Embed Size (px)
Citation preview

APPLIED AND ENVIRONMENTAL MICROBIOLOGY, May 1983, P. 1614-1621 Vol. 45, No. 50099-2240/83/051614-08$02.00/0Copyright © 1983, American Society for Microbiology
Growth Characteristics of Saccharomyces rouxii Isolated fromChocolate Syrup
L. RESTAINO,* STEPHANIE BILLS, KARIN TSCHERNEFF, AND LAWRENCE M. LENOVICHHershey Foods Corporation, Hershey, Pennsylvania 17033
Received 2 November 1982/Accepted 19 January 1983
We investigated the growth parameters of Saccharomyces rouxii isolated fromspoiled chocolate syrup. The optimum pH range for S. rouxii was 3.5 to 5.5,whereas the minimum and maximum pH values that permitted growth were 1.5and 10.5, respectively. For cells grown in 0 and 60% sucrose the optimum wateractivity (a,) values were 0.97 and 0.96, respectively. The optimum temperaturefor S. rouxii increased with a decreasing a, regardless of whether glucose orsucrose was used as the humectant. The optimum temperatures for S. rouxii were28°C at an a. of >0.995 and 35°C at an a, of 0.96 to 0.90 in 2x potato dextrosebroth with sucrose. Increasing the sorbate concentration (from 0.03 to 0.10%)caused the growth of S. rouxii to become more inhibited between a,s of >0.995and 0.82. S. rouxii did not grow when the sorbate level was 0.12% (wt/vol). Atlower sorbate levels, the effect of sorbate on the growth of S. rouxii depended onthe a, level. Lowering the a, enhanced the resistance of S. rouxii to increasingconcentrations of potassium sorbate. Permeability and polyol production arediscussed with respect to sorbate tolerance of S. rouxii at different a, levels.
The microorganisms that spoil foods contain-ing a low water activity (aw), high acidity, lowredox potential, and high carbon/nitrogen ratioare osmotolerant yeasts (31, 37). Since theseyeasts compete poorly with other microorga-nisms in nonselective environments, these pa-rameters become important in favoring thegrowth of osmotolerant yeasts. Osmotolerantyeasts are the main spoilage organisms in foodssuch as honey, maple syrup, raw sugar cane,fruit syrups, candy, jams, jellies (37), soymashes (24), fondants (28), fruit juice concen-trates (30), dried fruits (21, 22), and chocolatesyrup (unpublished data). Contamination ofthese food products could originate from highlycontaminated raw ingredients, inadequate sani-tation due to the difficulty of cleaning a high-sugar commodity from equipment, improperpackaging and storage facilities, and insects dur-ing all phases of production. The spoiled productusually becomes turbid with a strong alcoholicaroma. If the product is enclosed in a hermeti-cally sealed container, the package will swelldue to carbon dioxide production. Growth ofosmotolerant yeasts could reduce the solid con-tent and alter the solubility of solutes that causethe aw of the food to increase.Osmotolerant yeasts comprise, primarily,
members of the genus Saccharomyces, the ma-jor species of which include S. rouxii, S. bailii,and S. bisporus. Strains tolerant to very highconcentrations of solutes belong to the species
S. rouxii (27, 36), the most common spoilageorganism of the osmotolerant yeasts. The mini-mum a, range of this yeast in a fructose syrup atpH 4.8 (35) and in glucose broths (32) is 0.62 to0.65, whereas the optimum aw in sucrose brothsis 0.98 (R. H. Tilbury, M.S. thesis, University ofBristol, England). The solute has an influence onthe growth of S. rouxii, for which the limitingconcentrations are 20 to 22% for NaCl, 80% forglucose, and 80% for sucrose (24). S. rouxii andother osmotolerant yeasts are classified as me-sophiles which can grow at refrigeration tem-peratures (32). The pH range for most osmoto-lerant and non-osmotolerant yeasts is 2.0 to 7.0with an optimum pH of 4.0 to 4.5 (32). Ingram(16) has shown a direct relationship between thetolerance of osmotolerant yeasts to high acidityand reduced aw levels, where the lower the aw,the less tolerant the microorganism is to acidity.Further, Baird-Parker and Kooiman (3) havestudied the coupled effects of sorbic acid, pH,and a, on the growth of yeasts. Their datashowed that an increase in sucrose (decrease ina,) and a decrease in pH cause sorbate tobecome more effective against the growth ofyeasts. However, the antimycotic effects of po-tassium sorbate to the growth of S. rouxii de-pend on the preconditioning of this yeast tosorbate or other weak organic acids (5; J. I. Pitt,personal communication).The objective of this investigation was to
elucidate various growth parameters of an osmo-
1614
on June 25, 2018 by guesthttp://aem
.asm.org/
Dow
nloaded from

GROWTH PARAMETERS OF S. ROUXII 1615
tolerant, sorbate-resistant S. rouxii strain isolat-ed from chocolate syrup.
MATERIALS AND METHODSYeast strain and identification scheme. S. rouxii
NRRL Y-12691, isolated from chocolate syrup con-taining 0.1% potassium sorbate, was used in thisinvestigation. Stock cultures were adapted and main-tained on slants of 2x potato dextrose agar (PDA[Difco Laboratories]) (12% glucose added) without orwith 60%o (wt/vol) sucrose through at least four trans-fers and stored at 4°C. Stock cultures were transferredonce every 2 months.The identification of the yeast isolate was made by
using the carbohydrate fermentation scheme describedby Lodder (18), a DNA hybridization assay (17), andsensitivity to 0.5% acetic acid in malt extract agar plus2% glucose (26; J. I. Pitt, personal communication).The yeast isolate rapidly (within 5 days) fermentedglucose, maltose, and fructose and slowly (28 days)utilized sucrose. After 7 days of incubation at 25°C onmalt extract agar containing 0.5% acetic acid, novisible growth of this yeast strain was observed,indicating sensitivity to this weak organic acid. TheDNA hybridization assay as described by Kurtzman etal. (17) showed a 98% DNA base sequence comple-mentarity with the type strain, S. rouxii NRRL Y-229.
Preparation of inoculum. S. rouxii cells were trans-ferred from slants to 2 x potato dextrose broth (PDB).Cells maintained on 2x PDA-12% glucose were trans-ferred to 2x PDB-12% glucose, whereas cells from 2xPDA-12% glucose slants with 60o (wt/vol) sucrosewere added to 2x PDB-12% glucose containing thecorresponding sucrose level. For the experiments in-volving optimum aw, optimum pH, and pH range,inocula grown in the presence and absence of 60%osucrose were used, whereas for the other experiments,only cells grown in 2x PDB-12% glucose withoutsucrose were used. The flasks containing the mediumplus the inoculum were incubated at 25°C in a shakingwater bath (160 rpm) for 2 days. Stationary-phase cellswere transferred to sterile 500-ml polypropylene cen-trifuge bottles. Cells grown in 60%o sucrose werediluted 1:10 with sterile distilled water to reduce theviscosity. The inocula were centrifuged at 4 to 8°C for30 min at 6,000 x g. The supernatant liquid wasdiscarded, and the pellet was resuspended in 10 to 15ml of sterile distilled water. The absorbancy of thecells was adjusted to 0.500 at 420 nm, using a Bausch& Lomb Spectronic 20 colorimeter, which corre-sponded to a cell concentration ranging from 4.0 x 105to 1.0 x 106 cells per ml. Volumes of 0.3 to 0.35 ml and0.7 ml of cells grown in medium containing 0 and 60%osucrose, respectively, were used to inoculate 80-mlvolumes of the various growth media.
Preparation of growth media. The basal medium forall growth experiments was 2x PDB-12% glucose,which was prepared according to manufacturer'sspecifications. For the experiments involving optimumaw, 200-ml volumes of the basal growth medium wereformulated with various sucrose levels (expressedthroughout as wt/vol). Glucose and sucrose wereadded separately and dissolved with the aid of heat.After boiling, duplicate 80-ml volumes for each su-crose level were dispensed into 250-ml Fernbachflasks and autoclaved at 121°C for 10 min. The pH of
the media was adjusted to 5.0 by using sterile HCI orNaOH. After adjustments, a, levels were determinedfor each sucrose level by using a Beckman Hygrolinerecorder (model VFB). The corresponding sucroseand a, levels were 0% sucrose, >0.995 a,; 6% su-crose, 0.995 a,; 12% sucrose, 0.99 a,; 18% sucrose,0.98 a,; 23% sucrose, 0.97 a,; 27% sucrose, 0.96 a,;30% sucrose, 0.95 a,; 40% sucrose, 0.92 a,; and 50%sucrose, 0.90 a,. The flasks were stored at roomtemperature with the caps wrapped with Parafilm toprevent a change in aw. After inoculation, the flaskswere incubated in a shaking water bath (160 rpm) at250C.For the experiments involving optimum, minimum,
and maximum pH values, 3 liters of the basal mediumsupplemented with 23 or 27% sucrose was formulatedand autoclaved at 121°C for 10 min. With a sterile 250-ml graduated cylinder, 160-ml volumes were dispensedinto sterile 250-ml Fembach flasks. pH values rangingfrom 1.0 to 12.0 (in increments of 0.5 pH units) wereobtained with sterile HCl or NaOH. Different concen-trations of HCl and NaOH were used to add thesmallest possible amount of the base or acid to obtainthe desired pH. After pH adjustment, the a, wasdetermined; 23% sucrose gave an aw of 0.97, and 27%sucrose gave an aw of 0.96. S. rouxii cells previouslygrown in 0 or 60%o glucose were inoculated into thepH-adjusted medium, and the flasks were incubated at25°C in a shaking water bath (160 rpm). The presenceof visible growth was monitored daily for 30 days.
Double-strength PDB with 0% sucrose-0o glucose,23% sucrose-0o glucose, 09%o sucrose-12% glucose, orthe above-mentioned sucrose and glucose levels nec-essary to obtain a, levels of 0.98, 0.97, 0.96, 0.95,0.92, and 0.90 was used to determine the optimumtemperature versus the a, level of humectant. Thegrowth medium was formulated as in the experimentsinvolving optimum a,. The flasks were placed in 600-ml beakers containing 30 ml of distilled water toreduce temperature fluctuations during incubation.Flasks (one from each growth medium) were placed inincubators at 22, 25, 28, 32, 35, 37, and 40°C. Afterequilibrating for 24 h, the media were inoculated withS. rouxii previously grown in 0% sucrose.For studying the effects of the potassium sorbate
concentration on the growth of S. rouxii at various awlevels, the basal growth medium plus various sucroseconcentrations as described for the experiments onoptimum a, was used. In addition, sucrose concentra-tions of 60 and 70o (wtlvol), corresponding to aws of0.86 and 0.82, respectively, were used. Portions (79ml) of the basal growth medium with appropriatesucrose concentrations were dissolved in 250-ml Fern-bach flasks and autoclaved at 121°C for 10 min.Potassium sorbate granules were dissolved in distilledwater at concentrations of 2.4, 4.8, 6.4, 8.0, 9.6, and12.0o (wt/vol) on the day of use. These solutions werefilter sterilized through a 0.2-,um membrane. After thegrowth media equilibrated to room temperature, 1.0 mlwas added to each aw level to yield final sorbateconcentrations of 0.03, 0.06, 0.08, 0.10, 0.12, and0.15%. An equal volume of sterile distilled water wasadded to media in duplicate flasks to yield a controlwithout sorbate. The media were adjusted to pH 5.0,and the aw levels were calculated. The flasks contain-ing the various media were inoculated with S. rouxiiand incubated statically in an air incubator at 320C.
VOL. 45, 1983
on June 25, 2018 by guesthttp://aem
.asm.org/
Dow
nloaded from

1616 RESTAINO ET AL.
2.3(
2.4(
2.5(
2.6(
45-2.70-w
2.80-
Z 2.90
0
13.00
W 3.10z
w
O3.20
3.30
3.40
3.50
1.00 .98 .96 .94 .92 .90 .88
WATER ACTIVITYFIG. 1. Optimum a, levels for S. rouxii cells pre-
conditioned in 0 or 60% sucrose. Cells were grown tothe stationary phase in 2x PDB-12% glucose plus 0%(@) or 60% (U) sucrose.
The presence of visible growth was noted daily for 47days.
Plating media, diluent, and enumeration procedures.Plating media and diluents with sucrose were utilizedto reduce osmotic shock. Basal phosphate buffer (0.3mM, pH 7.2) diluent and PDA (with 2% glucose added)plating medium were used in all experiments. Regulardiluent and plating medium were used when thegrowth medium contained 0 or 6% sucrose. A 40%(wt/vol) sucrose-phosphate buffer (pH 5.1; a,, 0.95)was used as the diluent with 12, 18, 23, 27, or 30%sucrose in the growth medium, whereas 60% (wt/vol)sucrose-phosphate buffer (pH 5.1; aw, 0.93) was usedto dilute growth medium containing 40, 50, 60, or 70%sucrose. PDA plating medium containing 30% sucrose(pH 5.2; aw, 0.99), 45% sucrose (pH 5.2; aw, 0.95), 50%sucrose (pH 5.2; a,, 0.94), 60% sucrose (pH 5.2; aw,0.92), and 65% sucrose (pH 5.2; aw, 0.90) was used toenumerate S. rouxii cells from growth media havingaws of 0.97 to 0.98, 0.96, 0.95, 0.90 and 0.92, and 0.82and 0.86, respectively.The pour plate technique was utilized to measure
the growth of the osmotolerant S. rouxii cells. Atvarious time intervals, 1-ml volumes were pipettedfrom the different growth media, serially diluted in theappropriate diluent, and plated in duplicate on thecorresponding PDA with sucrose. Plates were incubat-ed at 25°C for a minimum of 5 days. Yeast counts were
transformed to loglo yeast cells per milliliter. The datawere analyzed statistically by comparing the geomet-ric means of duplicate samples. When appropriate,linear regression curves were determined from data forexponentially growing S. rouxii cells. A 95% confi-
dence level was used on all linear regression curves.Growth and death rates (generations per hour) andgeneration times (in hours [reciprocals of growth ordeath rates]) were computed from the linear regressioncurves.
RESULTS
Optimum a, levels. The optimum a, levels ofS. rouxii preconditioned in 0 and 60% sucroseare presented in Fig. 1. S. rouxii cells werepreconditioned in the supplemented basal medi-um with 0 or 60% sucrose for at least fourtransfers. The preconditioning of S. rouxii cellsto 0 or 60% sucrose influenced the growth ratepattern over a pH range. At pH 5.0, the optimumaWs for S. rouxii cells were 0.97 and 0.96 for cellspreviously grown in 0 and 60% sucrose, respec-tively. Optimum a, levels were determined withglucose and sucrose as the humectants.Growth response to S. rouxii at various pH
levels. The effects of different pH levels on thegrowth of S. rouxii cells previously grown in 0and 60% sucrose are presented in Table 1. Thepreconditioning of S. rouxii in 0 or 60% sucrose
did not influence the growth response of S.rouxii to different pH levels. The minimum andmaximum pH values for growth of S. rouxii in2x PDB-12% glucose were 1.5 and 10.5, respec-tively. Since the generation times were similarthrough several consecutive pH values for S.rouxii cells, an optimum pH could not be calcu-lated; instead, comparable growth rates wereobtained over a wide pH range. The optimumpH range was 3.5 to 5.5 for S. rouxii.
TABLE 1. Growth response of S. rouxii at differentpH levels in 2x PDB-12% glucose
Responsea of inoculum grown:
pH Without In 60%sucrose sucrose
1.0 - NT1.5 + +2.0 NT + (8.78)2.5 ++ (5.13) + + (2.85)3.0 + + (2.27) ++ (2.68)3.5 ++ (2.20) + + (2.47)4.0 + + (2.27) + + (2.60)4.5 + + (2.26) ++ (2.50)5.0 ++ (2.16) + + (2.57)5.5 + + (2.15) + + (2.60)6.0 + + (2.23) + + (2.67)6.5 ++ (2.25) NT7.0 + + + + (2.67)
10.5 + + NT11.0 -11.5 - NT12.0 - NT
a Symbols: + +, normal growth; +, poor growth;-,no growth; NT, not tested. Numbers within parenthe-ses are generation times (in hours).
APPL. ENVIRON. MICROBIOL.
on June 25, 2018 by guesthttp://aem
.asm.org/
Dow
nloaded from
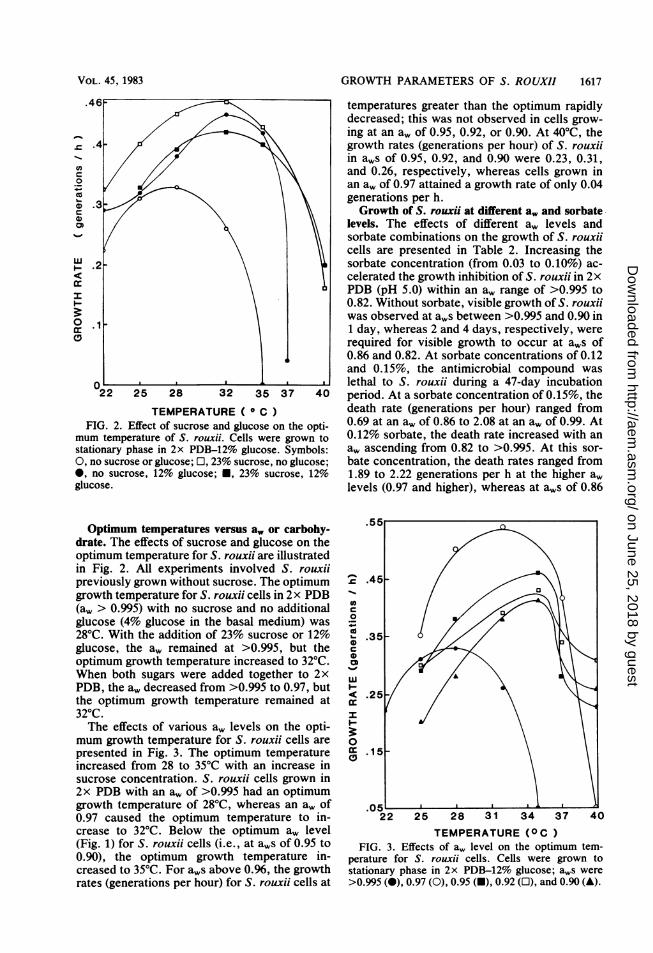
GROWTH PARAMETERS OF S. ROUXII 1617
0
CD .3c
0
cm
2
01
022 25 28 32 35 37 40
TEMPERATURE ( ° C)FIG. 2. Effect of sucrose and glucose on the opti-
mum temperature of S. rouxii. Cells were grown tostationary phase in 2x PDB-12% glucose. Symbols:0, no sucrose or glucose; E, 23% sucrose, no glucose;0, no sucrose, 12% glucose; U, 23% sucrose, 12%glucose.
Optimum temperatures versus a, or carbohy-drate. The effects of sucrose and glucose on theoptimum temperature for S. rouxii are illustratedin Fig. 2. All experiments involved S. rouxiipreviously grown without sucrose. The optimumgrowth temperature for S. rouxii cells in 2 x PDB(a, > 0.995) with no sucrose and no additionalglucose (4% glucose in the basal medium) was28°C. With the addition of 23% sucrose or 12%glucose, the a, remained at >0.995, but theoptimum growth temperature increased to 32°C.When both sugars were added together to 2xPDB, the a, decreased from >0.995 to 0.97, butthe optimum growth temperature remained at320C.The effects of various a, levels on the opti-
mum growth temperature for S. rouxii cells arepresented in Fig. 3. The optimum temperatureincreased from 28 to 35°C with an increase insucrose concentration. S. rouxii cells grown in2x PDB with an a, of >0.995 had an optimumgrowth temperature of 28°C, whereas an a, of0.97 caused the optimum temperature to in-crease to 320C. Below the optimum a, level(Fig. 1) for S. rouxii cells (i.e., at aWs of 0.95 to0.90), the optimum growth temperature in-creased to 35°C. For aWs above 0.96, the growthrates (generations per hour) for S. rouxii cells at
temperatures greater than the optimum rapidlydecreased; this was not observed in cells grow-ing at an a. of 0.95, 0.92, or 0.90. At 40°C, thegrowth rates (generations per hour) of S. rouxiiin aWs of 0.95, 0.92, and 0.90 were 0.23, 0.31,and 0.26, respectively, whereas cells grown inan a, of 0.97 attained a growth rate of only 0.04generations per h.Growth of S. rouxii at different a, and sorbate
levels. The effects of different a, levels andsorbate combinations on the growth of S. rouxiicells are presented in Table 2. Increasing thesorbate concentration (from 0.03 to 0.10%) ac-celerated the growth inhibition of S. rouxii in 2xPDB (pH 5.0) within an a, range of >0.995 to0.82. Without sorbate, visible growth of S. rouxiiwas observed at a,s between >0.995 and 0.90 in1 day, whereas 2 and 4 days, respectively, wererequired for visible growth to occur at a,s of0.86 and 0.82. At sorbate concentrations of 0.12and 0.15%, the antimicrobial compound waslethal to S. rouxii during a 47-day incubationperiod. At a sorbate concentration of 0.15%, thedeath rate (generations per hour) ranged from0.69 at an a, of 0.86 to 2.08 at an a, of 0.99. At0.12% sorbate, the death rate increased with ana, ascending from 0.82 to >0.995. At this sor-bate concentration, the death rates ranged from1.89 to 2.22 generations per h at the higher a,levels (0.97 and higher), whereas at a,s of 0.86
2.45-
0
0~
.0s , , .22 25 28 31 34 37 40
TEMPERATURE ( °C )FIG. 3. Effects of a, level on the optimum tem-
perature for S. rouxii cells. Cells were grown tostationary phase in 2x PDB-12% glucose; a,s were>0.995 (0), 0.97 (0), 0.95 (-), 0.92 ([), and 0.90 (A).
VOL. 45, 1983
on June 25, 2018 by guesthttp://aem
.asm.org/
Dow
nloaded from

1618 RESTAINO ET AL.
TABLE 2. Effects of a, levels versus sorbateconcentrations on growth of S. rouxiia
Effectb with the following potassium sorbateaw concn:
0.00%0 0.03% 0.06% 0.08% 0.10%90.12% 0.15%
>0.995 1 d 3 d 1.41 1.82 1.92 2.22 1.750.99 1 d 2 d 15 d 1.85 2.38 2.17 2.080.98 1 d 2 d 6 d 14 d 1.47 1.82 1.720.97 1 d 2 d 3 d 8 d 0.30 1.89 1.790.96 1 d 2 d 3 d 5 d 20 d 0.72 1.790.95 1 d 2 d 3 d 5 d 0.92 1.720.92 1 d 2 d 4 d 5 d 13 d 0.77 1.850.90 1 d 3 d 5 d 6 d 13 d 0.84 1.240.86 2 d 5 d 6 d 8 d 0.60 0.55 0.690.82 4 d 8 d 0.40 0.43 0.48 0.61 1.41
a The experiment was performed for 47 days at320C.bNumbers followed by the letter d (days) indicate
the number of days before visible growth could beobserved. Numbers without the letter d indicate thedeath rate (number of generations per hour).
and 0.82, the death rates for S. rouxii cells were0.55 and 0.61, respectively.At sorbate concentrations of 0.03 to 0.10%,
the day that visible growth of S. rouxii cells wasobserved depended on the aw level. Growth wasinitially observed in 2 days in 2x PDB contain-ing 0.03% sorbate at aws ranging from 0.92 to0.99, whereas 3, 5, and 8 days, respectively,were required for visible growth at aws of>0.995 and 0.90, 0.86, and 0.82. At 0.06%sorbate, aws of 0.95 to 0.97 required 3 daysbefore there was visible growth of S. rouxii,whereas death rates of 0.40 and 0.41 generationsper h, respectively, occurred for S. rouxii at awsof 0.82 and >0.995. At 0.08 and 0.10% sorbate,the width of the aw range that supported S.rouxii growth decreased compared with that atthe lower sorbate concentrations. At 0.08% sor-bate, aws of 0.92, 0.95, and 0.96 supported thefastest growth rate of S. rouxii, but death oc-curred at aws of >0.99. At 0.10% sorbate, thisyeast propagated fastest at aws of 0.90 and 0.92,exemplified by visible growth in 13 days, where-as death occurred for S. rouxii at an aw above0.97.
DISCUSSIONThe most common of the osmotolerant yeasts
is S. rouxii (32, 37), which can grow at an aw of0.62 when other extrinsic conditions are optimal(27). Using sucrose broths, Tilbury (M.S. thesis)showed that the optimum aw for growth of S.rouxii is 0.98. In the present investigation, theoptimum aw of the S. rouxii strain isolated fromchocolate syrup was 0.97 and 0.96, respectively,for cells previously grown in 0 and 60% sucrose.Anand and Brown (1) have shown slight varia-
tions in growth rate patterns at different a,s fortwo S. rouxii strains. Slight discrepancies inoptimum a,s could be due to the basal growthmedia, slight genetic variations within species,preconditioning with respect to aw, or a combi-nation of these. The data from this investigationshow the influence of acclimation in determiningthe optimum aw for an osmotolerant yeast strain.When investigating the optimum aw for an osmo-tolerant yeast, the preconditioning becomes ex-tremely important.The pH growth range of S. rouxii is very wide
(1.5 to 10.5) (Table 1), enabling this microorgan-ism to survive or grow in a wide range of foods.This wide pH tolerance can allow this yeast tocross-contaminate a variety of foods under con-ditions where sofne commodities could containparameters favorable for osmotolerant yeastspoilage. The optimum pH range of this S. rouxiistrain was 3.5 to 5.5, which relates to the pHrange (4.0 to 4.5) of S. rouxii calculated byTilbury (M.S. thesis). However, the pH opti-mum range in this investigation was substantial-ly larger than the pH range determined by Til-bury (M.S. thesis), indicating influences ofgrowth media or strain variations or both.The optimum pH range and aw levels of S.
rouxii do not coincide with the pH and awparameters for the types offoods spoiled by thisyeast. Other intrinsic or extrinsic characteristicsmust be involved. Acidic foods such as tomatopaste, canned fruits in heavy syrups, margarine,canned cured meats, Gouda cheese, and low-saltfish, pork, and beef products (10, 33) are notnormally spoiled by S. rouxii even though theiraw levels (0.92 to 0.98) are near the optimum forS. rouxii. However, products that are usuallyspoiled by S. rouxii have lower aw levels (0.80 to0.85); these products include honey, maple syr-up, fruit syrups, jams, liquid sugar, soy mashes,and chocolate syrups (24, 37; unpublished data).The pHs of these low-aw foods are in the rangeoptimal for S. rouxii growth. Since a variety ofbacterial strains can proliferate faster than os-motolerant yeasts at aws ranging from 0.97 to>0.995, this yeast cannot compete and spoil aproduct suitable for bacterial growth. In aharsher environment, i.e., in products contain-ing a low aw and pH, most bacteria either cannotgrow or grow very slowly. Spoilage will mostlikely be caused by yeasts or mold. Additionalfactors involving the numbers and types of os-motolerant yeasts initially infecting the food, thetypes of nutrients, the redox potential (liquid orsolid food), and the temperature during storage(37) can enable the osmotolerant yeasts to out-grow the xerotolerant molds. As the yeastsproliferate, the production of ethanol and CO2further creates an environment detrimental toxerotolerant molds (32).
APPL. ENVIRON. MICROBIOL.
on June 25, 2018 by guesthttp://aem
.asm.org/
Dow
nloaded from

GROWTH PARAMETERS OF S. ROUXII 1619
As noted by other researchers (16, 23), adecrease in a. causes the optimum temperatureof S. rouxii to increase (Fig. 2 and 3). AlthoughS. rouxii can rapidly metabolize glucose, where-as sucrose either is not fermented or is ferment-ed extremely slowly (18), the optimum tempera-ture for S. rouxii increases regardless of whethersucrose or glucose is used as the humectant.Recently, increased recovery rates of osmoto-lerant yeasts from intermediate-a, foods, e.g.,fruit juice concentrates, have been reported onmedia with elevated sugar concentrations (40).Since the optimum growth temperature range forS. rouxii in a broth containing an a, between0.90 and 0.97 is 32 to 35°C, an incubation tem-perature of 32°C, instead of the traditional 22 to25°C (34) for 5 days, would be more efficient forenumerating osmotolerant yeasts in the pres-ence of elevated sugar concentrations. In addi-tion, S. bailii, a yeast less osmotolerant than S.rouxii, has an optimum temperature of 32°C atan aw of 0.96 (unpublished data), increasing theflexibility of this incubation temperature forenumerating a wide range of osmotolerantyeasts.Why was an increased optimum temperature
of S. rouxii observed when it was inoculated in agrowth medium containing reduced a, levels?One explanation could involve the protectiveeffect of the solute on the metabolic mechanismsand cellular integrity of S. rouxii. The heatresistance of microorganisms (vegetative cellsand spores) increases with elevated concentra-tions of solutes (4, 11, 12, 14). Doyle and Marth(12) and Gibson (14) have explained this in-creased heat resistance as a dehydrating phe-nomenon resulting in a greater stability of cellu-lar components, specifically, proteins. In thepresent investigation, a similar explanationcould be proposed for the elevated optimumtemperature of S. rouxii with a decreasing aw.However, since a greater concentration of glu-cose (30%) or sucrose (45%) in the heatingmenstruum was required to protect S. rouxiifrom heat (12, 14) than was needed to increasethe optimum temperature, a simple dehydrationphenomenon of cells may not be the total expla-nation. A second explanation, involving the pro-duction of intracellular polyols (9) by S. rouxii,is also feasible. Intracellular polyols have beenshown to function as compatible solutes (6, 7, 9),which relates to the protection of enzymes inyeasts against inhibition or inactivation. Theconcentrations of intracellular polyols in osmo-tolerant yeasts can accumulate rapidly in cellsgrowing in decreasing a, levels (6, 13). Only aslight reduction in a, can cause a rapid increaseof intracellular polyols in osmotolerant yeasts.When S. rouxii grows in a high sugar concentra-tion, the polyol that accumulates is arabitol (20),
whose concentration is directly proportional tothe a, level. Although the mechanism of polyolprotection toward enzymes had not been eluci-dated, this system could be related to heat-sensitive enzymes with respect to elevated tem-peratures. Thus, intracellular polyols couldenable S. rouxii to proliferate at a higher tem-perature, resulting in an increased optimum tem-perature.With the relationship between potassium sor-
bate tolerance of S. rouxii and different a,levels, some hypotheses or thoughts can beformulated on the mechanism of sorbate resist-ance in osmotolerant yeasts, specifically, S.rouxii. In 1977, Warth (38) proposed a mecha-nism of resistance of S. bailii to benzoic andsorbic acids, involving an inducible energy-re-quiring system which transports the preserva-tive out of the cell. Since elevated glucose levelsin the growth media cause S. bailii to becomemore resistant to the preservative (25), aerobicrespiration of glucose could provide the energyneeded to pump the preservative out of the cell(38). In the present investigation, each a, levelcontained 16% glucose, and the cultures werevigorously shaken to permit sufficient aeration.If Warth's theory (38) can be applied to S.rouxii, the sorbate resistance of this yeastshould not be markedly influenced by moderateaw changes. For a particular sublethal sorbatelevel, the growth rate of S. rouxii should befastest near the optimum aw level. However, thehigher aws (0.98 to >0.995) enhanced the anti-mycotic activity of sorbate against the growth ofS. rouxii, whereas lower aws (0.90 to 0.92,farther from the optimum) increased the toler-ance of this yeast to higher concentrations ofsorbate.From the data obtained in this investigation,
there are two other possibilities involving themechanism of resistance of S. rouxii to sorbate.At lower aw levels (higher solute concentrations)the cells shrink (11), causing the pore size on thecell membrane to become smaller, which mayretard the flow of sorbate into the cell. If thistheory is valid, the induced energy system(Warth's theory [38]) to transport sorbate out ofthe cell could still be plausible. At lower awlevels the flow of sorbate into the cell will beretarded, allowing the energy formed by glucoserespiration to pump the sorbate out at a rate atleast equivalent to that of the uptake of thepreservative. At the higher aw values and sor-bate levels (>0.06%), the uptake of sorbatewould be too rapid, causing an intracellularaccumulation of the preservative.The resistance of S. rouxii to sorbate at differ-
ent aw values could also involve the productionof polyols (6) functioning as a compatible solute(7). Potassium sorbate inhibits various enzyme
VOL. 45, 1983
on June 25, 2018 by guesthttp://aem
.asm.org/
Dow
nloaded from

1620 RESTAINO ET AL.
systems (2, 15, 19, 39). With decreasing awlevels, increasing amounts of polyols will beproduced or retained intracellularly to maintainosmoregulation (8) and will function as a com-patible solute protecting various enzymes fromthe inhibition or inactivation of certain antimi-crobial agents. At the higher aws (0.98 to>0.995), the intracellular polyol levels would betoo low to effectively counteract the antimycoticeffectiveness of the preservative. When the aw isgreatly reduced (<0.85), the growth rate of S.rouxii is extremely retarded (low metabolicrate), which could cause a depressed rate ofpolyol production, allowing the yeast cells tobecome more sensitive to the preservative. Thistheory of sorbate resistance is further supportedby the data of Bills et al. (5), who have deter-mined the various growth responses of precondi-tioned S. rouxii cells in media containing differ-ent levels of sucrose and sorbate. Furtherstudies are being conducted to analyze intracel-lular polyols at different aw levels and relate thisto sorbate resistance of S. rouxii.As noted above, the aw level of the medium
influenced the resistance of S. rouxii to potassi-um sorbate. S. rouxii can tolerate increasingconcentrations of potassium sorbate at decreas-ing aw levels. The addition of potassium sorbate(0.1%) to a liquid, intermediate-moisture food(pH 4.5 to 5.5) such as syrup toppings will notensure that the growth of osmotolerant yeastscan be retarded. Consequently, a variety offactors must be considered, including the chemi-cal constituents of the food (sugar concentra-tion, salt level, fat content, and pH), processing,packaging, storage temperature and length, anduse of additional antimicrobial agents that mightenhance or hinder the effectiveness of sorbate(29).
ACKNOWLEDGMENT
We greatly appreciate the DNA hybridization experimentsperformed by C. P. Kurtzman.
LITERATURE CITED
1. Anand, J. C., and A. D. Brown. 1968. Growth rate pat-terns of the so-called osmophilic and non-osmophiliclyeasts in solutions of polyethylene glycol. J. Gen. Micro-biol. 52:205-212.
2. Azukas, J. J., R. N. Costilow, and H. L. Sadoff. 1961.Inhibition of alcoholic fermentation by sorbic acid. J.Bacteriol. 81:189-194.
3. Baird-Parker, A. C., and W. J. Kooiman. 1980. Softdrinks, fruit juices, concentrates, and fruit preserves, p.
643-668. In International Commission on MicrobiologicalSpecifications for Foods (ed.), Microbial ecology offoods,vol. 2. Academic Press, Inc., New York.
4. Beuchat, L. R. 1981. Combined effects of solutes and foodpreservatives on rates of inactivation of and colony forma-tion by heated spores and vegetative cells of molds. Appl.Environ. Microbiol. 41:472-477.
5. Bills, S., L. Restaino, and L. M. Lenovich. 1982. Growthresponse of an osmotolerant sorbate-resistant yeast, Sac-charomyces rouxii, at different sucrose and sorbate levels.
J. Food Prot. 45:1120-1125.6. Brown, A. D. 1974. Microbial water relations: features of
the intracellular composition of sugar-tolerant yeasts. J.Bacteriol. 118:769-777.
7. Brown, A. D. 1976. Microbial water stress. Bacteriol Rev.40:803-846.
8. Brown, A. D., and M. Edgley. 1980. Osmoregulation inyeast, p. 75-90. In D. W. Rains, R. C. Valentine, and A.Hollaender (ed.), Genetic engineering of osmoregulation.Plenum Publishing Corp., New York.
9. Brown, A. D., and J. R. Simpson. 1972. Water relations ofsugar-tolerant yeasts: the role of intracellular polyols. J.Gen. Microbiol. 72:589-591.
10. Christian, J. H. B. 1980. Reduced water activity, p. 70-91. In International Commission on MicrobiologicalSpecifications for Foods (ed.), Microbial ecology offoods, vol. 1. Academic Press, Inc., New York.
11. Corry, J. E. L. 1976. The effect of sugars and polyols onthe heat resistance and morphology of osmophilic yeasts.J. Appl. Bacteriol. 40:269-276.
12. Doyle, M. P., and E. H. Marth. 1975. Thermal inactiva-tion of condia from Aspergillus flavus and Aspergillusparasiticus. J. Milk Food Technol. 38:750-758.
13. Edgley, M., and A. D. Brown. 1978. Response of xeroto-lerant and non-tolerant yeasts to water stress. J. Gen.Microbiol. 104:343-345.
14. Gibson, B. 1973. The effect of high sugar concentrationson the heat resistance of vegetative microorganisms. J.Appl. Bacteriol. 36:365-376.
15. Harada, K., R. Higuchi, and I. Utsunii. 1968. Studies onsorbic acid. IV. Inhibition of the respiration of yeast.Agric. Biol. Chem. 32:936-940.
16. Ingram, M. 1959. Physiological properties of osmophilicyeasts. Rev. Ferment. Ind. Aliment. 14:23-33.
17. Kurtzman, C. P., M. J. Smiley, C. J. Johnson, L. J.Wickerham, and G. B. Fuson. 1980. Two new and closelyrelated heterothallic species, Pichia amylophila and Pi-chia mississippiensis: characterization by hybridizationand deoxyribonucleic acid reassociation. Int. J. Syst.Bacteriol. 30:208-216.
18. Lodder, J. 1971. The yeasts. North-Holland PublishingCo., Amsterdam.
19. Martoadipraivito, W., and J. R. Whitaker. 1963. Potassi-um sorbate inhibition of yeast alcohol dehydrogenase.Biochim. Biophys. Acta 77:536-544.
20. Moran, J. W., and L. D. Witter. 1979. Effects of sugars onD-arabitol production and glucose metabolism in Saccha-romyces rouxii. J. Bacteriol. 138:823-831.
21. Mrak, E. M., H. J. Phaff, R. H. Vaughn, and H. N.Hansen. 1942. Yeasts occurring in souring figs. J. Bacteri-ol. 44:441-450.
22. Nury, F. S., M. W. Miller, and J. E. Brekke. 1960. Preser-vative effect of some antimicrobial agents on high-mois-ture dried fruits. Food Technol. (Chicago) 14:113-115.
23. Onishi, H. 1959. Studies on osmophilic yeasts. V. Factorsaffecting growth of soy yeasts and others in the environ-ment of a high concentration of sodium chloride (2). Bull.Agric. Chem. Soc. Jpn. 23:351-360.
24. Onishi, H. 1963. Osmophilic yeasts. Adv. Food Res.12:53-94.
25. Pitt, J. I. 1974. Resistance of some food spoilage yeasts topreservatives. Food Technol. Aust. 26:238-241.
26. Pitt, J. I., and K. C. Richardson. 1973. Spoilage by pre-servative-resistant yeasts. CSIRO Food Res. Q. 33:80-85.
27. Scott, W. J. 1957. Water relations of food spoilage micro-organisms. Adv. Food Res. 7:83-127.
28. Seiler, D. A. L. 1980. Yeast spoilage of bakery products,p. 135-152. In F. A. Skinner, S. M. Passmore, and R. R.Davenport (ed.), Biology and activities of yeasts. Aca-demic Press, Inc., New York.
29. Sofos, J. N., and F. F. Busta. 1981. Antimicrobial activityof sorbate. J. Food Prot. 44:614-622.
30. Splittstoesser, D. F. 1978. Fruits and fruit products, p. 83-110. In L. R. Beuchat (ed.), Food and beverage mycolo-gy. AVI Publishing Co., Westport, Conn.
APPL. ENVIRON. MICROBIOL.
on June 25, 2018 by guesthttp://aem
.asm.org/
Dow
nloaded from

VOL. 45, 1983
31. Tilbury, R. H. 1976. The microbial stability of intermedi-ate moisture foods with respect to yeasts, p. 138-165. InR. Davies, G. G. Birch, and K. J. Parker (ed.), Intermedi-ate moisture foods. Applied Sciences, London.
32. Tilbury, R. H. 1980. Xerotolerant (osmophilic) yeasts, p.153-179. In F. A. Skinner, S. M. Passmore, and R. R.Davenport (ed.), Biology and activities of yeasts. Aca-demic Press, Inc., New York.
33. Troller, J. A., and J. H. B. Christian. 1978. Water activityand food. Academic Press, Inc., New York.
34. U.S. Food and Drug Administration. 1978. Bacteriologicalanalytical manual. Bureau of Foods, U.S. Food and DrugAdministration, Washington, D.C.
35. Von Schelhorn, M. 1950. Untersuchungen uber den Ver-derb wasserarmer Lebensmittel durch osmophile Mik-roorganismen. I. Verberb von Lebensmittel durch osmo-
GROWTH PARAMETERS OF S. ROUXII 1621
phile Hefen. Z. Lebensm. Unters. Forsch. 91:117-124.36. Walker, H. W. 1977. Spoilage of food by yeasts. Food
Technol. (Chicago) 31:57-61.37. Walker, H. W., and J. C. Ayres. 1970. Yeasts as spoilage
organisms, p. 463-527. In A. H. Rose and J. S. Harrison(ed.), The yeasts, vol. 3. Academic Press, Inc., NewYork.
38. Warth, A. D. 1977. Mechanism of resistance of Saccharo-myces bailii to benzoic, sorbic, and other weak acids usedas food preservatives. J. Appl. Bacteriol. 43:215-230.
39. Whitaker, J. R. 1959. Inhibition of sulfhydryl enzymeswith sorbic acid. Food Res. 24:37-43.
40. Zottola, E. A. 1976. Osmophilic microorganisms, p. 203-207. In M. L. Speck (ed.), Compendium of methods forthe microbiological examinations of foods. American Pub-lic Health Association, Washington, D.C.
on June 25, 2018 by guesthttp://aem
.asm.org/
Dow
nloaded from





























