Embed Size (px)
Citation preview

Vol. 49, No. 6APPLIED AND ENVIRONMENTAL MICROBIOLOGY, June 1985, p. 1502-15060099-2240/85/061502-05$02.00/0Copyright C) 1985, American Society for Microbiology
Growth of Agaricus campestris NRRL 2334 in the Form of PelletsANTONIO M. MARTIN* ANDVALERIE I. BAILEY
Department ofBiochemistry, Memorial University of Newfoundland, St. John's, Newfoundland, Canada AIB 3X9
Received 11 October 1984/Accepted 19 March 1985
The production of pellets of the fungus Agaricus campestris NRRL 2334 was studied in submergedfermentation with peat extract as the main substrate source. Pellets up to 6mm in diameter were obtained whenthe peat extract was diluted to reduce the concentration of growth inhibitors. Yeast extract and yeast extractplus glucose were the most effective nutrient supplements in the diluted peat extract media and stimulated theformation of large pellets which contained 44.4% crude protein, 2.8% fat, and 9% ash (dry weight basis). Nosolid supports were required for the growth of the pellets. The effects on the growth morphology of severaldilution ratios of the peat extract, rates of agitation and aeration, and time were investigated.
The submerged growth- of molds can adopt two mainforms, diffused filamentous mycelia or agglomerated myce-lia; the latter is usually called fungal pellets. The formationof pellets has been reported in the growth of many genera ofmolds (1, 21). In industrial fermentation processes, thegrowth of molds in the form of pellets could have importantconsequences. For example, the viscosity of a fermentationbroth could be lower in a pellet suspension than in afilamentous one, enhancing the mass transfer and mixingcharacteristics of the medium. However, there are fewindustrial processes in which pellet growth has been re-ported (4, 13, 19). A possible disadvantage in the formationof pellet biomass is the increased diffusional limitation ofoxygen and other nutrients to the interior of the pellet (20).
Pellet formation has been studied in several fermentationprocesses, and different mechanisms and models have beenproposed to explain pellet growth (14). In some cases, theappearance of pellets is not desired because they lead to anonhomogeneous mycelium. However, pellets have a poten-tial application in mainly two areas: (i) as a support or carriermaterial for immobilized enzymes and cells and (ii) in thesubmerged production of edible mushroom mycelium (7). Inthe latter, pellet formation is important for ease of separationof the biomass from the fermentation medium and for thedevelopment of flavor (8), which is apparently due to auto-lysis in the center of the pellet (20). In addition, the pellettexture could be more acceptable as a mushroom substitutethan the texture of filamentous mycelia. Agaricus campestrishas been utilized to a great extent in research aimed at thesubmerged growth of mushroom mycelium (8, 22), from theinitial works of Humfeld (5) to the utilization of peat extractas a substrate source (10). However, the original strains usedby Humfeld were questioned by Molitoris (15), who claimedthey probably were strains of Beauveria tenella.The mixture of organic compounds in peat and its rela-
tively low price make this product a potential source ofnutrients in fermentation processes. Heat treatment of peatin an acid solution produces a liquid extract which contains3 to 5% total carbohydrates. In the submerged growth ofmushroom mycelium, it has been found that yeast extract-supplemented peat extract improves growth. However, in-hibitory effects have also been detected and, in previousexperiments, pellet formation was not observed in the sub-
* Corresponding author.
merged growth of A. campestris (A. M. Martin and V. I.Bailey, 27th Ann. Conf. Can. Inst. Food Sci. Technol., 1984,abstr. 87, p. 55). The complex composition of peat and itsextracts has made difficult the identification of many oftheir growth-promoting and growth-inhibiting components.Therefore, any attempt to remove the latter could result inthe simultaneous removal of nutrients from the medium.This was the case in the growth of Candida utilis in a peatextract medium (17). The growth rate and biomass concen-tration decreased when the peat extract was treated withactivated carbon. In this work, ways to produce pellets of A.campestris with peat extract as a major nutrient source andthe reduction of the inhibiting effect of this substrate onmycelial growth were investigated.
MATERIALS AND METHODS
Organism. The fungus A. campestris NRRL 2334 wasobtained from the American Type Culture Collection, Beth-esda, Md., and was adapted to use peat extract as the mainsubstrate source (11).Growth medium. The fungus was maintained on peat
extract agar slants and was aseptically transferred into 100ml of sterile culture medium in 250-ml shake flasks. Thefollowing medium compositions were used. Peat extract wasdiluted with distilled water in the ratios 3:1, 1:1, and 1:3. Thesupplements 0.5% (wt/wt) yeast extract, 0.5% (wt/wt)(NH4)2HP04, and 1% glucose, when added, were used inaddition to the nutrients already present in the peat extract.Combinations of the diluted peat extract and the supple-ments were used as growth media at 24°C and pH 6 (10). ThepH of the peat extract was adjusted from its original value of1.5 with 15 N NaOH.Source and preparation of peat extract. Sphagnum peat
moss from bogs near St. John's, Newfoundland, Canada,was hydrolyzed with 1.5% (vol/vol) H2SO4, and the extractwas obtained as previously described (12). The extractionprocess was designed to obtain a peat extract with a totalcarbohydrate concentration of ca. 3% (wt/wt).
Fermentation and separation of biomass. The inoculatedshake flasks were incubated in a Gyrotory water batchshaker (model G76; New Brunswick Scientific Co., Inc.,Edison, N.J.) for 8 days at 150 rpm. The inoculum added tothe shake flasks contained at least 0.5 cm2 of the myceliumfrom the stock culture and was obtained by blending themycelium slants with 50 ml of distilled water in a Waring
1502
Dow
nloa
ded
from
http
s://j
ourn
als.
asm
.org
/jour
nal/a
em o
n 31
Jan
uary
202
2 by
46.
70.3
8.15
3.

AGARICUS CAMPESTRIS PELLETS 1503
A - .0
Q
0
0
0.5 1.0
3:1 1:1 1:3Peat Extract:Water Ratio
FIG. 1. Final dry biomass concentration (Conc.) and mycelialmorphology of A. campestris grown in diluted and nutrient-supple-mented peat extract. Symbols indicate the predominant morphologyas follows: O and M, filamentous; A and A, small pellets; and 0 and0, large pellets. Solid symbols represent media containing 1%glucose, solid lines indicate supplementation with yeast extract, andbroken lines indicate supplementation with (NH4)2HPO4. Processeswere conducted in shake flasks.
blender for 30 s under aseptic conditions. An inoculum ratioof 5% (vol/vol) was used in all the processes. The necessityof having the mycelium from the shake flasks in a homoge-neous form at the start of each experiment made blendingagain necessary when transferring the inoculum to theaerated and agitated fermentation stage, which was con-
ducted in a 2-liter batch fermentor (model Bioflo; NewBrunswick Scientific Co.) with a 1-liter working volume offermentation medium.The pH of the fermentor medium was maintained by using
automatic control equipment, which added 0.5 N NaOH or0.5 N H2SO4 when the pH deviated from the desired value.The fermentor was equipped with two sets of a standardsix-flat-blade turbine and four baffle plates. Two levels ofmechanical agitation (100 and 200 rpm) were used, in addi-tion to the processes conducted without mechanical agita-tion. Three levels of aeration rates were tested: 0.5, 1, and1.5 volumes of air per volume of medium per minute. Thefermentor cultures, except when indicated otherwise, were
grown for 8 days.Analytical methods. The nitrogen content of the washed
mycelium was calculated by the Kjeldahl method, and thepercent protein content was determined by using the nitro-gen content (N x 6.25). Total lipids were determined byether extraction. Ash was determined by combustion in a
muffle furnace at 600°C. The mycelial dry weight was
calculated by filtering the culture medium through oven-
dried, weighed Whatman no. 1 filter paper, washing the filterpaper and mycelium with distilled water, and drying (105°C)the filter paper and mycelium to a constant weight, with thedifference being the mycelial dry weight. The total carbohy-drate concentration was determined by the anthrone reagent
Aeration rates (vvm)FIG. 2. Effect of aeration and agitation on the final dry biomass
concentration (Conc.) and mycelial morphology of A. campestrisgrown in diluted (1:1) peat extract supplemented with 0.5% yeastextract and 1.0% glucose. Symbols indicate the predominant mor-
phology as follows: OI, filamentous; A, small pellets; and 0, largepellets. The revolutions per minute used were as follows: -, none;
---, 100; and -, 200. vvm, Volume of air per volume of mediumper minute. Processes were conducted in a 2-liter fermentor.
method (16). Analytical grade chemicals were used in theexperiments. All reported results are the averages of at leastthree determinations.
RESULTS AND DISCUSSIONShake flask fermentations. In an attempt to avoid the
observed inhibitory effects of some of the peat extractcomponents on mycelium growth. diluted peat extract was
used as the substrate source. Based on the results ofprevious experiments on the growth of A. c ampestris in peatextract (A. M. Martin and V. 1. Bailey. 27th Ann. Conf. Can.Inst. Food Sci. Technol., 1984. abstr. 87, p. 55), yeastextract or (NH4)2HP04 was used as a nutrient supplement.Figure 1 shows the growth obtained at three dilution levels.The dilution ratio of 3:1 (peat extract/distilled water) pro-duced results similar to those obtained when nondiluted peatextract was used. Diluting the peat extract in a 1:1 ratiogenerally enhanced mycelial growth. indicating that thedilution of the growth inhibitors in the peat extract produceda stronger effect on the growth of A. (anmpestris than did thereduction of the substrate concentration in the medium.Further dilution of the peat extract (1:3) caused a lowering inthe biomass concentration because of an increased reductionof the available nutrients in the medium. The addition ofglucose increased the dry mycelial concentration obtained,especially in the more dilute peat extract.
Besides the effect on the biomass concentration of dilutingand supplementing the peat extract, a collateral effect on theform of the mycelial growth was observed. In general, threetypes of growth were observed: (i) filamentous growth, suchas that observed when nondiluted peat extract was used, (ii)small pellets with an approximate diameter of 2 mm or less,and (iii) larger pellets with an approximate diameter of 3 to 6mm. A combination of filamentous and pellet growth was
0
U
4.0-J
.3.5Uc00
w3.00E0
j:2.5L,.0
2.01
AiS -
0-,
I
67.0%-o
U)c
0ro'6(A
tn
E.*5.0
4.C
A
0o
a
3.01.5
VOL. 49, 1985
M.- .-A 1%
El-
Dow
nloa
ded
from
http
s://j
ourn
als.
asm
.org
/jour
nal/a
em o
n 31
Jan
uary
202
2 by
46.
70.3
8.15
3.

1504 MARTIN AND BAILEY
FIG. 3. Pellets of A. campestris grown in diluted (1:1) and nutrient-supplemented peat extract in an aerated fermentor (1.0 volume of airper volume of medium per minute).
observed, specifically when small pellets were produced.Quantitatively, a specific type of growth was consideredpredominant when it accounted for more than 65% of the drybiomass concentration. The types of growth in differentmedia are shown in Fig. 1.As in nondiluted peat extract, irrespective of nutrient
supplementation, growth in peat extract diluted 3:1 in dis-tilled water was filamentous. More dilute peat extract al-lowed the production of mycelial pellets, larger pellets beingproduced when yeast extract and yeast extract plus glucosewere incorporated into the medium. Because the addition ofglucose alone did not produce the pellet form of growth, itwas concluded that yeast extract, when added to dilutedpeat extract, was the main factor in the production of thelarger pellets.
Aerated and agitated fermentations. The effects of aerationrates and mechanical agitation on the production of pellets
are shown in Fig. 2. Because of the results in shake flasks,yeast extract-plus-glucose supplement and peat extract di-luted 1:1 were used as the growth medium.The effect of aeration on the growth of mushroom myce-
lium, specifically in the development of a pelleted mycelium,is complex and, in some cases, contradictory (8). However,it appears that lower aeration rates facilitate pellet growth(9). The oxygen mass transfer in the fermentation suspensionis enhanced when pellets are formed because of the lowerresistance produced by lowering the viscosity of the me-dium, as compared with filamentous growth in the same typeof medium. However, the oxygen supply to the interior ofthe pellets decreases because of the condensation of myce-lium characteristic of the pellet structure and as a function ofTthe pellet diameter.
Figure 2 shows that increased aeration rates enhanced thefinal dry mycelium concentration obtained in the fermenta-
APPL. ENVIRON. MICROBIOL.
Dow
nloa
ded
from
http
s://j
ourn
als.
asm
.org
/jour
nal/a
em o
n 31
Jan
uary
202
2 by
46.
70.3
8.15
3.
AGARICUS CAMPESTRIS PELLETS 1505
-J
C)1
%-O
c0
n
0
E0
L-a
4.0
30
2.0
1.0
//^//////
0.'/
//
/
/c// (
/
00
2 4 6 10 12Time (days)
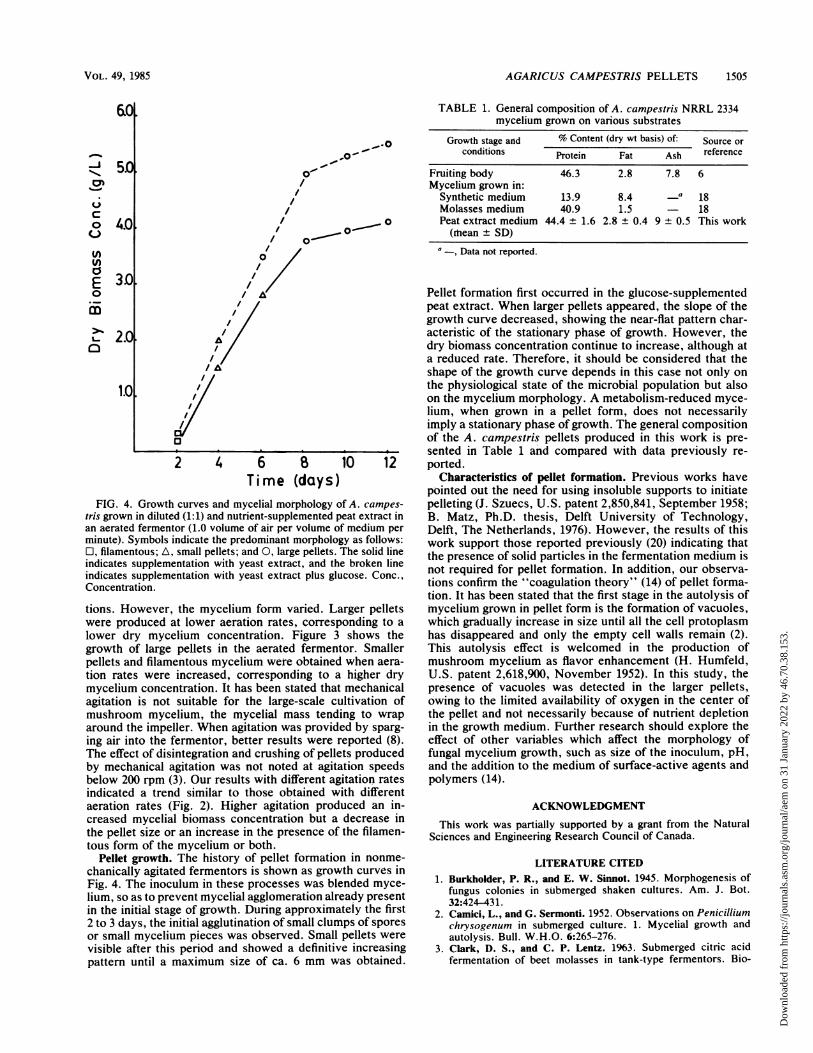
FIG. 4. Growth curves and mycelial morphology of A. campes-
tris grown in diluted (1:1) and nutrient-supplemented peat extract inan aerated fermentor (1.0 volume of air per volume of medium per
minute). Symbols indicate the predominant morphology as follows:O, filamentous; A, small pellets; and 0, large pellets. The solid lineindicates supplementation with yeast extract, and the broken lineindicates supplementation with yeast extract plus glucose. Conc.,Concentration.
tions. However, the mycelium form varied. Larger pelletswere produced at lower aeration rates, corresponding to a
lower dry mnycelium concentration. Figure 3 shows thegrowth of large pellets in the aerated fermentor. Smallerpellets and filamentous mycelium were obtained when aera-tion rates were increased, corresponding to a higher drymycelium concentration. It has been stated that mechanicalagitation is not suitable for the large-scale cultivation ofmushroom mycelium, the mycelial mass tending to wraparound the impeller. When agitation was provided by sparg-ing air into the fermentor, better results were reported (8).The effect of disintegration and crushing of pellets producedby mechanical agitation was not noted at agitation speedsbelow 200 rpm (3). Our results with different agitation ratesindicated a trend similar to those obtained with differentaeration rates (Fig. 2). Higher agitation produced an in-creased mycelial biomass concentration but a decrease inthe pellet size or an increase in the presence of the filamen-tous form of the mycelium or both.
Pellet growth. The history of pellet formation in nonme-
chanically agitated fermentors is shown as growth curves in
Fig. 4. The inoculum in these processes was blended myce-lium, so as to prevent mycelial agglomeration already presentin the initial stage of growth. During approximately the first2 to 3 days, the initial agglutination of small clumps of sporesor small mycelium pieces was observed. Small pellets were
visible after this period and showed a definitive increasingpattern until a maximum size of ca. 6 mm was obtained.
TABLE 1. General composition of A. campestris NRRL 2334mycelium grown on various substrates
Growth stage and % Content (dry wt basis) of: Source orconditions Protein Fat Ash reference
Fruiting body 46.3 2.8 7.8 6Mycelium grown in:
Synthetic medium 13.9 8.4 _a 18Molasses medium 40.9 1.5 - 18Peat extract medium 44.4 + 1.6 2.8 ± 0.4 9 ± 0.5 This work
(mtean ± SD)
a, Data not reported.
Pellet formation first occurred in the glucose-supplementedpeat extract. When larger pellets appeared, the slope of thegrowth curve decreased, showing the near-flat pattern char-acteristic of the stationary phase of growth. However, thedry biomass concentration continue to increase, although ata reduced rate. Therefore, it should be considered that theshape of the growth curve depends in this case not only onthe physiological state of the microbial population but alsoon the mycelium morphology. A metabolism-reduced myce-lium, when grown in a pellet form, does not necessarilyimply a stationary phase of growth. The general compositionof the A. campestris pellets produced in this work is pre-sented in Table 1 and compared with data previously re-ported.
Characteristics of pellet formation. Previous works havepointed out the need for using insoluble supports to initiatepelleting (J. Szuecs, U.S. patent 2,850,841, September 1958;B. Matz, Ph.D. thesis, Delft University of Technology,Delft, The Netherlands, 1976). However, the results of thiswork support those reported previously (20) indicating thatthe presence of solid particles in the fermentation medium isnot required for pellet formation. In addition, our observa-tions confirm the "coagulation theory'" (14) of pellet forma-tion. It has been stated that the first stage in the autolysis ofmycelium grown in pellet form is the formation of vacuoles,which gradually increase in size until all the cell protoplasmhas disappeared and only the empty cell walls remain (2).This autolysis effect is welcomed in the production ofmushroom mycelium as flavor enhancement (H. Humfeld,U.S. patent 2,618,900, November 1952). In this study, thepresence of vacuoles was detected in the larger pellets,owing to the limited availability of oxygen in the center ofthe pellet and not necessarily because of tnutrient depletionin the growth medium. Further research should explore theeffect of other variables which affect the morphology offungal mycelium growth, such as size of the inoculum, pH,and the addition to the medium of surface-active agents andpolymers (14).
ACKNOWLEDGMENTThis work was partially supported by a grant from the Natural
Sciences and Engineering Research Council of Canada.
LITERATURE CITED1. Burkholder, P. R., and E. W. Sinnot. 1945. Morphogenesis of
fungus colonies in submerged shaken cultures. Am. J. Bot.32:424-431.
2. Camici, L., and G. Sermonti. 1952. Observations on Penicilliumchrysogenum in submerged culture. 1. Mycelial growth andautolysis. Bull. W.H.O. 6:265-276.
3. Clark, D. S., and C. P. Lentz. 1%3. Submerged citric acidfermentation of beet molasses in tank-type fermentors. Bio-
VOL. 49, 1985
.10,.,.o
.0-1.0,
Dow
nloa
ded
from
http
s://j
ourn
als.
asm
.org
/jour
nal/a
em o
n 31
Jan
uary
202
2 by
46.
70.3
8.15
3.

1506 MARTIN AND BAILEY
technol. Bioeng. 5:193-199.4. Clark, S. 1962. Submerged citric acid fermentation of ferrocya-
nide-treated beet molasses: morphology of pellets of Aspergillusniger. Can. J. Microbiol. 8:133-136.
5. Humfeld, H1. 1948. The production of mushroom mycelium(Agaricus campestris) in submerged culture. Science 107:373.
6. Kosaric, N., A. LeDuy, and J. E. Zajic. 1973. Submerged culturegrowth of edible mushrooms on waste sulphite liquors. Can. J.Chem. Eng. 51:186-190.
7. Kosaric, N., and N. Miyata. 1981. Growth of morel mushroommycelium in cheese whey. J. Dairy Res. 148:149-162.
8. Litchfield, J. H. 1967. Submerged production of mushroommycelium, p. 107-144. In H. J. Peppler (ed.), Microbial tech-nology. Reinhold Publishing Corp., Amsterdam.
9. Litchfield, J. H., R. C. Overbeck, and R. S. Davidson. 1963.Factors affecting the growth of morel mushroom mycelium insubmerged culture. J. Agric. Food Chem. 11:158-162.
10. Martin, A. M. 1983. Submerged production of Agaricus cam-
pestris mycelium in peat extracts. J. Food Sci. 48:206-207.11. Martin, A. M. 1983. Adaptation of Agaricus campestris NRRL
2334 to a peat extract medium. Can. J. Microbiol. 29:108-110.12. Martin, A. M., and V. I. Bailey. 1984. Study on the acid
hydrolysis of peat. 1. Extraction of carbohydrates. J. Chem.Technol. Biotechnol. 34:70-74.
13. Martin, S. M., and W. R. Waters. 1952. Production of citric acid
by submerged fermentation. Ind. Eng. Chem. 39:2229-2233.14. Metz, B., and N. W. Kossen. 1977. The growth of molds in the
form of pellets. A literature review. Biotechnol. Bioeng.19:781-799.
15. Molitoris, J. T. 1962. Bestimmung der Agaricus campestrisStamme NRRL 2334, 2335, 2336 als Beaveria tenella (De-lacroix, Siemenon). Mushroom Sci. 5:218-230.
16. Morris, D. L. 1948. Quantitative determination of carbohydrateswith Dreyword's anthrone reagent. Science 107:254-255.
17. Quierzy, P., N. Therien, and A. LeDuy. 1979. Production ofCandida utilis protein from peat extracts. Biotechnol. Bioeng.21:1175-1190.
18. Reusser, F., J. F. T. Spencer, and H. R. Sallans. 1958. Proteinand fat content of some mushrooms grown in submergedculture. Appl. Microbiol. 6:1-4.
19. Steel, R., S. M. Martin, and C. P. Lentz. 1954. A standardinoculum for citric acid production in submerged culture. Can.J. Microbiol. 1:150-157.
20. van Suijdam, J. C., N. W. F. Kossen, and P. G. Paul. 1980. Aninoculum technique for the production of pellets. Eur. J. Appl.Microbiol. Biotechnol. 10:211-221.
21. Whitaker, A., and P. A. Long. 1973. Fungal pelleting. ProcessBiochem. 8:27-31.
22. Worgan, J. T. 1968. Culture of the higher fungi. Prog. Ind.Microbiol. 8:73-139.
APPL. ENVIRON. MICROIBIOL.
Dow
nloa
ded
from
http
s://j
ourn
als.
asm
.org
/jour
nal/a
em o
n 31
Jan
uary
202
2 by
46.
70.3
8.15
3.





























