Embed Size (px)
Citation preview

Molecular Plant • Volume 6 • Number 3 • Pages 975–977 • May 2013 LETTER TO THE EDITOR
H3K36 Methylation Is Involved in Promoting Rice Flowering
Dear Editor,Histone H3 lysine 36 (H3K36) methylation is a conserved
epigenetic mark in all eukaryotes (Berr et al., 2011; Wagner and Carpenter, 2012). Reverse genetic analysis in Arabidopsis had uncovered a crucial role of H3K36 di- and tri-methylation (H3K36me2 and H3K36me3) in flowering-time regulation (reviewed in Berr et al., 2011). The Arabidopsis SET DOMAIN GROUP 8 (SDG8), also known as EARLY FLOWERING IN SHORT DAYS (EFS), encodes a SET-domain protein show-ing similarity to the yeast SET2 and the Drosophila ASH1 and the human NSD1 proteins. The sdg8 loss-of-function mutants exhibit an early-flowering phenotype associated with reduced levels of H3K36me2 and H3K36me3 at chro-matin and suppression of FLOWERING LOCUS C (FLC), a key flowering repressor in Arabidopsis (Kim et al., 2005; Zhao et al., 2005; Xu et al., 2008). Flowering time is a critical trait in plant life history and is fine-tuned by complex gene net-works (Tsuji et al., 2011; He, 2012). Because of a lack of FLC homolog in rice, it is of great interest to examine the role of H3K36 methylation in flowering-time control in this eco-nomic important plant.
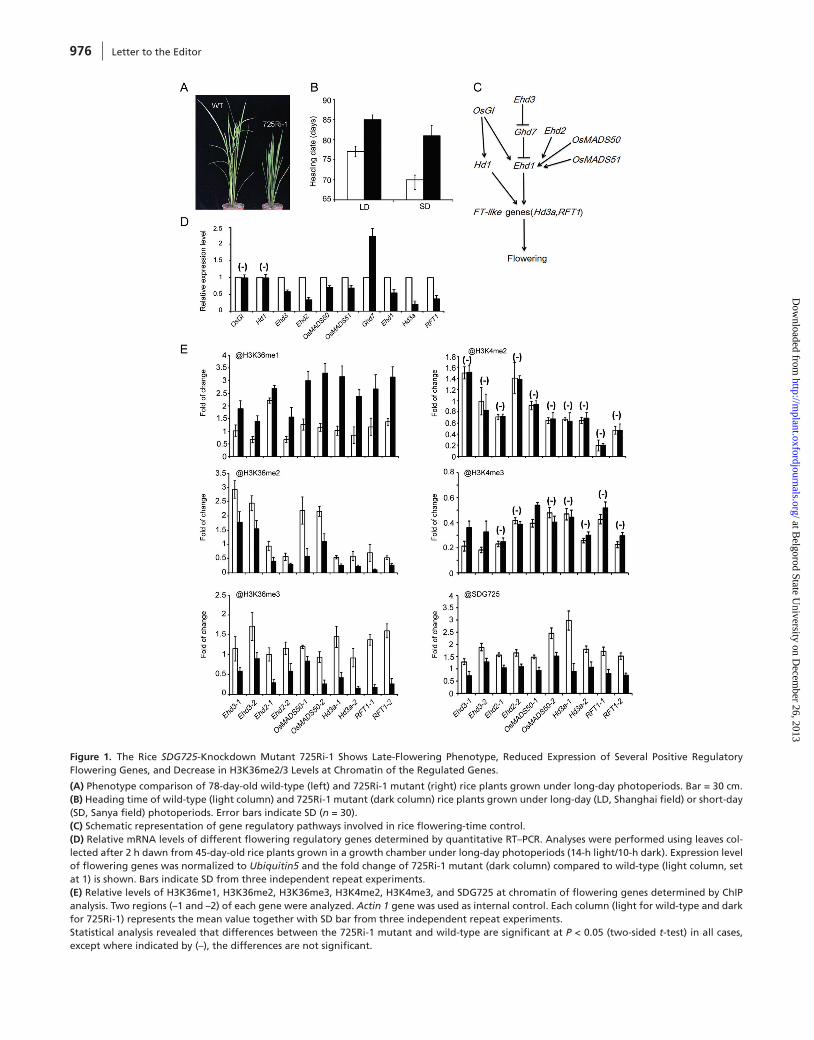
Previously, we reported that the rice SDG725 encodes a close homolog of SDG8 exhibiting H3K36-specific histone methyltransferase activity in vitro and in vivo, and is involved in promotion of brassinosteroid-related gene expression (Sui et al., 2012). In this study, we show that SDG725 is involved in promoting rice flowering. To investigate flowering time, we grew side by side the 725Ri-1 mutant line, which exhibits a knockdown of over 80% of SDG725 transcripts by a RNAi construct (Sui et al., 2012), together with wild-type (WT) in two latitude places: Shanghai with a subtropical climate and long-day (LD) photoperiods and Sanya with tropical climate and short-day (SD) photoperiods. At both places, the 725Ri-1 mutant plants showed a late-flowering phenotype compared to WT (Figure 1A) and the average heading time of the 725Ri-1 mutant compared to WT was considerably delayed (Figure 1B).
We next investigated expression levels of several genes, including OsGI, Hd1, Ehd3, Ehd2 (also named RID1 or OsId1), OsMADS50, OsMADS51, Ghd7, Ehd1, Hd3a, and RFT1, from different flowering regulatory pathways (Figure 1C; Tsuji et al., 2011). Leaf samples were collected 2 h after dawn from 45-day-old plants grown in a growth chamber under LD (14-h light/10-h dark) as well as from 35-day-old plants grown under SD (10-h light/14-h dark) growth conditions.
They were subjected to quantitative RT–PCR analysis using gene-specific primers. No significant differences in the levels of OsGI and Hd1 transcripts were observed between 725Ri-1 and WT under either SD or LD conditions (shown for LD in Figure 1D). Remarkably, the expression levels of Ehd3, Ehd2, OsMADS50, OsMADS51, Ehd1, Hd3a, and RFT1 all were dras-tically reduced in 725Ri-1 compared with WT under either SD or LD conditions (shown for LD in Figure 1D). In agree-ment with a previous report showing that Ehd3 functions as a repressor of Ghd7 (Matsubara et al., 2011), Ghd7 expression was elevated in 725Ri-1 (Figure 1D).
To investigate histone methylation status in chromatin of flowering genes, we performed chromatin immunoprecipitation (ChIP) experiments using commercially available antibodies specifically recognizing H3K36me1, H3K36me2, H3K36me3, H3K4me2, or H3K4me3. We chose to analyze histone methylation levels at Ehd3, Ehd2, OsMADS50, Hd3a, and RFT1 genes located at different positions within the flowering regulatory pathways (Figure 1C). As shown in Figure 1E, 725Ri-1 mutant compared with WT exhibited reduced H3K36me2 and H3K36me3 levels accompanied by increased H3K36me1 levels at all examined flowering genes, with each gene analyzed at two different regions using specific PCR primers. In contrast, H3K4me2 levels were unaffected and H3K4me3 levels were increased in some regions in 725Ri-1 mutant compared with WT (Figure 1E). Using the SDG725-specific antibody (Sui et al., 2012), we found that the chromatin-binding levels of the SDG725 protein at flowering genes were significantly reduced in 725Ri-1 mutant compared with WT (Figure 1E). These results indicate that SDG725 is recruited and is specifically required for deposition of H3K36me2/3, which is associated with active transcription of flowering genes.
Taken together, this study establishes a crucial func-tion of SDG725 in rice flowering-time regulation via H3K36me2/3-mediated activation of several genes within photoperiodic pathways. Recently, Sun and colleagues
© The Author 2012. Published by the Molecular Plant Shanghai Editorial Office in association with Oxford University Press on behalf of CSPB and IPPE, SIBS, CAS.
doi:10.1093/mp/sss152, Advance Access publication 13 December 2012
Received 28 November 2012; accepted 4 December 2012
at Belgorod State U
niversity on Decem
ber 26, 2013http://m
plant.oxfordjournals.org/D
ownloaded from
976 Letter to the Editor
Figure 1. The Rice SDG725-Knockdown Mutant 725Ri-1 Shows Late-Flowering Phenotype, Reduced Expression of Several Positive Regulatory Flowering Genes, and Decrease in H3K36me2/3 Levels at Chromatin of the Regulated Genes.
(A) Phenotype comparison of 78-day-old wild-type (left) and 725Ri-1 mutant (right) rice plants grown under long-day photoperiods. Bar = 30 cm.(B) Heading time of wild-type (light column) and 725Ri-1 mutant (dark column) rice plants grown under long-day (LD, Shanghai field) or short-day (SD, Sanya field) photoperiods. Error bars indicate SD (n = 30).(C) Schematic representation of gene regulatory pathways involved in rice flowering-time control.(D) Relative mRNA levels of different flowering regulatory genes determined by quantitative RT–PCR. Analyses were performed using leaves col-lected after 2 h dawn from 45-day-old rice plants grown in a growth chamber under long-day photoperiods (14-h light/10-h dark). Expression level of flowering genes was normalized to Ubiquitin5 and the fold change of 725Ri-1 mutant (dark column) compared to wild-type (light column, set at 1) is shown. Bars indicate SD from three independent repeat experiments.(E) Relative levels of H3K36me1, H3K36me2, H3K36me3, H3K4me2, H3K4me3, and SDG725 at chromatin of flowering genes determined by ChIP analysis. Two regions (–1 and –2) of each gene were analyzed. Actin 1 gene was used as internal control. Each column (light for wild-type and dark for 725Ri-1) represents the mean value together with SD bar from three independent repeat experiments.Statistical analysis revealed that differences between the 725Ri-1 mutant and wild-type are significant at P < 0.05 (two-sided t-test) in all cases, except where indicated by (–), the differences are not significant.
at Belgorod State U
niversity on Decem
ber 26, 2013http://m
plant.oxfordjournals.org/D
ownloaded from

Letter to the Editor 977
(2012) reported that SDG724, encoding a SET-domain pro-tein much smaller than SDG725, is required for deposition of H3K36me2/3 at OsMADS50 and RFT1 but not Hd3a chro-matin. Thus, SDG724 and SDG725 show both overlapped and specific functions in regulation of different flowering genes. Both studies on SDG724 and SDG725 demonstrate that H3K36me2/3 promotes rice flowering, which is in sharp contrast to the previously known function of these epigenetic marks in Arabidopsis flowering suppression (Zhao et al., 2005; Xu et al., 2008). The difference resides in H3K36me2/3-mediated transcription activation of FLC, a key flowering repressor present in Arabidopsis but absent in rice. This comparative observation remarkably illustrates an example of how a similar epigenetic regulatory mecha-nism could lead to distinct physiological/phenotypical out-comes between two plant species. The finding of histone methylation in rice flowering-time control intrigues future research about their roles in rice domestication and possi-ble application in rice breeding.
FUNDING
This work was funded by the National Basic Research Program of China (973 Program, grant no. 2012CB910500). No conflict of interest declared.
Pengfei Suia,2, Jinlei Shia,2, Xueying Gaoa, Wen-Hui Shenb and Aiwu Donga,1
a State Key Laboratory of Genetic Engineering, Institute of Plant Biology, Department of Biochemistry, School of Life Sciences, Fudan University,
Shanghai 200433, P.R. Chinab Institut de Biologie Moléculaire des Plantes du CNRS, Université de
Strasbourg, 12 rue du Général Zimmer, 67084 Strasbourg Cedex, France1 To whom correspondence should be addressed. E-mail aiwudong@fudan.
edu.cn, tel. +86-21–55665673.2 These authors contributed equally to this work.
REFERENCES
Berr, A., Shafiq, S., and Shen, W.H. (2011). Histone modifications in transcriptional activation during plant development. Biochim. Biophys. Acta. 1809, 567–576.
He, Y. (2012). Chromatin regulation of flowering. Trends Plant Sci. 17, 556–562.
Kim, S.Y., He, Y., Jacob, Y., Noh, Y.S., Michaels, S., and Amasino, R. (2005). Establishment of the vernalization-responsive, win-ter-annual habit in Arabidopsis requires a putative histone H3 methyl transferase. Plant Cell. 17, 3301–3310.
Matsubara, K., Yamanouchi, U., Nonoue,Y., Sugimoto, K., Wang, Z., Minobe, Y., and Yano, M. (2011). Ehd3, encoding a plant homeodomain finger-containing protein, is a critical promoter of rice flowering. Plant J. 66, 603–612.
Sui, P., Jin, J., Ye, S., Mu, C., Gao, J., Feng, H., Shen, W.H., Yu, Y., and Dong, A. (2012). H3K36 methylation is critical for brassi-nosteroid-regulated plant growth and development in rice. Plant J. 70, 340–347.
Sun, C., Fang, J., Zhao, T., Xu, B., Zhang, F., Liu, L., Tang, J., Zhang, G., Deng, X., Chen, F., (2012). The histone methyltransferase SDG724 mediates H3K36me2/3 deposition at MADS50 and RFT1 and promotes flowering in rice. Plant Cell. 24, 3235–3247.
Tsuji, H., Taoka, K., and Shimamoto, K. (2011). Regulation of flow-ering in rice: two florigen genes, a complex gene network, and natural variation. Curr. Opin. Plant Biol. 14, 45–52.
Wagner, E.J., and Carpenter, P.B. (2012). Understanding the lan-guage of Lys36 methylation at histone H3. Nat. Rev. Mol. Cell Biol. 13, 115–126.
Xu, L., Zhao, Z., Dong, A., Soubigou-Taconnat, L., Renou, J.P., Steinmetz, A., and Shen, W.H. (2008). Di- and tri- but not mon-omethylation on histone H3 lysine 36 marks active transcrip-tion of genes involved in flowering time regulation and other processes in Arabidopsis thaliana. Mol. Cell Biol. 28, 1348–1360.
Zhao, Z., Yu, Y., Meyer, D., Wu, C., and Shen, W.H. (2005). Prevention of early flowering by expression of FLOWERING LOCUS C requires methylation of histone H3 K36. Nat. Cell Biol. 7, 1256–1260.
at Belgorod State U
niversity on Decem
ber 26, 2013http://m
plant.oxfordjournals.org/D
ownloaded from