Embed Size (px)
Citation preview

2
Normally Occurring Environmentaland Behavioral Influences on Gene
ActivityFrom central dogma to probabilistic epigenesis
Gilbert Gottlieb
The central dogma of molecular biology holds that ‘‘information’’ flows from thegenes to the structure of the proteins that the genes bring about through theformula DNA ! RNA ! Protein. In this view, a set of master genes activatesthe DNA necessary to produce the appropriate proteins that the organism needsduring development. In contrast to this view, probabilistic epigenesis holds thatnecessarily there are signals from the internal and external environment thatactivate DNA to produce the appropriate proteins. To support this view, asubstantial body of evidence is reviewed showing that external environmentalinfluences on gene activation are normally occurring events in a large variety oforganisms, including humans. This demonstrates how genes and environmentswork together to produce functional organisms, thus extending the author’smodelof probabilistic epigenesis.
The new discipline of the genetics of behaviour, to judge by some recent books, iscaught in the dogmas of Mendelian genetics without regard to developments inmodemgenetics during the last ten years, and tomodern experimental approaches tothe genetic roots of behaviour. Books on the subject usually begin with an account ofthe principles of Mendelian genetics. The material on behaviour deals mainly withmutated animals and their observed changes in behaviour. That is exactly whatgenetic principles predict. If an important mutation should not be followed by achange in behaviour – then geneticists would have to worry about the validity ofthe principles.
Handbook of Developmental Science, Behavior, and GeneticsEdited by Kathryn E. Hood, Carolyn Tucker Halpern, Gary Greenberg, and Richard M. Lerner© 2010 Blackwell Publishing Ltd. ISBN: 978-1-405-18782-4

What these books fail to pay attention to is the trend in modern genetics which dealswith the activation of gene areas, with the influence of external factors on theactualization of gene-potentials and their biochemical correlates in behaviour . . ..I would venture to guess that, apart from the dogma, the main reason for thissilence is the fear of even the slightest suspicion that one might misinterpret suchfacts to mean that a Lamarckian mechanism were at work. (Hyd�en, 1969,pp. 114–115).
In the ensuing decades sinceHyd�enmade the above observation, things have notchanged very much. A virtual revolution has taken place in our knowledge ofenvironmental influences on gene expression that has not yet seeped into the socialsciences in general and the behavioral sciences in particular. Aside from the fearedmisinterpretation of Lamarckian mechanisms at work, there is an explicit dogma,formulated as such that does not permit environmental influences on gene activity:the ‘‘central dogma of molecular biology,’’ first enunciated by Crick in 1958.
Although the central dogma may seem quite remote from psychology, I thinkit lies behind some psychological and behavioral theories that emphasizethe sheerly endogenous construction of the nervous system and early behavior(e.g., Elman et al., 1996; Spelke&Newport, 1998) and the ‘‘innate foundation of thepsyche’’ (e.g., Tooby & Cosmides, 1990), independent of experience or functionalconsiderations: The essentially dichotomous view that genes and other endogen-ous factors construct part of the organism and environment determines otherfeatures of the organism. This article attempts to show how genes and environ-ments necessarily cooperate in the construction of organisms, specifically, howgenes require environmental and behavioral inputs to function appropriatelyduring the normal course of individual development.
Predetermined and Probabilistic Epigenesis
In earlier articles, I described two concepts of epigenetic development: predeter-mined and probabilistic epigenesis (Gottlieb, 1970, 1976). In these early formula-tions, the difference between the two points of view hinged largely on how theyconceived of the structure–function relationship. In predeterminism, it wasunidirectional (S ! F), whereas in probabilism it was bidirectional (S$ F).Subsequently, I (Gottlieb, 1976, p. 218; 1983, p. 13; 1991, p. 13) extended the uni-and bidirectionality to include genetic activity:
Predetermined EpigenesisUnidirectional Structure—Function Development
Genetic activity (DNA ! RNA ! Protein) !structural maturation ! function, activity, or experience
14 Gilbert Gottlieb

Probabilistic EpigenesisBidirectional Structure—Functional DevelopmentGenetic activity (DNA$RNA$ Protein)$
structural maturation$ function, activity, or experience
As it applies to the nervous system, structural maturation refers to neurophy-siological and neuroanatomical development, principally the structure andfunction of nerve cells and their synaptic interconnections. The unidirectionalstructure-function view assumes that genetic activity gives rise to structuralmaturation that then leads to function in a nonreciprocal fashion, whereas thebidirectional view holds that there are reciprocal influences among geneticactivity, structural maturation, and function. In the unidirectional view, theactivity of genes and the maturational process are pictured as relativelyencapsulated or insulated, so that they are uninfluenced by feedback from thematuration process or function, whereas the bidirectional view assumes thatgenetic activity and maturation are affected by function, activity, or experience.The bidirectional or probabilistic view applied to the usual unidirectionalformula calls for arrows going back to genetic activity to indicate feedbackserving as signals for the turning on and off of genetic activity. The usual view, asis discussed below in the section on the central dogma of molecular biology, callsfor genetic activity to be regulated by the genetic system itself in a strictly feed-forward manner. In this article, I (a) present the central dogma as a version ofpredetermined epigenesis, and (b) elaborate on the prior description of proba-bilistic epigenesis to bring it up to date on what is now known about the details ofthe bidirectional effects among genetic activity, structural maturation, neuraland behavioral function, and experience.
The Central Dogma
The central dogma asserts that ‘‘information’’ flows in only one direction from thegenes to the structure of the proteins that the genes bring about through theformula DNA ! RNA ! Protein. (Messenger RNA [mRNA] is the intermediaryin the process of protein synthesis. In the lingo ofmolecular biology,DNA ! RNAis called transcription and RNA ! Protein is called translation.) After retroviruses(RNA ! DNA) were discovered in the 1960s, Crick wrote a postscript to his 1958report inwhich he congratulated himself for not claiming that reverse transcriptionwas impossible: ‘‘In looking back I am struck not only by the brashness whichallowed us to venture powerful statements of a very general nature, but also by therather delicate discrimination used in selecting what statements to make’’(Crick, 1970, p. 562). He then went on to consider the central dogma formula,DNA ! RNA ! Protein, in much more explicit detail than in his earlier paper.
From Central Dogma to Probabilistic Epigenesis 15

In particular, he wrote, ‘‘These are the three [information] transfers which thecentral dogma postulates never occur:
Protein ! ProteinProtein ! DNA
Protein ! RNA’’ (p. 562).
I suppose if one is going to be brash about making proposals in largely uncharteredwaters, it stands to reason one might err, even given the otherwise acknowledgedinsight of the author regarding other scientific issues. In the present case, Crick waswrong in two of the three central-dogmatic postulates described above. Regardingprotein–protein interactions, it is now known that in neurodegenerative disorderssuch as Creutzfeldt–Jakob disease, prions (abnormally conformed proteins) cantransfer their abnormal conformation to other proteins (meaning Protein !Protein transfer of information), without the benefit of nucleic acid participation(RNA or DNA) (Telling et al., 1996). The strength of the dogma that nucleic acidsare required for ‘‘information transfer’’ is so compelling that some people believethere must be something like an RNA-transforming virus that brings about thechanged protein conformation, even though there is no evidence for such a virus(Chesebro, 1998; Grady, 1996).
Regarding Protein ! DNA transfer, there has long been recognized a class ofregulative proteins that bind toDNA, serving to activate or inhibit DNA expression(i.e., turning genes on or off; reviews in Davidson, 1986; Pritchard, 1986).
With respect to the third prohibited information transfer (Protein ! RNA),which would amount to reverse translation, to my knowledge, that phenomenonhas not yet been observed.
Any ambiguity about the controlling factors in gene expression in the centraldogma was removed in a later article by Crick, in which he specifically said thatthe genes of higher organisms are turned on and off by other genes (Crick, 1982,p. 515). Figure 2.1 shows the central dogma of molecular biology in the form of adiagram.
The Genome According to Central Dogma
The picture of the genome that emerges from the central dogma is (a) one ofencapsulation, setting the genome off from supragenetic influences, and (b) alargely feed-forward informational process in which the genes contain a blueprintormaster plan for the construction and determination of the organism. In this view,the genome is not seen as part of the development-physiological system of theorganism, responsive to signals from internal cellular sources such as the cytoplasmof the cell, cellular adhesion molecules (CAMs), or to extracellular influences such
16 Gilbert Gottlieb

as hormones, and certainly not to extraorganismic influcences such as stimuli orsignals from the external environment. Witness the well-known biologist ErnstMayr’s (1982) view ‘‘that the DNA of the genotype does not itself enter intothe developmental pathway but simply serves as a set of instructions’’ (p. 824).Mae-WanHo (1984) characterized this viewof the genes as the unmovedmovers ofdevelopment and themasters of the cellular slavemachinery of the organism. Ho’swork on the transgenerational effects of altered cytoplasmic influences seriouslyfaults Mayr’s view, as does the research reviewed by Jablonka and Lamb (1995).
Genes are conserved during evolution, therefore, some of the same genes arefound in many different species. What this has demonstrated is that there is not aninvariable association between the activity of a specific gene and the part of thebody in which it is active. One of the best demonstrations is the activity of the so-called Hox genes that are found in a number of species (Grenier, Garber, Warren,Whitington,&Carroll, 1997). As shown in Figure 2.2, in fruit flies theHox genes are
Figure 2.1. Central dogma of molecular biology. The right-going arrows represent thecentral dogma. The discovery of retroviruses (represented by the left-going arrow fromRNA to DNA) was not part of the dogma but, after the discovery, was said by Crick (1970)not to be prohibited in the original formulation of the dogma (Crick, 1958).
Figure 2.2. Darkened areas indicate different parts of the body in which the same Hoxgenes are active in three species: abdominal segment inDrosophila, virtually the entire bodyin centipede, and only the single hindmost segment inOnychophora. From ‘‘Evolution of theEntire Arthropod Hox Gene Set Predated the Origin and Radiation of the Onychoran/Arthropod Clade,’’ by J. K. Grenier, T. L. Garber, R. Warren, P. M. Whitington, andS. Carrol, 1997, Current Biology, 7, p. 552. Copyright 1997 by Jennifer Grenier. Adapted withpermission.
From Central Dogma to Probabilistic Epigenesis 17
Publisher's Note:Image not availablein the electronic edition
Publisher's Note:Image not availablein the electronic edition

active only in the abdominal segment of the body, whereas in centipedes thesame Hox genes are active in all segments of the body except the head. And, in arelated wormlike creature, Onychophora, the Hox genes are active only in a singlesegment of the organism in its hindmost region. Because these are nothomologous parts of these three species, this example demonstrates that thespecific developmental contributions of the same genes vary as a consequence ofthe developmental system in which they find themselves. Genes that play a rolein the abdominal segment of fruit flies are active in virtually all the bodilysegments of centipedes, but only in a single segment in Onychophora.
The main point of this article is to extend the normally occurring influences ongenetic activity to the external environment, thereby further demonstrating that agenome is not encapsulated and is in fact a part of an organism’s generaldevelopmental-physiological adaptation to environmental stresses and signals:Genes express themselves appropriately only in responding to internally andexternally generated stimulation. Further, in this view, although genes participatein themaking of protein, protein is also subject to other influences (Davidson, 1986;Pritchard, 1986), and protein must be further stimulated and elaborated to becomepart of the nervous system (or other systems) of the organism, so that genes operateat the lowest level of organismic organization and they do not, in and ofthemselves, produce finished traits or features of the organism.1
Thus, there is no correlation between genome size and the structural complexityof organisms (reviewed in Gottlieb, 1992, pp. 154–157), nor is there a correlationbetween numbers of genes and numbers of neurons in the brains of a variety oforganisms (see Table 2.1). The organism is a product of epigenetic development,which includes the genes as well as many other supragenetic influences. Sincethis latter point has been the subject of numerous contributions (reviewed inGottlieb, 1992, 1997), I shall not deal with it further here, but, rather restrict thisarticle to documenting that the activity of genes is regulated the same way as the
Table 2.1. Approximate number of genes and neurons in the brains of organisms indifferent lineages
Lineage and organism Genes Neurons
ChordatesMus musculus 70,000 40 millionHomo sapiens 70,000 85 billion
NematodesCaenorhabdhitis elegans 14,000 302
ArthropodsDrosophila melanogaster 12,000 250,000
Note. The exact number of neurons in the brain of C. elegans is known to be 302. From ‘‘Evolution andModification of Brains and Sensory Systems,’’ by G. L. GaborMiklos, 1998, reprinted by permission ofDaedalus, Journal of the American Academy of Arts and Sciences, from the issue titled ‘‘The Brain,’’Spring 1998, Vol. 127, No. 2, p. 200.
18 Gilbert Gottlieb

rest of the organism; the activity of genes is called forth by signals from thenormally occurring external environment, as well as the internal environment(Nijhout, 1990; Pritchard, 1986). Although this fact is not well known in the socialand behavioral sciences, it is surprising to find that it is also not widely appreciatedin biology proper (Strohman, 1997). In biology, the external environment is seen asthe agent of natural selection in promoting evolution, not as a crucial feature ofindividual development (van der Weele, 1995). Many biologists subscribe to thenotion that ‘‘the genes are safely sequestered inside the nucleus of the cell and out ofreach of ordinary environmental effects’’ (Wills, 1989, p. 19).
Normally Occurring Environmental Influences on Gene Activity
As can be seen in Table 2.2, a number of different naturally occurring environ-mental signals can stimulate gene expression in a large variety of organisms fromnematodes to humans. The earliest demonstration of this regularly occurringphenomenon that I could find in intact organisms is in the work of H. Hyd�en(Hyd�en & Egyh�azi, 1962). In this rarely cited study, hungry rats had to learn totraverse a narrow rod from an elevated starting platform to an elevated feedingplatform–a veritable balancing act. The nuclear base ratios in their vestibular nervecells were then compared with an untrained control group and a control groupgiven passive vestibular stimulation. The RNA base ratios in the experimentalgroups differed from both control groups. There was no difference between thecontrol groups.
I think theHyd�en and Egyh�azi (1962) study is rarely cited because the results notonly do not fit into any existing paradigm, they also seem to raise the Lamarckianspectre mentioned by Hyd�en (1969) in the opening quotation.2 If that is the case,there is an elementary misunderstanding. First, environmental stimulation of geneactivity in the organ of balance does notmean the genes were necessarily altered inthe process or, second, if they were altered, there is no reason to assume that thealteration was passed on to the progeny, as would be required by the way Lamarckused the notion of the inheritance of acquired characters in his theory ofevolution.3 In the Hyd�en and Egyh�azi study, themost conservative and acceptableexplanation is that genes (DNA)were turned on in the experimental group in awaythat they were not turned on in the control groups, resulting in an alteration ofRNA base ratios in the experimental group.
To understand the findings summarized in Table 2.2, the nongeneticist willneed to recall that the sequence of amino acids in proteins is determined by thesequence of nucleotides in the gene that ‘‘codes’’ for it, operating through theintermediary of mRNA. So there are three levels of evidence of genetic activity inTable 2.2: protein expression or synthesis, mRNA activity, and genetic activityitself. A difference in number of brain cells as a consequence of environmental
From Central Dogma to Probabilistic Epigenesis 19
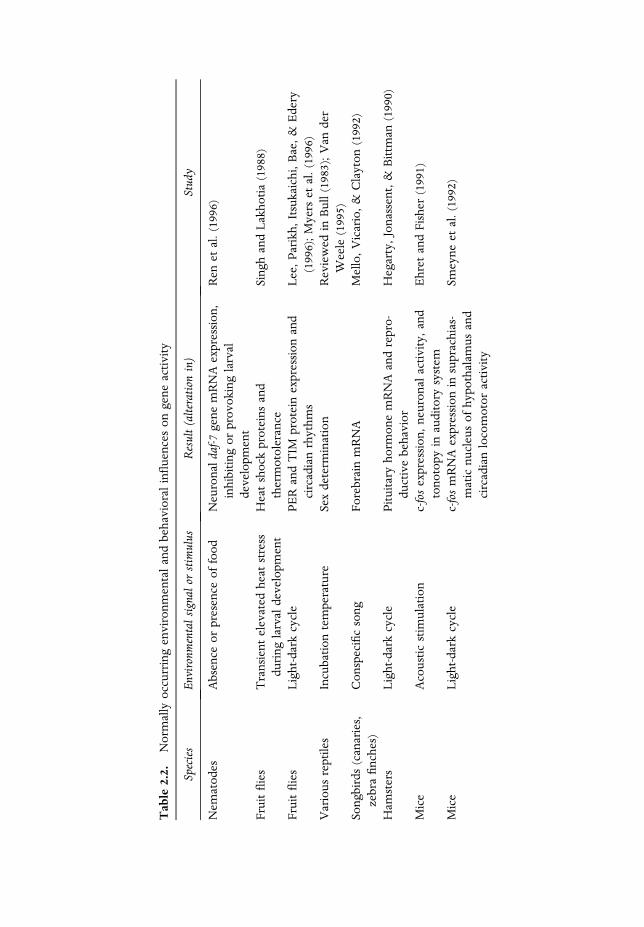
Table
2.2.
Normally
occurringenvironm
entalandbehavioralinfluences
ongene
activity
Species
Environm
entalsignalor
stimulus
Result(alteration
in)
Study
Nem
atodes
Absence
orpresence
offood
Neuronaldaf-7
gene
mRNAexpression
,inhibiting
orprovokinglarval
developm
ent
Ren
etal.(1996)
Fruitflies
Transient
elevated
heat
stress
during
larvaldevelopm
ent
Heatshockproteins
and
thermotolerance
SinghandLakho
tia(1988)
Fruitflies
Light-darkcycle
PERandTIM
proteinexpression
and
circadianrhythm
sLee,Parikh,
Itsukaichi,Bae,&
Edery
(1996);Myerset
al.(1996)
Various
reptiles
Incubation
temperature
Sexdeterm
ination
Reviewed
inBull(1983);Van
der
Weele(1995)
Songbirds(canaries,
zebrafin
ches)
Con
specificsong
ForebrainmRNA
Mello,Vicario,&
Clayton
(1992)
Ham
sters
Light-darkcycle
Pituitary
horm
onemRNAandrepro-
ductivebehavior
Hegarty,Jonassent,&
Bittm
an(1990)
Mice
Acousticstim
ulation
c-fosexpression
,neuron
alactivity,and
tono
topy
inauditory
system
Ehret
andFisher
(1991)
Mice
Light-darkcycle
c-fosmRNAexpression
insuprachias-
maticnu
cleusof
hypothalam
usand
circadianlocomotor
activity
Smeyne
etal.(1992)

Rats
Tactilestim
ulation
c-fosexpression
andnu
mberof
soma-
tosensorycorticalneuron
sMack&
Mack(1992)
Rats
Learningtask
involvingves-
tibu
larsystem
Nuclear
RNAbase
ratios
investibular
nervecells
Hyd�en
&Egyh�azi(1962)
Rats
Visualstim
ulation
RNAandproteinsynthesisin
visual
cortex
Rose(1967)
Rats
Environ
mentalcomplexity
Brain
RNAdiversity
Uphou
se&
Bon
ner(1975);review
inRosenzw
eig&
Bennett(1978)
Rats
Prenatalnu
trition
CerebralDNA(cerebralcellnu
mber)
Zam
enho
f&
vanMarthens(1978)
Rats
Infantile
handling;
separation
from
mother
Hypothalm
icmRNAsforcorticotropin-
releasingho
rmon
ethrougho
utlife
Meaneyet
al.(1996)
Cats
Visualstim
ulation
Visualcortex
RNAcomplexity
(diversity)
Grouse,Scheier,Letendre,&
Nelson
(1980)
Hum
ans
Academicexam
inations
taken
bymedicalstud
ents
(psychologicalstress)
Interleukin2receptor
mRNA(immun
esystem
respon
se)
Glaseret
al.(1990)
Note.mRNA¼messenger
RNA;PERandTIM
areproteins
arisingfrom
per(period)
andtim(timeless)gene
activity.

influences, as in theMack andMack (1992), andZamenhof and vanMarthens (1978)studies, means that DNA activity has been turned on by the environmentalstimulation. In the case of the more recent of these two studies, Mack and Mackwere able to measure fos activity as well as count the number of cortical cells,whereas in the earlier study, Zamenhof & van Marthens were able only to countthe number of cerebral cells as evidence of DNA activity.
As noted in Table 2.2, there are important neural and behavioral correlations togenetic activity, even though the activity of the genes is quite remote from theseeffects. The posttranslational expression of genes beyond the initial synthesis ofprotein involves the intervention of many factors before the end product of geneactivity is realized (review in Pritchard, 1986, p. 179).
The fact that normally occurring environmental events stimulate gene activityduring the usual course of development in a variety of organismsmeans that genesand genetic activity are part of the developmental-physiological system and do notreside outside of that system as some biologists and others have assumed on thebasis of the central dogma. The mechanisms by which environmental signals turnon genetic activity during the normal course of development is being activelyexplored in a number of laboratories. The interested reader is referred to thereviews by Campbell and Zimmermann (1982), Curran, Smeyne, Robertson,Vendrell, and Morgan (1994), Holliday (1990), Jablonka and Lamb (1995), Morganand Curran (1991), and Rosen and Greenberg (1994). Psychologists may beparticularly interested in the fact that environmentally provoked gene expressionis thought to be required for long-term memory (review in Goelet, Castellucci,Schacher, & Kandel, 1986).
From Central Dogma of Molecular Biologyto Probabilistic Epigenesis
Themain purpose of this article is to place genes and genetic activity firmlywithin adevelopmental-physiological framework, one in which genes not only affect eachother and mRNA, but are affected by activities at other levels of the system up toand including the external environment. This developmental system of bidirec-tional, coactional influences is captured schematically in Figure 2.3. In contrast tothe unidirectional and encapsulated genetic predeterminism of the central dogma,a probabilistic view of epigenesis holds that the sequence and outcomes ofdevelopment are probabilistically determined by the critical operation of variousendogenous and exogenous stimulative events (Gottlieb, 1970, p. 111; recentreview in Gottlieb, 1997). The probabilistic-epigenetic framework presented inFigure 2.3 is based not only onwhatwe now know aboutmechanisms of individualdevelopment at all levels of analysis, but the framework also derives from ourunderstanding of evolution and natural selection. Natural selection serves as a
22 Gilbert Gottlieb

filter and preserves reproductively successful phenotypes. These successful pheno-types are products of individual development and thus are a consequence of theadaptability of the organism to its developmental conditions. Therefore, naturalselection has preserved (favored) organisms that are adaptably responsive bothbehaviorally and physiologically to their developmental conditions.
Organisms with the same genes can develop very different phenotypes underdifferent ontogenetic conditions, as demonstrated by the two extreme variants of asingle parasiticwasp species shown in Figure 2.4, and by identical twins reared apartin the human species (Figure 2.5; these twinswere first described by Shields in 1962,pp. 43–44, 178–180, and later by Tanner, 1978, p. 119).4
Since the probabilistic-epigenetic view presented in Figure 2.3 does not portrayenough detail at the level of genetic activity, it is useful to flesh that out incomparison to the previously described central dogma of molecular biology.
BIDIRECTIONAL INFLUENCES
ENVIRONMENT
BEHAVIOR
NEURAL ACTIVITY
GENETIC ACTIVITY
(Physical, Social, Cultural)
Individual Development
Figure 2.3. Probabilistic-epigenetic framework: Depiction of the completely bidirectionaland coactional nature of genetic, neural, behavioral, and environmental influences over thecourse of individual development. From Individual Development and Evolution: The Genesis ofNovel Behavior (p. 186) by Gilbert Gottlieb, 1992, New York: Oxford University Press.Copyright 1992 by Oxford University Press, Inc. Reprinted with permission.
Butterfly Host Alder Host
Figure 2.4. Two very different morphological outcomes of development in the minuteparasiticwasp. The outcomes depended on the host (butterfly or alder fly) inwhich the eggswere laid. The insects are of the same species of parasitic wasp (Trichogramma semblidis).Adapted on the basis of Wigglesworth (1964).
From Central Dogma to Probabilistic Epigenesis 23

As shown in Figure 2.6, the original central dogma explicitly posited one-waytraffic from DNA ! RNA ! Protein and was silent about any other flows ofinformation (Crick, 1958). Later, after the discovery of retroviruses (RNA ! DNAinformation transfer), Crick (1970) did not claim to have predicted that phenom-enon, but, rather that the original formulation did not expressly forbid it. In thebottom of Figure 2.6, probabilistic epigenesis, being inherently bidirectional inthe horizontal and vertical levels (Figure 2.3), has information flowing not onlyfrom RNA ! DNA but between Protein$ Protein and DNA$DNA. The onlyrelationship that is not yet supported is Protein ! RNA, in the sense of reversetranslation (protein altering the structure of RNA), but there are other influences ofprotein on RNA activity (not its structure) that would support such a directionalflow. For example, a process known as phosphorylation can modify proteinssuch that they activate (or inactivate) other proteins (Protein ! Protein), which,when activated, trigger rapid association of mRNA (Protein ! RNA activity).When mRNAs are transcribed by DNA, they do not necessarily become imme-
Figure 2.5. Remarkable illustration of the enormous phenotypic variation that can resultwhen monozygotic (single egg) identical twins are reared apart in very different familyenvironments from birth. From Foetus Into Man (p. 120) by J. M. Tanner, 1978, Cambridge,MA: Harvard University Press. Copyright 1978 by Harvard University Press. Reprintedwith permission
24 Gilbert Gottlieb
Publisher's Note:Image not availablein the electronic edition

diately active but require a further signal to do so. The consequences of phosphory-lation could provide that signal (Protein ! Protein ! mRNAactivity ! Protein),A process like this appears to be involved in the expression of ‘‘fragile X mentalretardation protein’’ under normal conditions and proves disastrous to neuraland psychological development when it does not occur (Weiler et al., 1997).5
An excellent overview of the various roles of phosphorylation in the nervous systemis provided by Hyman and Nestler (1993, Chapter 4).
Amplifying the left side of the bottom of Figure 2.6, it is known that geneexpression is affected by events in the cytoplasm of the cell, which is the immediateenvironment of the nucleus andmitochondria of the cell whereinDNA resides, andby hormones that enter the cell and its nucleus. This feed-downward effect can bevisualized thusly:
Genetic Activity According To Central Dogma
DNA DNA ProteinRNA
?
DNA DNA ProteinRNA
?
Genetic Activity According To Probabilistic Epigenesis
DNA
DNA
RNA
RNA
Protein
Protein
Internal andExternal
Environment
Figure 2.6. Different views of influences on genetic activity in the central dogma andprobabilistic epigenesis. The filled arrows indicate documented sources of influence,whereas the open arrow from Protein back to RNA remains a theoretical possibility inprobabilistic epigenesis and is prohibited in the central dogma (as are Protein$ Proteininfluences). Protein ! Protein influences occur (a) when prions transfer their abnormalconformation to other proteins and (b) when, during normal development, proteinsactivate or inactivate other proteins as in the phosphorylation example described in text.The filled arrows from Protein to RNA represent the activation of mRNA by protein as aconsequence of phosphorylation, for example. DNA$DNA influences are termed‘‘epistatic,’’ referring to the modification of gene expression depending on the geneticbackground in which they are located. In the central dogma, genetic activity is dictatedsolely by genes (DNA ! DNA), whereas in probabilistic epigenesis internal and externalenvironmental events activate genetic expression through proteins (Protein ! DNA),hormones, and other influences. To keep the diagram manageable, the fact that behaviorand the external environment exert their effects on DNA through internal mediators(proteins, hormones, etc.) is not shown; nor is it shown that the protein products of somegenes regulate the expression of other genes. (See text for further discussion.)
From Central Dogma to Probabilistic Epigenesis 25

Gene expression influenced by
external environment behavior/psychological function/experience
cytoplasm
hormones
According to this view, different proteins are formed depending on the particularfactors influencing gene expression. Concerning the effect of psychological func-tioning on gene expression, we have the evidence in Table 2.2 of heightenedinterleukin 2 receptor mRNA, an immune system response, in medical studentstaking academic examinations (Glaser et al., 1990). More recently, in an elegantstudy that traverses all levels from psychological functioning to neural activity toneural structure to gene expression, Cirelli, Pompeiano, and Tononi (1996) showedthat genetic activity in certain areas of the brain is higher during waking than insleeping in rats. In that study, the stimulation of gene expression was influenced bythe hormone norepinephrine flowing from locus coeruleus neurons that fire at verylow levels during sleep and at high levels during waking and when triggered bysalient environmental events. Norepinephrine modifies neural activity and excit-ability, as well as the expression of certain genes. So, in this case, we have evidencefor the interconnectedness of events relating the external environment andpsychological functioning to genetic expression by a specifiable hormone emanatingfrom the activity of a specific neural structurewhose functioningwaxes andwanes inrelation to the psychological state of the organism.
Importance of Behavioral and Neural Activityin Determining Gene Expression, Anatomical Structure,
and Physiological Function
Many, if not all, of the normally occurring environmental influences on geneticactivity summarized in Table 2.2 involve behavioral and neural mediation. In thespirit of this article, I want to emphasize the contribution of events above thegenetic level (thewhole organism and environmental context) byway of redressingthe balance to the way many think about the overriding importance of molecularbiology. The earliest synaptic connections in the embryonic and fetal nervoussystem are created by spontaneous activity of nerve cells (reviews in Corner, 1994;Katz & Shatz, 1996). This early, ‘‘exuberant’’ phase produces a very large arrayof circuits that are then pared down by the organism’s encounters with its prenataland postnatal environments. In the absence of behavioral and neural activity(e.g., experimentally induced paralysis), cells do not die, and circuits do not becomepruned in an adaptive way that fits the organism to the demands of its physical,
26 Gilbert Gottlieb

social, and cultural environments (Pittman & Oppenheim, 1979). A recent reviewof the development and evolution of brain plasticity may be found in Black andGreenough (1998).
Sometimes one reads the perfectly reasonable-sounding suggestion that,although genes do not make anatomical, physiological, or behavioral traits, thegenes constrain the outer limits of variation in such traits. It is, of course, thedevelopmental system, of which the genes are a part (Figure 2.3), and not solelythe genes, that constrains development. It is not possible to predict in advancewhatthe outcome of development will be when the developing organism is faced withnovel environmental or behavioral challenges never before faced by a species orstrain of animal. This has been known since 1909 when Woltereck did the firstexperiments that resulted in the open-ended concept of the norm of reaction, anidea that has beenmisunderstood by some behavior geneticists who think of genesas setting up a too-narrow range of reaction (reviews in Gottlieb, 1995; Piatt &Sanislow, 1988).
A very striking example of the role of novel behavior bringing about an entirelynew anatomical structure can be seen in Slijper’s (1942) goat in Figure 2.7. Thisanimal was bom with undeveloped forelimbs and adopted a kangaroolike form oflocomotion. As a result, its skeleton and musculature became modified, with apelvis and lower spinal column like that of a biped instead of a quadruped(Figure 2.7). Thus, although there can be no doubt that genes and other factorsplace constraints on development, Slijper’s goat shows that it is not possible toknow the limits of these constraints in advance, even though it might seem quitereasonable to assume, in advance of empirical inquiry, that a quadruped is notcapable of bipedality. Although an open-ended, empirically based norm of reaction
(a) (b)
Figure 2.7. Modification of pelvic and spinal anatomy consequent to bipedalism. Thefigure shows (a) the pelvis and lower spine of a normal quadrupedal goat, and (b) the pelvisand lower spine of a goat born without forelimbs and that adopted a form of locomotionsimilar to a kangaroo. From Foundation of Developmental Genetics (p. 310) by D. J.Pritchard, 1986, London and Philadelphia: Taylor & Francis. Copyright 1986 by DorianPritchard. Reprinted with permission.
From Central Dogma to Probabilistic Epigenesis 27

can accommodate Slijper’s goat, a narrowly constrained, rationally based range ofreaction cannot, no matter how reasonable it seems. It may very well be that allquadruped species cannot adapt bipedally, but we cannot know that withoutperturbing the developmental system.
Summary and Conclusions
It is tempting to show the nice link between probabilistic epigenesis and anepigenetic behavioral theory of evolution; however, that topic has been reviewedin depth in several recent publications (Gottlieb, 1992, 1997), so I will forego thattemptation here in favor of sticking to the main point of this article. The centraldogma lies behind the persistent trend in biology and psychology to view genes andenvironments as making identifiably separate contributions to the phenotypicoutcomes of development. Quantitative behavior genetics is based on thiserroneous assumption. It is erroneous because animal experiments have shownagain and again that it is not possible to identify the genetic and environmentalcomponents of any phenotype, whether behavioral, anatomical, or physiological(extensive review in Wahlsten & Gottlieb, 1997).6 Although genes no doubt playa constraining role in development, the actual limits of these constraints arequite wide and, most important, cannot be specified in advance of experimentalmanipulation or accidents of nature as documented in Figures 2.3, 2.4 and 2.7. (Theprenatal environment also plays a constraining role that cannot be known inadvance of experimental ormanipulative inquiry; Gottlieb, 1971, 1997.) There is nodoubt that not only genes and environments constrain development at all levels ofthe system (Figure 2.3).
The theoretical crux of this article is that the internal and external environ-ments supply the necessary signals to the genes that instigate the eventualproduction of the requisite proteins under normal as well as unusual conditions ofdevelopment. There is no genetic master plan or blueprint that is self-actualizedduring the course of development, as was assumed by the central dogma.Without doubt, there are unusual developmental conditions to which genescannot respond adaptably, but the range of possible adaptable genetic responsesto strange environmental conditions is truly astounding, as when bird oralepithelial cells mixed with mouse oral mesenchyme cells resulted in the produc-tion of a fully enameledmolar tooth (Kollar& Fisher, 1980). The phrase ‘‘scarce asa hen’s tooth’’ is based on the fact that bird oral epithelial cells never produceteeth when in conjunction with bird oral mesenchyme cells, as would be the caseunder normal conditions of development. If this finding is ‘‘clean’’ (nomouse oralepithelial cells accidentally contaminating the mix), it involves the appropriatereactivation of a genetic combination that had been latent for 80 million yearswhen birds’ last toothed ancestor existed (Pritchard, 1986, pp. 308–309). Also, the
28 Gilbert Gottlieb

finding that a crucial nutriment experimentally deleted from the environment ofbacterial cells could lead to the production of that nutriment by a genetic recombina-tion (adaptive mutation) caused a storm of disbelief in the biological community untilit was shown that there was indeed a molecular basis for this ‘‘theoreticallyimpossible’’ finding (Harris, Longerich, & Rosenberg, 1994; Thaler, 1994).
It will be interesting to see how probabilistic epigenesis becomes modified inthe ensuing years as more information accrues through the necessarily interdisci-plinary and multidisciplinary efforts of future researchers. The contrastingideas of predetermined and probabilistic epigenesis were first put forward inGottlieb (1970). Although the central dogma as depicted in Figure 2.6 is consistentwith the formulation of predetermined epigenesis, it is too much to claim that thecontrasting formulation of probabilistic epigenesis in 1970 predicted all the detailsof the relationships in the lower half of Figure 2.6. One can only say that thoserelationships are consistent with the bidirectional influences stated in the prob-abilistic formula Genetic Activity$ Structure$ Function presented in Gottlieb(1976, p. 218) and elaborated in Gottlieb (1991, see especially Appendix, p. 13).As I have described in detail elsewhere (e.g., Gottlieb, 1992, 1997), the formulationof probabilistic epigenesis was built on the writings of Kuo (1976), Lehrman (1970),Montagu (1977), and Schneirla (1960).
Finally, in response to a concern raised by colleagues who have read this articlein manuscript form, I do hope that the emphasis on normally occurring environ-mental influences on gene activity does not raise the spectre of a new, subtle formof ‘‘environmentalism.’’ If I were to say organisms are often adaptably responsiveto their environments, I don’t think thatwould labelme as an environmentalist. So,by calling attention to genes being adaptably responsive to their internal andexternal environments, I am not being an environmentalist, but I am merelyincluding genetic activity within the probabilistic-epigenetic framework thatcharacterizes the organism and all of its constituent parts (Figure 2.3). Theprobabilistic-epigenetic view follows the open-systems view of developmentchampioned by the biologists Ludwig von Bertalanffy (1933/1962), Paul Weiss(1939/1969), and Sewall Wright (1968). Their writings were based on a highlyinteractive conception of embryology, and the central dogma simply overlookedthis tradition of biological theorizing, resulting in an encapsulated formulation ofgenetic activity at odds with the facts of embryological development. (The currentreductionist theoretical stance of molecular biology continues to disregard epige-netic considerations; Strohman, 1997.) Building on the insights of von Bertalanffy,Weiss, and Wright, the probabilistic-epigenetic view details the cooperativeworkings of the embryological open-systems view at the genetic and neurallevels, prenatal and postnatal behavior, and the external environment. This viewfleshes out at the prenatal and intraorganismic levels of analysis various otherapproaches in developmental psychology: ecological (Bronfenbrenner, 1979),transactional (Sameroff, 1983), contextual (Lerner & Kaufman, 1985), interactionalorholistic (Johnston, 1987;Magnusson, 1988), individual-sociological (Valsiner, 1997),
From Central Dogma to Probabilistic Epigenesis 29

structural-behavioral (Horowitz, 1987), dynamic systems (Thelen&Smith, 1994),and, most globally speaking, interdisciplinary developmental science (Cairns,Elder, & Costello, 1996).
Acknowledgments
Research and scholarly activities for this article were supported in part by NationalInstitute ofMental HealthGrantMH-52429. I have benefited fromdiscussions withJaan Valsiner and other members of the Carolina Consortium of Human Devel-opment, as well as from the comments of Dorian Pritchard, Lynda Uphouse,Richard C. Strohman, KathrynHood, andNora LeeWillis Gottlieb on earlier draftsof this article. James Black and William Greenough made very helpful substantivesuggestions for which I am most grateful. Ramona Rodriguiz provided generousbibliographic assistance.
Notes
This article first appeared in Psychological Review, (1998) 105, 792–802. Reprinted withpermission.
1. Among the most scholarly early critiques to make this point was that of G. Stent (1981),who wrote:
For the viewpoint that the structure and function of the nervous system of ananimal is specified by its genes provides too narrow a context for actuallyunderstanding developmental processes and thus sets a goal for the geneticapproach that is unlikely to be reached. Here too ‘‘narrow’’ is not to mean that abelief in genetic specification of the nervous system necessarily implies a lack ofawareness that in development there occurs an interaction between genes andenvironment, a fact of which all practitioners of the genetic approach arecertainly aware. Rather, ‘‘too narrow’’ means that the role of the genes, which,thanks to the achievements of molecular biology, we now know to be thespecification of the primary structure of protein molecules, is at too manyremoves from the processes that actually ‘‘build nerve cells and specify neuralcircuits which underlie behavior’’ to provide an appropriate conceptual frame-work for posing the developmental questions that need to be answered.(pp. 186–187)
Stent’s critique was taken a step further by Nijhout (1990), who wrote in a general wayabout the importance of interactions, above the genetic level, in the internal environ-ment of the organism to bring about growth and differentiation (morphogenesis).
30 Gilbert Gottlieb

Nijhout’s point was that ‘‘genes do not . . . ‘cause’ or ‘control’ morphogenesis; theyenable it to take place’’ (p. 443). Evenmore pertinent to the theme of this article, Nijhoutwrote that the geneswhose products are necessary during development are activated bystimuli that arise from the cellular and chemical processes of development. Thus thenetwork or pattern of gene activation does not constitute a program, it is both theconsequence of, and contributor to, development. (pp. 443)
In this article, I extend this point of view to the external environment.2. Due to the great advances in molecular techniques since 1962, some present-day
researchersmay question the results of Hyd�en and Egyh�azi onmethodological grounds.3. Although it is not a popular idea, and it is a separate question, genes can be altered by
internal (reverse transcription, for example) and external events during developmentand, under certain conditions, the activities of these altered genes can persist acrossgenerations (Campbell & Perkins, 1983; Campbell & Zimmermann, 1982; Holli-day, 1990; Jablonka & Lamb, 1995).
4. This great amount of phenotypic variation observed in identical twins (sharing thesame genotype) coordinates well with the enormous degree of phenotypic variation inthe human species, in which there is in fact only a very small degree of individual geneticvariation at the level of DNA. DNA is composed of two base pairs of nucleotides. Thereis such a small amount of variation in these base pairs in the human population that anytwo individuals selected at random from anywhere on earthwould exhibit differences inonly three or four base pairs out of 1,000 base pairs (i.e., .3% or .4%!; Cann, 1988;Merriwether et al., 1991).
5. The label of ‘‘fragile Xmental retardation protein’’ makes it sound as if there is a gene (orgenes) that produces a protein that predisposes tomental retardation, whereas, in actualfact, it is this protein that is absent from the brain of fragile Xmental retardates, and thusrepresents a failure of gene (or mRNA) expression rather than a positive geneticcontribution tomental retardation. The same is likely true for other ‘‘genetic’’ disorders,whether mental or physical. Such disorders most often represent biochemical defi-ciencies of one sort or another due to the lack of expression of the requisite genes andmRNAs to produce the appropriate proteins necessary for normal development. Thus,the search for ‘‘candidate genes’’ in psychiatric or other disorders is most often a searchfor genes that are not being expressed, not for genes that are being expressed and causingthe disorder. So-called ‘‘cystic fibrosis genes’’ and ‘‘manic-depression genes,’’ amongothers, are in this category. The instances that I know of in which the presence of genescauses a problem are Edward’s syndrome and Trisomy 21 (Down’s syndrome), whereinthe presence of an extra, otherwise normal, Chromosomes 18 and 21, respectively, causeproblems because the genetic system is adapted for two, not three, chromosomes ateach location. In some cases, it is of course possible that the expression ofmutated genescan be involved in a disorder, but, inmy opinion, it ismost often the lack of expression ofnormal genes that is the culprit.
6. This is not the same as saying one cannot pinpoint the participation of specific genes andspecific environments in contributing to phenotypic outcomes. However, because genesand environments always collaborate in the production of any phenotype, it is notpossible to say that a certain component of the phenotype was caused exclusively bygenes (independent of environmental considerations) and that some other componentwas caused exclusively by environment (independent of a genetic contribution).
From Central Dogma to Probabilistic Epigenesis 31

An understanding of developmental phenomena demands a relational or coactiveconcept of causality as opposed to singular causes acting in supposed isolation(discussed at length in Gottlieb, 1991, 1997). Overton (1998) has presented a historicaloverview on the topic of dualistic conceptions of causality versus the more recentrelational or coactive concept of causality. Further, with respect to the erroneousseparation of hereditary and environmental contributions to the phenotype byquantitative behavior geneticists, Wahlsten (1990) has shown that the absence ofheredity–environment interaction is a statistical artifact stemming from the insuffi-cient power of the analysis of variance to detect such interactions, not the empiricalabsence of such gene–environment interactions.
References
Black, J. E., & Greenough, W. T. (1998). Developmental approaches to the memoryprocess. In: J. L. Martinez&R. P. Kesner (Eds.), Learning and memory: A biological view(3rd ed., pp. 55–88). New York: Academic Press.
Bronfenbrenner, U. (1979). The ecology of human development: Experiments by nature anddesign. Harvard University Press,. Cambridge, MA.
Bull, J. J. (1983). Evolution of sex determining mechanisms. Menlo Park, CA: Benjamin/Cummings.
Cairns, R. B., Elder, G. H., & Costello, E. J. (Eds.). (1996). Developmental science. New York:Cambridge University Press.
Campbell, J. H., & Perkins, P. (1983). Transgenerational effects of drug and hormonaltreatments in mammals: A review of observations and ideas. Progress in BrainResearch, 73, 535–553.
Campbell, J. H., & Zimmermann, E. G. (1982). Automodulation of genes: A proposedmechanism for persisting effects of drugs and hormones inmammals.NeurobehavioralToxicology and Teratology, 4, 435–439.
Cann, R. L. (1988). DNA and human origins. Annual Review of Anthropology, 17, 127–143.
Chesebro, B. (1998). BSE and prions: Uncertainties about the agent. Science, 279, 42–43.
Cirelli, C., Pompeiano, M., & Tononi, G. (1996). Neuronal gene expression in the wakingstate: A role for locus coeruleus. Science, 274, 1211–1215.
Corner, M. A. (1994). Reciprocity of structure-function relations in developing neuralnetworks: The odyssey of a self-organizing brain through research fads, fallacies, andprospects. Progress in Brain Research, 102, 3–31.
Crick, F. H. C. (1958). On protein synthesis. Symposia of the Society for Experimental Biology,No. 12. The biological replication of macro-molecules (pp. 138–163). Cambridge, England:Cambridge University Press.
Crick, F. (1970). Central dogma of molecular biology. Nature, 227, 561–563.
Crick, F. (1982). DNA today. Perspectives in Biology and Medicine, 25, 512–517.
Curran, T., Smeyne, R., Robertson, L., Vendrell, M., & Morgan, J. I. (1994). FoslacZexpression: A transgenic approach to gene activation in the brain. In S. Zalcman,
32 Gilbert Gottlieb

R. Scheller, & R. Tsien (Eds.), Molecular neurobiology: Proceedings of the SecondNIMH Conference (pp. 165–172). Rockville, MD: National Institute of MentalHealth.
Davidson, E. H. (1986). Gene activity in early development. Orlando, FL: Academic Press.
Ehret, G., & Fisher, R. (1991). Neuronal activity and tonotopy in the auditory systemvisualized by c-fos gene expression. Brain Research, 567, 350–354.
Elman, J. L., Bates, E. A., Johnson, M. H., Karmiloff-Smith, A., Parisi, D., & Plunkett, K.(1996). Rethinking innateness: A connectionist perspective on development. Cambridge,MA: MIT Press.
Glaser, R., Kennedy, S., Lafuse, W. P., Bonneau, R. H., Speicher, C., Hillhouse, J., &Kiecolt-Glaser, J. K. (1990). Psychological-stress-induced modulation of interleukin 2receptor gene expression and interleukin 2 production in peripheral bloodleukocytes. Archives of General Psychiatry, 47, 707–712.
Goelet, P., Castelluci, V. F., Schacher, S., &Kandel, E. R. (1986). The long and short of long-term memory—A molecular framework. Nature, 322, 419–422.
Gottlieb, G. (1970). Conceptions of prenatal behavior. In L. R. Aronson, E. Tobach, D. S.Lehrman, & J. S. Rosenblatt (Eds.), Development and evolution of behavior: Essays inmemory of T. C. Schneirla (pp. 111–137). San Francisco: W. H. Freeman.
Gottlieb, G. (1971). Development of species identification in birds: An inquiry into the prenataldeterminants of perception. Chicago: University of Chicago Press.
Gottlieb, G. (1976). Conceptions of prenatal development: Behavioral embryology.Psychological Review, 83, 215–234.
Gottlieb, G. (1983). The psychobiological approach to developmental issues. In M. M. Haith &J. J. Campos (Eds.), Handbook of child psychology: Infancy and developmentalpsychobiology ( 4th ed., Vol. 2; pp. 1–26). New York: Wiley.
Gottlieb, G. (1991). Experiential canalization of behavioral development: Theory.Developmental Psychology, 27, 4–13.
Gottlieb, G. (1992). Individual development and evolution: The genesis of novel behavior.New York: Oxford University Press.
Gottlieb, G. (1995). Some conceptual deficiencies in ‘developmental’ behavior genetics.Human Development, 38, 131–141.
Gottlieb, G. (1997). Synthesizing nature–nurture: Prenatal roots of instinctive behavior.Mahwah, NJ: Erlbaum.
Grady, D. (1996). Ironing out the wrinkles in the prion strain problem. Science,274, 2010.
Grenier, J. K., Garber, T. L., Warren R., Whitington, P. M., & Carroll, S. (1997). Evolutionof the entire arthropod Hox gene set predated the origin and radiation of theonychophoran/arthropod clade. Current Biology, 7, 547–553.
Grouse, L. D., Schrier, B. K., Letendre, C. H., & Nelson, P. G. (1980). RNA sequencecomplexity in central nervous system development and plasticity. Current Topics inDevelopmental Biology, 16, 381–397.
Harris, R. S., Longerich, S., & Rosenberg, S. M. (1994). Recombination in adaptivemutation. Science, 264, 258–260.
From Central Dogma to Probabilistic Epigenesis 33

Hegarty, C.M., Jonassent, J. A., & Bittman, E. L. (1990). Pituitary hormone gene expressionin male golden hamsters: Interactions between photoperiod and testosterone.Journal of Neuroendocrinology, 2, 567–573.
Ho,M. -W. (1984). Environment and heredity in development and evolution. InM.-W.Ho,& P. T. Saunders (Eds.), Beyond neo-Darwinism: An introduction to the new evolutionaryparadigm (pp. 267–289). San Diego, CA: Academic Press.
Holliday, R. (1990). Mechanisms for the control of gene activity during development.Biological Reviews, 65, 431–471.
Horowitz, F. D. (1987). Exploring developmental theories: Toward a structural/behavioral modelof development. Hillsdale, NJ: Erlbaum.
Hyd�en,H. (1969). Afterthoughts. In A. Koestler & J. R. Smythies (Eds.), Beyond reductionism:New perspectives in the life sciences (pp. 114–117). London: Hutchinson.
Hyd�en, H., & Egyh�azi, E. (1962). Nuclear RNA changes of nerve cells during a learningexperiment in rats. Proceedings of the National Academy of Sciences, USA, 48, 1366–1373.
Hyman, S. E., & Nestler, E. J. (1993). The molecular foundations of psychiatry. Washington,DC: American Psychiatric Press.
Jablonka, E., & Lamb, M. J. (1995). Epigenetic inheritance and evolution: The Lamarckiandimension. Oxford, England: Oxford University Press.
Johnston, T. D. (1987). The persistence of dichotomies in the study of behavioraldevelopment. Developmental Review, 7, 149–182.
Katz, L. C., & Shatz, C. J. (1996). Synaptic activity and the construction of cortical circuits.Science, 274, 1133–1138.
Kollar, E. J., & Fisher, C. (1980). Tooth induction in chick epithelium: Expression ofquiescent genes for enamel synthesis. Science, 207, 993–995.
Kuo, Z. -Y. (1976). The dynamics of behavior development (enlarged ed). New York: Plenum.
Lee, C., Parikh, V., Itsukaichi, T., Bae, K., & Edery, I. (1996). Resetting the Drosophilaclock by photic regulation of PER and a PER-TIM complex. Science, 271,1740–1744.
Lehrman, D. S. (1970). Semantic and conceptual issues in the nature-nurture problem. InL. R. Aronson, E. Tobach, D. S. Lehrman, & J. S. Rosenblatt (Eds,), Developmentand evolution of behavior: Essays in memory of T. C. Schneirla (pp. 17–52). San Francisco:W. H. Freeman.
Lerner, R. M., & Kaufman, M. B. (1985). The concept of development in contextualism.Developmental Review, 5, 309–333.
Mack, K. J., &Mack, P. A. (1992). Induction of transcription factors in somatosensory cortexafter tactile stimulation. Molecular Brain Research, 12, 141–147.
Magnusson, D. (1988). Individual development from an interactional perspective: A longitudinalstudy. Hillsdale, NJ: Erlbaum.
Mayr, E. (1982). The growth of biological thought. Harvard University Press: Cambridge, MA.
Meaney, M. J., Diorio, J., Francis, D., Widdowson, J., LaPlante, P., Caldji, C., Sharma, S.,Secki, J. P., & Plotsky, P. M. (1996). Early environmental regulation of forebrainglucocorticoid receptor gene expression: Implications for adrenocortical responses tostress. Developmental Neuroscience, 18, 49–72.
34 Gilbert Gottlieb

Mello, C. V., Vicario, D. S., & Clayton, D. F. (1992). Song presentation induces geneexpression in the songbird forebrain. Proceedings of the National Academy of Sciences,USA, 89, 6818–6822.
Merriwether, D. A., Clark, A. G., Ballinger, S. W., Schurr, T. G., Soodyall, H., Jenkins, T.,Sherry, S. T., &Wallace, D. C. (1991). The structure of human mitochondrial DNAvariation. Journal of Molecular Evolution, 33, 543–555.
Miklos, G. L..Gabor (in press). Evolution and modification of brains and sensory systems.Daedalus.
Montagu, A. (1977). Sociogenic brain damage. In S. Arieti & G. Chrzanowski (Eds.),New dimensions in psychiatry: A world view (Vol. 2, pp. 4–25). New York: Wiley.
Morgan, J. I., & Curran, T. (1991). Stimulus-transcription coupling in the nervous system:Involvement of the inducible proto-oncogenes fos and jun. Annual Review ofNeurosciences, 14, 421–451.
Myers, M. P., Wager-Smith, K., Rothenfluh-Hilfiker, A., & Young, M. W. (1996). Light-induced degradation of TIMELESS and entertainment of the Drosophila circadianclock. Science, 271, 1736–1740.
Nijhout, H. F. (1990). Metaphors and the role of genes in development. BioEssays, 12,441–446.
Overton, W. F. (1998). Developmental psychology: Philosophy, concepts, andmethodology. In R. M. Lerner (Ed.), Handbook of child psychology: Theoreticalmodels of human development ( 5th ed., vol. 1, pp. 107–189). New York: Wiley.
Pittman, R., & Oppenheim, R. W. (1979). Cell death of motoneurons in the chick embryospinal cord. IV: Evidence that a functional neuromuscular interaction is involved inthe regulation of naturally occurring cell death and the stabilization of synapses,Journal of Comparative Neurology, 187, 425–446.
Platt, S. A., & Sanislow, C. A. (1988). Norm-of-reaction: Definition andmisinterpretation ofanimal research. Journal of Comparative Psychology, 102, 254–261.
Pritchard, D. J. (1986). Foundations of developmental genetics. London and Philadelphia:Taylor and Francis.
Ren, P., Lin, C.-S., Johnson, R., Albert, P. S., Pilgrim, D., & Riddle, D. L. (1996). Control ofC. elegans larval development by neuronal expression of a TGFb homolog. Science,274, 1389–1391.
Rose, S. P. R. (1967). Changes in visual cortex on first exposure of rats to light: Effect onincorporation of tritiated lysine into protein. Nature, 215, 253–255.
Rosen, L. B.,&Greenberg,M.E. (1994). Regulationof c-fos andother immediate-early genesin PC-12 cells as amodel for studying specificity in neuronal signaling. In S. Zalcman,R. Scheller, & R. Tsein (Eds.), Molecular neurobiology: Proceedings of the Second NIMHConference (pp. 203–216). Rockville, MD: National Institute of Mental Health.
Rosensweig, M. R., & Bennett, E. L. (1978). Experiential influences on brain anatomy andbrain chemistry in rodents. In G. Gottleib (Ed.), Early influences (pp. 289–327).New York: Academic Press.
Sameroff, A. J. (1983). Developmental systems: Contexts and evolution. InW. Kessen (Ed.),Handbook of child psychology: History, theory, and methods. (4th ed., Vol. 1, pp. 237–294).New York: Wiley.
From Central Dogma to Probabilistic Epigenesis 35

Schneirla, T. C. (1960). Instinctive behavior, maturation—Experience and development.In B. Kaplan & S.Wapner (Eds.), Perspectives in psychological theory—Essays in honor ofHeinz Werner (pp. 303–334). New York: International Universities Press.
Shields, J. (1962). Monozygotic twins. London: Oxford University Press.
Singh, A. K., & Lakhotia, S. C. (1988). Effect of low-temperature rearing on heat shockprotein synthesis and heat sensitivity in Drosophila melanogaster. DevelopmentalGenetics, 9, 193–201.
Slijper, E. J. (1942). Biologic-anatomical investigations on the bipedal gait and uprightposture in mammals, with special reference to a little goat, bomwithout forelegs. II.Proceedings Series C Biological andMedical Sciences: Koninklijke Nederlandse Akademie vimWetenschappen Amsterdam, 45, 407–415.
Smeyne, R. J., Schilling, K., Robertson, L., Luk, D., Oberdick, J., Curran, T., &Morgan, J. I.(1992). Fos-lacZ transgenic mice: Mapping sites of gene induction in the centralnervous system. Neuron, 8, 13–23.
Spelke, E. S., & Newport, E. L. (1998). Nativism, empiricism, and the development ofknowledge. In R. M. Lerner (Ed.), Handbook of child psychology: Theoretical models ofhuman development (5th ed., Vol. 1, pp. 275–340). New York: Wiley.
Stent, G. (1981). Strength and weakness of the genetic approach to the development of thenervous system. Annual Review of Neuroscience, 4, 163–194.
Strohman, R. C. (1997). The coming Kuhnian revolution in biology. Nature Biotechnology,15, 194–200.
Tanner, J. M. (1978). Foetus into man: Physical growth from conception to maturity. Cambridge,MA: Harvard University Press.
Telling, G. C., Parchi, P., DeArmond, S. J., Cortelli, P., Montagna, P., Gabizon, R.,Mastrianni, J., Lugaresi, E., Gambetti, P., & Pruisner, S. B. (1996). Evidence for theconformation of the pathologic isoform of the prion protein enciphering andpropagating prion diversity. Science, 274, 2079–2082.
Thaler, D. S. (1994). The evolution of genetic intelligence. Science, 264, 224–225.
Thelen, E., & Smith, L. B. (1994). A dynamic systems approach to cognition and action.Cambridge, MA: MIT Press.
Tooby, J., & Cosmides, L. (1990). The past explains the present: Emotional adaptationsand the structure of ancestral environments. Ethology and Sociobiology, 11,375–424.
Uphouse, L. L., & Bonner, J. (1975). Preliminary evidence for the effects of environmentalcomplexity on hybridization of rat brain RNA to rat unique DNA. DevelopmentalPsychobiology, 8, 171–178.
Valsiner, J. (1997). Culture and the development of children’s action: A theory of humandevelopment. (2nd ed.). New York: Wiley.
Van der Weele, C. (1995). Images of development: Environmental causes in ontogeny.Unpublished doctoral dissertation, Vrije University, Amsterdam.
von Bertalanffy, L. (1962).Modern theories of development: An introduction to theoretical biology.New York: Harper. (Original work published 1933)
36 Gilbert Gottlieb

Wahlsten, D. (1990). Insensitivity of the analysis of variance to heredity-environmentinteraction. Behavioral and Brain Sciences, 13, 109–120; open peer commentary,121–161.
Wahlsten, D., & Gottlieb, G. (1997). The invalid separation of nature and nurture:lessons from animal experimentation. In R. J. Sternberg & E. Grigorenko (Eds.),Intelligence, heredity, and environment (pp. 163–192). New York: CambridgeUniversity Press.
Weiler, I. J., Irwin, S. A., Klintsova, A. Y., Spencer, C. M., Brazelton, A. D., Miyashiro, K.,Comery, T., Patel, B., Eberwine, J., & Greenough, W. T. (1997). Fragile X mentalretardation protein is translated near synapses in response to neurotransmitteractivation. Proceedings of the National Academy of Sciences USA, 94, 5395–5400.
Weiss, P. (1939). Principles of development: A text in experimental embryology. New York: Holt,Rinehart and Winston. (Reprinted in 1969 by Hafner.)
Wigglesworth, V. B. (1964). The life of insects. Cleveland, OH: World Publishing.
Wills, C. (1989). The wisdom of the genes: New pathways in evolution. New York: Basic Books.
Woltereck, R. (1909). Weitere experimentelle Untersuchungen €uber Artver€anderung,speziell €uber das Wesen quantitativer Artunterschiede bei Daphniden. [Furtherexperimental investigations of species modification, particularly the nature ofquantitative species differences in daphnia]. Verhandlungen der DeutschenZoologischen Gesellschaft, 19, 110–173.
Wright, S. (1968). Evolution and the genetics of population: Vol. 1: Genetic and biometricfoundations. Chicago: University of Chicago Press.
Zamenhof, S., & van Marthens, E. (1978). Nutritional influences on prenatal braindevelopment. In G. Gottlieb (Ed.), Early influences (pp. 149–186). New York:Academic Press.
From Central Dogma to Probabilistic Epigenesis 37