Embed Size (px)
Citation preview

Copyright © 2009 Pearson Education, Inc.
Chapter 18 Developmental Genetics
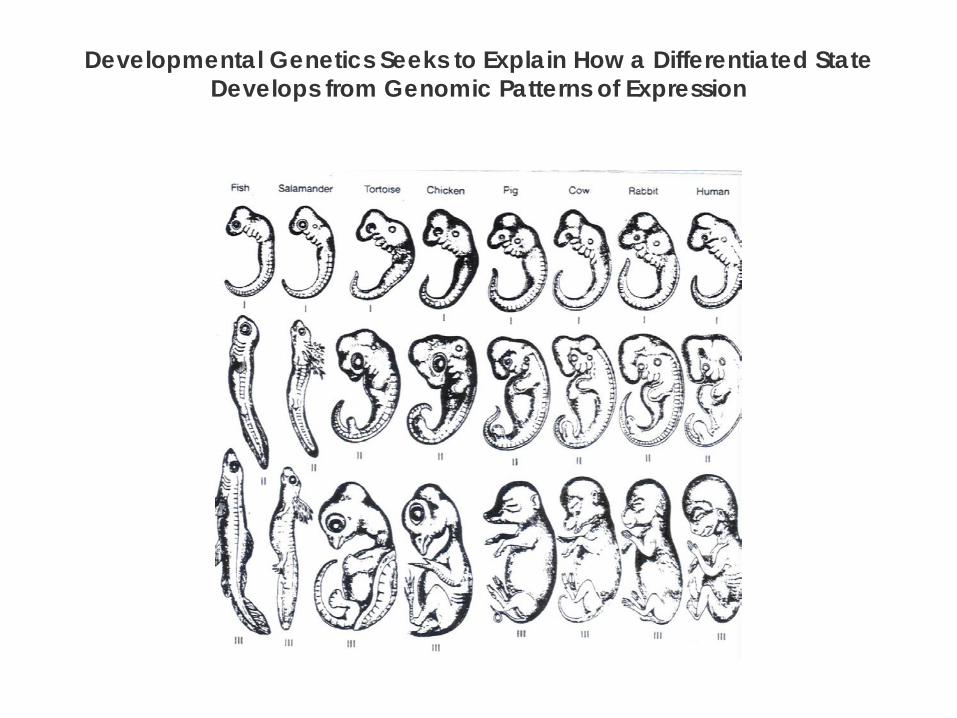
Developmental Genetics Seeks to Explain How a Differentiated State Develops from Genomic Patterns of Expression

Genetic questions in development
What genes are expressed?
When are they expressed?
In what parts of the developing embryo are they expressed?
How is the expression of these genes regulated?
What happens when these genes are defective?

Evolutionary conservation of development
Most developmental mechanisms and signaling systems are used in all multicellular organisms.
Pax6 is a gene required for eye formation humans, mice, frogs, flies, octopi, mollusks, sea squirts
Gehring, 1996

FGF-10 causes outgrowth of turtle’s shell
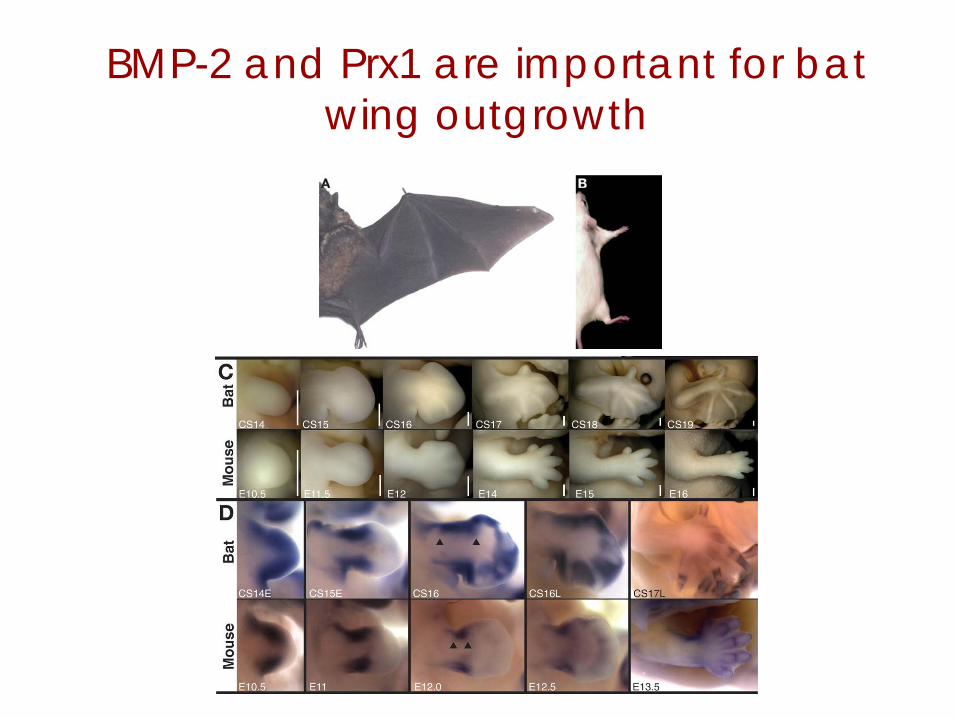
BMP-2 and Prx1 are important for bat wing outgrowth

The primary model organisms used for the genetic study of development
Drosophila, C. elegans, Mice, Zebrafish, Arabidopsis

Developmental concepts
Laying out a body plan: Drosophila
Programs of differentiation: Drosophila, Arabadopsis
Cell-Cell communication: C. elegans

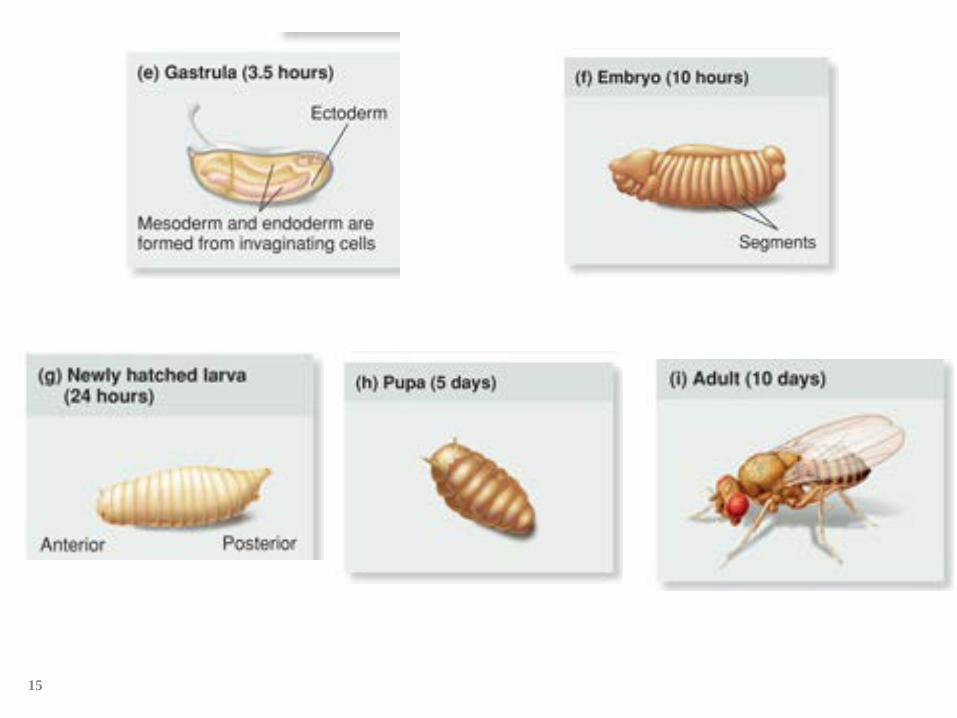
Drosophila life cycle
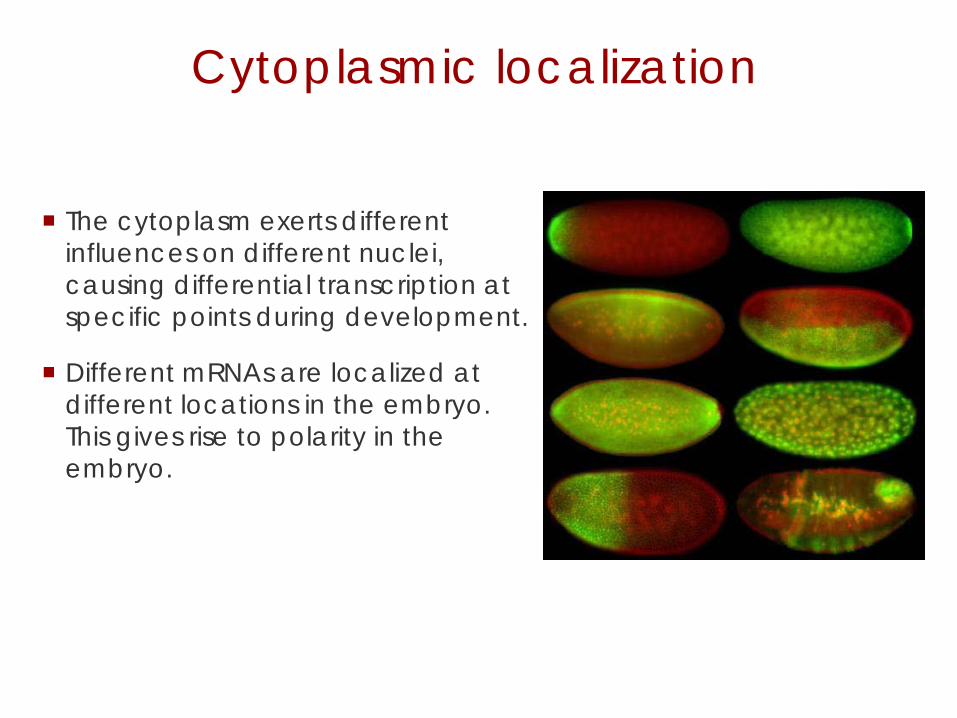
Cytoplasmic localization
The cytoplasm exerts different influences on different nuclei, causing differential transcription at specific points during development.
Different mRNAs are localized at different locations in the embryo. This gives rise to polarity in the embryo.

Maternal effect genes
When mutation occurs in a cytoplasmic gene or affects its localization, the flies carrying this mutation are not affected.
But the offspring of female flies with the mutation will be affected. Their eggs will not have proper localization of the cytoplasmic components.

Early embryo development
In the early stages of embryonic development, the zygote nucleus undergoes a number of divisions without cytokinesis.
These nuclei arrange themselves around the periphery of the egg and are exposed to different gradients of maternally derived transcripts and proteins.


Body plan is set up early
Insects have a segmented body plan
Segmentation is determined early in development
Cells within the segment have an orientation
The adult structures form from each segment of the embryo
15

Genes controlling embryonic development are either
Maternal-effect genes: mRNAs in the egg from the
mother
Zygotic genes Transcribed in the embryo from its DNA Two copies: One from mom, One from dad Often regulated by the maternal effect genes
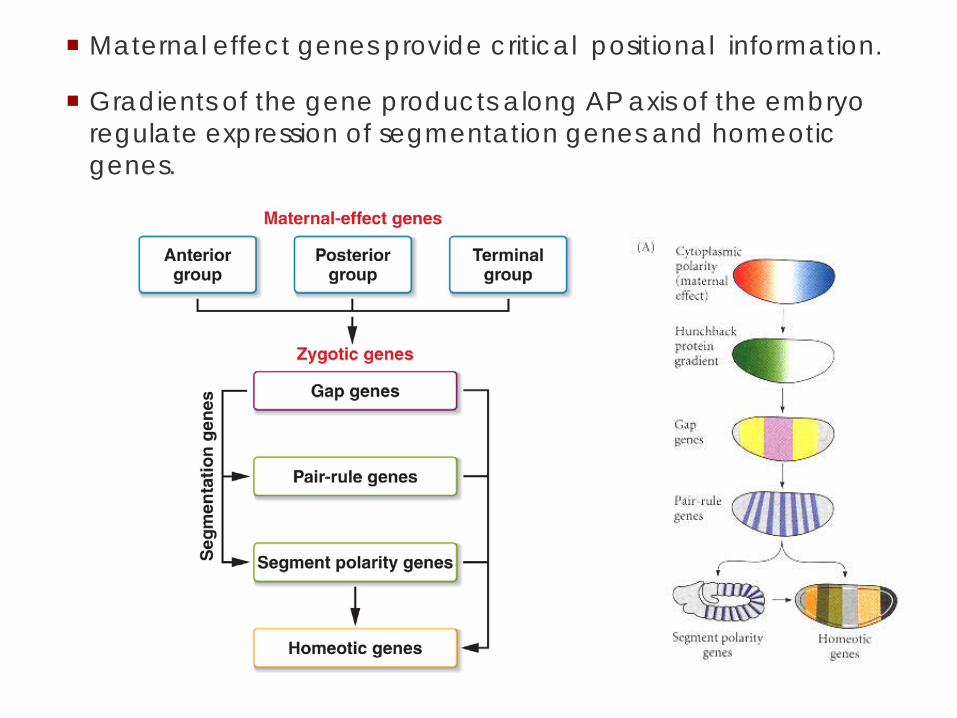
Maternal effect genes provide critical positional information.
Gradients of the gene products along AP axis of the embryo regulate expression of segmentation genes and homeotic genes.
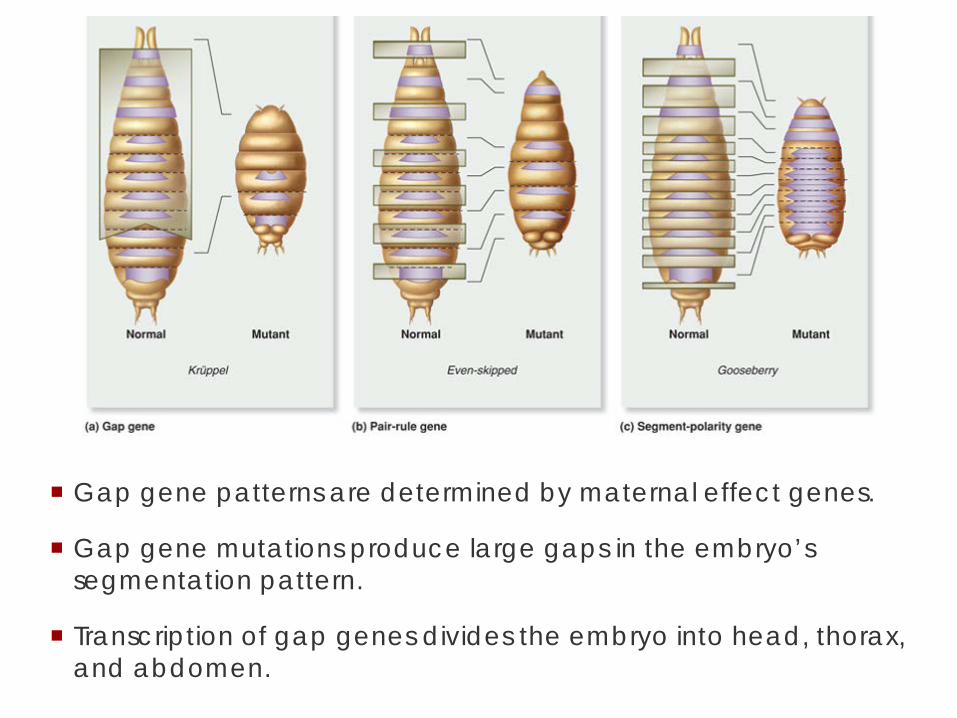
Gap gene patterns are determined by maternal effect genes.
Gap gene mutations produce large gaps in the embryo’s segmentation pattern.
Transcription of gap genes divides the embryo into head, thorax, and abdomen.

19
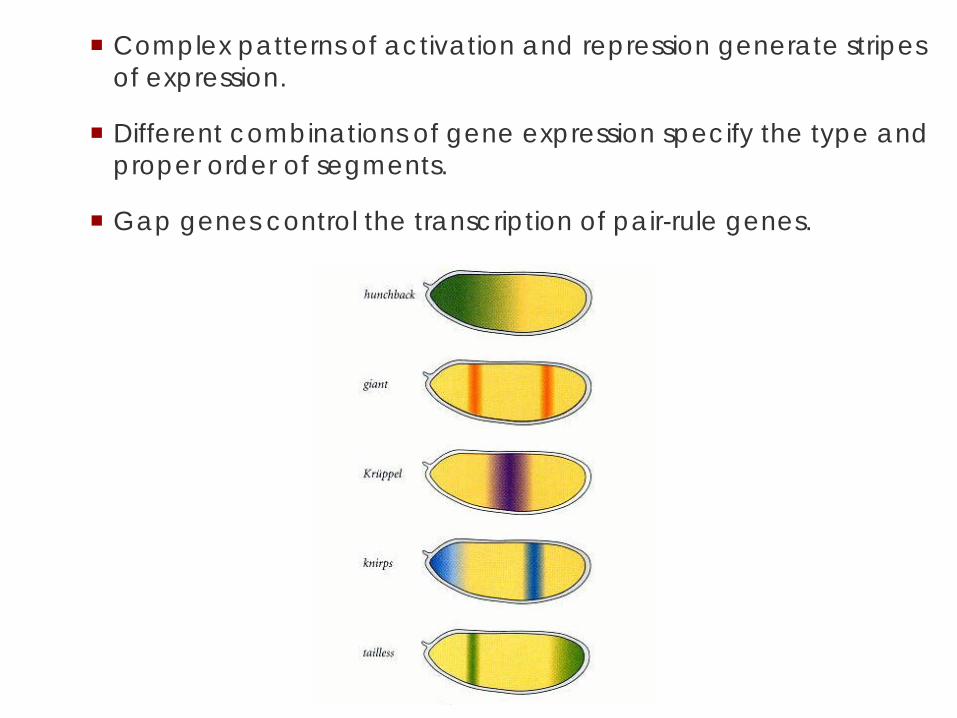
Complex patterns of activation and repression generate stripes of expression.
Different combinations of gene expression specify the type and proper order of segments.
Gap genes control the transcription of pair-rule genes.

Mutations in pair-rule genes eliminate segment-size sections at every other segment.
At least eight pair-rule genes act to divide the embryo into a series of stripes that overlap.

Complex regulation for each stripe of the pair-rule genes
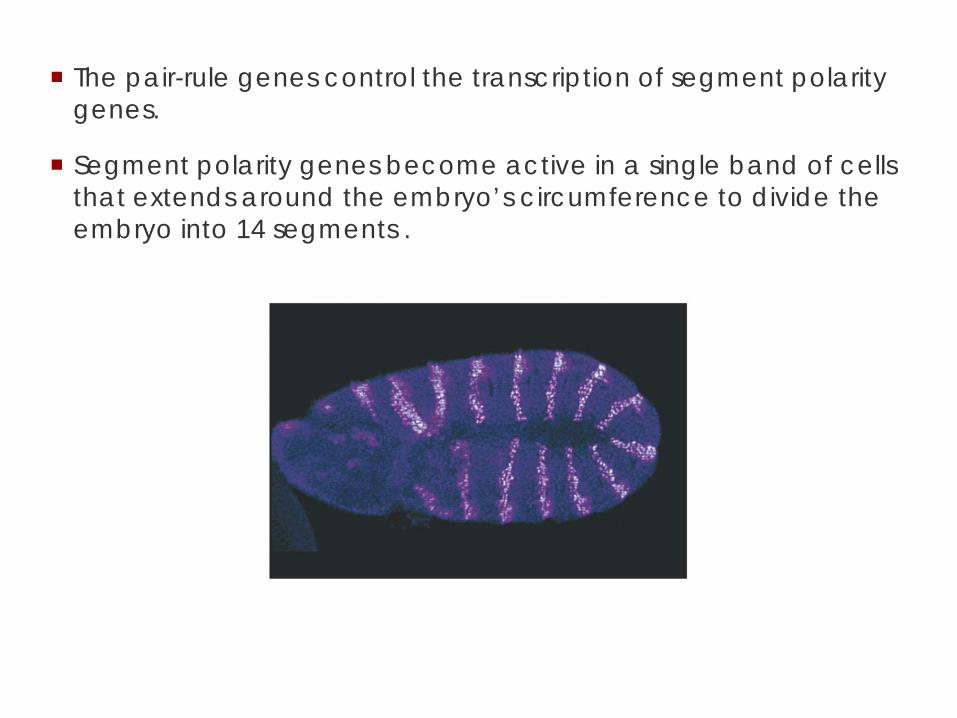
The pair-rule genes control the transcription of segment polarity genes.
Segment polarity genes become active in a single band of cells that extends around the embryo’s circumference to divide the embryo into 14 segments .

Many drosophila genes have human homologues.
Cleidocranial dysplasia (CCD) is the result of a mutation in the human CBFA (RUNX2) gene, the homolog of runt.
It is characterized by numerous skeletal defects.

Mice with one mutant copy of the runt homolog gene have skeletal abnormalities as seen with the human.
Those with two mutant copies have complete absence of bone formation.
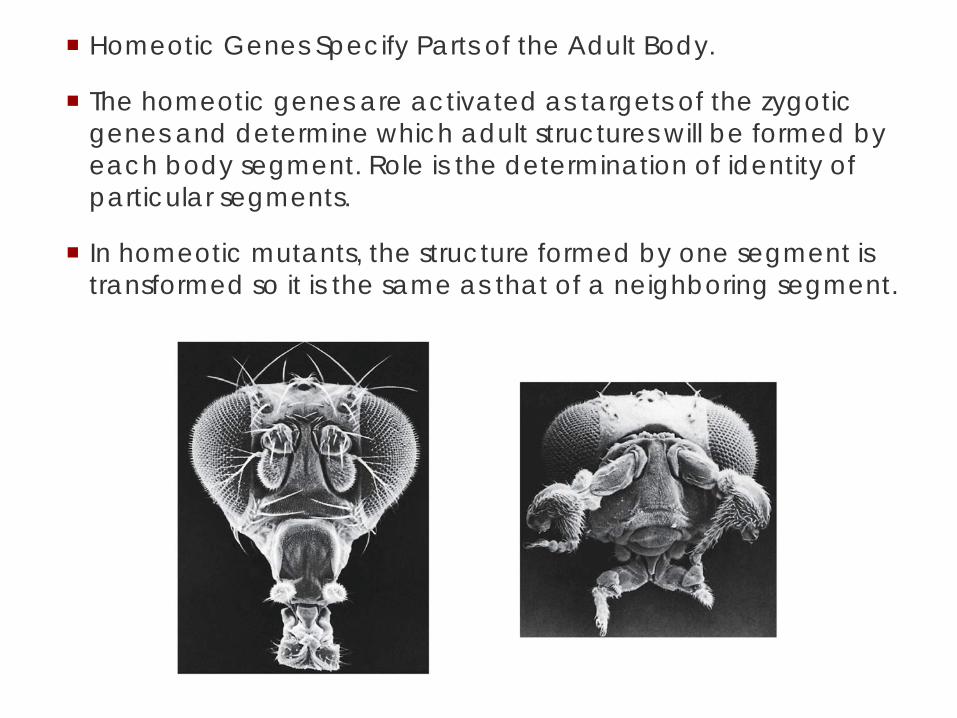
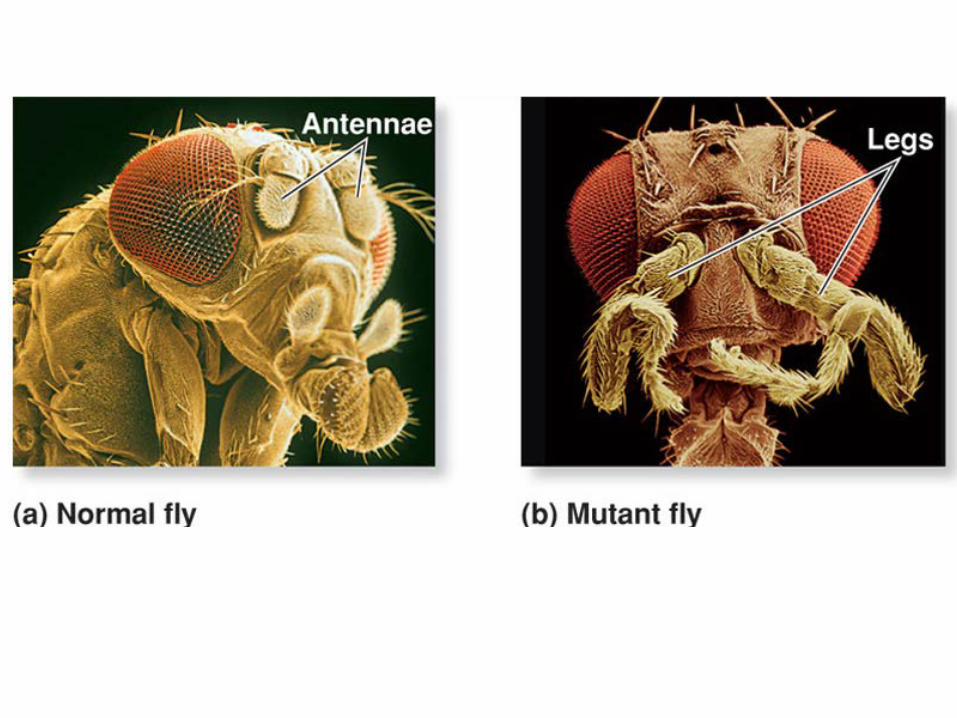
Homeotic Genes Specify Parts of the Adult Body.
The homeotic genes are activated as targets of the zygotic genes and determine which adult structures will be formed by each body segment. Role is the determination of identity of particular segments.
In homeotic mutants, the structure formed by one segment is transformed so it is the same as that of a neighboring segment.

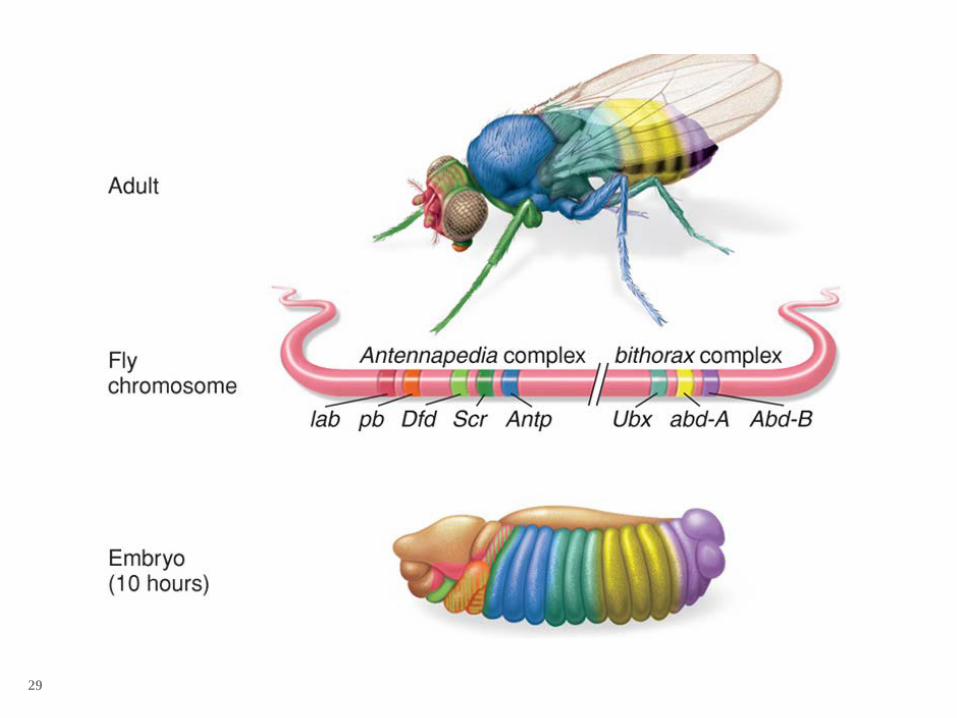
Hox genes are found adjacent to each other.
The Drosophila genome contains two clusters of homeotic selector (Hox) genes on chromosome 3.
The Antennapedia complex contains five genes that specify structures in the head and the first two thoracic segments.
The Bithorax complex contains three genes that specify the posterior segments .
29

30
Bithorax gene complex Normal – wings on 2nd thoracic segment and 2
halteres on 3rd thoracic segment Mutant – 3rd segment has wings so 2 sets of wings
and no halteres

Hox genes encode transcription factors that include a 60 aa DNA-binding homeodomain.
Expression of the Hox genes is colinear with position in embryo.

Hox genes are evolutionarily conserved.
Hox genes are found in the genomes of most eukaryotes with a segmented body plan.
Humans have four copies of the Hox cluster.
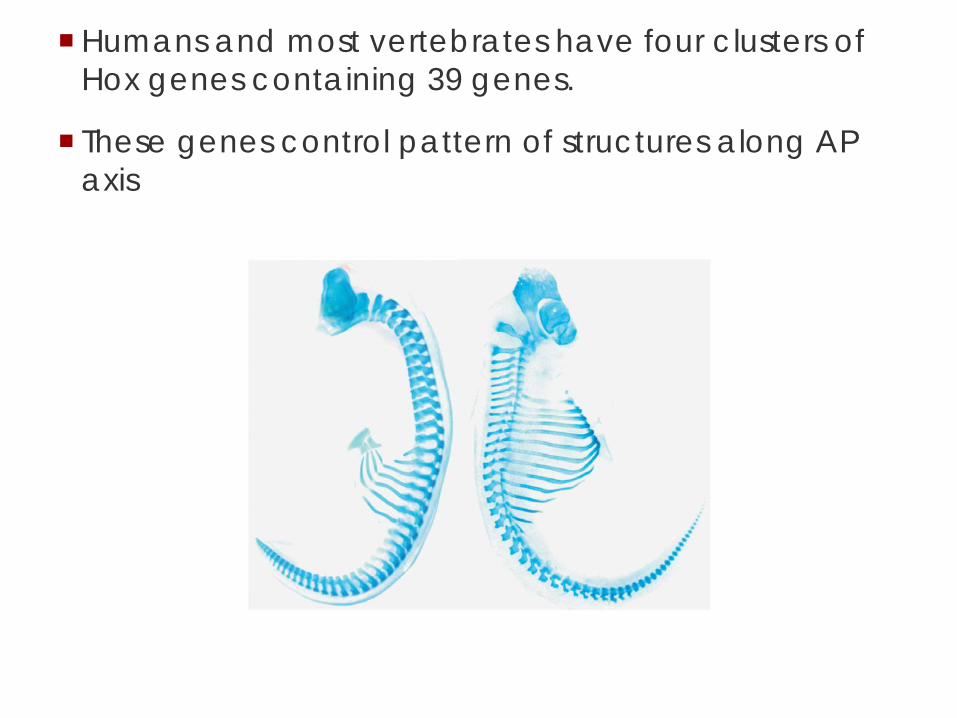
Humans and most vertebrates have four clusters of Hox genes containing 39 genes.
These genes control pattern of structures along AP axis
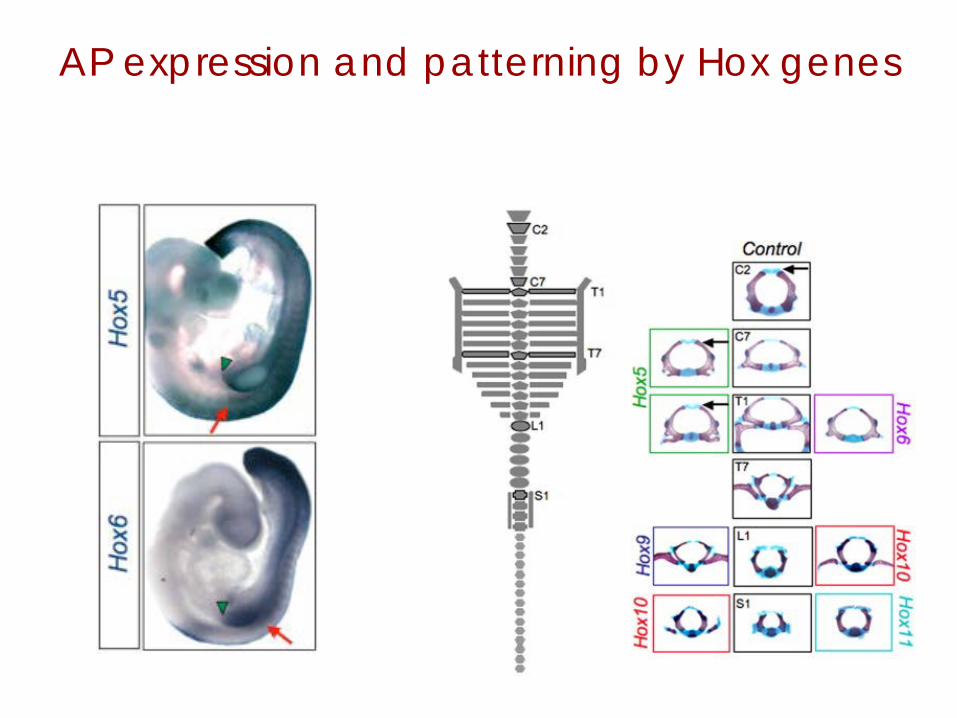
AP expression and patterning by Hox genes

Cell-cell signaling in C. elegans development
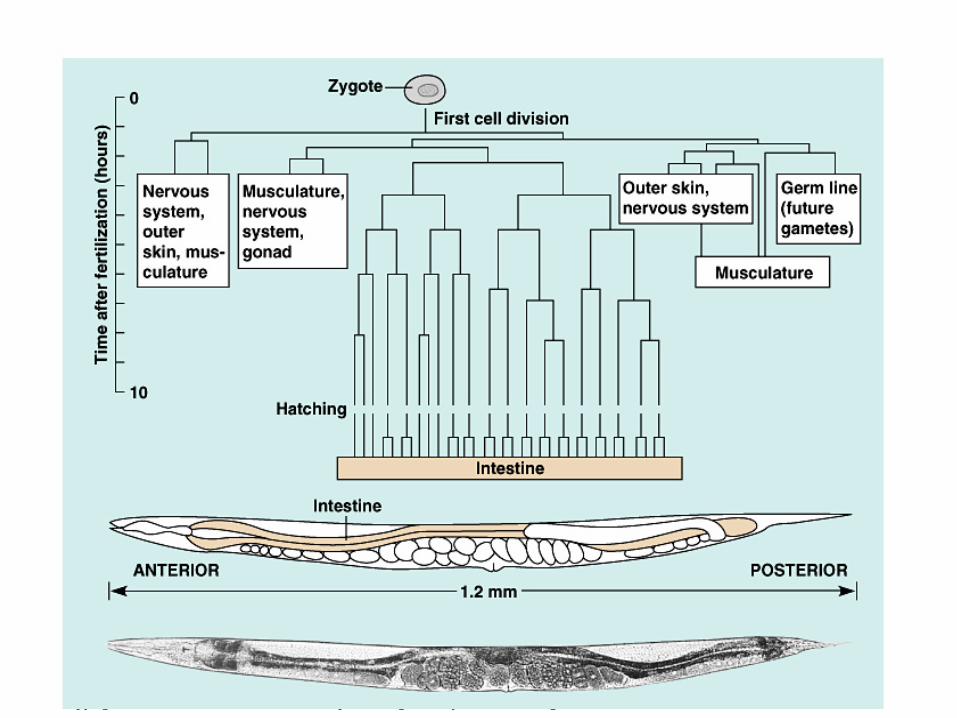
C. elegans has well-understood genetics and a completed genome sequence.
Adults are formed from 959 somatic cells, for each of which the developmental lineage from fertilized egg to adult has been mapped.
Copyright © 2002 Pearson Education, Inc., publishing as Benjamin Cummings


Vulval development involves three rounds of cell-cell interactions that transmit or receive signals from other cells.
Notch-delta signaling

Notch-Delta pathway signals between adjacent cells
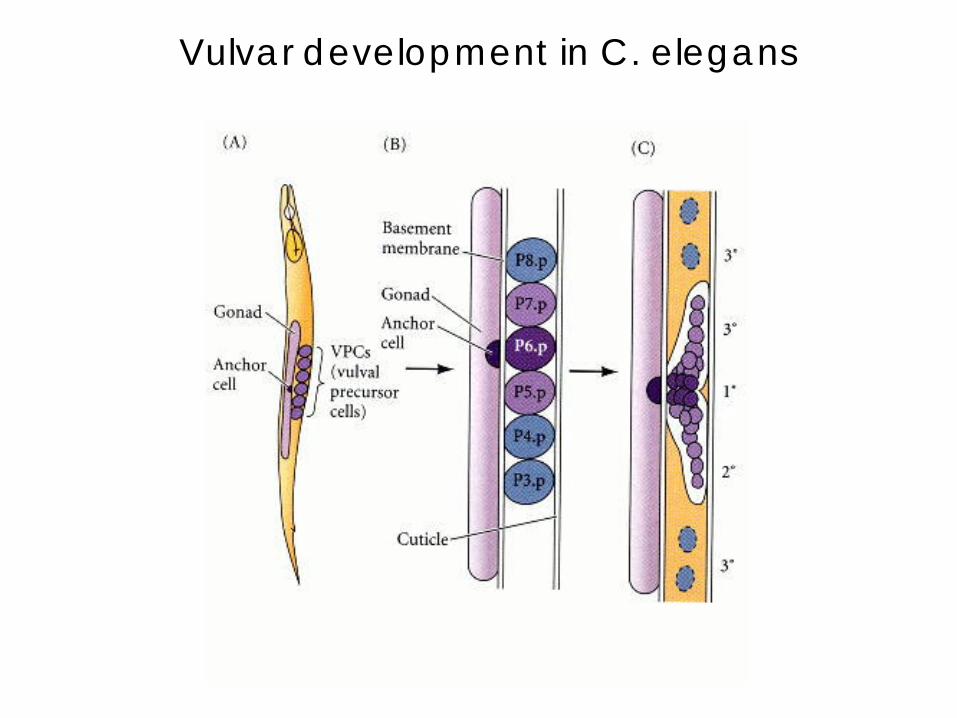
Vulvar development in C. elegans

• Already present on the ventral surface of the second-stage larva are six cells from which the vulva will arise.
• A single cell in the embryonic gonad, the anchor cell, initiates a cascade of signals that establishes the fate of the vulval precursor cells.
• It does so by producing an inducer protein that binds to receptors on the closest cells.

• The closer to the anchor cell, the more inducer binds; these precursor cells form the inner vulva.
• The inducer is a growth factor: the high levels of inducer probably cause division and differentiation of this cell to form this structure. It also activates a gene for a second inducer.
• Receptors on the two adjacent vulval precursor cells bind the
second inducer, which stimulates these cells to divide and develop into the outer vulva.
• Because the three remaining vulval precursor cells are too far away to receive either signal, they give rise to epidermal cells.