Embed Size (px)
Citation preview



Dedicated to Yvonne, Mariam and Hemed. For their care & inspiration.


List of Papers
This thesis is based on the following papers, which are referred to in the text by their Roman numerals.
I Jeneby, M. M., Suleman, M. A., and Gichuki, C.(2002). Sero-epizootiologic survey of Trypanosoma brucei in Kenyan nonhu-man primates. Part I. Journal of Zoo and Wildlife Medicine. 33(4):337-41.
II Gicheru, M.M., Jeneby, M. M., Macharia, J. C., Carlsson, H.E. and Suleman M. A. (2009). Prevalence of antibodies and cell mediated immune response against Leishmania major in feral non-human primates from Kenya. Part I. Acta Tropica. 109(2):136-40.
III Maamun M. Jeneby, M. Ngeiywa, D. S. Yole, J. Mwenda, M. A. Suleman, Carlsson HE. 2008. Enzootic simian piroplasm (Entopo-lypoides macaci) in wild-caught Kenyan nonhuman primates. J Med Primatol. Dec; 37(6):329-36.
IV Jeneby M. Maamun, Mbaruk A. Suleman, Mercy Akinyi, Hast-ings Ozwara, Thomas Kariuki and Hans-Erik Carlsson. 2011. Pre-valence of Babesia microti in free-ranging Kenyan baboons and African green monkeys. Journal of Parasitology 97(1):63-67.
V Jeneby M.M., Suleman MA, Ozwara HS, Carlsson HE. Detection and molecular characterization of haemoprotozoan parasites of wild-caught nonhuman primates in Kenya. Manuscript.
Reprints were made with permission from the respective publishers.


Contents
CHAPTER 1: GENERAL INTRODUCTION ............................................. 11 1.1 General introduction ........................................................................... 11 1.2 Parasites aetiology .............................................................................. 11 1.3 Non human primates (NHPs) ............................................................. 12 1.4 Haemoprotozoan parasites of NHPs ................................................... 13 1.5 Clinical disease presentation .............................................................. 14 1.6 Use of the NHPs ................................................................................. 14 1.7 Economic burden and zoonotic haemoprotozoa of NHPs .................. 15 1.8 Diagnosis ............................................................................................ 15
CHAPTER 2: OBJECTIVES AND RATIONAL FOR THE STUDY ......... 17 2.1 Objective overview ............................................................................ 17 2.2 Measurable specific objectives ........................................................... 17
2.2.1 To assess the prevalence of haemoflagellates of medical importance (Trypanosoma and Leishmania) in baboons, vervets and Syke’s monkeys caught and relocated away from densely populated rural areas (study I and II). ................................................................ 17 2.2.2 To determine the presence and distribution of malaria parasites and piroplasms in wild-caught baboons, vervets and Syke’s monkeys in Kenya (study III and IV). .............................................................. 17 2.2.3 To detect and characterize haemoprotozoan parasites in wild free-ranging NHPs using molecular tools and microscopic evaluation of blood smears (study V). .................................................................. 17
2.3 Hypothesis .......................................................................................... 17 2.4 Rationale............................................................................................. 18
CHAPTER 3: MATERIALS AND METHODOLOGY ............................... 19 3.1 Study sites .......................................................................................... 19 3.2 Capture of NHPs ................................................................................ 19 3.3 Collection of blood samples from NHPs ............................................ 20 3.4 Parasitological methods ...................................................................... 21 3.5 Haematological parameters ................................................................ 21 3.6 Antigen preparations .......................................................................... 21 3.7 Immunological assays ........................................................................ 22

3.8 Molecular identification of haemoparasites ....................................... 22 3.8.1Extraction of parasites DNA ........................................................ 22 3.8.2 PCR amplifications ..................................................................... 22 3.8.3 Sequencing and phylogenetic analysis ....................................... 23
3.9 Statistical Analysis ............................................................................ 23 3.9 Photography ....................................................................................... 24
CHAPTER 4: RESULTS .............................................................................. 25 4.1 Parasitological results ......................................................................... 25
4.1.1 Active infections ......................................................................... 25 4.1.2 Prevalence of E. macaci and H. kochi ........................................ 27
4.2. Immunological results ....................................................................... 29 4.2.1 Serological detection and characterization of piroplasm infecting NHPs .................................................................................... 29
4.3. Seroprevalence of T. brucei .............................................................. 29 4.4. L. major seroprevalence .................................................................... 30 4.5. Leishmania recall lymphocyte proliferation assay ............................ 32 4.8 Molecular detection and sequence analysis ........................................ 32
4.8.1 Molecular survey of B. microti ................................................... 32 4.9 Molecular survey and parasite characterization ................................. 34
5.0 Discussion ............................................................................................... 38 5.1 T. brucei prevalence ........................................................................... 38 5.2. L. major seroprevalence .................................................................... 39 5.3 Detection and distribution of E. macaci and H. kochi ........................ 40 5.4 Molecular detection, prevalence and characterization of haemoparasites ......................................................................................... 41 5.4 Conclusion .......................................................................................... 43
5.4.1 The main conclusions drawn from this study are; ...................... 44
Acknowledgements ....................................................................................... 45
Funding ......................................................................................................... 46
References ..................................................................................................... 47

Abbreviations
NHPs Nonhuman primates Ab-ELISA Antibody enzyme-linked immunosorbent assay Con A Concanavlin A PBMCs Peripheral blood mononuclear cells AGMs African green monkeys LAT Latex agglutination test WB Western blot CL Cutaneous leishmaniasis HAT Human African trypanosomiasis PCR Polymerase chain reaction Cyt-b Cytochrome b 18 SSrRNA 18 small subunit ribosomal RNA


11
CHAPTER 1: GENERAL INTRODUCTION
1.1 General introduction This study investigated on the presence, distribution and characterization of haemoprotozoan parasites of wild-caught African green monkeys (AGMs) or vervet monkeys (Chlorocebus aethiops), olive baboons (Papio cyno-cephalus anubis) and Syke's monkeys (Cercopithecus mitis) commonly found in Kenya. The importance of information regarding parasites of non human primates (NHPs) has previously been ignored by biomedical re-searchers (Ananias et al., 1997). However, there is a renewed interest in the role played by wild animal population as hosts of parasites for the benefit of wild animal conservation research (Gulland, 1995; Wolfe et al., 1998) and surveillance of emerging zoonotic pathogens (Wolfe et al., 1998; Seetham-chai et al., 2008). NHPs in Kenya are part of wild fauna, and in some regions they are mostly found in habitats close to human settlements (Eley, 1998).
1.2 Parasites aetiology The term ‘haemoprotozoa’ is derived from haem (blood) and protozoa, the unicellular parasitic organisms under the sub-kingdom Protozoa (Molyneux and Ashford, 1987). The genera Plasmodium and phylogeneticaly closely related parasite, Hepatocystis belong to the Phylum: Apicomplexa, Order: Coccidiida and Family: Plasmodiidae. The genera Entopolypoides, Babesia and Theileria are popularly known as ‘piroplasms’. They are classified un-der the Phylum: Apicomplexa, Order: Pyroplasmida and Family: Babesii-dae. Classification of the genera Leishmania and Trypanosoma, popularly known as ‘haemoflagellates’ (Molyneux and Ashford, 1983) is as follows: Phylum: Sarcomastigophora, Class: Zoomastigophora, Order: Kinetoplasti-da, Family: Trypanosomatidae, Genus: Trypanosoma and Leishmania. Para-sites in the phylum Apicomplexa are commonly referred to as ‘apicomplex-ans’.

12
1.3 Non human primates (NHPs) Old World monkeys (Catirrhini) and apes (Hominoidea) are those found in Africa, the Indian sub-continent and in East Asia. New World or neotropical NHPs (Platirrhini) are found in South and Central America. Old world mon-keys are comprised of 16 genera, of which 10 are found in Africa (Oates, 1996). Among the Old World African monkeys, Cercopithecus species (AGMs and Syke’s monkeys) compete with Papio species (baboons) as the most widely distributed monkeys in South and Central Africa. Cercopithecus spp. are widely spread in terms of ecological habitats, being found in the rain forests, woodlands and savannah in sub-Saharan Africa, from Senegal in the West and Ethiopia in the East to the Cape in the South. There is also a signif-icant population of AGMs on the island of St. Kitts in the Caribbian brought there from Africa by sailors during the 17th and 18th ceturies. Baboon’s dis-tribution stretches from the arid desert fringes in Arabia in the North, to the tip of Cape in the South Africa (Haltenorth and Diller, 1977).
In Kenya, AGMs and baboons are the most widely distributed and com-monest monkeys. They are also regarded as pests by farmers (Eley, 1989; Eley et al., 1989b). The AGMs are found in savanna, woodland, and rive-rine, gallery, lakeshore and coastal forests. Syke’s monkeys are arboreal and are widespread throughout forested regions in Kenya. Baboons are adaptable opportunists and do well almost anywhere including near human habitation. They are found in a wide variety of habitats, from forest through grasslands to semi-arid areas (Eley, 1989; Jonathan, 1974). According classification by Jonathan (1974), the genera, Papio, Chlorocebus and Cercopithecus are classified under the Phylum: Chordata, Order: Primates, and Family: Cer-copithidae. Species of the genus Papio found in Kenya include (i) olive ba-boon (P. cynocephalus anubis), and yellow baboon (P. cynocephalus cyno-cephalus). Species of the genus Chlorocebus (formally Cercopithecus), African green monkey or vervet monkey (C. aethiops) and Cercopithecus spp. namely Syke’s monkey (C. mitis), De Brazza’s (C. neglectus) and crested mangaby (C. galeritis).
From the description of Haltennorth and Diller (1977), vervet monkey is a grey-brown monkey with a green back. They also have white fringed black face, long whitish cheek whiskers, and white surrounds to the eyes, black feet, and black tipped tail. Males have blue scrotum and red penis. The body length is 40 - 60cm and tail is about 15cm longer than the body. Syke’s monkey is same size as the vervet monkey but much heavier with males weighing 5 - 10 Kg and females 3 - 6 Kg. Syke’s or mitis monkey is black or blue - black with its back and face flecked with silver or grey fur, round black face, and black limbs and tail. The muzzle and chin hairs are grey and the naked nose and area around the eyes is blue-grey, the males have whitish ear tufts. Baboons are the largest terrestrial monkeys. They have prominent muzzle, a naked dog - like face, close -set eyes and large chick pouches.

13
They have deep barrel chests and walk on their fingers and soles of their feet (plantigrade). Both males and females have grey skinned hairless buttocks and large flesh coloured ischial callosities (sitting pads) which are separated in the females but continuous in males. As the name implies the Olive ba-boon is a grey - brown in colour with a naked black muzzle and black hands and feet. Yellow baboon has yellow-brown fur, and grey hands and feet. Compared with olive baboons, the yellow baboons have a slender appear-ance and its upper lips are not as prominent and its nose is not up-turned.
1.4 Haemoprotozoan parasites of NHPs Parasitic haemoprotozoan parasites which include species of Plasmodium, Leishmania, Trypanosoma, Hepatocystis, Giardia and Babesia infect various species of New World monkeys from South America and Old World mon-keys from Asia (Otsuru and Sekikawa, 1979; Collins and Aikawa 1993; Gozalo and Tantalean, 1996; Ziccardi and Lourenco-de-Oliveira, 1997). Old World monkeys and apes from Africa are hosts to Hepatocystis kochi, Ento-polypoides macaci and various species of Plasmodium (Garnham, 1966). Previous epidemiological surveys of blood parasites reported P. reichnowi and P. shwetzi as natural malaria parasites of African apes, chimpanzee (Pan troglodytes) and gorillas (Gorilla gorilla). Mangabey monkeys (Cercocebus albigena, C. atys and C. albegena) in Central and West Africa were found to harbour P. georgesi, P. gonderi and P. petersi infections (Collins and Ai-kawa, 1993). Baboons and AGMs in Kenya are naturaly parasitized by H. kochi, E. macaci (Leathers, 1978; Turner et al., 1982; Jeneby et al., 2008) and Babesia microti-like parasites (Jeneby et al., 2011). H. Kochi was for-merly called Plasmodium Kochi and was first observed on the livers of AGMs in the form of macroscopic cysts (Garnham, 1966). Simian piro-plasm, E. macaci was first described in Macaca irus monkeys from Java by Mayer in 1934 (Gleason and Wolf, 1974). It is synonymous to the economi-cally important livestock and human piroplasm of the genus Babesia (Bronsdon et al., 1999).
AGMs are experimentally susceptible to the African salivarian trypano-somes group, Trypanosoma brucei rhodesiense (Poltera, 1985; Pearson et al., 1986). However, natural infection of wild-caught NHPs from East Africa has not been established. Nevertheless, theoretically some baboons and AGMs are perceived to be reservoir hosts of trypanosomes (Fiennes, 1967; Kuntz, 1973). A study on seroprevalence of T. brucei in wild-caught ba-boons, AGMs and Syke’s monkeys in Kenya detected high titres of anti-T. brucei antibodies (Jeneby et al., 2003). A case report identified a natural and active Leishmania major infection in a Kenyan wild-caught AGM (Binhazim et al., 1987). Seroprevalence survey of L. major in wild-caught baboons,

14
AGMs and Syke’s monkeys in Kenya reported positive titres of anti- L. ma-jor antibodies (Gicheru et al., 2009).
1.5 Clinical disease presentation Most NHPs malarias and infections with piroplasms such as E. macaci or B. microti are generally asymptomatic. However, the infections may cause debilitation and overt diseases can be precipitated by stress, concurrent dis-eases, splenectomy, or immunosupression (Coatney et al., 1971; Bronsdon et al., 1999; Voorberg-van der Wel et al., 2008). Although E. macaci is claimed to be a non-pathogenic, eliciting mild fever in infected monkeys (Gleason & Wolf, 1974) it has however been shown to cause acute anaemia in immunosupressed or splenectomized monkeys (Bronsdon et al., 1999). H. kochi is known to be non pathogenic in the monkey host. It however elicits macroscopic milky white cysts in the liver of the infected animal (Garnham, 1966). Experimental infection of AGMs with species of Trypano-soma and Leishmania produce a variety of clinical diseases depending on the virulence or tropism of the parasite and differential host immune responses (Pearson et al., 1986; Olobo et al., 2001).
1.6 Use of the NHPs Baboons, AGMs and to a lesser extent Syke's monkeys have figured promi-nently in various disciplines of biomedical research (Suleiman et al., 2000; Farah et al., 2001; Olobo et al., 2001; Carlson et al., 2004). Biomedical re-search with NHPs plays a crucial role in the advancement of human and animal health. The NHPs are genetically and physiologically similar to hu-mans. These similarities are critical to biomedical research targeting the causes, progression, prevention, and treatment of a wide variety of diseases (Carlson et al., 2004). Because nonhuman primates reflect the anatomical, physiological, and behavioural makeup of humans, they provide an indispen-sable, and currently irreplaceable, bridge between basic laboratory studies and clinical use. Much of what we know about the brain, heart disease, Alz-heimer’s, AIDS, viruses, hepatitis, and cancer has come from nonhuman primates (Brack, 1987). Besides their use in research, NHPs are also kept as pets and to a large extent they are hunted for their meat in central African countries (Harder et al., 2002; Njiokou et al., 2004).

15
1.7 Economic burden and emerging zoonoses The genera Plasmodium, Trypanosoma and Leishmania are the causative agents of human diseases, malaria, trypanosomiasis and leishmaniasis re-spectively. Trypanosoma, Babesia and Theileria parasitize wild and domes-tic animals causing debilitating diseases such as trypanosomosis, babesiosis and theileriosis respectively (Uilenberg, 1995; Shaw, 2004). NHPs are sub-ject to contact with vectors which transmit various human and livestock pa-rasitic protozoa (kuntz, 1970; Brack, 1985). In regard to potential zoonoses between NHPs and humans, approximately 13 (40 %) parasites affecting neotropical NHPs have been identified to be pathogenic to humans. (Levine, 1970 ; Toft, 1986; Brack, 1987). Going through the whole list of protozoan parasite hosts, one can draw parallels between humans and NHPs. The paral-lels would be much closer if more information about the parasites of the monkeys were known (Levine, 1970).
There is increasing awareness of the impact of nonhuman primate-borne malaria zoonoses in Malaysia (Dean, 1992; White, 2008; Seethamchai et al., 2008). This raises the question as to whether monkeys living in the rainfo-rests are natural reservoirs for both simian and human malaria. The results may have implications for the interpretation of the current epidemiological situation of malaria (Fandeur et al., 2000). B. microti-like parasites have been detected in a rhesus monkey (Macaca mulatta) and in wild-caught Ke-nyan baboons, AGMs and Sykes’ monkeys (Voorberg-vd Wel et al 2008; Jeneby et al., 2011). Although the transmission from NHPs to humans has not been established, B. microti has emerged as significant human pathogen in recent years. The rodent species B. microti in North America, the bovine species B. divergens in Europe, and as-yet-unnamed species referred to as WA1 are now recognized as important zoonotic agents of human babesiosis (Chisolm et al., 1986; Herwaldt, 1995; Tabara et al., 2007).
Human infections by Trypanosoma and Leihsmania spp. are largely con-sidered anthropophilic. Humans are in most cases regarded as accidental hosts of Leishmania sp., with gerbils and dogs being the sylvatic and domes-tic reservoirs, respectively (Beach et al., 1984; Ryan et al., 2006). The etio-logical agents of sleeping sickness, T. brucei rhodesiense is a zoonotic infec-tion with animal reservoirs, primarily game animals such as waterbuck, har-tebeest, reedbuck, duiker, and antelope (Harder et al., 2002; Njiokou et al., 2004). In West Africa, the domestic pig is considered the only important reservoir host of T. B. Gambiense. (Schares et al., 1996).
1.8 Diagnosis Microscopy remains the ‘gold’ standard for the diagnosis of many protozoan parasites. However, it is labour intensive and requires specialized micro-

16
scopists for accurate identification and interpretation of results. Definitive identification is particularly difficult when parasites are morphologically similar, very small in size or parasitemia is low (Etkind et al., 1980; Singh, 1997). Cultivation of parasites in vitro and parasite amplification by inocu-lation of infected host blood into naïve hamsters or mice is time consuming and often expensive (Morgan and Thompson, 1998). Serological techniques such as immunofluorescence antibody test (IFAT) and enzyme-linked im-munosorbent assay (ELISA) have been a method of choice in epidemiology and diagnosis of many haemoprotozoan parasites (Aiyedun et al., 1976; Homer et al., 2000; Berman, 1997). However, these techniques may pose a problem because of cross-reactions with other closely related organisms resulting to differences in interpretation between laboratories (Passos et al., 1998; Hanscheid et al., 2003). To circumvent the limitations of conventional diagnostic tests, highly sensitive molecular tools such as polymerase chain reaction (PCR) tests are now preferred methods. PCR tests have been uti-lized for conducting epidemiological surveys of simian malaria parasites (Snounou et al., 1993; Gubbels et al., 1999) and piroplasms infecting NHPs (Figueroa, 1995; Caccio et al., 2000). In recent years, PCR-based diagnostic methods have been described for detection of trypanosomiasis (Chappuis et al., 2005) and leishmaniasis pathogens (Kumar et al., 2007). The use of primers targeting Cty-b and 18SrRNA genes of haemoparasites is widely used in molecular surveys for NHPs haemoparasites and their molecular characterization (Escalante and Ayala, 1994; Qari et al., 1996; Morgan and Thompson, 1998; Seethamchai, 2008).

17
CHAPTER 2: OBJECTIVES AND RATIONAL FOR THE STUDY
2.1 Objective overview This study reports on a series of field and laboratory investigations focusing on evaluating the occurrence and distribution of haemoprotozoa parasitizing wild-caught Papio cynocephalus anubis (olive baboons) and Chlorocebus aethiops (African green monkeys, AGMs) and Cercopithecus mitis (Syke's monkeys) in Kenya. In addition, parasites detected in non human primates were PCR amplified, DNA isolates sequenced and their phylogeny deter-mined.
2.2 Measurable specific objectives 1. To assess the prevalence of haemoflagellates of medical importance
(Trypanosoma and Leishmania) in baboons, vervets and Syke’s monkeys caught and relocated away from densely populated rural areas (study I and II).
2. To determine the presence and distribution of malaria parasites and pi-roplasms in wild-caught baboons, vervets and Syke’s monkeys in Kenya (study III and IV).
3. To detect and characterize haemoprotozoan parasites in wild free-ranging NHPs using molecular tools and microscopic evaluation of blood smears (study V).
2.3 Hypothesis The present study is based on the null hypotheses that wild free ranging non human primates in Kenya are not reservoirs of haemoprotozoan parasites of medical and veterinary importance.

18
2.4 Rationale Studies on pathogens of wildlife are critical in providing basic information about the life history traits such as virulence, host specificity and the evolu-tion of the genetic diversity of pathogens (Grenfell and Gulland, 1995). As potential carriers of various pathogenic agents capable of causing disease in humans, wild free-ranging NHPs are now recognized as important reservoir hosts for human pathogens. This is important especially in situations where human population is living close to monkey habitats (Wolfe et al., 1998). A case in point is the major public health concern over sporadic human infec-tions with P. knowlesi, a parasite of the macaque monkeys in Southern Thai-land and Malaysian peninsular is (Singh et al., 2004; White, 2008; Cox-Singh et al., 2008). As reservoir hosts for various zoonotic pathogens (Brack, 1987), NHPs populations may also hold valuable clues to the origins and evolution of some important human pathogens (Wolfe et al., 1998) such as P. falciparum (Liu et al., 2010). In addition, the presence of haemopro-tozoan parasites in NHPs might result in substantial, unanticipated impact on research programs by skewing the immunological profiles of experimental animals (Bronsdon et al., 1999; Voorberg-van der Wel et al., 2008). There-fore, there is need for parasite surveillance in NHPs living in close proximity to human settlements and in NHPs used in biomedical research. Previous surveys on haemoprotozoan parasite infecting NHPs from the E. African region were conducted using mainly parasitological techniques (Kuntz and Moore, 1975; Turner et al., 1982, Jeneby et al., 2008). Although highly use-ful, these surveys left an information gap on definitive molecular identifica-tion of haemoprotozoan parasites of NHPs in Kenya, East Africa. The main objective of this study was therefore to utilize the conventional diagnostic methods together with the molecular techniques to identify and characterize haemoprotozoan parasites circulating in a population of wild free-ranging baboons, AGMs and Syke’s monkeys in Kenya.

19
CHAPTER 3: MATERIALS AND METHODOLOGY
3.1 Study sites Sampling locations included Juja and Kilimanbogo in Thika District Central Province, Marigat in Baringo Districts Rift Valley province, Watamu in Kili-fi District Coast province, Kibwezi in Makueni District Eastern province and Kisamis and Nguruman in and Kajiado Districts in Rift Valley Province and Mt. Elgon in Elgon District in Western Province (Fig. 1). Apart from being endowed with different kinds of NHPs populations, among them vervets, Sykes’ monkeys, and baboons (Eley, 1987), some of these districts are also known to be malaria, leishmaniasis and trypanosomiasis endemic foci. In Thika, Baringo, Makueni and Kilifi districts, troops of NHPs frequently in-vade farms. This phenomenon has generated intense conflict between the NHPs and people in these areas making it necessary to translocate the mon-keys to other parts of the country. The translocation exercise provided the opportunity to examine the animals for protozoan haemoparasites.
3.2 Capture of NHPs Permit for trapping the NHPs and collection of specimens from them was obtained from the Kenya Wildlife Service, Nairobi. Animal acquisition, care, and maintenance were undertaken with the approval of and according to the guidelines of Institute of Primate Research (IPR) Institutional Scientific Review Committe (ISERC). Rectangular traps (1.5m high and 0.9m wide) made of iron bars, mesh wire and weighted trap doors designed by IPR staff were used (Papers I, II and III ). These kind of traps have been successful-ly used in the trapping of baboons for translocation project (Eleyet et al., 1987b). Briefly, monkeys were baited for a week or two with fresh whole maize cobs, carrots and bananas. After the monkeys became habituated, the traps were set in the evening after the animals had scampered to their sleep-ing site with the aim of capturing them the next day. After each trapping round, animals were transferred from the traps into smaller holding cages (0.8 m high by 0.6 m wide) before moving them to the holding site.
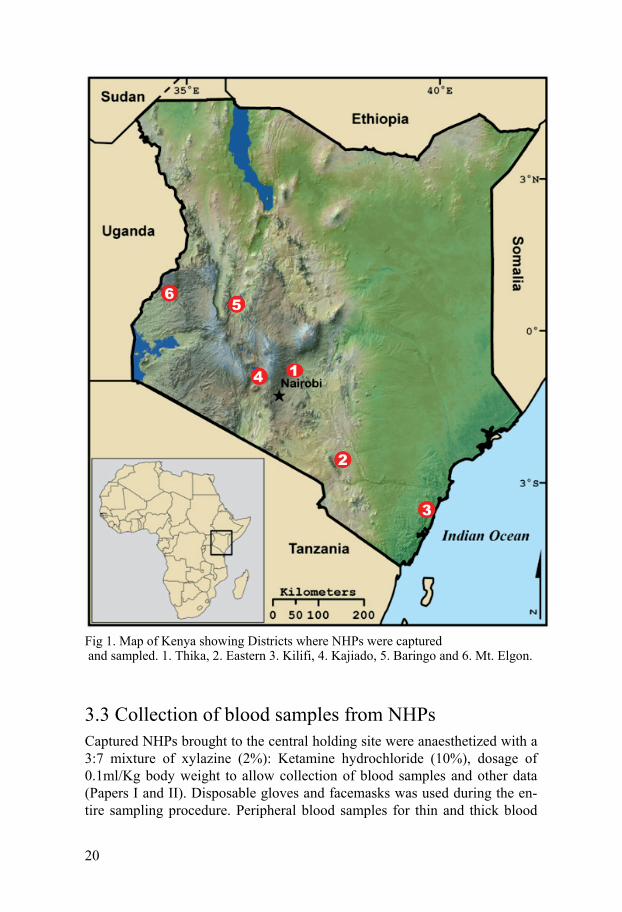
20
Fig 1. Map of Kenya showing Districts where NHPs were captured and sampled. 1. Thika, 2. Eastern 3. Kilifi, 4. Kajiado, 5. Baringo and 6. Mt. Elgon.
3.3 Collection of blood samples from NHPs Captured NHPs brought to the central holding site were anaesthetized with a 3:7 mixture of xylazine (2%): Ketamine hydrochloride (10%), dosage of 0.1ml/Kg body weight to allow collection of blood samples and other data (Papers I and II). Disposable gloves and facemasks was used during the en-tire sampling procedure. Peripheral blood samples for thin and thick blood

21
smears was collected as follows; ear or fingertips of NHPs was first steri-lized using clean cotton swab soaked with 70% ethanol. The sterilized area was pricked using a sterile blood lancet. Blood drops were used for prepara-tion of thin and thick smears, wet blood films and DNA extraction. Venous blood (5 ml) for serum preparation was collected in sterile 9 ml serum tubes (Greiner bio-one, France). Blood (5 ml) for lymphocyte recall was collected in sterile 9 ml EDTA tubes (Greiner bio-one, France) and used immediately for cell separation and proliferation assay. Serum was stored at -20oC until tested.
3.4 Parasitological methods Thin and thick blood smears were prepared by conventional method with blood from finger or ear prick of NHPs and examined under light micro-scope at x 1000 magnification (Paper I and II). Wet blood films were pre-pared from EDTA blood and examined under light microscop at x 40 (Paper I). Mouse sub-inoculation and microhematocrit (buffy coat) techniques were utilized to detect trypanosomes in fresh EDTA blood samples (Paper I). Swiss mice were inoculated intraperitoneally with 0.2 ml of fresh EDTA blood samples drawn from 23 AGMs captured in Kibwezi which were trans-ferred to the Institute of Primate Research quarantine facility. Blood smears and wet blood films were prepared from the tail blood and examined for trypanosomes by light microscopy.
3.5 Haematological parameters Routine haematological parameters were determined using EDTA blood drawn from the wild-caught NHPs (Paper II). The erythron indices, packed cell volume (PCV), red blood cell (RBC), haemoglobin (Hb) level and plate-lets were recorded using microhematocrit capillary tubes centrifuged for 5 min and read on a Hawksley microhematocrit reader (Hawksley, England). Total leucococyte count, differential white cell counts (WBC), total plasma protein, mean corpuscular volume (MCV) and plasma fibrinogen was deter-mined manually.
3.6 Antigen preparations Crude soluble Trypanosoma brucei brucei antigen was prepared from try-panosomes acquired from rats inoculated with T. brucei brucei (Paper I). Antigen for Leishmania detection was prepared from L. major (NLB-144) strain (Paper II). It was originally isolated from a sand fly (Phlebotomus

22
duboscqi) vector in Baringo District and maintained in BALB/c mice by serial subcutaneous passage. Determination of Leishmania and Trypanosoma protein concentrations were done using modified Bradford-Protein Dye-Binding assay reaction as described in Paper II.
3.7 Immunological assays Antibodies against T. brucei and L. major were determined by indirect anti-body ELISA assays (Paper I and II). Sera antibody reactivity to L. major was also determined by sodium dodecyl sulphate polyacrylamide gel elec-trophoresis (SDS-PAGE) as described in Paper II. Recall proliferation as-say was performed using formalin fixed L. major promastigotes to probe peripheral blood leucocytes (PBL) from AGMs (Paper II). T. brucei anti-gens were detected by the latex agglutination test (LAT) using suspension of latex particles coated with monoclonal antibodies against invariant internal antigens for T. brucei (Paper I). Sero-reactivity to whole parasite antigens of E. macaci, B. bovis, B. bigemina and B. microti were determined by indirect fluorescent antibody test (IFAT) (Paper III).
3.8 Molecular identification of haemoparasites
3.8.1Extraction of parasites DNA Two methods of DNA extraction were used (Paper IV). DNA from dried blood samples absorbed on filter papers were extracted using Chelex-100 method (Sigma Chemical Co, st. Louis, US) while DNA from EDTA blood (0.5 ml) was extracted by QIAamp DNA extraction kit (Qiagen, Hilden, Germany).
3.8.2 PCR amplifications Detection of B. microti in NHPs blood samples and ticks was carried out with two step nested PCR (N-PCR) with primers targetting fragment of -tubulin gene of Babesia and Theileria sp. N-PCR was performed essentialy as described by Caccio et al., 2000 (Paper IV). Briefly, primary amplification, was performed with primers F34 (5'-TGTGGTAACCAGAT(t/c)GG(a/t)GCCAA-3'), and R323 (5'-TCnGT (a/g)TA(a/g) TG nCC(t/c)TT(a/g)GCCCA-3') in a 50 µL reaction. Secondary amplification used 1 µL of primary amplification product with internal primers, F79 (5'-GA(a/g)CA(t/c)GGnAT nGA(t/c)CCnGTAA-3') and R206 (5'-AC(a/t/g)GA(a/g) TC CATGGT(a/t/g) CCnGG (t/c)T-3') in a

23
25 µL reaction. Amplifications were performed in DNA engine (PTC-200 Peltier thermal cycler MJ Research INC. Massachusetts, USA).
For the detection of Plasmodium and Hepatocystis spp., a set of primers targeting sections within the mitochondrial cytochrome b (Cty-b) gene were utilized (Paper V). PCR was performed using forward primer, MTCbF0, (5-GTAATGC CTAGACGTATTCCT-3) and reverse primer, MTCbR0, (5-ACTCCCTATCATGTC TTGC-3). Nested amplification of the 18 small subunit ribosomal RNA (18SS rRNA) gene of Babesia, Theileria and the related piroplasm of the genera Entopolypoides and Cytauxzoon was de-tected using two newly designed set of primers for nested PCR (BecA-Hub, International Livestock Research Institute, Kenya). Primary amplification was performed by forward primer, IL0-9029 (5-AACCTGGTTGATCTTG CCAG-3) and reverse primer, ILO-9030 (5-GAACCTGCG GAAG GATC ATTA-3). Secondary amplification utilized forward primer ILO-MWG 4/70 (5-TGGTTGATCTTGCCAGTA-3) and reverse primer ILO-7782 (5-GAACCTGCG GAAG GATC ATTA-3) (Paper V).
3.8.3 Sequencing and phylogenetic analysis PCR products from amplified fragment of Cyt-b gene and 18SS rRNA gene were extracted from their respective agarose gels and isolates purified by Gene Elute PCR clean-up kit (Sigma, St. Louis, Missouri, USA) as per manufacturers protocol. Purified genomic DNA isolates with > 10 pM of DNA concentrations were directly sequenced by BigDye® Terminator v3.1 (Applied Biosystems) (Paper V). Sequences from the BLAST and those indentified in this study were aligned by Clustal X 2.0 software and phylogenetic tree constructs created from the aligned sequences. The phylogenetic tree construction was determined by the bootstrap neighbor-joining (NJ) Tree method using Kimura-2 parameter, maximum likelihood and tamura 3-parameter with 1000 bootstrap iterations.
3.9 Statistical Analysis The parasites infection rate differences between the NHPs species, age groups, sexes and sampling locations was assessed using the Pearson Chi-square test ( 2). Statistical difference P < 0.05% was regarded as being sig-nificant. SPSS version 9.0 (Chicago, IL, USA) for windows was used for data processing.

24
3.9 Photography Parasite photographs were taken at x100 objectives using a Leitz Autholux 11TM camera system fitted onto a BMX 50 microscope. Colour prints (ASA size 200) of NHPs were taken using Nikon F30111TM (Nikon, Japan) cam-era mounted on BMX 50 microscope.

25
CHAPTER 4: RESULTS
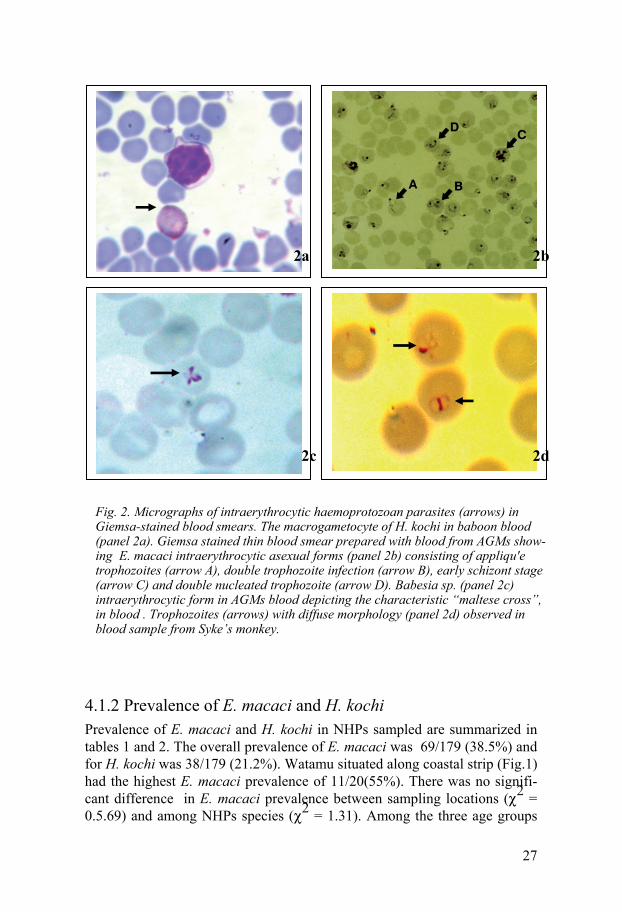
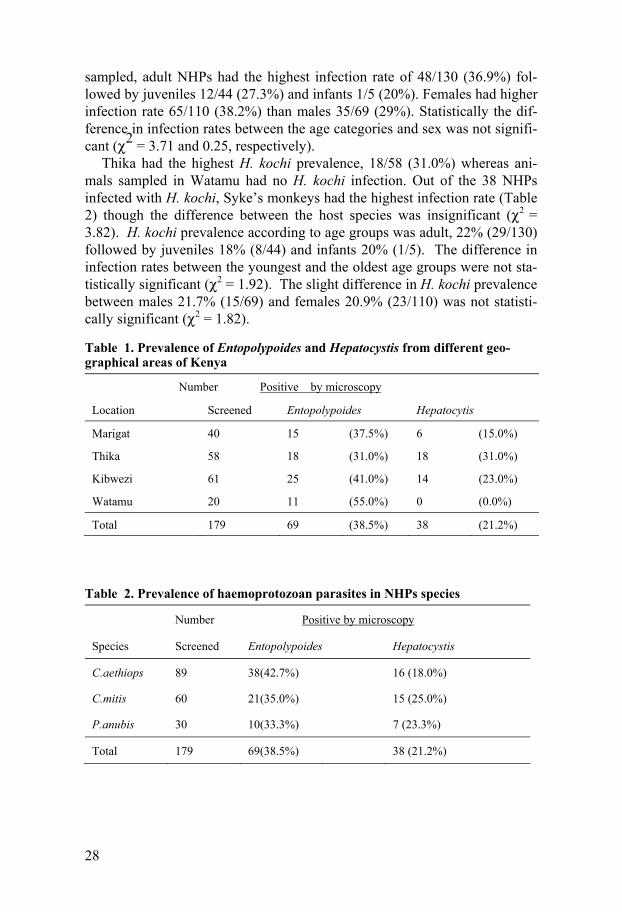
4.1 Parasitological results 4.1.1 Active infections Microscopic evaluation of Giemsa-stained blood smears (BS) detected ga-metocytes stages of H. kochi and intraerythrocytic asexual stages of E. ma-caci and B. microti in blood samples from AGMs, baboons and Syke’s mon-keys (Fig. 2). In this study, microscopic distinction between E. macaci and B. microti was possible only in cases were parasitemia was over 0.5% and the asexual stages were large in size. In addition, definitive characteristics such as the formation of “maltese cross” by Babesia species, E. macaci ap-pliqué trophozoite features and brown malaria parasite pigment inside the infected cells were important morphological features used for identification.
H. kochi macrogametocytes (female) measured 6 – 8 m in diameter. The cytoplasm of macrogametocyte cytoplasm stained brownish yellow with pink nucleus lying in an unstained zone in the cytoplasm (Fig. 2a). Micro-gametocytes measured 5 – 7 m in diameter. Its cytoplasm stained brownish yellow and they have a large pink stained nuclei peripherally located.
E. macaci trophozoite stages of various morphological features (Fig. 2b) were detected. Some of the morphological features displayed by the in-traerythrocytic stages were exclusive to each host species (Study III). Small rings measuring between 1 - 1.5 m with thin blue cytoplasmic rings at-tached to one or two red accessory chromatin dot enclosing a vacuole were frequently observed. Blood smears from AGMs with > 1% parasitemia, dis-played trophozoites with thick blue cytoplasmic rings and red chromatin dot inside the ring giving a bird’s eye view. Horse shoe- shaped trophozoites with double nucleus were also common. AGMs with parasitemia above 2% displayed mixed stage infection composed of small rings, trophozoites and early shizont stages (Fig. 2b). Accole’ forms on the surface of erythrocytes are markedly conical with the nucleus at the pointed end (Fig. 2b). This ac-cole’ shape is different from those on baboon’s erythrocytes which appear as pale-blue vesicles giving a smooth blue bump (Study III). Also noted in some of the infected AGMs with up to 2% parasitemia were the banded (striated) trophozoites which resembled the band forms of Plasmodium ma-larie (Study III). Typical large rings (2 – 3.8 m in diameter) with a small

26
red chromatin dot occupying nearly a quarter of the cell were observed in the infected RBCs of Syke’s monkeys (Study III).
B. microti-like intraerythrocytic trophozoites displaying various morpho-logical features were observed in thin blood smears prepared from baboons and AGMs blood (Study IV). Characteristic Babesia trophozoites arranged as tetrad, “Maltese cross” (Fig. 2c) and others appered as thin small rings with thin cytoplasmic membrane and a very small red stained nucleus were observed.
27
4.1.2 Prevalence of E. macaci and H. kochi Prevalence of E. macaci and H. kochi in NHPs sampled are summarized in tables 1 and 2. The overall prevalence of E. macaci was 69/179 (38.5%) and for H. kochi was 38/179 (21.2%). Watamu situated along coastal strip (Fig.1) had the highest E. macaci prevalence of 11/20(55%). There was no signifi-cant difference in E. macaci prevalence between sampling locations ( 2 = 0.5.69) and among NHPs species ( 2 = 1.31). Among the three age groups
Fig. 2. Micrographs of intraerythrocytic haemoprotozoan parasites (arrows) in Giemsa-stained blood smears. The macrogametocyte of H. kochi in baboon blood (panel 2a). Giemsa stained thin blood smear prepared with blood from AGMs show-ing E. macaci intraerythrocytic asexual forms (panel 2b) consisting of appliqu'e trophozoites (arrow A), double trophozoite infection (arrow B), early schizont stage (arrow C) and double nucleated trophozoite (arrow D). Babesia sp. (panel 2c) intraerythrocytic form in AGMs blood depicting the characteristic “maltese cross”, in blood . Trophozoites (arrows) with diffuse morphology (panel 2d) observed in blood sample from Syke’s monkey.
2a 2b
2c 2d
28
sampled, adult NHPs had the highest infection rate of 48/130 (36.9%) fol-lowed by juveniles 12/44 (27.3%) and infants 1/5 (20%). Females had higher infection rate 65/110 (38.2%) than males 35/69 (29%). Statistically the dif-ference in infection rates between the age categories and sex was not signifi-cant ( 2 = 3.71 and 0.25, respectively).
Thika had the highest H. kochi prevalence, 18/58 (31.0%) whereas ani-mals sampled in Watamu had no H. kochi infection. Out of the 38 NHPs infected with H. kochi, Syke’s monkeys had the highest infection rate (Table 2) though the difference between the host species was insignificant ( 2 = 3.82). H. kochi prevalence according to age groups was adult, 22% (29/130) followed by juveniles 18% (8/44) and infants 20% (1/5). The difference in infection rates between the youngest and the oldest age groups were not sta-tistically significant ( 2 = 1.92). The slight difference in H. kochi prevalence between males 21.7% (15/69) and females 20.9% (23/110) was not statisti-cally significant ( 2 = 1.82).
Table 1. Prevalence of Entopolypoides and Hepatocystis from different geo-graphical areas of Kenya
Number Positive by microscopy
Location Screened Entopolypoides Hepatocytis
Marigat 40 15 (37.5%) 6 (15.0%)
Thika 58 18 (31.0%) 18 (31.0%)
Kibwezi 61 25 (41.0%) 14 (23.0%)
Watamu 20 11 (55.0%) 0 (0.0%)
Total 179 69 (38.5%) 38 (21.2%)
Table 2. Prevalence of haemoprotozoan parasites in NHPs species
Number Positive by microscopy
Species Screened Entopolypoides Hepatocystis
C.aethiops 89 38(42.7%) 16 (18.0%)
C.mitis 60 21(35.0%) 15 (25.0%)
P.anubis 30 10(33.3%) 7 (23.3%)
Total 179 69(38.5%) 38 (21.2%)

29
4.2. Immunological results 4.2.1 Serological detection and characterization of piroplasm infecting NHPs Sera from thirty NHPs sampled were screened for antibodies against E. macaci, B. bovis, and B. bigemina whole parasite antigen by IFAT assay (Study III). Seven sera samples from animals positive for Entopolypoides by microscopy reacted positively at titre of 1:625 to the E. macaci antigen. Five samples which were negative for Entopolypoides using microscopy (three from Sykes’ monkeys, one each from AGM and baboon) reacted positively to E. macaci antigen. None of the sera samples reacted with either B. Bigemina or B. Bovis antigens. Three sera samples from baboons which were positive for E. macaci by microscopy and IFAT, showed a weak positive reaction to B. bigemina antigen at 1:25 antibody titer, which was regarded as inconclusive. However, the same sera did not react with B. bovis antigens.
4.3. Seroprevalence of T. brucei Ab-ELISA and LAT methods revealed presence of both anti-T. brucei anti-bodies and T. brucei antigens respectively in 121 sera samples analyzed (Study I). Indirect Ab-ELISA revealed positive titres of antibodies against T. brucei among the NHPs species from all the sampling locations (Table 3). The prevalence of trypanosomiasis as determined by Ab-ELISA was 9% (11/121). Sykes’ monkeys caught in Watamu, Coast province had the high-est anti-T. brucei antibodies prevalence of 15% followed by AGMs, 7.4% and baboons 4.1% (Table 3). Circulating T. brucei antigen was detected in 6.6% (8/121) of sera tested using LAT (Fig. 3). All sera samples positive by LAT were sero-positive by Ab-ELISA (Table 3). However, these animals did not show any clinical signs of morbidity.

30
Fig. 3. A special plastic test card with pre-marked areas for agglutination tests. LAT performed on 10 serum samples. Samples 1 to 3 are strongly positive, sample 4 is weakly positive, and samples 5 to 10 are negative.
Table3. Prevalence of Trypanosoma brucei in monkey species as determined by serological techniques
Geographical C. aethiops C. mitis P. anubis Location No. positive by positive by positive by Examined LAT Ab-ELISA LAT Ab-ELISA LAT Ab-ELISA Marigat 29 1 (3.4%) 2 (6.9%) nc nc nc nc Thika 49 2 (4.1%) 2 (4.1%) 2 (4.1%) 2 (4.1%) 2 (4.1%) 2 (4.1%) Watamu 20 nc nc 3 (15%) 3 (15%) nc nc Kibwezi 23 0 (0%) 2 (8.7%) nc nc nc nc Total 121
Key; nc-not captured, LAT- latex agglutination test, Ab-ELISA-enzyme-linked antibody assay.
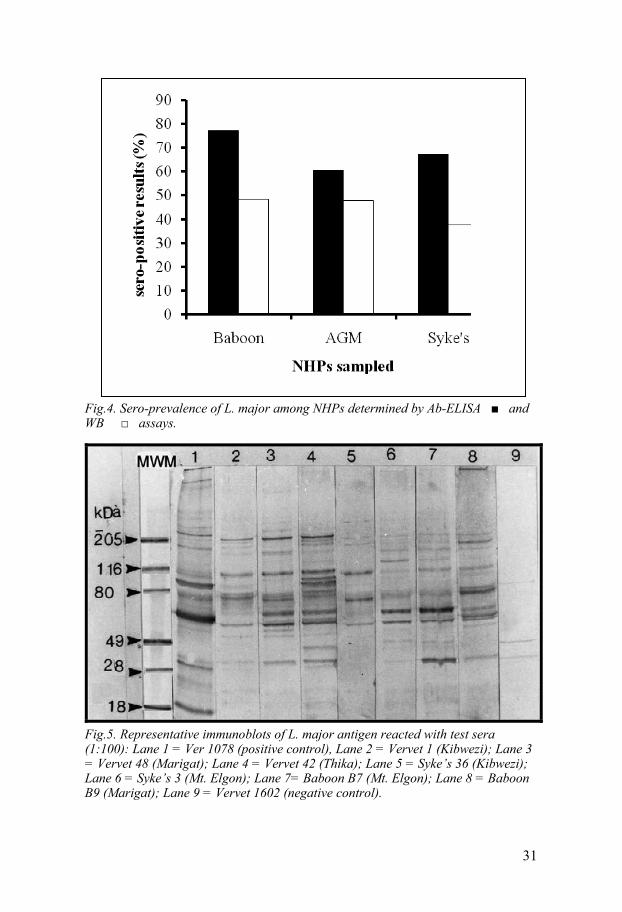
4.4. L. major seroprevalence Of the 378 NHPs serum samples tested in Ab-ELISA for anti-L. major anti-bodies, 250 (66.1%) were positive for antibodies against L. major (Study III). As determined by Ab-ELISA, baboons (Fig 4) had the highest seropre-valence for circulating anti-Leishmania antibodies, (77.2%), statistically different ( 2 = 0.04) from AGMs (60.6%). There was no statistical differ-ence ( 2 = 0.63) in positive frequencies of anti-Leishmania antibodies in the three NHPs species sampled using WB assay (Table 2). Also, results from WB assays showed baboons (48.5%) and AGMs (47.8%) to have similar prevalence of anti-L. major antibodies. Positive control test sera recognized prominent bands observed at 18, 28, 63, 80, 116 and 205 kD. Sero-positive sera reacted with specific L. major antigenic bands at 28, 80,116 and 205 kD (Fig. 5). Anti-L. major antibodies seroprevalence rates by Ab-ELISA were Coast (60%), Eastern (64.3%), Rift valley (63.8%), Central (64.8%) and Western (82.5%). L. major seroprevalence rates as determined by WB analysis of sera from the total number of animals sampled from each prov-ince were 70% in Western, 54.4% in Rift valley, 50% in Coast, 47.4% in Eastern and 31.5% in Central Province.
31
Fig.4. Sero-prevalence of L. major among NHPs determined by Ab-ELISA and WB assays.
Fig.5. Representative immunoblots of L. major antigen reacted with test sera (1:100): Lane 1 = Ver 1078 (positive control), Lane 2 = Vervet 1 (Kibwezi); Lane 3 = Vervet 48 (Marigat); Lane 4 = Vervet 42 (Thika); Lane 5 = Syke’s 36 (Kibwezi); Lane 6 = Syke’s 3 (Mt. Elgon); Lane 7= Baboon B7 (Mt. Elgon); Lane 8 = Baboon B9 (Marigat); Lane 9 = Vervet 1602 (negative control).

32
4.5. Leishmania recall lymphocyte proliferation assay Comparison of results from Ab-ELISA, WB and proliferation assay are summarized in table 4, (Study III). Using the more specific PBMC prolife-ration assay, the high positive frequency observed in animals from Marigat, 42.3% compared to samples from Kibwezi was statistically significant ( 2 = 0.01).
Table 4. Prevalence of anti-L. major antibodies and cell mediated immune re-sponse in wild-caught vervet monkeys from Marigat and Kibwezi
Sampling site
Ab-ELISA (% +Ve)
Western blot (% +Ve)
Proliferation assay (% +Ve)
Marigat Kibwezi
21/26 (80.1%) 19/31 (61.3%)
19/26 (73.1%) 12/31 (38.7 %)
11/26 (42.3%) 6/31 (19.4%)
4.8 Molecular detection and sequence analysis Two molecular surveys utilizing PCR detection techniques were carried out for the purpose of detecting and determining the distribution of piroplasm, plasmodia and genetically closely related parasites such as E. macaci, H. kochi, and Theileria spp.
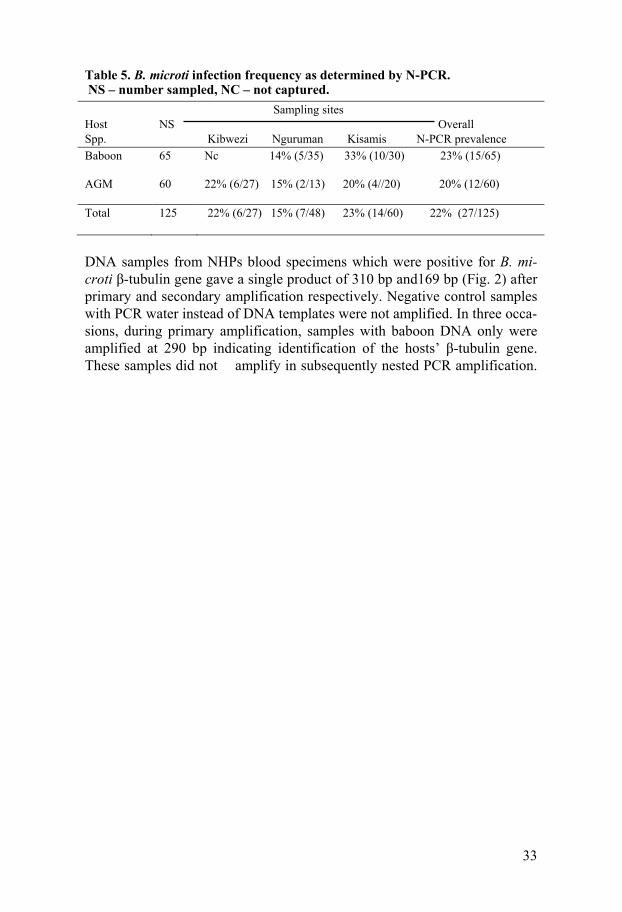
4.8.1 Molecular survey of B. microti Nested PCR tests indicated 22% (27/125) B. microti infection frequency in Kenyan wild caught NHPs (Table 5, study IV). In regard to gender preva-lence, 25% (11/44) female baboons were positive for B. microti genomic DNA compared to 19% (4/21) in male baboons. Unlike in baboons, more males 21% (5/24) than female 19% (7/36) AGMs were positive for B. micro-ti. Overall there was no significant difference in B. microti infection rates between baboons and AGMs ( 2 = 0.655) or between males and females ( 2 = 0.744). The highest infection frequency of 33% was recorded in baboons from Kisamis location. Also, this location had the highest prevalence of 23% (14/60). The difference in Babesia prevalence between Kibwezi 22% (6/27) and Nguruman 15% (7/48) was statistically significant ( 2 = 0.02) but not between Kibwezi and Kisamis 23% (14/60) ( 2 = 0.06).
33
Table 5. B. microti infection frequency as determined by N-PCR. NS – number sampled, NC – not captured.
Host Spp.
NS
Sampling sites Overall Kibwezi Nguruman Kisamis N-PCR prevalence
Baboon 65 Nc 14% (5/35) 33% (10/30) 23% (15/65)
AGM 60 22% (6/27) 15% (2/13) 20% (4//20) 20% (12/60)
Total 125 22% (6/27) 15% (7/48) 23% (14/60) 22% (27/125)
DNA samples from NHPs blood specimens which were positive for B. mi-croti -tubulin gene gave a single product of 310 bp and169 bp (Fig. 2) after primary and secondary amplification respectively. Negative control samples with PCR water instead of DNA templates were not amplified. In three occa-sions, during primary amplification, samples with baboon DNA only were amplified at 290 bp indicating identification of the hosts’ -tubulin gene. These samples did not amplify in subsequently nested PCR amplification.

34
Fig. 6. Gel electrophoresis separation of primary (lanes 2-8) and secondary (lanes 9-15) PCR products of a fragment of -tubulin gene. Lanes 1, 100-bp ladder. Lane 2-3, parasite DNA sample from baboons. Lane 4-5, parasite DNA samples from AGM. Lane 6, positive B. microti DNA sample. Lane 7, negative control (PCR water). Lane 8, Host (baboon) DNA sample, Lane 9-10, nested amplification of parasite DNA samples from baboons. Lane 11-12, parasite DNA samples from AGMs. Lane 13, positive control and lane 14, host (baboon) DNA sample.
4.9 Molecular survey and parasite characterization In another molecular survey, Plasmodium and related parasites were sur-veyed using PCR tests which utilized MTCbF0 and MTCbR0 primers target-ing fragment of Cty-b gene, amplified single bands of approximately 950 base pair (bp) (Study V). More PCR tests with primers, ILO-MWG 4/70 and ILO-7782 targeting the 18SSrRNA gene of piroplasms amplified samples at approximately 500 bp and 405 bp mark (Fig. 7).
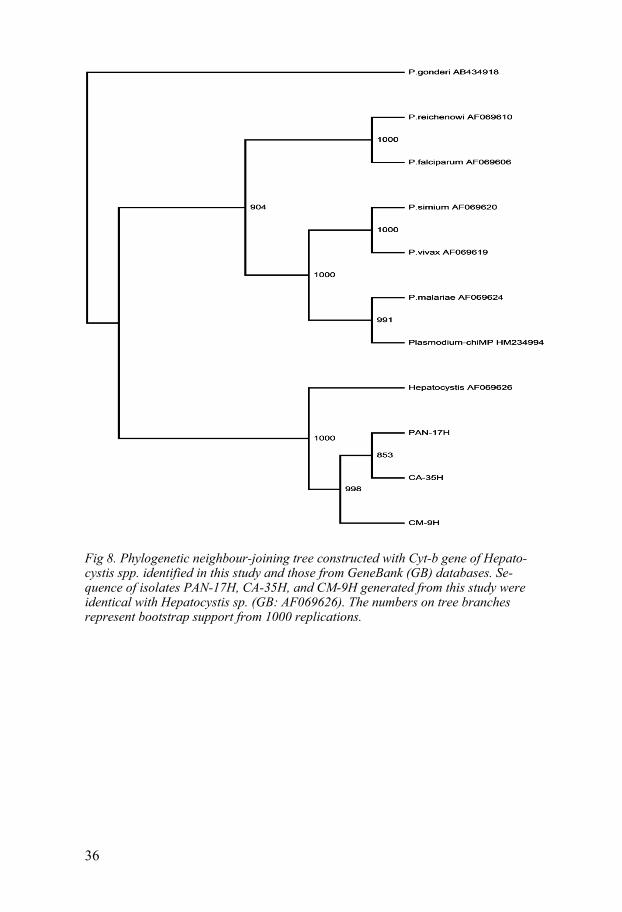
The Cyt-b gene sequences generated in this study were 96.6% identical to Hepatocystis sp. isolated from a baboon, Papio nubensis in Ethiopia (Gen-Bank Accession no. GB: AF069626). None of these sequences identified with Plasmodium spp. Construction of phylogenetic tree inferred from the Cyt-b gene reaffirmed the isolates identity with Hepatocystis sp. (Fig. 8) only. All sequences clustered within the H. kochi lineage supported with a bootstrap value of 1000. The three representative sequences from this study were almost identical with p-distance averaging 0.001 calculated using all pair-wise comparisons (BioEdit Sequence Alignment Editor).
BLAST search from six sequences generated by N-PCR amplification of the 18SS rRNA gene were 95% identical to Babesia sp. KMG 2009a (GB:
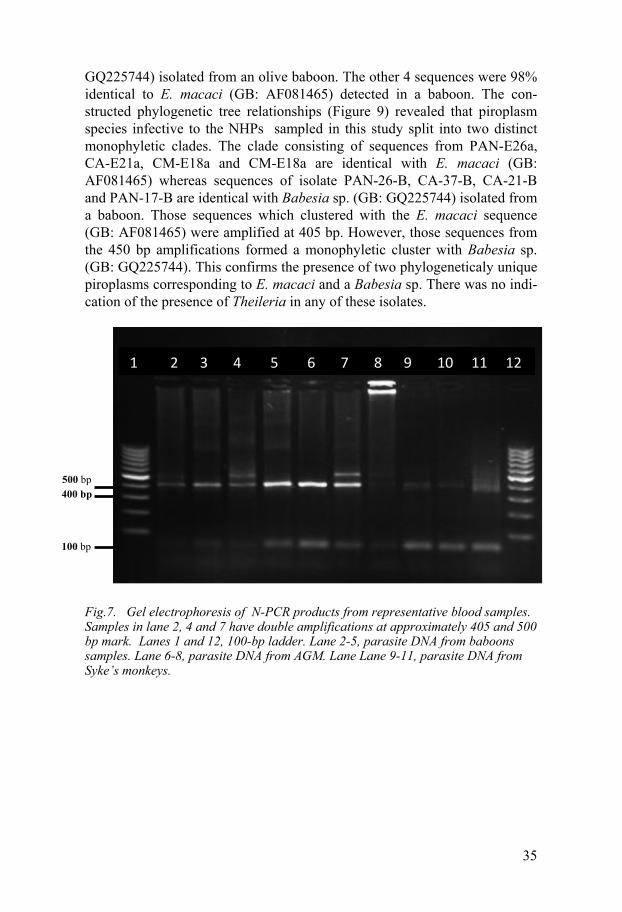
35
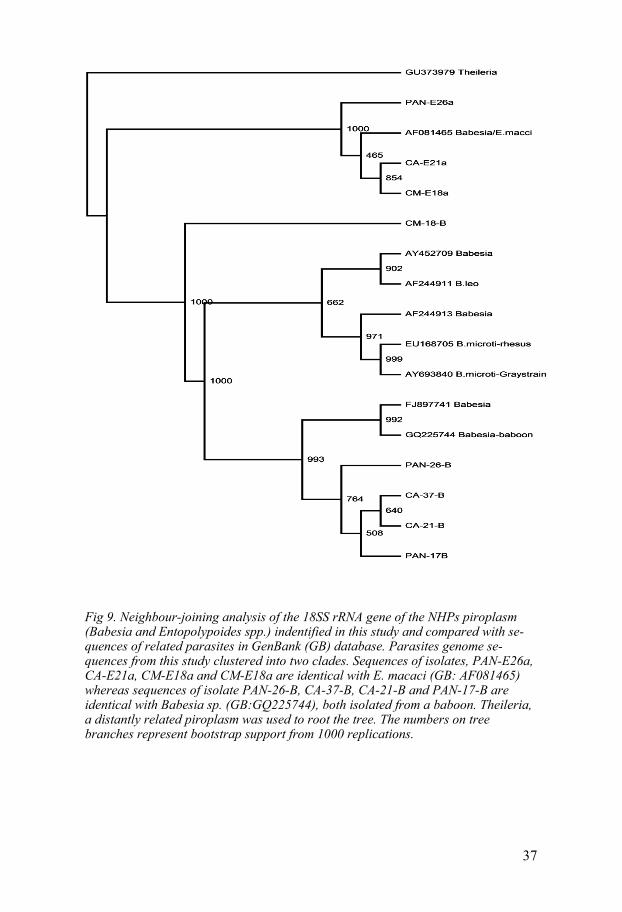
GQ225744) isolated from an olive baboon. The other 4 sequences were 98% identical to E. macaci (GB: AF081465) detected in a baboon. The con-structed phylogenetic tree relationships (Figure 9) revealed that piroplasm species infective to the NHPs sampled in this study split into two distinct monophyletic clades. The clade consisting of sequences from PAN-E26a, CA-E21a, CM-E18a and CM-E18a are identical with E. macaci (GB: AF081465) whereas sequences of isolate PAN-26-B, CA-37-B, CA-21-B and PAN-17-B are identical with Babesia sp. (GB: GQ225744) isolated from a baboon. Those sequences which clustered with the E. macaci sequence (GB: AF081465) were amplified at 405 bp. However, those sequences from the 450 bp amplifications formed a monophyletic cluster with Babesia sp. (GB: GQ225744). This confirms the presence of two phylogeneticaly unique piroplasms corresponding to E. macaci and a Babesia sp. There was no indi-cation of the presence of Theileria in any of these isolates.
Fig.7. Gel electrophoresis of N-PCR products from representative blood samples. Samples in lane 2, 4 and 7 have double amplifications at approximately 405 and 500 bp mark. Lanes 1 and 12, 100-bp ladder. Lane 2-5, parasite DNA from baboons samples. Lane 6-8, parasite DNA from AGM. Lane Lane 9-11, parasite DNA from Syke’s monkeys.
1 2 3 4 5 6 7 8 9 10 11 12
100 bp
500 bp 400 bp
36
Fig 8. Phylogenetic neighbour-joining tree constructed with Cyt-b gene of Hepato-cystis spp. identified in this study and those from GeneBank (GB) databases. Se-quence of isolates PAN-17H, CA-35H, and CM-9H generated from this study were identical with Hepatocystis sp. (GB: AF069626). The numbers on tree branches represent bootstrap support from 1000 replications.
37
Fig 9. Neighbour-joining analysis of the 18SS rRNA gene of the NHPs piroplasm (Babesia and Entopolypoides spp.) indentified in this study and compared with se-quences of related parasites in GenBank (GB) database. Parasites genome se-quences from this study clustered into two clades. Sequences of isolates, PAN-E26a, CA-E21a, CM-E18a and CM-E18a are identical with E. macaci (GB: AF081465) whereas sequences of isolate PAN-26-B, CA-37-B, CA-21-B and PAN-17-B are identical with Babesia sp. (GB:GQ225744), both isolated from a baboon. Theileria, a distantly related piroplasm was used to root the tree. The numbers on tree branches represent bootstrap support from 1000 replications.

38
5.0 Discussion
In this study, sero-prevalence surveys revealed that wild free-ranging ba-boons, AGMs and Syke’s monkeys are naturally exposed to T. brucei and L. major (Study I and II). The NHPs were also found to be naturaly infected with H. kochi, E. macaci and Babesia sp (Study III and IV). Active infec-tions by H. kochi, E. macaci and Babesia were mostly chronic, characterized by low parasite numbers and frequent mixed infections (Study V). Detection of chronic Babesia infection and confirmation of mixed parasite infections in Kenyan wild-caught monkeys is significant information to the conservation and biomedical utilization of these animals. In addition, the assessment of haemoprotozoan parasites of medical and economic importance such as Plasmodium, Babesia, Trypanosoma and Leishmania in wild NHPs found in habitats close to human settlements is of importance in the control and eradi-cation of probable human zoonotic pathogens.
5.1 T. brucei prevalence Active Trypanosoma infection was not demonstrated in sampled NHPs (Pa-per I). There are several reasons suggested for the absence of active infec-tions in Kenyan NHPs. Self-cure has been observed in AGMs after experi-mental infection with T. b. rhodesience and persistence of antigens after curative treatment demonstrated (Gichuki et al., 1994). This attributes ex-plains the probable reason to the presence of anti-T. brucei antibodies and T. brucei antigens with absence of active infection (Chappuis et al., 2005). Another reason is the presence of trypanocidal factors in the serum of NHPs such as the baboon, which would lyse infecting trypanosomes before a de-monstrable parasitemia develops (Baker, 1972; Poltera, 1985) and interfere with the tissue invasive nature of the infecting parasites, especially those of T. brucei group (Gichuki et al., 1994; Molyneux and Ashford, 1983).
Although active trypanosome infections could not be demonstrated in this study, Ab-ELISA assays revealed 9% (11/121) prevalence whereas LAT detected parasite antigenemia in 6.6 % (8/121) of the samples. This indicates natural exposure to T. brucei in the primate species sampled. The Ab-ELISA used in this study has been used widely to study T. brucei infection in do-mestic animals and humans (Van Knappen et al., 1977) and disease progres-sion in NHPs (Gichuki et al., 1994). The field oriented LAT test has been

39
shown to have sensitivity of 98.3% in cattle and 100% in sheep and goats. Specificity was 97.2% in cattle, 99% in sheep and 100% in goats (Kayang et al., 1997). In this study, Syke’s monkeys caught in Watamu, in Coast Prov-ince had the highest trypanosomiasis prevalence of 15% followed by AGMs from Kibwezi in Eastern Province, 7.4% and baboons from Thika in Central Province, 4.1% and AGMs, 4.1%. These results are comparable to trypano-somosis seroprevalence in cattle from Coastal, 15.6%, Rift valley 12.9% and Western 8.3% regions of Kenya (Bourn et al., 2001). A more recent epide-miological study (Thumbi et al., 2010) of cattle trypanosomiasis in different parts Western Province of Kenya reported T. brucei prevalence ranging from 8-17% as determined by PCR tests. Previous sero-epidemiological and recent molecular studies of human trypanosomiasis in Kenya from 1977 to 2007 show variable prevalence in different parts of Western Province rang-ing from 0.3% to 53.2% and a median value of 2.1% (Bourn et al., 2001; Thumbi et al., 2010).
Cases of natural infections (Ziccardi et al., 2000) and experimental trypa-nosome infections of Neotropical NHPs from South America have been do-cumented (Weinman et al., 1978; Ziccardi andLourenco-de-Oliveira, 1997) but, AGMs from East Africa are only known to be experimentally suscepti-ble to T. b. rhodesiense (Poltera, 1985; Pearson et al., 1986). Theoretically some East African NHPs are perceived to be reservoir hosts of trypanosomes (Fiennes, 1967; Kuntz, 1973) although this is yet to be demonstrated.
5.2. L. major seroprevalence Precense of active Leishmania infection was not detected in sampled NHPs (Paper II), unlike in the previous study where a single AGM from Thika District was diagnosed with an active infection through in vitro culture of biopsy material taken from a suspicious ulceration on the forehead (Binha-zim et al., 1987). In this study, conducted surveys did not detect any suspi-cious ulceration on any sampled NHPs to warrant biopsy and in vitro culture as was the case in the previous study (Binhazim et al., 1987). The failure to take skin or spleen biopsy could have contributed to the failure to detect active Leishmania infection in this study. According to Baker, (1972), Leishmania group of parasites are mostly tissue invasive in nature and re-quires the use of in vitro culture with biopsy materials taken from skin or spleen (Berman, 1997). WB assays revealed 46.3% (175/378) L. major prevalence compared to 66.1% (250/378) by Ab-ELISA assay. In addition, WB tests showed baboons and AGMs to have comparable frequency of anti-L. major antibodies (48.5% and 47.8%) respectively while Sykes’ (37.5%) had lower antibodies frequency. This is contrary to Ab-ELISA tests which showed baboons and Sykes’ monkeys having higher frequency of anti-L. major antibodies than AGMs. This discrepancy could be attributed to using

40
crude L. major antigen which reacts with antibodies from several protozoan parasites (Harrith et al., 1987). Compared to ELISA using crude antigen, WB has been shown to be more specific (Isaza et al., 1997; Sakru et al., 2007). Also, the high seroprevalence of leishmaniasis in baboons and AGMs as revealed by WB analysis might be attributed to the high permissiveness of these NHPs to Leishmania parasites as compared to Sykes’ monkey (Githure et al., 1986; Binhazim et al., 1993).
Prevalence of L. major (82%) around Mt. Elgon in Western Province was significantly high ( 2 = 0.012) compared to Thika, 34.3%, and Kibwezi 70%. This is not surprising since Mt. Elgon region is known to sustain an active focus of L. major transmission (Mutinga, 1975; Muigai et al., 1987). In Marigat and Tot, Rift valley province visceral leishmaniasis (VL) caused by L. donovani is known to be co-endemic with cutaneous leishmaniasis (Mui-gai et al., 1987; Perkin et al., 1988). Since the WB assay used in this study could not differentiate between the two Leismania species, the high anti-L. major sero-prevalence in vervet monkeys, 52.9% in Marigat and 62.1% in Tot could be due to exposure by either or both parasite species. These sites are also known to have high human VL prevalence of up to 52.5% (Ryan et al., 2006). Positive results from recall proliferation assay ascertained the exposure of AGMs to L. major (Paper II). Together with the absence of skin lesions, the results can be interpreted as either these monkeys were subclini-cally infected or infected and self-cured. A previous study on experimentally infected NHPs had shown that lymphocytes from asymptomatic and self-cured animals had high proliferation index as compared to naïve control animals (Gicheru et al., 1997).
L. major prevalence of 46.3% was significantly higher ( 2 = 0.075) com-pared to T. brucei prevalence of 6.6% as determined by WB. The low T. brucei prevalence could be attributed to the continuous concerted effort by the Kenyan government and its partners (WHO) to eradicate both human and animal trypanosomiasis (Rutto and Karuga, 2009). These efforts are not replicated in leishmaniasis control because the disease is regarded as neg-lected (WHO, 2007). In addition, trypanosomiasis in Kenya is mostly found in the Western regions of the country (Rutto and Karuga, 2009) while leish-maniasis is widely dispersed in small foci (Mutinga et al., 1990).
5.3 Detection and distribution of E. macaci and H. kochi Microscopic evaluation of Giemsa-stained blood smears detected single in-fections by E. macaci and H. kochi (Study III and IV). Failure to detect mixed parasite infections these wild-caught Kenyan NHPs is consistent with the results of previous epidemiological studies (Kuntz and Moore, 1973; Turner et al. 1982). According to Moore and Kuntz, (1975), E. macaci and H. kochi infections have been shown to occur in same populations of Papio

41
cynocephalus in Kenya, but not in the same host. Failure to detect mixed infections could be attributed to the solo use of microscopic evaluation of Giemsa-stained blood samples. This method has been shown to face sensitiv-ity and specificity challenges when detecting piroplasms or malaria parasites in chronic infections characterized by low parasitemia and mixed infections (Persing et al., 1992; Amiri, 2010).
Prevalence of E. macaci as determined by microscopy in NHPs sampled show high overall infection rate of 38.5% (Study III) compared to earlier studies which reported a 12% infection rate in Kenyan baboons (Moore and Kuntz, 1975). This variation could be due to the difference in sample size or the localities in which the animals were captured and sampled. Sampling by Moore and Kuntz was concentrated in Naivasha in central highlands of Kenya. In contrast, the present sampling was done in several regions experi-encing different climatic conditions. Also, our sampling was conducted after the rainy season (January-March) which could be sequel to increased vector population and activity on the abundant vegetations.
H. kochi prevalence of 21% as determined by microscopy in all three NHPs species sampled. Leathers, (1978) reported a 29% infection rate with H. ko-chi in 249 AGMs. This was higher than infection prevalence observed in AGMs (18%) sampled in this study. The discrepancy could be attributed to sampling locations which were different in each study. Prevalence of H. kochi has been shown to be as high as 40% in NHPs from west central Afri-ca and from central East Africa (Voller, 1972; Loeb, 1978). Although H. kochi infection seemed slightly more frequent in males (21.7%) than females (20.9%), these results were not statistically significant ( 2 = 1.92). These results conform to a more general picture in which the occurrence of this parasite was equal between the sexes of NHPs (Leathers, 1978).
5.4 Molecular detection, prevalence and characterization of haemoparasites In this study, molecular findings revealed single and mixed haemoprotozoan parasite infections composed of Hepatocystis, Entopolypoides and Babesia species in wild-caught baboons, AGMs and Syke’s monkeys in Kenya. As-sessment of haemoprotozoan parasites of economic importance such as ma-laria parasites or Babesia species in wild NHPs found close to human set-tlements in Kenya is of importance. Our results on detection of mixed infec-tion are not in agreement with the previous studies which detected single infections by either Hepatocystis (Turner et al., 1982: Phillips-Conroy et al., 1988) or E. macaci in wild-caught NHPs from East Africa (Kuntz and Moore, 1975: Jeneby et al., 2008). The piroplasm, E. macaci infections have

42
been shown to occur in different populations of Papio cynocephalus in Kenya, but not in the same host together with H. kochi (Moore and Kuntz, 1975). Unlike in previous surveys, the use of multiplex PCR primers capable of detecting closely related parasite species contributed to the detection of mixed infection in this study. The diagnoses of haemoparasites in the earlier studies were based on microscopic examination of Giemsa-stained blood smears. This method faces sensitivity and specificity challenges when de-tecting piroplasms or malaria parasites in chronic infections characterized by low parasitemia and mixed infections (de Vos et al., 2000; Amiri, 2010).
Overall H. kochi prevalence of 61% as determined by PCR was approx-imately two-fold higher than 33% by E. macaci and 27% by Babesia. Mi-croscopy detected 51% H. kochi infection, double the 24% piroplasm infec-tions. The difference in the parasites infection rates could be attributed to the diference in animal numbers trapped and sampled from the different locations. In addition, specific vector density in the different ecological zones where the animals were trapped and sampled may influence chances of a specific haemoparasite infection, suggesting transmission in circum-scribed foci. PCR results detected variable mixed infections composed of 17% infection by H. kochi and E. macaci, 12% by H. kochi and Babesia, and 7% by E. macaci and B. microti. There was also 2 % multiple infections by B. microti, E. macaci and H. kochi detected by PCR techniques. PCR data on mixed infection suggests that Hepatocystis, a Plasmodium-like natu-ral parasite of NHPs has a marked suppressive effects on Entopolypoides and Babesia species infecting NHPs. Samples which were positive for Hepato-cystis as determined by microscopy were subsequently found to be co-infected with either Babesia or Entopolypoides as detected by PCR. Howev-er, all the samples which were positive for piroplasm trophozoites as eva-luated by microscopy were also PCR positive for either Entopolypoides or Babesia but not for Hepatocystis. This indicates whenever Hepatocystis in-fection occurs, Entopolypoides or Babesia infections is suppressed to sub clinical infections with very low parasitemia which makes microscopic eval-uation of blood smears ineffective. Cross-protection leading to immunologi-cal suppression of a malaria parasite by a piroplasm has been established (Duivenvoorde et al., 2010). A natural B. microti-like infection in rhesus monkey was shown to have marked suppressive effect on subsequent ex-perimental infections by Plasmodium cyanomolgi in the same animal (Voor-berg-van der Wel et al., 2008). To demonstrate cross-protection between Hepatocystis and Entopolypoides or Babesia requires more elaborate immu-nological investigations (Cox, 1978). These investigations will provide valuable information which could be useful for new vaccines development especially in the development of attenuated vaccines for human babesiosis or malaria (Wykes and Good, 2007; Shkap et al., 2007).
The microscopy method detected more H. kochi infections, 51% com-pared to PCR 30% because microscopy was not able to detect co-infections

43
like PCR tets. However, PCR revealed higher (61%) overall H. kochi infec-tions (single plus mixed) compared with the 51% by microscopy. Also, PCR detected more Entopolypoides, 33% and Babesia, 27% compared with 24% piroplasm infection rate diagnosed by microscopy. These results confirms that the molecular detection offer higher sensitivity and specificity than mi-croscopic methods (Snounou et al., 1993: Duarte et al., 2008). Using prim-ers targeting the 18SS rRNA gene and cyt-b gene, more Plasmodium and Hepatocystis cases and multiple infections was detected in Asian macaques monkeys as compared to the microscopic evaluation of blood smears (Duarte et al., 2008: Seethamchai et al., 2008). The 18SS rRNA and cyt-b loci are recommended for epidemiological studies of haemoprotozoan parasites be-cause of their intra-species conservation associated with appropriate inters-pecies variation (Steenkeste et al., 2009).
Phylogenetic analyses inferred from Cty-b sequences from this study con-firmed the presence of Hepatocystis and the absence of Plasmodium infec-tions in baboons AGMs and Syke’s monkeys. Although wild free-ranging baboons harbor parasites closely related to Plasmodium, they are not natural-ly susceptible to Plasmodium in the wild (Garnham, 1966). In this study, phylogenetic sequence analysis indicates that there are two distinct pirop-lasm species circulating in the NHPs populations. Some of the piroplasm sequences identified with Babesia sp. (GB: GQ225744 and FJ897741) earli-er isolated from a baboon blood sample in Ethiopia. Others identified with E. macaci also isolated from baboons (Bronsdon et al., 1999). Our results com-plements a previous study that analyzed the 18SS rRNA gene of a Babesia-like parasite detected in a baboon and concluded that Entopolypoides is closely related to Babesia microti (97.9% sequence similarity) however, they represent different species (Bronsdon et al., 1999). Sequence analysis of Cty-b genes of Hepatocystis showed sequence variation in isolates taken from same NHPs hosts and those from different hosts. There was less sequence heterogeneity in Babesia and Entopolypoides isolates. Analysis of Cty-b gene has consistently shown sequence difference among isolates of Hepato-cystis and Plasmodium. (Seethamchai et al., 2008).
5.4 Conclusion The main conclusions drawn from this study are: The serological assays used in this study provide strong circumstantial evidence that wild-caught AGMs, olive baboons and Syke’s monkeys captured close to human settlements are naturally exposed to T. brucei and L. major infections. These infections could be sub-clinical or cases of infection and self cure as seen in controlled experimental animal studies (Gichuki et al., 1994; Gicheru et al., 1997) and in humans CL (Sakru, et al., 2007). AGMs, olive baboons and Syke’s mon-keys are equally susceptible to T. brucei and L. major. Both T. brucei and L.

44
major co-exist in Marigat, Thika, Watamu and Kibwezi. The evidence of specific immunological involvement towards natural T. brucei and L. major infections as shown by serological assays (Study I and II) indicates that AGMs, olive baboons and Syke’s monkeys are suitable animal models for human trypanosomiasis and leishmaniasis.
The detection and molecular characterization of haemoprotozoan para-sites indicate that there are cases of either single or multiple infections by Hepatocystis, Babesia and Entopolypoides naturally infecting NHPs in Kenya. Molecular diagnosis and monitoring of infectious haemoprotozoa in NHPs living close to human settlements in Africa is a step towards control of probable zoonotic diseases. In Asia, molecular epidemiology was able to detect deadly P. knowlesi infections in human blood samples thought to have been the less lethal human malaria parasite, P. malariae (Cox-Singh et al., 2008). With the close morphological similarities within the phylum Api-complexa, precise diagnosis is important for accurate treatment of a person or a NHP destined for biomedical research (Bronsdon et al., 1999). Infec-tion of NHPs with Babesia may also hold potential for zoonotic transmission to individuals who work closely with these animals (Bronsdon et al., 1999; Wachtman and Mansfield, 2008). Besides zoonotic challenges, reports have shown that chronic B. microti infections suppresses P. cynomolgy experi-mental infection in rhesus monkeys (Wel et al., 2008; Duivenvoorde et al., 2010) posing a significant confounder for biomedical research.
5.4.1 The main conclusions drawn from this study are; 1. Old World NHPs (Baboon, AGMs and Sykes’ monkey) from Kenya
harbor H. kochi, E. macaci and B. microti-like parasites. 2. PCR-based tests confirmed that mixed infection of H. kochi, E. macaci
and B. microti-like parasites occur in NHPs. 3. Molecular survey using seminested multiplex PCR confirmed that wild-
caught Kenyan baboon, AGMs and Sykes’ monkeys are not infected with malaria parasites in the wild.
4. Although active trypanosomosis or leishmaniasis was not established, the presence of anti-T. brucei and anti-L. major antibodies in sera from wild-caught Kenyan baboon, AGMs and Sykes’ monkeys indicated ex-posure.

45
Acknowledgements
This study was part of collaborative research project between Division of comparative medicine (DCM) Uppsala University, and Institute of Primate Research (IPR), National museums of Kenya (NMK).
I salute all staff members in these two institutions who in many ways facili-tated the completion of this work. Specifically I would like to acknowledge the following;
Prof. Svante Winberg for accepting to supervise this study and train in his department.
Prof. Hans-Erik Carlsson for being my initial supervisor, for his initial facilitation of this study, providing guidance and critical assistance in prepa-ration of manuscripts thoughout this study.
Dr. Suleiman Mbaruk for accepting me to work in his department, and his constant guidance in collection of samples from the field, laboratory supervi-sion and preparation of manuscripts.
Dr. Thomas Kariuki, Director IPR, for the encouragement and providing financial solutions for purchasing of reagents and financing thenumerous field excursions.
I am also grateful to Dr. Maina Ngotho, head of animal science department and all field technicians for providing invaluable technical support in regard to the trapping of NHPs and collection of samples.
I wish to express my sincere gratitude to my family, Yvonne, Mariam and Hemed for their constant support and love.

46
Funding
This study was funded in part by grant from the INDEVELOP foundation University of Uppsala and by the Directors’ Research Initiative Fund of In-stitute of Primate Research, Kenya.

47
References
Aiyedun, B. A., A. A. Amodu, D. Bidwell, G. Bone, A. A. Buck, J. Coulm, J. L. Frezil, N. H. Kent, P. Mattern, A. R. Njogu, P. de Raadt, A. Voller, and M. Wery. (1976). Parallel evaluation of serological tests applied in African trypa-nosomiasis: a WHO collaborative study. Bull. W. H. O. 54:141-147.
Altmann, J., Altmann, S.A. and Hausfater, G. (1981). Physical maturation and age estimates of yellow baboons. American Journal of Primatology. 1: 389-399.
Ameri M. (2010). Laboratory diagnosis of malaria in nonhuman primates. Vet Clin Pathol; 39: 5–19.
Anderson, J. F., L. A. Magnarelli, C. S. Donner, A. Spielman, and J. Piesman. (1979). Canine Babesia new to North America. Science 204:1431-1432.
Ashford, R.W., Reid, G.D.F., Wrangham, R.W. (2000). Intestinalparasites of the chimpanzee Pan troglodytes in Kibale forest, Uganda. Ann Trop Med Parasi-tol. 94:–9.
Baker, J.R. (1972). Protozoa of tissue and blood. In R. N. T-W- Fiennes. (Ed.) Pathology of Simian Primates, Part II; Infectious and Paarasitic Diseaeses (Pp. 29-56). New York: S. Karger.
Berman, J.D. (1997). Human leishmaniasis: clinical, diagnostic, and chemotherapeu-tic developments in the last 10 years. Clin Infect Dis 24: 684–703
Binhazim, A.A., Githure, J.I., Reid, G.D.F. and Muchemi, G. R. (1987). Isolation of L. major from a naturally infected vervet monkey (C. aethiops) caught in Kiambu District, Kenya. Journal of Parasitology, 73:1278-9.
Binhazim, A.A., Sung, S.S., Chapman, L.W. Jr and Olobo, J. (1993). Comparative susceptibility of African green monkeys (Cecopithecus aethiops) to experimen-tal infection with Leishmania leishmania donovani and Leishmania leishmania infatum. Laboratory Animal Science, 43: 37-47.
Brack M. (1985). Agents transmissible from simians to man. Berlin. Sprin-ger-Verlag. Pp. 24-89 & 189-206.
Bronsdon M. A., Homer M. J., Magera M. H. J, Harison C, Andrews G. R, Bielitzk J. T. (1999). Detection of enzootic babesiosis in baboons (Papio cyanocepha-lus) and phylogenetic evidence surpporting synonym of the genera Entopoly-poides and Babesia. J clin microbiol, 37:1548-53.
Brown, R.C., Collins, W.E., Warren, McW and Contacos, P.G. (1973). The primate malarias. USA Government Printing Office. Washington DC, U.S.A. (Pp.1-336).
Bruce-Chwatt, L. J. (1985). Essential Malariology. 2nd Edn. William Heinmann Medical Books Ltd. London.
Burridge, (1975). The role of wild mammals in the epidemiology of bovine theil-erioses in East Africa. Journal of Wildlife Diseases. 11: 68-75.
Caccio, S., Camma, C., Onumac, M., Severinia, C. (2000). The -tubulin gene of Babesia and Theileria parasites is an informative marker for species discrimi-nation. International Journal for Parasitology. 30: 1181-1185.

48
Carlsson, H. E., Schapiro,S. J., Farah, I. and Hau, J. (2004). Use of primates in re-search:a global overview. American Journal of Primatology. 63:25-237.
Chappuis, F., Loutan, L., Simarro, P., Lejon, V and Bu¨scher, P. 2005. Review: Options for Field Diagnosis of Human African Trypanosomiasis. Clinical Mi-crobiology. 18:133-146.
Cheesbrough, M. (1987). Medical Laboratory Manual for Tropical Countries, 2nd Edn. Oxford. Butterworth-Heinemann Ltd. (Pp. 164-272)
Chiarelli, A.B. (1973). Taxonomic Atlas of Living Primates. London and NY. Aca-demic Press.
Chisolm, E. S., Sulzer, A. J., and Ruebush II, T. K. (1986). Indirect immunofluores-cence test for human Babesia microti infection: antigenic specificity. Am. J. Trop. Med. Hyg. 35:921–925.Contemp Top Lab Anim Sci. 40(1):41-2.
Coatney, G. R., Collins, W. E., Warren, McW. and Contacos, P. G. (1971). The Primate Malarias. Washington D C: USA Govertent Printing Office. (Pp. 1-336).
Coatney, G.R. (1971). The simian malarias: zoonoses, anthroponoses, or both. American Journal of Medicine and Hygiene, 20: 795-803.
Collins, K. W. (1974). Primate malarias. Advance in vetenary science and compara-tive medicine ,Vol .18 . New York . Academic Press Inc (Pp.1-23).
Collins, W. E.; Contacos, P. G and Richardson, B. B. (1975). Studies on human malaria in Aotus monkeys. Vi. Infectivity of Plasmodium malariae to different Anophilines. Journal of Parsitology. 61: 941-943.
de Vos AJ, Molloy J.B, and Jorgensen W.K. (2000). Bovine babesiosis. In: Office International des Epizooties. (Ed) Manual of recommended diagnostic tech-niques and requirements for biological products, Vol. IV, Pp. 412-422.
Duarte, A.M., Malafronte Rdos S, Cerutti C Jr, Curado I, de Paiva BR, Maeda AY, Yamasaki T, Summa ME, Neves Ddo V, de Oliveira SG, Gomes Ade C. (2008). Natural Plasmodium infections in Brazilian wild monkeys: reservoirs for human infections?. Acta Trop, 107:179-85.
Duivenvoorde L.M, Voorberg-van der Wel A, Werff N.M, Braskamp G, Remarque E J, Kondova I, Kocken C.H.M, Thomas A.W (2010). Suppression of Plasmo-dium cynomolgi in rhesus macaques by co-infection with Babesia microti. In-fection and Immunity, 78: 1032–1039.
Dvoraková , H.M., Dvorácková, M. (2007). Babesiosis, a little known zoonosis. Epidemiol Mikrobiol Imunol. 56:176-80.
Eley, R.M. (1989). Know Your Monkey: A Guide to the Primates of Kenya. Nai-robi, Kenya: Institute of Primate Research, National Museums of Kenya. (Pp. 1-50).
Eley, R.M., Strum, S. C., Muchemi, G. and Reid, G.D.F. (1989). Nutrition, body condition, activity patterns, and parasitism of free- ranging troops of olive ba-boons (Papio anubis) in Kenya. American Journal of Primatology. 18:209-219.
Escalante, A. A., and Ayala, F. J. (1994). Phylogeny of the malarial genus Plasmo-dium, derived from rRNA gene sequences. Proc. Natl. Acad. Sci. USA 91: 11373–11377.
Escalante, A. A., and Ayala, F. J. (1994). Phylogeny of the malarial genus Plasmo-dium, derived from rRNA gene sequences. Proc. Natl. Acad. Sci. USA 91: 11373–11377.
Escalante, A., Freeland, D., Collins, W., and Lal, A. (1998). The evolution of pri-mate malaria parasites based on the gene encoding cytochrome b from the li-near mitochondrial genome. PNAS. 95: 8124-8129.
Escalante, A.A. and Ayala, J. (1995). Evolutionary origin of plasmodium and other Apicomplexa based on Rrna genes. Prac. Nah Acad. Sci. USA. 92: 5793-5797.

49
Etkind, P., J. Piesman, T. K. Ruebush II, A. Spielman, and D. D. Juranek. 1980. Methods for detection of Babesia microti infection in wild rodents. J. Parasitol. 66:107-110.
Fandeur, T., Volney, B., Peneau, C., and de Thoisy, B. (2000).Monkeys of the rain-forest in French Guiana are natural reservoirs for P. brasilianum/P. malariae malaria. Parasitology. 120 ( Pt 1):11-21.
Farah I.O., Kariuki T.M., King C. L., Hau J. (2001). An overview of animal models in experimental schistosomiasis and refinements in the use of non-human pri-mates. Lab Anim 35:-212.
Fiennes, R. (1967). Zoonosis of Primates. Cornell University Press. Great Britain, Part II. (Pp. 69-71).
Figueroa, J.V. and Buening, G.M. (1995). Nucleis acid probes as a diagnostic method for tick-borne hemoparasites of veterinary importance. Vet Parasitol. 57: 75-92.
Flynn., R.J. (1973). Parasites of Laboratory Animals. Lower State University Press. America, (Ia. Pp.3-32).
Garnham, P. (1966). Malaria Parasites and Other Haemosporidia. Oxford: Black-well Scientific Publications.
Gicheru, M.M., Jeneby, M.M., Macharia, J.C., Carlsson, H.E., Sulemana, M.A. 2009. Prevalence of antibodies and cell mediated immune response against Leishmania major in feral nonhuman primates from Kenya. Acta Tropica 109: 136–140.
Gicheru, M.M., Olobo, J.O., Kariuki, T.M. and Adthiambo, C. (1995). Visceral leishmaniasis in vervet monkeys:Immunological responses during asympto-matic infections. Scandinavian Journal of Immunology. 41: 202-208.
Githure J. I., Shatry A.M., Tarara R., Chulay J.D., Suleman M.A., and Chunge C.N. (1986). The suitability of East African primates as animal models for leishma-niasis. Transaction of Royal Society of Tropical Medicine and Hygiene, 80(4): 575-6.
Gozalo, A. and Tantalean, M. (1996). Parasitic protozoa in neotropical primates. Laboratory Primates Newsletter, No3. 35: 1-7.
Gubbels J. M., De Vos A. P., Van Der Weide M., Viseras J., Schouls L. M., De Vries E., and Jongejan F. (1999). Simultaneous detection of bovine theileria and babesia species by reverse line blot hybridization. Journal of Clinical Mi-crobiology, 37: 1782–1789.
Gulland, F.M.D. (1995). Impact of infectious diseases in wild populations, a review. In ecology of infectios diseases in natural populations. Grenfell, B.T and A.P, Dobson(Eds). Cambridge University Press.
Haltenorth, T. and Diller, H. (1977). Mammals of Africa, a field guide. William Collins and Sons Ltd. London. Pp 199-207.
Hanscheid T. Current strategies to avoid misdiagnosis of malaria. Clin Microbiol Infect. 2003;9:497–504.
Hau J, Van Hoosier G. L. (2003). Handbook of Laboratory Animal Science: Animal Models, Volume III. Pp. 169-249.javascript:void(0)
Herder S, Simo G, Nkinin S, Njiokou F. (2002). Identification of trypanosomes in wild animals from southern Cameroon using the polymerase chain reaction (PCR). Parasite. 4:345-9.
Homer, M. J., I. Aguilar-Delfin, S. R. Telford Iii, P. J. Krause, And D. H. Persing. Babesiosis. 2000. Clinical Microbiology Reviews. 13: 451–469.
Jeneby M. Maamun, Mbaruk A. Suleman, Mercy Akinyi, Hastings Ozwara, Thomas Kariuki and Hans-Erik Carlsson. (2011). Prevalence of Babesia mi-

50
croti in free-ranging Kenyan baboons and African green monkeys. Journal of Parasitology 97:63-67.
Jeneby MM, Ngeiywa M, Yole D.S, Mwenda J.M, Suleman M.A, Carlson H.E. (2008). Enzootic simian piroplasm (Entopolypoides macaci ) in wild-caught Kenyan non-human primates. J Med Primatol. 37:329-36.
Kayang, B.B., Bosompem, K.M., Assoku, R.K., Awumbila, B. (1997). Detection of Trypanosoma brucei, T. congolense and T. vivax infections in cattle, sheep and goats using latex agglutination. Int J Parasitol. 27:83-7.
Kingdon, J. (1971). East African Mammals, Vol(I). The University of Chicago Press. Pp.177-182, 213-15.
Kumar, R., Bumb, R.A., Ansari, N.A., Mehta, R.D., Salotra, P. (2007). Cutaneous leishmaniasis caused by Leishmania tropica in Bikaner, India: parasite identi-fication and characterization using molecular and immunologic tools. Am. J. Trop. Med. Hyg. 76, 896-901.
Kuntz, E. R. (1973). Models for Investigation in Parasitology. In H. G. Bourne (Ed). Nonhuman Primates and Medical Research. New York and London: Academ-ic Press. Pp. 167-175.
Kuntz, E.R and Moore, J.A (1975). Commensals and parasites of African baboons (Papio cyanocephalus, L.1766) captured in Rift Valley province of central Kenya. J Med Primatol. 2:236-241.
Leathers, C. W. (1978). The Prevalence of Hepatocystis kochi in African Green Monkeys. Laboratory Animal Science. 28:186-189.
Leeflang, P., Buys, J. and Blotkamp, C. (1978). Studies on Trypanosoma vivax. Comparison of Parasitological Diagnostic Methods. International Journal of Parasitology, Pp 8: 15-18
Levine, N.D. (1970). Protozoan parasites of nonhuma primates as zoonotic agents. Laboratory Animal Care, Part II. No.2. 20:372-382.
Losos, J. George. (1986). Infectious Tropical Diseases of Domestic Animals. NY, USA, Churchil Livingstone Inc, Pp I: 3-319.
Molyneux, D.H. and Ashford, R.W. (1983). The Biology of Trypanosoma and Leishmania. Taylor and Francis Ltd. London, (Pp 5-89).
Morgan U.M and Thompson R.C. (1998). Molecular detection of parasitic protozoa. Parasitology, 117:73-85.
Njiokou F, Simo G, Nkinin S.W, Laveissière C, Herder S. (2004). Infection rate of Trypanosoma brucei s.l., T. vivax, T. congolense "forest type", and T. simiae in small wild vertebrates in south Cameroon. Acta Trop. 92:139-46
Oates, F. J. (1996). African Primates. States Survey and Conservation Action Plan. Revised Edn. IUCN, Gland Switzerland (Pp. 2-4).
Oliveira, R.L. and Dean, L.M. (1995). Simian Malaria at Two Sites in the Brazilian Amazon. The Infection Rates of Plasmodium Brasilianum in non-human pri-mates Memorias do Instituto Oswaldo Cruz, 90(3): 331-339.
Olobo J.O., Reid, G.D.F., Githure J.I., Anjili, C.O., and Koech, D. K. (1990). Vervet Monkey Model for Human Cutaneous Leishmaniasis. XIV Congress of the In-ternational Primatological Society.
Otsuru, M. and Sekikawa, H. (1979). Surveys of simian malaria in Japan. Zentrallatt fur Bakteriologie and Hygiene, 345-350.
Passos L.M., Bell-Sakyi L., and Brown C.G. (1998). Immunochemical characteriza-tion of in vitro culture-derived antigens of Babesia bovis and Babesia bigemi-na. Vet. Parasitol. 76: 239-249.
Pearson, R.D., Wheler, D.A., Harison, L.H. and Kay, D.H. (1983). Review of Infec-tious Diseases. Vol 5. The University of Chicago. (Pp. 907-923).

51
Poltera, A.A., Sayer, P.D., Rudinm, W and Bovel, D. (1985). Trypanosomal cardiac valvulitis in vervet monkeys. Journal of Tropical Medicine and Parasitology, 36(2):77-80.
Qari, S.H., Shi, Y. P., Pieniazek, N.J., Collins, W. E. and Lal, A.A. (1996). Phylo-genetic relationship among the malaria parasites based on small subunit rRNA gene sequences: Monophyletic nature of the human malaria parasite, Plasmo-dium falciparum. Molecular phylogenetics and evolution, 6:–165.
Rutto, J.J. and Karuga, J. W. (2009). Temporal and spatial epidemiol-ogy of sleeping sickness and use of geographical information sys-tem (GIS) in Kenya. J. Vector Borne Dis. 46: 18-25.
Seethamchai, S., Putaporntip, C., Malaivijitnond, S., Cui. L., and Jongwutiwes, S. (2008). Malaria and Hepatocystis species in wild macaques, southern Thailand. Am J Trop Med Hyg. 78(4):646-53.
Shkap, V., de Vos, A. J., Zweygarth E., and Jongejan, F. (2007). Attenuated vac-cines for tropical theileriosis, babesiosis and heartwater: the continuing neces-sity. Trends Parasitol. 23:420–426.
Singh, B., Cox-Singh, J., Miller, A.O., Abdullah, M.S., Snounou, G., Rahman, H.A., (1996). Detection of malaria in Malaysia by nested polymerase chain re-action amplification of dried blood spots on filter papers. Trans R Soc Trop Med Hyg. 90: 519–521.
Snounou, G., Pinheiro, L., Gonc¸alves A, Fonseca, L., Dias, F., Brown, K.N., Do Rosario, V., (1993). The importance of sensitive detection of malaria parasites in the human and insect hosts in epidemiological studies, as shown by the analysis of field samples from Guinea Bissau. Trans R Soc Trop Med Hyg. 87:649–653.
Steenkeste N, Incardona S, Chy S, Duval L, Ekala MT, Lim P, Hewitt S, Sochantha T, Socheat D, Rogier C, Mercereau-Puijalon O, Fandeur T, Ariey F (2009). Towards high-throughput molecular detection of Plasmodium: new approaches and molecular markers. Malar J. 29: 80-86.
Steirger, R.F. (1973). The Ultrastructure of T. brucei in The Course of its Life Cycle and Some Related Aspects. Acta Tropica, 30(1-2):64-168.
Suleman M. A., Wango, E., Farah, I. O., Hau, J. (2000). Adrenal cortex and stomach lesions associated with stress in wild male African green monkeys (Cercopithecus aethiops) in the post-capture period. J. Med. Primatol 29:388-342.
Telford, S.R III., Gorenflot, A., Brasseur, P., and Spielman, A. (1993). Babesial. infections in humans and wildlife. In: Kreier JP, ed. Parasitic protozoa. 2nd ed. Vol. 5. San Diego, Calif. Academic Press. Pp.1-47.
Turner, T.R., Lambrrecht, F.L. and Jolly, C.L. (1982). Hepatocystis Parasitemia In Wild Kenya Vervet Monkeys (Cecopithecus Aethiops). Journal of Medical Primatology,11,191-194.
Uilenberg, G. (1995). International collaborative research: significance of tick-borne hemoparasitic diseases to world animal health. Vet. Parasitol. 57, 19–41.
Vial, H.J. and Gorenflot , A. (2006). Chemotherapy against babesiosis. Vet Parasi-tol. 31;138(1-2):147-60.
Voller, A. (1972). Plasmodium and Hepatocystis. In R. N. T. W Fiennes, (Ed.). Pa-thology of simian primates, Part II: Infectious and Parasitic Diseases. New York.
Voller, A. and Bidwell, D. (1995). Enzyme-Linked Immunosorbent Assay. Clinical Immunology, 17:99 - 107.
Warhurst, D.C and Williams, J.E. (1996). Laboratory Diagnosis of Malaria. Journal of Clinical Pathology, 49:533-38.

52
Warren, Mc W. (1970). Simian and Anthropoid Malarias: Their Role in Human Duseases. Laboratory Animal Care. No. 2. Part II. American Association of Laboratory Animal Care, 20:368-376.
Weinman, D., Wallis, R. C., Cheong, W.H. and Mahadevan, S. (1978) Triatomines as experimental vectors of trypanosomes of Asian monkeys. American Journal of Medical Hygiene, (Pp. 232-237).
White, N. J. (2008). Plasmodium knowlesi: The Fifth Human Malaria Parasite . Clinical Infectious Diseases. 46:172–3.
WHO, (2001). African trypanosomiasis or sleeping sickness. WHO. http://www.who.int/ mediacentre/factsheets/
WHO, 1990. Control of Leishmaniasis. Report of a WHO Expert Committee. Geneva; World Health Organization. (WHO/ LEISH/98.9).
WHO, 2001. Report of the Scientific Working Group meeting on African trypano-somiasis Geneva, 4-8.
Wijers, D.J.B. (1971). A Ten Years Study of Kala Azar in Tharaka (Meru District, Kenya) Part1. Incidence Studies from Marimanti. East African Medical Jour-nal. 48: 533.
Wolfe N.D, Escalante A.A, Karesh W.B, Kilbourn A, Spielman A, Lal A.A. (1998).Wild primate populations in emerging infectious disease research: the missing link? Emerg Infect Dis. 4: 149–158.
World Health Organization (1991). Tropical Diseases: Progress in Research 1989-1990. W. H.O, Geneva.
Wykes M, Good MF, (2007). A case for whole-parasite malaria vaccines. Int J Pa-rasitol 37: 705–712.
Ziccardi, M, and R. Lourenco-de-Oliveira. 1997. The infection rates of trypano-somes in squirrel monkeys at two sites in the Brazilian Amazon. Mem Inst Oswaldo Cruz. 92 (4): 465-470.

















![[TITLE WITH CAPITAL LETTERS]pure.au.dk/portal/files/79630819/AW_Bacteria_and_protozoa_in_soil... · Protozoa ›Protista: unicellular eukaryotic organisms: protozoa, unicellular algae,](https://img.pdfslide.us/doc/110x75/606ca0b8d91e76743244800e/title-with-capital-letterspureaudkportalfiles79630819awbacteriaandprotozoainsoil.jpg)

![Protozoa: [s., protozoan; Greek protos, first, and zoon, animal] A protozoan can be defined as a usually motile, eukaryotic, unicellular protest. the](https://img.pdfslide.us/doc/110x75/56649d965503460f94a7fbc1/protozoa-s-protozoan-greek-protos-first-and-zoon-animal-a-protozoan.jpg)