hirata M, Tanaka H, Takahashi Y, Murata T, Kabashima K, Sugimoto
Y, Kobayashi T, Ushikubi F, Aze Y, Eguchi N, Urade Y, Yoshida N, Kimura K,
Mizoguchi A, Honda Y, Nagai H, Narumiya S. Prostaglandin d2 as a mediator of
allergic asthma. Science. 2000;287:2013-2017.
4. Satoh T, Moroi R, Aritake K, Urade Y, Kanai Y, Sumi K, Yokozeki H, Hirai H,
Nagata K, Hara T, Utsuyama M, Hirokawa K, Sugamura K, Nishioka K, Nakamura
M. Prostaglandin d2 plays an essential role in chronic allergic inflammation of the
skin via crth2 receptor. J Immunol. 2006;177:2621-2629.
5. Itoh K, Chiba T, Takahashi S, Ishii T, Igarashi K, Katoh Y, Oyake T, Hayashi N,
Satoh K, Hatayama I, Yamamoto M, Nabeshima Y. An nrf2/small maf heterodimer
mediates the induction of phase ii detoxifying enzyme genes through antioxidant
response elements. Biochem Biophys Res Commun. 1997;236:313-322.
6. Guney S, Schuler A, Ott A, Hoschele S, Zugel S, Baloglu E, Bartsch P, Mairbaurl H.
Dexamethasone prevents transport inhibition by hypoxia in rat lung and alveolar
epithelial cells by stimulating activity and expression of na+-k+-atpase and
epithelial na+ channels. Am J Physiol Lung Cell Mol Physiol.
2007;293:L1332-1338.
7. Kandil E, Lin YY, Bluth MH, Zhang H, Levi G, Zenilman ME. Dexamethasone
mediates protection against acute pancreatitis via upregulation of
pancreatitis-associated proteins. World J Gastroenterol. 2006;12:6806-6811.
8. Tokudome S, Sano M, Shinmura K, Matsuhashi T, Morizane S, Moriyama H,
Tamaki K, Hayashida K, Nakanishi H, Yoshikawa N, Shimizu N, Endo J, Katayama
T, Murata M, Yuasa S, Kaneda R, Tomita K, Eguchi N, Urade Y, Asano K,
Utsunomiya Y, Suzuki T, Taguchi R, Tanaka H, Fukuda K. Glucocorticoid protects
rodent hearts from ischemia/reperfusion injury by activating lipocalin-type
prostaglandin d synthase-derived pgd2 biosynthesis. J Clin Invest.

2009;119:1477-1488.
9. Tohyama S, Hattori F, Sano M, Hishiki T, Nagahata Y, Matsuura T, Hashimoto H,
Suzuki T, Yamashita H, Satoh Y, Egashira T, Seki T, Muraoka N, Yamakawa H,
Ohgino Y, Tanaka T, Yoichi M, Yuasa S, Murata M, Suematsu M, Fukuda K.
Distinct metabolic flow enables large-scale purification of mouse and human
pluripotent stem cell-derived cardiomyocytes. Cell Stem Cell. 2013;12:127-137.
10. Zhang Y, Sano M, Shinmura K, Tamaki K, Katsumata Y, Matsuhashi T, Morizane S,
Ito H, Hishiki T, Endo J, Zhou H, Yuasa S, Kaneda R, Suematsu M, Fukuda K.
4-hydroxy-2-nonenal protects against cardiac ischemia-reperfusion injury via the
nrf2-dependent pathway. J Mol Cell Cardiol. 2010;49:576-586.
11. Kita Y, Takahashi T, Uozumi N, Shimizu T. A multiplex quantitation method for
eicosanoids and platelet-activating factor using column-switching reversed-phase
liquid chromatography-tandem mass spectrometry. Anal Biochem.
2005;342:134-143.
12. Nagahama R, Matoba T, Nakano K, Kim-Mitsuyama S, Sunagawa K, Egashira K.
Nanoparticle-mediated delivery of pioglitazone enhances therapeutic
neovascularization in a murine model of hindlimb ischemia. Arterioscler Thromb
Vasc Biol. 2012;32:2427-2434.
13. Okamatsu S, Lefer AM. Cardiac effects of prostaglandins during global ischemia in
isolated perfused cat hearts. Prostaglandins Leukot Med. 1982;8:1-10.
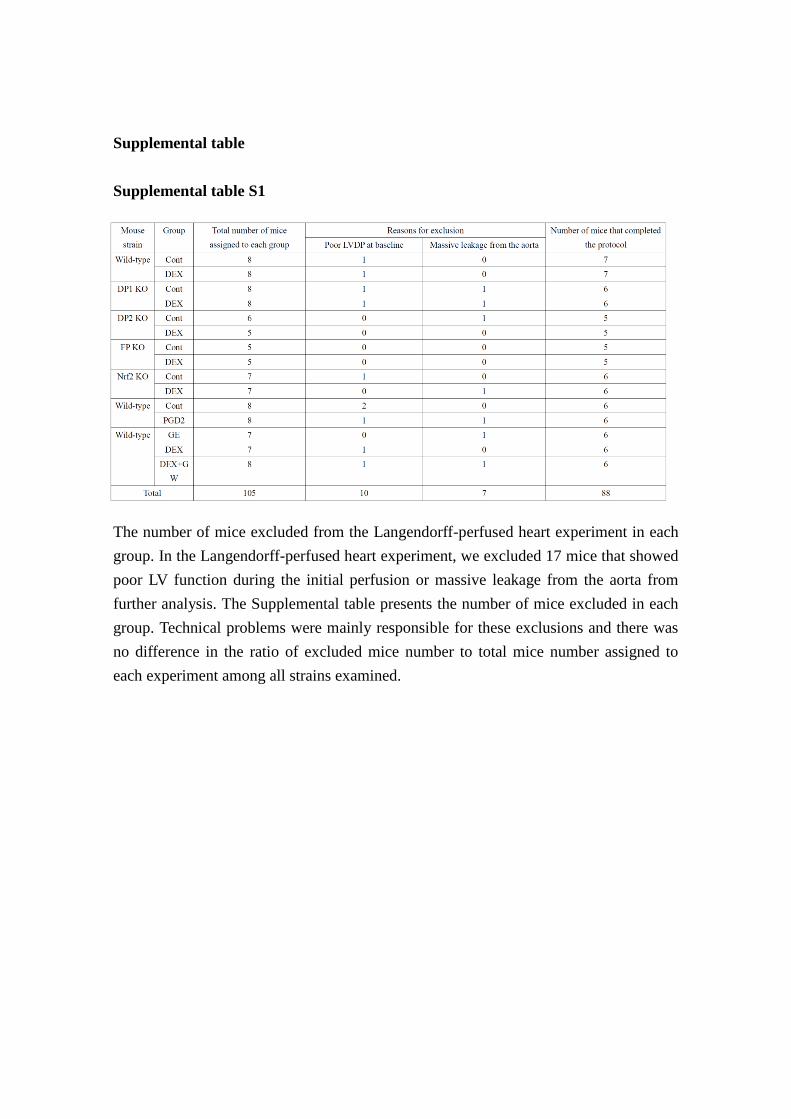
Supplemental table
Supplemental table S1
The number of mice excluded from the Langendorff-perfused heart experiment in each
group. In the Langendorff-perfused heart experiment, we excluded 17 mice that showed
poor LV function during the initial perfusion or massive leakage from the aorta from
further analysis. The Supplemental table presents the number of mice excluded in each
group. Technical problems were mainly responsible for these exclusions and there was
no difference in the ratio of excluded mice number to total mice number assigned to
each experiment among all strains examined.
0
20
40
60
80
100
120Con (n=6)PGD2 (n=6)
6050403020100BSL0
20
40
60
80
6050403020100BSL0
150
300
450
600
750
6050403020100BSL
0
1000
2000
3000
4000
5000
6050403020100BSL0
1000
2000
3000
4000
5000
6050403020100BSL
LVD
P (m
mH
g)
Reperfusion (min)
Ischemia
LVE
DP
(mm
Hg)
Reperfusion (min)
Ischemia
HR
(bpm
)
Reperfusion (min)
Ischemia+
LVdP
/dt (
mm
Hg/
s)
Reperfusion (min)
- LV
dP/d
t (m
mH
g/s)
Reperfusion (min)
Ischemia Ischemia
Tota
l LD
H a
ctiv
ity (I
U)
Con PGD2
S1
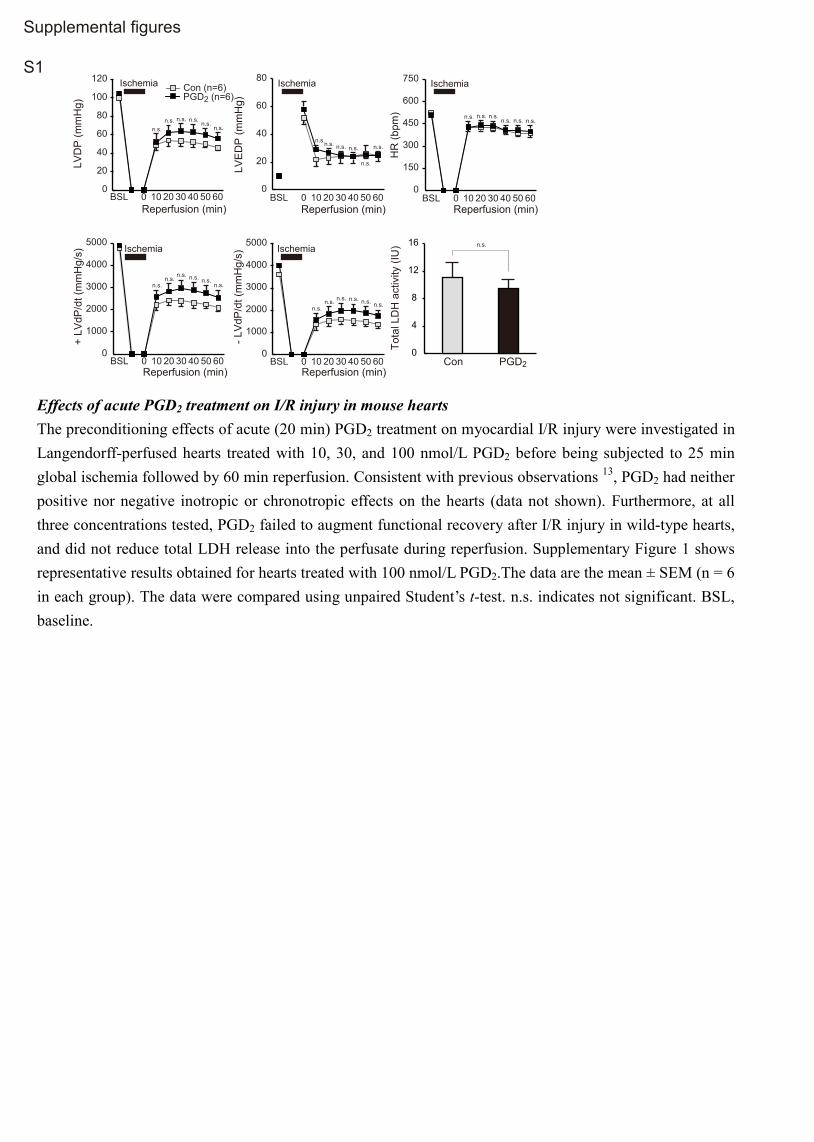
n.s.
Supplemental figures
0
4
8
12
16
n.s.n.s. n.s. n.s.
n.s.n.s.
n.s.n.s. n.s. n.s.
n.s.
n.s.
n.s. n.s. n.s.n.s. n.s. n.s.
n.s.n.s.
n.s. n.s. n.s.n.s.
n.s.n.s. n.s. n.s. n.s. n.s.
Effects of acute PGD2 treatment on I/R injury in mouse hearts The preconditioning effects of acute (20 min) PGD2 treatment on myocardial I/R injury were investigated in Langendorff-perfused hearts treated with 10, 30, and 100 nmol/L PGD2 before being subjected to 25 min global ischemia followed by 60 min reperfusion. Consistent with previous observations 13, PGD2 had neither positive nor negative inotropic or chronotropic effects on the hearts (data not shown). Furthermore, at all three concentrations tested, PGD2 failed to augment functional recovery after I/R injury in wild-type hearts, and did not reduce total LDH release into the perfusate during reperfusion. Supplementary Figure 1 shows representative results obtained for hearts treated with 100 nmol/L PGD2.The data are the mean ± SEM (n = 6 in each group). The data were compared using unpaired Student’s t-test. n.s. indicates not significant. BSL, baseline.

S2
Dexamethasone (DEX) induces Nrf2-related gene expression in hearts in a lipocalin-type prostaglandin (PG) D synthase (L-PGDS)-dependent manner. Adult 10-week-old male mice (C57BL6J) or Ptgds–/– (L-PGDS-knockout [KO]) mice were injected with 2 mg/kg, i.p., DEX and the time course of changed in gene expression was determined by quantitative polymerase chain reaction. The data are the mean ± SEM (n = 6). #P < 0.05, compared with WT control (unpaired Student’s t-test). *P < 0.05 compared with L-PGDS-KO control (unpaired Student’s t-test). Gclc, glutamate-cysteine ligase, catalytic subunit; Gsr, glutathione reductase; G6pdx, glucose-6-phosphate dehydrogenase X-linked; Hmox-1, heme oxygenase 1; Gclm, glutamate-cysteine ligase, modifier subunit; Cat, catalase; Nqo1, NAD(P)H dehydrogenase, quinone 1.
0 0
1
2
3 L-PGDS-KOWT
h
gene
exp
ress
ion
(fo
ld c
hang
e)Gclc
0
1
2Gsr
0
1
2G6pdx
3 6 12 24 0 h
gene
exp
ress
ion
(fo
ld c
hang
e)
3 6 12 24 0 h
gene
exp
ress
ion
(fo
ld c
hang
e)
3 6 12 24
**
* * ** * *
0
1
2
0
1
2
3
0
4
8
0
1
2Gclm Cat
0 h
gene
exp
ress
ion
(fo
ld c
hang
e)
3 12 24 0 h
gene
exp
ress
ion
(fo
ld c
hang
e)
3 6 12 24 0
1
2Nqo1
0 h
gene
exp
ress
ion
(fo
ld c
hang
e)
3 6 12 24
Nfe2l2
Hmox1
0 h
gene
exp
ress
ion
(fo
ld c
hang
e)
3 6 12 24
0 h
gene
exp
ress
ion
(fo
ld c
hang
e)
3 6 12 24 6
* #
* *
*
*
*
**
**#
#
#
#
##
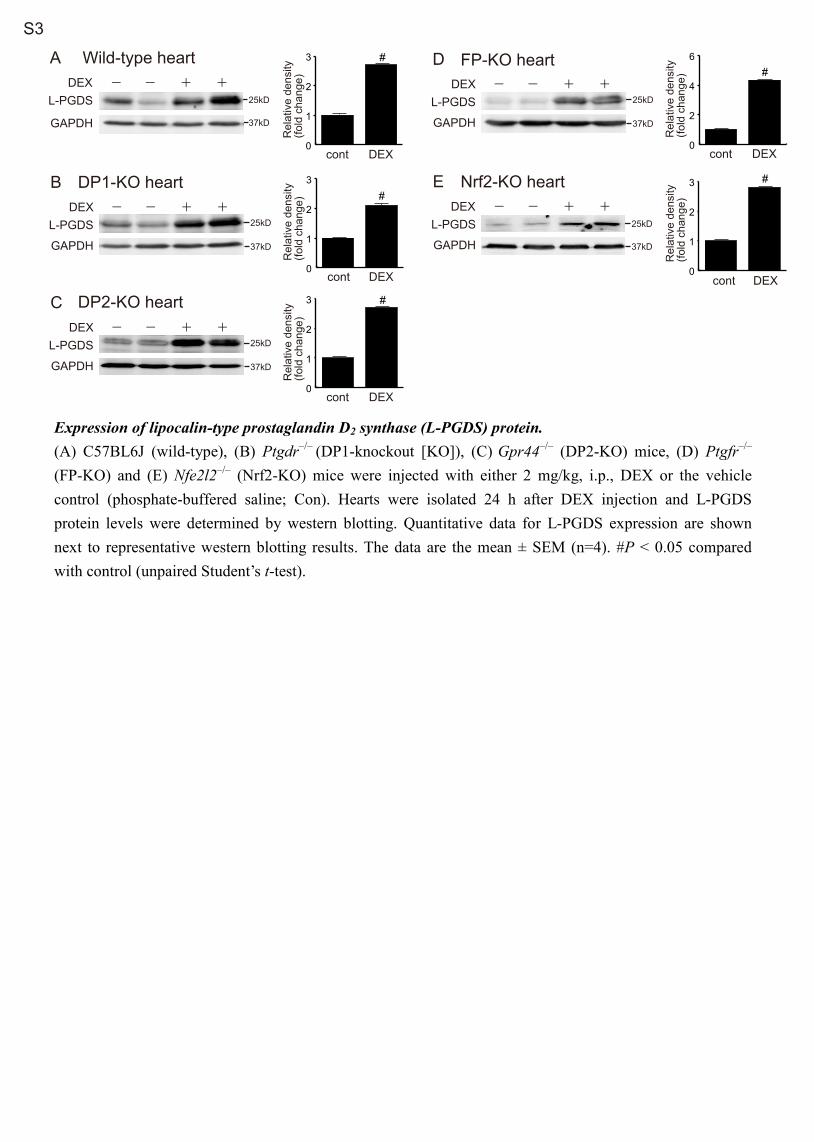
37kD
25kD
S3
A
L-PGDS
GAPDH
Wild-type heart - - + +DEX
37kD
25kD
B
L-PGDS
GAPDH
DEX
DP1-KO heart - - + +
DEXcontR
elat
ive
dens
ity
(fold
cha
nge) #
37kD
25kD
C
L-PGDS
GAPDH
DEX
DP2-KO heart - - + +
0
1
2
3
DEXcont
Rel
ativ
e de
nsity
(fo
ld c
hang
e)#
37kD
25kD
D
L-PGDS
GAPDH
DEX - - + +
0
2
4
6
DEXcont
Rel
ativ
e de
nsity
(fo
ld c
hang
e)
#
37kD
25kD
E
L-PGDS
GAPDH
- - + +DEX
0
1
2
3
DEXcont
Rel
ativ
e de
nsity
(fo
ld c
hang
e)
#
0
1
2
3
0
1
2
3
DEXcont
Rel
ativ
e de
nsity
(fo
ld c
hang
e)
# FP-KO heart
Nrf2-KO heart
Expression of lipocalin-type prostaglandin D2 synthase (L-PGDS) protein. (A) C57BL6J (wild-type), (B) Ptgdr–/– (DP1-knockout [KO]), (C) Gpr44–/– (DP2-KO) mice, (D) Ptgfr–/– (FP-KO) and (E) Nfe2l2–/– (Nrf2-KO) mice were injected with either 2 mg/kg, i.p., DEX or the vehicle control (phosphate-buffered saline; Con). Hearts were isolated 24 h after DEX injection and L-PGDS protein levels were determined by western blotting. Quantitative data for L-PGDS expression are shown next to representative western blotting results. The data are the mean ± SEM (n=4). #P < 0.05 compared with control (unpaired Student’s t-test).
0
20
40
60
80
100
120Cont (n=5)DEX (n=5)
6050403020100BSL0
20
40
60
6050403020100BSL0
150
300
450
600
750
6050403020100BSL
0
1000
2000
3000
4000
5000
6050403020100BSL0
1000
2000
3000
4000
5000
6050403020100BSL
LVD
P (m
mH
g)
Reperfusion (min)
Ischemia
+ LV
dP/d
t (m
mH
g/s)
Reperfusion (min)
- LV
dP/d
t (m
mH
g/s)
Reperfusion (min)
LVE
DP
(mm
Hg)
Reperfusion (min)
Ischemia
HR
(bpm
)
Reperfusion (min)
Ischemia
Tota
l LD
H a
ctiv
ity (I
U)Ischemia Ischemia
Con DEX
#
0
4
8
12
16
B DP2-KO heart
# #
#
#
#
0
20
40
60
80
100
120Cont (n=6)DEX(n=6)
6050403020100BSL0
20
40
60
6050403020100BSL0
150
300
450
600
750
6050403020100BSL
0
1000
2000
3000
4000
5000
6050403020100BSL0
1000
2000
3000
4000
5000
6050403020100BSL
LVD
P (m
mH
g)
Reperfusion (min)
Ischemia
+ LV
dP/d
t (m
mH
g/s)
Reperfusion (min)
- LV
dP/d
t (m
mH
g/s)
Reperfusion (min)
LVE
DP
(mm
Hg)
Reperfusion (min)
Ischemia
HR
(bpm
)
Reperfusion (min)
Ischemia
Tota
l LD
H a
ctiv
ity (I
U)Ischemia Ischemia
Con DEX
n.s.
0
4
8
12
16
A DP1-KO heart
n.s. n.s. n.s.n.s. n.s.
n.s.
n.s. n.s.n.s.
n.s. n.s. n.s.
n.s.
n.s. n.s.
n.s.n.s.
n.s.
n.s. n.s. n.s.n.s. n.s.
n.s.
n.s. n.s. n.s. n.s.
n.s.n.s.
S4
Cardioprotective effects of dexamethasone (DEX) were significantly blunted in mice lacking DP1 receptors. Mice were injected with either 2 mg/kg, i.p., DEX or vehicle control (Con) and then, 24 h later, hearts were isolated and subjected to 25 min total global ischemia followed by aerobic reperfusion. Data show the time course of recovery of left ventricular (LV) developed pressure (LVDP), LV end-diastolic pressure (LVEDP), heart rate (HR), and ±dP/dt, as well as lactate dehydrogenase (LDH) release into the perfusate, in hearts from (A) Ptgdr–/– (DP1-knockout [KO]) mice (n = 6) and (B) Gpr44–/– (DP2-KO) mice (n = 5). Data are the mean ± SEM. #P < 0.05 compared with vehicle-treated control hearts (unpaired Student’s t-test). n.s. indicates not significant. BSL, baseline.
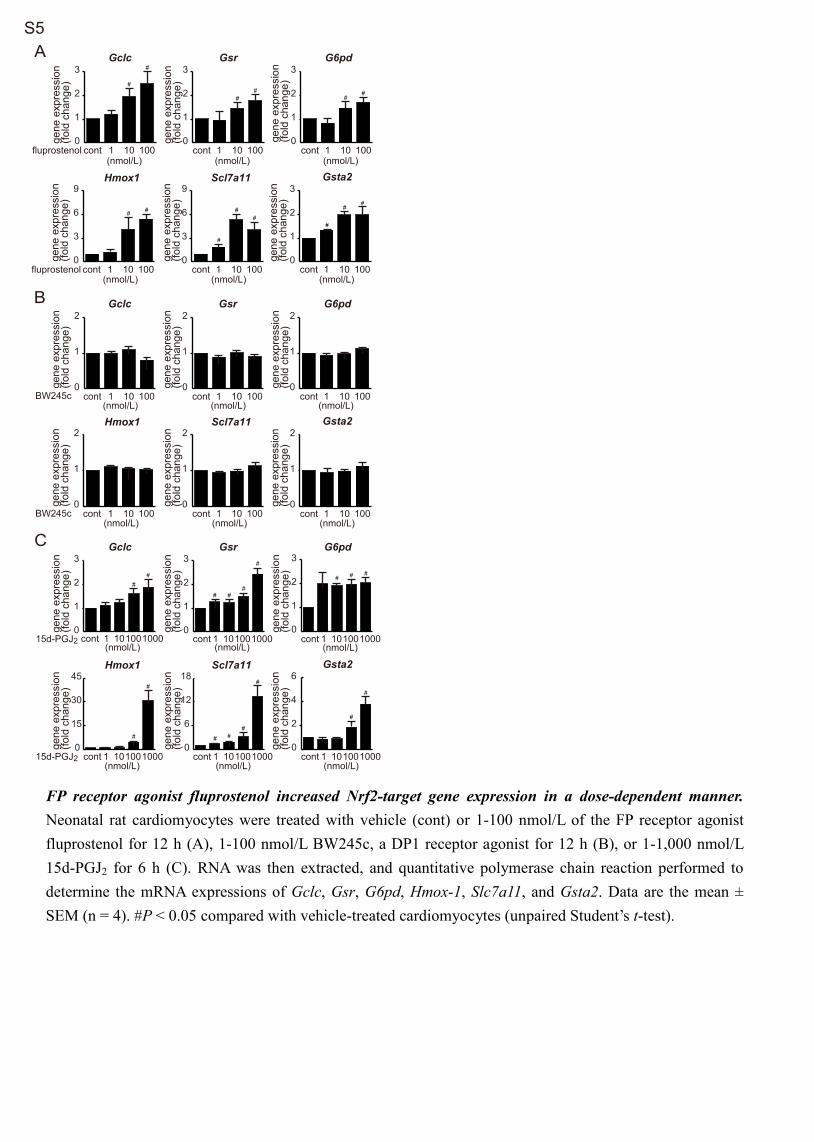
fluprostenol
fluprostenol
BW245c
BW245c
S5
B
gene
exp
ress
ion
(fold
cha
nge)
Gclc
0
1
2
cont 1 10 100(nmol/L)
0
1
2
0
1
2
0
1
2
cont 1 10 100(nmol/L)
Gsta2
cont 1 10 100(nmol/L)
cont 1 10 100(nmol/L)
Gsr G6pd
gene
exp
ress
ion
(fold
cha
nge)
gene
exp
ress
ion
(fold
cha
nge)
gene
exp
ress
ion
(fold
cha
nge)
gene
exp
ress
ion
(fold
cha
nge)
Hmox1
0
1
2
cont 1 10 100(nmol/L)
0
1
2
cont 1 10 100(nmol/L)
Scl7a11
gene
exp
ress
ion
(fold
cha
nge)
Age
ne e
xpre
ssio
n (fo
ld c
hang
e)Gclc
0
1
2
3
cont 1 10 100(nmol/L)
0
1
2
3
0
1
2
3
0
1
2
3
cont 1 10 100(nmol/L)
Gsta2
cont 1 10 100(nmol/L)
cont 1 10 100(nmol/L)
Gsr G6pd
gene
exp
ress
ion
(fold
cha
nge)
gene
exp
ress
ion
(fold
cha
nge)
gene
exp
ress
ion
(fold
cha
nge)
gene
exp
ress
ion
(fold
cha
nge)
Hmox1
0
3
6
9
cont 1 10 100(nmol/L)
0
3
6
9
cont 1 10 100(nmol/L)
Scl7a11ge
ne e
xpre
ssio
n (fo
ld c
hang
e)
#
#
#
##
##
##
# # ##
#
15d-PGJ2
15d-PGJ2
C
gene
exp
ress
ion
(fold
cha
nge)
Gclc
0
2
4
6
cont 1 10100(nmol/L)
0
1
2
3
0
1
2
3
0
1
2
3
Gsta2
Gsr G6pd
gene
exp
ress
ion
(fold
cha
nge)
gene
exp
ress
ion
(fold
cha
nge)
gene
exp
ress
ion
(fold
cha
nge)
gene
exp
ress
ion
(fold
cha
nge)
Hmox1
0
6
12
18
0
15
30
45Scl7a11
gene
exp
ress
ion
(fold
cha
nge)
# # ##
###
##
#
##
##
#
#
#
1000 cont 1 10100(nmol/L)
1000 cont 1 10100(nmol/L)
1000
cont 1 10100(nmol/L)
1000 cont 1 10100(nmol/L)
1000 cont 1 10100(nmol/L)
1000
FP receptor agonist fluprostenol increased Nrf2-target gene expression in a dose-dependent manner. Neonatal rat cardiomyocytes were treated with vehicle (cont) or 1-100 nmol/L of the FP receptor agonist fluprostenol for 12 h (A), 1-100 nmol/L BW245c, a DP1 receptor agonist for 12 h (B), or 1-1,000 nmol/L 15d-PGJ2 for 6 h (C). RNA was then extracted, and quantitative polymerase chain reaction performed to determine the mRNA expressions of Gclc, Gsr, G6pd, Hmox-1, Slc7a11, and Gsta2. Data are the mean ± SEM (n = 4). #P < 0.05 compared with vehicle-treated cardiomyocytes (unpaired Student’s t-test).
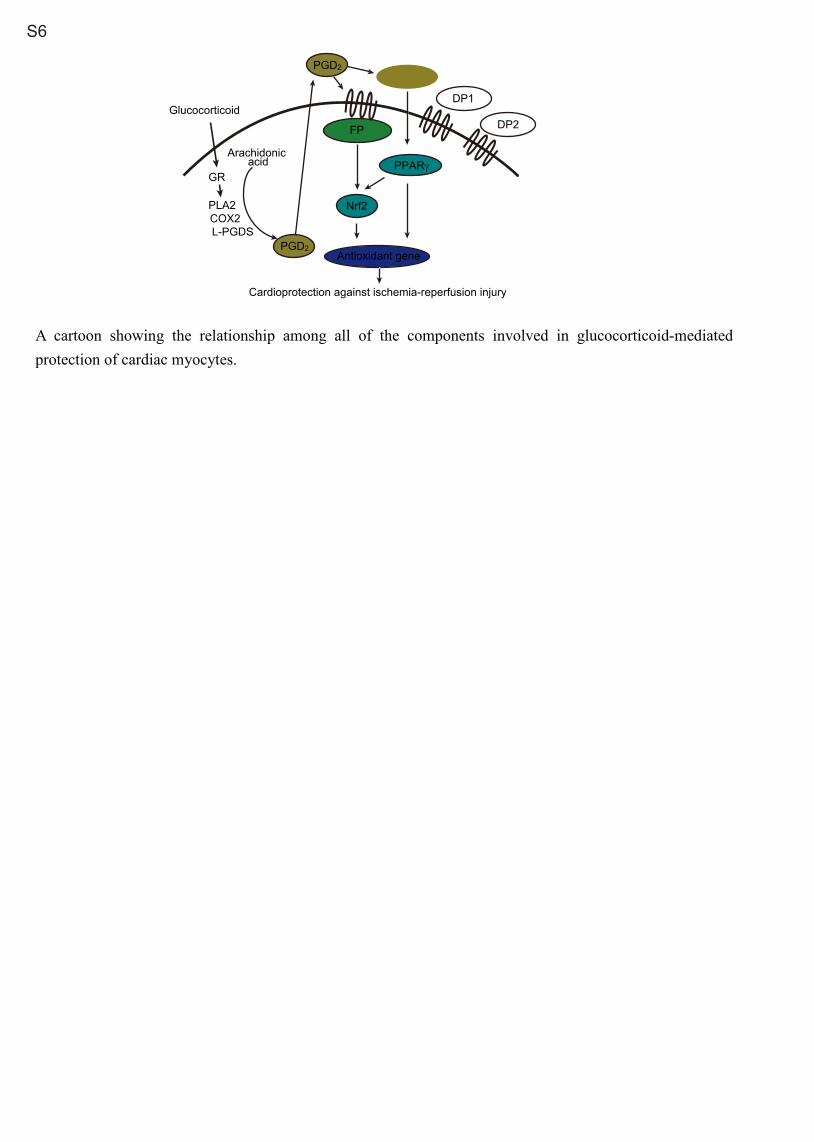
S6
Glucocorticoid
L-PGDS
PLA2COX2
15d-PGJ2
Arachidonic acid
Cardioprotection against ischemia-reperfusion injury
GR
PGD2
FP
PGD2
PPARγ
Nrf2
DP1
DP2
Antioxidant gene
A cartoon showing the relationship among all of the components involved in glucocorticoid-mediated protection of cardiac myocytes.





























