Embed Size (px)
DESCRIPTION
cinetica
Citation preview

JOURNAL OF BACTERIOLOGY, Feb. 2005, p. 923–929 Vol. 187, No. 30021-9193/05/$08.00�0 doi:10.1128/JB.187.3.923–929.2005Copyright © 2005, American Society for Microbiology. All Rights Reserved.
Growth Kinetics of Extremely Halophilic Archaea (FamilyHalobacteriaceae) as Revealed by Arrhenius Plots
Jessie L. Robinson,1 Brandy Pyzyna,1 Rachelle G. Atrasz,1 Christine A. Henderson,1Kira L. Morrill,1 Anna Mae Burd,1 Erik DeSoucy,1 Rex E. Fogleman III,1
John B. Naylor,1 Sarah M. Steele,1 Dawn R. Elliott,1Kathryn J. Leyva,2 and Richard F. Shand1*
Department of Biological Sciences, Northern Arizona University, Flagstaff,1 and Department ofMicrobiology, Arizona College of Osteopathic Medicine,
Midwestern University, Glendale,2 Arizona
Received 22 August 2004/Accepted 31 October 2004
Members of the family Halobacteriaceae in the domain Archaea are obligate extreme halophiles. They occupya variety of hypersaline environments, and their cellular biochemistry functions in a nearly saturated saltymilieu. Despite extensive study, a detailed analysis of their growth kinetics is missing. To remedy this,Arrhenius plots for 14 type species of the family were generated. These organisms had maximum growthtemperatures ranging from 49 to 58°C. Nine of the organisms exhibited a single temperature optimum, whilefive grew optimally at more than one temperature. Generation times at these optimal temperatures rangedfrom 1.5 h (Haloterrigena turkmenica) to 3.0 h (Haloarcula vallismortis and Halorubrum saccharovorum). Allshared an inflection point at 31 � 4°C, and the temperature characteristics for 12 of the 14 type species werenearly parallel. The other two species (Natronomonas pharaonis and Natronorubrum bangense) had significantlydifferent temperature characteristics, suggesting that the physiology of these strains is different. In addition,these data show that the type species for the family Halobacteriaceae share similar growth kinetics and arecapable of much faster growth at higher temperatures than those previously reported.
The family Halobacteriaceae in the domain Archaea pres-ently is comprised of 18 genera and 49 validly described species(International Committee on Systematics of Prokaryotes[http://www.the-icsp.org]). All members are extreme halo-philes, requiring at least 1.5 M NaCl for growth, but growoptimally in 2.0 to 4.5 M NaCl (6, 20). All have exceptionallyhigh internal cation concentrations that approach 6 M in somespecies (e.g., Halobacterium salinarum) (21). In addition toexhibiting halophilicity, four genera (Natronococcus, Natrono-monas, Natronorubrum, and Natronobacterium) are also alka-liphilic, growing optimally between pH 9.5 and 10.0 (20). Theseorganisms have been isolated from a wide variety of environ-ments, including the sediment of a cold, hypersaline lake inAntarctica (Halorubrum lacusprofundi) (3), the Dead Sea(Haloferax volcanii) (16), the Great Salt Lake (Halorabdusutahensis) (31), a hypersaline soda lake in Egypt (Natronomo-nas pharaonis) (27), and a salt crystal from a Permian halitedeposit (Halosimplex carlsbadense) (30). There is also a reportof haloarchaeal 16S rDNA amplified from inside a blacksmoker (28), but no haloarchaeons have been cultured fromthis source. All haloarchaea grow aerobically, and one species(Halobacterium salinarum) can grow phototrophically usingbacteriorhodopsin as a light-driven proton pump (19).
Since “. . .growth is the core of bacterial physiology. . .” (11)and since all experiments in cellular regulation are fundamen-tally physiological, it is essential to understand the growth
physiology of the organism one is studying. Toward this goalwe generated a complete Arrhenius plot for the haloarchaeonHaloferax mediterranei (22, 25; J. L. Robinson and R. F. Shand,unpublished data [http://jan.ucc.nau.edu/�shand]). Thisgrowth physiology experiment revealed the following: (i) theorganism is eurythermal, growing between 12 and 55°C withgeneration times of 752.5 and 1.67 h, respectively, (ii) it has anoptimal temperature plateau (from 47 to 51°C; gavg � 1.20 �0.01 h, where gavg represents average generation time) insteadof a single optimum temperature, and (iii) it has an inflectionpoint (i.e., a deviation from the Arrhenius portion of the plot)at 33°C. We also discovered that all physiologically based ex-periments involving this organism have been conducted undersuboptimal growth temperatures. The first (and only other)correlation between growth rate and temperature for a halo-archaeon was produced for Halorubrum lacusprofundi (3) byuse of a temperature gradient incubator. Unfortunately, bal-anced growth conditions were not established and optical den-sity readings at the higher growth temperatures were con-founded by flocculent growth.
Extremely halophilic Archaea possess eukaryote-like fea-tures such as multisubunit RNA polymerases, homologues toeukaryotic transcription factors, TATA-box promoters (18,29), and leaderless transcripts (4); these features make themdistinct from Bacteria at the molecular level. Moreover, giventhe diversity of environments from which the haloarchaea areisolated and the lack of detailed information regarding theirgrowth physiology, we generated Arrhenius plots for 14 typespecies. These data provide critical information for elucidatingtheir physiological, metabolic, and genetic features.
* Corresponding author. Mailing address: Department of BiologicalSciences, Northern Arizona University, Flagstaff, AZ 86011-5640.Phone: (928) 523-9970. Fax: (928) 523-7500. E-mail: [email protected].
923
on January 27, 2015 by guesthttp://jb.asm
.org/D
ownloaded from

MATERIALS AND METHODS
Haloarchaeal strains, media and growth conditions. The type species and theappropriate medium for each are listed in Table 1. At the start of this projectthere were 14 genera (20); since then, four new genera (Halorhabdus, Halomi-crobium, Halobiforma, and Halosimplex [http://www.the-icsp.org]) have beenadded to the family and are not represented in this study. Cultures were grownat 200 rpm in model G76 and C76 orbital shaking water baths (New BrunswickScientific Co. Inc., Edison, N.J.) with gabled covers (essential for reducingevaporation). Temperature was monitored with ASTM (American Society forTesting and Materials) thermometers accurate to �0.2°C. To promote efficientaeration, 125-ml tribaffled flasks (Kimble Glass, Vineland, N.J.) with Morton-type closures containing 25 ml of medium were used. Growth was monitoredspectrophotometrically using a dual-beam BioSpec 1601 (Shimadzu ScientificInstruments, Inc., Columbia, Md.). All samples with an optical density at 600nanometers (OD600) �0.10 were diluted to an OD600 between 0.05 and 0.10before reading. To mitigate volume effects (especially in cultures with slowgrowth rates), no more than 15 ml of culture was removed from the culture flask.A survey of growth media at 41°C for Haloarcula vallismortis, Halococcus mor-rhuae, and Natrinema pellirubrum included ATCC 1863, GN101, and NRC817, inaddition to the recommended medium (see Table 1 and Table 2) (8).
Criteria for establishing balanced growth and determining generation times.Prior to collection of Arrhenius plot data, cultures were placed into balancedgrowth (1): a minimum of eight generations in exponential phase (at least twosubcultures). To ensure they remained in balanced growth, the cultures weregrown from an OD600 of 0.01 to an OD600 of no more than 0.20 (�4.5 doublings).The starting temperature for each Arrhenius plot was 41°C. Once in balancedgrowth, the 41°C cultures were subcultured bidirectionally into aerated media in2°C increments; balanced growth was then reestablished. This was repeated untilboth the maximum temperature and 23°C were reached. Due to the prolongedgeneration times at lower temperatures, it was impractical to grow these organ-isms below 23°C (3, 22).
Minimum requirements for growth curves from which generation times weredetermined were as follows: (i) samples were taken at least once every genera-tion, (ii) each growth curve had a minimum of six time points, (iii) R2 was �0.995,and (iv) the generation times of the last two growth curves differed by �20%.Typically, it took one or two growth curves after balanced growth had beenestablished to meet these criteria; however, in some cases it took as many as 10.An organism was considered not able to grow when the OD600 remained �0.03and no growth occurred for 48 h.
Statistical analysis. A Student t test was used to determine whether differ-ences in temperature characteristics were significant (see Fig. 2). Since thecorrelation coefficients were remarkably high (R2 � 0.984) (see Table 3), thewithin-group variance was negligible. Therefore, the slopes were compared.
RESULTS AND DISCUSSION
Methodological rationale. Cultures were shifted from onetemperature to another in 2°C increments to prevent long lagsin growth and a heat shock or cold shock response. Heat shockgenes are essential for growing cells, and “[i]nduction of theheat shock proteins during pre-adaptation at moderate tem-peratures increases survival of the organism at higher temper-atures” (14).
The necessity for placing cultures into balanced growth priorto collecting data has been stated by Neidhardt et al. (17):“Unless growth is monitored throughout a physiological exper-iment, the results may not be reproducible. . . . In fact, a phys-iological experiment done with a poorly characterized cultureis all but useless.” We have shown that the growth rate ofHalobacterium salinarum NRC817 decreases from its initialgrowth rate as early as OD600 � �0.1 (25). However, this is anextreme example, and all organisms examined in this studygrew linearly to an OD600 of �0.2. Consequently, an OD600 of�0.2 was chosen as a conservative maximum at which to sub-culture in order to maintain balanced growth. This had theadditional benefits of (i) preventing spectrophotometric inter-ference due to accumulation of gas vesicles (26) and (ii) pre-venting physiological perturbations due to decreased oxygenavailability at higher temperatures (5) and pH changes in me-dia that were relatively unbuffered.
In context of microbial physiology, an Arrhenius plot takesthe form shown in Fig. 1D. The linear portion of the plot wheregrowth rate correlates with temperature is called the Arrhe-nius, physiological, or “normal” portion, and the slope of thisportion is called the temperature characteristic (11). Tradition-ally, temperature is plotted as 1,000/T (in degrees Kelvin) (9),however, we have converted to the more familiar degrees Cel-sius. The log of the specific growth rate constant (k � 0.693/g)is plotted on the abscissa.
Growth medium survey. Although we used published me-dium recipes for growing the organisms, we found that threespecies grew poorly (g � 8 h) in the recommended medium.For these organisms a survey was done to determine whethersome other medium would be better at supporting growth (see
TABLE 1. Strains, media, and sources
Type species Medium Sourcea
Haloarcula vallismortis ATCC 1863b,c ATCC 29715Halobacterium salinarum (NRC-1)d GN101(8) S. DasSarmaHalobaculum gomorrense ATCC 2169b ATCC 700876Halococcus morrhuae ATCC 1863b,c ATCC 43103Haloferax volcanii (DS70)d ATCC 1176b C. DanielsHalogeometricum borinquense ATCC 213b,e R. VreelandHalorubrum saccharovorum ATCC 876b,f R. VreelandHaloterrigena turkmenica ATCC 1863b,c R. VreelandNatrialba asiatica ATCC 805b,g C. LitchfieldNatrinema pellirubrum ATCC 1863b,c R. VreelandNatronobacterium gregoryi ATCC 1590h P. JablonskiNatronococcus occultus ATCC 1590h P. JablonskiNatronomonas pharaonis ATCC 1590h P. JablonskiNatronorubrum bangense ATCC 1590h JCM 10635
a ATCC, American Type Culture Collection; JCM, Japan Collection of Mi-croorganisms.
b These media were prepared by using ATCC recipes (http://www.atcc.org)with the following modifications: media were filter sterilized, and trace elements(0.2 �M CuSO4 · 5H2O, 11.6 �M Fe(NH4)2(SO4)2 · 6H2O, 1.8 �M MnSO4 ·H2O, 1.5 �M ZnSO4 · 7H2O) were added. Trace elements were aliquoted, storedat �20°C, and added just prior to inoculation. Trace elements were thawed onlyonce.
c FeCl2 and MnCl2 were excluded.d NRC-1 was chosen because its genome has been sequenced (18); DS70 was
chosen because it is widely used to study haloarchaeal genetics.e Made as one solution. CaCl2 · 2H2O was used in place of CaCl2 · 6H2O (filter
sterilized using a prefilter).f Autoclaved, cooled to room temperature, and then filter sterilized using a
prefilter.g FeSO4 was excluded.h These media were prepared by using ATCC recipes with the following modifi-
cations: media were filter sterilized and contained SL-6 trace elements (3.5 �MZnSO4 · 7H2O, 1.5 �M MnCl2 · 4H2O, 48.5 �M H3BO3, 8.4 �M CoCl2 · 6H2O, 0.59�M CuCl2 · 2H2O, 0.84 �M NiCl2 · 6H2O, 1.3 �M Na2MoO4 · H2O). Traceelements were aliquoted, stored at �20°C, and added just prior to inoculation. Traceelements were thawed only once. The medium was incubated at 41°C overnight,cooled to room temperature, and then filter sterilized using a prefilter.
TABLE 2. Growth medium survey
Strain Recommendedmedium (g in h)
Medium used(g in h)
Haloarcula vallismortis ATCC 112 (8.5) ATCC 1863 (4.1)Halococcus morrhuae ATCC 112a ATCC 1863 (3.4)Natrinema pellirubrum ATCC 330a ATCC 1863 (3.8)
a No growth after 20 h (Natrinema pellirubrum) or 48 h (Halococcus morrhuae)of incubation, but both eventually grew.
924 ROBINSON ET AL. J. BACTERIOL.
on January 27, 2015 by guesthttp://jb.asm
.org/D
ownloaded from
Materials and Methods). Table 2 shows that all three of thesestrains grew faster (g � 4.1 h) in a medium other than thatusually associated with these organisms, and these media wereused in this study.
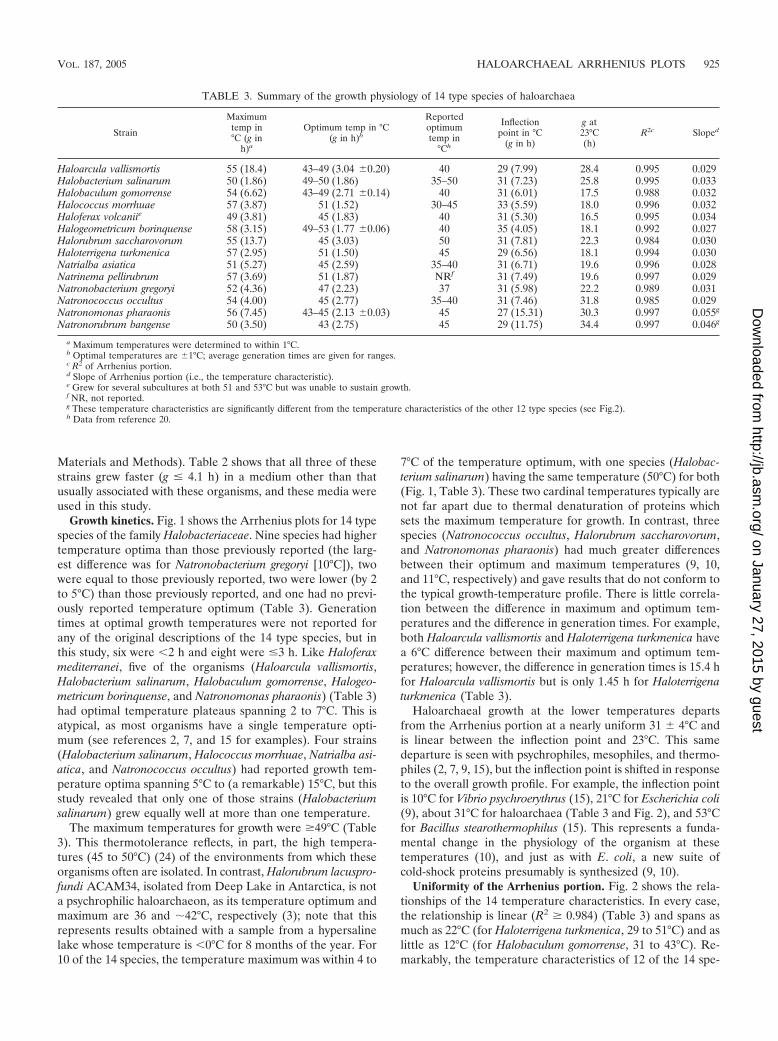
Growth kinetics. Fig. 1 shows the Arrhenius plots for 14 typespecies of the family Halobacteriaceae. Nine species had highertemperature optima than those previously reported (the larg-est difference was for Natronobacterium gregoryi [10°C]), twowere equal to those previously reported, two were lower (by 2to 5°C) than those previously reported, and one had no previ-ously reported temperature optimum (Table 3). Generationtimes at optimal growth temperatures were not reported forany of the original descriptions of the 14 type species, but inthis study, six were �2 h and eight were �3 h. Like Haloferaxmediterranei, five of the organisms (Haloarcula vallismortis,Halobacterium salinarum, Halobaculum gomorrense, Halogeo-metricum borinquense, and Natronomonas pharaonis) (Table 3)had optimal temperature plateaus spanning 2 to 7°C. This isatypical, as most organisms have a single temperature opti-mum (see references 2, 7, and 15 for examples). Four strains(Halobacterium salinarum, Halococcus morrhuae, Natrialba asi-atica, and Natronococcus occultus) had reported growth tem-perature optima spanning 5°C to (a remarkable) 15°C, but thisstudy revealed that only one of those strains (Halobacteriumsalinarum) grew equally well at more than one temperature.
The maximum temperatures for growth were �49°C (Table3). This thermotolerance reflects, in part, the high tempera-tures (45 to 50°C) (24) of the environments from which theseorganisms often are isolated. In contrast, Halorubrum lacuspro-fundi ACAM34, isolated from Deep Lake in Antarctica, is nota psychrophilic haloarchaeon, as its temperature optimum andmaximum are 36 and �42°C, respectively (3); note that thisrepresents results obtained with a sample from a hypersalinelake whose temperature is �0°C for 8 months of the year. For10 of the 14 species, the temperature maximum was within 4 to
7°C of the temperature optimum, with one species (Halobac-terium salinarum) having the same temperature (50°C) for both(Fig. 1, Table 3). These two cardinal temperatures typically arenot far apart due to thermal denaturation of proteins whichsets the maximum temperature for growth. In contrast, threespecies (Natronococcus occultus, Halorubrum saccharovorum,and Natronomonas pharaonis) had much greater differencesbetween their optimum and maximum temperatures (9, 10,and 11°C, respectively) and gave results that do not conform tothe typical growth-temperature profile. There is little correla-tion between the difference in maximum and optimum tem-peratures and the difference in generation times. For example,both Haloarcula vallismortis and Haloterrigena turkmenica havea 6°C difference between their maximum and optimum tem-peratures; however, the difference in generation times is 15.4 hfor Haloarcula vallismortis but is only 1.45 h for Haloterrigenaturkmenica (Table 3).
Haloarchaeal growth at the lower temperatures departsfrom the Arrhenius portion at a nearly uniform 31 � 4°C andis linear between the inflection point and 23°C. This samedeparture is seen with psychrophiles, mesophiles, and thermo-philes (2, 7, 9, 15), but the inflection point is shifted in responseto the overall growth profile. For example, the inflection pointis 10°C for Vibrio psychroerythrus (15), 21°C for Escherichia coli(9), about 31°C for haloarchaea (Table 3 and Fig. 2), and 53°Cfor Bacillus stearothermophilus (15). This represents a funda-mental change in the physiology of the organism at thesetemperatures (10), and just as with E. coli, a new suite ofcold-shock proteins presumably is synthesized (9, 10).
Uniformity of the Arrhenius portion. Fig. 2 shows the rela-tionships of the 14 temperature characteristics. In every case,the relationship is linear (R2 � 0.984) (Table 3) and spans asmuch as 22°C (for Haloterrigena turkmenica, 29 to 51°C) and aslittle as 12°C (for Halobaculum gomorrense, 31 to 43°C). Re-markably, the temperature characteristics of 12 of the 14 spe-
TABLE 3. Summary of the growth physiology of 14 type species of haloarchaea
Strain
Maximumtemp in°C (g in
h)a
Optimum temp in °C(g in h)b
Reportedoptimumtemp in
°Ch
Inflectionpoint in °C
(g in h)
g at23°C(h)
R2c Sloped
Haloarcula vallismortis 55 (18.4) 43–49 (3.04 �0.20) 40 29 (7.99) 28.4 0.995 0.029Halobacterium salinarum 50 (1.86) 49–50 (1.86) 35–50 31 (7.23) 25.8 0.995 0.033Halobaculum gomorrense 54 (6.62) 43–49 (2.71 �0.14) 40 31 (6.01) 17.5 0.988 0.032Halococcus morrhuae 57 (3.87) 51 (1.52) 30–45 33 (5.59) 18.0 0.996 0.032Haloferax volcaniie 49 (3.81) 45 (1.83) 40 31 (5.30) 16.5 0.995 0.034Halogeometricum borinquense 58 (3.15) 49–53 (1.77 �0.06) 40 35 (4.05) 18.1 0.992 0.027Halorubrum saccharovorum 55 (13.7) 45 (3.03) 50 31 (7.81) 22.3 0.984 0.030Haloterrigena turkmenica 57 (2.95) 51 (1.50) 45 29 (6.56) 18.1 0.994 0.030Natrialba asiatica 51 (5.27) 45 (2.59) 35–40 31 (6.71) 19.6 0.996 0.028Natrinema pellirubrum 57 (3.69) 51 (1.87) NRf 31 (7.49) 19.6 0.997 0.029Natronobacterium gregoryi 52 (4.36) 47 (2.23) 37 31 (5.98) 22.2 0.989 0.031Natronococcus occultus 54 (4.00) 45 (2.77) 35–40 31 (7.46) 31.8 0.985 0.029Natronomonas pharaonis 56 (7.45) 43–45 (2.13 �0.03) 45 27 (15.31) 30.3 0.997 0.055g
Natronorubrum bangense 50 (3.50) 43 (2.75) 45 29 (11.75) 34.4 0.997 0.046g
a Maximum temperatures were determined to within 1°C.b Optimal temperatures are �1°C; average generation times are given for ranges.c R2 of Arrhenius portion.d Slope of Arrhenius portion (i.e., the temperature characteristic).e Grew for several subcultures at both 51 and 53°C but was unable to sustain growth.f NR, not reported.g These temperature characteristics are significantly different from the temperature characteristics of the other 12 type species (see Fig.2).h Data from reference 20.
VOL. 187, 2005 HALOARCHAEAL ARRHENIUS PLOTS 925
on January 27, 2015 by guesthttp://jb.asm
.org/D
ownloaded from

cies are nearly parallel (Fig. 2 and Table 3), with the sloperanging between 0.027 and 0.034, a difference of only 0.007.The temperature characteristic for Haloferax mediterranei(slope � 0.033) also falls within this group (data not shown).The two notable exceptions are haloalkaliphiles, Natronomo-nas pharaonis and Natronorubrum bangense (Fig. 2 and Table3). The difference in the temperature characteristics for thesetwo species from the results obtained with the other 12 isstatistically significant (P � 0.001) and suggests that the phys-iology of these two organisms is fundamentally different. Whilealkaliphilicity might be a contributing factor, the other twohaloalkaliphiles (Natronobacterium gregoryi and Natronococcusoccultus) group with the 10 neutrophiles. For comparison, wedetermined the temperature characteristics for Halorubrumlacusprofundi (slope � 0.040; R2 � 0.943) and E. coli (slope �0.032; R2 � 0.981) from published data (3, 9). The tempera-ture characteristic for Halorubrum lacusprofundi was signifi-
cantly different (P � 0.001) from those of the 12 haloarchae-ons, but, interestingly, that of E. coli was not.
Significance. The heavy lifting has been done with respect todetermining optimum growth temperatures for several speciesin the family Halobacteriaceae. Optimization experiments forany other parameter can now be done in a single water bath.Although we did not conduct an extensive survey to determinewhether other species would grow better in a medium differentthan that recommended, we suspect that for many of the or-ganisms listed in Table 1 there are media that will yield fastergeneration times.
The prevailing view is that the haloarchaea “. . .have rela-tively long generation times (e.g., 3 to 4 h for Haloferax volcanii;8 to 12 h for Halobacterium spp.)” (Halohandbook [http://www.microbiol.unimelb.edu.au/micro/staff/mds/HaloHandbook]; seealso references 12 and 13). However, under optimal growth tem-peratures, these organisms grow much faster than has been as-
FIG. 1. Arrhenius plots for 14 type species of the family Halobacteriaceae (k � 0.693/g). — � —, regression line for the Arrhenius portion; � � � � ,extrapolation toward minimum growth temperature. (D) a, maximum temperature; b, optimum temperature; c, temperature at the inflection point;AP, Arrhenius portion.
926 ROBINSON ET AL. J. BACTERIOL.
on January 27, 2015 by guesthttp://jb.asm
.org/D
ownloaded from
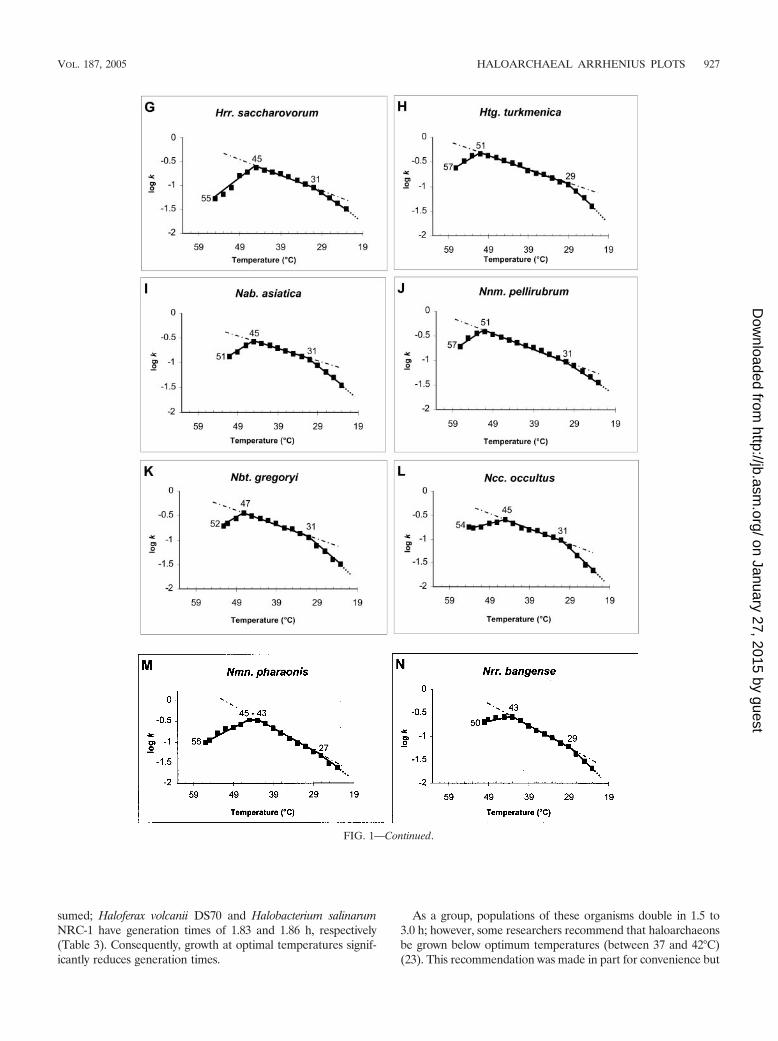
sumed; Haloferax volcanii DS70 and Halobacterium salinarumNRC-1 have generation times of 1.83 and 1.86 h, respectively(Table 3). Consequently, growth at optimal temperatures signif-icantly reduces generation times.
As a group, populations of these organisms double in 1.5 to3.0 h; however, some researchers recommend that haloarchaeonsbe grown below optimum temperatures (between 37 and 42°C)(23). This recommendation was made in part for convenience but
FIG. 1—Continued.
VOL. 187, 2005 HALOARCHAEAL ARRHENIUS PLOTS 927
on January 27, 2015 by guesthttp://jb.asm
.org/D
ownloaded from

also for concern about oxygen solubility in high salt. While oxygensolubility might be an issue at temperatures near the maximum,practical steps for enhancing oxygen diffusion at optimal temper-atures (e.g., the use of baffled culture flasks) have been describedpreviously (25).
In addition to cardinal growth temperature and generationtime results, careful growth studies can reveal much about thephysiology of an organism. The temperature characteristics forHalorubrum lacusprofundi, Natronomonas pharaonis, and Na-tronorubrum bangense suggest there is something fundamen-tally different about their physiology that deserves furtherstudy.
ACKNOWLEDGMENTS
This work was supported by Public Health Service grantGM05600-02 from the National Institutes of Health and a grant fromthe Arizona Board of Regents’ Technology and Research InitiativeFunds (Proposition 301) to R.F.S. K.J.L. was supported through In-tramural Grants from Midwestern University.
We thank Chuck Daniels, Shil DasSarma, Peter Jablonski, CarolLitchfield, and Russ Vreeland for supplying strains and Stephen Shus-ter for help with statistical analysis.
REFERENCES
1. Campbell, A. 1957. Synchronization of cell division. Bacteriol. Rev. 21:263–272.
2. Chablain, P. A., G. Philippe, A. Groboillot, N. Truffaut, and J. F. Guespin-Michel. 1997. Isolation of a soil psychrotrophic toluene-degrading Pseudo-monas strain: influence of temperature on the growth characteristics ondifferent substrates. Res. Microbiol. 148:153–161.
3. Franzmann, P. D., E. Stackebrandt, K. Sanderson, J. K. Volkman, D. E.Cameron, P. L. Stevenson, T. A. McKeen, and H. R. Burton. 1988. Halobac-terium lacusprofundi sp. nov., a halophilic bacterium isolate from Deep Lake,Antarctica. Syst. Appl. Microbiol. 11:20–27.
4. Fuglsang, A. 2004. Compositional nonrandomness upstream of start codonsin archaebacteria. Gene 332:89–95.
5. Gary, K. 1989. On-line electrochemical sensors in fermentation. Am. Bio-technol. Lab. 7:26–33.
6. Grant, W. D., and H. Larsen. 1989. Family Halobacteriaceae Gibbons 1974,269AL, p. 2218–2232. In J. T. Stanley, M. P. Bryant, N. Pfennig, and J. G.Holt (ed.), Bergey’s manual of systematic bacteriology, vol. 3. Williams &Wilkins, Baltimore, Md.
7. Guillou, C., and J. F. Guespin-Michel. 1996. Evidence for two domains ofgrowth temperature for the psychrotrophic bacterium Pseudomonas fluore-scens MF0. Appl. Environ. Microbiol. 62:3319–3324.
8. Haseltine, C., T. Hill, R. Montalvo-Rodriguez, S. K. Kemper, R. F. Shand,and P. Blum. 2001. Secreted euryarchaeal microhalocins kill hyperthermo-philic crenarchaea. J. Bacteriol. 183:287–291.
9. Herendeen, S. L., R. A. VanBogelen, and F. C. Neidhardt. 1979. Levels ofmajor proteins of Escherichia coli during growth at different temperatures. J.Bacteriol. 139:185–194.
10. Ingraham, J. L., and A. G. Marr. 1996. Effect of temperature, pressure, pH,and osmotic stress on growth, p. 1570–1578. In F. C. Neidhardt, R. CurtissIII, J. L. Ingraham, E. C. C. Lin, K. B. Low, B. Magasanik, W. S. Reznikoff,M. Riley, M. Schaechter, and H. E. Umbarger (ed.), Escherichia coli andSalmonella cellular and molecular biology, 2nd ed., vol. 2. ASM Press, Wash-ington, D.C.
11. Ingraham, J. L., O. Maaløe, and F. C. Neidhardt. 1983. Growth of thebacterial cell, p. ix, 227–265. Sinauer Associates, Inc., Sunderland, Mass.
12. Juez, G. 1988. Taxonomy of extremely halophilic archaebacteria, p. 3–24. InF. Rodriguez-Valera (ed.), Halophilic bacteria, vol. II. CRC Press, BocaRaton, Fla.
13. Kushner, D. J. 1993. Growth and nutrition of halophilic bacteria, p. 87–103.In R. H. Vreeland and L. I. Hochstein (ed.), The biology of halophilicbacteria. CRC Press, Boca Raton, Fla.
14. Lengeler, J. W., G. Drews, and H. G. Schlegel (ed.). 1999. Biology of theprokaryotes, p. 658–660. Thieme, Stuttgart, Germany.
15. Mohr, P. W., and S. Krawiec. 1980. Temperature characteristics and Arrhe-nius plots for nominal psychrophiles, mesophiles and thermophiles. J. Gen.Microbiol. 121:311–317.
16. Mullakhanbhai, M. F., and H. Larsen. 1975. Halobacterium volcanii spec.nov., a Dead Sea halobacterium with a moderate salt requirement. Arch.Microbiol. 104:201–214.
17. Neidhardt, F. C., J. L. Ingraham, and M. Schaechter. 1990. Physiology of thebacterial cell, p. 216, 227–229. Sinauer Associates, Inc., Sunderland, Mass.
18. Ng, W. V., S. P. Kennedy, G. G. Mahairas, B. Berquist, M. Pan, H. D.Shukla, S. R. Lasky, N. S. Baliga, V. Thorsson, J. Sbrogna, S. Swartzell, D.Weir, J. Hall, T. A. Dahl, R. Welti, Y. A. Goo, B. Leithauser, K. Keller, R.Cruz, M. J. Danson, D. W. Hough, D. G. Maddocks, P. E. Jablonski, M. P.Krebs, C. M. Angevine, H. Dale, T. A. Isenbarger, R. F. Peck, M. Pohlschro-der, J. L. Spudich, K. H. Jung, M. Alum, T. Freitas, S. Hou, C. J. Daniels,P. P. Dennis, A. D. Omer, H. Ebhardt, T. M. Lowe, P. Liang, M. Riley, L.
FIG. 2. Comparison of the temperature characteristics of the 14 type species of Halobacteriaceae listed in Table 1. ■, Natronomonas pharaonis;Œ, Natronorubrum bangense; ----, temperature characteristics of the other 12 type species (see Fig. 1 and Table 3 for details).
928 ROBINSON ET AL. J. BACTERIOL.
on January 27, 2015 by guesthttp://jb.asm
.org/D
ownloaded from

Hood, and S. DasSarma. 2000. Genome sequence of Halobacterium speciesNRC-1. Proc. Natl. Acad. Sci. USA 97:12176–12181.
19. Oesterhelt, D., and G. Krippahl. 1983. Phototrophic growth of halobacteriaand its use for isolation of photosynthetically-deficient mutants. Ann. Inst.Pasteur Microbiol. 134B:137–150.
20. Oren. A. 2001. The order Halobacteriales, p. 2, 6–19. In M. Dworkin, S.Falkow, E. Rosenberg, K.-H. Schleifer, and E. Stackebrandt (ed.), Theprokaryotes. A handbook on the biology of bacteria: ecophysiology, isola-tion, identification, applications, 3rd ed. Springer-Verlag, New York, N.Y.[Online.]
21. Oren, A. 2002. Halophilic microorganisms and their environments, p. 211.Kluwer Academic Publishers, Dordrecht, The Netherlands.
22. Perez, A. M. 2000. Growth physiology of Haloferax mediterranei R4 andpurification of halocin H4. M.S. thesis. Northern Arizona University, Flag-staff, Ariz.
23. Rodriguez-Valera, F. 1995. Cultivation of halophilic archaea, p. 13–16. InF. T. Robb, A. R. Place, K. R. Sowers, H. J. Schreier, S. DasSarma, andE. M. Fleishmann (ed.), Archaea, a laboratory manual. Cold Spring HarborLaboratory Press, Plainview, N.Y.
24. Rodriguez-Valera, F. 1993. Introduction to saline environments, p. 1–23. InR. H. Vreeland and L. I. Hochstein (ed.), The biology of halophilic bacteria.CRC Press, Boca Raton. Fla.
25. Shand, R. F., and A. M. Perez. 1999. Haloarchaeal growth kinetics, p.
411–424. In J. Seckbach (ed.), Enigmatic microorganisms and life in extremeenvironments. Kluwer Academic Publishers, Dordrecht, The Netherlands.
26. Shand, R. F., and M. C. Betlach. 1991. Expression of the bop gene cluster ofHalobacterium halobium is induced by low oxygen tension and by light. J.Bacteriol. 173:4692–4699.
27. Soliman, G. S. H., and H. G. Truper. 1982. Halobacterium pharaonis sp. nov.,a new, extremely haloalkaliphilic archaebacterium with low magnesium re-quirement. Zentbl. Bakteriol. Parasitenkd. Infektionskr. Hyg. Abt. 1 Orig.3:318–329.
28. Takai, K., T. Komatsu, F. Inagaki, and K. Horikoshi. 2001. Distribution ofarchaea in a black smoker chimney structure. Appl. Env. Microbiol. 67:3618–3629.
29. Thompson, D. K., J. R. Palmer, and C. J. Daniels. 1999. Expression andheat-responsive regulation of a TFIIB homologue for the archaeon Haloferaxvolcanii. Mol. Microbiol. 33:1081–1092.
30. Vreeland, R. H., S. Straight, J. Krammes, K. Dougherty, W. D. Rosenzweig, andM. Kamekura. 2002. Halosimplex carlsbadense gen. nov., sp. nov., a uniquehalophilic archaeon, with three 16S rRNA genes, that grows only in definedmedium with glycerol and acetate or pyruvate. Extremophiles 6:445–452.
31. Waino, M., B. J. Tindall, and K. Ingvosen. 2000. Halorhabdus utahensis gen.nov., sp. nov., an aerobic, extremely halophilic member of the Archaea fromGreat Salt Lake, Utah. Int. J. Syst. Evol. Microbiol. 50:183–190.
VOL. 187, 2005 HALOARCHAEAL ARRHENIUS PLOTS 929
on January 27, 2015 by guesthttp://jb.asm
.org/D
ownloaded from